Submitted:
27 June 2025
Posted:
30 June 2025
You are already at the latest version
Abstract
Variegated Cymbidium lancifolium is a highly valued ornamental plant, sought after in both local and international markets. The commercial production of variegated C. lancifolium through traditional propagation methods faces significant challenges, such as low propagation rates and prolonged growth timeframes. This study aims to develop effective in vitro propagation techniques for variegated C. lancifolium through asymbiotic seed germination to enhance production efficiency and meet market demand. We examined the effects of various plant growth regulators and coconut water (CW) on in vitro seed germination. The highest germination percentage (46.8%) was recorded in Murashige and Skoog (MS) medium supplemented with 50 mL/L CW, 4.0 µM α-naphthalene acetic acid (NAA), 2.3 µM kinetin (KN), and 2.9 µM gibberellic acid (GA3). The seed-derived rhizomes were placed on MS medium containing indole-3-acetic acid (IAA), indole-3-butyric acid (IBA), and NAA for proliferation. Among the auxins, NAA proved to be the most effective, significantly increasing rhizome proliferation, with the highest number (17.4) and length (2.1 cm) observed at 5.0 µM. The rhizome explants were cultured on MS medium enriched with Kinetin (KN), N6-(2-isopentenyl)adenine (2-IP), and N6-benzyladenine (BA) to promote plantlet regeneration. Of the cytokinins tested, BA at 10.0 µM resulted in the highest rate of plantlet regeneration (79.4%), the greatest number of plantlets (4.4 per culture), and notable plantlet height (8.5 cm). We obtained plantlets characterized by dark green leaves, light green leaves, and distinct variegation patterns. They were transferred into three different substrate mixtures for acclimatization. The substrate made of orchid stone (30%), wood bark (30%), coconut husk chip (20%), and perlite (20%) supported the highest survival rate (95.9%). The study successfully established optimized in vitro propagation techniques for variegated C. lancifolium, enabling enhanced germination, rhizome proliferation, and plantlet regeneration to meet the growing market demand.
Keywords:
asymbiotic seed germination
; auxin
; cytokinin
; gibberellic acid 3
; coconut water
; orchids
1. Introduction
Orchids are among the most advanced monocotyledonous plants and have been well-known to humans for a considerable time. The genus Cymbidium Swartz comprises 70 species [1], often referred to as the orchids of the East. These species are found in temperate regions such as Korea, Japan, and China, and have been valued as noble plants since ancient times [1,2]. Cymbidium species are prized ornamental plants that are frequently sold as cut flowers and potted plants. Moreover, they are used in landscape gardening and can be incorporated into hanging baskets [3,4]. Cymbidium lancifolium, a small terrestrial herb, sometimes grows saxicolous or epiphytic at elevations ranging from 300 to 2200 m. It belongs to the Orchidaceae family and is native to Korea as well as the tropical and subtropical regions of Asia [1]. C. lancifolium is recognized as a critically endangered species and is listed in the Red Data [5]. Therefore, conservation efforts for C. lancifolium are necessary.
The conventional propagation of C. lancifolium through both asexual and sexual methods is relatively slow due to its slow growth and the challenges associated with seed germination under natural conditions [5]. Therefore, effective techniques for the large-scale propagation of this species for commercial purposes are essential. Tissue culture methods have been widely used to propagate various orchids. Among these in vitro propagation techniques, asymbiotic seed germination is often employed to produce rare and commercially important orchids, including Cymbidium species [6]. This technique is also useful for introducing new isolates, genotypes, and cultivars in a short period. Numerous reports detail the in vitro propagation of Cymbidium species via asymbiotic seed germination; however, this method is significantly influenced by several factors, including genotype, seed age, type and composition of the culture medium, plant growth regulators (PGRs), organic additives, and the culture environment [6,7,8,9].
Li et al. [10] reported that seeds of Cymbidium faberi ‘Jiepeimei’ × Cymbidium sinense ‘Qijianheimo’ collected 90 and 105 days after pollination failed to germinate in vitro. However, seeds collected 120, 135, or 150 days after pollination successfully germinated in Knudson medium. Hossain et al. [11] noted that seeds of Cymbidium giganteum achieved an impressive 100% germination rate in vitro when cultured on Phytomax or Mitra media supplemented with either peptone (2 g/L) or 6-benzylaminopurine (BAP, 1 mg/L). Similarly, Cymbidium elegans seeds germinated best on MS medium with 1 mg/L BAP [12]. In contrast, Cymbidium iridioides seeds germinated optimally in MS medium with 1 mg/L each of BAP and α-naphthalene acetic acid (NAA) [13]. Seeds obtained from a nine-month-old seed capsule of Cymbidium aloifolium exhibited the highest seed germination rate of 49% on Murashige and Skoog (MS) medium, irrespective of the addition of 0.5 mg/L BAP [14]. In contrast, seeds from seven-month-old Cymbidium aloifolium seed capsules germinated best (98.3%) on MS medium enriched with 0.5 mg/L each of BAP and NAA [15]. Furthermore, the highest in vitro seedling germination for Cymbidium findlaysonianum was achieved by culturing seeds on Vacin and Went medium enriched with a combination of activated charcoal (AC, 0.2%), banana homogenate (5%), coconut water (CW, 15%), potato homogenate (5%), and 20 g/L sucrose [16]. In another study, Cymbidium nanutum immature seeds germinated best (45%) on half-strength MS medium containing 0.5 mg/L NAA, 30 g/L sucrose, AC (1 g/L), and coconut milk (100 mL/L) [17].
Currently, only two reports exist in the literature regarding the in vitro propagation of C. lancifolium [18,19]. The authors propagate C. lancifolium by culturing rhizomes obtained from asymbiotically germinated seeds on a semi-solid Hyponex medium containing 2 g/L peptone and 30 g/L sucrose. However, the percentage of seed germination has not been disclosed. Additionally, the in vitro propagation of variegated C. lancifolium has not been documented. Variegated C. lancifolium plants (Figure 1A and B) exhibit unique leaf variegation characterized by a mix of green, yellow, and white sectors distributed along the leaf blade. The variegation pattern is primarily striated, featuring longitudinal stripes that run parallel to the leaf veins. Some leaves present broader yellowish-white sectors, while others showcase finer green and yellow striping. Although C. lancifolium is known for its attractive foliage, the variegated form is particularly sought after due to its distinctive leaf patterns. The commercial production of variegated C. lancifolium through conventional propagation methods poses challenges, including low propagation rates and extended time requirements. Therefore, this study aims to establish efficient in vitro propagation techniques for variegated C. lancifolium via asymbiotic seed germination.
2. Results
2.1. Effects of PGRs and CW on Asymbiotic Seed Germination
Only 0.2% of the variegated C. lancifolium seeds germinated in MS medium without PGRs and CW. The addition of auxins (indole-3-acetic acid (IAA) or NAA) enhanced the seed germination rate. Variegated C. lancifolium seeds (Figure 1C,D) swelled within a month of cultivation (Figure 1E). The formation of white and green protocorms was observed after two months (Figure 1E,F), and the protocorm subsequently developed into a green rhizome (Figure 1G) after four months of cultivation. We also obtained many yellowish-green rhizomes and a few white ones. The percentage of seed germination increased in a dose-dependent manner with IAA supplementation. Raising the concentration of IAA from 0.5 to 8.0 µM raised the germination rate from 0.7% to 10.2%. Similarly, NAA enhanced germination, reaching a maximum of 16.8% at 4.0 µM. However, increasing NAA concentration to 8.0 µM reduced the germination percentage to 12.6% (Table 1). Further enhancement occurred with the addition of CW to the medium containing NAA (4.0 µM). Among the tested CW concentrations (25–100 mL/L), 50 mL/L yielded the highest germination rate (26.2%), while both lower (25 mL/L) and higher (75 and 100 mL/L) concentrations were less effective. The combination of NAA (4.0 µM) and CW (50 mL/L) with kinetin (KN) further boosted germination, with the highest response (32.2%) recorded at 2.3 µM KN. Increasing KN concentrations beyond this level did not significantly enhance germination (Table 1). Notably, adding gibberellic acid (GA3 0.3-2.9 µM) to the optimized NAA (4.0 µM) + KN (2.3 µM) + CW (50 mL/L) treatment significantly improved germination. A progressive increase in germination occurred with rising GA3 concentrations, with the highest percentage (46.8%) achieved at 2.9 µM GA3 (Table 1).
2.2. Effects of Auxins on Rhizome Proliferation
The rhizomes (dark green, yellowish-green, and white) obtained from the seed germination medium [MS + NAA (4.0 µM) + KN (2.3 µM) + CW (50 mL/L) + GA3 (2.9 µM)] served as explants for rhizome proliferation. The rhizomes of variegated C. lancifolium (Figure 1H) developed branches within a month of cultivation (Figure 1F). After 6 months of cultivation on MS medium lacking auxins, the number (1.3) and length (0.5 cm) of the rhizomes were lower. The addition of auxin enhanced rhizome growth and proliferation. Among the three auxins tested, NAA proved to be the most effective, particularly at 5.0 µM, which yielded the highest number of rhizomes (17.4) with a corresponding length of 2.1 cm (Table 2, Figure 1J). This was followed by 10.0 µM NAA and 10.0 µM IBA, which also resulted in substantial rhizome multiplication (12.4 and 11.0, respectively), although with slightly shorter lengths (1.6 and 1.7 cm). The longest rhizomes were observed at 5.0 µM IBA (2.5 cm) and 2.5 µM NAA (2.4 cm), indicating that both IBA and NAA effectively promoted rhizome elongation and multiplication at specific concentrations. In contrast, IAA was the least effective auxin, producing relatively fewer and shorter rhizomes across all tested concentrations. The maximum response for IAA was recorded at 15.0 µM, resulting in 6.2 rhizomes with an average length of 1.1 cm. ANOVA results confirmed significant main effects of auxin type, concentration, and their interaction on both rhizome number (R2 = 0.748) and length (R2 = 0.491) (Table 2).
The type of auxin and its concentration significantly influenced both the number and length of rhizomes. Among the tested auxins, NAA produced the highest average number of rhizomes (10.0), followed by IBA (8.3), while IAA resulted in the lowest number (3.8). A similar trend was observed for rhizome length, with NAA (1.9 cm) and IBA (1.8 cm) promoting significantly greater elongation than IAA (1.1 cm) (Figure 2A). Regarding concentration, the highest rhizome number was recorded at 5.0 µM (9.4) and 10.0 µM (9.2), both significantly greater than 2.5 µM (4.2) and 15.0 µM (7.1). Rhizome length peaked at 5.0 µM (2.0 cm) and 2.5 µM (1.8 cm), while higher concentrations (10.0 and 15.0 µM) led to reduced elongation (Figure 2B). These results indicate that both auxin type and concentration interact to affect rhizome induction, with NAA at moderate concentrations (5.0–10.0 µM) being the most effective for promoting both rhizome number and length.
2.3. Effects of Cytokinins on Plantlet Regeneration
Rhizome segments cultured on MS medium without cytokinin were found to be ineffective in inducing shoot buds. However, the addition of cytokinin promoted plantlet regeneration. The rhizome explants produced many rhizomes, and the growing tips developed shoot buds within four months, subsequently developing roots at the bottom of the shoots. The rhizome explants yielded 95% green plantlets, 4% light green plantlets, and 1% variegated plantlets (Figure 1K-R). Furthermore, the green plantlets (Figure 1N) grew faster, followed by light-green (Figure 1L,M,O) and variegated plantlets (Figure 1K,P,Q,R). The regeneration of plantlets was significantly affected by 2-IP, BA and KN (Table 3). Among the cytokinins tested, N6-benzyladenine (BA) proved to be the most effective in promoting plantlet regeneration. The highest regeneration percentage (79.4%) occurred at 10.0 µM BA, which also resulted in the greatest number of plantlets per rhizome (4.4) and significant plantlet height (8.5 cm). Increasing BA concentration from 2.5 to 10.0 µM enhanced regeneration and plantlet proliferation, while a further increase to 20.0 µM caused a decline in both parameters. KN at 5.0 µM also supported high regeneration (46.7%) and plantlet height (9.5 cm), although the number of plantlets per rhizome (2.2) was lower compared to BA. Similarly, N6-(2-isopentenyl)adenine (2-IP) induced moderate responses, with 5.0 µM yielding 38.9% regeneration and 2.1 plantlets per rhizome. However, at higher concentrations (10.0–20.0 µM), both 2-IP and KN showed reduced effectiveness (Table 3). Plantlet height varied depending on cytokinin type and concentration, with the tallest plantlets recorded in BA (10.2 cm at 5.0 µM) and KN (9.5 cm at 5.0 µM) treatments. Statistical analysis revealed highly significant effects of cytokinin type, concentration, and their interaction on all three parameters, with the highest R2 value observed for plantlet regeneration (0.919), indicating strong treatment responsiveness (Table 3).
Cytokinin type and concentration significantly affected plantlet regeneration, the number of plantlets per rhizome, and plantlet height. Among the cytokinins tested, BA proved to be the most effective, leading to the highest plantlet regeneration percentage (53.8%), the greatest number of plantlets (2.4), and the tallest plantlets (7.8 cm), followed by KN and 2-IP, which displayed moderate responses. KN promoted 33.5% regeneration, yielding 2.0 plantlets per culture and a mean height of 6.9 cm, while 2-IP was the least effective, showing 20.8% regeneration with shorter plantlets (6.2 cm) and fewer shoots (1.6) (Figure 3A). In terms of concentration, 5.0 µM cytokinin produced the most favorable results across all parameters, including the highest plantlet height (8.9 cm), a high regeneration percentage (51.1%), and a significant number of plantlets (2.2). A concentration of 10.0 µM also supported a high number of plantlets (2.6), although with slightly lower regeneration (42.6%) and plantlet height (7.2 cm). Both lower (2.5 µM) and higher (20.0 µM) concentrations were less effective, particularly at 20.0 µM, where plantlet height significantly decreased (5.0 cm) (Figure 3B). These findings suggest that BA at 5.0–10.0 µM is optimal for plantlet regeneration and growth in rhizome-derived cultures.
2.4. Acclimatization of Plantlets
The survival rate of regenerated orchid plantlets during the acclimatization phase varied significantly depending on the potting substrate composition. The highest survival percentage (95.9%) was observed in a medium composed of orchid stone (30%), wood bark (30%), coconut husk chip (20%) and perlite (20%) (Figure 4), indicating that a well-balanced mix of moisture retention and aeration components supports optimal plantlet establishment. A substrate consisting of orchid stone and wood bark in equal proportions (50:50) also resulted in high survival (82.1%), while a mixture of orchid stone (50%), coconut husk chip (10%), coconut fiber (20%) and perlite (20%) showed reduced survival (66.4%), possibly due to over-retention of moisture or inadequate aeration. After three months, the plantlets were transferred into pots containing orchid stone (50%) and wood bark (50%) because the plantlets grew best in this substrate mixture compared to the other two mixtures studied. These results underscore the importance of optimizing substrate composition to enhance the post-in vitro survival of orchid plantlets.
3. Discussion
Although orchid seed capsules contain many seeds, germination in nature is difficult because the seeds have undifferentiated embryos and lack the endosperm and a fungal partner required for orchid seed germination [20]. This issue can be addressed through an in vitro seed germination technique, which allows orchid seeds to germinate asymbiotically in vitro, leading to successful plant regeneration [15,21]. However, the asymbiotic seed germination of terrestrial Cymbidium species native to temperate regions is known to be challenging [22]. In this study, we demonstrate, for the first time, the in vitro propagation of variegated C. lancifolium through asymbiotic seed germination. The successful in vitro germination of this terrestrial orchid under asymbiotic conditions is influenced by PGRs and CW (Table 1). Auxin regulates germination and protocorm development in orchids during asymbiotic in vitro germination [23]. The inclusion of auxin, particularly NAA, promoted the germination of variegated C. lancifolium seeds in vitro. The beneficial effect of NAA on asymbiotic seed germination has been reported in various Cymbidium species, including Cymbidium aloifolium [24], Cymbidium elegans [12], and Cymbidium. nanutum [17].
Various organic supplements, such as banana pulp, casein hydrolysate, CW, peptone, pineapple juice, potato homogenate, tomato juice, and yeast extract, are often added to the culture medium to enhance asymbiotic seed germination of orchids in vitro and subsequent growth and development [25,26]. In this study, incorporating CW into medium containing auxin (NAA 4.0 µM) improved the germination of variegated C. lancifolium seeds in vitro. CW, known as liquid endosperm, promotes orchid seed germination due to its reported content of various growth-promoting substances such as abscisic acid, amino acids, IAA, GA, minerals, proteins, sugars, vitamins, and zeatin riboside [25,27,28,29]. The beneficial effect of CW on asymbiotic seed germination has also been noted in Cymbidium aloifolium [30], Cymbidium devonianum [31], Cymbidium findlaysonianum [16], and Cymbidium hybrid [32].
It has been reported that the supplementation of auxin (NAA) and cytokinin (BA/BAP) in the culture medium demonstrated a better response for asymbiotic seed germination of Cymbidium aloifolium [15,33], Cymbidium faberi [34], Cymbidium devonianum [31], Cymbidium eburneum [35], Cymbidium elegans [12], Cymbidium iridioides [13,36], and Cymbidium lowianum [37]. On the other hand, a combination of BAP and 2,4-dichlorophenoxyacetic acid (2,4-D) decreased the percentage of seed germination in Cymbidium giganteum [11]. In this study, a combination of NAA (4.0 µM) and KN (2.3 µM) enhanced the germination of variegated C. lancifolium seeds. The result is in accordance with Kang and Yang [38], who reported that 14 Cymbidium goeringii F1 hybrid seeds germinated on a medium containing 0.1 mg/L NAA and 0.01 mg/L KN. GA3 is one of the gibberellic acids used to promote in vitro asymbiotic seed germination of orchids [39,40]; however, the effect of GA3 on seed germination in Cymbidium species has not been disclosed. In this study, variegated C. lancifolium seeds germinated best (46.8%) on medium containing 50 mL CW, 4.0 µM NAA, 2.3 µM KN, and 2.9 µM GA3. Similarly, Comparetia falcate seeds achieved the highest germination rate (100%) on medium with 15 µM of both KN and GA3 [41]. In another study, Gastrochilus matsuran seeds germinated best (93.3%) on medium containing 5% CW, 1 µM NAA, and 1.5 µM GA3 [42].
The formation of rhizomes is a key step in the in vitro propagation of terrestrial orchids, particularly temperate Cymbidium species. Auxins, especially NAA and IBA, enhance rhizome proliferation and growth in variegated C. lancifolium, likely by promoting cell elongation of rhizomes. The best proliferation of variegated C. lancifolium rhizomes (17.4 rhizomes with a mean length of 2.1 cm per explant) occurred on MS medium containing 5.0 µM NAA (Table 2). The beneficial effects of NAA on rhizome multiplication and growth were also reported in other Cymbidium species [43]; however, the optimal concentration of NAA varied among the species: Cymbidium aloifolium (27.0 µM) [44], Cymbidium faberi (5.4 µM) [45], Cymbidium forrestii (10.8 µM) [46], and Cymbidium sinense (5.4 µM) [47]. Additionally, rhizome proliferation and growth of C. kanran were observed to be best on medium containing 54.0 µM and 5.4 µM of NAA, respectively [48].
Regeneration of variegated C. lancifolium plantlets from rhizome explants occurred only when the medium was supplemented with cytokinin; however, the production of plantlets was influenced by the type and concentration of cytokinins. Among the cytokinins studied, BA at 10.0 µM was determined to be the most effective for plantlet regeneration. With this treatment, 79.4% of rhizome explants developed an average of 4.4 plantlets that reached a height of 8.5 cm (Table 3). The requirement of cytokinin (BA) for shoot or plantlet regeneration from rhizome tips has been documented in Cymbidium aloifolium [49], Cymbidium faberi [45], Cymbidium forrestii [46], C. lancifolium [18], and Cymbidium sinense [47]. The plantlets obtained from this study exhibited a range of striking and distinct variegation patterns, each reminiscent of the variegated C. lancifolium mother plant. Some plantlets displayed prominent marginal and interveinal variegation with creamy-yellow stripes along the leaf edges and between the veins, while the midrib remained green (Figure 1K), suggesting a moderately consistent chimeric expression with ornamental appeal. Others featured a broad central yellow stripe along the midrib, flanked by green margins (Figure 1P), indicating potential genetic fixation and enhancing their value as uniform ornamental cultivars. Additionally, certain plantlets showcased both central and marginal variegation, characterized by vivid yellow coloration along the midrib and thinner yellow bands along the leaf edges (Figure 1Q), making them strong candidates for commercial propagation. Finally, some exhibited mild and irregular sectorial variegation (Figure 1R), suggesting partial expression or unstable chimerism that may offer opportunities for further selection and stabilization through clonal propagation.
Acclimatization of plantlets is a crucial final step in micropropagation, and the survival rate of Cymbidium species after hardening in different substrates varies significantly based on the medium used. For instance, Cymbidium aloifolium plantlets have demonstrated an impressive survival rate of 83% when acclimatized in a mixture of peat moss and small pieces of brick [49]. In contrast, Park et al. [2] reported a 97% survival rate for Cymbidium goeringii plantlets hardened on sphagnum moss. In another study, Cymbidium eburneum plantlets showed a 70% survival rate when hardened on a mixture of brick, charcoal, and decaying litter in a ratio of 1:1:1, along with a layer of moss [35]. Meanwhile, Cymbidium findlaysonianum plantlets displayed a survival rate of 70% when acclimatized in coconut peat [16]. Additionally, a study by Wang et al. [37] found that Cymbidium lowianum plantlets achieved a 92% survival rate when hardened on moss. In the present study, among the three different substrate mixes tested, the highest survival rate (95.9%) of variegated C. lancifolium plantlets was obtained on a mixture of orchid stone (30%), wood bark (30%), coconut husk chip (20%), and perlite (20%). These findings highlight the significant impact that substrate choice has on the survival of Cymbidium plantlets during the acclimatization process.
4. Materials and Methods
4.1. Plant Materials and Surface Disinfection
The five-year-old variegated Cymbidium lancifolium plants, grown in a shaded polyhouse, were self-pollinated on November 19, 2021 (Figure 1A), and the seed capsules were collected on September 14, 2022 (Figure 1B). The seed capsules were placed in a 500 mL glass beaker containing 5 mL of liquid detergent (Pril, Chongju, Korea) and then kept under running tap water for 20 minutes. They were subsequently washed with distilled water and air-dried. The washed capsules were surface-disinfected in a laminar airflow cabinet by soaking them in 70% ethanol (95%) for 3 minutes, followed by treatment with 10% sodium hypochlorite (9%, Daejung, Daejeon, Korea) for 15 minutes. Each treatment was followed by washing twice and four times, respectively, with sterile distilled water. Finally, the seed capsules were dipped in ethanol (99.9%, Daejung, Daejeon, Korea) for 30 seconds and then flamed off.
4.2. Asymbiotic Seed Germination
The single rib of the surface-disinfected seed capsule was dissected longitudinally, and the seeds were placed in a 500 mL culture bottle containing 150 mL of MS medium with vitamins, supplemented with 500 mg/L AC, 30 g/L sucrose, and 7 g/L plant agar (control medium), along with various concentrations of PGRs and coconut water (Table 1). The pH was adjusted to 5.5-5.6 using 1N potassium hydroxide or 1N hydrochloric acid before autoclaving for 23 minutes at 123°C. GA3 was filter-sterilized and added to the autoclaved medium. The AC, MS medium, PGRs, plant agar, and sucrose were sourced from Duchefa, Haarlem, The Netherlands. Nine culture bottles were used for each treatment, with each containing 500 ± 80 seeds. The cultures were maintained at 23 ± 2°C under an 8-hour photoperiod using white light-emitting diodes (WLEDs) with a photosynthetic photon flux density (PPFD) of 6-10 µmol s−1 m−2. The percentage of seed germination was recorded after four months of culture.
4.3. Rhizome Proliferation
The rhizomes (0.1 mm long) obtained from the germinated seeds were cultured on MS medium containing 500 mg/L AC, 50 mL/L CW, 7 g/L plant agar, 30 g/L sucrose, and 0-15 µM of auxins (Table 2) for rhizome proliferation. After 3 months of culture, the rhizomes were transferred to fresh medium. The cultures were maintained at 23 ± 2°C under a 12-hour photoperiod using WLEDs with a PPFD of 10-14 µmol s−1 m−2. Fifteen culture bottles were used for each treatment, with each bottle containing 13 rhizomes. The number and length of the rhizomes were recorded after 6 months of culture.
4.4. Plantlet Regeneration
The rhizomes separated from the clusters grown on MS medium containing 5.0 µM NAA were cut into segments measuring 1.0-1.3 cm long and placed on MS medium supplemented with 500 mg/L AC, 50 mL/L CW, 7 g/L plant agar, 30 g/L sucrose, and 0-20 µM of cytokinins to induce plantlets. The cultures were maintained at 23 ± 2°C under a 12-hour photoperiod, with WLEDs providing a PPFD of 30-34 µmol s−1 m−2. Fifteen culture bottles were used for each treatment, with each containing 10 rhizomes. After 3 months of culturing, the rhizomes were transferred to fresh medium. The percentage of plantlet regeneration, along with the number and height of plantlets, was recorded after 6 months of culturing.
4.5. Acclimatization
The well-developed plantlets (4–11 cm tall) were transplanted into trays containing a mixture of orchid stone and wood bark (50:50), orchid stone, wood bark, coconut husk chip, and perlite (30:30:20:20), or orchid stone, coconut husk chip, coconut fiber, and perlite (50:10:20:20). These were fertigated with a ¼ MS basal salt solution at 2-day intervals and maintained in the polyhouse at 23 ± 2°C under a 16-hour photoperiod at a 200 µmol m−2 s−1 PPFD supplied by a high-pressure sodium lamp (LU400W/PSL, GE lighting, Cleveland, OH) and a relative humidity of 70–80%. After 2 weeks, the plants were fertigated with Hyponex (N-P-K; 20-20-20, Hyponex Japan Crop., Osaka, Japan) solution (2.0 g/L). Three trays were used for each treatment, with each tray containing 75 plantlets. The survival rate was recorded after 2 months. The acclimatized plantlets (3 months old) were transferred into plastic pots containing a mixture of orchid stone and wood bark (50:50) for further growth and development. They were maintained in a shaded polyhouse greenhouse and fertigated with Hyponex solution.
4.6. Statistical Analysis
All experiments were conducted using a completely random design. The data were analyzed with ANOVA, and the means were separated using DMRT at the 5% level in SAS Release 9.4.
5. Conclusions
This study demonstrated that PGRs combined with CW, auxins, cytokinins, and substrates play a crucial role in in vitro seed germination, rhizome proliferation, plantlet regeneration, and plantlet survival. NAA, especially at 5.0–10.0 µM, was the most effective in stimulating rhizome multiplication in terms of both number and length. For plantlet regeneration from rhizomes, BA at 10.0 µM proved to be optimal, promoting the highest regeneration percentage, plantlet number, and plantlet height. Among the tested substrates, a mixture of orchid stone (30%), wood bark (30%), coconut husk chip (20%), and perlite (20%) provided the highest acclimatization success (95.9%) and is recommended for ensuring the efficient transition of in vitro-regenerated plantlets to ex vitro conditions. These findings offer a reliable framework for developing an effective micropropagation protocol for variegated C. lancifolium. The stable and visually distinct variegation patterns observed among in vitro-derived plantlets from a variegated mother plant suggest the potential fixation of variegation traits during plantlet development. This indicates that the variegation may have a genetic or chimeric basis, rather than being solely epigenetic or stress-induced. The uniform expression of central, marginal, or interveinal variegation in individual plantlets enhances their ornamental value and provides a platform for selecting elite lines for commercial propagation. These findings also open new avenues for studying the mechanisms of variegation in orchids and their heritability through sexual and asexual propagation.
Author Contributions
Conceptualization, I.S.; methodology, I.S.; software, I.K.; validation, I.S. and I.K.; formal analysis, I.K.; investigation, I.S.; data curation, I.K.; writing—original draft preparation, I.K. and I.S.; writing—review and editing, I.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data is available within the article.
Acknowledgments
This article was supported by the KU Research Professor Program of Konkuk University.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AC | Activated charcoal |
| CW | Coconut water |
| GA3 | Gibberellic acid |
| IAA | Indole-3-acetic acid |
| IBA | Indole-3-butyric acid |
| KN | Kinetin |
| MS | Murashige and Skoog |
| 2-IP | N6-(2-isopentenyl)adenine |
| BA | N6-benzyladenine |
| PPFD | Photosynthetic photon flux density |
| PGRs | Plant growth regulators |
| WLEDs | White light-emitting diodes |
| NAA | α-naphthalene acetic acid |
References
- Teoh, E.S. Cymbidium Sw. In Orchid Species from Himalaya and Southeast Asia, Teoh, E.S., Ed.; Springer Nature: Switzerland AG, Cham, Sw. In Orchid Species from Himalaya and Southeast Asia, Teoh, E.S., Ed.; Springer Nature: Switzerland AG, Cham, Switzerland, 2021; Volume 1, pp. 265–286. [Google Scholar]
- Park, H.Y.; Kang, K.W.; Kim, D.H.; Sivanesan, I. In vitro propagation of Cymbidium goeringii Reichenbach fil. through direct adventitious shoot regeneration. Physiol. Mol. Biol. Plants 2018, 24, 307–313. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, H.J.; Kim, K.S. Night interruption promotes vegetative growth and flowering of Cymbidium. Sci. Hortic. 2011, 130, 887–893. [Google Scholar] [CrossRef]
- De, L.C.; Singh, R. Organic Production of Cymbidium Orchids. Acta Sci. Agric. 2018, 2, 30–35. [Google Scholar]
- Lee, J.S.; Choe, B.H. Distributions and red data of wild orchids in the Korean peninsula. Korean J. Plant Taxon. 2006, 36, 335–360. [Google Scholar] [CrossRef]
- Balilashaki, K.; Martinez-Montero, M.E.; Vahedi, M.; Cardoso, J.C.; Silva Agurto, C.L.; Leiva-Mora, M.; Feizi, F.; Musharof Hossain, M. Medicinal Use, Flower Trade, Preservation and Mass Propagation Techniques of Cymbidium Orchids—An Overview. Horticulturae 2023, 9, 690. [Google Scholar] [CrossRef]
- Hussien, M.; Kryuchkova, V.; Raeva-Bogoslovskaya, E.; Molkanova, O. Clonal Micropropagation of Cymbidium erythrostylum Rolfe. Int. J. Plant Biol. 2023, 14, 28–38. [Google Scholar] [CrossRef]
- Yao, M.; Wu, C.; Huang, W.; Fang, Z. Plant Growth Regulator-based Tissue Culture System Optimization for Cymbidium faberi Rolfe. HortScience 2024, 59, 1358–1368. [Google Scholar] [CrossRef]
- Monica, H.; Kumaria, S. Exogenous application of chitosan, a potent biotic elicitor enhances micropropagation efficiency of Cymbidium aloifolium (L.) Sw., an orchid of medicinal and horticultural importance. Vegetos 2025, 38, 58–69. [Google Scholar] [CrossRef]
- Li, F.; Bao, J.; Sun, Y.; Liu, C.; Ma, H.; Zhang, T.; Chen, X. Embryo development and corresponding factors affecting in vitro germination of Cymbidium faberi × C. sinense hybrid seeds. Arch. Biol. Sci. 2016, 68, 541–550. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sharma, M.; da Silva, J.A.T.; Pathak, P. Seed germination and tissue culture of Cymbidium giganteum Wall. ex Lindl. Sci. Hortic. 2010, 123, 479–487. [Google Scholar] [CrossRef]
- Pradha, S.; Pant, B. In vitro seed germination in Cymbidium elegans Lindl. and Dendrobium densiflorum Lindl. ex Wall. (Orchidaceae). Botanica Orientalis. J. Plant Sci. 2010, 6, 100–102. [Google Scholar] [CrossRef]
- Pant, B.; Swar, S. Micropropagation of Cymbidium iridioides. Nepal J. Sci. Technol. 2011, 12, 91–96. [Google Scholar] [CrossRef]
- Kumar, A.; Chauhan, S.; Rattan, S.; Warghat, A.R.; Kumar, D.; Bhargava, B. In vitro propagation and phyto-chemical assessment of Cymbidium aloifolium (L.) Sw.: An orchid of pharma-horticultural importance. S. Afr. J. Bot. 2022, 144, 261–269. [Google Scholar] [CrossRef]
- Pradhan, S.; Regmi, T.; Parmar, G.; Pant, B. Effect of different media on in vitro seed germination and seedling development of Cymbidium aloifolium (L.) Sw. Nepal J. Sci. Technol. 2013, 14, 51–56. [Google Scholar] [CrossRef]
- Tawara, S.; Suraninpong, P.; Chanprame, S. Germination and Regeneration of Cymbidium findlaysonianum Lindl. on a medium supplemented with some organic sources. Walailak J. Sci. Technol. 2008, 5, 125–135. [Google Scholar]
- Fang, Z.; Huang, W.; Zeng, S.; Wu, K. In vitro propagation of Cymbidium nanutum YS Wu et SC Chen. Propag. Ornam. Plants 2011, 11, 149–155. [Google Scholar]
- Kim, J.Y.; Lee, J.S. Effect of growth regulators on rhizome growth and differentiation of Cymbidium lancifolium, a native Korean bamboo orchid. Korean Soc. Hortic. Sci. Acad. Pres. Summ. 1991, 9(1), 174–175. [Google Scholar]
- Kim, J.Y.; Lee, J.S. Effect of cultural conditions on rhizome growth and organogenesis of Cymbidium lancifolium native to Korea in vitro. J. Korean Soc. Hortic. Sci. 1992, 33, 471–476. [Google Scholar]
- Yeung, E.C. A perspective on orchid seed and protocorm development. Bot. Stud. 2017, 58, 33. [Google Scholar] [CrossRef]
- Tikendra, L.; Singh, A.R.; Vendrame, W.A.; Nongdam, P. In Vitro Propagation of Endangered Vanda coerulea Griff. ex Lindl.: Asymbiotic Seed Germination, Genetic Homogeneity Assessment, and Micro-Morpho-Anatomical Analysis for Effective Conservation. Agronomy 2025, 15, 1195. [Google Scholar] [CrossRef]
- Paek, K.Y.; Murthy, H.N. Temperate oriental Cymbidium species. In Orchid biology reviews and perspectives, Kull, T., Arditti, J., Eds.; Kluwer Academic Publishers: Dordrecht, Netherlands, 2002; Volume 8. [Google Scholar]
- Li, Y.; Chen, H.; Kong, X.; Yin, Y.; Li, J.; Wu, K.; Zeng, S.; Fang, L. Excessive accumulation of auxin inhibits protocorm development during germination of Paphiopedilum spicerianum. Plant Cell Rep. 2025, 44, 23. [Google Scholar] [CrossRef] [PubMed]
- Deb, C.R.; Pongener, A. Asymbiotic seed germination and in vitro seedling development of Cymbidium aloifolium (L.) Sw: A multipurpose orchid. J. Plant Biochem. Biotechnol. 2011, 20, 90–95. [Google Scholar] [CrossRef]
- Utami, E.S.W.; Hariyanto, S. Organic compounds: Contents and their role in improving seed germination and protocorm development in orchids. Int. J. Agron. 2020, 2020, 2795108. [Google Scholar] [CrossRef]
- Anghelescu, N.E.; Vafaee, Y.; Ahmadzadeh, K.; Chen, J.T. Asymbiotic Seed Germination in Terrestrial Orchids: Problems, Progress, and Prospects. In: Advances in Orchid Biology Biotechnology and Omics, Tiwari, P., Chen, J.T., Eds.; Springer, Singapore, 2023, pp 221–260.
- Tan, S.N.; Yong, J.W.H.; Ge, L. Analyses of Phytohormones in Coconut (Cocos nucifera L.) Water Using Capillary Electrophoresis-Tandem Mass Spectrometry. Chromatography 2014, 1, 211–226. [Google Scholar] [CrossRef]
- Huh, Y.S.; Lee, J.K.; Nam, S.Y.; Paek, K.Y.; Suh, G.U. Improvement of asymbiotic seed germination and seedling development of Cypripedium macranthos Sw. with organic additives. J. Plant Biotechnol. 2016, 43, 138–145. [Google Scholar] [CrossRef]
- Zhang, Y.; Kan, J.; Liu, X.; Song, F.; Zhu, K.; Li, N.; Zhang, Y. Chemical Components, Nutritional Value, Volatile Organic Compounds and Biological Activities In Vitro of Coconut (Cocos nucifera L.) Water with Different Maturities. Foods 2024, 13, 863. [Google Scholar] [CrossRef]
- Rohmah, K.N.; Taratima, W. Effect of chitosan, coconut water and potato extract on protocorm growth and plantlet regeneration of Cymbidium aloifolium (L.) Sw. Curr. Appl. Sci. Technol. 2022, 22, 1–10. [Google Scholar]
- Parmar, G. In vitro seed germination and seedling development of Cymbidium devonianum Paxton (Orchidaceae). Bull. Dept. Plant Res. 2014, 36, 61–64. [Google Scholar]
- Kim, D.H.; Kang, K.W.; Sivanesan, I. In vitro propagation of Cymbidium hybrid. Propag. Ornam. Plants 2017, 17, 48–54. [Google Scholar]
- Paul, M.; Islam, T.; Sarker, R.H.; Hoque, M.I. In vitro mass propagation of Cymbidium aloifolium (L.) Sw. Plant Tissue Cult. Biotechnol. 2019, 29, 73–79. [Google Scholar] [CrossRef]
- Tao, J.; Yu, L.; Kong, F.; Zhao, D. Effects of plant growth regulators on in vitro propagation of Cymbidium faberi Rolfe. Afr. J. Biotechnol. 2011, 10, 15639–15646. [Google Scholar] [CrossRef]
- Gogoi, K.; Kumaria, S.; Tandon, P. Ex situ conservation of Cymbidium eburneum Lindl.: A threatened and vulnerable orchid, by asymbiotic seed germination. 3 Biotech. 2012, 2, 337–343. [Google Scholar] [CrossRef]
- Nyorak, J.; Sonar, C.B. In vitro mass multiplication of Cymbidium iridioides D. Don–a medicinal orchid from Arunachal Pradesh, India. Pleione 2020, 14, 49–55. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Z.; Huang, L.; Su, J. In Vitro Mass Scale Propagation of Wild Cymbidium lowianum with a Rare and Endangered Plant. Am. J. Plant Sci. 2013, 4, 34755. [Google Scholar] [CrossRef]
- Kang, T.J.; Yang, D.C. Days to germination and effect of growth regulator on rhizome growth in Cymbidium goeringii Hybrid. Korean J. Plant Res. 2003, 6, 144–148. [Google Scholar]
- Teixeira da Silva, J.A.; Tsavkelova, E.A.; Ng, T.B.; Parthibhan, S.; Dobránszki, J.; Cardoso, J.C.; Rao, M.V.; Zeng, S. Asymbiotic in vitro seed propagation of Dendrobium. Plant Cell Rep. 2015, 34, 1685–1706. [Google Scholar] [CrossRef]
- Thummavongsa, T.; Musimun, C.; Watthana, S.; Gale, S.; Choeyklin, R.; Wiriyathanawudhiwong, N.; Muangsan, N. Enhancing Germination of Habenaria janellehayneana (Orchidaceae): Insight from Asymbiotic and Symbiotic Methods. J. Ornam. Plants 2024, 14, 11–23. [Google Scholar]
- Manrique, J.P.; Fernandex-Lizarazo, C.; Suarez-Silva, A. Evaluation of the effect of three growth regulators in the germination of Comparetia falcate seeds under in vitro conditions. In Vitro Cell. Dev. Biol. Plant 2005, 41, 838–843. [Google Scholar] [CrossRef]
- Kang, H.; Kang, K.W.; Kim, D.H.; Sivanesan, I. In Vitro Propagation of Gastrochilus matsuran (Makino) Schltr., an Endangered Epiphytic Orchid. Plants 2020, 9, 524. [Google Scholar] [CrossRef]
- Paek, K.Y.; Kozai, T. Micropropagation of temperate Cymbidium via rhizome culture. HortTechnology 1998, 8, 283–288. [Google Scholar] [CrossRef]
- Nayak, N.R.; Rath, S.P.; Patnaik, S. In vitro propagation of three epiphytic orchids, Cymbidium aloifolium (L.) Sw., Dendrobium aphyllum (Roxb.), Fisch. and Dendrobium moschatum (Buch-Ham) Sw. through thidiazuron-induced high frequency shoot proliferation. Sci. Hortic. 1997, 71, 243–250. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, X.; Liu, Y. In vitro plant regeneration from the immature seeds of Cymbidium faberi. Plant Cell Tissue Organ Cult. 2005, 81, 247–251. [Google Scholar] [CrossRef]
- Paek, K.Y.; Yeung, E.C. The effects of 1-naphthaleneacetic acid and N6-benzyladenine on the growth of Cymbidium forrestii rhizomes in vitro. Plant. Cell Tissue Organ. Cult. 1991, 24, 65–71. [Google Scholar] [CrossRef]
- Huang, C. L.; Okubo, H. In vitro morphogenesis from rhizomes of Cymbidium sinense. J. Fac. Agr., Kyushu Univ. 2005, 50, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, K.; Uemoto, S. Micropropagation of a terrestrial Cymbidium species using rhizomes developed from seeds and pseudobulbs. Plant Cell Tissue Organ Cult. 1990, 22, 237–244. [Google Scholar] [CrossRef]
- Nayak, N.R.; Chand, P.K.; Rath, S.P.; Patnaik, S. Influence of some plant growth regulators on the growth and organogenesis of Cymbidium aloifolium (L.) Sw. seed derived rhizomes in vitro. In Vitro Cell. Dev. Biol. Plant 1998, 34, 185–188. [Google Scholar] [CrossRef]
Figure 1.
In vitro propagation of variegated C. lancifolium. (A) self-pollinated flowering plant of variegated C. lancifolium; (B) self-pollinated plant of variegated C. lancifolium developing mature seed capsules; (C) longitudinally cut seed capsule showing numerous seeds; (D) close-up view of mature seeds containing embryos; (E) formation of swollen seeds and protocorms; (F) seeds developing white and green protocorms; (G) formation of rhizomes from the protocorms; (H-J) various stages of rhizome growth and proliferation; (K) plantlets regenerated from the rhizome displaying prominent marginal and interveinal variegation; (L) regeneration of light green plantlets; (M) regeneration of yellow-green plantlets; (N) regeneration of dark green plantlets; (O) well-developed light green plantlets; (P) plantlets exhibiting central striped variegation; (Q) plantlets showing a combination of central and marginal variegation; (R) plantlets displaying sectorial and mild marginal variegation; (S) plantlets acclimatized in medium consisting of orchid stone (30%), wood bark (30%), coconut husk chip (20%), and perlite (20%); (T) plantlets acclimatized in medium consisting of orchid stone (50%) and wood bark (50%); (U) plantlets acclimatized in medium consisting of orchid stone (50%), coconut husk chip (10%), coconut fiber (20%), and perlite (20%); (V) acclimatized plantlet transferred into a pot containing orchid stone (50%) and wood bark (50%). Bar: (A-C, F-V) 1 cm; (D,E) 0.5 mm.
Figure 1.
In vitro propagation of variegated C. lancifolium. (A) self-pollinated flowering plant of variegated C. lancifolium; (B) self-pollinated plant of variegated C. lancifolium developing mature seed capsules; (C) longitudinally cut seed capsule showing numerous seeds; (D) close-up view of mature seeds containing embryos; (E) formation of swollen seeds and protocorms; (F) seeds developing white and green protocorms; (G) formation of rhizomes from the protocorms; (H-J) various stages of rhizome growth and proliferation; (K) plantlets regenerated from the rhizome displaying prominent marginal and interveinal variegation; (L) regeneration of light green plantlets; (M) regeneration of yellow-green plantlets; (N) regeneration of dark green plantlets; (O) well-developed light green plantlets; (P) plantlets exhibiting central striped variegation; (Q) plantlets showing a combination of central and marginal variegation; (R) plantlets displaying sectorial and mild marginal variegation; (S) plantlets acclimatized in medium consisting of orchid stone (30%), wood bark (30%), coconut husk chip (20%), and perlite (20%); (T) plantlets acclimatized in medium consisting of orchid stone (50%) and wood bark (50%); (U) plantlets acclimatized in medium consisting of orchid stone (50%), coconut husk chip (10%), coconut fiber (20%), and perlite (20%); (V) acclimatized plantlet transferred into a pot containing orchid stone (50%) and wood bark (50%). Bar: (A-C, F-V) 1 cm; (D,E) 0.5 mm.

Figure 2.
Effect of auxin type (A) and concentration (B) on rhizome growth and proliferation. Different letters show significant differences by DMRT at p<0.5.
Figure 2.
Effect of auxin type (A) and concentration (B) on rhizome growth and proliferation. Different letters show significant differences by DMRT at p<0.5.
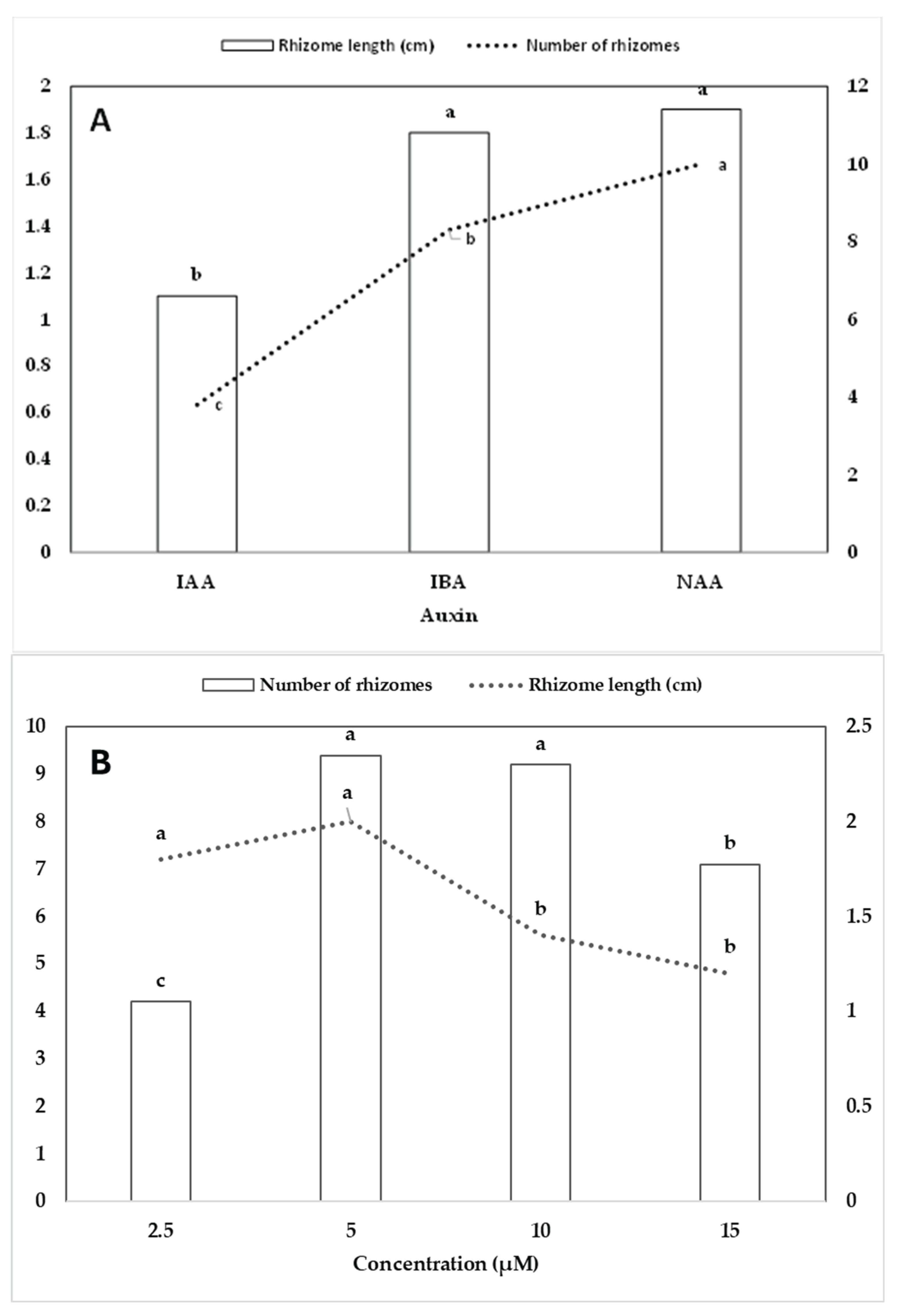
Figure 3.
Effect of cytokinin type (A) and concentration (B) on plantlet regeneration (%), number of plantlets and plantlet height (cm). Different letters show significant differences by DMRT at p<0.5.
Figure 3.
Effect of cytokinin type (A) and concentration (B) on plantlet regeneration (%), number of plantlets and plantlet height (cm). Different letters show significant differences by DMRT at p<0.5.
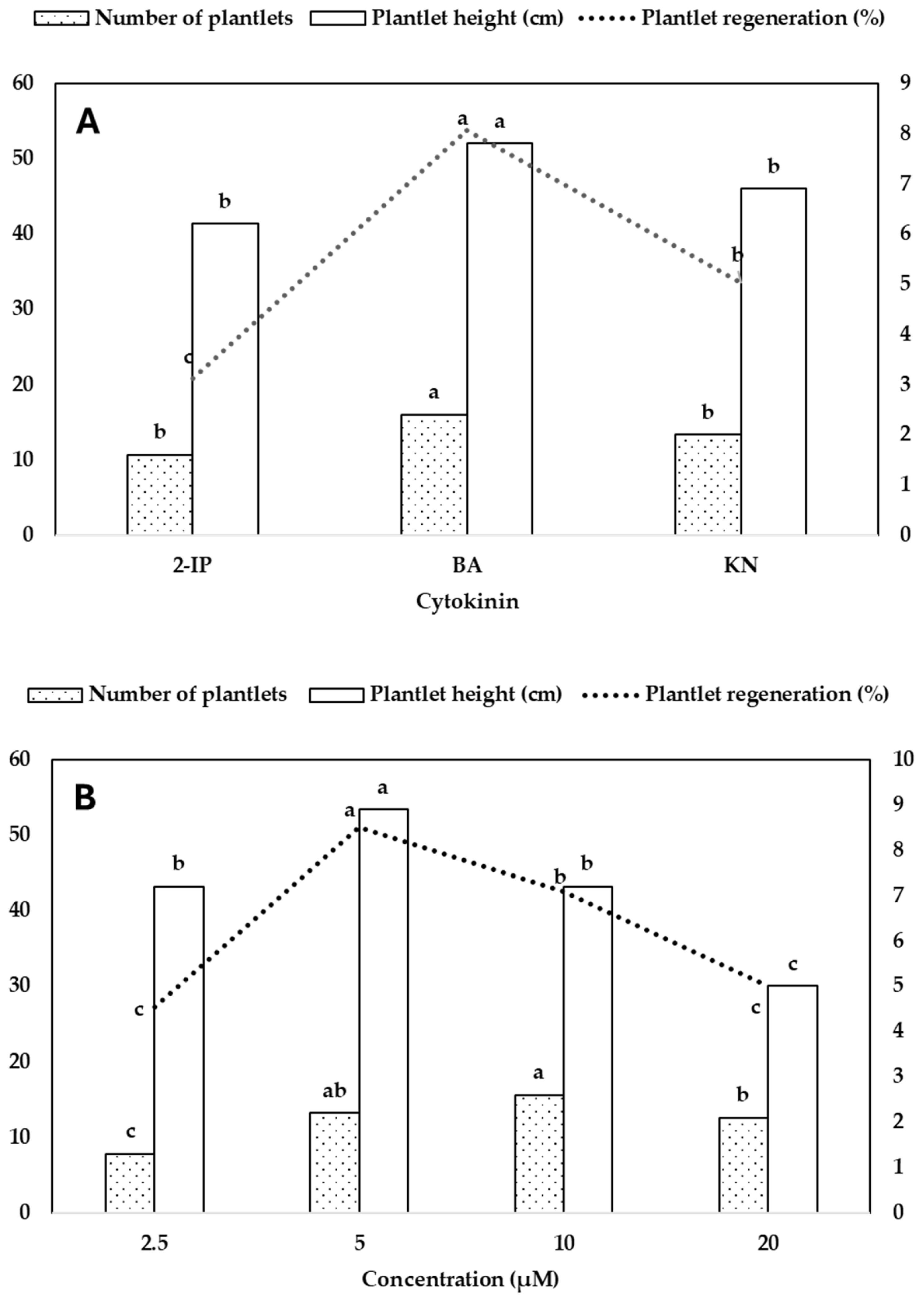
Figure 4.
Effect of growing media on the survival rate of variegated C. lancifolium plantlets during acclimatization. Different letters show significant differences by DMRT at p<0.5.
Figure 4.
Effect of growing media on the survival rate of variegated C. lancifolium plantlets during acclimatization. Different letters show significant differences by DMRT at p<0.5.
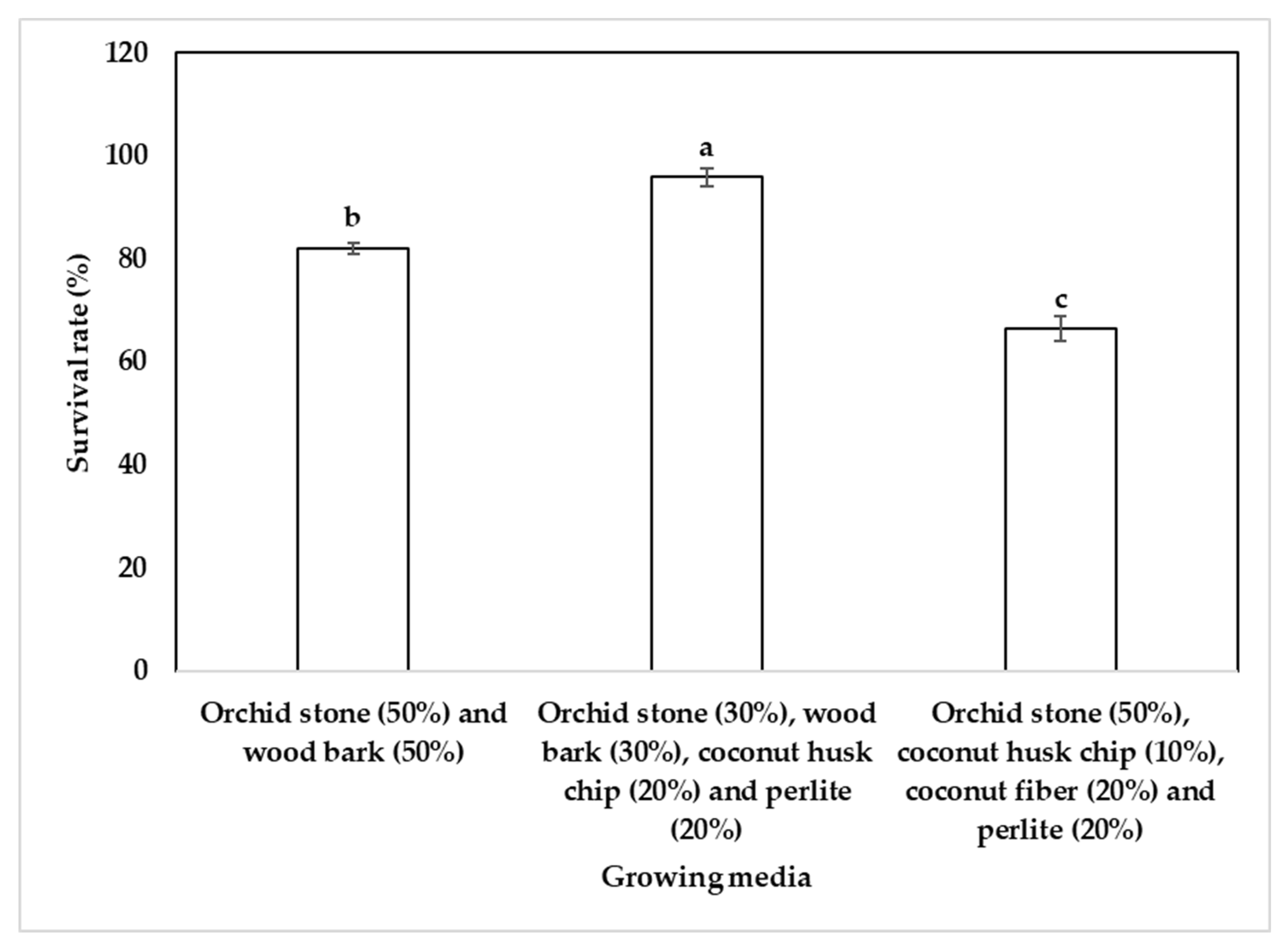

Table 1.
Effect of PGRs and CW on asymbiotic seed germination in variegated C. lancifolium.
| PGRs (µM) | Coconut water (mL/L) | Seed germination (%) | |||
| IAA | NAA | KN | GA3 | ||
| 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.2 ± 0.1 l |
| 0.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.7 ± 0.3 l |
| 1.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.2 ± 0.1 kl |
| 2.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.4 ± 0.5 jk |
| 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.3 ± 1.2 ij |
| 8.0 | 0.0 | 0.0 | 0.0 | 0.0 | 10.2 ± 1.1 hi |
| 0.0 | 0.5 | 0.0 | 0.0 | 0.0 | 3.0 ± 0.4 kl |
| 0.0 | 1.0 | 0.0 | 0.0 | 0.0 | 6.2 ± 0.8 ijk |
| 0.0 | 2.0 | 0.0 | 0.0 | 0.0 | 8.9 ± 1.1 hij |
| 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 16.8 ± 1.5 f |
| 0.0 | 8.0 | 0.0 | 0.0 | 0.0 | 12.6 ± 1.3 gh |
| 0.0 | 4.0 | 0.0 | 0.0 | 25 | 18.1 ± 1.1 ef |
| 0.0 | 4.0 | 0.0 | 0.0 | 50 | 26.2 ± 2.0 d |
| 0.0 | 4.0 | 0.0 | 0.0 | 75 | 20.9 ± 1.6 e |
| 0.0 | 4.0 | 0.0 | 0.0 | 100 | 14.3 ± 1.3 fg |
| 0.0 | 4.0 | 2.3 | 0.0 | 50 | 32.2 ± 1.7 c |
| 0.0 | 4.0 | 4.7 | 0.0 | 50 | 27.8 ± 1.5 d |
| 0.0 | 4.0 | 9.4 | 0.0 | 50 | 28.3 ± 1.7 d |
| 0.0 | 4.0 | 2.3 | 0.3 | 50 | 33.3 ± 2.0 c |
| 0.0 | 4.0 | 2.3 | 1.4 | 50 | 40.6 ± 1.8 b |
| 0.0 | 4.0 | 2.3 | 2.9 | 50 | 46.8 ± 1.8 a |
Means ± SE followed by different letters(a-l) are significantly different according to Duncan’s multiple range test (DMRT) at p<0.5.
Table 2.
Effect of auxins on growth and proliferation of rhizomes of C. lancifolium.
| Auxin (µM) | Number of rhizomes | Rhizome length (cm) | ||
| IAA | IBA | NAA | ||
| 0.0 | 0.0 | 0.0 | 1.3 ± 0.2 h | 0.5 ± 0.1 f |
| 2.5 | 0.0 | 0.0 | 1.9 ± 0.4 gh | 1.0 ± 0.1 ef |
| 5.0 | 0.0 | 0.0 | 2.8 ± 0.4 gh | 1.4 ± 0.2 cde |
| 10.0 | 0.0 | 0.0 | 4.1 ± 0.6 fg | 1.0 ± 0.1 ef |
| 15.0 | 0.0 | 0.0 | 6.2 ± 1.0 ef | 1.1 ± 0.2 de |
| 0.0 | 2.5 | 0.0 | 3.0 ± 0.6 gh | 2.0 ± 0.2 ab |
| 0.0 | 5.0 | 0.0 | 8.0 ± 1.0 de | 2.5 ± 0.1 a |
| 0.0 | 10.0 | 0.0 | 11.0 ± 1.0 bc | 1.7 ± 0.2 bc |
| 0.0 | 15.0 | 0.0 | 5.8 ± 0.7 ef | 1.2 ± 0.2 cde |
| 0.0 | 0.0 | 2.5 | 7.8 ± 0.8 de | 2.4 ± 0.2 a |
| 0.0 | 0.0 | 5.0 | 17.4 ± 0.9 a | 2.1 ± 0.2 ab |
| 0.0 | 0.0 | 10.0 | 12.4 ± 1.0 b | 1.6 ± 0.2 bcd |
| 0.0 | 0.0 | 15.0 | 9.4 ± 0.9 cd | 1.3 ± 0.2 cde |
| ANOVA | R-Square | 0.748 | 0.491 | |
| Coefficient of variation | 34.86 | 32.94 | ||
| Auxin | p<0.0001 | p<0.0001 | ||
| Concentration | p<0.0001 | p<0.0001 | ||
| Auxin*Concentration | p<0.0001 | p<0.0208 | ||
Means ± SE followed by different letters(a-h) are significantly different according to DMRT at p<0.5.
Table 3.
Effect of cytokinins on inducing plantlets from C. lancifolium rhizomes obtained through asymbiotic seed germination.
Table 3.
Effect of cytokinins on inducing plantlets from C. lancifolium rhizomes obtained through asymbiotic seed germination.
| Cytokinin (µM) | Plantlet regeneration (%) | Number of plantlets per rhizome culture |
Plantlet height (cm) |
||
| 2-IP | BA | KN | |||
| 0.0 | 0.0 | 0.0 | 0.0 ± 0.0 j | 0.0 ± 0.0 g | 0.0 ± 0.0 1 |
| 2.5 | 0.0 | 0.0 | 21.7 ± 2.0 fg | 1.1 ± 0.1 f | 5.9 ± 0.6 e-h |
| 5.0 | 0.0 | 0.0 | 38.9 ± 1.6 e | 2.1 ± 0.3 b-e | 7.0 ± 0.4 c-e |
| 10.0 | 0.0 | 0.0 | 13.9 ± 1.6 hi | 1.7 ± 0.2 c-f | 6.3 ± 0.6 e-g |
| 20.0 | 0.0 | 0.0 | 8.9 ± 1.4 i | 1.6 ± 0.2 c-f | 5.5 ± 0.5 f-h |
| 0.0 | 2.5 | 0.0 | 33.9 ± 3.2 e | 1.3 ± 0.2 ef | 7.6 ± 0.5 cde |
| 0.0 | 5.0 | 0.0 | 67.8 ± 3.0 b | 2.3 ± 0.2 bc | 10.2 ± 0.9 a |
| 0.0 | 10.0 | 0.0 | 79.4 ± 1.8 a | 4.4 ± 0.5 a | 8.5 ± 0.5 a-c |
| 0.0 | 20.0 | 0.0 | 61.7 ± 2.0 c | 2.7 ± 0.3 b | 4.9 ± 0.5 gh |
| 0.0 | 0.0 | 2.5 | 26.1 ± 2.3 f | 1.4 ± 0.2 def | 7.9 ± 0.7 bcd |
| 0.0 | 0.0 | 5.0 | 46.7 ± 2.4 d | 2.2 ± 0.4 bcd | 9.5 ± 0.8 ab |
| 0.0 | 0.0 | 10.0 | 34.4 ± 1.5 e | 1.8 ± 0.3 c-f | 6.7 ± 0.5 d-g |
| 0.0 | 0.0 | 20.0 | 19.4 ± 2.1 gh | 2.0 ± 0.3 b-e | 4.4 ± 0.3 h |
| ANOVA | R-Square | 0.919 | 0.537 | 0.503 | |
| Coefficient of variation | 17.62 | 40.01 | 25.37 | ||
| Cytokinin | p<0.0001 | p<0.0001 | p<0.0003 | ||
| Concentration | p<0.0001 | p<0.0001 | p<0.0001 | ||
| Cytokinin*Concentration | p<0.0001 | p<0.0001 | p<0.0171 | ||
Means ± SE followed by different letters(a-l) are significantly different according to DMRT at p<0.5.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



