Submitted:
23 June 2025
Posted:
26 June 2025
You are already at the latest version
Abstract
Zebrafish are model organisms for drug screening owing to their transparent bodies, rapid embryonic development, and genetic similarities with humans. However, using standard polystyrene culture plates can limit the oxygen supply, potentially affecting embryo survival and the reliability of assays conducted in zebrafish. In this study, we evaluated the application of a novel, highly oxygen-permeable culture plate (InnoCellTM) in zebrafish development and drug screening assays. Under both normal and oxygen-restricted conditions, zebrafish embryos cultured on InnoCellTM plates exhibited significantly improved developmental parameters, including heart rate and body length, compared with those cultured on conventional polystyrene plates. The InnoCellTM plate enabled a significant reduction in medium volume without compromising zebrafish embryo viability, thereby demonstrating its advantages, particularly in high-throughput 384-well formats. Drug screening tests using antiangiogenic receptor tyrosine kinase inhibitors (TKIs) revealed enhanced sensitivity and more pronounced biological effects in InnoCellTM plates, as evidenced by quantification of intersegmental blood vessels and gene expression analysis of the vascular endothelial growth factor receptor (kdrl). These results indicate that the InnoCellTM high oxygen-permeable plate markedly improves zebrafish-based drug screening efficiency and assay reliability, highlighting its potential for widespread application in biomedical research.
Keywords:
zebrafish
; drug screening
; oxygen permeability
; high-throughput
; culture plate
; angiogenesis inhibitors
1. Introduction
Cell culture and drug screening are indispensable tools in the research domains of biology, physiology, pharmacology, toxicology, and medicine. Cell culture involves techniques for cultivating cells outside their natural habitats, under controlled and appropriate conditions. In basic scientific research, cell cultures are typically employed to analyze cellular structure and function, elucidate the mechanisms underlying cell proliferation and differentiation, and investigate the molecular pathways that regulate gene and protein expression. Cell cultures are crucial in pathological research, regenerative medicine, personalized therapies, and safety evaluations, directly affecting human health [1,2]. Drug screening is a crucial step in drug development, used to efficiently assess a wide range of compounds to identify those that specifically interact with target receptors or other biological entities. This process is vital for the discovery of new therapeutic agents, the evaluation of pharmacological effectiveness, and the assessment of toxicity. By employing cultured cells and model organisms, drug screening substantially contributes to the creation of safe and effective pharmaceuticals [3,4].
Sterile polystyrene (PS) plates are commonly used in cell culture drug screening owing to their convenience. However, cells in culture can swiftly generate hypoxic conditions in vitro, with a notable reduction in dissolved oxygen levels occurring within 1 h of either seeding or medium replacement [5]. An adequate supply of oxygen is essential for the metabolism and function of almost all cell types. For example, hepatocytes, especially primary hepatocytes, require relatively high levels of oxygen to sustain a physiological biomimetic environment [6]. Therefore, the gap between cell culture conditions and the actual physiological settings hinders biotechnological research. An uncomplicated yet efficient approach has been introduced to enhance oxygen delivery by incorporating oxygen-permeable substances, such as polydimethylsiloxane (PDMS) or 4-polymethyl-1-pentene polymer (PMP) at the base of culture plates. PDMS, an organic polymer derived from silicone, is recognized for its optical clarity and ability to allow gas passage. Its high oxygen permeability makes it a popular choice for cell culture and serves as an alternative to traditional PS-bottom surfaces [6,7]. Nevertheless, a major drawback is that hydrophobic molecules can be adsorbed on both the surface and within the bulk of PDMS [8]. Nile red has been shown to infiltrate PDMS, leading to an increase in fluorescence over time [9]. Moreover, PMP is increasingly utilized in cell culture applications owing to its excellent oxygen permeability, biocompatibility, chemical stability, and transparency [10]. Mitsui Chemicals, Inc. developed a next-generation cell culture solution called the InnoCellTM T-plate (hereafter referred to as the InnoCellTM plate). This plate features a base constructed from PMP, which is known for its excellent oxygen permeability and release properties. Comparing the oxygen consumption rates of primary rat hepatocytes cultured on InnoCell™ plates with those on conventional PS-bottom plates, PMP was found to exhibit an oxygen permeability coefficient up to 15 times greater than that of PS [11]. This corresponds to a 190-fold increase in the theoretical oxygen supply, calculated as the product of the bottom thickness and oxygen permeability coefficient. Additionally, the InnoCellTM plate exhibits low levels of chemical sorption [11]. The InnoCellTM plate-induced environment can also boost cytochrome P450 activity and enhance mitochondrial function. Therefore, this cutting-edge plate is considered a promising option for various drug-testing applications.
Zebrafish (Danio rerio) is an ideal vertebrate model extensively used in traditional developmental biology and has recently gained prominence in human disease research [12,13,14]. Zebrafish are effective tools for drug screening owing to their distinct biological and practical benefits. As small tropical freshwater fish, zebrafish are genetically manageable and share approximately 70% of their genome with humans, including orthologs for approximately 84% of the genes associated with human diseases. Zebrafish embryos develop rapidly and are optically transparent, enabling real-time, noninvasive observation of internal organs and drug-induced phenotypic changes. Zebrafish can be bred in large numbers and maintained at a low cost, making them well-suited for high-throughput screening in multi-well plate formats. Notably, their functional organ systems, including cardiovascular, nervous, immune, and metabolic systems, are similar to those of mammals. This similarity enables the efficient assessment of drug effectiveness and toxicity, with substantial translational relevance. Because the early embryonic stages are not heavily regulated by animal welfare laws, rapid testing of compounds can be performed with fewer ethical concerns. These characteristics highlight the potential of zebrafish as a highly efficient, cost-effective, and predictive model for drug discovery and biomedical research [15,16,17].
In the current study, we aimed to evaluate the application of InnoCellTM plates in zebrafish development and drug screening. We first assessed the impact of the high oxygen permeability of the plate on zebrafish embryos under both normal and oxygen-deprived conditions and compared the results with those obtained using conventional PS plates. Furthermore, we investigated the minimum culture medium volume required in 96- and 384-well plates to support normal development and effective drug activity.
2. Materials and Methods
2.1. Ethics Statement
Animal handling was conducted in accordance with Japan's Act on Welfare and Management of Animals, established by the Ministry of the Environment of Japan, and adhered to international standards.
2.2. Zebrafish Maintenance and Embryo Collection
Wild-type AB zebrafish were obtained from the Zebrafish International Research Center (ZFIN; Eugene, OR, USA). The transgenic strain Tg(kdrl:EGFP) was generously provided by Professor Stefan Schulte-Merker (WWU Münster) [18]. Adult zebrafish were housed and nurtured in a cultivation system set to 28 ± 0.5°C, with a 14/10 h light/dark cycle, following the standard ZFIN protocols (https://zfin.atlassian.net/wiki/spaces/prot/overview; accessed on April 8, 2025). Fish were provided GEMMA Micro 75, 150, or 300 (Skretting, Fontaine-les-Vervins, France) twice daily, tailored to their developmental stages and sizes.
To collect embryos, adult zebrafish were placed in a spawning tank with a female-to-male ratio of 3:2 one day prior to mating. Following natural spawning, embryos were gathered and incubated in 0.3 × Danieau’s solution (17.4 mM NaCl, 0.21 mM KCl, 0.12 mM MgSO4, 0.18 mM Ca(NO3)2, and 1.5 mM 4-(2-hydroxyethyl) 1-piperazinyl-ethane-2-sulfonic acid [HEPES]; pH 7.6). Healthy fertilized embryos were selected at the shield stage (6 hours post-fertilization [hpf]) using a stereomicroscope (Nikon SMZ745T; Tokyo, Japan).
2.3. Zebrafish Embryo Culture on Plates
PS control culture plates, available in 24-, 96-, and 384-well configurations, were obtained from Corning Scientific Products, Inc. (Corning, NY, USA). InnoCellTM plates were supplied by Mitsui Chemicals, Inc., Tokyo, Japan, and are distinguished by the attachment of 50 μm PMP sheets at the base to enhance oxygen permeability.
2.3.1. Impact of Fish Density and Oxygen Conditions on Development in 24-Well Plates
Zebrafish embryos at 6 hpf were selected and placed in each well at densities of 1, 5, 10, 20, 30, and 50 fish per well per milliliter of medium. To assess how the oxygen permeability of the InnoCellTM plate influences zebrafish development, two distinct conditions were established: one with a standard plate lid, referred to as the "No sealing" condition, and the other with plate seals to prevent oxygen exchange from the top of the well, termed the "Sealing" condition. Embryos were incubated at 28°C for 72 h.
2.3.2. Identify the minimum medium volume for 96- and 384-well plates
A standard control setup was implemented, consisting of 25 embryos per 100 mL of culture medium, consistent with standard zebrafish culture guidelines [19]. For the 96-well plate, healthy embryos were placed in medium volumes of 10, 15, 20, 30, 40, 50, 100, and 200 μL/well/fish. For the 384-well plate, embryos were cultured in medium volumes of 8, 10, 15, 20, 30, 40, 60, and 80 μL/well/fish. Both the "No sealing" and "Sealing" conditions were employed, as described previously. The experiment was conducted over a period ranging from 6 to 72 hpf at a stable temperature of 28°C.
2.4. Assessment of Developmental Effects
Heart rate was recorded at 56 hpf using a stereomicroscope, and both survival rates and morphological changes were assessed at 72 hpf. Additionally, zebrafish larvae were imaged under a stereomicroscope at 72 hpf. Body length was measured from the anterior tip of the snout to the posterior end of the notochord using the ImageJ software (National Institutes of Health, Bethesda, USA).
2.5. Drug Screening Assay
Tyrosine kinase inhibitors (TKIs), including SU4312 (CAS#: 5812-07-7; Cayman Chemical, Ann Arbor, MI, USA), sorafenib (CAS#: 284461-73-0; Sigma-Aldrich, St. Louis, MO, USA), axitinib (CAS#: 319460-85-0; Selleck Chemicals, Houston, TX, USA), and valproic acid (VPA; CAS#: 99-66-1; TCI Chemicals, Tokyo, Japan), were dissolved in 100% dimethyl-sulfoxide (DMSO) to prepare the stock solution and diluted in 0.3 × Danieau’s solution culture medium.
For chemical exposure, Tg(kdrl:EGFP) embryos at 24 hpf underwent dechorionation by treatment with protease (Roche Diagnostics, Indianapolis, IN, USA) at a concentration of 1 mg/mL for 5 min. Thereafter, the embryos were allocated to 96-well culture plates, with each well containing either 40 or 100 μL per embryo under no sealing conditions, and 20 or 100 μL per embryo under sealing conditions. In 384-well plates, embryos were distributed with 20 and 80 μL per embryo per well under no sealing conditions, and 15 and 80 μL per embryo per well under sealing conditions. The embryos were then exposed to the vehicle (0.1% DMSO) and various TKIs. The final concentrations for SU4312, sorafenib, axitinib, and VPA were set at 6, 1.5, 1, and 5 μM, respectively. After a 3-day treatment period, the embryos were anesthetized using 500 ppm 2-phenoxyethanol (Wako Pure Chemicals, Osaka, Japan), and fluorescence images were captured using a BZ-X710 fluorescence microscope (Keyence, Tokyo, Japan). The number of intersegmental blood vessels (ISVs) was then counted. Each experiment was conducted with at least two replicates on different days and during breeding sessions.
2.6. Real-Time Quantitative PCR (qPCR)
Following chemical exposure, zebrafish larvae were collected at 96 hpf and homogenized using beads. Total RNA was isolated and purified using TRIzol reagent (Life Technologies, Carlsbad, CA, USA) and the QIAGEN RNeasy Mini-prep Kit (Qiagen, Hilden, Germany). The total RNA concentration was determined using a spectrophotometer (BioPhotometer, Eppendorf, Hamburg, Germany), and cDNA was synthesized from 200 ng of total RNA using the ReverTra Ace qPCR RT Kit (Toyobo, Osaka, Japan). Quantitative PCR was performed using the Power SYBR Green Master Mix (Applied Biosystems, Foster City, CA, USA) on an ABI StepOnePlus Real-Time PCR System (Applied Biosystems). mRNA expression levels of target genes were normalized to β-actin (bact). The primer sequences used were as follows:
kdrl forward primer: 5’-CGCAAAGGAGACGCTAGACT-3’;
kdrl reverse primer: 5’-TGTAAGCCAGGGTAAGGGGA-3’;
bact forward primer: 5’-CATCCATCGTCCACAGGAAGTG-3’;
bact reverse primer: 5’-TGGTCGTTCGTTTGAATCTCAT-3’
2.7. Statistical Analysis
The results are presented as mean values with standard deviation (SD). Statistical significance was evaluated using one-way analysis of variance (ANOVA), followed by Bonferroni-Dunn multiple comparison tests. Data analyses were performed using GraphPad Prism version 10.4.2 (GraphPad Software Inc., San Diego, CA, USA). Statistical significance was defined as a p-value of <0.05.
3. Results
3.1. InnoCellTM Plate Rescued Zebrafish Embryo Development in Low Oxygen Environments
To evaluate the potential of the InnoCellTM plate for culturing zebrafish embryos, we selected healthy embryos at 6 hpf and placed them in a 24-well standard PS culture plate (Control) and an InnoCellTM plate, varying the densities and oxygen conditions (Figure 1A). Embryos were incubated at 28℃ until 72 hpf, and heart rates were measured at 56 hpf. Under normal oxygen conditions covered with a plate lid (no sealing), the heart rate was significantly elevated in the InnocellTM plate compared with that in the control plate at densities of 5, 10, 20, 30, and 50 fish/mL (Figure 1B). However, in an extreme hypoxic condition, by cutting oxygen using a cohere plate seal (sealing), the heart rates of embryos cultured in the InnoCellTM plate were significantly slower than those of the embryos cultured in the control plate at the densities of 5, 10, 20, 30, and 50 fish/mL (Figure 1C). The average body length of embryos cultured on the control plate was shorter than that of embryos cultured on the InnoCellTM plate at 5, 20, 30, and 50 fish/mL (Figure 1D). Furthermore, under extremely limited oxygen conditions, the body length of embryos cultured in the InnoCellTM plate at 5, 10, 20, 30, and 50 fish/mL was significantly longer than that of embryos cultured in the control plate (Figure 1E). Representative images of these conditions are presented in Figure 1F and 1G. Under the "No sealing" condition, larvae cultured in control plates exhibited signs of developmental delay compared with those cultured in InnoCellTM plates, particularly at higher densities (20–50 fish/mL) (Figure 1F; Supplementary Figure S1A). These signs include shorter body length, lighter body pigmentation, and larger yolk sacs (indicating slower nutrient absorption post-hatching). Furthermore, when the oxygen supply was restricted by sealing, larvae cultured in control plates displayed markedly more pronounced developmental delays (Figure 1G; Supplementary Figure S1B). Typically, zebrafish embryos emerge from their chorions at 72 hpf. However, when maintained under sealed conditions at a concentration of 50 fish/mL, the hatching rate in the control plates was 13%, which increased significantly to 84% in the InnoCellTM plates.
3.2. Developmental Assessment Using 96-well Plates
A 96-well plate was used for drug screening in zebrafish. Initially, we assessed the effectiveness of the InnoCellTM plate for embryo development and determined the minimum culture medium volume required. According to the guidelines outlined in The Zebrafish Book for nurturing zebrafish larvae, a density of 25‒50 embryos per 100 mL (4000 µL/fish) should be maintained [19]. Consequently, we incorporated a “Petri dish” condition as the standard control environment. Embryos were cultured individually in wells of both control and InnoCellTM plates with varying liquid volumes. Figure 2B illustrates that zebrafish in the InnoCellTM plate had significantly higher heart rates, reaching 190 ± 2 bpm, compared to those of embryos on the control plate (179 ± 6 bpm; p < 0.05) at a volume of 50 μL per well per fish. Additionally, at a volume of 200 μL per well per fish without sealing, embryo heart rates were 175 ± 2 and 166 ± 5 bpm for the InnoCellTM and control plates, respectively. At volumes of 30 and 40 μL/well/fish under sealing conditions, embryos cultured in InnoCellTM plates had higher average heart rates than those cultured on control plates (p < 0.05; Figure 2C). Similar trends were observed for body length measurements at 72 hpf (Figure 2D-E). Embryos cultured in the InnoCellTM plate exhibited a notable increase in body length at volumes of 15, 10, 100, and 200 µL/well/fish without sealing compared with those cultured in the control plate (Figure 2D). Additionally, an increase in embryo body length was observed at 20, 40, 100, and 200 µL/well/fish under sealed conditions in the InnoCellTM plate compared with that in the control plate (Figure 2E). Although 10, 15, 20, and 30 µL/well/fish under no sealing condition did not induce any issues in embryo development, a three-day culture resulted in notable evaporation. Accordingly, we identified ≥40 µL/well/fish as the minimum volume for normal embryo development under no sealing condition using the 96-well InnoCellTM plate. Based on body length results at 72 hpf, ˃20 µL/well/fish was identified as the optimal minimum volume for embryo development under sealing conditions using the 96-well InnoCellTM plate
3.3. Developmental Assessment Using 384-well Plates
For zebrafish-based drug screening, 384-well plates are primarily used for high-throughput compound screening and automated image analysis, depending on the experimental objectives and specific conditions. To determine the applicability of the 384-well InnoCellTM plate, we performed a developmental assessment similar to that performed using 96-well plates. Compared with the standard normal conditions (Petri dish), embryos cultured in the control plates exhibited markedly delayed development, although this phenomenon was not observed in embryos cultured in InnoCellTM plates. For culture volumes of 40, 60, 80 µL/well/fish under no sealing condition (Figure 3A) and 30, 40, 60, 80 µL/well/fish under sealing condition (Figure 3B), embryos cultured in InnoCellTM plate exhibited significantly higher heart rates than those cultured in control plates. Under both no sealing and sealing conditions, the body length of embryos at 72 hpf was significantly increased when cultured in volumes exceeding 20 µL/well/fish (Figure 3C and 3D). Accordingly, we concluded that the minimum culture volume should be 20 and 15 µL/well/fish under no sealing and sealing conditions, respectively, when using the 384-well InnoCellTM plate.
3.4. Application for Drug Screening
Given the successful performance of the InnoCellTM plate in supporting zebrafish embryo development, we explored its effectiveness in drug screening applications. Antiangiogenic receptor TKIs were selected owing to their well-characterized and quantifiable biological effects in zebrafish models. The TKIs used in this study are listed in Table 1.
For 96-well plates, the minimum and maximum volume selected were 40 and 100 µL/well/fish, respectively, under the no sealing condition; for the sealing condition, the minimum and maximum volumes selected were 20 and 100 µL/well/fish, respectively. After drug exposure from 24 to 96 hpf, the zebrafish were anesthetized, and GFP fluorescent images were obtained to enumerate ISVs (Figure 4A). Embryos exposed to PTK787, SU5416, SU4312, or the negative control VPA showed no significant differences in the number of ISVs between the control and InnoCellTM plates, regardless of oxygen conditions and drug volume. Exposure to sorafenib and axitinib exerted strong angiogenic effects under almost all conditions. Embryos exposed to sorafenib showed a decreasing trend in the number of ISVs when cultured at 40 µL/well/fish in InnoCellTM plates under the no sealing condition (p = 0.09; Figure 4B), although no differences were observed between the InnoCellTM and control plates at 100 µL/well/fish (Figure 4C). Conversely, embryos cultured at 20 µL/well/fish in the InnoCellTM plate exhibited a significantly lower number of ISVs than those cultures in control plates under sealing conditions (p < 0.05; Figure 4D). However, no differences between these two plates were observed at 100 µL/well/fish (Figure 4E).
Given their anti-angiogenic capacity, we selected SU4312, sorafenib, and axitinib for evaluation in 384-well plates. The minimum and maximum volumes were 20 and 80 µL/well/fish for no sealing conditions and 15 and 80 µL/well/fish for sealing conditions. Similar to the observations in 96-well plates, ISV numbers in embryos exposed to the maximum TKI volumes did not differ significantly between InnoCellTM and control 384-well plates under both no sealing and sealing conditions (Figure 5B and D). However, exposure to SU4312 significantly reduced the number of ISVs at 20 µL/well/fish volume under no sealing conditions in the InnoCellTM plate compared with that in the control plate (Figure 5A). Axitinib also exhibited greater activity in the InnoCellTM plate under sealing conditions at a volume of 15 µL/fish (Figure 5C).
3.5. Expression Levels of the kdrl Gene
As the manual assessment of angiogenesis inhibition may lack sufficient sensitivity, we quantified the expression levels of kdrl (homolog of the human VEGFR2) under the minimum volume and sealing conditions using 96- and 384-well plates. Consistent with the observations in Figure 5A, mRNA expression levels were significantly reduced in embryos exposed to sorafenib and axitinib at 20 µL in the control plates (p < 0.01 and p < 0.05, respectively; Figure 6A). Notably, a more significant inhibitory effect was observed in InnoCellTM plates (p < 0.0001; Figure 6B), especially upon exposure to SU4312, with no significant suppression of ISV numbers. Assessment in 384-well plates using the minimum volume of 15 µL/well/fish revealed no significant differences in the kdrl expression upon exposure to SU4312 in both the control and InnoCellTM plates (Figure 6C and D). Exposure to sorafenib significantly downregulated kdrl expression in control plates (Figure 6C), whereas a trend toward inhibition was observed in the InnoCellTM plates (Figure 6D). Additionally, axitinib exhibited a robust inhibitory effect on kdrl expression in both the control and InnoCellTM plates.
4. Discussion
This study highlights the substantial benefits of employing a highly oxygen-permeable culture plate, known as the InnoCellTM plate, for drug screening using zebrafish. Sufficient oxygen supply is crucial for embryonic development, as it is vital for metabolism, growth, and differentiation, especially in fast-developing organisms such as zebrafish. Hypoxic conditions can rapidly arise in traditional PS culture plates, negatively affecting cellular metabolism and viability [5,6]. The innovative InnoCellTM plate, made from PMP, substantially enhances oxygen permeability compared with standard PS plates, thereby markedly improving cellular metabolism and function [11]. In this study, we found that the InnoCellTM plate demonstrated notably enhanced developmental outcomes in zebrafish embryos, including heart rate and body length, under both normal and oxygen-restricted conditions, compared with conventional PS plates (Figure 1). These results align with earlier findings in mammalian cell cultures and extend the positive effects of PMP materials on vertebrate embryogenesis and organism-level development.
Notably, under oxygen-restricted (sealed) conditions, the advantages of the InnoCellTM plates were more notable. Embryos cultured on conventional PS plates exhibited marked developmental delays, including reduced body length, delayed yolk absorption, impaired pigmentation, and significantly lower hatching rates at higher densities (Figure 1E and 1G). Conversely, embryos in the InnoCellTM plate maintained relatively normal developmental profiles, suggesting that even limited oxygen transfer through the PMP base was sufficient to alleviate severe hypoxic stress and support normal developmental trajectories. Interestingly, under the same conditions (Figure 1C), embryos grown on PS plates unexpectedly demonstrated elevated heart rates in almost all medium volumes. This may reflect a transient compensatory response to acute hypoxia, as previously reported in zebrafish embryos [20,21]. Hypoxia can trigger tachycardia to enhance oxygen delivery before cardiac function declines. In contrast, embryos in the InnoCellTM plate maintained stable heart rates, indicating adequate oxygen availability. Collectively, these results emphasize the hypoxia-induced physiological stress in PS plates and the advantages of utilizing oxygen-permeable systems, such as the InnoCellTM plate. Moreover, these findings highlight the critical importance of oxygen availability during zebrafish development and demonstrate that PMP plates with high oxygen permeability offer substantial benefits under difficult conditions, such as high embryo densities or sealed cultures. This technology enhances the assay consistency and has considerable potential to improve the reliability of zebrafish-based drug screening.
In studies using 96-well plates (Figure 2), the InnoCellTM plate demonstrated superior support for the development of zebrafish embryos compared with traditional control plates across various medium volumes. Embryos grown in the InnoCellTM plate displayed notably higher heart rates and increased body lengths, especially at medium volumes ranging from 50 to 200 μL per well, under both sealed and unsealed conditions. Remarkably, the InnoCellTM plate supported normal development even at lower volumes, down to 40 μL per well without sealing and 20 μL per well with sealing. In contrast, PS plates necessitated larger volumes to avoid developmental delays. This benefit is likely attributable to the improved oxygen permeability of the InnoCellTM plate, which compensates for the smaller medium volumes by ensuring adequate oxygen delivery. Determining the optimal minimum medium volume in 96-well plates is essential for balancing cost, efficiency, and embryo health. If the volume is extremely low, oxygen depletion, nutrient deficiency, and pH changes can hinder development [5]. Zebrafish embryos can reportedly withstand oxygen levels as low as about 3.33 mg/L without major developmental issues; however, a drop below this level may lead to stunted growth or death [22]. Moreover, evaporation in small volumes can concentrate substances, thereby increasing the risk of toxicity. Generally, medium volumes of 100–200 μL per well are employed in zebrafish drug screening to ensure adequate oxygen and nutrients [23,24,25]. However, certain protocols allow this volume to be reduced to 50 or 70 μL per well [26,27], which can heighten the risk of hypoxia and evaporation, especially in PS plates [28]. Based on our findings, the InnoCellTM plate facilitates normal development even at volumes as low as 20–40 μL per well, highlighting its potential for economical, high-throughput screening. This has substantial implications for drug screening processes, given that reducing the medium volume can decrease reagent expenses and increase screening efficiency. Additionally, the InnoCellTM plate offers a more stable and physiologically relevant setting, enhancing the reliability and consistency of zebrafish-based assays, even in high-throughput scenarios. Our findings underscore the importance of integrating plate design with volume optimization to enhance zebrafish drug screening systems.
The 96-well plates are commonly employed in zebrafish drug screening owing to their appropriate size and ease of use, whereas the 384-well plates are less frequently used because of technical difficulties, such as smaller wells, limited medium volume, and evaporation, which can affect embryo survival and data integrity [17]. Nonetheless, 384-well plates have notable benefits, including lower reagent costs, increased throughput, and compatibility with automated imaging systems, making them a promising option for large-scale chemical or genetic screening [15,16,29]. In our study involving a 384-well plate setup (Figure 3), the InnoCellTM plate exhibited distinct advantages over the standard PS plates. Embryos cultured in the InnoCellTM plate maintained substantially higher heart rates and greater body lengths, even when the medium volume was reduced to 20 μL per well without sealing and 15 μL per well with sealing conditions that typically impede development in conventional plates. This is likely due to the high oxygen permeability of the PMP material, which helps overcome the oxygen constraints frequently encountered in the 384-well plate setup. Notably, this feature allows for a substantial reduction in medium volume without impacting embryo viability or assay sensitivity, which is crucial for high-throughput screening. These findings emphasize the potential of the InnoCellTM plate to enhance screening efficiency, cost-effectiveness, and data reproducibility in automated zebrafish-based systems.
Moreover, drug screening tests using TKIs demonstrated the effectiveness of the InnoCellTM plate in identifying antiangiogenic effects in zebrafish embryos compared with the traditional PS plates. In both 96- and 384-well formats, the InnoCellTM plate enabled the accurate evaluation of ISV formation and vascular gene expression (kdrl), even with reduced medium volumes and under oxygen-limited conditions (sealing). Exposure to sorafenib and axitinib substantially inhibited ISVs, with consistent suppression observed on the InnoCellTM plate, particularly at lower volumes. Although SU4312 demonstrated less consistent effects on ISV morphology, it substantially reduced kdrl mRNA levels, indicating that molecular responses can occur before or independently of visible vascular changes. This finding is consistent with previous studies that identified kdrl as a sensitive early marker of antiangiogenic drug activity in zebrafish models [30,31]. These results underscore the need to incorporate both phenotypic and molecular analyses to assess drug efficacy in zebrafish. Relying solely on vessel counts may lead to an underestimation of the biological activity of certain compounds, whereas combining morphological and gene expression evaluations would offer a more comprehensive assessment. Additionally, the data revealed that the high oxygen permeability of the InnoCellTM plate maintained assay sensitivity even under challenging conditions, such as reduced volume or sealing, which are crucial for high-throughput screening. Overall, the InnoCellTM platform enhances assay consistency, sensitivity, and scalability and advances zebrafish-based pharmacological assessment.
5. Conclusions
Utilization of the InnoCellTM culture plate, which allows for high oxygen permeability, greatly improves the development of zebrafish embryos and the sensitivity of assays in the drug screening context. Compared with conventional PS plates, InnoCellTM plates fostered better physiological conditions, as demonstrated by increased heart rates, longer body lengths, and higher developmental success rates under both normal and oxygen-restricted environments. Notably, the volume of culture medium required was markedly reduced without negatively impacting embryonic viability, making these plates especially beneficial for high-throughput formats such as 384-well assays. Additionally, drug screening outcomes revealed heightened sensitivity in identifying the antiangiogenic effects of TKIs, which was corroborated by both phenotypic analysis and molecular gene expression assessments. Collectively, these results strongly indicate that the use of InnoCellTM plates can markedly improve experimental results, optimize resource utilization, and enhance data reliability, thereby advancing zebrafish application in pharmaceutical and biomedical research.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Representative images of embryos at 72 hpf cultured in the control and InnoCellTM 24-well plates.
Author Contributions
Conceptualization, L. Z. and N. N.; methodology, L. Z.; validation, S. K. and L. Z.; formal analysis, S. K. and L. Z.; investigation, S. K. and L. Z.; resources, Y. K.; data curation, S. K. and L. Z.; writing—original draft preparation, S. K. and L. Z.; writing—review and editing, Y. K. and N. N.; supervision, N. N.; project administration, L. Z.; funding acquisition, L. Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Mitsui Chemicals, Inc.
Institutional Review Board Statement
This study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Animal Care and Use Committee of Mie University (2024-19).
Informed Consent Statement
Not applicable.
Data Availability Statement
All of the relevant data are presented within the paper.
Acknowledgments
The authors thank Masami Yamamura for her assistance with animal management, and Ms. Chikako Araki and Mrs. Yukako Demiya for their secretarial assistance.
Conflicts of Interest
The authors declare that this study received funding from Mitsui Chemicals Inc. The funder provided the resources but was not involved in the study design, collection, analysis, interpretation of data, or decision to submit it for publication.
Abbreviations
The following abbreviations are used in this manuscript:
| TKIs | tyrosine kinase inhibitors |
| kdrl | vascular endothelial growth factor receptor |
| PS | polystyrene |
| PDMS | polydimethylsiloxane |
| PMP | 4-polymethyl-1-pentene polymer |
| VPA | valproic acid |
| ISVs | intersegmental blood vessels |
| VEGFR | vascular endothelial growth factor receptor |
| PDGF | platelet-derived growth factor |
| FIt-4 | Fms-like tyrosine kinase 4 |
| c-Kit | mast/stem cell growth factor receptor Kit |
| PDGFR | platelet-derived growth factor receptor |
| EGFR | epidermal growth factor receptor |
| HER-2 | human epidermal growth factor receptor 2 |
| IGF | insulin-like growth factor |
| FGFR1 | fibroblast growth factor receptor 1 |
| RAF | rapidly accelerated fibrosarcoma |
| HDAC1 | histone deacetylase 1 |
References
- Abuwatfa, W.H.; Pitt, W.G.; Husseini, G.A. Scaffold-based 3D cell culture models in cancer research. J. Biomed. Sci. 2024, 31, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, H.; Kimmel, H.; Rondo, E.; Underhill, G.H. Advances in high throughput cell culture technologies for therapeutic screening and biological discovery applications. Bioeng. Transl. Med. 2023, 9, e10627. [Google Scholar] [CrossRef]
- Sharma, K.; Dey, S.; Karmakar, R.; Rengan, A.K. A comprehensive review of 3D cancer models for drug screening and translational research. Cancer Innov. 2023, 3, e102. [Google Scholar] [CrossRef]
- Esparza, A.; Jimenez, N.; Borrego, E.A.; Browne, S.; Natividad-Diaz, S.L. Review: Human stem cell-based 3D in vitro angiogenesis models for preclinical drug screening applications. Mol. Biol. Rep. 2024, 51, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kieninger, J.; Aravindalochanan, K.; Sandvik, J.A.; Pettersen, E.O.; Urban, G.A. Pericellular oxygen monitoring with integrated sensor chips for reproducible cell culture experiments. Cell Prolif. 2014, 47, 180–188. [Google Scholar] [CrossRef]
- Scheidecker, B.; Shinohara, M.; Sugimoto, M.; Danoy, M.; Nishikawa, M.; Sakai, Y. Induction of in vitro Metabolic Zonation in Primary Hepatocytes Requires Both Near-Physiological Oxygen Concentration and Flux. Front. Bioeng. Biotechnol. 2020, 8, 524. [Google Scholar] [CrossRef]
- Xiao, W.; Shinohara, M.; Komori, K.; Sakai, Y.; Matsui, H.; Osada, T. The importance of physiological oxygen concentrations in the sandwich cultures of rat hepatocytes on gas-permeable membranes. Biotechnol. Prog. 2014, 30, 1401–1410. [Google Scholar] [CrossRef]
- Wang, J.D.; Douville, N.J.; Takayama, S.; Elsayed, M. Quantitative Analysis of Molecular Absorption into PDMS Microfluidic Channels. Ann. Biomed. Eng. 2012, 40, 1862–1873. [Google Scholar] [CrossRef]
- Toepke, M.W.; Beebe, D.J. PDMS absorption of small molecules and consequences in microfluidic applications. Lab Chip 2006, 6, 1484–1486. [Google Scholar] [CrossRef]
- Sticker, D.; Rothbauer, M.; Ehgartner, J.; Steininger, C.; Liske, O.; Liska, R.; Neuhaus, W.; Mayr, T.; Haraldsson, T.; Kutter, J.P.; et al. Oxygen Management at the Microscale: A Functional Biochip Material with Long-Lasting and Tunable Oxygen Scavenging Properties for Cell Culture Applications. ACS Appl. Mater. Interfaces 2019, 11, 9730–9739. [Google Scholar] [CrossRef]
- Nishikawa, M.; Ito, H.; Tokito, F.; Hirono, K.; Inamura, K.; Scheidecker, B.; Danoy, M.; Kawanishi, T.; Arakawa, H.; Kato, Y.; et al. Accurate Evaluation of Hepatocyte Metabolisms on a Noble Oxygen-Permeable Material With Low Sorption Characteristics. Front. Toxicol. 2022, 4, 810478. [Google Scholar] [CrossRef] [PubMed]
- Fox, S.C.; Waskiewicz, A.J. Transforming growth factor beta signaling and craniofacial development: modeling human diseases in zebrafish. Front. Cell Dev. Biol. 2024, 12, 1338070. [Google Scholar] [CrossRef] [PubMed]
- Zang, L.; Maddison, L.A.; Chen, W. Zebrafish as a Model for Obesity and Diabetes. Front. Cell Dev. Biol. 2018, 6, 91. [Google Scholar] [CrossRef]
- Kolesnikova, T.O.; Demin, K.A.; Costa, F.V.; de Abreu, M.S.; Kalueff, A.V. Zebrafish models for studying cognitive enhancers. Neurosci. Biobehav. Rev. 2024, 164, 105797. [Google Scholar] [CrossRef] [PubMed]
- Chapela, D.; Sousa, S.; Martins, I.; Cristovao, A. M.; Pinto, P.; Corte-Real, S.; Saude, L. , A zebrafish drug screening platform boosts the discovery of novel therapeutics for spinal cord injury in mammals. Sci Rep 2019, 9, (1), 10475. [Google Scholar] [CrossRef]
- Habjan, E.; Schouten, G.K.; Speer, A.; van Ulsen, P.; Bitter, W. Diving into drug-screening: zebrafish embryos as an in vivo platform for antimicrobial drug discovery and assessment. FEMS Microbiol. Rev. 2024, 48. [Google Scholar] [CrossRef]
- Wang, W.; Gao, X.; Liu, L.; Guo, S.; Duan, J.-A.; Xiao, P. Zebrafish as a Vertebrate Model for High-Throughput Drug Toxicity Screening: Mechanisms, Novel Techniques, and Future Perspectives. J. Pharm. Anal. 2025. [Google Scholar] [CrossRef]
- Bussmann, J.; Bakkers, J.; Schulte-Merker, S. , Early endocardial morphogenesis requires Scl/Tal1. PLoS Genet 2007, 3, (8), e140. [Google Scholar] [CrossRef]
- Westerfield, M. A guide for the laboratory use of zebrafish (Danio rerio). In THE ZEBRAFISH BOOK, 5th ed.; University of Oregon Press: Eugene, 2007. [Google Scholar]
- Padilla, P.A.; Roth, M.B. Oxygen deprivation causes suspended animation in the zebrafish embryo. Proc. Natl. Acad. Sci. 2001, 98, 7331–7335. [Google Scholar] [CrossRef]
- Barrionuevo, W. R.; Burggren, W.W. O2 consumption and heart rate in developing zebrafish (Danio rerio): influence of temperature and ambient O2. Am J Physiol 1999, 276, (2), R505–13. [Google Scholar]
- Strecker, R.; Seiler, T.-B.; Hollert, H.; Braunbeck, T. Oxygen requirements of zebrafish (Danio rerio) embryos in embryo toxicity tests with environmental samples. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2011, 153, 318–327. [Google Scholar] [CrossRef]
- van Bree, N.; Oppelt, A.-S.; Lindström, S.; Zhou, L.; Boutin, L.; Coyle, B.; Swartling, F.J.; Johnsen, J.I.; Bräutigam, L.; Wilhelm, M. Development of an orthotopic medulloblastoma zebrafish model for rapid drug testing. Neuro-Oncology 2024, 27, 779–794. [Google Scholar] [CrossRef]
- Truong, L.; Bieberich, A. A.; Fatig, R. O., 3rd; Rajwa, B.; Simonich, M. T.; Tanguay, R. L. , Accelerating antiviral drug discovery: early hazard detection with a dual zebrafish and cell culture screen of a 403 compound library. Arch Toxicol 2025, 99, (3), 1029–1041. [Google Scholar] [CrossRef]
- Sturtzel, C.; Grissenberger, S.; Wenninger-Weinzierl, A.; Distel, M. High-Content Imaging-Based Screening for Anticancer Compounds in Zebrafish Xenografts. Methods Mol Biol 2025, 2905, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Widder, M.; Carbaugh, C.; van der Schalie, W.; Miller, R.; Brennan, L.; Moore, A.; Campbell, R.; Akers, K.; Ressner, R.; Martin, M.; et al. Identification of Potential Sepsis Therapeutic Drugs Using a Zebrafish Rapid Screening Approach. Life 2024, 14, 1689. [Google Scholar] [CrossRef] [PubMed]
- McGown, A.; Shaw, D.P.J.; Ramesh, T. ZNStress: a high-throughput drug screening protocol for identification of compounds modulating neuronal stress in the transgenic mutant sod1G93R zebrafish model of amyotrophic lateral sclerosis. Mol. Neurodegener. 2016, 11, 1–11. [Google Scholar] [CrossRef]
- Ali, S.; van Mil, H.G.J.; Richardson, M.K.; Mueller, F. Large-Scale Assessment of the Zebrafish Embryo as a Possible Predictive Model in Toxicity Testing. PLOS ONE 2011, 6, e21076. [Google Scholar] [CrossRef] [PubMed]
- Wiley, D. S.; Redfield, S. E.; Zon, L. I. , Chemical screening in zebrafish for novel biological and therapeutic discovery. Methods Cell Biol 2017, 138, 651–679. [Google Scholar] [CrossRef]
- Bussmann, J.; Bos, F.L.; Urasaki, A.; Kawakami, K.; Duckers, H.J.; Schulte-Merker, S. Arteries provide essential guidance cues for lymphatic endothelial cells in the zebrafish trunk. Development 2010, 137, 2653–2657. [Google Scholar] [CrossRef]
- Chan, J.; Bayliss, P.E.; Wood, J.M.; Roberts, T.M. Dissection of angiogenic signaling in zebrafish using a chemical genetic approach. Cancer Cell 2002, 1, 257–267. [Google Scholar] [CrossRef]
Figure 1.
Evaluation of the zebrafish embryo development cultured in the 24-well control and InnoCellTM plates at different densities. (A) Experimental design and schema. (B-C) Heart rate measurement at 56 hours post-fertilization (hpf) cultured in control and InnoCellTM plates under no sealing (B) and sealing (C) conditions. (D-E) Body length at 72 hpf cultured in the control and InnoCellTM plates under no sealing; (D) and sealing (E) conditions. (F-G) Representative images of embryos at 72 hpf cultured in the control and InnoCellTM plates at different densities under no sealing (F) and sealing (G) conditions. * p < 0.05, ** p < 0.01, *** p < 0.001, n = 5. Data are presented as the mean ± standard deviation (SD).
Figure 1.
Evaluation of the zebrafish embryo development cultured in the 24-well control and InnoCellTM plates at different densities. (A) Experimental design and schema. (B-C) Heart rate measurement at 56 hours post-fertilization (hpf) cultured in control and InnoCellTM plates under no sealing (B) and sealing (C) conditions. (D-E) Body length at 72 hpf cultured in the control and InnoCellTM plates under no sealing; (D) and sealing (E) conditions. (F-G) Representative images of embryos at 72 hpf cultured in the control and InnoCellTM plates at different densities under no sealing (F) and sealing (G) conditions. * p < 0.05, ** p < 0.01, *** p < 0.001, n = 5. Data are presented as the mean ± standard deviation (SD).
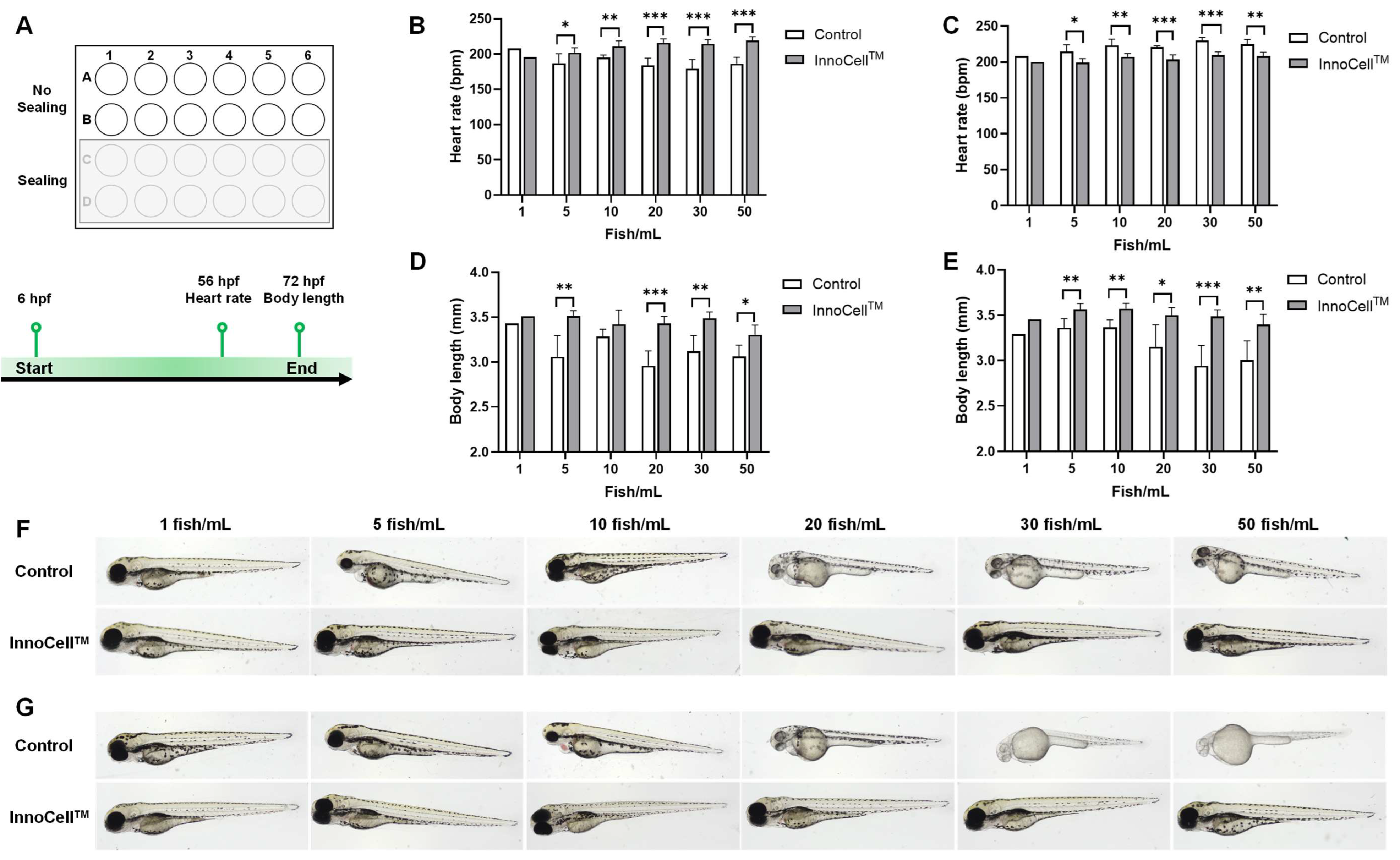
Figure 2.
Figure 2. Evaluation of embryo development cultured in the 96-well control and InnoCellTM plates at different volumes. (A) Experimental design and schema. (B-C) Heart rate measurement at 56 hours post-fertilization (hpf) cultured in control and InnoCellTM plates under no sealing (B) and sealing (C) conditions. (D-E) Body length at 72 hpf cultured in the control and InnoCellTM plates under no sealing (D) and sealing (E) conditions. * p < 0.05, ** p < 0.01, *** p < 0.001, n = 4‒20. Data are presented as the mean ± standard deviation (SD).
Figure 2.
Figure 2. Evaluation of embryo development cultured in the 96-well control and InnoCellTM plates at different volumes. (A) Experimental design and schema. (B-C) Heart rate measurement at 56 hours post-fertilization (hpf) cultured in control and InnoCellTM plates under no sealing (B) and sealing (C) conditions. (D-E) Body length at 72 hpf cultured in the control and InnoCellTM plates under no sealing (D) and sealing (E) conditions. * p < 0.05, ** p < 0.01, *** p < 0.001, n = 4‒20. Data are presented as the mean ± standard deviation (SD).
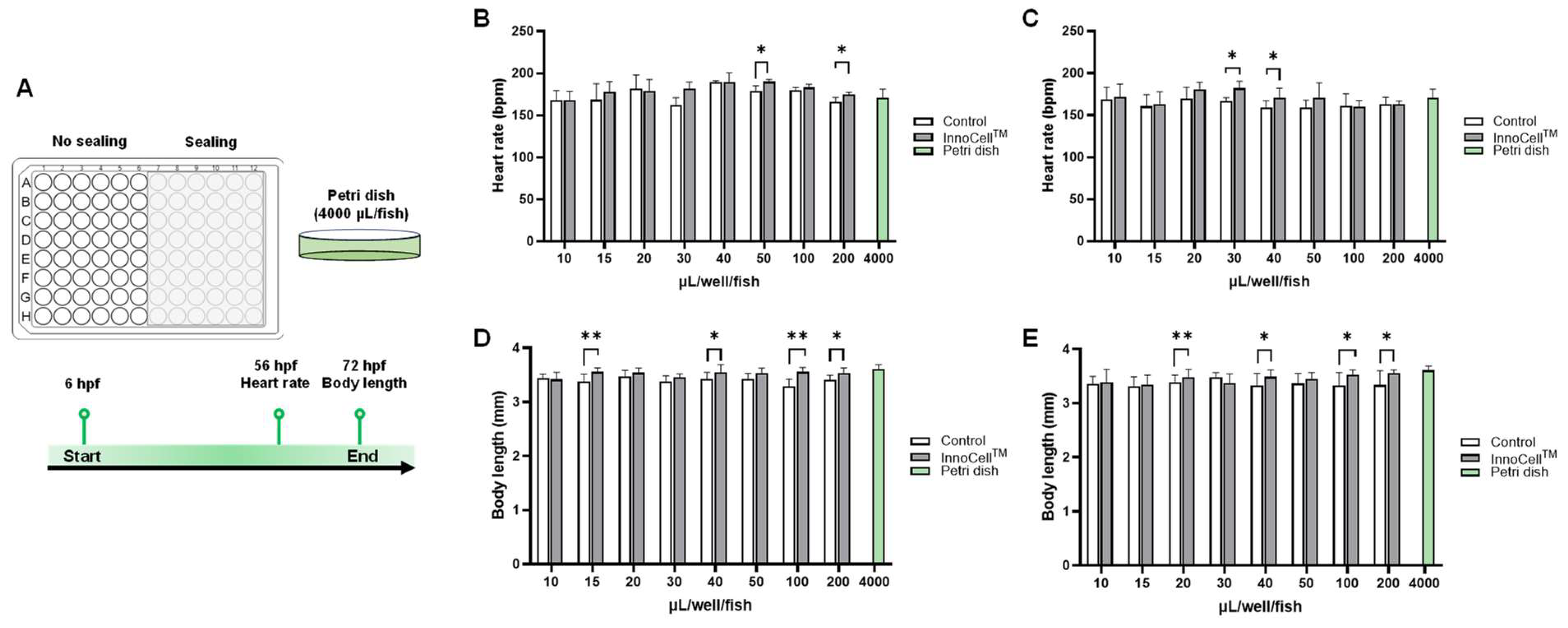
Figure 3.
Figure 3. Evaluation of the zebrafish embryo development cultured in the 384-well control and InnoCellTM plates at different volumes. (A-B) Heart rate measurement at 56 hours post-fertilization (hpf) cultured in the control and InnoCellTM plates under no sealing (A) and sealing (B) conditions. (C-D) Body length at 72 hpf cultured in the control and InnoCellTM plates under no sealing (C) and sealing (D) conditions. * p < 0.05, ** p < 0.01, *** p < 0.001, n = 4‒8. Data are presented as the mean ± standard deviation (SD).
Figure 3.
Figure 3. Evaluation of the zebrafish embryo development cultured in the 384-well control and InnoCellTM plates at different volumes. (A-B) Heart rate measurement at 56 hours post-fertilization (hpf) cultured in the control and InnoCellTM plates under no sealing (A) and sealing (B) conditions. (C-D) Body length at 72 hpf cultured in the control and InnoCellTM plates under no sealing (C) and sealing (D) conditions. * p < 0.05, ** p < 0.01, *** p < 0.001, n = 4‒8. Data are presented as the mean ± standard deviation (SD).
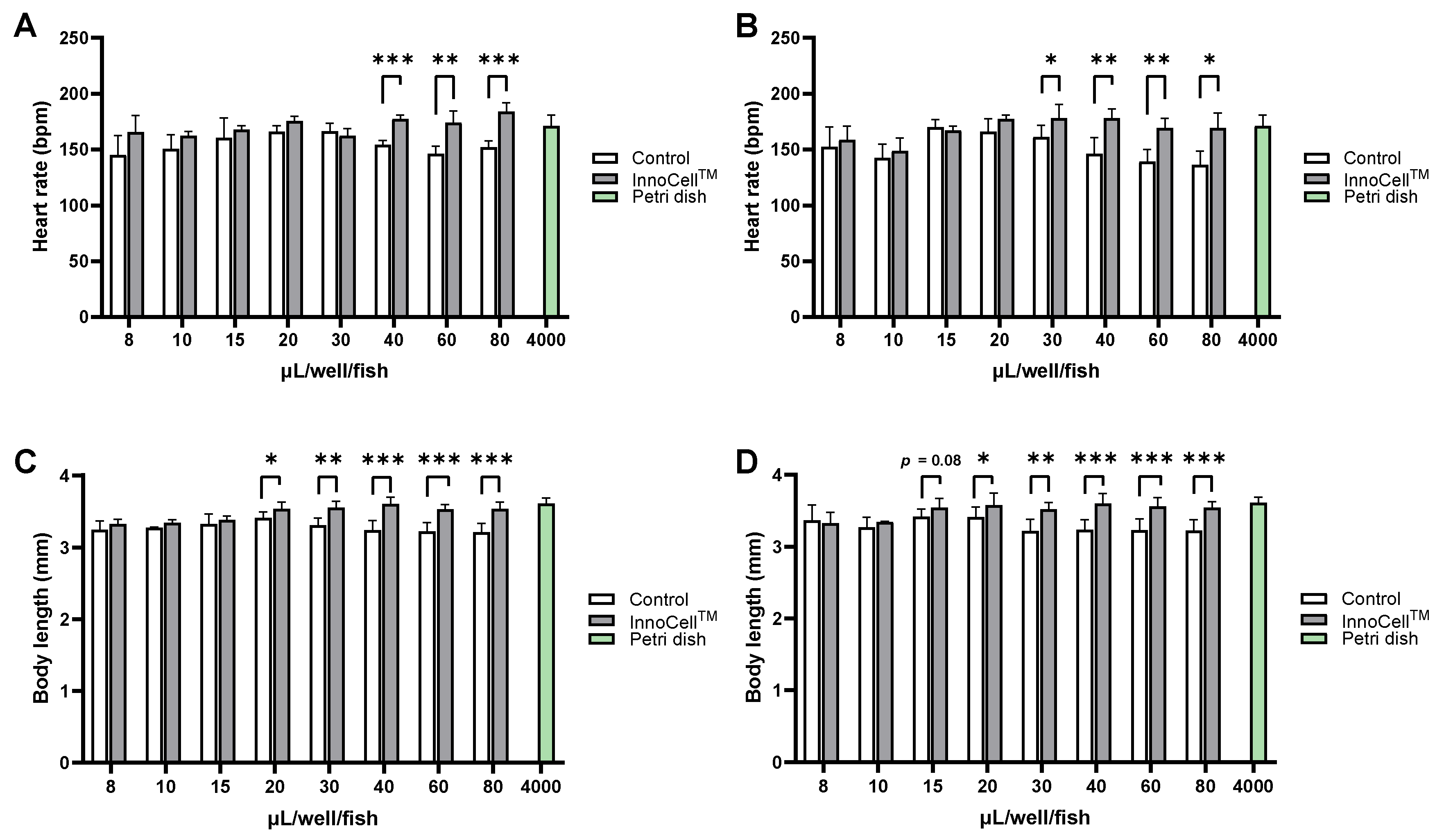
Figure 4.
Drug screening using the 96-well plates. (A) Experimental design and schema. The upper panels are the bright field (upper) and GFP fluorescent (lower) images of a 96 hours post-fertilization (hpf) zebrafish embryo. Arrows indicate the venous intersegmental blood vessels (ISVs). (B-C) Numbers of ISV after drug exposure at 96 hpf cultured in the control and InnoCellTM plates under no sealing conditions with a culture medium volume of 40 µL/well/fish (B) and 100 µL/well/fish (C). (D-E) Numbers of ISV after drug exposure at 96 hpf cultured in the control and InnoCellTM plates under sealing conditions with a culture medium volume of 20 µL/well/fish (D) and 100 µL/well/fish (E). (F) Representative images of ISVs in the trunk at 96 hpf after exposure to each TKI. * p < 0.05, n = 5-16. Data are presented as the mean ± standard deviation (SD). TKI, tyrosine kinase inhibitor; VA, valproic acid.
Figure 4.
Drug screening using the 96-well plates. (A) Experimental design and schema. The upper panels are the bright field (upper) and GFP fluorescent (lower) images of a 96 hours post-fertilization (hpf) zebrafish embryo. Arrows indicate the venous intersegmental blood vessels (ISVs). (B-C) Numbers of ISV after drug exposure at 96 hpf cultured in the control and InnoCellTM plates under no sealing conditions with a culture medium volume of 40 µL/well/fish (B) and 100 µL/well/fish (C). (D-E) Numbers of ISV after drug exposure at 96 hpf cultured in the control and InnoCellTM plates under sealing conditions with a culture medium volume of 20 µL/well/fish (D) and 100 µL/well/fish (E). (F) Representative images of ISVs in the trunk at 96 hpf after exposure to each TKI. * p < 0.05, n = 5-16. Data are presented as the mean ± standard deviation (SD). TKI, tyrosine kinase inhibitor; VA, valproic acid.
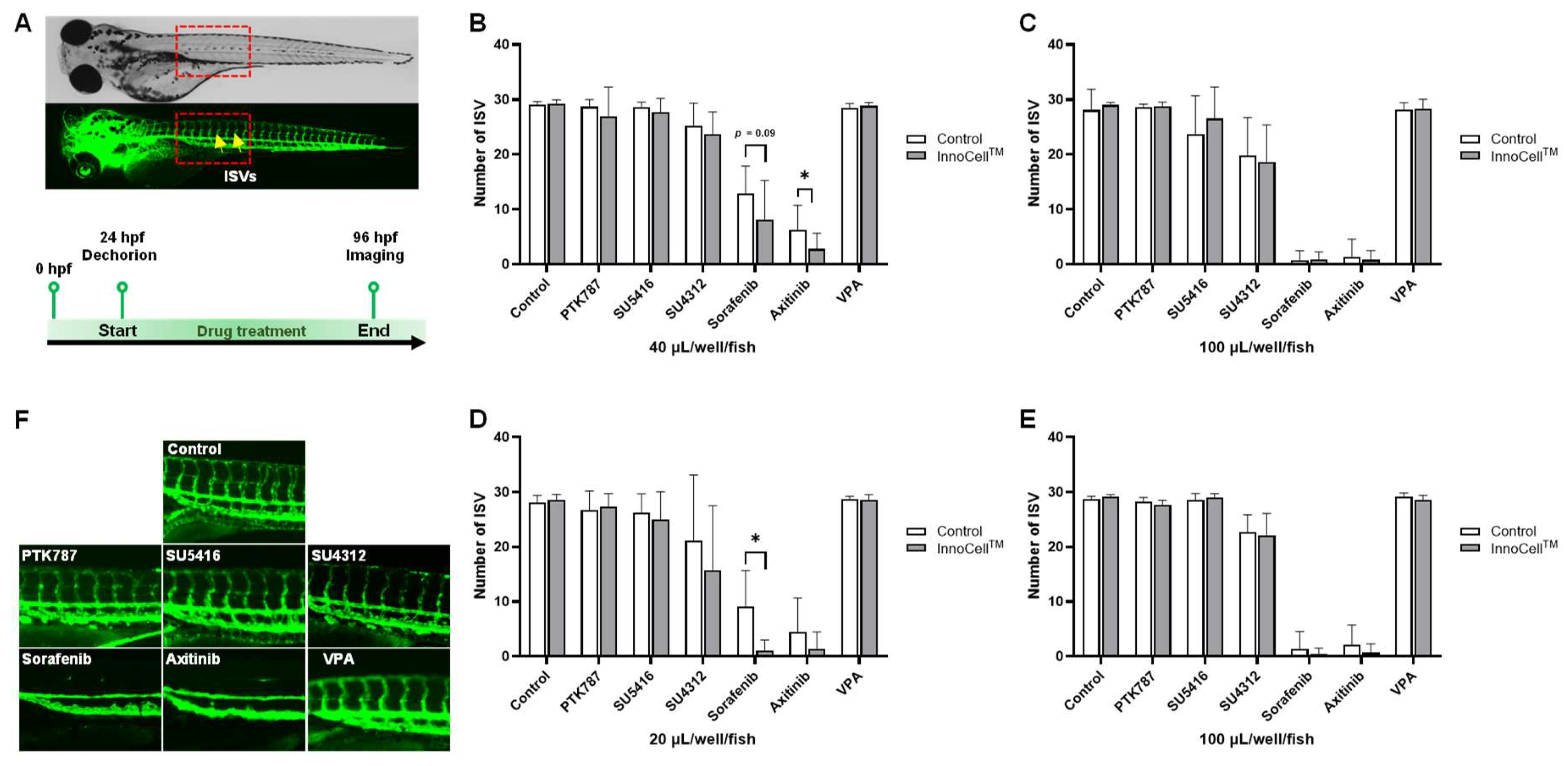
Figure 5.
Drug screening using the 384-well plates. (A-B) Numbers of ISV after drug exposure at 96 hpf cultured in control and InnoCellTM plates under no sealing conditions with a culture medium volume of 20 µL/well/fish (A) and 80 µL/well/fish (B). (C-D) Numbers of ISV after drug exposure at 96 hpf cultured in the control and InnoCellTM plates under sealing conditions with a culture medium volume of 15 µL/well/fish (C) and 80 µL/well/fish (D). ** p < 0.01, n = 7. Mean ± standard deviation (SD). VA, valproic acid.
Figure 5.
Drug screening using the 384-well plates. (A-B) Numbers of ISV after drug exposure at 96 hpf cultured in control and InnoCellTM plates under no sealing conditions with a culture medium volume of 20 µL/well/fish (A) and 80 µL/well/fish (B). (C-D) Numbers of ISV after drug exposure at 96 hpf cultured in the control and InnoCellTM plates under sealing conditions with a culture medium volume of 15 µL/well/fish (C) and 80 µL/well/fish (D). ** p < 0.01, n = 7. Mean ± standard deviation (SD). VA, valproic acid.
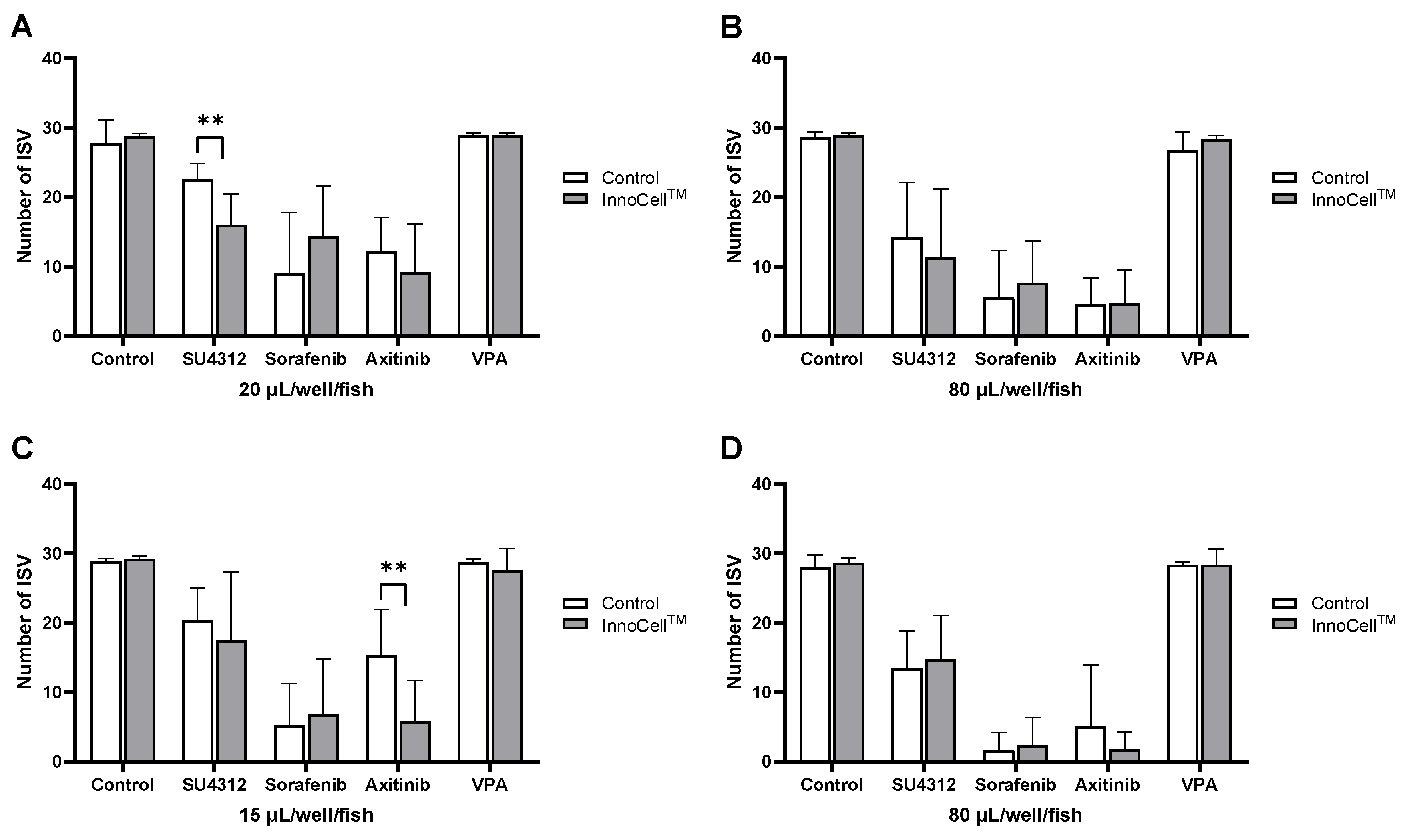
Figure 6.
Expression levels of kdrl upon exposure to TKIs under sealing conditions. (A, B) Relative mRNA expression levels of kdrl at 96 hours post-fertilization (hpf) exposed to 20 µL TKIs under sealing conditions using the 96-well control (A) and the InnoCellTM (B) plates. (C, D) Relative mRNA expression levels of kdrl at 96 hpf exposed to 15 µL TKIs under sealing conditions using the 384-well control (C) and the InnoCellTM (D) plates. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, n = 4. Data are presented as the mean ± standard deviation (SD). TKIs, tyrosine kinase inhibitors; VA, valproic acid.
Figure 6.
Expression levels of kdrl upon exposure to TKIs under sealing conditions. (A, B) Relative mRNA expression levels of kdrl at 96 hours post-fertilization (hpf) exposed to 20 µL TKIs under sealing conditions using the 96-well control (A) and the InnoCellTM (B) plates. (C, D) Relative mRNA expression levels of kdrl at 96 hpf exposed to 15 µL TKIs under sealing conditions using the 384-well control (C) and the InnoCellTM (D) plates. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, n = 4. Data are presented as the mean ± standard deviation (SD). TKIs, tyrosine kinase inhibitors; VA, valproic acid.
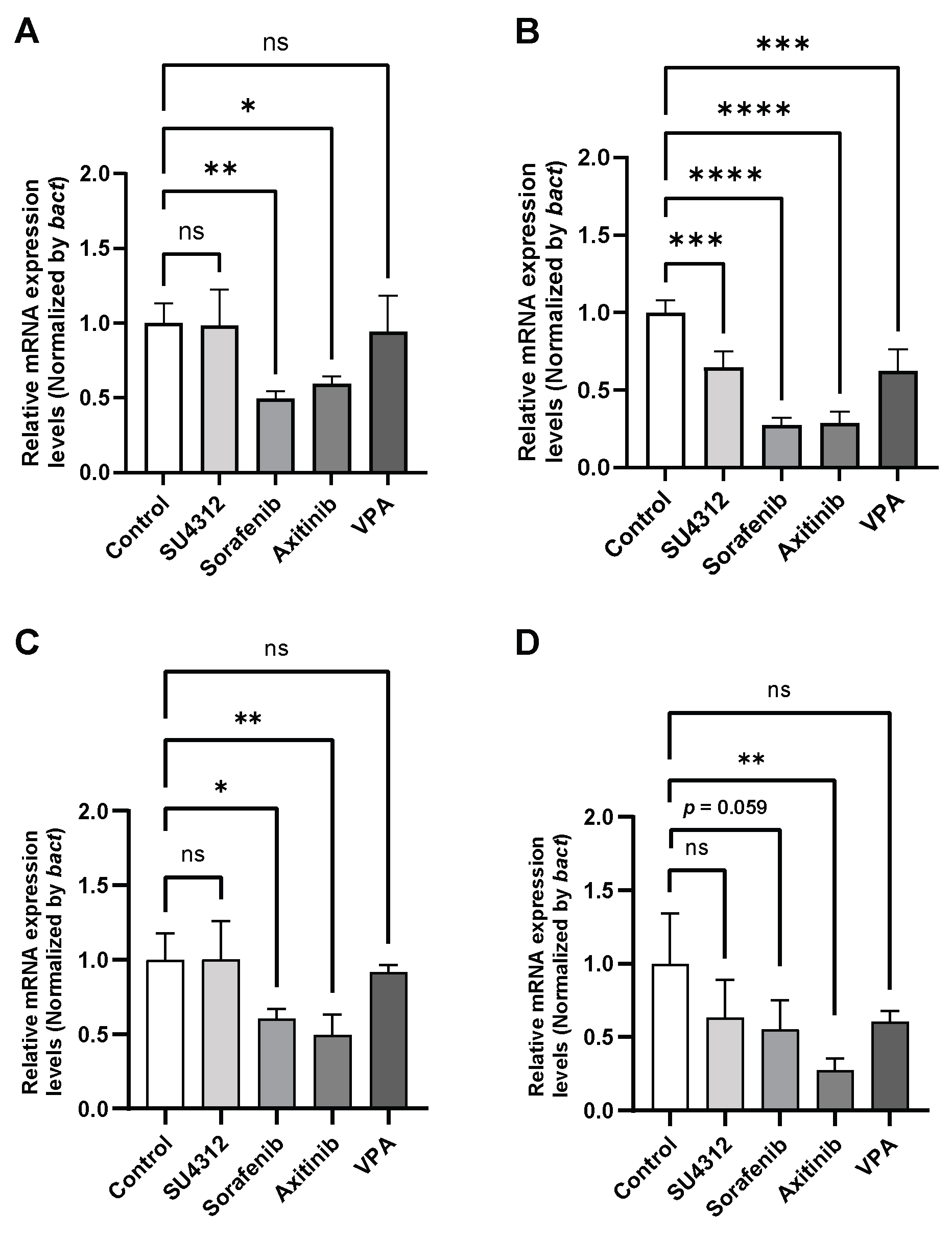

Table 1.
Properties of tyrosine kinase inhibitors.
| Drug | CAS No. | Molucular Weight (Da) | Targets | Exposure Concentration (µM) |
|---|---|---|---|---|
| PTK787 | 212141-51-0 | 419.73 | VEGFR2, VEGFR-1, PDGF, FIt-4, c-Kit | 0.15 |
| SU5416 | 204005-46-9 | 238.29 | VEGFR2, PDGFR, FIt-1, FIt-4, c-kit |
2.5 |
| SU4312 | 5812-07-7 | 264.3 | VEGFR2, PDGFR, EGFR, HER-2, IGF | 6 |
| Sorafenib | 284461-73-0 | 464.82 | VEGFR, PDGFR, FGFR1, KIT, RAF | 1.5 |
| Axitinib | 319460-85-0 | 386.47 | VEGFR1, VEGFR2, VEGFR3, PDGFRβ, c-Kit | 0.5 |
| Valproic acid | 99-66-1 | 144.21 | HDAC1 | 5 |
VEGFR, vascular endothelial growth factor receptor; PDGF, platelet-derived growth factor; FIt-4, Fms-like tyrosine kinase 4; c-Kit, mast/stem cell growth factor receptor Kit; PDGFR, platelet-derived growth factor receptor; EGFR, epidermal growth factor receptor; HER-2, human epidermal growth factor receptor 2; IGF, insulin-like growth factor; FGFR1, fibroblast growth factor receptor 1; RAF, rapidly accelerated fibrosarcoma; HDAC1, histone deacetylase 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



