Submitted:
14 June 2025
Posted:
16 June 2025
You are already at the latest version
Abstract
Highly pathogenic avian influenza (HPAI) H5N1, particularly clade 2.3.4.4b, has demonstrated an unprecedented capacity for cross-species transmission, with recent reports confirming its presence in dairy cattle in the United States in 2024. This unexpected spillover challenges traditional understanding of the virus's host range and raises serious public health and veterinary concerns. Infected cattle presented with clinical signs such as decreased milk production, thickened or discolored milk, respiratory symptoms, and lethargy. Pathological findings revealed inflammation of the mammary glands and the detection of a virus in nasal secretions and raw milk, suggesting a potential for both intra- and interspecies transmission. While the current risk of human-to-human transmission remains low, the detection of H5N1 in a human exposed to infected cattle highlights the need for heightened surveillance and protective measures. Moreover, the presence of infectious viruses in the food chain, particularly in unpasteurized milk, introduces a new dimension of zoonotic risk. This review synthesizes emerging evidence on the epidemiology, pathology, diagnostic findings, and zoonotic implications of HPAI H5N1 infection in cattle. It also highlights the importance of genomic surveillance, intersectoral collaboration, and One Health approaches in managing this evolving threat. As the virus continues to circulate and adapt across diverse hosts, including wild birds, domestic poultry, and now mammals, the potential for reassortment and emergence of novel strains remains a significant concern. Immediate actions to strengthen biosecurity, monitor viral evolution, and protect both animal and human populations are critical to mitigate the global risk posed by this expanding panzootic.
Keywords:
H5N1
; cattle
; highly pathogenic avian influenza
; zoonosis
; bovine
; One Health
Introduction
The influenza virus belongs to the family Orthomyxoviridae and is classified into four types: A, B, C, and D. In particular, influenza A virus presents a wide variety of combinations between two surface proteins: hemagglutinin (H) and neuraminidase (N), with 18 known subtypes of H and 11 of N [1,2]. However, the five subtypes of most significant epidemiological relevance are H1N1, H2N2, H3N2, H5N1, and H7N9 [3,4,5,6].
Influenza A has been responsible for most of the pandemics documented in human history [7], including: the Spanish flu (H1N1) in 1918, the Asian flu (H2N2) in 1957, the Hong Kong flu (H3N2) in 1968 [8], and swine flu (H1N1), the result of a mutation of the original virus, in 2009 [9]. The first outbreaks of avian influenza by the H5N1 subtype were also recorded in poultry in Scotland, United Kingdom (UK), in 1959 [10] and geese in Guangdong Province, China, in 1996 [11,12].
It has been reported that the avian influenza virus is classified into two groups based on the intravenous pathogenicity index (IVPI), with highly pathogenic avian influenza (HPAI) and low pathogenic avian influenza (LPAI) [10,13,14]. In particular, the avian influenza virus subtype H5N1 is considered highly pathogenic (HPAI) [15] being the migratory birds of the order Anseriformes (e.g., geese, swans, ducks) and Chatradriformes (e.g., gulls, terns), their main reservoir [16]. In addition, H5N1 is characterized by having several clades, or genetic subdivisions, while clade 2.3.4.4b is the one that has been recognized since 2016 [17].
Clade 2.3.4.4b spread globally through migratory birds; however, initial outbreaks and mortalities were mainly limited to avian species, including poultry [17]. HPAI H5N1 clade 2.3.4.4b is currently distributed in Europe, Africa, Asia, North America (including the United States of America [USA] and Canada), and South America (comprising Colombia, Peru, Ecuador, Venezuela, Chile, Argentina, Bolivia, and Uruguay) (Figure 1) [18,19,20,21]. The mutation has allowed the virus to adapt to various conditions and spillover to multiple especies and humans [22], since it is responsible for the unusual outbreaks that have affected more than 43 species of mammals, such as minks, seals, rodents, opossums, raccoons, lions, cats, goats, and, with great concern, cattle [15,16,19,23,24,25,26,27,28]. Similarly, it has raised alarms due to its zoonotic potential after it was isolated as a causal agent of infection in humans [29,30].
In 2024, an outbreak of HPAI H5N1 clade 2.3.4.4b started in dairy cows in the USA (Figure 2) [31,32,33,34,35,36,37]. This event is considered concerning as well as unusual, given that cows routinely contract influenza viruses, but when they do, the one involved is type D. Therefore, this is the first time that a highly pathogenic strain has been found in this animal species [38]. However, although the first global report of an outbreak in cows occurred in 2024 in the USA, evidence of cattle susceptibility to HPAI H5N1 has been present since 2008, as shown in experimental research conducted by Kalthoff et al. [39]. The virus was inoculated in four calves, and although the animals remained healthy, seroconversion occurred in all of them, thus demonstrating that the virus had the potential to infect bovine calves.
Since the first notification of HPAI H5N1 infection in dairy cows in Texas, USA, in March 2024, rapid spread has occurred to more than 380 farms in 14 states [40]. It was determined that the infected animals cohabited or were close to poultry or wild bird populations [32]. It has also been suggested that cow-to-cow transmission could have occurred through respiratory fluids [37]. The clinical manifestations developed by infected cows were fever, lethargy, loss of appetite, runny nose, mastitis, decreased milk production, thickening of milk, reduced rumination, and changes in fecal consistency [26]. This outbreak has represented substantial economic losses for the livestock sector due to the decrease in the productive and reproductive parameters of the animals [19,33].
Cases of human infection with HPAI H5N1 in the U.S. have raised multiple questions due to the zoonotic characteristics of the virus (Figure 3) [41]. In August 2024, the country reported 14 cases of sick people, whose symptoms ranged from conjunctivitis to mild or moderate respiratory injuries, and they also reported having had previous exposure to cattle [42]. In December of that year, the figure rose to 59 people [43]. By January 2025, the Centers for Disease Control and Prevention (CDC) reported that a total of 67 confirmed cases of H5N1 avian influenza in humans were registered in the U.S., with the news that in the state of Louisiana the death of a person previously hospitalized with a severe case of avian influenza (H5N1) was reported. Becoming the first human fatal case in the country (Figure 3) [44].
The current panzootic caused by the HPAI H5N1 virus that has infected cattle in the U.S. could become one of the most important due to its geographic range, number of infected animal species, zoonotic risk, and significant economic losses [45]. Although to date the transmission of the virus from person to person has not been proven, the characteristics of genetic mutation and ecological adaptation exhibited by the virus have made it an agent of great concern due to its pandemic potential [46].
The outbreak reported in cattle has generated multiple reactions about the imminent health risks that loom over the human population, since this virus has been detected in unpasteurized milk and wastewater, threatening to enter the food chain [47,48]. Highly pathogenic avian influenza (HPAI) H5N1 highlights the need for One Health approaches by generating multisectoral collaborations involving entities at the municipal, departmental, national, and international levels [49]. In addition to the above, cattle farming is one of the most critical livestock activities worldwide, which increases the risk of zoonoses to humans, either by direct exposure of workers on farms with infected animals or by the consumption of unpasteurized milk and derived dairy products [50]. In this article, we will discuss the zoonotic, epidemiological, ecological, and clinical aspects, as well as the origin, distribution, and nature of the avian influenza (H5N1) virus in cattle.
Nature and Classification of Influenzavirus
The National Center for Biotechnology Information (NCBI) proposes that the taxonomy of influenza viruses is classified as Domain: Riboviria; Kingdom: Orthornavirae; Phylum: Negarnaviricota; Class: Insthoviricetes; Order: Articulavirales, and Family: Orthomyxoviridae (NCBI:txid11320) [51]. Regarding the genera, five were initially described within this family: Influenzavirus A, Influenza B, Influenzavirus C, Thogotovirus, and Isavirus [52].
Today, the virus is recognized as Alphainfluenzavirus (influenza A virus), the Beta-influenza (influenza B virus), the Gamma-influenza (influenza C virus), and the Delta-influenza (influenza D virus) (Figure 4) [1]. Influenza A and B viruses have eight segments of negative-sense RNA with two major surface glycoproteins: hemagglutinin (HA) and neuraminidase (NA), whereas influenza viruses C and D consist of seven segments of negative-sense RNA with only one major surface glycoprotein, hemagglutinin-esterase-fusion (HEF) [53].
The influenza D virus causes respiratory disease in pigs and ruminants; the influenza C virus colonizes the upper respiratory tract and induces influenza mainly in children; the influenza B virus produces respiratory symptoms having tropism through the upper and lower respiratory tract in adults; and the influenza A virus, whose symptoms are similar to B, is of global concern to health authorities since the antigenic drift of its main glycoprotein (hemagglutinin) allows it to mutate easily, adapting to various ecological and host contexts (humans and animals) [54].
Influenza A viruses encode at least ten major proteins: hemagglutinin (HA) (Figure 5), basic polymerase 1 and 2 (PB1-PB2), acid polymerase (PA), nucleoprotein (NP), neuraminidase (NA), matrix 1 and 2 (M1-M2), nonstructural protein 1 and 2 (NS1-NS2), nuclear export protein (NEP) and others with specific or as yet unknown functions such as PB1-F2, PB1-N40, PA-X, and PA-N155 and PA-N182 [55].
The replication cycle of this virus is a complex process [56,57]. It begins when the virus attaches to the host cell via hemagglutinin (HA), which interacts with sialic acid receptors on the cell surface [58]. After the virus enters through endocytosis, the low pH in the endosome facilitates the fusion of the viral envelope with the endosome membrane, releasing the viral genome into the cytoplasm [59]. The genetic material is transported to the nucleus where the viral RNA polymerase initiates the transcription and replication of the viral RNA; transcription generates positive sense mRNA for viral protein synthesis in the cytoplasm and complementary RNA (cRNA) that will serve as a template for new copies of viral genomics (vRNA) [60]. The viral proteins hemagglutinin (HA), Neuraminidase (NA), and Matrix 2 (M2) are synthesized in the endoplasmic reticulum and then transported to the Golgi apparatus, where they are directed to the plasma membrane [1].
The assembly of the virion occurs near the plasma membrane, where the viral ribonucleoprotein complex (vRNP) and structural proteins bind to the cell’s plasma membrane to use it as an envelope, while neuraminidase facilitates the release of viral particles by cutting the sialic acid bonds between the virus and the host cell [61]. As a result, new viral particles are released by budding and can infect other cells, starting a new cycle of infection [62].
The key factor in virus entry into an organism is the hemagglutinin surface glycoprotein HA (strongly expressed in influenza A virus), which contains the host receptor binding site to allow the virus particle to attach to specific host cells, the fusion peptide that is inserted into the target cell membrane during membrane fusion, and other structural elements that can retract during the Membrane Fusion Process [63]. Since HA is a surface glycoprotein of the virus particle, it is easily recognized by antibodies, however, the virus can evade the host’s immune system through unique mechanisms, as well, during its replication, viral RNA polymerase makes frequent mistakes, allowing the creation of HA variants that will allow the virus to become resistant to existing antibodies, facilitating immune system evasion and virus replication in the infected host [63,64].
Undoubtedly, influenza A viruses, which can cause severe pneumonia and ultimately lead to death, pose a significant threat to human health, as the virus can recombine segments, resulting in the emergence of pandemic strains for which there is little pre-existing immunity in the human population [53,65,66]. This condition of mutation has allowed it to be the virus with the highest number of pandemics in the history of humanity, as well as being responsible for significant human mortality figures in recent times [8], such is the case of the Spanish flu of 1918 that left a balance of around 21 million people dead [67], surpassing even COVID-19, which until February 2025 registered more than 7 million deaths [68].
Within the set of influenza A viruses, the H5, H6, H7, H9 and H10 subtypes are associated with avian influenza, which is distinguished from seasonal/pandemic influenza or influenza (H1N1, H3N2) because the former may have highly pathogenic strains that are capable of inducing greater damage to the organs and systems of the body and therefore a more serious disease in animals and humans [10,15]. The H5N1 strain of highly pathogenic avian influenza (HPAI) shows a basic reproductive index (R0) which varies between 0.1-1.14, as well as a prevalence of 58 – 84% in the human population exposed to poultry, additionally, HPAI H5N1 and HPAI H7N9 show higher mortality rates than other subtypes such as H1N1, H2N2 and H3N2 [3]. In the same way, the evidence of HPAI H5N1’s ability to adapt and mutate is striking and indicates that this virus has evolved in recent years, since in 2008 clade 2.3.4.4 was registered, and since 2016 another genetic subdivision had already been isolated, clade 2.3.4.4b [17]. Also, it is known that until before 2020, the neuraminidase (NA) protein of subtype N1 of clade 2.3.4.4b had a truncated peduncular domain that mediated virulence factors in poultry; recently, with new infections, it has been possible to determine that the vast majority of highly pathogenic H5N1 viruses of clade 2.3.4.4b currently circulating worldwide have the extended peduncular version of neuraminidase, which may increase the risk of these viruses being transmitted between humans [69].
Worldwide Distribution of H5N1
Historically, since their first appearances in Scotland (UK) and China in 1959 and 1996, respectively, H5N1 HPAI had been limited to poultry; however, the mutation allowed the virus to adapt and infect wild birds [10,11,47]. This situation has led to an increasing number of infections in waterfowl, raptors, and peridomestic species in recent years [16]. The rapid intercontinental spread of the virus, facilitated by the migration of infected birds, the genomic rearrangements of the virus, and the unusual spread in terrestrial and marine mammals, has triggered a panzootic of HPAI H5N1 and raised significant pandemic risk concerns [7]. More than two decades after HPAI H5N1 became established in poultry in Southeast Asia, the virus has spread to various countries in Africa, South America, North America, Europe, and Antarctica [70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88].
As previously reported, the HPAI H5N1 virus is distributed globally and has infected multiple animal species [89]. In Chile, important implications in wild birds were reported with the death of Peruvian pelicans (Pelecanus thagus), Franklin’s gull (Larus pipixcan), grey gull (Leucophaeus modestus), elegant tern (Thalasseus elegans) and black grater (Rynchops niger) [90,91]. In Peru, mass deaths of common dolphins (Delphinus delphis) and South American sea lion (Otaria flavescens)[77]. Similarly, in this country, a case was documented in neotropical cormorants (Nannopterum brasilianum) and a lion (Panthera leo) [15]. In Punta Bermeja, the largest colony of South American sea lions in Argentina, more than 800 animals of this species died [21]. Also, rodents Rattus norvegicus and Rattus rattus were HIV-positive in the Giza region, Egypt [92].
Regarding domestic mammals, there is a history of HPAI H5N1 seropositivity in cats, dogs, and, more recently, in cattle [26,93]. In Poland, a case was registered of a male cat, neutered, approximately 6 years old, with a history of feeding raw chicken meat, who was taken to the clinic for symptoms of apathy and anorexia that, after three days, evolved into respiratory distress and neurological signs that led to the death of the animal [94]. In France, the virus was detected in a domestic cat that lived near a duck farm [95]. In South Dakota, USA, 10 cats died, whose postmortem study reflected lesions and viral antigens in different organs, mainly in the brain [96]. Similarly, on a dairy farm in the USA, cats of different ages died after being fed raw milk from cows [17]. Curiously, some countries (e.g., Slovenia) even still sell fresh raw milk in public street vending machines in the city of Ljubljana (Figure 6).
The concern of H5N1 transmission in cats focuses on the consumption of infected birds and contaminated milk; however, the anthropozoonotic risk is often overlooked. In Michigan, USA a cat that did not have access to the outside world tested positive for the virus and developed clinical manifestations with neurological deterioration, the owner reported that he worked on H5N1-positive dairy farms and transported unpasteurized milk, did not wear protective equipment and when he arrived at the house the cat rolled around in his clothes [97].
In Washington, USA, antibodies to HPAI H5N1 virus were detected in 4/194 (2%) dogs that hunted or participated in hunting and training trials with wild birds [98]. It was also shown that inoculation of the virus in Beagle dogs resulted in excretion of the virus and rapid seroconversion without disease [99]. For their part, in an investigation carried out in China, they found that, after infecting dogs with the virus intranasally and intratracheally, they developed disease, including anorexia, fever, conjunctivitis, respiratory distress, cough, and even death [100].
Something particular happens with HPAI H5N1 in pigs, because even though in experimental infections it has been shown that it causes interstitial pneumonia with necrotizing bronchiolitis, with high viral titers in the lower respiratory tract and 100% seroconversion, infected animals eliminate a limited amount of the virus, in addition to not transmitting by contact [101]. For this reason, pigs are considered to serve as intermediate hosts and play an essential role in the genetic recombination of the virus, to make an inter-species leap that can easily trigger an avian influenza pandemic [102].
History of Influenza Virus Infection in Cattle
It should be noted that, although the first natural infection with influenza A virus in cattle was recorded during the 2024 outbreak, cases of natural infection with influenza D (IDV) had already been documented in these animals [103]. This virus was initially isolated in Oklahoma, USA, from a 15-week-old pig that showed influenza-like symptoms [104]. Later evidence showed that the IDV is more common in cattle than in pigs, which is why it is believed that cattle act as the main reservoir of the virus [53]. Although the initial detection occurred in pigs in 2011, serological surveillance suggests that IDV has been present in cattle since at least 2004[105].
Calves and fattening cattle are often the most susceptible to IDV infection [105,106]. In recent years there has been increasing evidence supporting the theory that, along with the virus Bovine Viral Diarrhea (BVDV), Bovine Herpesvirus 1 (BHV-1), Bovine Respiratory Syncytial Virus (BRSV) and Parainfluenza Virus Type 3 (PI3), Influenza D virus (IDV) is linked to the bovine respiratory disease complex [107]. However, findings of positive samples for IDV not only from cattle showing clinical signs associated with bovine respiratory disease, but also from healthy cattle, would demonstrate that the virus can cause asymptomatic infection [108].
IDV has been distributed around the world and has been associated with livestock infections in several countries such as the USA, Japan, China, France, Italy, Ireland, Luxembourg, and Brazil [106,109,110,111,112]. In turn, in addition to cattle and pigs, IDV antibodies have been detected in goats, sheep, horses, camels, and buffalo [54,108]. Similarly, there is evidence of infection in exposed and non-exposed persons to livestock, indicating that influenza virus type D, like type A, has zoonotic potential [53].
H5N1 in Cattle
Due to the outbreak of the HPAI H5N1 virus in cattle in the United States during 2024, there was considerable concern about the implications for public health and food safety [48]. However, it should be clarified that in this country, detection of the virus in more than 200 mammals has been reported since 2022 [113]. These facts support the evolutionary line of HPAI H5N1 in the US, as the virus has been detected in poultry and wild birds since its entry through the Canadian border in 2021, and subsequently in cats, goats, and cattle [19]. On March 20, 2024, the Minnesota Board of Animal Health reported that a kid raised on a farm in Stevens County, west-central Minnesota, tested positive for highly pathogenic avian influenza (HPAI); an outbreak of HPAI H5N1 in poultry had recently been detected on this same farm [114]. In turn, on March 25, 2024, the first outbreak in the world of HPAI H5N1 clade 2.3.4.4b virus was reported in dairy cows in three US states: Texas, Kansas and New Mexico, subsequently, on March 29, 2024, the virus was detected in cattle from a farm in Michigan that had recently imported animals from Texas and from this cases were reported in different states of the country [113,114,115,116,117,118]. Phylogenetic analysis of HPAI H5N1-positive samples, clade 2.3.4.4b, in dairy cattle indicated that the reassociation of the virus likely occurred in wild birds, followed by transmission to dairy cattle at the end of 2023 [119]. Following the introduction of the virus in cattle, its spread in the U.S. was facilitated by the movement of asymptomatic dairy cattle, which infected other animal species and humans [19].
In dairy cattle, HPAI H5N1 clade 2.3.4.4b infection presented with fever, lethargy, dehydration, loss of appetite, clear nasal discharge, increased breathing rate, difficulty breathing, reduced rumination, changes in stool consistency, such as diarrhea or dry stools, involution of the mammary gland in several of the affected cows, in addition to a notable drop in milk production that even became with an abnormal yellowish color similar to colostrum, thick and sometimes curdled consistency [19,26,29,33,37]. In a study carried out by Caserta et al. [25]. The virus was reported to have caused mortality in some infected cows. Meanwhile, cattle that managed to recover experienced a clinical illness that lasted between 5 and 14 days, but they continued to exhibit decreased milk production for at least 4 weeks.
One of the main determining factors in the virus-host relationship is the availability of receptors that allow viral development, particularly, influenza A viruses use sialic acids from the host as their receptors for the initial binding of the hemagglutinin protein (HA) and entry into cells, in that sense, it has been reported that the respiratory and mammary glands of dairy cattle naturally infected in the USA with HPAI H5N1 are rich in avian influenza virus-specific sialic acid, suggesting virus epitheliotropism in the mammary gland of cattle [27,120]. It has also been observed that the bovine H5N1 virus replicates efficiently in the epithelium of the glandular cistern and teats of dairy cows, demonstrating that these viruses can invade the mammary gland through the teat canal [121].
Raw milk from infected animals has been reported to contain a high viral load, which is a potential source of dissemination and spread of this virus [122]. While it is true that the milk of sick cows is not commercialized [38]. It was recently announced that milk sold in 10 U.S. states contained H5N1 fragments [33]. In addition, it has been found that the virus can remain infectious on fomites and milking equipment materials for several hours [123,124]. In addition, research has concluded that pasteurization is the only effective method to achieve viral inactivation of HPAI H5N1 in cattle milk [122,125]. However, it cannot be ignored that cattle infected during the outbreak in the U.S. also presented high viral titers in nasal swab samples; therefore, while it is true that milk and fomites are essential sources of infection, respiratory fluids also generate concern [37].
Something very striking is that two years before the detection of HPAI H5N1 in dairy cattle, the virus was already detected in wastewater in the U.S., with no relation or presence of dairy processing facilities or dairy farms within the sewer basin [126]. Subsequently, in investigations carried out during the 2024 outbreak, the virus was again detected in wastewater whose basin included milk processing sources [48].
Although the mechanism by which cattle were infected in the U.S. is unclear, official reports indicate that the farms where the outbreaks occurred had reported simultaneous mortality events in poultry and wild birds [24]. So far, the most reasonable explanation for HPAI H5N1 infection in cattle is related to contact with fluids or tissues from infected birds, as different studies have shown that the excretion of the virus in birds occurs through the oropharyngeal and cloacal pathways, with maximum expression in the oropharyngeal pathway [127,128,129]. On the other hand, the entry of the virus into mammals can occur nasally or oropharyngeally [39,130].
H5N1 in Humans
The first case of HPAI H5N1 infection in the world was reported in the Hong Kong Special Administrative Region (China) in 1997. Since then, the virus reappeared in 2003 and continues to infect people around the world, posing a potential pandemic threat due to its continued global spread and evolution [131]. Although H5N1 has not adapted to human-to-human transmission in nearly thirty years, currently, the abundant circulation in various animal species, including mammals, increases the possibility of recombination of new pandemic strains [132].
The World Health Organization (WHO) has reported a total of 954 cases of H5N1 infection in humans with 464 deaths, from January 2003 to December 2024 worldwide (Table 1). Cases have been reported in 24 countries, with Egypt (n = 359), Indonesia (n = 200), Vietnam (n = 129), Cambodia (n = 72), and in the US (n = 59) being the countries with the highest number of cases, accounting for 85% of the total cases reported worldwide. Indonesia is the country with the highest mortality rate compared to its reported cases [43].
According to CDC figures, by January 6, 2025, the cumulative number of human HPAI H5N1 infections in the U.S. had risen from 59 to 67 cases, and the first death was reported [44]. It is estimated that most documented human infections in the U.S. resulted from dairy farm workers’ exposure to unpasteurized milk during milking [32,124,133,134]. People infected with HPAI H5N1 developed mild respiratory symptoms and conjunctivitis [42], or were asymptomatic, as in the case of three veterinarians with positive serological results for the highly pathogenic avian influenza A(H5) virus [135]. On the other hand, a case was reported of a 65-year-old deceased person with underlying medical conditions who, after being exposed to a flock of non-commercial backyard birds and wild birds, developed severe respiratory complications [136].
On April 01, 2024, the first case of suspected transmission of HPAI H5N1 virus from cow to human was reported in Texas, USA, after CDC confirmed that a worker on a commercial dairy farm tested positive by real-time reverse transcriptase polymerase chain reaction (RT-qPCR) for HPAI H5N1 clade 2.3.4.4b infection; the patient only experienced conjunctivitis with no other signs or symptoms, was treated with oseltamivir and recovered; however, no disease was identified among the patient’s household members, all of whom received post-exposure prophylaxis with the same antiviral [32].
Subsequently, in May 2024, two adult dairy farm workers in Michigan USA were identified with infection with the virus; one of these, presented discomfort in his right eye a day after milk splashed in his eye while milking a cow, meanwhile, the other worker, from another farm, presented cough, difficulty breathing, headache, sore throat, fatigue, nasal congestion and rhinitis; The tasks of this worker mainly included the care of sick cows, including administration of drugs orally, it should be mentioned that the worker used eye protection and gloves, but did not use a respirator or mask [137].
Concerning the human immune response against HPAI H5N1, studies are limited, however, research has been carried out in animals such as the study in ferrets, in which they found that animals infected with H5N1 and that had previously been exposed to the H1N1 virus had a cross-reaction with the neuraminidase (NA) protein of H5N1. This was evidenced by the low viral load present in nasal secretions and organs external to the respiratory tract, thus suggesting that humans with immunity to the H1N1 virus may experience milder illness due to the influenza A H5N1 virus strain [134]. Likewise, it has been reported that the H5N1 vaccines available in the U.S., which are of a different strain and clade than the one presented in the 2024 outbreak, generate neutralizing crossover antibodies against H5N1 of the clade 2.3.4.4b circulating in humans and may be helpful as temporary alternate vaccines while a specific one for the emerging clade 2.3.4.4b is developed [31].
In this sense, the unprecedented intercontinental spread that since 2020 has been occurring with the highly pathogenic avian influenza (H5N1) virus to multiple species of animals such as birds, mammals and humans has generated alarms and concerns to public health due to its zoonotic and pandemic potential, therefore, it is essential to continue implementing and strengthening research, international cooperation and active surveillance of the virus in different hosts to establish prevention and control strategies that safeguard the world population, including H5N1 in cattle (Table 2) [138].
Limitations
This review is limited by the rapidly evolving nature of the HPAI H5N1 outbreak in cattle, with much of the available data being preliminary and subject to revision. Many findings are based on case reports, early surveillance data, and experimental studies, which may not capture the full scope of viral transmission dynamics or clinical outcomes. Additionally, there is a limited amount of peer-reviewed literature specifically on bovine H5N1 infections, which constrains comprehensive analysis. Future studies with broader epidemiological and molecular data will be essential to validate and expand upon current observations.
Conclusions
The emergence of highly pathogenic avian influenza (HPAI) H5N1 in cattle, particularly the clade 2.3.4.4b strain identified in the United States in 2024, marks a concerning development in the epidemiology of zoonotic diseases. Historically restricted to avian species and select mammals, H5N1’s confirmed ability to infect bovines underscores the virus’s expanding host range and adaptive capacity. Clinical evidence, including significant declines in milk production, mammary gland pathology, and respiratory distress, highlights the impact on livestock health and productivity. Moreover, the detection of infectious viral particles in raw milk and nasal secretions introduces potential risks to the food chain and public health.
Although human-to-human transmission remains unproven, confirmed human infections linked to exposure to infected cattle and unpasteurized milk demand heightened vigilance. The possibility of further interspecies transmission and viral reassortment elevates the risk of a future pandemic scenario. Given the significant economic losses and public health implications, coordinated surveillance, rapid diagnostics, and stringent biosecurity measures are imperative.
This situation calls for a robust One Health approach—integrating human, animal, and environmental health disciplines—to monitor and mitigate the ongoing panzootic threat. Continued genomic surveillance and international collaboration will be crucial for tracking viral evolution and adapting preventive strategies. In conclusion, the unprecedented spillover of HPAI H5N1 into cattle not only challenges current paradigms of influenza virus ecology but also highlights urgent gaps in preparedness and response capacities that must be addressed globally.
Author Contributions
I.C.S.R: Conceptualisation, methodology, formal analysis, investigation, writing—original draft preparation, and writing—review and editing; C.L.S.J.: Conceptualisation, methodology, formal analysis, investigation, writing—original draft preparation, and writing—review and editing; J.L.B.A: Conceptualisation, methodology, formal analysis, investigation, writing—original draft preparation, and writing—review and editing; J.D.A-E: Conceptualisation, formal analysis, investigation, writing—original draft preparation, and writing—review and editing; D.K.B.A.: Conceptualisation, formal analysis, investigation, writing—original draft preparation, and writing—review and editing; A.J.RM.: Conceptualisation, investigation, writing—original draft preparation, and writing review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
Pontificia Universidad Católica del Ecuador covered the APC in this article.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Available upon reasonable request.
Acknowledgements
This article has been registered in the Research Proposal Registration of the Coordination of Scientific Integrity and Surveillance of Universidad Cientifica del Sur, Lima, Peru.
Conflicts of Interest
No conflicts of interest.
References
- Hutchinson, E.C. Influenza Virus. Trends Microbiol 2018, 26, 809–810. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Jayaswal, N.; Kumar, S.; Rao, G.; Budha, R.R.; Mohanty, A.; Mehta, R.; Apostolopoulos, V.; Sah, S.; Bonilla-Aldana, D.K.; et al. Targeting H3N2 influenza: advancements in treatment and vaccine strategies. Expert Rev Anti Infect Ther 2025, 23, 5–18. [Google Scholar] [CrossRef]
- Park, J.E.; Ryu, Y. Transmissibility and severity of influenza virus by subtype. Infect Genet Evol 2018, 65, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Morales, A.J.; Hui, D.S.; Ippolito, G.; Traore, T.; Satta, G.; Everett, D.B.; Zumla, A. Avian Influenza - The next travel-associated pandemic? Proactive One Health surveillance is required to reduce the risk of the spread. Travel Med Infect Dis 2025, 65, 102829. [Google Scholar] [CrossRef]
- Chaudhary, R.K.; L, A.; Patil, P.; Mateti, U.V.; Sah, S.; Mohanty, A.; Rath, R.S.; Padhi, B.K.; Malik, S.; Jassim, K.H.; et al. System Biology Approach to Identify the Hub Genes and Pathways Associated with Human H5N1 Infection. Vaccines (Basel) 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, *!!! REPLACE !!!*; Khandia, R.; Chopra, H.; Choudhary, O.P.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. The re-emergence of H3N2 influenza: An update on the risk and containment. New Microbes New Infect 2023, 53, 101147. [Google Scholar] [CrossRef]
- Peacock, T.P.; Moncla, L.; Dudas, G.; VanInsberghe, D.; Sukhova, K.; Lloyd-Smith, J.O.; Worobey, M.; Lowen, A.C.; Nelson, M.I. The global H5N1 influenza panzootic in mammals. Nature 2025, 637, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Kilbourne, E.D. Influenza pandemics of the 20th century. Emerg Infect Dis 2006, 12, 9–14. [Google Scholar] [CrossRef]
- Ankomah, A.A.; Moa, A.; Chughtai, A.A. The long road of pandemic vaccine development to rollout: A systematic review on the lessons learnt from the 2009 H1N1 influenza pandemic. Am J Infect Control 2022, 50, 735–742. [Google Scholar] [CrossRef]
- Charostad, J.; Rezaei Zadeh Rukerd, M.; Mahmoudvand, S.; Bashash, D.; Hashemi, S.M.A.; Nakhaie, M.; Zandi, K. A comprehensive review of highly pathogenic avian influenza (HPAI) H5N1: An imminent threat at doorstep. Travel Med Infect Dis 2023, 55, 102638. [Google Scholar] [CrossRef]
- Guan, Y.; Smith, G.J. The emergence and diversification of panzootic H5N1 influenza viruses. Virus Res 2013, 178, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulos, V.; Chavda, V.P.; Mehta, R.; Rodriguez-Morales, A.J.; Henao-MartÍnez, A.F.; Sah, R. Alert and surveillance on H5N1 influenza virus: risks to agriculture and public health. Ther Adv Infect Dis 2024, 11, 20499361241266521. [Google Scholar] [CrossRef]
- Rodriguez-Morales, A.J.; Bonilla-Aldana, D.K.; Paniz-Mondolfi, A.E. Concerns about influenza H5N8 outbreaks in humans and birds: Facing the next airborne pandemic? Travel Med Infect Dis 2021, 41, 102054. [Google Scholar] [CrossRef] [PubMed]
- Bonilla-Aldana, D.K.; Aguirre-Florez, M.; Villamizar-Peña, R.; Gutiérrez-Ocampo, E.; Henao-Martínez, J.F.; Cvetkovic-Vega, A.; Dhama, K.; Rabaan, A.; Sah, R.; Rodriguez-Morales, A.J.; et al. After SARS-CoV-2, will H5N6 and other influenza viruses follow the pandemic path? Infez Med 2020, 28, 475–485. [Google Scholar] [PubMed]
- Cruz, C.D.; Icochea, M.E.; Espejo, V.; Troncos, G.; Castro-Sanguinetti, G.R.; Schilling, M.A.; Tinoco, Y. Highly Pathogenic Avian Influenza A(H5N1) from Wild Birds, Poultry, and Mammals, Peru. Emerg Infect Dis 2023, 29, 2572–2576. [Google Scholar] [CrossRef]
- Neumann, G.; Kawaoka, Y. Highly pathogenic H5N1 avian influenza virus outbreak in cattle: the knowns and unknowns. Nature Reviews Microbiology 2024, 22, 525–526. [Google Scholar] [CrossRef]
- Hu, X.; Saxena, A.; Magstadt, D.R.; Gauger, P.C.; Burrough, E.R.; Zhang, J.; Siepker, C.; Mainenti, M.; Gorden, P.J.; Plummer, P.J.; et al. Genomic characterization of highly pathogenic avian influenza A H5N1 virus newly emerged in dairy cattle. Emerg Microbes Infect 2024, 13, 2380421. [Google Scholar] [CrossRef]
- Cui, P.; Shi, J.; Wang, C.; Zhang, Y.; Xing, X.; Kong, H.; Yan, C.; Zeng, X.; Liu, L.; Tian, G.; et al. Global dissemination of H5N1 influenza viruses bearing the clade 2.3.4.4b HA gene and biologic analysis of the ones detected in China. Emerg Microbes Infect 2022, 11, 1693–1704. [Google Scholar] [CrossRef]
- Sreenivasan, C.C.; Li, F.; Wang, D. Emerging Threats of Highly Pathogenic Avian Influenza A (H5N1) in US Dairy Cattle: Understanding Cross-Species Transmission Dynamics in Mammalian Hosts. Viruses 2024, 16. [Google Scholar] [CrossRef]
- Ruiz-Saenz, J.; Martinez-Gutierrez, M.; Pujol, F.H. Multiple introductions of highly pathogenic avian influenza H5N1 clade 2.3.4.4b into South America. Travel Med Infect Dis 2023, 53, 102591. [Google Scholar] [CrossRef]
- Rimondi, A.; Vanstreels, R.E.T.; Olivera, V.; Donini, A.; Lauriente, M.M.; Uhart, M.M. Highly Pathogenic Avian Influenza A(H5N1) Viruses from Multispecies Outbreak, Argentina, August 2023. Emerg Infect Dis 2024, 30, 812–814. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Haque, S.; Tawil, S.; Husni, R.; Bonilla-Aldana, D.K.; Montenegro-Idrogo, J.J.; Rodriguez-Morales, A.J. Avian influenza spillover to humans: Are we prepared to deal with another potential pandemic? Travel Med Infect Dis 2023, 55, 102634. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Yeom, M.; Vu, T.T.H.; Do, H.Q.; Na, W.; Lee, M.; Jeong, D.G.; Cheon, D.S.; Song, D. Characterization of highly pathogenic avian influenza A (H5N1) viruses isolated from cats in South Korea, 2023. Emerg Microbes Infect 2024, 13, 2290835. [Google Scholar] [CrossRef]
- Mostafa, A.; Naguib, M.M.; Nogales, A.; Barre, R.S.; Stewart, J.P.; García-Sastre, A.; Martinez-Sobrido, L. Avian influenza A (H5N1) virus in dairy cattle: origin, evolution, and cross-species transmission. mBio 2024, 15, e0254224. [Google Scholar] [CrossRef] [PubMed]
- Caserta, L.C.; Frye, E.A.; Butt, S.L.; Laverack, M.; Nooruzzaman, M.; Covaleda, L.M.; Thompson, A.C.; Koscielny, M.P.; Cronk, B.; Johnson, A.; et al. Spillover of highly pathogenic avian influenza H5N1 virus to dairy cattle. Nature 2024, 634, 669–676. [Google Scholar] [CrossRef]
- Burrough, E.R.; Magstadt, D.R.; Petersen, B.; Timmermans, S.J.; Gauger, P.C.; Zhang, J.; Siepker, C.; Mainenti, M.; Li, G.; Thompson, A.C.; et al. Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus Infection in Domestic Dairy Cattle and Cats, United States, 2024. Emerg Infect Dis 2024, 30, 1335–1343. [Google Scholar] [CrossRef]
- Nelli, R.K.; Harm, T.A.; Siepker, C.; Groeltz-Thrush, J.M.; Jones, B.; Twu, N.C.; Nenninger, A.S.; Magstadt, D.R.; Burrough, E.R.; Piñeyro, P.E.; et al. Sialic Acid Receptor Specificity in Mammary Gland of Dairy Cattle Infected with Highly Pathogenic Avian Influenza A(H5N1) Virus. Emerg Infect Dis 2024, 30, 1361–1373. [Google Scholar] [CrossRef]
- Bonilla-Aldana, D.K.; Bonilla-Aldana, J.L.; Acosta-España, J.D.; Rodriguez-Morales, A.J. Highly Pathogenic Avian Influenza H5N1 in Cats (Felis catus): A Systematic Review and Meta-Analysis. Animals (Basel) 2025, 15. [Google Scholar] [CrossRef]
- Eisfeld, A.J.; Biswas, A.; Guan, L.; Gu, C.; Maemura, T.; Trifkovic, S.; Wang, T.; Babujee, L.; Dahn, R.; Halfmann, P.J.; et al. Pathogenicity and transmissibility of bovine H5N1 influenza virus. Nature 2024, 633, 426–432. [Google Scholar] [CrossRef]
- Honein, M.A.; Olsen, S.J.; Jernigan, D.B.; Daskalakis, D.C. Challenges and Opportunities for Wastewater Monitoring of Influenza Viruses During the Multistate Outbreak of Highly Pathogenic Avian Influenza A(H5N1) Virus in Dairy Cattle and Poultry. Am J Public Health 2024, 114, 1309–1312. [Google Scholar] [CrossRef]
- Khurana, S.; King, L.R.; Manischewitz, J.; Posadas, O.; Mishra, A.K.; Liu, D.; Beigel, J.H.; Rappuoli, R.; Tsang, J.S.; Golding, H. Licensed H5N1 vaccines generate cross-neutralizing antibodies against highly pathogenic H5N1 clade 2.3.4.4b influenza virus. Nat Med 2024, 30, 2771–2776. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.; Reed, C.; Davis, C.T.; Uyeki, T.M.; Behravesh, C.B.; Kniss, K.; Budd, A.; Biggerstaff, M.; Adjemian, J.; Barnes, J.R.; et al. Outbreak of Highly Pathogenic Avian Influenza A(H5N1) Viruses in U.S. Dairy Cattle and Detection of Two Human Cases - United States, 2024. MMWR Morb Mortal Wkly Rep 2024, 73, 501–505. [Google Scholar] [CrossRef]
- Sah, R.; Srivastava, S.; Kumar, S.; Mehta, R.; Donovan, S.; Sierra-Carrero, L.; Luna, C.; Woc-Colburn, L.; Cardona-Ospina, J.A.; Hinestroza-Jordan, M.; et al. Concerns on H5N1 avian influenza given the outbreak in U.S. dairy cattle. Lancet Reg Health Am 2024, 35, 100785. [Google Scholar] [CrossRef]
- Lowen, A.C.; Baker, A.L.; Bowman, A.S.; García-Sastre, A.; Hensley, S.E.; Lakdawala, S.S.; Moncla, L.H.; Nelson, M.I.; Pekosz, A.; Poulson, R.L.; et al. Pandemic risk stemming from the bovine H5N1 outbreak: an account of the knowns and unknowns. J Virol 2025, e0005225. [Google Scholar] [CrossRef]
- Chakraborty, C.; Bhattacharya, M. Evolution and mutational landscape of highly pathogenic avian influenza strain A(H5N1) in the current outbreak in the USA and global landscape. Virology 2024, 600, 110246. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, C.; Jensen, H.E.; Trebbien, R.; Webby, R.J.; Larsen, L.E. Avian and Human Influenza A Virus Receptors in Bovine Mammary Gland. Emerg Infect Dis 2024, 30, 1907–1911. [Google Scholar] [CrossRef] [PubMed]
- Oguzie, J.U.; Marushchak, L.V.; Shittu, I.; Lednicky, J.A.; Miller, A.L.; Hao, H.; Nelson, M.I.; Gray, G.C. Avian Influenza A(H5N1) Virus among Dairy Cattle, Texas, USA. Emerg Infect Dis 2024, 30, 1425–1429. [Google Scholar] [CrossRef]
- Cohen, J. Worries about bird flu in U.S. cattle intensify. Science 2024, 384, 12–13. [Google Scholar] [CrossRef]
- Kalthoff, D.; Hoffmann, B.; Harder, T.; Durban, M.; Beer, M. Experimental infection of cattle with highly pathogenic avian influenza virus (H5N1). Emerg Infect Dis 2008, 14, 1132–1134. [Google Scholar] [CrossRef]
- Halwe, N.J.; Cool, K.; Breithaupt, A.; Schön, J.; Trujillo, J.D.; Nooruzzaman, M.; Kwon, T.; Ahrens, A.K.; Britzke, T.; McDowell, C.D.; et al. H5N1 clade 2.3.4.4b dynamics in experimentally infected calves and cows. Nature 2025, 637, 903–912. [Google Scholar] [CrossRef]
- Perez-Acle, T.; Ravello, C.; Rosemblatt, M. Are we cultivating the perfect storm for a human avian influenza pandemic? Biol Res 2024, 57, 96. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Maemura, T.; Guan, L.; Eisfeld, A.J.; Biswas, A.; Kiso, M.; Uraki, R.; Ito, M.; Trifkovic, S.; Wang, T.; et al. A human isolate of bovine H5N1 is transmissible and lethal in animal models. Nature 2024, 636, 711–718. [Google Scholar] [CrossRef]
- WHO. Cumulative number of confirmed human cases for avian influenza A(H5N1) reported to WHO, 2003-2024. 12 December 2024. Available online: https://www.who.int/publications/m/item/cumulative-number-of-confirmed-human-cases-for-avian-influenza-a(h5n1)-reported-to-who--2003-2024--20-december-2024 (accessed on 7 March 2025).
- CDC. First H5 Bird Flu Death Reported in United States. Available online: https://www.cdc.gov/media/releases/2025/m0106-h5-birdflu-death.html (accessed on 7 March 2025).
- Plaza, P.I.; Gamarra-Toledo, V.; Euguí, J.R.; Lambertucci, S.A. Recent Changes in Patterns of Mammal Infection with Highly Pathogenic Avian Influenza A(H5N1) Virus Worldwide. Emerg Infect Dis 2024, 30, 444–452. [Google Scholar] [CrossRef]
- Parums, D.V. Editorial: Rapid Testing for the Avian Influenza A(H5N1) Virus is Urgently Required as Infections in Poultry and Dairy Cows are on the Rise, and so is Transmission to Humans. Med Sci Monit 2025, 31, e949109. [Google Scholar] [CrossRef]
- Guan, L.; Eisfeld, A.J.; Pattinson, D.; Gu, C.; Biswas, A.; Maemura, T.; Trifkovic, S.; Babujee, L.; Presler, R., Jr.; Dahn, R.; et al. Cow’s Milk Containing Avian Influenza A(H5N1) Virus - Heat Inactivation and Infectivity in Mice. N Engl J Med 2024, 391, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Louis, S.; Mark-Carew, M.; Biggerstaff, M.; Yoder, J.; Boehm, A.B.; Wolfe, M.K.; Flood, M.; Peters, S.; Stobierski, M.G.; Coyle, J.; et al. Wastewater Surveillance for Influenza A Virus and H5 Subtype Concurrent with the Highly Pathogenic Avian Influenza A(H5N1) Virus Outbreak in Cattle and Poultry and Associated Human Cases - United States, May 12-July 13, 2024. MMWR Morb Mortal Wkly Rep 2024, 73, 804–809. [Google Scholar] [CrossRef]
- Koopmans, M.P.G.; Barton Behravesh, C.; Cunningham, A.A.; Adisasmito, W.B.; Almuhairi, S.; Bilivogui, P.; Bukachi, S.A.; Casas, N.; Cediel Becerra, N.; Charron, D.F.; et al. The panzootic spread of highly pathogenic avian influenza H5N1 sublineage 2.3.4.4b: a critical appraisal of One Health preparedness and prevention. Lancet Infect Dis 2024, 24, e774–e781. [Google Scholar] [CrossRef]
- Spackman, E.; Jones, D.R.; McCoig, A.M.; Colonius, T.J.; Goraichuk, I.V.; Suarez, D.L. Characterization of highly pathogenic avian influenza virus in retail dairy products in the US. J Virol 2024, 98, e0088124. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; McVeigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: a comprehensive update on curation, resources and tools. Database (Oxford) 2020, 2020. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Kash, J.C. Influenza virus evolution, host adaptation, and pandemic formation. Cell Host Microbe 2010, 7, 440–451. [Google Scholar] [CrossRef]
- Liu, R.; Sheng, Z.; Huang, C.; Wang, D.; Li, F. Influenza D virus. Curr Opin Virol 2020, 44, 154–161. [Google Scholar] [CrossRef]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel Influenza D virus: Epidemiology, pathology, evolution and biological characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef]
- Sonnberg, S.; Webby, R.J.; Webster, R.G. Natural history of highly pathogenic avian influenza H5N1. Virus Res 2013, 178, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nature Reviews Disease Primers 2018, 4, 3. [Google Scholar] [CrossRef]
- Carter, T.; Iqbal, M. The Influenza A Virus Replication Cycle: A Comprehensive Review. Viruses 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Te Velthuis, A.J.; Fodor, E. Influenza virus RNA polymerase: insights into the mechanisms of viral RNA synthesis. Nat Rev Microbiol 2016, 14, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Lakadamyali, M.; Rust, M.J.; Zhuang, X. Endocytosis of influenza viruses. Microbes Infect 2004, 6, 929–936. [Google Scholar] [CrossRef]
- Kash, J.C.; Goodman, A.G.; Korth, M.J.; Katze, M.G. Hijacking of the host-cell response and translational control during influenza virus infection. Virus Res 2006, 119, 111–120. [Google Scholar] [CrossRef]
- Wagner, R.; Matrosovich, M.; Klenk, H.D. Functional balance between haemagglutinin and neuraminidase in influenza virus infections. Rev Med Virol 2002, 12, 159–166. [Google Scholar] [CrossRef]
- Nayak, D.P.; Hui, E.K.; Barman, S. Assembly and budding of influenza virus. Virus Res 2004, 106, 147–165. [Google Scholar] [CrossRef]
- Luo, M. Influenza virus entry. Adv Exp Med Biol 2012, 726, 201–221. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, S.; Blijleven, J.S.; Roos, W.H.; Onck, P.R.; van der Giessen, E.; van Oijen, A.M. Hemagglutinin-Mediated Membrane Fusion: A Biophysical Perspective. Annu Rev Biophys 2018, 47, 153–173. [Google Scholar] [CrossRef]
- Dadonaite, B.; Gilbertson, B.; Knight, M.L.; Trifkovic, S.; Rockman, S.; Laederach, A.; Brown, L.E.; Fodor, E.; Bauer, D.L.V. The structure of the influenza A virus genome. Nat Microbiol 2019, 4, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Pleschka, S. Overview of influenza viruses. Curr Top Microbiol Immunol 2013, 370, 1–20. [Google Scholar] [CrossRef] [PubMed]
- PAHO. La muerte púrpura: la gran gripe de 1918. Available online: https://www.paho.org/es/quienes-somos/historia-ops/muerte-purpura-gran-gripe-1918 (accessed on 11 March 2025).
- WHO. Number of COVID-19 deaths reported to WHO (cumulative total). Available online: https://data.who.int/dashboards/covid19/deaths (accessed on 11 March 2025).
- Hermann, E.; Krammer, F. Clade 2.3.4.4b H5N1 neuraminidase has a long stalk, which is in contrast to most highly pathogenic H5N1 viruses circulating between 2002 and 2020. mBio 1128, e0398924. [Google Scholar] [CrossRef]
- Bennett-Laso, B.; Berazay, B.; Muñoz, G.; Ariyama, N.; Enciso, N.; Braun, C.; Krüger, L.; Barták, M.; González-Aravena, M.; Neira, V. Confirmation of highly pathogenic avian influenza H5N1 in skuas, Antarctica 2024. Front Vet Sci 2024, 11, 1423404. [Google Scholar] [CrossRef]
- Banyard, A.C.; Bennison, A.; Byrne, A.M.P.; Reid, S.M.; Lynton-Jenkins, J.G.; Mollett, B.; De Silva, D.; Peers-Dent, J.; Finlayson, K.; Hall, R.; et al. Detection and spread of high pathogenicity avian influenza virus H5N1 in the Antarctic Region. Nat Commun 2024, 15, 7433. [Google Scholar] [CrossRef]
- Lisovski, S.; Günther, A.; Dewar, M.; Ainley, D.; Aldunate, F.; Arce, R.; Ballard, G.; Bauer, S.; Belliure, J.; Banyard, A.C.; et al. Unexpected Delayed Incursion of Highly Pathogenic Avian Influenza H5N1 (Clade 2.3.4.4b) Into the Antarctic Region. Influenza Other Respir Viruses 2024, 18, e70010. [Google Scholar] [CrossRef]
- Fasanmi, O.G.; Odetokun, I.A.; Balogun, F.A.; Fasina, F.O. Public health concerns of highly pathogenic avian influenza H5N1 endemicity in Africa. Vet World 2017, 10, 1194–1204. [Google Scholar] [CrossRef]
- Tassoni, L.; Fusaro, A.; Milani, A.; Lemey, P.; Awuni, J.A.; Sedor, V.B.; Dogbey, O.; Commey, A.N.; Meseko, C.; Joannis, T.; et al. Genetically Different Highly Pathogenic Avian Influenza A(H5N1) Viruses in West Africa, 2015. Emerg Infect Dis 2016, 22, 2132–2136. [Google Scholar] [CrossRef]
- Abolnik, C.; Phiri, T.; Peyrot, B.; de Beer, R.; Snyman, A.; Roberts, D.; Ludynia, K.; Jordaan, F.; Maartens, M.; Ismail, Z.; et al. The Molecular Epidemiology of Clade 2.3.4.4B H5N1 High Pathogenicity Avian Influenza in Southern Africa, 2021-2022. Viruses 2023, 15. [Google Scholar] [CrossRef]
- Abolnik, C.; Roberts, L.C.; Strydom, C.; Snyman, A.; Roberts, D.G. Outbreaks of H5N1 High Pathogenicity Avian Influenza in South Africa in 2023 Were Caused by Two Distinct Sub-Genotypes of Clade 2.3.4.4b Viruses. Viruses 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Leguia, M.; Garcia-Glaessner, A.; Muñoz-Saavedra, B.; Juarez, D.; Barrera, P.; Calvo-Mac, C.; Jara, J.; Silva, W.; Ploog, K.; Amaro, L.; et al. Highly pathogenic avian influenza A (H5N1) in marine mammals and seabirds in Peru. Nat Commun 2023, 14, 5489. [Google Scholar] [CrossRef]
- Bruno, A.; Alfaro-Núñez, A.; de Mora, D.; Armas, R.; Olmedo, M.; Garcés, J.; Vaca, M.S.; De la Torre, E.; Jarrin, D.; Burbano, L.; et al. Phylogenetic analysis reveals that the H5N1 avian influenza A outbreak in poultry in Ecuador in November 2022 is associated with the highly pathogenic clade 2.3.4.4b. Int J Infect Dis 2023, 133, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Paternina, D.; Herazo, R.; Oviedo, M.; Mattar, S. Dramatic re-emergence of avian influenza in Colombia and Latin America. Travel Med Infect Dis 2024, 59, 102711. [Google Scholar] [CrossRef]
- Pulit-Penaloza, J.A.; Brock, N.; Belser, J.A.; Sun, X.; Pappas, C.; Kieran, T.J.; Basu Thakur, P.; Zeng, H.; Cui, D.; Frederick, J.; et al. Highly pathogenic avian influenza A(H5N1) virus of clade 2.3.4.4b isolated from a human case in Chile causes fatal disease and transmits between co-housed ferrets. Emerg Microbes Infect 2024, 13, 2332667. [Google Scholar] [CrossRef]
- Kandeil, A.; Patton, C.; Jones, J.C.; Jeevan, T.; Harrington, W.N.; Trifkovic, S.; Seiler, J.P.; Fabrizio, T.; Woodard, K.; Turner, J.C.; et al. Rapid evolution of A(H5N1) influenza viruses after intercontinental spread to North America. Nat Commun 2023, 14, 3082. [Google Scholar] [CrossRef]
- Ly, H. Highly pathogenic avian influenza H5N1 virus infections of dairy cattle and livestock handlers in the United States of America. Virulence 2024, 15, 2343931. [Google Scholar] [CrossRef] [PubMed]
- Tawidian, P.; Torchetti, M.K.; Killian, M.L.; Lantz, K.; Dilione, K.E.; Ringenberg, J.M.; Bevins, S.N.; Lenoch, J.B.; Ip, H.S. Genotypic Clustering of H5N1 Avian Influenza Viruses in North America Evaluated by Ordination Analysis. Viruses 2024, 16. [Google Scholar] [CrossRef]
- Pabbaraju, K.; Tellier, R.; Wong, S.; Li, Y.; Bastien, N.; Tang, J.W.; Drews, S.J.; Jang, Y.; Davis, C.T.; Fonseca, K.; et al. Full-genome analysis of avian influenza A(H5N1) virus from a human, North America, 2013. Emerg Infect Dis 2014, 20, 887–891. [Google Scholar] [CrossRef]
- Verhagen, J.H.; Fouchier, R.A.M.; Lewis, N. Highly Pathogenic Avian Influenza Viruses at the Wild-Domestic Bird Interface in Europe: Future Directions for Research and Surveillance. Viruses 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Engelsma, M.; Heutink, R.; Harders, F.; Germeraad, E.A.; Beerens, N. Multiple Introductions of Reassorted Highly Pathogenic Avian Influenza H5Nx Viruses Clade 2.3.4.4b Causing Outbreaks in Wild Birds and Poultry in The Netherlands, 2020-2021. Microbiol Spectr 2022, 10, e0249921. [Google Scholar] [CrossRef]
- Muzyka, D.; Rula, O.; Tkachenko, S.; Muzyka, N.; Köthe, S.; Pishchanskyi, O.; Stegniy, B.; Pantin-Jackwood, M.; Beer, M. Highly Pathogenic and Low Pathogenic Avian Influenza H5 Subtype Viruses in Wild Birds in Ukraine. Avian Dis 2019, 63, 219–229. [Google Scholar] [CrossRef]
- King, J.; Harder, T.; Conraths, F.J.; Beer, M.; Pohlmann, A. The genetics of highly pathogenic avian influenza viruses of subtype H5 in Germany, 2006-2020. Transbound Emerg Dis 2021, 68, 1136–1150. [Google Scholar] [CrossRef]
- Bonilla-Aldana, D.K.; Calle-Hernández, D.M.; Ulloque-Badaracco, J.R.; Alarcón-Braga, E.A.; Hernández-Bustamante, E.A.; Cabrera-Guzmán, J.C.; Quispe-Vasquez, S.M.; Huayta-Cortez, M.A.; Benites-Zapata, V.A.; Rodriguez-Morales, A.J. Highly pathogenic avian influenza A(H5N1) in animals: A systematic review and meta-analysis. New Microbes New Infect 2024, 60-61, 101439. [Google Scholar] [CrossRef]
- Ariyama, N.; Pardo-Roa, C.; Muñoz, G.; Aguayo, C.; Ávila, C.; Mathieu, C.; Almonacid, L.I.; Medina, R.A.; Brito, B.; Johow, M.; et al. Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus in Wild Birds, Chile. Emerg Infect Dis 2023, 29, 1842–1845. [Google Scholar] [CrossRef]
- Jimenez-Bluhm, P.; Siegers, J.Y.; Tan, S.; Sharp, B.; Freiden, P.; Johow, M.; Orozco, K.; Ruiz, S.; Baumberger, C.; Galdames, P.; et al. Detection and phylogenetic analysis of highly pathogenic A/H5N1 avian influenza clade 2.3.4.4b virus in Chile, 2022. Emerg Microbes Infect 2023, 12, 2220569. [Google Scholar] [CrossRef] [PubMed]
- Kutkat, O.; Gomaa, M.; Moatasim, Y.; El Taweel, A.; Kamel, M.N.; El Sayes, M.; GabAllah, M.; Kandeil, A.; McKenzie, P.P.; Webby, R.J.; et al. Highly pathogenic avian influenza virus H5N1 clade 2.3.4.4b in wild rats in Egypt during 2023. Emerg Microbes Infect 2024, 13, 2396874. [Google Scholar] [CrossRef]
- Parums, D.V. Editorial: Concerns as Highly Pathogenic Avian Influenza (HPAI) Virus of the H5N1 Subtype is Identified in Dairy Cows and Other Mammals. Med Sci Monit 2024, 30, e945315. [Google Scholar] [CrossRef]
- Szaluś-Jordanow, O.; Golke, A.; Dzieciątkowski, T.; Chrobak-Chmiel, D.; Rzewuska, M.; Czopowicz, M.; Sapierzyński, R.; Kardas, M.; Biernacka, K.; Mickiewicz, M.; et al. A Fatal A/H5N1 Avian Influenza Virus Infection in a Cat in Poland. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Briand, F.X.; Souchaud, F.; Pierre, I.; Beven, V.; Hirchaud, E.; Hérault, F.; Planel, R.; Rigaudeau, A.; Bernard-Stoecklin, S.; Van der Werf, S.; et al. Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus in Domestic Cat, France, 2022. Emerg Infect Dis 2023, 29, 1696–1698. [Google Scholar] [CrossRef] [PubMed]
- Chothe, S.K.; Srinivas, S.; Misra, S.; Nallipogu, N.C.; Gilbride, E.; LaBella, L.; Mukherjee, S.; Gauthier, C.H.; Pecoraro, H.L.; Webb, B.T.; et al. Marked neurotropism and potential adaptation of H5N1 clade 2.3.4.4.b virus in naturally infected domestic cats. Emerg Microbes Infect 2025, 14, 2440498. [Google Scholar] [CrossRef] [PubMed]
- Naraharisetti, R.; Weinberg, M.; Stoddard, B.; Stobierski, M.G.; Dodd, K.; Wineland, N.; Beal, M.; Morse, J.; Hatter, S.; Sledge, D.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Infection of Indoor Domestic Cats Within Dairy Industry Worker Households - Michigan, May 2024. MMWR. Morbidity and mortality weekly report 2025, 74, 61–65. [Google Scholar] [CrossRef]
- Brown, J.D.; Black, A.; Haman, K.H.; Diel, D.G.; Ramirez, V.E.; Ziejka, R.S.; Fenelon, H.T.; Rabinowitz, P.M.; Stevens, L.; Poulson, R.; et al. Antibodies to Influenza A(H5N1) Virus in Hunting Dogs Retrieving Wild Fowl, Washington, USA. Emerg Infect Dis 2024, 30, 1271–1274. [Google Scholar] [CrossRef]
- Maas, R.; Tacken, M.; Ruuls, L.; Koch, G.; van Rooij, E.; Stockhofe-Zurwieden, N. Avian influenza (H5N1) susceptibility and receptors in dogs. Emerg Infect Dis 2007, 13, 1219–1221. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhong, G.; Wang, G.; Deng, G.; Li, Y.; Shi, J.; Zhang, Z.; Guan, Y.; Jiang, Y.; Bu, Z.; et al. Dogs are highly susceptible to H5N1 avian influenza virus. Virology 2010, 405, 15–19. [Google Scholar] [CrossRef]
- Kwon, T.; Trujillo, J.D.; Carossino, M.; Lyoo, E.L.; McDowell, C.D.; Cool, K.; Matias-Ferreyra, F.S.; Jeevan, T.; Morozov, I.; Gaudreault, N.N.; et al. Pigs are highly susceptible to but do not transmit mink-derived highly pathogenic avian influenza virus H5N1 clade 2.3.4.4b. Emerg Microbes Infect 2024, 13, 2353292. [Google Scholar] [CrossRef]
- Imai, M.; Herfst, S.; Sorrell, E.M.; Schrauwen, E.J.; Linster, M.; De Graaf, M.; Fouchier, R.A.; Kawaoka, Y. Transmission of influenza A/H5N1 viruses in mammals. Virus Res 2013, 178, 15–20. [Google Scholar] [CrossRef]
- Yu, J.; Li, T.; Wen, Z.; Wu, S.; Wang, Z.; Zheng, J.; Chen, M.; Chen, F.; Wei, W.K.; Zhai, S.L.; et al. Identification of D/Yama2019 Lineage-Like Influenza D Virus in Chinese Cattle. Front Vet Sci 2022, 9, 939456. [Google Scholar] [CrossRef]
- Sreenivasan, C.; Thomas, M.; Sheng, Z.; Hause, B.M.; Collin, E.A.; Knudsen, D.E.; Pillatzki, A.; Nelson, E.; Wang, D.; Kaushik, R.S.; et al. Replication and Transmission of the Novel Bovine Influenza D Virus in a Guinea Pig Model. J Virol 2015, 89, 11990–12001. [Google Scholar] [CrossRef]
- Ferguson, L.; Eckard, L.; Epperson, W.B.; Long, L.P.; Smith, D.; Huston, C.; Genova, S.; Webby, R.; Wan, X.F. Influenza D virus infection in Mississippi beef cattle. Virology 2015, 486, 28–34. [Google Scholar] [CrossRef]
- Flynn, O.; Gallagher, C.; Mooney, J.; Irvine, C.; Ducatez, M.; Hause, B.; McGrath, G.; Ryan, E. Influenza D Virus in Cattle, Ireland. Emerg Infect Dis 2018, 24, 389–391. [Google Scholar] [CrossRef]
- Collin, E.A.; Sheng, Z.; Lang, Y.; Ma, W.; Hause, B.M.; Li, F. Cocirculation of two distinct genetic and antigenic lineages of proposed influenza D virus in cattle. J Virol 2015, 89, 1036–1042. [Google Scholar] [CrossRef]
- Silveira, S.; Falkenberg, S.M.; Kaplan, B.S.; Crossley, B.; Ridpath, J.F.; Bauermann, F.B.; Fossler, C.P.; Dargatz, D.A.; Dassanayake, R.P.; Vincent, A.L.; et al. Serosurvey for Influenza D Virus Exposure in Cattle, United States, 2014-2015. Emerg Infect Dis 2019, 25, 2074–2080. [Google Scholar] [CrossRef] [PubMed]
- Quast, M.; Sreenivasan, C.; Sexton, G.; Nedland, H.; Singrey, A.; Fawcett, L.; Miller, G.; Lauer, D.; Voss, S.; Pollock, S.; et al. Serological evidence for the presence of influenza D virus in small ruminants. Vet Microbiol 2015, 180, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Ducatez, M.F.; Pelletier, C.; Meyer, G. Influenza D virus in cattle, France, 2011-2014. Emerg Infect Dis 2015, 21, 368–371. [Google Scholar] [CrossRef] [PubMed]
- Snoeck, C.J.; Oliva, J.; Pauly, M.; Losch, S.; Wildschutz, F.; Muller, C.P.; Hübschen, J.M.; Ducatez, M.F. Influenza D Virus Circulation in Cattle and Swine, Luxembourg, 2012-2016. Emerg Infect Dis 2018, 24, 1388–1389. [Google Scholar] [CrossRef]
- da Silva, M.S.; Mosena, A.C.S.; Baumbach, L.; Demoliner, M.; Gularte, J.S.; Pavarini, S.P.; Driemeier, D.; Weber, M.N.; Spilki, F.R.; Canal, C.W. Cattle influenza D virus in Brazil is divergent from established lineages. Arch Virol 2022, 167, 1181–1184. [Google Scholar] [CrossRef]
- CDC. Situación actual: influenza aviar en vacas lecheras. Available online: https://espanol.cdc.gov/bird-flu/situation-summary/mammals.html#:~:text=El%201%20de%20abril%2C%20los,(H5N1)%20de%20la%20HPAI (accessed on 13 March 2025).
- Goujgoulova, G.; Koev, K. Risk Assessment of Spread of the Influenza A Virus in Cows in South Bulgaria. Viruses 2025, 17. [Google Scholar] [CrossRef]
- Rubin, E.J.; Baden, L.R.; Goldstein, R.; Shuford, J.A.; Morrissey, S. NEJM Outbreaks Update - H5N1: A View from the States. N Engl J Med 2025. [Google Scholar] [CrossRef]
- Hawman, D.W.; Tipih, T.; Hodge, E.; Stone, E.T.; Warner, N.; McCarthy, N.; Granger, B.; Meade-White, K.; Leventhal, S.; Hatzakis, K.; et al. Clade 2.3.4.4b but not historical clade 1 HA replicating RNA vaccine protects against bovine H5N1 challenge in mice. Nat Commun 2025, 16, 655. [Google Scholar] [CrossRef] [PubMed]
- Alexakis, L.; Buczkowski, H.; Ducatez, M.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Ståhl, K.; Staubach, C.; Svartström, O.; Terregino, C.; et al. Avian influenza overview September-December 2024. Efsa j 2025, 23, e9204. [Google Scholar] [CrossRef]
- Mostafa, A.; Barre, R.S.; Allué-Guardia, A.; Escobedo, R.A.; Shivanna, V.; Rothan, H.; Castro, E.M.; Ma, Y.; Cupic, A.; Jackson, N.; et al. Replication kinetics, pathogenicity and virus-induced cellular responses of cattle-origin influenza A(H5N1) isolates from Texas, United States. Emerg Microbes Infect 2025, 14, 2447614. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.-Q.; Hutter, C.R.; Markin, A.; Thomas, M.; Lantz, K.; Killian, M.L.; Janzen, G.M.; Vijendran, S.; Wagle, S.; Inderski, B.; et al. Emergence and interstate spread of highly pathogenic avian influenza A(H5N1) in dairy cattle in the United States. Science 2025, 388, eadq0900. [Google Scholar] [CrossRef]
- Song, H.; Hao, T.; Han, P.; Wang, H.; Zhang, X.; Li, X.; Wang, Y.; Chen, J.; Li, Y.; Jin, X.; et al. Receptor binding, structure, and tissue tropism of cattle-infecting H5N1 avian influenza virus hemagglutinin. Cell 2025, 188, 919–929.e919. [Google Scholar] [CrossRef]
- Imai, M.; Ueki, H.; Ito, M.; Iwatsuki-Horimoto, K.; Kiso, M.; Biswas, A.; Trifkovic, S.; Cook, N.; Halfmann, P.J.; Neumann, G.; et al. Highly pathogenic avian H5N1 influenza A virus replication in ex vivo cultures of bovine mammary gland and teat tissues. Emerg Microbes Infect 2025, 14, 2450029. [Google Scholar] [CrossRef] [PubMed]
- Crossley, B.M.; Miramontes, C.C.; Rejmanek, D.; Gallardo, R.; Pereira, R. In-laboratory inactivation of H5N1 in raw whole milk through milk acidification: Results from a pilot study. J Dairy Sci 2025, 108, 2264–2275. [Google Scholar] [CrossRef]
- Kaiser, F.; Cardenas, S.; Yinda, K.C.; Mukesh, R.K.; Ochwoto, M.; Gallogly, S.; Wickenhagen, A.; Bibby, K.; de Wit, E.; Morris, D.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Stability in Irradiated Raw Milk and Wastewater and on Surfaces, United States. Emerg Infect Dis 2025, 31. [Google Scholar] [CrossRef]
- Le Sage, V.; Campbell, A.J.; Reed, D.S.; Duprex, W.P.; Lakdawala, S.S. Persistence of Influenza H5N1 and H1N1 Viruses in Unpasteurized Milk on Milking Unit Surfaces. Emerg Infect Dis 2024, 30, 1721–1723. [Google Scholar] [CrossRef]
- Schafers, J.; Warren, C.J.; Yang, J.; Zhang, J.; Cole, S.J.; Cooper, J.; Drewek, K.; Kolli, B.R.; McGinn, N.; Qureshi, M.; et al. Pasteurisation temperatures effectively inactivate influenza A viruses in milk. Nat Commun 2025, 16, 1173. [Google Scholar] [CrossRef]
- Falender, R.; Radniecki, T.S.; Kelly, C.; Cieslak, P.; Mickle, D.; Hall, H.; Scholz, R.; Sutton, M. Avian Influenza A(H5) Subtype in Wastewater - Oregon, September 15, 2021-July 11, 2024. MMWR Morb Mortal Wkly Rep 2025, 74, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.K.; Kumar, M.; Murugkar, H.V.; Nagarajan, S.; Tosh, C.; Namdeo, P.; Singh, R.; Mishra, S.; Senthilkumar, D.; Singh, V.P.; et al. Highly pathogenic avian influenza (H5N1) infection in crows through ingestion of infected crow carcasses. Microb Pathog 2023, 183, 106330. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.S.; Franson, J.C.; Gill, R.E.; Meteyer, C.U.; TeSlaa, J.L.; Nashold, S.; Dusek, R.J.; Ip, H.S. Experimental challenge and pathology of highly pathogenic avian influenza virus H5N1 in dunlin (Calidris alpina), an intercontinental migrant shorebird species. Influenza Other Respir Viruses 2011, 5, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Takadate, Y.; Tsunekuni, R.; Kumagai, A.; Mine, J.; Kikutani, Y.; Sakuma, S.; Miyazawa, K.; Uchida, Y. Different Infectivity and Transmissibility of H5N8 and H5N1 High Pathogenicity Avian Influenza Viruses Isolated from Chickens in Japan in the 2021/2022 Season. Viruses 2023, 15. [Google Scholar] [CrossRef]
- Vahlenkamp, T.W.; Teifke, J.P.; Harder, T.C.; Beer, M.; Mettenleiter, T.C. Systemic influenza virus H5N1 infection in cats after gastrointestinal exposure. Influenza Other Respir Viruses 2010, 4, 379–386. [Google Scholar] [CrossRef]
- Xu, C.; Dong, L.; Xin, L.; Lan, Y.; Chen, Y.; Yang, L.; Shu, Y. Human avian influenza A (H5N1) virus infection in China. Sci China C Life Sci 2009, 52, 407–411. [Google Scholar] [CrossRef]
- Galli, M.; Giacomelli, A.; Lai, A.; Zehender, G. H5N1 influenza A virus: lessons from past outbreaks and emerging threats. Infez Med 2025, 33, 76–89. [Google Scholar] [CrossRef]
- Uyeki, T.M.; Milton, S.; Abdul Hamid, C.; Reinoso Webb, C.; Presley, S.M.; Shetty, V.; Rollo, S.N.; Martinez, D.L.; Rai, S.; Gonzales, E.R.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Infection in a Dairy Farm Worker. N Engl J Med 2024, 390, 2028–2029. [Google Scholar] [CrossRef]
- Le Sage, V.; Werner, B.D.; Merrbach, G.A.; Petnuch, S.E.; O’Connell, A.K.; Simmons, H.C.; McCarthy, K.R.; Reed, D.S.; Moncla, L.H.; Bhavsar, D.; et al. Influenza A(H5N1) Immune Response among Ferrets with Influenza A(H1N1)pdm09 Immunity. Emerg Infect Dis 2025, 31, 477–487. [Google Scholar] [CrossRef]
- Leonard, J.; Harker, E.J.; Szablewski, C.M.; Margrey, S.F.; II, K.F.G.; Crossley, K.; Fletcher, E.; McCreavy, C.J.; Weis-Torres, S.; Wang, D.; et al. Seroprevalence of Highly Pathogenic Avian Influenza A(H5) Virus Infections Among Bovine Veterinary Practitioners — United States, September 2024. MMWR Morb Mortal Wkly Rep 2025 2025, 74, 50–52. [Google Scholar] [CrossRef]
- Mahase, E. Bird flu: US reports first human death in person infected with H5N1. Bmj 2025, 388, r28. [Google Scholar] [CrossRef] [PubMed]
- Morse, J.; Coyle, J.; Mikesell, L.; Stoddard, B.; Eckel, S.; Weinberg, M.; Kuo, J.; Riner, D.; Margulieux, K.; Stricklen, J.; et al. Influenza A(H5N1) Virus Infection in Two Dairy Farm Workers in Michigan. N Engl J Med 2024, 391, 963–964. [Google Scholar] [CrossRef] [PubMed]
- Pulit-Penaloza, J.A.; Belser, J.A.; Brock, N.; Kieran, T.J.; Sun, X.; Pappas, C.; Zeng, H.; Carney, P.; Chang, J.; Bradley-Ferrell, B.; et al. Transmission of a human isolate of clade 2.3.4.4b A(H5N1) virus in ferrets. Nature 2024, 636, 705–710. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
H5N1 Bird Flu in the United States, according to GISAID, June 11, 2025 (https://gisaid.org/resources/gisaid-in-the-news/highly-pathogenic-avian-influenza-outbreak-in-the-united-states/#c5130). Subsampled phylogenetic trees with focus on recent U.S. H5N1 samples shown for HA, NA, and PB2 (as of 11 June 2025). Clade 2.3.4.4b of the highly pathogenic avian influenza (HPAI) virus, which has been responsible for outbreaks among wild and domestic birds globally, continues to spread among dairy cattle, poultry, and other animal species in the United States. Since April 2024, the U.S. CDC has confirmed 70 human infections based on genomic sequencing. In three of these cases, the NA-S247N amino acid substitution was detected, which may slightly reduce the virus’s sensitivity to the neuraminidase inhibitor oseltamivir in laboratory settings. Additionally, the CDC reported a separate mutation in the polymerase acidic (PA) protein in a virus isolated from a recently confirmed human H5N1 case in California.
Figure 1.
H5N1 Bird Flu in the United States, according to GISAID, June 11, 2025 (https://gisaid.org/resources/gisaid-in-the-news/highly-pathogenic-avian-influenza-outbreak-in-the-united-states/#c5130). Subsampled phylogenetic trees with focus on recent U.S. H5N1 samples shown for HA, NA, and PB2 (as of 11 June 2025). Clade 2.3.4.4b of the highly pathogenic avian influenza (HPAI) virus, which has been responsible for outbreaks among wild and domestic birds globally, continues to spread among dairy cattle, poultry, and other animal species in the United States. Since April 2024, the U.S. CDC has confirmed 70 human infections based on genomic sequencing. In three of these cases, the NA-S247N amino acid substitution was detected, which may slightly reduce the virus’s sensitivity to the neuraminidase inhibitor oseltamivir in laboratory settings. Additionally, the CDC reported a separate mutation in the polymerase acidic (PA) protein in a virus isolated from a recently confirmed human H5N1 case in California.
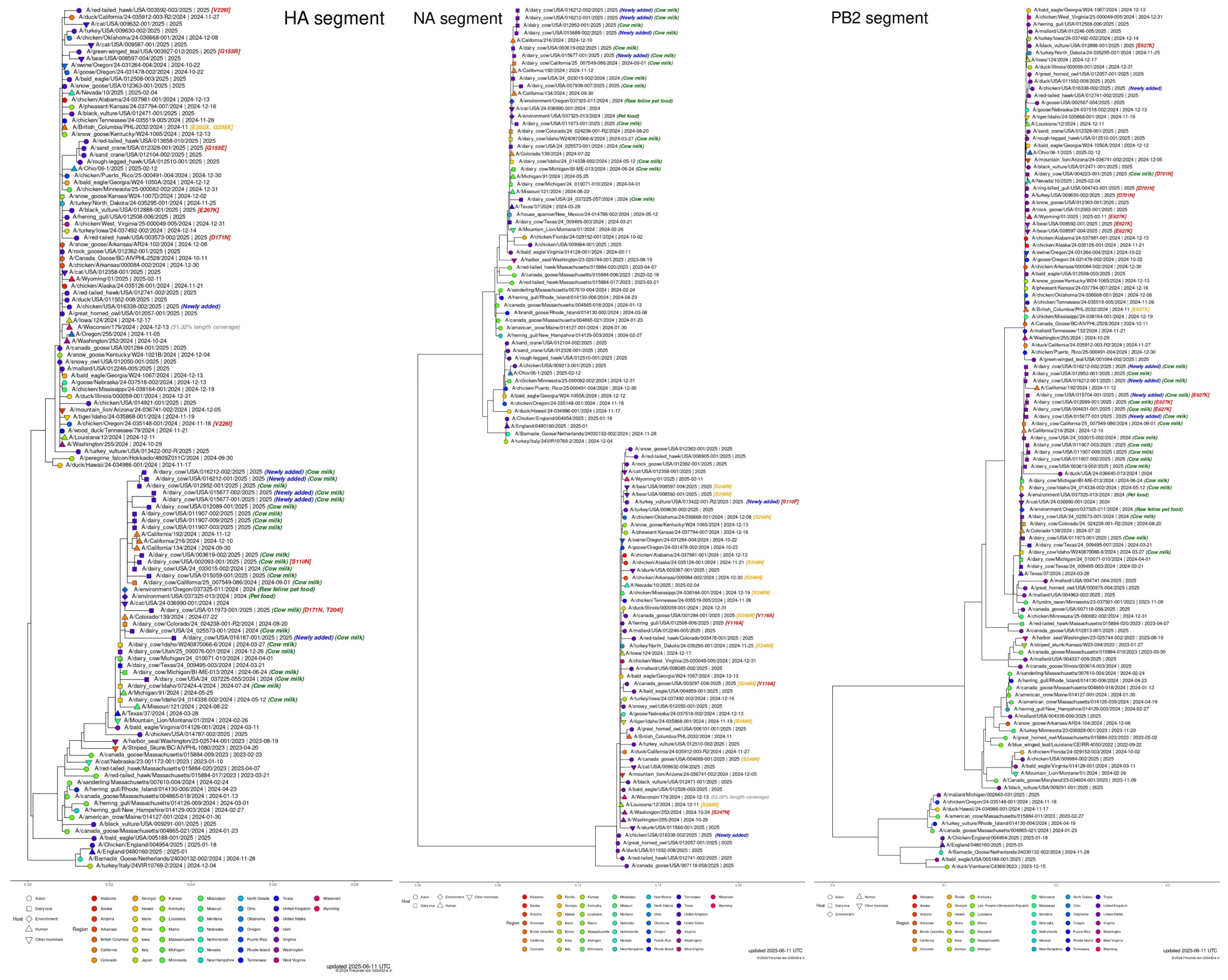
Figure 2.
Number of Confirmed Cases of H5N1 in Cattle by State, Total Outbreak, United States of America, March 2024-June 2025, according to the animal and plant health inspection service (APHIS) of the U.S. Department of agriculture (USDA). Reproduced from https://www.aphis.usda.gov/livestock-poultry-disease/avian/avian-influenza/hpai-detections/hpai-confirmed-cases-livestock.
Figure 2.
Number of Confirmed Cases of H5N1 in Cattle by State, Total Outbreak, United States of America, March 2024-June 2025, according to the animal and plant health inspection service (APHIS) of the U.S. Department of agriculture (USDA). Reproduced from https://www.aphis.usda.gov/livestock-poultry-disease/avian/avian-influenza/hpai-detections/hpai-confirmed-cases-livestock.
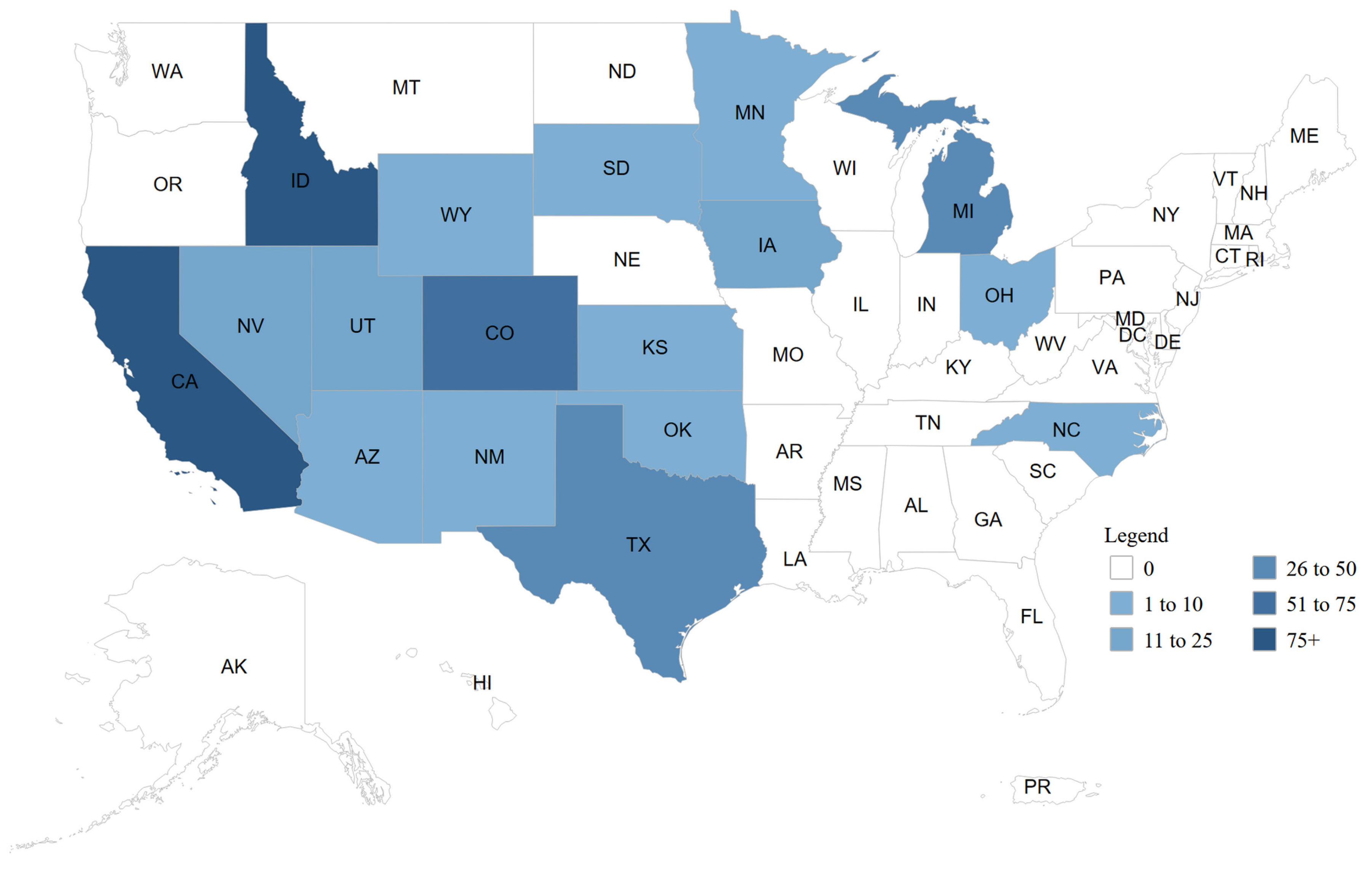
Figure 3.
Influenza H5 virus infections in humans exposed to affected animals by state (January 1, 2024-June 6, 2025), according to the Centers for Disease Control and Prevention (CDC). Reproduced from https://www.cdc.gov/bird-flu/h5-monitoring/index.html.
Figure 3.
Influenza H5 virus infections in humans exposed to affected animals by state (January 1, 2024-June 6, 2025), according to the Centers for Disease Control and Prevention (CDC). Reproduced from https://www.cdc.gov/bird-flu/h5-monitoring/index.html.
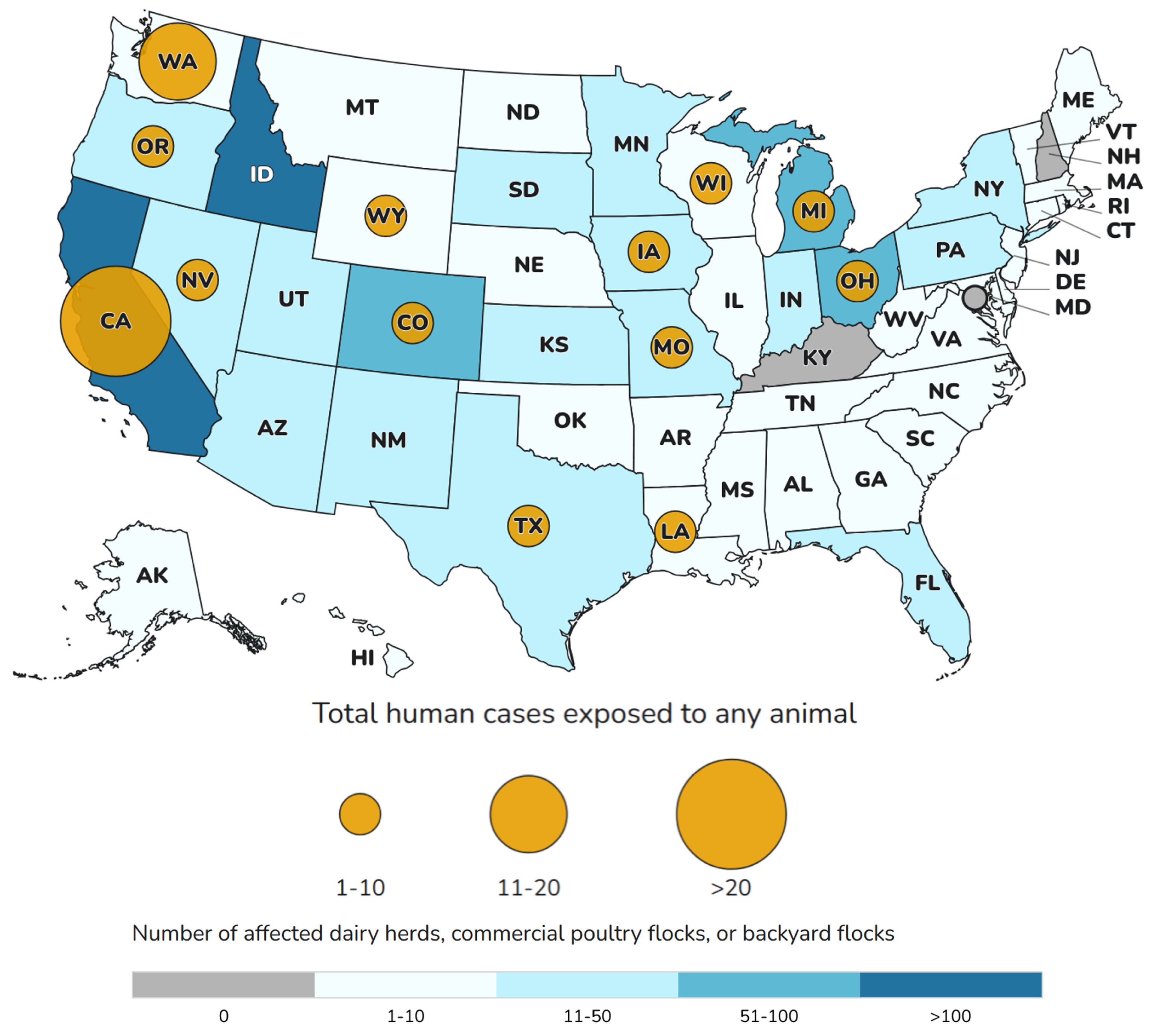
Figure 4.
Taxonomic classification of the influenza viruses, according to the International Committee of Viral Taxonomy (ICTV), https://ictv.global/taxonomy. Accessed June 12, 2025.
Figure 4.
Taxonomic classification of the influenza viruses, according to the International Committee of Viral Taxonomy (ICTV), https://ictv.global/taxonomy. Accessed June 12, 2025.

Figure 5.
Changes in the Hemagglutinin for the top emerging variants based on sequences collected in the past 100 days. Influenza Hemagglutinin mutation surveillance, EpiFlu Update, January 2025 (www.gisaid.org).
Figure 5.
Changes in the Hemagglutinin for the top emerging variants based on sequences collected in the past 100 days. Influenza Hemagglutinin mutation surveillance, EpiFlu Update, January 2025 (www.gisaid.org).
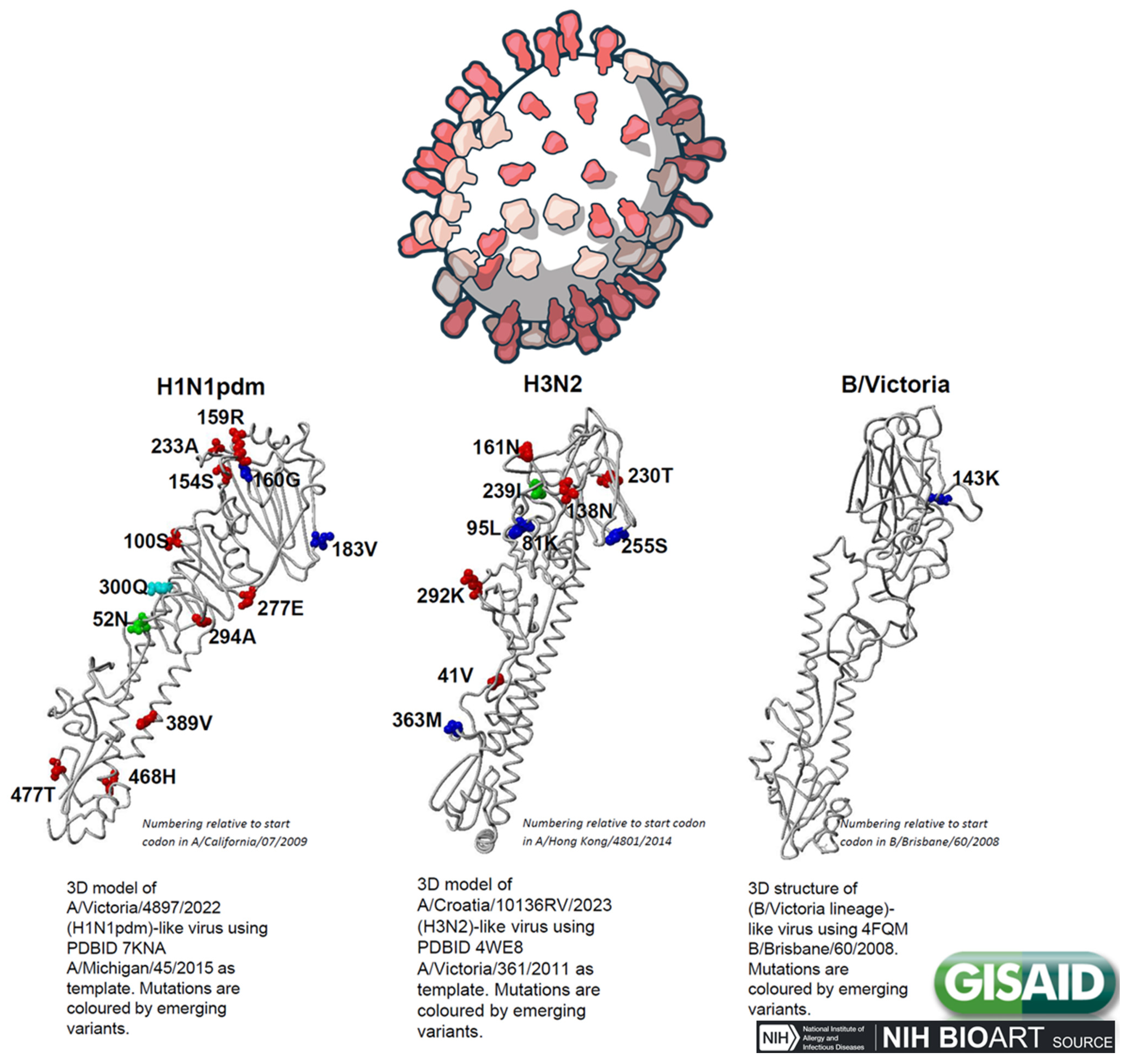
Figure 6.
Fresh raw milk vending machine in Ljubljana, Slovenia. Photo taken by AJRM on January 28, 2024. The text says: MLEKOmat – every day fresh raw milk (stored at 4°C up to three days) from the Miš farm. Logo: "Authentic fresh milk from Zavrh under Šmarna gora". The price of a liter is 1.30 euros. A note at the bottom left says: “Note: Use clean bottles only! Milk must be boiled before use!” (https://www.mleko-mat.si/mlekomat/).
Figure 6.
Fresh raw milk vending machine in Ljubljana, Slovenia. Photo taken by AJRM on January 28, 2024. The text says: MLEKOmat – every day fresh raw milk (stored at 4°C up to three days) from the Miš farm. Logo: "Authentic fresh milk from Zavrh under Šmarna gora". The price of a liter is 1.30 euros. A note at the bottom left says: “Note: Use clean bottles only! Milk must be boiled before use!” (https://www.mleko-mat.si/mlekomat/).


Table 1.
Number of confirmed human H5N1 cases and deaths worldwide during 2003-2024.
| Country | 2003-2009 | 2010-2014 | 2015-2019 | 2020 | 2021 | 2022 | 2023 | 2024 | Total | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | |
| Australia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Azerbaijan | 8 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 5 |
| Bangladesh | 1 | 0 | 6 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 1 |
| Cambodia | 9 | 7 | 47 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 4 | 10 | 2 | 72 | 43 |
| Canada | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 1 |
| Chile | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| China | 38 | 25 | 9 | 5 | 6 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 56 | 32 |
| Djibouti | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Ecuador | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Egypt | 90 | 27 | 120 | 50 | 149 | 43 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 359 | 120 |
| India | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Indonesia | 162 | 134 | 35 | 31 | 3 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 200 | 168 |
| Iraq | 3 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 2 |
| Lao | 2 | 2 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 2 |
| Myanmar | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Nepal | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Nigeria | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Pakistan | 3 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1 |
| Spain | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 2 | 0 |
| Thailand | 25 | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 17 |
| Turkey | 12 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 4 |
| UK | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 5 | 0 |
| USA | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 58 | 0 | 59 | 0 |
| Vietnam | 112 | 57 | 15 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 129 | 65 |
| Total | 468 | 282 | 233 | 125 | 160 | 48 | 1 | 0 | 2 | 1 | 6 | 1 | 12 | 4 | 72 | 3 | 954 | 464 |
Table 2.
Challenges and Strategies in Surveillance, Diagnostics, Control and Prevention of H5N1 in Cattle.
Table 2.
Challenges and Strategies in Surveillance, Diagnostics, Control and Prevention of H5N1 in Cattle.
| Domain | Challenges | Strategies |
|---|---|---|
| Surveillance |
|
|
| Diagnostics |
|
|
| Control |
|
|
| Prevention |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



