Submitted:
03 July 2025
Posted:
04 July 2025
You are already at the latest version
Abstract
It has been demonstrated that the influenza virus possesses substantial global epidemic potential, with higher incidence rates observed in obese individuals and prolonged recovery times. Obesity affects millions of individuals globally, and a deeper understanding of its associated physiological disturbances is essential for addressing key public health concerns. Obese individuals exhibit impaired immune organ function, decreased immune cell activity, disrupted metabolism characterized by mitochondrial dysfunction and an imbalance in gut microbiota associated with intestinal mucosal barrier damage. The gut microbiota and their metabolic composition in obese patients differ from those in non-obese individuals, potentially promoting viral replication and exacerbating disease severity. These factors collectively contribute to more severe tissue damage and heightened immune responses in obese patients during influenza infection. Therefore, understanding the impact of obesity on influenza virus infection dynamics enables the development of strategies promoting healthy lifestyles to manage body weight and enhance immunity against viral infections. Additionally, given that this special population may not respond optimally to antimicrobial drugs and vaccination, it is necessary to consider how treatment strategies for this group are managed. This review illustrates findings concerning impact of obesity on the immune response to influenza virus infection, including potential underlying mechanisms.
Keywords:
influenza virus
; obesity
; gut microbiota
; immunity
1. Introduction
Influenza virus is a highly contagious pathogen that imposes a considerable socio-economic burden of morbidity and mortality globally. Epidemiological reports show, periodically occurring influenza pandemics can result in higher mortality rates compared to seasonal outbreaks [1,2]. Since the outbreak of the H1N1 Spanish influenza in 1918 [3], numerous influenza pandemics have occurred, such as the H2N2 Asian influenza in 1957 [4], the H3N2 Hong Kong influenza in 1968 [5], and the H1N1 influenza pandemic in 2009 [6]. Since then, influenza virus infection has been responsible for numerous global pandemics, resulting in tens of millions of deaths and extensive lung injury, suggesting that the virus was significantly associated with severe respiratory pathology.
Obesity is the product of the combined influence of biological and environmental factors, resulting in the increase of excess adipose tissue in the body, which will lead to the increase of morbidity and mortality of certain diseases [7]. Despite the implementation of nutritional interventions and physical education programs, the global prevalence of obesity continues to increase, with the World Health Organization estimating that 20% adults worldwide will be overweight by 2035 [8]. The most recent data from the "China Nutrition and Chronic Disease Status Report" indicates that the prevalence of overweight and obesity among Chinese residents aged 18 and above has surpassed 50%, with the obesity rate among children and adolescents aged 6 to 17 approaching 20%, and the obesity rate among children under 6 years old reaching 10% [9]. The data reflects a significant increase in the risk of chronic diseases, such as cardiovascular disease, diabetes, and cancer. In particular, the risk of developing diabetes increases by 27% for every 5 kilograms gained [10]. Although nutritional status has long been recognized as a critical determinant of the body's response to infections, weight increase in the progression and transmission of infectious diseases has recently gained increasing attention. It has been documented that H1N1 can replicate in human primary adipocytes, indicating that obesity may serve as an independent risk factor for increased severity of influenza infection [11].Therefore, considering escalating obesity epidemic and its association with influenza complications, it is essential to investigate the key metabolic, gut microbiota, and immune dysregulations during influenza virus infection, as well as their impact on promoting intra-host viral diversity.
2. The Epidemiological Association Between Obesity and Influenza Infection
Obesity has been associated with a variety of respiratory disorders, such as chronic obstructive pulmonary disease, asthma, pulmonary embolism, and aspiration pneumonia. An in-depth data analysis has shown that obesity is an independent risk factor linked to morbidity and mortality during the influenza A (H1N1) pandemic [12]. Concretely, obesity can influence the host's resistance to the influenza virus in multiple aspects, including the lung, liver, intestines, and immunity (Figure 1). These effects manifest as reduced lung volume, abnormalities in respiratory muscle function, and impaired gas exchange [13]. Impaired lung function significantly promotes influenza virus infection and subsequent lung injury, which are further aggravated by sleep apnea and chronic inflammation [14].
Higher BMI has been identified as an additional risk factor for hospitalization. Clinical evidence demonstrates that the degree of adiposity correlates with disease severity following infection. Influenza viral RNA has been detected in the aerosols with a positive correlation observed between aerosol viral load and BMI [15]. In addition to infections mentioned before, increase BMI has been positively correlated with adiposity in the airway wall, wall thickness, and inflammation, which can lead to asthma-related deaths [16].
During influenza season, the hospitalization incidences and hospital stay among obese individuals were found to increase, suggesting obesity hinders the recovery process [17]. Furthermore, obesity delays the clearance of the influenza virus load and extends the shedding duration, thereby leading to long-term transmission [18]. A severe case confirmed that obese patients infected with the H1N1 virus have a two-fold greater likelihood of being admitted to the Intensive Care Unit (ICU) [19]. A multicenter study involving 144 ICUs in Spain indicated that obesity was linked to higher ICU resource consumption and longer hospitalizations among individuals infected with H1N1 [20]. Among patients admitted to the ICU because of H1N1 infection, obese and morbidly obese patients are more prone to developing pneumonitis than non-obese patients [21]. Even younger patients over the age of 20 years with underlying co-morbidities such as obesity and diabetes had a higher hospitalization rate and mortality [12,22,23]. In addition, obesity rates of up to 6.8% have been found among the pediatric patients hospitalized due to influenza, including young children aged under 5 years [24]. Fever is the main symptom in up to 90% of elderly patients and 97-99% of children, lasting approximately 5 days [25]. Individuals who are obese also frequently experience fever during influenza, and the duration of the fever is notably longer. Thus, data from the influenza pandemic indicates that obesity poses a risk regardless of age.However, the exact mechanism by which obesity affects influenza virus is not yet clear. We summed up some potential new protein targets such as α/β-hydrolase domain 6, Histone deacetylase 6, G-protein-coupled receptor 4 and so on can affect obesity and influenza(Table 1).
3. The Relationship Between Obesity and Gut Microbiota
Obesity not only leads to an increased hospitalization rate and prolonged hospital stays among influenza patients, but also causes intestinal flora imbalance in these individuals. The association between obesity and gut microbiota has been investigated in animal models and human patients. Since the early stages of human microbiome research, a remarkable difference in the composition of the gut microbiome has been detected in obese individuals [36]. Menni et al. pointed out that patients with a more diverse gut microbiota exhibited a lower likelihood of experiencing long-term weight gain, suggesting that a stable gut microbiota may potentially play a role in preventing excessive weight fluctuations, including both gain and loss [37]. Of note, a decrease in Akkermansiamuciniphila has most consistently been observed to be associated with obesity [38].Other studies concentrated on the disparities detected in the Bacillota/Bacteroidota ratio among obese and lean individuals [39]. However, the use of the Bacillota/Bacteroidota ratio as a marker for obesity is somewhat controversial. As reviewed, other studies reported discrepancies where obese patients presented a decreased Bacillota/Bacteroidota ratio [40]. Another element contributing to these inconsistencies is linked to the metabolic endotoxemia hypothesis. This hypothesis suggests that elevated adiposity and the development of low-grade inflammation might be associated with the spread of LPS. Obesity is intricately associated with low-grade chronic inflammation. A high-fat diet has the potential to modify the composition of the gut microbiota. This modification results in an elevation of endotoxins in the intestine, subsequently augmenting intestinal permeability. The gut microbiota activates immune signaling pathways by modulating intestinal permeability. This activation elicits chronic, low-grade inflammatory responses, which in turn heightens the risk of obesity-related disorders [41]. However, this theory conflicts with the decreased levels of Bacteroidota found in obese patients. The reason is that this phylum mainly consists of Gram-negative bacteria, which are rich in LPS [40]. These discrepancies might also be associated with the experimental procedures employed in various studies, such as the number of participants, the methods utilized, and even the heterogeneity among participants.
Furthermore, scientific evidence suggests that gut microorganisms may extract additional energy from the diet, contributing to the link between gut microbiota and obesity. The gut microbiota breaks down certain dietary compounds that would not be digested otherwise, like some carbohydrates and fibers. Subsequently, the intestinal microbiota transforms these into short-chain fatty acids (SCFA). SCFA account for 10% of the daily energy intake of humans and are crucial for colon and liver cells. It has been discovered that when both lean and obese adults consume a diet with an excessive calorie amount, lean individuals expel more energy through feces compared to obese individuals [42]. However, a central theme regarding the relationship between obesity and microbiota is that diet significantly influences the composition of the intestinal microbiota, and as a result, affects the host's health. A Western-style diet, which is high in saturated fats and low in fiber, can trigger changes in gut microbiota. This leads to a decrease in the abundance of beneficial microorganisms (dysbiosis) and an increase in inflammation. Such a pattern is frequently associated with the development of metabolic disorders like obesity [43]. Therefore, metabolic disorders in obese individuals can disrupt the intestinal microbiota, ultimately resulting in an elevated risk of influenza virus infection.
4. Mechanisms of the Interaction Between Gut Microbes and the Respiratory Immune System
Obesity not only affects influenza virus infection through metabolic disorders, but also indirectly increases the risk of virus invasion by reshaping the structure of intestinal flora. Gut microbiota is the "second brain" of the host immune system, and its imbalance may break the gut-lung axis, leading to systemic immunosuppression and local mucosal defense defects.
The gut microbiota can regulate respiratory immune responses in two primary ways. First, it generates systemic factors that influences the airways. Second, it directs the recirculation of immune cells from the gut, thereby modulating immune responses in the lungs. For instance, when type 2 innate lymphoid cells (ILC2) are activated either by IL-25 or as a result of gut helminth infection, they migrate to the airways, thereby facilitating both tissue repair and immune responses against helminths [44]. Microbial colonization in early life also leads to the seeding of invariant natural killer (NK) T cells in both the gut and lungs. These cells offer long-term protection against inflammatory bowel disease (IBD) and allergy [45]. The migration of immune cells to the lung, which is also known as homing, is intricate and mainly relies on inflammatory signals [46]. Interestingly, at the steady state, the homing of immune cells is affected by the circadian rhythm [47]. Even though the microbiota has been demonstrated to have a definite role in the trafficking of immune cells to the gut, it remains uncertain whether the microbiome controls the trafficking of immune cells to the airways under normal, steady-state conditions. [48].
In addition, the gut microbiota can secrete a diverse array of metabolites that are not inherently present in the human body. Elaborate experiments involving the administration of isotype-labeled bacteria to germ-free (GF) mice revealed that metabolites derived from microorganisms can be detected in nearly all tissues. These tissues include the bone marrow, brain, and adipose tissue. Regrettably, there was no evidence indicating that lung tissue was examined in that study [49]. To further corroborate this phenomenon, it was recently demonstrated that gut-derived butyrate can reach the skin and affect keratinocyte function within 45 minutes after administration [50]. The gut-lung-immune axis might also operate indirectly and be mediated by other organs. For example, multiple studies indicate that the gut-lung axis achieves its effect by regulating hematopoiesis in the bone marrow [51,52,53,54]. The gut microbiome can communicate with distal organs by generating various types of factors. Hence, the microbiome undergoes continuous cycles of division and death, which leads to the substantial release of bacterial cell wall components and lung injury [55].
Moreover, the gut microbiota exhibits high metabolic activity and synthesizes a wide range of microbial by-products. These microbe-derived molecules are not naturally present in the human body. Conversely, these by-products are absorbed into the peripheral tissues and may interact with the immune system. Before reaching the respiratory tract, they can be further metabolized by the host. Together, gut microbiota is likely to exert a broader influence on immune homeostasis, which in turn affects influenza infection.
5. Gut-Lung Axis in Influenza Virus Infection
Respiratory viral infections exert bidirectional regulatory effects on the gut microbiota. In turn, the gut microbiota modulates immune defense mechanisms against viral pathogens in the respiratory tract. It has been reported that Influenza A infection can cause dysbiosis in the gut and allow the overgrowth of pathogenic bacteria [56,57]. Although the influenza virus exhibits relatively low replication activity in the gastrointestinal tract, its infection can still modulate the composition of the intestinal microbiota through indirect mechanisms, such as impaired nutrient absorption [57]. This process occurs upon the activation of immune cells in the lung, after which the activated immune cells migrate back to the gut [58]. During an immune response, effector T cells migrate to nonlymphoid tissue (NLT) depending on chemokine receptor expression and chemokine production at the infection site [59]. It was discovered that the production of IFN-γ and IL-17 in the lungs, which was induced by influenza, enhanced the expression of CCR9 on CD4+ T cells. This increase enabled the CD4+ T cells to migrate to the gut. Once in the gut, they facilitated Th17-mediated intestinal inflammation and tissue damage, ultimately resulting in microbial dysbiosis [58].
The most well-established way by which the gut microbiota conveys information to the lung is through the generation of microbial metabolites. Among these metabolites, SCFAs have been acknowledged to regulate antiviral immunity in the lung via multiple mechanisms [60]. Firstly, it has been reported that SCFAs and other metabolites derived from microbes enhance the metabolism and effector function of CD8+ T cells throughout the body. This includes the ability to kill virus-infected cells in the lung [53,61]. Mechanistically, butyrate metabolically reprograms CD8+ T cells by promoting glutamine utilization and fatty acid oxidation, positioning them for optimal recall responses to secondary infection [62]. Environmental factors that change food intake can have an indirect impact on SCFA levels. Interestingly, a high ambient temperature enhanced autophagy in the lungs, weakened inflammasome-dependent cytokine secretion, and affected virus-specific CD8+ T cell responses [61]. In addition to the aforementioned findings, other CD markers have demonstrated significant efficacy in combating viral infections. Our review integrates extensive studies that highlight the critical roles played by various T cells-associated CDs during viral infections (Table 2).
A parallel mechanism by which mice supplemented with butyrate or inulin were safeguarded against influenza infection is related to the bone marrow [70]. Additionally, mice fed a high-fiber diet exhibited augmented macrophage-dendritic cell progenitors in the bone marrow [71]. This led to the production of Ly6c patrolling monocytes and an increase in the number of alternatively activated macrophages in the airways. Upon viral exposure, these cells generated less CXCL1. As a consequence, neutrophil infiltration was diminished, and immunopathology was restricted. Nevertheless, this attenuated response of alveolar macrophages might make the immune system more vulnerable to secondary infections. In fact, it has been demonstrated that reduced systemic acetate levels can impair the function of alveolar macrophages, indicating that susceptibility to secondary pneumococcal infection may increase following influenza infection [57].
Another way by which the gut microbiota safeguards against respiratory viruses is by boosting type I IFNs. Under normal conditions, IFNs keep the body in a state of vigilance and control the production of various cytokines via feedforward and feedback mechanisms [72]. Microbial components, such as cell wall products or the release of RNA and DNA, can trigger the production of IFNs via intracellular Rig-I-like receptors, cyclic GMP-AMP synthase, and extracellular TLR [73,74,75]. Previous research has shown that animals treated with antibiotics experience impaired inflammasome activation and IFN responses, leading to a defective adaptive immune system and an inability to effectively clear viruses from the airways [76,77]. Several recent studies have conducted follow-up investigations on these reports. They have pinpointed mechanisms by which commensal bacteria act, either directly or via trans-kingdom interactions with other microorganisms like helminths [78], can sustain these systemic tonic interferon signals and safeguarding against pulmonary viral infections. Firstly, outer membrane products derived from Bacteroides, a genus that ranks among the most prevalent ones in the human gastrointestinal (GI) tract, can trigger the production of IFN-β by colonic dendritic cells (DCs) via TLR4, thereby safeguarding against systemic viral infections, such as influenza infection [79]. Secondly, the microbiota is capable of triggering tonic IFN production in plasmacytoid DCs (pDCs), releasing of single-stranded (ss)RNA, following a TLR7-MyD88-dependent mechanism [80,81]. Meanwhile, the production of IFN by pDCs maintains conventional DCs in a metabolic and transcriptomic state of readiness, enabling them to respond to stimuli and activate T cells [80]. Although the second mechanism has been demonstrated to limit the systemic spread of viruses, it still needs to be explored whether the interactions between microbiota and pDCs also have a role in the control of respiratory viral infections.
Only a small number of microbial metabolites or individual microbes have been identified as specific triggers tonic IFN production. Studies in gnotobiotic animal have indicated that colonization with Clostridium scindens—a bacterium expressing the enzyme 7α-dehydroxylation necessary for converting primary bile acids (BAs) into secondary BAs—or direct administration of the secondary BA deoxycholic acid (DCA) can protect antibiotic-treated or germ-free (GF) mice from systemic chikungunya virus infection. This protection occurs by restoring pDCs function [81]. While the precise mechanism by which secondary BAs safeguard against systemic viral infection remains mostly unknown, one proposed pathway involves virus-induced activation of NF-κB. This activation promotes TGR5-mediated cellular uptake and intracellular accumulation of secondary Bas.Subsequently, this accumulation induces the phosphorylation of key antiviral signaling components, facilitating viral clearance [82].
The role of tryptophan-derived metabolites in respiratory viral infections has been extensively explored. Desaminotyrosine(DAT), is produced by Clostridium orbiscindens during the conversion of dietary flavonoids and amino acid metabolism [83]. DAT can induce tonic IFN signaling and enhance macrophage phagocytic activity, thereby protecting against lethal influenza infection. In addition, viruses like SARS-CoV-2 are known to exploit the aryl hydrocarbon receptor (AhR) to evade immune activation, enabling viral replication, triggering mucus production, and dysregulating gas exchange [84,85]. On the contrary, a recent study demonstrated that selectively reducing AhR expression in lung endothelium protected against influenza-induced pathology, highlighting that the tissue-protective function of AhR plays a dual role at different stages of the disease [86]. These studies prove that microbial metabolites could regulate host immune to influence viral infections, and more metabolites are listed in Table 3.
Another mechanism through which the gut microbiota can systemically trigger the cGAS-STING-IFN pathway involves the release of membrane vesicles that deliver bacterial DNA to immune cells. The lipid encapsulation of microbial DNA facilitates its transport, enabling immune modulation at distant sites, providing protection against respiratory viral infections [96]. It has been demonstrated that a gain-of-function STING mutation model can cause autoimmune vasculopathy and fibrosing lung disease, suggesting that microbiota-driven tonic STING activation may also exacerbate autoimmunity [97].Furthermore, the gut microbiota regulates IFN production by lung epithelial cells, offering protection against influenza infection. Specifically, the Clostridium strain C. butyricum promotes the release of 18-hydroxyeicosapentaenoic acid (18-HEPE), which activates tonic IFN-λ production in lung epithelial cells through the G protein-coupled receptor GPR120, thereby enhancing resistance to influenza infection [95]. However, some contradictory studies have suggested that IFN-λ production may be detrimental during chronic lung viral infections, indicating that IFN-λ plays a dual role depending on the context [98]. Additionally, short-chain fatty acids (SCFAs) like acetate and butyrate can trigger an antiviral immune defense against respiratory syncytial virus (RSV) by stimulating airway epithelial cells to produce IFNs [87,88]. Collectively, these recent studies highlight multiple mechanisms by which gut microbial factors sustain systemic tonic IFN production. Moreover, they demonstrate how these factors regulate the metabolic functions of both innate and adaptive immune cells within the lung, thereby providing protection against respiratory viral infections.
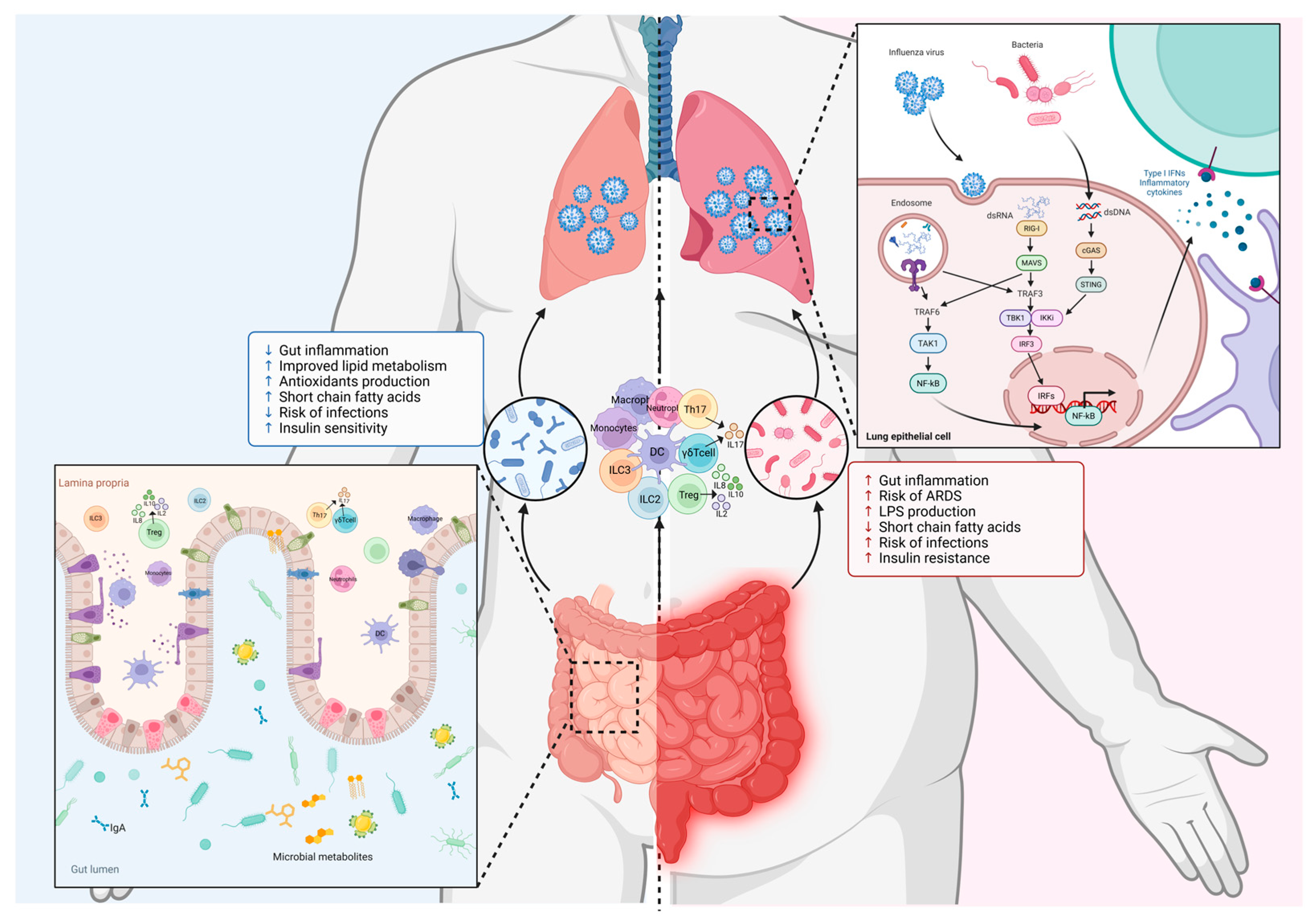
Figure 2.
Mechanisms of Gut-Lung axis in influenza virus infection. The dsDNA from gut microbiota activates the cGAS-STING signaling pathway, while dsRNA from influenza virus activates the RIG-I-MAVS signaling pathway in respiratory epithelial cells. The dsDNA and dsRNA can phosphorylate and activate TBK1 and its downstream IRF3 transcription factor. The activation of IRF3 dimerizes and enters the nucleus to initiate the transcription of type I interferon genes. Gut microbiota produced metabolites that activate the immune system. In addition, Tregs, Th17 cells, ILC2s, ILC3s, and γδ T cells migrate from the gut to the lungs to impact respiratory immunity. The disorder of intestinal flora in obese individuals is more likely to cause intestinal inflammation and increase the severity of influenza virus infection.
Figure 2.
Mechanisms of Gut-Lung axis in influenza virus infection. The dsDNA from gut microbiota activates the cGAS-STING signaling pathway, while dsRNA from influenza virus activates the RIG-I-MAVS signaling pathway in respiratory epithelial cells. The dsDNA and dsRNA can phosphorylate and activate TBK1 and its downstream IRF3 transcription factor. The activation of IRF3 dimerizes and enters the nucleus to initiate the transcription of type I interferon genes. Gut microbiota produced metabolites that activate the immune system. In addition, Tregs, Th17 cells, ILC2s, ILC3s, and γδ T cells migrate from the gut to the lungs to impact respiratory immunity. The disorder of intestinal flora in obese individuals is more likely to cause intestinal inflammation and increase the severity of influenza virus infection.
6. Targeting the Gut Microbiota to Treat Influenza
Influenza A virus results in 3 to 5 million severe cases and 250,000 to 500,000 deaths annually. Vaccination remains the primary preventive strategy, but its effectiveness is limited by Influenza A virus high susceptibility to antigenic drift and delays in vaccine production [99]. Moreover, a great deal of information has been reported regarding the resistance to antiviral drugs such as Oseltamivir and Baloxavir, which are used to treat viral infections [100]. Consequently, these limitations justify exploring alternative influenza therapies. Recent research has discovered that modulating the gut microbiota can enhance the immune responses against IAV through the gut-lung axis, suggesting the therapeutic potential of the lung microbiome should be taken into consideration as well [101]. Critical unanswered questions include whether endogenous factors—particularly those derived from the gut microbiome—can regulate its composition and function [102]. Conducting in-depth research on the transient characteristics of the lung microbiome, which probably varies with aging and upon exposure to environmental antigens, might enable us to pinpoint potential lung-resident microbes or components that possess therapeutic potential.
Figure 3.
The novel therapeutic targeting the gut microbiota to treat influenza.
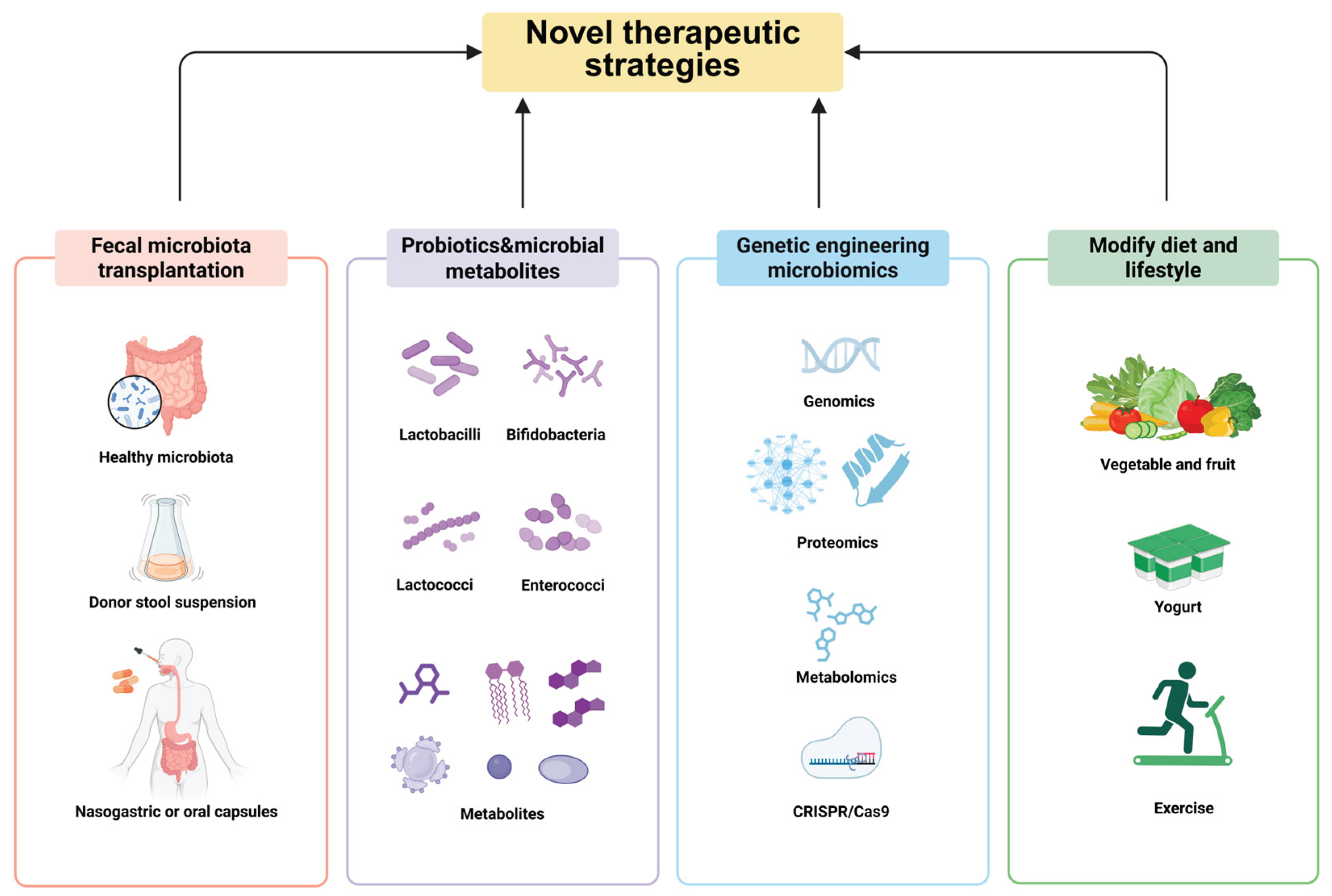
6.1. Fecal Microbiota Transplantation (FMT)
Fecal microbiota transplantation (FMT) is a method that involves transplanting the entire intestinal microbiota of healthy individuals into other patients. As the most direct strategy for regulating the intestinal microbiome at present, FMT can effectively alter the composition of the intestinal microbiota. Multiple studies have confirmed that FMT has beneficial effects on many diseases, such as intestinal graft-versus-host disease, irritable bowel syndrome, inflammatory bowel disease, and multidrug-resistant bacterial infections [103,104,105]. In addition, FMT for the treatment of recurrent Clostridioides difficile infection has been included in the clinical guidelines in the United States [106]. Liu et al. conducted oral FMT treatment on 11 discharged patients with COVID-19 to analyze the potential impact of FMT on the intestinal microbiota and immune system after SARS-CoV-2 infection. The results showed that all five patients with gastrointestinal symptoms improved after treatment. FMT corrected intestinal microbiota dysbiosis by increasing the relative abundance of Actinobacteria at the phylum level, reducing the relative abundance of Proteobacteria, and increasing the relative abundance of Bifidobacterium and Faecalibacterium at the genus level [107]. A case report described two patients with risk factors for severe COVID-19 (Patient 1: an 80-year-old male with pneumonia and sepsis; Patient 2: a 19-year-old male with ulcerative colitis receiving immunosuppressive therapy) who underwent FMT for Clostridioides difficile infection and were later diagnosed with COVID-19. Notably, both patients had mild clinical symptoms and did not develop severe COVID-19 but recovered rapidly, suggesting FMT can microbiota homeostasis and promote immune recovery [108]. Together, these studies provide preliminary clues for the application of FMT in the treatment of viral pneumonia, but more research is needed for clinical application.
6.2. Targeted Therapy of Probiotics and Microbial Metabolites
Recent studies suggest that probiotics can confer health benefits to the host by enhancing intestinal barrier function, modulating the interaction between the host and microbiota, producing antimicrobial substances and organic acids, resisting pathogens, regulating immune responses, and influencing metabolic processes [109]. Therefore, probiotics can serve as an adjuvant therapy for various diseases, and there are well-established guidelines for their application in pediatric populations [110]. As previously discussed, the gut microbiota in individuals with COVID-19 undergoes significant dysbiosis, much of which is detrimental to host health. Supplementing probiotics to restore intestinal microecological balance shows promise as an adjunctive therapy for COVID-19, potentially enhancing immune function and promoting recovery from infection [111]. A bibliometric analysis of 84 studies on microecological interventions for COVID-19 revealed that probiotics or prebiotics could reinforce the mucosal barrier, regulate the host's immune system, and enhance the interaction between the gut-lung axis, thereby increasing resistance to SARS-CoV-2 infection and reducing disease duration and severity [112]. Clinically, probiotics have been shown to alleviate symptoms associated with COVID-19, such as diarrhea, fatigue, and fever, while also reducing the need for intensive care [113,114]. Furthermore, microbiota influence disease progression via bioactive metabolites including SCFAs, aromatic compounds, amino acids, bile acids, vitamins, and lipids [115,116]. T The human microbiota influences SARS-CoV-2 infection both directly and indirectly through its complex metabolome. Notably, numerous studies have reported a reduced abundance of SCFA-producing bacteria in patients with COVID-19 [117,118]. Zhang et al. identified reduced capacity for SCFA and L-isoleucine biosynthesis in the gut microbiome of COVID-19 patients, alongside increased urea synthesis. This suggests SCFAs play multifaceted roles, including anti-inflammatory effects, immune modulation, and metabolic regulation [119]. In addition to SCFAs, three intestinal microbiota-derived metabolites—tryptamine, 2,5-bis(3-indolylmethyl)pyrazine, and N6-(D2-isopentenyl)adenosine—have been identified. These metabolites function as 5-hydroxytryptamine receptor agonists, demonstrate anti-SARS-CoV-2 activity, and show structural and functional parallels with clinically used antiviral drugs, suggesting targeting probiotics and microbial metabolites may aid in treating viral pneumonia [120].
6.3. Transgenic Microbial Therapy
In the past several years, the Food and Drug Administration (FDA) has given approval to a significant number of biologic drugs, including monoclonal antibodies, therapeutic proteins, and vaccines. It has been demonstrated that the large interindividual variation in the composition of the lung and gut microbiomes may reflect functional redundancy. Specifically, metagenomic studies have further emphasized that the host exerts selection pressure based on metabolic functional traits rather than specific taxa, suggesting that future therapeutic strategies should focus on targeting the metabolic output of bacteria rather than their taxonomic classification [121]. Most of these products utilized in the treatment of various human diseases are genetically modified and produced through biological pathways. However, the high cost of production, particularly during downstream processing, the short half-life of the drugs, and the limitations in drug delivery collectively hinder their therapeutic applications [122,123]. Thus, engineered microbiomics serves as a promising alternative with the potential to secrete therapeutic proteins, deliver antigens, monitor the gut environment, eradicate pathogens, modulate the immune system, and metabolize harmful substances [124,125]. It has been reported that the development of a genome-scale library for probiotics using CRISPR can facilitate mapping genotype-phenotype relationships [126]. For instance, Crook et al. found that glycoside hydrolases enhanced cell viability in the gut [127]. Recently, the FDA's approval of RBX2660 for treating Clostridioides difficile infection has paved the way for the market of microbiome-based products. These cases suggest that advances in bacterial genetic engineering technology have made it feasible to modify the intestinal microbiome to enhance resistance to influenza infection. In addition, genetically engineered microorganisms can achieve a higher level of specificity when compared to FMT. In the coming years, many startups, well-established biotechnology companies, and research institutions are expected to make substantial investments in the research and development of live biotherapeutic products for treating human diseases, thereby enriching their product pipelines with bacterial therapies.
6.4. Modify Diet and Lifestyle
Dietary intervention can regulate gut microbiota and enhance the efficacy of immunotherapy with minimal harmful risks, suggesting that it may represent a more promising approach. Many diets that have been proven to be beneficial for human health typically contain high levels of dietary fiber. Studies have shown that short-term consumption of diets consisting exclusively of plant products can alter microbial community structure and interindividual differences in microbial gene expression [128]. However, randomized controlled trials of specific foods or nutrients have shown limited long-term effects of these interventions on microbiota and immune function [129]. Gouez et al. have confirmed that severe malnutrition was strongly associated with lower survival [130]. Moreover, Gao et al. found that Resveratrol (RESV) can up-regulate the cytotoxic effect of CD8+ T cells and improve the tumor immunosuppressive microenvironment [131]. In daily life, we can also supplement probiotics by drinking yogurt, which helps to regulate the balance of intestinal flora, promote the digestion and absorption of food, and thus maintain the normal physiological function of the intestine [132]. More importantly, we should also focus on strengthening the positive impact of exercise on physical health. Continuous and moderate exercise can effectively enhance the body's resistance to external bacterial invasions, thereby maintaining a good state of health. At present, dietary adjuvant therapy is still in its infancy, and more experiments are needed to explore the impact of dietary habits and lifestyle on the gut microbiome.
7. Conclusions and Future Perspectives
In recent years, the relationship between gut microbiota and immunotherapy has become a hot topic in immunology research. Obesity weakens anti-influenza immunity through multiple mechanisms, and the regulation of the microbiome provides a new target for reversing immune suppression. The composition and function of the gut microbiota are significantly altered in influenza patients. As an important pathway connecting the gut and lung, the regulation of gut-lung axis has a profound impact on the immune microenvironment of the lung. Future research is required to better understand how the composition of the gut microbiome affects the response to immunotherapy and drug resistance, as well as to investigate strategies for enhancing immunotherapy efficacy by restoring microbiome balance, such as through probiotics and FMT. In particular, large sample and multi-center clinical trials are needed to verify the effectiveness and safety of microbiome intervention and provide reliable basis for clinical practice. Moreover, with advancements in medicine, it is anticipated that the potential of gut microbiota in influenza immunotherapy will be fully harnessed, leading to improved therapeutic outcomes for patients. Taken together, it is necessary to combine basic research and clinical translation to develop comprehensive treatment strategies based on the microbiota-metabolic-immune axis to reduce the burden of influenza in obese people.
Author Contributions
Conceptualization, X.J. and J.S.; formal analysis, X.J.; resources, J.S.; writing—original draft preparation, X.J.; writing—review and editing, J.S.; visualization, X.J.; supervision, J.S.. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), China Postdoctoral Science Foundation (2015M581873), Natural Science Foundation of Jiangsu Province (BK20150450), the Scientific Research Foundation for the Returned Overseas Chinese Scholars, State Education Ministry (2015311) to J.S. and the postgraduate Research & Practice Innovation Program of Yangzhou University (XKYCX19132) to X. J..
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
During the preparation of this manuscript, the authors used BioRender. We thank BioRender (biorender.com) for providing items for drawing schematic model.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Centers for Disease, C. and Prevention, Estimates of deaths associated with seasonal influenza --- United States, 1976-2007. MMWR Morb Mortal Wkly Rep, 2010. 59(33): p. 1057-62.
- Shrestha, S.S.; Swerdlow, D.L.; Borse, R.H.; Prabhu, V.S.; Finelli, L.; Atkins, C.Y.; Owusu-Edusei, K.; Bell, B.; Mead, P.S.; Biggerstaff, M.; et al. Estimating the Burden of 2009 Pandemic Influenza A (H1N1) in the United States (April 2009-April 2010). Clin. Infect. Dis. 2010, 52, S75–S82. [CrossRef]
- Tumpey, T.M.; Basler, C.F.; Aguilar, P.V.; Zeng, H.; SolórZano, A.; Swayne, D.E.; Cox, N.J.; Katz, J.M.; Taubenberger, J.K.; Palese, P.; et al. Characterization of the Reconstructed 1918 Spanish Influenza Pandemic Virus. Science 2005, 310, 77–80. [CrossRef]
- Xu, R.; McBride, R.; Paulson, J.C.; Basler, C.F.; Wilson, I.A. Structure, Receptor Binding, and Antigenicity of Influenza Virus Hemagglutinins from the 1957 H2N2 Pandemic. J. Virol. 2010, 84, 1715–1721. [CrossRef]
- West, J., et al., Characterization of changes in the hemagglutinin that accompanied the emergence of H3N2/1968 pandemic influenza viruses. PLoS Pathog, 2021. 17(9): p. e1009566.
- Taubenberger, J.K. and D.M. Morens, 1918 Influenza: the mother of all pandemics. Emerg Infect Dis, 2006. 12(1): p. 15-22.
- Janssen, I., P.T. Katzmarzyk, and R. Ross, Body mass index, waist circumference, and health risk: evidence in support of current National Institutes of Health guidelines. Arch Intern Med, 2002. 162(18): p. 2074-9.
- Alexopoulos, S.J.; Chen, S.-Y.; Brandon, A.E.; Salamoun, J.M.; Byrne, F.L.; Garcia, C.J.; Beretta, M.; Olzomer, E.M.; Shah, D.P.; Philp, A.M.; et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nat. Commun. 2020, 11, 1–13. [CrossRef]
- Yu, Z.; Zhao, Z.; Ding, C.; Liu, Q.; Ma, T.; Han, X.; Lu, D.; Zhang, L. The association between sarcopenia and cardiovascular disease: An investigative analysis from the NHANES. Nutr. Metab. Cardiovasc. Dis. 2025, 35, 103864. [CrossRef]
- Caballero, B., Humans against Obesity: Who Will Win? Adv Nutr, 2019. 10(suppl_1): p. S4-S9.
- Hornung, F.; Schulz, L.; Köse-Vogel, N.; Häder, A.; Grießhammer, J.; Wittschieber, D.; Autsch, A.; Ehrhardt, C.; Mall, G.; Löffler, B.; et al. Thoracic adipose tissue contributes to severe virus infection of the lung. Int. J. Obes. 2023, 47, 1088–1099. [CrossRef]
- Louie, J.K.; Acosta, M.; Samuel, M.C.; Schechter, R.; Vugia, D.J.; Harriman, K.; Matyas, B.T. A Novel Risk Factor for a Novel Virus: Obesity and 2009 Pandemic Influenza A (H1N1). Clin. Infect. Dis. 2011, 52, 301–312. [CrossRef]
- Mafort, T.T., et al., Obesity: systemic and pulmonary complications, biochemical abnormalities, and impairment of lung function. Multidiscip Respir Med, 2016. 11: p. 28.
- Falagas, M.E.; Athanasoulia, A.P.; Peppas, G.; Karageorgopoulos, D.E. Effect of body mass index on the outcome of infections: a systematic review. Obes. Rev. 2009, 10, 280–289. [CrossRef]
- Yan, J.; Grantham, M.; Pantelic, J.; de Mesquita, P.J.B.; Albert, B.; Liu, F.; Ehrman, S.; Milton, D.K. Infectious virus in exhaled breath of symptomatic seasonal influenza cases from a college community. Proc. Natl. Acad. Sci. 2018, 115, 1081–1086. [CrossRef]
- Elliot, J.G.; Donovan, G.M.; Wang, K.C.; Green, F.H.; James, A.L.; Noble, P.B. Fatty airways: implications for obstructive disease. Eur. Respir. J. 2019, 54, 1900857. [CrossRef]
- Kwong, J.C.; Campitelli, M.A.; Rosella, L.C. Obesity and Respiratory Hospitalizations During Influenza Seasons in Ontario, Canada: A Cohort Study. Clin. Infect. Dis. 2011, 53, 413–421. [CrossRef]
- E Maier, H.; Lopez, R.; Sanchez, N.; Ng, S.; Gresh, L.; Ojeda, S.; Burger-Calderon, R.; Kuan, G.; Harris, E.; Balmaseda, A.; et al. Obesity Increases the Duration of Influenza A Virus Shedding in Adults. J. Infect. Dis. 2018, 218, 1378–1382. [CrossRef]
- Fezeu, L.; Julia, C.; Henegar, A.; Bitu, J.; Hu, F.B.; Grobbee, D.E.; Kengne, A.-P.; Hercberg, S.; Czernichow, S. Obesity is associated with higher risk of intensive care unit admission and death in influenza A (H1N1) patients: a systematic review and meta-analysis. Obes. Rev. 2011, 12, 653–659. [CrossRef]
- Díaz, E.; Rodríguez, A.; Martin-Loeches, I.; Lorente, L.; Del Mar Martín, M.; Pozo, J.C.; Montejo, J.C.; Estella, A.; Arenzana, .; Rello, J. Impact of Obesity in Patients Infected With 2009 Influenza A(H1N1). Chest 2011, 139, 382–386. [CrossRef]
- Kok, J.; Blyth, C.C.; Foo, H.; Bailey, M.J.; Pilcher, D.V.; Webb, S.A.; Seppelt, I.M.; Dwyer, D.E.; Iredell, J.R.; Semple, M.G. Viral Pneumonitis Is Increased in Obese Patients during the First Wave of Pandemic A(H1N1) 2009 Virus. PLOS ONE 2013, 8, e55631. [CrossRef]
- Vaillant, L.; La Ruche, G.; Tarantola, A.; Barboza, P.; for the epidemic intelligence team at InVS Epidemiology of fatal cases associated with pandemic H1N1 influenza 2009. Eurosurveillance 2009, 14, 19309. [CrossRef]
- Investigators, A.I., et al., Critical care services and 2009 H1N1 influenza in Australia and New Zealand. N Engl J Med, 2009. 361(20): p. 1925-34.
- Andrew, M.K.; Pott, H.; Staadegaard, L.; Paget, J.; Chaves, S.S.; Ortiz, J.R.; McCauley, J.; Bresee, J.; Nunes, M.C.; Baumeister, E.; et al. Age Differences in Comorbidities, Presenting Symptoms, and Outcomes of Influenza Illness Requiring Hospitalization: A Worldwide Perspective From the Global Influenza Hospital Surveillance Network. Open Forum Infect. Dis. 2023, 10, ofad244. [CrossRef]
- E Cohen, L.; Hansen, C.L.; Andrew, M.K.; A McNeil, S.; Vanhems, P.; Kyncl, J.; Domingo, J.D.; Zhang, T.; Dbaibo, G.; Laguna-Torres, V.A.; et al. Predictors of Severity of Influenza-Related Hospitalizations: Results From the Global Influenza Hospital Surveillance Network (GIHSN). J. Infect. Dis. 2024, 229, 999–1009. [CrossRef]
- Lau, D.; Tobin, S.; Pribiag, H.; Nakajima, S.; Fisette, A.; Matthys, D.; Flores, A.K.F.; Peyot, M.-L.; Madiraju, S.R.M.; Prentki, M.; et al. ABHD6 loss-of-function in mesoaccumbens postsynaptic but not presynaptic neurons prevents diet-induced obesity in male mice. Nat. Commun. 2024, 15, 1–22. [CrossRef]
- Guan, D.; Men, Y.; Bartlett, A.; Hernández, M.A.S.; Xu, J.; Yi, X.; Li, H.-S.; Kong, D.; Mazitschek, R.; Ozcan, U. Central inhibition of HDAC6 re-sensitizes leptin signaling during obesity to induce profound weight loss. Cell Metab. 2024, 36, 857–876.e10. [CrossRef]
- Liang, Y.; Luo, C.; Sun, L.; Feng, T.; Yin, W.; Zhang, Y.; Mulholland, M.W.; Zhang, W.; Yin, Y. Reduction of specific enterocytes from loss of intestinal LGR4 improves lipid metabolism in mice. Nat. Commun. 2024, 15, 1–12. [CrossRef]
- Sass, F.; Ma, T.; Ekberg, J.H.; Kirigiti, M.; Ureña, M.G.; Dollet, L.; Brown, J.M.; Basse, A.L.; Yacawych, W.T.; Burm, H.B.; et al. NK2R control of energy expenditure and feeding to treat metabolic diseases. Nature 2024, 635, 987–1000. [CrossRef]
- Petersen, J., et al., GLP-1-directed NMDA receptor antagonism for obesity treatment. Nature, 2024. 629(8014): p. 1133-1141.
- Fadahunsi, N.; Petersen, J.; Metz, S.; Jakobsen, A.; Mathiesen, C.V.; Buch-Rasmussen, A.S.; Kurgan, N.; Larsen, J.K.; Andersen, R.C.; Topilko, T.; et al. Targeting postsynaptic glutamate receptor scaffolding proteins PSD-95 and PICK1 for obesity treatment. Sci. Adv. 2024, 10, eadg2636. [CrossRef]
- Wei, W.; Lyu, X.; Markhard, A.L.; Fu, S.; Mardjuki, R.E.; Cavanagh, P.E.; Zeng, X.; Rajniak, J.; Lu, N.; Xiao, S.; et al. PTER is a N-acetyltaurine hydrolase that regulates feeding and obesity. Nature 2024, 633, 182–188. [CrossRef]
- Zhang, J.; Huang, Y.; Li, H.; Xu, P.; Liu, Q.; Sun, Y.; Zhang, Z.; Wu, T.; Tang, Q.; Jia, Q.; et al. B3galt5 functions as a PXR target gene and regulates obesity and insulin resistance by maintaining intestinal integrity. Nat. Commun. 2024, 15, 1–16. [CrossRef]
- Zhang, J.; Wu, Y.; Wang, Y.; Liu, P.; Liu, K.; Sun, J.; Zhang, P.; Wang, X.; Liu, X.; Xu, X. Influenza A virus infection activates STAT3 to enhance SREBP2 expression, cholesterol biosynthesis, and virus replication. iScience 2024, 27, 110424. [CrossRef]
- Xing, M.; Li, Y.; Zhang, Y.; Zhou, J.; Ma, D.; Zhang, M.; Tang, M.; Ouyang, T.; Zhang, F.; Shi, X.; et al. Paraventricular hypothalamic RUVBL2 neurons suppress appetite by enhancing excitatory synaptic transmission in distinct neurocircuits. Nat. Commun. 2024, 15, 1–21. [CrossRef]
- Thingholm, L.B., et al., Obese Individuals with and without Type 2 Diabetes Show Different Gut Microbial Functional Capacity and Composition. Cell Host Microbe, 2019. 26(2): p. 252-264 e10.
- Menni, C.; A Jackson, M.; Pallister, T.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Gut microbiome diversity and high-fibre intake are related to lower long-term weight gain. Int. J. Obes. 2017, 41, 1099–1105. [CrossRef]
- Xu, Z.; Jiang, W.; Huang, W.; Lin, Y.; Chan, F.K.; Ng, S.C. Gut microbiota in patients with obesity and metabolic disorders — a systematic review. Genes Nutr. 2022, 17, 1–18. [CrossRef]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison of the gut microbiota composition between obese and non-obese individuals in a Japanese population, as analyzed by terminal restriction fragment length polymorphism and next-generation sequencing. BMC Gastroenterol. 2015, 15, 1–10. [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [CrossRef]
- Cox, A.J., N.P. West, and A.W. Cripps, Obesity, inflammation, and the gut microbiota. Lancet Diabetes Endocrinol, 2015. 3(3): p. 207-15.
- Jumpertz, R.; Le, D.S.; Turnbaugh, P.J.; Trinidad, C.; Bogardus, C.; Gordon, J.I.; Krakoff, J. Energy-Balance Studies Reveal Associations between Gut Microbes, Caloric Load, and Nutrient Absorption in Humans. Am. J. Clin. Nutr. 2011, 94, 58–65. [CrossRef]
- Kim, B.; Choi, H.-N.; Yim, J.-E. Effect of Diet on the Gut Microbiota Associated with Obesity. J. Obes. Metab. Syndr. 2019, 28, 216–224. [CrossRef]
- Huang, Y.; Mao, K.; Chen, X.; Sun, M.-A.; Kawabe, T.; Li, W.; Usher, N.; Zhu, J.; Urban, J.F., Jr.; Paul, W.E.; et al. S1P-dependent interorgan trafficking of group 2 innate lymphoid cells supports host defense. Science 2018, 359, 114–119. [CrossRef]
- Olszak, T.; An, D.; Zeissig, S.; Vera, M.P.; Richter, J.; Franke, A.; Glickman, J.N.; Siebert, R.; Baron, R.M.; Kasper, D.L.; et al. Microbial Exposure During Early Life Has Persistent Effects on Natural Killer T Cell Function. Science 2012, 336, 489–493. [CrossRef]
- Alon, R.; Sportiello, M.; Kozlovski, S.; Kumar, A.; Reilly, E.C.; Zarbock, A.; Garbi, N.; Topham, D.J. Leukocyte trafficking to the lungs and beyond: lessons from influenza for COVID-19. Nat. Rev. Immunol. 2020, 21, 49–64. [CrossRef]
- Pick, R.; He, W.; Chen, C.-S.; Scheiermann, C. Time-of-Day-Dependent Trafficking and Function of Leukocyte Subsets. Trends Immunol. 2019, 40, 524–537. [CrossRef]
- Zundler, S.; Günther, C.; Kremer, A.E.; Zaiss, M.M.; Rothhammer, V.; Neurath, M.F. Gut immune cell trafficking: inter-organ communication and immune-mediated inflammation. Nat. Rev. Gastroenterol. Hepatol. 2022, 20, 50–64. [CrossRef]
- Uchimura, Y.; Fuhrer, T.; Li, H.; Lawson, M.A.; Zimmermann, M.; Yilmaz, B.; Zindel, J.; Ronchi, F.; Sorribas, M.; Hapfelmeier, S.; et al. Antibodies Set Boundaries Limiting Microbial Metabolite Penetration and the Resultant Mammalian Host Response. Immunity 2018, 49, 545–559.e5. [CrossRef]
- Trompette, A.; Pernot, J.; Perdijk, O.; Alqahtani, R.A.A.; Domingo, J.S.; Camacho-Muñoz, D.; Wong, N.C.; Kendall, A.C.; Wiederkehr, A.; Nicod, L.P.; et al. Gut-derived short-chain fatty acids modulate skin barrier integrity by promoting keratinocyte metabolism and differentiation. Mucosal Immunol. 2022, 15, 908–926. [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [CrossRef]
- Clarke, T.B.; Davis, K.M.; Lysenko, E.S.; Zhou, A.Y.; Yu, Y.; Weiser, J.N. Recognition of peptidoglycan from the microbiota by Nod1 enhances systemic innate immunity. Nat. Med. 2010, 16, 228–231. [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Pattaroni, C.; Lopez-Mejia, I.C.; Riva, E.; Pernot, J.; Ubags, N.; Fajas, L.; Nicod, L.P.; Marsland, B.J. Dietary Fiber Confers Protection against Flu by Shaping Ly6c− Patrolling Monocyte Hematopoiesis and CD8+ T Cell Metabolism. Immunity 2018, 48, 992–1005.e8. [CrossRef]
- A Hill, D.; Siracusa, M.C.; Abt, M.C.; Kim, B.S.; Kobuley, D.; Kubo, M.; Kambayashi, T.; LaRosa, D.F.; Renner, E.D.; Orange, J.S.; et al. Commensal bacteria–derived signals regulate basophil hematopoiesis and allergic inflammation. Nat. Med. 2012, 18, 538–546. [CrossRef]
- Maurice, C.F.; Haiser, H.J.; Turnbaugh, P.J. Xenobiotics Shape the Physiology and Gene Expression of the Active Human Gut Microbiome. Cell 2013, 152, 39–50. [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.Y.; Yeoh, Y.K.; Li, A.Y.L.; Zhan, H.; Wan, Y.; Chung, A.C.K.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients With COVID-19 During Time of Hospitalization. Gastroenterology 2020, 159, 944–955.e948. [CrossRef]
- Sencio, V.; Barthelemy, A.; Tavares, L.P.; Machado, M.G.; Soulard, D.; Cuinat, C.; Queiroz-Junior, C.M.; Noordine, M.-L.; Salomé-Desnoulez, S.; Deryuter, L.; et al. Gut Dysbiosis during Influenza Contributes to Pulmonary Pneumococcal Superinfection through Altered Short-Chain Fatty Acid Production. Cell Rep. 2020, 30, 2934–2947.e6. [CrossRef]
- Wang, J., et al., Respiratory influenza virus infection induces intestinal immune injury via microbiota-mediated Th17 cell-dependent inflammation. J Exp Med, 2014. 211(12): p. 2397-410.
- Laidlaw, B.J.; Zhang, N.; Marshall, H.D.; Staron, M.M.; Guan, T.; Hu, Y.; Cauley, L.S.; Craft, J.; Kaech, S.M. CD4+ T Cell Help Guides Formation of CD103+ Lung-Resident Memory CD8+ T Cells during Influenza Viral Infection. Immunity 2014, 41, 633–645. [CrossRef]
- Mann, E.R.; Lam, Y.K.; Uhlig, H.H. Short-chain fatty acids: linking diet, the microbiome and immunity. Nat. Rev. Immunol. 2024, 24, 577–595. [CrossRef]
- Moriyama, M.; Ichinohe, T. High ambient temperature dampens adaptive immune responses to influenza A virus infection. Proc. Natl. Acad. Sci. 2019, 116, 3118–3125. [CrossRef]
- Bachem, A.; Makhlouf, C.; Binger, K.J.; de Souza, D.P.; Tull, D.; Hochheiser, K.; Whitney, P.G.; Fernandez-Ruiz, D.; Dähling, S.; Kastenmüller, W.; et al. Microbiota-Derived Short-Chain Fatty Acids Promote the Memory Potential of Antigen-Activated CD8+ T Cells. Immunity 2019, 51, 285–297 e285. [CrossRef]
- Alarcon, P.C.; Ulanowicz, C.J.; A Damen, M.S.M.; Eom, J.; Sawada, K.; Chung, H.; Alahakoon, T.; Oates, J.R.; Wayland, J.L.; E Stankiewicz, T.; et al. Obesity Uncovers the Presence of Inflammatory Lung Macrophage Subsets With an Adipose Tissue Transcriptomic Signature in Influenza Virus Infection. J. Infect. Dis. 2024, 231, e317–e327. [CrossRef]
- Castro, I.A.; Jorge, D.M.M.; Ferreri, L.M.; Martins, R.B.; Pontelli, M.C.; Jesus, B.L.S.; Cardoso, R.S.; Criado, M.F.; Carenzi, L.; Valera, F.C.P.; et al. Silent Infection of B and CD8 + T Lymphocytes by Influenza A Virus in Children with Tonsillar Hypertrophy. J. Virol. 2020, 94. [CrossRef]
- Hensen, L.; Illing, P.T.; Clemens, E.B.; Nguyen, T.H.O.; Koutsakos, M.; van de Sandt, C.E.; Mifsud, N.A.; Nguyen, A.T.; Szeto, C.; Chua, B.Y.; et al. CD8+ T cell landscape in Indigenous and non-Indigenous people restricted by influenza mortality-associated HLA-A*24:02 allomorph. Nat. Commun. 2021, 12, 1–20. [CrossRef]
- Koutsakos, M., et al., Human CD8(+) T cell cross-reactivity across influenza A, B and C viruses. Nat Immunol, 2019. 20(5): p. 613-625.
- van de Wall, S., et al., Dynamic landscapes and protective immunity coordinated by influenza-specific lung-resident memory CD8(+) T cells revealed by intravital imaging. Immunity, 2024. 57(8): p. 1878-1892 e5.
- Schmidt, A.; Fuchs, J.; Dedden, M.; Kocher, K.; Schülein, C.; Hübner, J.; Antão, A.V.; Irrgang, P.; Oltmanns, F.; Viherlehto, V.; et al. Inflammatory conditions shape phenotypic and functional characteristics of lung-resident memory T cells in mice. Nat. Commun. 2025, 16, 1–18. [CrossRef]
- Pritzl, C.J.; Luera, D.; Knudson, K.M.; Quaney, M.J.; Calcutt, M.J.; Daniels, M.A.; Teixeiro, E. IKK2/NFkB signaling controls lung resident CD8+ T cell memory during influenza infection. Nat. Commun. 2023, 14, 1–14. [CrossRef]
- Lyu, Z.; Yuan, G.; Zhang, Y.; Zhang, F.; Liu, Y.; Li, Y.; Li, G.; Wang, Y.; Zhang, M.; Hu, Y.; et al. Anaerostipes caccae CML199 enhances bone development and counteracts aging-induced bone loss through the butyrate-driven gut–bone axis: the chicken model. Microbiome 2024, 12, 1–24. [CrossRef]
- Dang, A.T.; Begka, C.; Pattaroni, C.; Caley, L.R.; Floto, R.A.; Peckham, D.G.; Marsland, B.J. Butyrate regulates neutrophil homeostasis and impairs early antimicrobial activity in the lung. Mucosal Immunol. 2023, 16, 476–485. [CrossRef]
- Gough, D.J.; Messina, N.L.; Clarke, C.J.; Johnstone, R.W.; Levy, D.E. Constitutive Type I Interferon Modulates Homeostatic Balance through Tonic Signaling. Immunity 2012, 36, 166–174. [CrossRef]
- Sun, L.; Wu, J.; Du, F.; Chen, X.; Chen, Z.J. Cyclic GMP-AMP Synthase Is a Cytosolic DNA Sensor That Activates the Type I Interferon Pathway. Science 2013, 339, 786–791. [CrossRef]
- Loo, Y.-M.; Gale, M., Jr. Immune Signaling by RIG-I-like Receptors. Immunity 2011, 34, 680–692. [CrossRef]
- Kawai, T.; Akira, S. Toll-like Receptors and Their Crosstalk with Other Innate Receptors in Infection and Immunity. Immunity 2011, 34, 637–650. [CrossRef]
- Abt, M.C.; Osborne, L.C.; Monticelli, L.A.; Doering, T.A.; Alenghat, T.; Sonnenberg, G.F.; Paley, M.A.; Antenus, M.; Williams, K.L.; Erikson, J.; et al. Commensal Bacteria Calibrate the Activation Threshold of Innate Antiviral Immunity. Immunity 2012, 37, 158–170. [CrossRef]
- Ichinohe, T.; Pang, I.K.; Kumamoto, Y.; Peaper, D.R.; Ho, J.H.; Murray, T.S.; Iwasaki, A. Microbiota regulates immune defense against respiratory tract influenza A virus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 5354–5359. [CrossRef]
- McFarlane, A.J.; McSorley, H.J.; Davidson, D.J.; Fitch, P.M.; Errington, C.; Mackenzie, K.J.; Gollwitzer, E.S.; Johnston, C.J.; MacDonald, A.S.; Edwards, M.R.; et al. Enteric helminth-induced type I interferon signaling protects against pulmonary virus infection through interaction with the microbiota. J. Allergy Clin. Immunol. 2017, 140, 1068–1078.e6. [CrossRef]
- Stefan, K.L.; Kim, M.V.; Iwasaki, A.; Kasper, D.L. Commensal Microbiota Modulation of Natural Resistance to Virus Infection. Cell 2020, 183, 1312–1324.e10. [CrossRef]
- Schaupp, L.; Muth, S.; Rogell, L.; Kofoed-Branzk, M.; Melchior, F.; Lienenklaus, S.; Ganal-Vonarburg, S.C.; Klein, M.; Guendel, F.; Hain, T.; et al. Microbiota-Induced Type I Interferons Instruct a Poised Basal State of Dendritic Cells. Cell 2020, 181, 1080–1096.e19. [CrossRef]
- Winkler, E.S.; Shrihari, S.; Hykes, B.L.; Handley, S.A.; Andhey, P.S.; Huang, Y.-J.S.; Swain, A.; Droit, L.; Chebrolu, K.K.; Mack, M.; et al. The Intestinal Microbiome Restricts Alphavirus Infection and Dissemination through a Bile Acid-Type I IFN Signaling Axis. Cell 2020, 182, 901–918.e18. [CrossRef]
- Hu, M.-M.; He, W.-R.; Gao, P.; Yang, Q.; He, K.; Cao, L.-B.; Li, S.; Feng, Y.-Q.; Shu, H.-B. Virus-induced accumulation of intracellular bile acids activates the TGR5-β-arrestin-SRC axis to enable innate antiviral immunity. Cell Res. 2019, 29, 193–205. [CrossRef]
- Steed, A.L.; Christophi, G.P.; Kaiko, G.E.; Sun, L.; Goodwin, V.M.; Jain, U.; Esaulova, E.; Artyomov, M.N.; Morales, D.J.; Holtzman, M.J.; et al. The microbial metabolite desaminotyrosine protects from influenza through type I interferon. Science 2017, 357, 498–502. [CrossRef]
- Giovannoni, F.; Li, Z.; Remes-Lenicov, F.; Dávola, M.E.; Elizalde, M.; Paletta, A.; Ashkar, A.A.; Mossman, K.L.; Dugour, A.V.; Figueroa, J.M.; et al. AHR signaling is induced by infection with coronaviruses. Nat. Commun. 2021, 12, 1–12. [CrossRef]
- Liu, Y.; Lv, J.; Liu, J.; Li, M.; Xie, J.; Lv, Q.; Deng, W.; Zhou, N.; Zhou, Y.; Song, J.; et al. Mucus production stimulated by IFN-AhR signaling triggers hypoxia of COVID-19. Cell Res. 2020, 30, 1078–1087. [CrossRef]
- Major, J.; Crotta, S.; Finsterbusch, K.; Chakravarty, P.; Shah, K.; Frederico, B.; D’aNtuono, R.; Green, M.; Meader, L.; Suarez-Bonnet, A.; et al. Endothelial AHR activity prevents lung barrier disruption in viral infection. Nature 2023, 621, 813–820. [CrossRef]
- Antunes, K.H.; Singanayagam, A.; Williams, L.; Faiez, T.S.; Farias, A.; Jackson, M.M.; Faizi, F.K.; Aniscenko, J.; Kebadze, T.; Veerati, P.C.; et al. Airway-delivered short-chain fatty acid acetate boosts antiviral immunity during rhinovirus infection. J. Allergy Clin. Immunol. 2022, 151, 447–457.e5. [CrossRef]
- Antunes, K.H.; Fachi, J.L.; De Paula, R.; Da Silva, E.F.; Pral, L.P.; DOS Santos, A.; Dias, G.B.M.; Vargas, J.E.; Puga, R.; Mayer, F.Q.; et al. Microbiota-derived acetate protects against respiratory syncytial virus infection through a GPR43-type 1 interferon response. Nat. Commun. 2019, 10, 3273. [CrossRef]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [CrossRef]
- He, Y.; Zhao, C.; Su, N.; Yang, W.; Yang, H.; Yuan, C.; Zhang, N.; Hu, X.; Fu, Y. Disturbances of the gut microbiota-derived tryptophan metabolites as key actors in vagotomy-induced mastitis in mice. Cell Rep. 2024, 44, 114585. [CrossRef]
- Martin, A.M.; Yabut, J.M.; Choo, J.M.; Page, A.J.; Sun, E.W.; Jessup, C.F.; Wesselingh, S.L.; Khan, W.I.; Rogers, G.B.; Steinberg, G.R.; et al. The gut microbiome regulates host glucose homeostasis via peripheral serotonin. Proc. Natl. Acad. Sci. 2019, 116, 19802–19804. [CrossRef]
- E Griffin, M.; Hespen, C.W.; Wang, Y.; Hang, H.C. Translation of peptidoglycan metabolites into immunotherapeutics. Clin. Transl. Immunol. 2019, 8, e1095. [CrossRef]
- Yang, M.; Qi, X.; Li, N.; Kaifi, J.T.; Chen, S.; Wheeler, A.A.; Kimchi, E.T.; Ericsson, A.C.; Rector, R.S.; Staveley-O’cArroll, K.F.; et al. Western diet contributes to the pathogenesis of non-alcoholic steatohepatitis in male mice via remodeling gut microbiota and increasing production of 2-oleoylglycerol. Nat. Commun. 2023, 14, 1–17. [CrossRef]
- Yoo, W.; Zieba, J.K.; Foegeding, N.J.; Torres, T.P.; Shelton, C.D.; Shealy, N.G.; Byndloss, A.J.; Cevallos, S.A.; Gertz, E.; Tiffany, C.R.; et al. High-fat diet–induced colonocyte dysfunction escalates microbiota-derived trimethylamine N -oxide. Science 2021, 373, 813–818. [CrossRef]
- Hagihara, M.; Yamashita, M.; Ariyoshi, T.; Eguchi, S.; Minemura, A.; Miura, D.; Higashi, S.; Oka, K.; Nonogaki, T.; Mori, T.; et al. Clostridium butyricum-induced ω-3 fatty acid 18-HEPE elicits anti-influenza virus pneumonia effects through interferon-λ upregulation. Cell Rep. 2022, 41, 111755. [CrossRef]
- Erttmann, S.F.; Swacha, P.; Aung, K.M.; Brindefalk, B.; Jiang, H.; Härtlova, A.; Uhlin, B.E.; Wai, S.N.; Gekara, N.O. The gut microbiota prime systemic antiviral immunity via the cGAS-STING-IFN-I axis. Immunity 2022, 55, 847–861.e10. [CrossRef]
- Platt, D.J.; Lawrence, D.; Rodgers, R.; Schriefer, L.; Qian, W.; Miner, C.A.; Menos, A.M.; Kennedy, E.A.; Peterson, S.T.; Stinson, W.A.; et al. Transferrable protection by gut microbes against STING-associated lung disease. Cell Rep. 2021, 35, 109113–109113. [CrossRef]
- Broggi, A.; Ghosh, S.; Sposito, B.; Spreafico, R.; Balzarini, F.; Cascio, A.L.; Clementi, N.; De Santis, M.; Mancini, N.; Granucci, F.; et al. Type III interferons disrupt the lung epithelial barrier upon viral recognition. Science 2020, 369, 706–712. [CrossRef]
- Lopez, C.E.; Zacharias, Z.R.; Ross, K.A.; Narasimhan, B.; Waldschmidt, T.J.; Legge, K.L. Polyanhydride nanovaccine against H3N2 influenza A virus generates mucosal resident and systemic immunity promoting protection. npj Vaccines 2024, 9, 1–14. [CrossRef]
- Gaisina, I.; Li, P.; Du, R.; Cui, Q.; Dong, M.; Zhang, C.; Manicassamy, B.; Caffrey, M.; Moore, T.; Cooper, L.; et al. An orally active entry inhibitor of influenza A viruses protects mice and synergizes with oseltamivir and baloxavir marboxil. Sci. Adv. 2024, 10, eadk9004. [CrossRef]
- Ou, G.; Xu, H.; Wu, J.; Wang, S.; Chen, Y.; Deng, L.; Chen, X. The gut-lung axis in influenza A: the role of gut microbiota in immune balance. Front. Immunol. 2023, 14, 1147724. [CrossRef]
- Ross, F.C.; Patangia, D.; Grimaud, G.; Lavelle, A.; Dempsey, E.M.; Ross, R.P.; Stanton, C. The interplay between diet and the gut microbiome: implications for health and disease. Nat. Rev. Microbiol. 2024, 22, 671–686. [CrossRef]
- Ghani, R.; Mullish, B.H.; McDonald, J.A.K.; Ghazy, A.; Williams, H.R.T.; Brannigan, E.T.; Mookerjee, S.; Satta, G.; Gilchrist, M.; Duncan, N.; et al. Disease Prevention Not Decolonization: A Model for Fecal Microbiota Transplantation in Patients Colonized With Multidrug-resistant Organisms. Clin. Infect. Dis. 2020, 72, 1444–1447. [CrossRef]
- Johnsen, P.H.; Hilpüsch, F.; Cavanagh, J.P.; Leikanger, I.S.; Kolstad, C.; Valle, P.C.; Goll, R. Faecal microbiota transplantation versus placebo for moderate-to-severe irritable bowel syndrome: a double-blind, randomised, placebo-controlled, parallel-group, single-centre trial. Lancet Gastroenterol. Hepatol. 2018, 3, 17–24. [CrossRef]
- van Lier, Y.F., et al., Donor fecal microbiota transplantation ameliorates intestinal graft-versus-host disease in allogeneic hematopoietic cell transplant recipients. Sci Transl Med, 2020. 12(556).
- Surawicz, C.M., et al., Guidelines for diagnosis, treatment, and prevention of Clostridium difficile infections. Am J Gastroenterol, 2013. 108(4): p. 478-98; quiz 499.
- Liu, F.; Ye, S.; Zhu, X.; He, X.; Wang, S.; Li, Y.; Lin, J.; Wang, J.; Lin, Y.; Ren, X.; et al. Gastrointestinal disturbance and effect of fecal microbiota transplantation in discharged COVID-19 patients. J. Med Case Rep. 2021, 15, 1–9. [CrossRef]
- Biliński, J.; Winter, K.; Jasiński, M.; Szczęś, A.; Bilinska, N.; Mullish, B.H.; Małecka-Panas, E.; Basak, G.W. Rapid resolution of COVID-19 after faecal microbiota transplantation. Gut 2021, 71, 230–232. [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: from biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [CrossRef]
- Cameron, D.; Hock, Q.S.; Kadim, M.; Mohan, N.; Ryoo, E.; Sandhu, B.; Yamashiro, Y.; Jie, C.; Hoekstra, H.; Guarino, A. Probiotics for gastrointestinal disorders: Proposed recommendations for children of the Asia-Pacific region. World J. Gastroenterol. 2017, 23, 7952–7964. [CrossRef]
- Hu, J.; Zhang, L.; Lin, W.; Tang, W.; Chan, F.K.; Ng, S.C. Review article: Probiotics, prebiotics and dietary approaches during COVID-19 pandemic. Trends Food Sci. Technol. 2021, 108, 187–196. [CrossRef]
- Xavier-Santos, D.; Padilha, M.; Fabiano, G.A.; Vinderola, G.; Cruz, A.G.; Sivieri, K.; Antunes, A.E.C. Evidences and perspectives of the use of probiotics, prebiotics, synbiotics, and postbiotics as adjuvants for prevention and treatment of COVID-19: A bibliometric analysis and systematic review. Trends Food Sci. Technol. 2022, 120, 174–192. [CrossRef]
- D'ETtorre, G.; Ceccarelli, G.; Marazzato, M.; Campagna, G.; Pinacchio, C.; Alessandri, F.; Ruberto, F.; Rossi, G.; Celani, L.; Scagnolari, C.; et al. Challenges in the Management of SARS-CoV2 Infection: The Role of Oral Bacteriotherapy as Complementary Therapeutic Strategy to Avoid the Progression of COVID-19. Front. Med. 2020, 7, 389. [CrossRef]
- Ceccarelli, G.; Borrazzo, C.; Pinacchio, C.; Santinelli, L.; Innocenti, G.P.; Cavallari, E.N.; Celani, L.; Marazzato, M.; Alessandri, F.; Ruberto, F.; et al. Oral Bacteriotherapy in Patients With COVID-19: A Retrospective Cohort Study. Front. Nutr. 2021, 7, 613928. [CrossRef]
- McCarville, J.L.; Chen, G.Y.; Cuevas, V.D.; Troha, K.; Ayres, J.S. Microbiota Metabolites in Health and Disease. Annu. Rev. Immunol. 2020, 38, 147–170. [CrossRef]
- Van Treuren, W. and D. Dodd, Microbial Contribution to the Human Metabolome: Implications for Health and Disease. Annu Rev Pathol, 2020. 15: p. 345-369.
- Zuo, T.; Liu, Q.; Zhang, F.; Lui, G.; Tso, E.; Yeoh, Y.K.; Chen, Z.; Boon, S.; Chan, F.K.L.; Chan, P.; et al. Depicting SARS-CoV-2 faecal viral activity in association with gut microbiota composition in patients with COVID-19. Gut 2020, 70, 276–284. [CrossRef]
- Gu, S.; Chen, Y.; Wu, Z.; Chen, Y.; Gao, H.; Lv, L.; Guo, F.; Zhang, X.; Luo, R.; Huang, C.; et al. Alterations of the Gut Microbiota in Patients With Coronavirus Disease 2019 or H1N1 Influenza. Clin. Infect. Dis. 2020, 71, 2669–2678. [CrossRef]
- Zhang, F.; Wan, Y.; Zuo, T.; Yeoh, Y.K.; Liu, Q.; Zhang, L.; Zhan, H.; Lu, W.; Xu, W.; Lui, G.C.; et al. Prolonged Impairment of Short-Chain Fatty Acid and L-Isoleucine Biosynthesis in Gut Microbiome in Patients With COVID-19. Gastroenterology 2021, 162, 548–561.e4. [CrossRef]
- Piscotta, F.J.; Hoffmann, H.-H.; Choi, Y.J.; Small, G.I.; Ashbrook, A.W.; Koirala, B.; Campbell, E.A.; Darst, S.A.; Rice, C.M.; Brady, S.F.; et al. Metabolites with SARS-CoV-2 Inhibitory Activity Identified from Human Microbiome Commensals. mSphere 2021, 6, e0071121. [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [CrossRef]
- Chen, S.W.; Zhang, W. Current trends and challenges in the downstream purification of bispecific antibodies. Antib. Ther. 2021, 4, 73–88. [CrossRef]
- Mitragotri, S.; Burke, P.A.; Langer, R. Overcoming the challenges in administering biopharmaceuticals: formulation and delivery strategies. Nat. Rev. Drug Discov. 2014, 13, 655–672. [CrossRef]
- Charbonneau, M.R.; Isabella, V.M.; Li, N.; Kurtz, C.B. Developing a new class of engineered live bacterial therapeutics to treat human diseases. Nat. Commun. 2020, 11, 1–11. [CrossRef]
- Roslan, M.A.M.; Omar, M.N.; Sharif, N.A.M.; Raston, N.H.A.; Arzmi, M.H.; Neoh, H.-M.; Ramzi, A.B. Recent advances in single-cell engineered live biotherapeutic products research for skin repair and disease treatment. npj Biofilms Microbiomes 2023, 9, 1–9. [CrossRef]
- Cui, L.; Vigouroux, A.; Rousset, F.; Varet, H.; Khanna, V.; Bikard, D. A CRISPRi screen in E. coli reveals sequence-specific toxicity of dCas9. Nat. Commun. 2018, 9, 1–10. [CrossRef]
- Crook, N.; Ferreiro, A.; Gasparrini, A.J.; Pesesky, M.W.; Gibson, M.K.; Wang, B.; Sun, X.; Condiotte, Z.; Dobrowolski, S.; Peterson, D.; et al. Adaptive Strategies of the Candidate Probiotic E. coli Nissle in the Mammalian Gut. Cell Host Microbe 2019, 25, 499–512.e8. [CrossRef]
- Xin, Y.; Liu, C.-G.; Zang, D.; Chen, J. Gut microbiota and dietary intervention: affecting immunotherapy efficacy in non–small cell lung cancer. Front. Immunol. 2024, 15, 1343450. [CrossRef]
- McQuade, J.L.; Daniel, C.R.; A Helmink, B.; A Wargo, J. Modulating the microbiome to improve therapeutic response in cancer. Lancet Oncol. 2019, 20, e77–e91. [CrossRef]
- Gouez, M., et al., [Nutrition and physical activity (PA) during and after cancer treatment: Therapeutic benefits, pathophysiology, recommendations, clinical management]. Bull Cancer, 2022. 109(5): p. 516-527.
- Shan, G.; Minchao, K.; Jizhao, W.; Rui, Z.; Guangjian, Z.; Jin, Z.; Meihe, L. Resveratrol improves the cytotoxic effect of CD8 +T cells in the tumor microenvironment by regulating HMMR/Ferroptosis in lung squamous cell carcinoma. J. Pharm. Biomed. Anal. 2023, 229, 115346. [CrossRef]
- Saraiva, A.; Raheem, D.; Roy, P.R.; BinMowyna, M.N.; Romão, B.; Alarifi, S.N.; Albaridi, N.A.; Alsharari, Z.D.; Raposo, A. Probiotics and Plant-Based Foods as Preventive Agents of Urinary Tract Infection: A Narrative Review of Possible Mechanisms Related to Health. Nutrients 2025, 17, 986. [CrossRef]
Figure 1.
Obesity increases the body's susceptibility to influenza virus infection. Obesity can influence the host's resistance to the influenza virus in multiple aspects, including the lung, liver, intestines, and immunity.
Figure 1.
Obesity increases the body's susceptibility to influenza virus infection. Obesity can influence the host's resistance to the influenza virus in multiple aspects, including the lung, liver, intestines, and immunity.
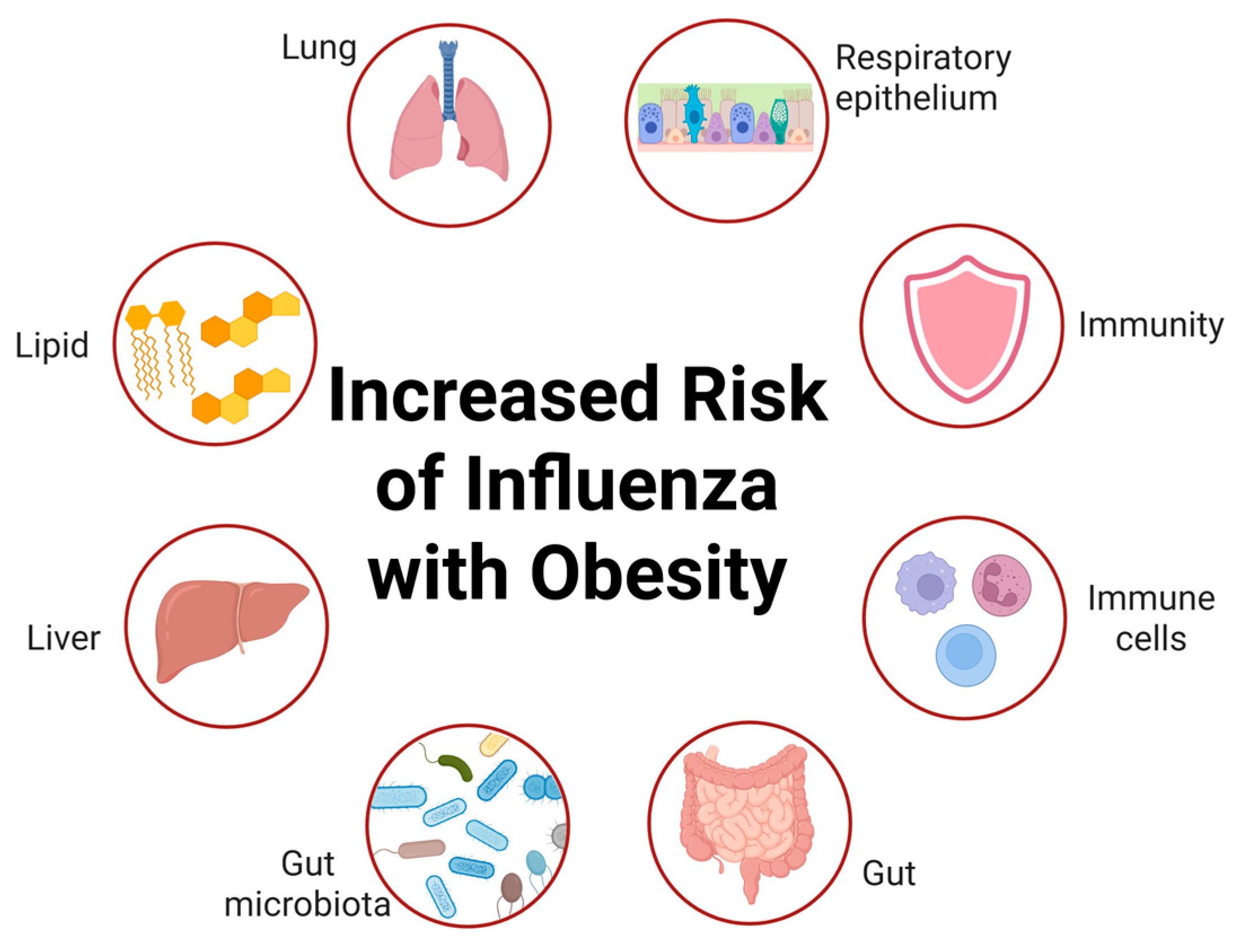

Table 1.
Potential targets of interaction between obesity and influenza.
| Target Protein/ Pathway | Function | References |
|---|---|---|
| ABHD6 | α/β-hydrolase domain 6 (ABHD6) is a lipase affecting energy metabolism. | [26] |
| HDAC6 | Histone deacetylase 6 (HDAC6)re-sensitizes leptin signaling during obesity. | [27] |
| LGR4 | G-protein-coupled receptor 4 (LGR4) impacts long-chain fatty acid-absorption. | [28] |
| NK2R | Neurokinin 2 receptor (NK2R) can increase energy expenditure peripherally. | [29] |
| NMDA receptor | The N-methyl-D-aspartate (NMDA) receptor antagonism can treat obesity. | [30] |
| PICK1, PSD95 | Protein interacting with C kinase 1 and postsynaptic density protein-95 targeting postsynaptic glutamate receptor for obesity treatment. | [31] |
| PTER | Orphan enzyme phosphotriesterase-related (PTER) is a N-acetyltaurine hydrolase. | [32] |
| PXR | Pregnane X receptor (PXR) can regulate glycolipid metabolism. | [33] |
| SREBP2, RORγ | Sterol regulatory element-binding protein 2 (SREBP2)and the retinoid acid receptor-related orphan receptor gamma (RORγ) regulate cholesterol metabolism. | [34] |
| RUVBL2 | Knockout of PVH RUVBL2 results in hyperphagic obesity. | [35] |
Table 2.
Cluster of Differentiation in viral infections.
| Cluster of Differentiation | Efficacy | References |
|---|---|---|
| CD4 | CD4 T cells play a multiplicity of roles in protective immunity to influenza, viral antigen specificity. | [58] |
| CD8 | CD8 T cells provide broadly cross-reactive immunity and alleviate disease severity by recognizing conserved epitopes. | [61] |
| CD11 | CD11b+ cDC2 subsets present in mice regulated by IRF4 during IAV infection. | [63,64] |
| CD27 | CD45RA−CD27− effector memory-like T-cells increase in IAV- and IBV-infected patients. | [65] |
| CD38 | CD38+Ki67+CD8+ effector T cells increase in IAV infected pediatric and adult subjects. | [66] |
| CD45 | The CD45-positive macrophages expressing mCherry increase in IAV-infected patients. | [63,65] |
| CD64 | Mice lacking myeloid TBK1 showed less recruitment of CD64+SiglecF-Ly6C inflammatory macrophages. | [63] |
| CD69 | CD69+CD103+ TRM cells preferentially localized to lung sites of prior IAV infection. | [67] |
| CD103 | Vaccine can induced lung tissue-resident memory T cells expressing high levels of CD103. | [67,68] |
| CD122 | Once memory to influenza is established and enhance NF-κB signaling in T cells can increases CD122 levels. | [69] |
Table 3.
The effect of microbial metabolites in host during virus infection.
| Microbial metabolites | Bacteria | Efficacy | References |
|---|---|---|---|
| Acetate | Acetobacter and Bifidobacterium pseudolongum | Acetate can trigger antiviral immune. | [87,88] |
| Butyrate | Clostridium butyricum and Butyrivibrio | Butyrate reprograms CD8+ T cells by promoting glutamine utilization and fatty acid oxidation. | [61] |
| LPS | Gram-negative bacteria | LPS can activate the TLR4 pathwayto trigger the NF-κB signaling pathway and regulate the inflammatory response. | [40] |
| BCAA |
Prevotellacopri and Bacteroides vulgatus |
Branched-chain amino acid can induce insulin resistance. | [89] |
| Indole derivatives (e.g. IAA, IPA, 5-HIAA) | Escherichia coli, Proteus and Vibrio cholerae | They can activate the AhR. | [90] |
| 5-HT | Enterochromaffin cells produce 5-HT influencing by gut microbiota | 5-hydroxytryptaminecan regulate glucose homeostasis. | [91] |
| PGN | All species of bacteria | Peptidoglycan can activate host immunity. | [92] |
| 2-octagenoate | Blautia bacterium | 2-octagenoate can lead to liver hypertrophy, steatosis, inflammation of liver cells, and fibrosis. |
[93] |
| DAT | Clostridium orbiscindens | DAT can trigger tonic IFN signaling and regulate the phagocytic activity of macrophages. | [83] |
| TMA | Gut microbiota | Trimethylamine converted to Trimethylamine-N-oxide (TMAO) in the liver. TMAO regulates glucose metabolism and causes adipose tissue inflammation. | [94] |
| Bile acids | Clostridium scindens | BAs activate virus-induced NF-κB. | [82] |
| 18-HEPE | Clostridium strain C. butyricum | 18-HEPE activates the production of tonic IFN-λ by lung epithelial cells via GPR120, leading to enhanced resistance to influenza infection. | [95] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



