Submitted:
05 June 2025
Posted:
06 June 2025
You are already at the latest version
Abstract
Prolactin (PRL) is a hormone primarily associated with lactation, but it plays various roles in both men and women. PRL belongs to the family of peptide hormones, in-cluding placental lactogen and growth hormone. Interestingly, PRL is a pleiotropic hor-mone affecting several physiological and pathological functions, including fertility. Moreover, several pathophysiological roles have been associated with such a hormone, including those of the immune system, autoimmune disorders, asthma, and ageing. Ad-ditionally, PRL receptors are ubiquitously expressed in tissues including the mammary gland, gonads, liver, kidney, adrenal gland, brain, heart, lungs, pituitary gland, uterus, skeletal muscle, skin, and blood cells, immune system. Therefore, in the present paper, we will resume the potential of the PRL may contribute to asthma by promoting inflamma-tion and modulating immune responses. The detection of its receptor in lung tissue sug-gests a direct role in airway smooth muscle contractility through activation of signaling pathways such as JAK2-STAT5, MAPK/ERK1/2, and PI3K/Akt, as well as influencing ion-ic currents that regulate cell contraction, proliferation, and survival. In this sense, this re-view aims to explore the potential involvement of PRL in asthma pathophysiology by examining its interactions with intracellular signaling pathways and its possible impact on airway smooth muscle contractility and immune modulation.
Keywords:
prolactin
; PRL
; asthma
; contraction
; hormonal regulation
1. Introduction
Asthma is a serious global health problem affecting approximately 300 million people of all age groups around the world, and causing about 1,000 deaths per day [1]. This ailment is a heterogeneous condition typically characterized by chronic airway inflammation, defined by the history of respiratory symptoms such as wheezing, shortness of breath, chest tightness, and cough that fluctuate over time and intensity, accompanied by variable limitation of expiratory air flow. According to the 2024 GINA [1], asthma can be classified in distinct clinical phenotypes, including allergic, nonallergic, late-onset, obesity-related, cough-predominant, and asthma with persistent airflow limitation. These phenotypes differ in onset, inflammatory profiles, and treatment responses, highlighting the need for personalized approaches for diagnosis and management.
Besides, it has been proposed that the hypophysis-pituitary-adrenal (HPA) axis worsens asthma due to a physiological stress response [2], and unfortunately, this response orchestrated by the HPA axis still holds many unidentified features regarding this ailment. Conceivably, the management of asthma might expand substantially by increasing the knowledge on the role played by HPA axis in asthma pathophysiology. In this sense, PRL, a hormone secreted by the pituitary gland, can modulate stress responses by inhibiting the HPA axis [3].
Growing evidence suggests that numerous hormonal factors play a role in lung development for optimal respiratory function, such as, by influencing respiratory mechanics and inflammation. Some relevant factors involved in airway and lung illnesses are, for example, ghrelin, leptin, and glucagon-like peptide-1 (GLP-1) [4]; retinoids and cholecalciferol [5,6]; sex steroids [7]; hormones such as insulin, prolactin [8] and glucagon [9], as well as growth factors like granulocyte/macrophage colony-stimulating factor (GM-CSF) [10].
Interestingly, sex hormones have been increasingly shown to play a substantial role in modulating smooth muscle contractility and affect asthma development. In this sense, during periods of changes in female sex hormones including puberty, menstruation, pregnancy, and menopause, these fluctuations have been associated with changes in asthma severity [11,12,13]. Pregnancy in asthmatic women becomes a particular concern, since during gestation, one-third of the pregnant women suffer worsening asthma symptoms, one-third improves, while the remaining one-third shows no change [13,14,15].
In this sense, prolactin (PRL), traditionally associated with lactation, is now recognized for its immunomodulatory effects [16,17]. It is currently recognized that PRL is a pleiotropic hormone that participates in more than 300 physiological functions, including reproduction, metabolism, immune response, and brain processes. PRL has been reported to cross the blood-brain barrier and exert its effects on various regions of the central nervous system. Furthermore, recent studies have demonstrated its involvement in immunological mechanisms, suggesting it plays a relevant role in inflammation [8,18]. On the other hand, PRL has been associated with many autoimmune diseases such as systemic lupus erythematosus (SLE), rheumatoid arthritis (RA) and multiple sclerosis (MS) [19,20,21]. Furthermore, it has been observed that, while aging, PRL levels fluctuate, and that both increases and decreases in these concentration have been linked to neurodegenerative diseases (including Huntington's, multiple sclerosis, Alzheimer's, and Parkinson's) [22,23].
However, in asthma, PRL might be involved in promoting inflammation and modulating immune cell activity [8,24]. Additionally, the presence of its PRL receptor (PRLR) has been observed in lung tissue in animal models [25], suggesting that this hormone could be directly involved in airway smooth muscle contraction, because PRLR stimulation activates the JAK2-STAT5 signaling pathway [26,27,28,29,30], which participates in processes such as contraction, proliferation, differentiation and cell survival. It has also been reported that PRL activates other signaling pathways such as MAPK/ERK1/2 and PI3K/Akt, expanding its impact on diverse cellular functions [29,31,32,33,34].
Interestingly, augmented plasma PRL concentration modulates ionic currents in multiple tissues from different species. For example, it modulates ATP-sensitive potassium channels to provoke an analgesic response in mice, significantly increasing the active transport of Ca²⁺ in duodenal enterocytes, an effect that is completely abolished by blocking L-type calcium channels with nifedipine or by inhibiting the main Ca²⁺ elimination systems in the basolateral membrane, e.g., plasma membrane Ca²⁺-ATPase (PMCA) and the Na⁺/Ca²⁺ exchanger (NCX) [35]. Meanwhile, in sensory neurons of female rats, PRL potentiates the activity of acid-sensitive ion channels in primary [36]. It has also been shown that these hormones induce small electrical currents in neurons through TRP-like calcium channels, and facilitate Ca²⁺ entry via voltage-gated L-type calcium channels. Furthermore, it has been suggested that its rapid responses in neuronal cells are mediated by the short isoform of the PRLR, activating intracellular signaling pathways such as PI3-kinase and PKC [33]. It stimulates sodium and chloride transport in renal epithelial cells (A6) by activating the epithelial sodium channel (ENaC) and a chloride channel (ClC4) type anion channel. This effect depends on the cAMP/PKA signaling pathway, since its inhibition blocks the response. PRL increases both the number and the probability of ENaC opening, thereby promoting amiloride-sensitive and -insensitive trans-epithelial currents [37]. Finally, PRL was observed to activate ENaC and ClC channels in A6 renal epithelial cells via the cAMP/PKA-dependent signaling pathway, too [37].
Despite the ample evidence on the role played by PRL in the lung and airways, its specific participation in asthma remains poorly understood. Conceivably, it might possess a potentially important role in the pathophysiology of asthma, which may be particularly relevant for women during pregnancy and lactation. The objective of this study is to analyze the possible role of prolactin in the modulation of airway smooth muscle excitation-contraction coupling.
2. The Hormone Prolactin
Currently, PRL interests the scientific community due to its multiple functions in the organism [30,38,39]. PRL was discovered in the 1930s by biologist Oscar Riddle; interestingly, at first, it was only recognized as a factor that controls milk production and secretion [38,40,41]. Nevertheless, it is now recognized as a pleiotropic hormone, with over 300 functions in numerous tissues [39,42,43].
PRL belongs to the family of peptide hormones, including placental lactogen and growth hormone, making up the family of somatolactogens, also known as class I helical cytokines (41, 44); these hormones are characterized by having a tertiary structure, composed of four antiparallel α helices [29,39]. Likewise, PRL is synthesized in the anterior lobe of the pituitary gland by specialized cells called lactotrophs [32,39,45,46]. However, it has also been reported that it is synthesized extrapituitarily in other tissues such as lymphocytes, skin fibroblasts, prostate cells, endothelial cells, adipose tissue cells, mammary gland, ovaries, decidua and in the brain [18,29,30,47,48].
Regarding its structural characteristics, it has been reported that PRL is encoded by a single gene (PRL), whose size is 10kb. This gene is composed of 5 exons and four introns in most species, including mammals, fish and birds [29,30,32,39,49,50]. Furthermore, it has been reported that the transcriptional regulation of pituitary and extrapituitary PRL expression is under the control of two independent promoter regions: the first is the proximal promoter region that modulates pituitary expression of PRL, and the second is a distal promoter region that promotes extrapituitary PRL expression [26,29,30,39].
However, at the molecular level, its expression and regulation are tissue-specific because several isoforms of PRL have been described. These result from alternative splicing, proteolytic cleavage, or post-translational modifications (such as phosphorylation, glycosylation, and deamidation) and the association with other circulating proteins. It is important to note that these modifications can modify the biological activity of PRL in the organism [29,32,39,51].
In addition, it has been informed that PRL is composed of 199 amino acid residues with a weight of 23 kDa. This form of the hormone is known as monomeric PRL, and it has three disulfide bonds present in similar places in rodents and humans [26,30,32,42]. Likewise, it has been reported that PRL can form dimers, polymers and aggregates [29,39,51,52].
Some examples of the modifications that PRL can have high molecular mass PRLs such as “big PRL” and “big PRL” (also known as macroPRL), which have been reported in the blood tissue of approximately 100 kDa [30,39,52,53]. Nevertheless, macroPRLs show lower biological activity and are suggested to participate in the storage, modification and release of PRL [29,39,54]. On the other hand, there are PRLs with low molecular weight, which can be 14 kDa, 16 kDa and 22 kDa; these hormones are generated from the proteolytic cleavage of the 23 kDa pituitary PRL [26,38,41,51,55]. An example of a molecular fragment derived from the proteolytic processing of PRL is vasoinhibine, generated by the action of proteases such as cathepsin D [51,55,56]. Finally, for it to exert its biological action, PRL requires interaction with its receptors, which are expressed in various tissues and organs (Figure 1a) [28,29,50,57].
3. Prolactin Receptors
As previously reported, the actions of PRL are initiated when this hormone binds to a homodimer of the PRLR, forming a heterotrimeric complex [28,29,30,31]. Therefore, it is essential to describe the characteristics of PRLRs, which are ubiquitously expressed in organisms [28,31,33,50], in tissues including the mammary gland, gonads, liver, kidney, adrenal gland, brain, heart, lungs, pituitary gland, uterus, skeletal muscle, skin, and blood cells immune system [28,39,41,58,59,60,61]. It has been reported that PRLRs are transmembrane proteins and belong to class I of the cytokine receptor superfamily. Interestingly, they lack intrinsic tyrosine kinase activity and can be phosphorylated by cytoplasmic proteins [28,30,33,50]. It should be noted that this family also includes receptors for GH (GHR), leptin (LEPR), leukaemia inhibiting factor (FIL) and erythropoietin (EPO), to mention a few [29,30,41].
PRLRs are composed of three domains: extracellular, transmembrane and intracellular [33,41,50], The extracellular domain includes two regions, called S1 and S2 (or D1 and D2), which together form the ligand binding site and are identical between species, as is the transmembrane domain, the only one that differs is the intracellular domain cytoplasmic, which can vary in length (Figure 1b) [29,30,33,41,50]. Modifications in the intracellular domain are key to recognizing the different isoforms that PRLR can display; this is because, like PRL, alternative mRNA splicing can occur in PRLR. Currently, three main isoforms for PRLR have been described in rodents and five in humans. However, the most studied, due to their distribution and expression, are the long isoform, the intermediate isoform and the short isoform in both species [28,33,41,54], To form homodimers or heterodimers, it is essential to note that the union of two PRLR monomers of the same isoform leads to the formation of homodimers or heterodimers when different isoforms join [41,50]. Interestingly, only in humans has a soluble isoform been described [27,30,54,62,63]. This soluble isoform of PRLR has been characterized in human breast cancer cell lines, being a regulatory mechanism for the bioavailability and signaling of extrapituitary PRL, which is why it is also known as "PRL binding proteins" in the extracellular domain [62,63,64].
On the other hand, PRLRs have been mainly related to the activation of the JAK2-STAT5 signaling pathway [26,27,28,29,30], this signaling pathway is activated in response to cytokines, growth factors, and PRL. Furthermore, it is implicated in multiple functions in secretory mammary epithelial cells, which include specifying, proliferating, differentiating, and surviving [26,27,50,61,64]. Also, it is essential to mention that the discovery of this signaling cascade was a significant advance in the understanding of the actions of PRL in the body [26,29,30,31,64]. Nevertheless, it has also been described that PRL can induce the activation of two other pathways, which are the MAPK/ERK1/2 pathway and the P13K/Akt pathway [29,31,32,33,34]. Activating all these pathways can influence the various described functions of PRL.
4. Prolactin Functions
The canonical function for which PRL is recognized is for its role in lactation; at this stage, the secretion of PRL is regulated by a delicate mechanism of endogenous factors, such as PRL-releasing factors (PRF) as well as thyrotropin-releasing hormone (THR) and by external factors such as photoperiod, food availability and physical processes associated with breast suction that stimulate its secretion. When there are high concentrations of PRL, PRL inhibitory factors (PIF) are released, which can be gamma-aminobutyric acid (GABA), dihydroxyphenylalanine (DOPA) and somatostatin [29,41,45,65,66]. However, currently, it is known that PRL regulates a large number of physiological processes, which is why it is known as a pleiotropic hormone with more than 300 functions described in the body, which are divided into categories such as reproduction, lactogenesis, brain processes, immune response, angiogenesis maternal behavior, growth, metabolism and osmoregulation [18,31,39,42,48,55,56,64,67].
Furthermore, it has been reported that PRL can cross the blood-brain barrier [38,45,48,68]. Therefore, the role that PRL plays in the brain is of interest for scientific research, where its expression by mRNA has been reported in several brain areas, such as the olfactory bulb, the corpus callosum, the choroid plexuses, the amygdala, the hypothalamus, thalamus, cerebral cortex and, hippocampus [26,28,29,31,32,38,50]. However, data on PRL expression in the brain remains limited and controversial. Since no conclusive studies demonstrate PRL secretion in the brain, it is proposed that all PRL effects arise from a pituitary source. However, a recent transcriptomic survey by Cabrera-Reyes et al., 2019 suggests effects on PRL in the hippocampus [69]. Likewise, the effects of PRL on the brain depend on factors such as age, sex, and the reproductive status of the species [27,29,30,31,33,34,61]. Finally, it is relevant to mention that PRL has been reported to participate in many brain functions, including maternal behavior, memory, energy balance, food intake, sleep, anxiety, neurogenesis, and neuroprotection [33,34,42,57,59,70,71,72,73,74,75], the latter being a field of interest for the scientific community.
5. Prolactin and Interactions
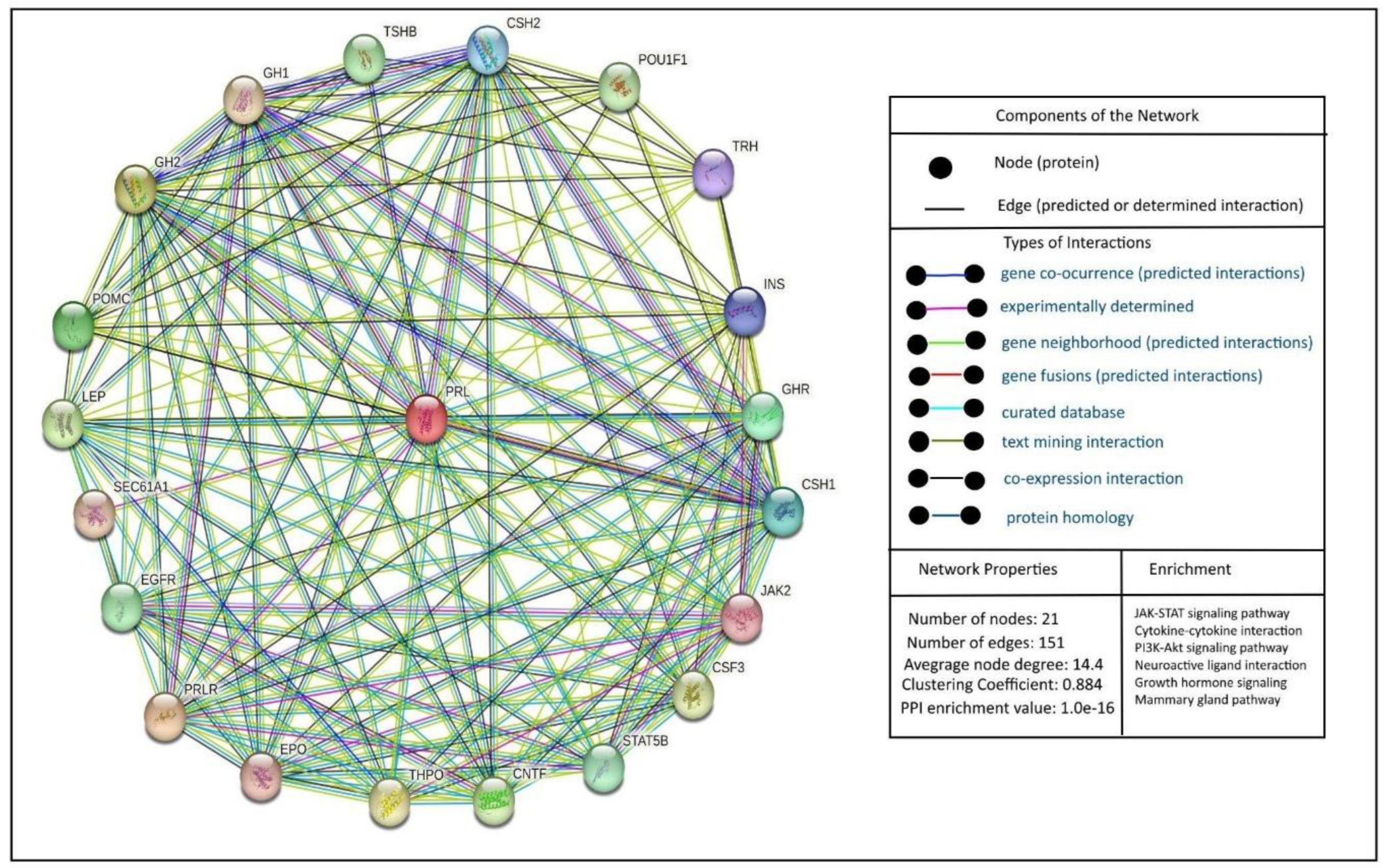
When discussing the protein-protein interactions of PRL, it is essential to focus on its interactions with its receptor and downstream signaling pathways. It is necessary to indicate that PRL's interactions are complex and involve multiple signaling pathways; among them, the JAK/STAT pathway is a central component of PRL signaling (Figure 2).
It is essential to mention that PRL is a pleiotropic hormone that can affect several physiological and pathological functions, including fertility. Moreover, PRLRs are widely expressed in several tissues, including brain regions and reproductive organs. Thus, PRLRs may exert prolactin's functions upon activation through several signaling pathways, as seen in Figure #.
6. Prolactin and the Immune System
Interestingly, the genes that encode PRL are located in the short arm of chromosome 6, in proximity to the HLA-DRB1 gene that is associated with autoimmune diseases, especially SLE [76,77]. Furthermore, prolactin has been shown to act as an immunomodulator. In splenic B cells from female Balb/c mice, prolactin treatment for 4 weeks led to a decrease in the BCR-mediated apoptosis of the T1 B cell subset, which was associated with an upregulation of the anti-apoptotic gene INF-γRII and downregulation of the pro-apoptotic Trp63 gene [78]. Additionally, the state or hyperprolactinemia also dysregulates receptor editing by causing the co-expression of more than one light chain in the cell’s surface; thus, the cell can escape clonal deletion and generating autoreactivity, as well as modifying the level of B cell anergy by lowering the BCR-mediated activation threshold [78]. In B cell hybridomas, PRL, in a dose-dependent manner, increases proliferation induced by IL-4, IL-5 and IL-6 and decreases the downregulation in proliferation induced by TGF-β [79,80].
In T cells, PRL treatment (2-200 ng/ml) incremented proliferation following IL-2 and phytohemagglutinin stimulation without modifying the subsets [81]. PRL also modulates dendritic cell (DC) differentiation and maturation. At physiological levels (10-20 ng/ml), PRL acts synergically with GM-CSF to inhibit DC maturation, comparable to the effect induced by IL-4, while supraphysiological concentrations (80 ng/ml) is stimulatory [77,82]. PRL modulates IFN-γ production through the JAK/Stat/IRF-1 pathway, in addition to the modulation of DC cells, thus leading to an inflammatory response that could be implicated in SLE patients [77,83,84,85,86]. Furthermore, in murine spleen CD1c-positive dendritic cells (SDCs), 24 hr PRL treatment incremented viability, stimulatory capacity, CD40 and MHC-11 expression, meanwhile decreasing the levels of CD54 and NF-κBp65 and endocytosis [87,88].
As for cytokine regulation, PRL is both subject to regulation and a mediator of cytokine production. While IL-1, IL-2 and IL-6 have been shown to stimulate PRL secretion, IFN-γ inhibits, and TNF-α has both stimulatory and inhibitory effects [80,89,90,91,92]. PRL has also been shown to enhance Th1 type cytokines in vivo and in vitro [80,82,93,94], increase the release of IL-12 [93], and IL-1 in mouse peritoneal macrophages [95], and increment IL-6 production via PRL-mediated IL-1 activity [96].
7. Prolactin and Autoimmune Diseases
Autoimmune diseases are influenced by hormonal regulation, with their higher prevalence in females suggesting a significant role for sex hormones in the underlying pathophysiology. Various diseases from this group have been associated with elevated circulating levels of PRL [76,77,97,98,99]. SLE activity has been positively associated with increased serum levels of PRL [77,97,100]. Treatment with bromocriptine, a dopamine receptor agonist that selectively inhibits prolactin secretion, has been demonstrated to effectively treat suppress disease activity in NZB/NZW (B/W) F1 mice, an SLE-like model, observing lower anti-DNA antibodies and circulating IgG levels, as well as an increase in longevity [101]. Similarly, in human SLE patients, bromocriptine treatment was associated with improved disease severity and lower anti-dsDNA. The patients presented flare-ups when the therapy was discontinued [77,102].
Hyperprolactinemia was found to be associated with antiphospholipid syndrome, especially with reproductive failure presented in the disease [98]. Furthermore, in a case report, hyperprolactinemia was found present in a patient with multiple autoimmune diseases (Jaccoud’s arthropathy, urticarial vasculitis, systemic lupus erythematosus and Sjögren’s syndrome) [76], correlating with the 20% of patients with SLE that present increased serum levels of prolactin [76,99].
8. Prolactin in Asthma
PRL, commonly recognized for its role in lactation, has gained attention as an immunomodulator and participant in the pathogenesis of immune diseases. Although little is known, emerging evidence suggests that PRL could be involved in asthma by modulating immune cell function and promoting a proinflammatory state. Interestingly, in one study, female rats were divided into three groups: virgins with no lung injury (n group), ovalbumin (OVA)-sensitized virgins (V group), and OVA-sensitized lactating females (L group). The L group had a lower bronchoalveolar lavage (BAL) leukocyte count and eosinophil and macrophage count compared with the V group; meanwhile, BAL interferon-γ were increased and corticosterone levels were lower. Interestingly, norepinephrine levels were higher in the L group compared to the N and V groups. These results indicate that lactation could protect the sensitized females from developing a pro-inflammatory response, and prolactin could contribute to this effect [103].
In another study, 86 children with mild asthma were either treated with sublingual immunotherapy (SLIT) or given a placebo for 6 months; at the end of the study, the treated group reported a significant improvement in asthma and rhinitis symptoms, lower serum levels of eosinophil cationic protein (ECP), IL-13, PRL and adrenocorticotropic hormone (ACTH). The reduction in the Th2 cytokine response and symptomology could be linked to an immunomodulatory effect of PRL suppressed by the SLIT treatment [24].
In women with perimenstrual asthma (PMA), serum prolactin levels tend to increase during both the luteal and follicular phases compared to asthmatic patients without PMA and healthy subjects [104]. Although a significant difference has not been observed, this does indicate a possible relation between prolactin and asthma symptomology that warrants further study. Recently, in OVA-induced asthmatic mice, the aqueous extract of Herba Houttuyniae reduced airway hyperresponsiveness (AHR) to methacholine challenge. Six metabolites were identified that could be attributed to the therapeutic effects. Of the targeted asthma-related genes that could be affected by the metabolite, the prolactin signaling pathway was identified as a prime targeted candidate, indicating that this pathway could be involved in the AHR modulation [105].
9. Prolactin and Aging
Aging is characterized by a subtle decline in all biological systems, including the endocrine ensembles, particularly at the central regulator of endocrine hypothalamic-pituitary units. In this sense, it has been reported that during aging, prolactin secretion decreases about 40% after menopause but declines less in older men. Since it has been reported that PRL-R is downregulated in the aged retina, it has been suggested that PRL signaling is impaired, leading to retinal function, suggesting that PRL is required for the homeostasis of aged retina [106]. On the other hand, PRL dysregulation has been associated with neurodegenerative diseases, including Huntington´s, multiple sclerosis [22], Alzheimer's (AD) and Parkinson's diseases (PD) [23]. In this section, we will focus only on AD and PD since both are the most common age-related neurodegenerative diseases. In the early stages of AD, it has been reported that PRL levels significantly increased [107], probably since individuals with AD show significant alterations in the tubero-infundibular pathway involved in the regulation of PRL secretion [108]; however, the levels decrease in the late phase of the disease. Similarly, in older adults with PD, it has been reported that serum levels of PRL increase as compared to age-matched controls [109]. In this sense, the rise of PRL in older men has been reported to influence cognition, mood and quality of life [110]. Finally, PRL has been recognized to influence bone [111], particularly postmenopausal women with hyperprolactinemia show multiple effects on bone metabolism, affecting both bone mass and density, which in turn is associated with a high risk of osteoporosis [112].
According to this section, PRL levels in aging are controversial since low or high levels lead to the impairment of multiple organs, contributing to the organism's decline. In this context, it is important to highlight that understanding the fluctuation of PRL levels during aging is imperative to designing more effective treatments that target the most common age-related diseases.
10. Conclusions
In addition to its conventional roles, there is ample evidence in a variety of models of the importance of prolactin in the modulation of the immune system and over ion membrane transport, either through direct or indirect mechanisms. All of these elements suggest the participation of prolactin in the pathogenesis of asthma by impacting the immune function and promoting airway smooth muscle contraction through important intracellular signaling pathways.
Author Contributions
Conceptualization: E.C., J.C.G.-V., E.F.S, B.S.M.R. and B.S.; development of themes: M.C., V.R.C., M.E.M.E., L.M.M., H.S.C., A.A.G., N.A.R,S., A.C.S.N., G.E.P.F., V.C., and G.G.A.; data curation, M.E.M.E., H.S.C., G.E.P.F., V.C., G.G.A and A.A.G., writing—original draft: E.C., J.C.G.-V., E.F.-S. and B.S.R.-M.; writing—review and editing: E.C., J.C.G.-V., N.A.R,S., B.S.R.-M., L.M.M., E.F.-S. and B.S.; funding acquisition, B.S. and EFS. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Instituto Nacional de Enfermedades Respiratorias (B.S) and supported by the División de Investigación, Facultad de Medicina, Universidad Nacional Autónoma de México, FM/DI/123/2024. (E.F.S.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would also like to thank Ms Dora Nely Martinez Aguirre for administrative and technical support (National Autonomous University of Mexico, Mexico City 04510).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| PRL | Prolactin |
| GINA | Global Initiative for Asthma |
| HPA | hypophysis-pituitary-adrenal |
| SLE | systemic lupus erythematosus |
| RA | rheumatoid arthritis |
| MS | multiple sclerosis |
| PRLR | Prolactin receptor |
| PMCA | Plasma membrane Ca²⁺-ATPase |
| NCX | Na⁺/Ca²⁺ exchanger |
| ENaC | Epithelial sodium channel |
| ClC4 | Chloride channel |
| FIL | Leukaemia inhibiting factor |
| EPO | Erythropoietin |
| PRF | PRL-releasing factors |
| PIF | PRL inhibitory factors |
| GABA | Gamma-aminobutyric acid |
| DOPA | Dihydroxyphenylalanine |
| DC | Dendritic cell |
| GM-CSF | Granulocyte/macrophage colony-stimulating factor |
| SDCs | CD1c-positive dendritic cells |
| OVA | ovalbumin |
| SLIT | sublingual immunotherapy |
| ECP | eosinophil cationic protein |
| ACTH | adrenocorticotropic hormone |
| PMA | perimenstrual asthma |
| AHR | airway hyperresponsiveness |
References
- GINA. global strategy for asthma management and prevention. 2024.
- Akcan, N.; Bahceciler, N.N. Headliner in Physiology and Management of Childhood Asthma: Hypothalamic-Pituitary-Adrenal Axis. Curr Pediatr Rev. 2020, 16(1), 43–52. [Google Scholar] [CrossRef]
- Parker, V.J.; Menzies, J.R.; Douglas, A.J. Differential changes in the hypothalamic-pituitary-adrenal axis and prolactin responses to stress in early pregnant mice. J Neuroendocrinol. 2011, 23(11), 1066–78. [Google Scholar] [CrossRef]
- Russjan, E. The Role of Peptides in Asthma–Obesity Phenotype. Int J Mol Sci. 2024, 25(6), 3213. [Google Scholar] [CrossRef]
- Afzal, M.; Kazmi, I.; Al-Abbasi, F.A.; Alshehri, S.; Ghoneim, M.M.; Imam, S.S.; Nadeem, M.S.; Al-Zahrani, M.H.; Alzarea, S.I.; Alquraini, A. Current Overview on Therapeutic Potential of Vitamin D in Inflammatory Lung Diseases. Biomedicines. 2021, 9(12), 1843. [Google Scholar] [CrossRef]
- Hernández-Díazcouder, A.; Romero-Nava, R.; Del-Río-Navarro, B.E.; Sánchez-Muñoz, F.; Guzmán-Martín, C.A.; Reyes-Noriega, N.; Rodríguez-Cortés, O.; Leija-Martínez, J.J.; Vélez-Reséndiz, J.M.; Villafaña, S.; et al. The Roles of MicroRNAs in Asthma and Emerging Insights into the Effects of Vitamin D3 Supplementation. Nutrients. 2024, 16(3), 341. [Google Scholar] [CrossRef]
- Sathish, V.; Martin, Y.N.; Prakash, Y.S. Sex steroid signaling: Implications for lung diseases. Pharmacol Ther. 2015, 150, 94–108. [Google Scholar] [CrossRef]
- Ochoa-Amaya, J.E.; Hamasato, E.K.; Tobaruela, C.N.; Queiroz-Hazarbassanov, N.; Anselmo Franci, J.A.; Palermo-Neto, J.; Greiffo, F.R.; de Britto, A.A.; Vieira, R.P.; Ligeiro de Oliveira, A.P.; et al. Short-term hyperprolactinemia decreases allergic inflammatory response of the lungs. Life Sci. 2015, 142, 66–75. [Google Scholar] [CrossRef]
- Insuela, D.B.R.; Daleprane, J.B.; Coelho, L.P.; Silva, A.R.; e Silva, P.M.; Martins, M.A.; Carvalho, V.F. Glucagon induces airway smooth muscle relaxation by nitric oxide and prostaglandin E2. J Endocrinol. 2015, 225(3), 205–17. [Google Scholar] [CrossRef]
- Gochicoa-Rangel, L.; Chávez, J.; Del-Río-Hidalgo, R.; Guerrero-Zúñiga, S.; Mora-Romero, U.; Benítez-Pérez, R.; Rodríguez-Moreno, L.; Torre-Bouscoulet, L.; Vargas, M.H. Lung function is related to salivary cytokines and hormones in healthy children. An exploratory cross-sectional study. Physiol Rep 2023, 11(23). [Google Scholar] [CrossRef]
- Bulkhi, A.A.; Shepard, K.V.; Casale, T.B.; Cardet, J.C. Elevated Testosterone Is Associated with Decreased Likelihood of Current Asthma Regardless of Sex. J Allergy Clin Immunol Pract 2020, 8(9), 3029–35.e4. [Google Scholar] [CrossRef]
- Radzikowska, U.; Golebski, K. Sex hormones and asthma: The role of estrogen in asthma development and severity. Allergy 2023, 78(3), 620–2. [Google Scholar] [CrossRef]
- Townsend, E.A.; Miller, V.M.; Prakash, Y.S. Sex Differences and Sex Steroids in Lung Health and Disease. Endocr Rev. 2012, 33(1), 1–47. [Google Scholar] [CrossRef]
- Romero-Martínez, B.S.; Flores-Soto, E.; Sommer, B.; Reyes-García, J.; Arredondo-Zamarripa, D.; Solís-Chagoyán, H.; Lemini, C.; Rivero-Segura, N.A.; Santiago-de-la-Cruz, J.A.; Pérez-Plascencia, C. 17β-estradiol induces hyperresponsiveness in guinea pig airway smooth muscle by inhibiting the plasma membrane Ca2+-ATPase. Mol Cell Endocrinol. 2024, 590, 112273. [Google Scholar] [CrossRef]
- Giles, W.; Murphy, V. Asthma in pregnancy: a review. Obstet Med. 2013, 6(2), 58–63. [Google Scholar] [CrossRef]
- Borba, V.V.; Zandman-Goddard, G.; Shoenfeld, Y. Prolactin and Autoimmunity. Front Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Dorshkind, K.; Horseman, N.D. The roles of prolactin, growth hormone, insulin-like growth factor-I, and thyroid hormones in lymphocyte development and function: insights from genetic models of hormone and hormone receptor deficiency. Endocr Rev. 2000, 21(3), 292–312. [Google Scholar] [CrossRef]
- Borba, V.V.; Zandman-Goddard, G.; Shoenfeld, Y. Prolactin and autoimmunity: The hormone as an inflammatory cytokine. Best Pract Res Clin Endocrinol Metab. 2019, 33(6), 101324. [Google Scholar] [CrossRef]
- Costanza, M.; Binart, N.; Steinman, L.; Pedotti, R. Prolactin: a versatile regulator of inflammation and autoimmune pathology. Autoimmun Rev. 2015, 14(3), 223–30. [Google Scholar] [CrossRef]
- Walker, S.E.; Jacobson, J.D. Roles of prolactin and gonadotropin-releasing hormone in rheumatic diseases. Rheum Dis Clin North Am. 2000, 26(4), 713–36. [Google Scholar] [CrossRef]
- Walker, S.E. Treatment of systemic lupus erythematosus with bromocriptine. Lupus. 2001, 10(3), 197–202. [Google Scholar] [CrossRef]
- Duc Nguyen, H.; Hoang, N.M.H.; Ko, M.; Seo, D.; Kim, S.; Jo, W.H.; Bae, J.W.; Kim, M.S. Association between Serum Prolactin Levels and Neurodegenerative Diseases: Systematic Review and Meta-Analysis. Neuroimmunomodulation. 2022, 29(2), 85–96. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Jabir, M.S.; Al-Gareeb, A.I.; Albuhadily, A.K. The conceivable role of prolactin hormone in Parkinson disease: The same goal but with different ways. Ageing Res Rev. 2023, 91, 102075. [Google Scholar] [CrossRef]
- Ippoliti, F.; De Santis, W.; Volterrani, A.; Lenti, L.; Canitano, N.; Lucarelli, S.; Frediani, T. Immunomodulation during sublingual therapy in allergic children. Pediatr Allergy Immunol. 2003, 14(3), 216–21. [Google Scholar] [CrossRef]
- Corbacho, A.M.; Valacchi, G.; Kubala, L.; Olano-Martín, E.; Schock, B.C.; Kenny, T.P.; Cross, C.E. Tissue-specific gene expression of prolactin receptor in the acute-phase response induced by lipopolysaccharides. Am J Physiol Endocrinol Metab. 2004, 287(4), E750–7. [Google Scholar] [CrossRef] [PubMed]
- Bole-Feysot, C.; Goffin, V.; Edery, M.; Binart, N.; Kelly, P.A. Prolactin (PRL) and its receptor: actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr Rev. 1998, 19(3), 225–68. [Google Scholar] [CrossRef]
- Sangeeta Devi, Y.; Halperin, J. Reproductive actions of prolactin mediated through short and long receptor isoforms. Mol Cell Endocrinol. 2014, 382(1), 400–10. [Google Scholar] [CrossRef]
- Gorvin, C.M. The prolactin receptor: Diverse and emerging roles in pathophysiology. J Clin Transl Endocrinol. 2015, 2(3), 85–91. [Google Scholar] [CrossRef]
- Molina-Salinas, G.; Rivero-Segura, N.A.; Cabrera-Reyes, E.A.; Rodríguez-Chávez, V.; Langley, E.; Cerbon, M. Decoding signaling pathways involved in prolactin-induced neuroprotection: A review. Front Neuroendocrinol. 2021, 61, 100913. [Google Scholar] [CrossRef]
- Costa-Brito, A.R.; Gonçalves, I.; Santos, C.R.A. The brain as a source and a target of prolactin in mammals. Neural Regen Res. 2022, 17(8), 1695–702. [Google Scholar] [CrossRef]
- Horseman, N.D.; Gregerson, K.A. Prolactin actions. J Mol Endocrinol. 2014, 52(1), R95–R106. [Google Scholar] [CrossRef]
- Marano, R.J.; Ben-Jonathan, N. Minireview: Extrapituitary Prolactin: An Update on the Distribution, Regulation, and Functions. Mol Endocrinol. 2014, 28(5), 622–33. [Google Scholar] [CrossRef]
- Patil, M.J.; Henry, M.A.; Akopian, A.N. Prolactin receptor in regulation of neuronal excitability and channels. Channels. 2014, 8(3), 193–202. [Google Scholar] [CrossRef]
- Molina-Salinas, G.; Rodríguez-Chávez, V.; Langley, E.; Cerbon, M. Prolactin-induced neuroprotection against excitotoxicity is mediated via PI3K/AKT and GSK3 β /NF- κB in primary cultures of hippocampal neurons. Peptides. 2023, 166, 171037. [Google Scholar] [CrossRef]
- Dorkkam, N.; Wongdee, K.; Suntornsaratoon, P.; Krishnamra, N.; Charoenphandhu, N. Prolactin stimulates the L-type calcium channel-mediated transepithelial calcium transport in the duodenum of male rats. Biochem Biophys Res Commun. 2013, 430(2), 711–6. [Google Scholar] [CrossRef]
- Liu, T.T.; Qu, Z.W.; Ren, C.; Gan, X.; Qiu, C.Y.; Hu, W.P. Prolactin potentiates the activity of acid-sensing ion channels in female rat primary sensory neurons. Neuropharmacology. 2016, 103, 174–82. [Google Scholar] [CrossRef]
- Greenlee, M.M.; Mitzelfelt, J.D.; Duke, B.J.; Al-Khalili, O.; Bao, H.F.; Eaton, D.C. Prolactin stimulates sodium and chloride ion channels in A6 renal epithelial cells. Am J Physiol Renal Physiol. 2015, 308(7), F697–705. [Google Scholar] [CrossRef]
- Cabrera-Reyes, E.A.; Limón-Morales, O.; Rivero-Segura, N.A.; Camacho-Arroyo, I.; Cerbón, M. Prolactin function and putative expression in the brain. Endocrine. 2017, 57(2), 199–213. [Google Scholar] [CrossRef]
- Bernard, V.; Young, J.; Binart, N. Prolactin-a pleiotropic factor in health and disease. Nat Rev Endocrinol. 2019, 15(6), 356–65. [Google Scholar] [CrossRef]
- Bates, R.W.; Lahr, E.L.; Riddle, O. The gross action of prolactin and follicle-stimulating hormone on the mature ovary and sex accessories of fowl. Am J Physiol Content. 1935, 111, 361–8. [Google Scholar] [CrossRef]
- Freeman, M.E.; Kanyicska, B.; Lerant, A.; Nagy, G. Prolactin: structure, function, and regulation of secretion. Physiol Rev. 2000, 80(4), 1523–631. [Google Scholar] [CrossRef]
- Grattan, D.R.; Kokay, I.C. Prolactin: A Pleiotropic Neuroendocrine Hormone. J Neuroendocrinol. 2008, 20(6), 752–63. [Google Scholar] [CrossRef]
- Bridges, R.S.; Grattan, D.R. 30 years after: CNS actions of prolactin: Sources, mechanisms and physiological significance. J Neuroendocrinol. 2019, 31(3), e12669. [Google Scholar] [CrossRef]
- Huising, M.O.; Kruiswijk, C.P.; Flik, G. Phylogeny and evolution of class-I helical cytokines. J Endocrinol. 2006, 189(1), 1–25. [Google Scholar] [CrossRef]
- Fitzgerald, P.; Dinan, T.G. Prolactin and dopamine: what is the connection? A review article. J Psychopharmacol. 2008, 22, 12–9. [Google Scholar] [CrossRef]
- Binart, N. Prolactin and pregnancy in mice and humans. Ann Endocrinol. 2016, 77(2), 126–7. [Google Scholar] [CrossRef]
- Carré, N.; Binart, N. Prolactin and adipose tissue. Biochimie. 2014, 97, 16–21. [Google Scholar] [CrossRef]
- Costanza, M.; Pedotti, R. Prolactin: Friend or Foe in Central Nervous System Autoimmune Inflammation? Int J Mol Sci. 2016, 17(12), 2026. [Google Scholar] [CrossRef]
- Guh, Y.J.; Lin, C.H.; Hwang, P.P. Osmoregulation in zebrafish: ion transport mechanisms and functional regulation. EXCLI J. 2015, 14, 627–59. [Google Scholar] [CrossRef]
- Carretero, J.; Sánchez-Robledo, V.; Carretero-Hernández, M.; Catalano-Iniesta, L.; García-Barrado, M.J.; Iglesias-Osma, M.C.; Blanco, E.J. Prolactin system in the hippocampus. Cell Tissue Res. 2019, 375(1), 193–9. [Google Scholar] [CrossRef]
- Triebel, J.; Robles, J.P.; Zamora, M.; Clapp, C.; Bertsch, T. New horizons in specific hormone proteolysis. Trends Endocrinol Metab. 2022, 33(6), 371–7. [Google Scholar] [CrossRef]
- Hattori, N.; Adachi, T.; Ishihara, T.; Shimatsu, A. The natural history of macroprolactinaemia. Eur J Endocrinol. 2012, 166(4), 625–9. [Google Scholar] [CrossRef]
- Bernard, V.; Villa, C.; Auguste, A.; Lamothe, S.; Guillou, A.; Martin, A.; Caburet, S.; Young, J.; Veitia, R.A.; Binart, N. Natural and molecular history of prolactinoma: insights from a Prlr-/– mouse model. Oncotarget. 2018, 9(5), 6144–55. [Google Scholar] [CrossRef]
- Bernard, V.; Young, J.; Chanson, P.; Binart, N. New insights in prolactin: pathological implications. Nat Rev Endocrinol. 2015, 11(5), 265–75. [Google Scholar] [CrossRef]
- Clapp, C.; Aranda, J.; González, C.; Jeziorski, M.C.; Martínez de la Escalera, G. Vasoinhibins: endogenous regulators of angiogenesis and vascular function. Trends Endocrinol Metab. 2006, 17(8), 301–7. [Google Scholar] [CrossRef]
- Macotela, Y.; Ruiz-Herrera, X.; Vázquez-Carrillo, D.I.; Ramírez-Hernandez, G.; Martínez De La Escalera, G.; Clapp, C. The beneficial metabolic actions of prolactin. Front Endocrinol. 2022, 13. [Google Scholar] [CrossRef]
- Vergara-Castañeda, E.; Grattan, D.R.; Pasantes-Morales, H.; Pérez-Domínguez, M.; Cabrera-Reyes, E.A.; Morales, T.; Cerbón, M. Prolactin mediates neuroprotection against excitotoxicity in primary cell cultures of hippocampal neurons via its receptor. Brain Res. 2016, 1636, 193–9. [Google Scholar] [CrossRef]
- Bu, G.; Liang, X.; Li, J.; Wang, Y. Extra-pituitary prolactin (PRL) and prolactin-like protein (PRL-L) in chickens and zebrafish. Gen Comp Endocrinol. 2015, 220, 143–53. [Google Scholar] [CrossRef]
- Torner, L. Actions of Prolactin in the Brain: From Physiological Adaptations to Stress and Neurogenesis to Psychopathology. Front Endocrinol. 2016, 7. [Google Scholar] [CrossRef]
- Farrar, V.S.; Harris, R.M.; Austin, S.H.; Nava Ultreras, B.; Booth, A.M.; Angelier, F.; Lang, A.S.; Feustel, T.; Lee, C.; Bond, A. Prolactin and prolactin receptor expression in the HPG axis and crop during parental care in both sexes of a biparental bird (Columba livia). Gen Comp Endocrinol. 2022, 315, 113940. [Google Scholar] [CrossRef]
- Wilson, A.B.; Whittington, C.M.; Meyer, A.; Scobell, S.K.; Gauthier, M.E. Prolactin and the evolution of male pregnancy. Gen Comp Endocrinol. 2023, 334, 114210. [Google Scholar] [CrossRef]
- Laud, K.; Gourdou, I.; Belair, L.; Peyrat, J.P.; Djiane, J. Characterization and modulation of a prolactin receptor mRNA isoform in normal and tumoral human breast tissues. Int J Cancer. 2000, 85(6), 771–6. [Google Scholar] [CrossRef]
- Fleming, J.M.; Ginsburg, E.; McAndrew, C.W.; Heger, C.D.; Cheston, L.; Rodriguez-Canales, J.; Vonderhaar, B.K.; Goldsmith, P. Characterization of Δ7/11, a functional prolactin-binding protein. J Mol Endocrinol. 2012, 50(2), 79–90. [Google Scholar] [CrossRef]
- Ben-Jonathan, N.; Mershon, J.L.; Allen, D.L.; Steinmetz, R.W. Extrapituitary prolactin: distribution, regulation, functions, and clinical aspects. Endocr Rev. 1996, 17(6), 639–69. [Google Scholar] [CrossRef] [PubMed]
- Grattan, D.R. 60 YEARS OF NEUROENDOCRINOLOGY: The hypothalamo-prolactin axis. J Endocrinol. 2015, 226(2), 101–22. [Google Scholar] [CrossRef]
- Gustafson, P.; Kokay, I.; Sapsford, T.; Bunn, S.; Grattan, D. Prolactin regulation of the HPA axis is not mediated by a direct action upon CRH neurons: evidence from the rat and mouse. Brain Struct Funct. 2017, 222(7), 3191–204. [Google Scholar] [CrossRef]
- Jayakumar, P.; Martínez-Moreno, C.G.; Lorenson, M.Y.; Walker, A.M.; Morales, T. Prolactin Attenuates Neuroinflammation in LPS-Activated SIM-A9 Microglial Cells by Inhibiting NF-κB Pathways Via ERK1/2. Cell Mol Neurobiol. 2022, 42(7), 2171–86. [Google Scholar] [CrossRef]
- Martinez-Campos, A.; Giovannini, P.; Parati, E.; Novelli, A.; Caraceni, T.; Müller, E.E. Growth hormone and prolactin stimulation by Madopar in Parkinson's disease. J Neurol Neurosurg Psychiatry. 1981, 44(12), 1116–23. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Reyes, E.A.; Vanoye–Carlo, A.; Rodríguez-Dorantes, M.; Vázquez-Martínez, E.R.; Rivero-Segura, N.A.; Collazo-Navarrete, O.; Cerbón, M. Transcriptomic analysis reveals new hippocampal gene networks induced by prolactin. Sci Rep 2019, 9(1). [Google Scholar] [CrossRef]
- Reem, G.H.; Ray, D.W.; Davis, J.R. The human prolactin gene upstream promoter is regulated in lymphoid cells by activators of T-cells and by cAMP. J Mol Endocrinol. 1999, 22(3), 285–92. [Google Scholar] [CrossRef]
- Torner, L.; Neumann, I.D. The Brain Prolactin System: Involvement in Stress Response Adaptations in Lactation. Stress. 2002, 5(4), 249–57. [Google Scholar] [CrossRef]
- Walker, T.L.; Vukovic, J.; Koudijs, M.M.; Blackmore, D.G.; Mackay, E.W.; Sykes, A.M.; Overall, R.W.; Hamlin, A.S.; Bartlett, P.F. Prolactin Stimulates Precursor Cells in the Adult Mouse Hippocampus. PLoS ONE. 2012, 7(9), e44371. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Segura, N.A.; Flores-Soto, E.; García De La Cadena, S.; Coronado-Mares, I.; Gomez-Verjan, J.C.; Ferreira, D.G.; Cabrera-Reyes, E.A.; Lopes, L.V.; Massieu, L.; Cerbón, M. Prolactin-induced neuroprotection against glutamate excitotoxicity is mediated by the reduction of [Ca2+]i overload and NF-κB activation. PLOS ONE. 2017, 12(5), e0176910. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Moratalla, A.; Martín, E.D. Prolactin enhances hippocampal synaptic plasticity in female mice of reproductive age. Hippocampus. 2021, 31(3), 281–93. [Google Scholar] [CrossRef]
- Rodríguez-Chávez, V.; Flores-Soto, E.; Molina-Salinas, G.; Martínez-Razo, L.D.; Montaño, L.M.; Cerbón, M. Prolactin reduces the kainic acid-induced increase in intracellular Ca2+ concentration, leading to neuroprotection of hippocampal neurons. Neurosci Lett. 2023, 810, 137344. [Google Scholar] [CrossRef] [PubMed]
- Anaya, J.M.; Shoenfeld, Y. Multiple autoimmune disease in a patient with hyperprolactinemia. Isr Med Assoc J. 2005, 7(11), 740–1. [Google Scholar]
- Vieira Borba, V.; Sharif, K.; Shoenfeld, Y. Breastfeeding and autoimmunity: Programing health from the beginning. Am J Reprod Immunol 2018, 79(1). [Google Scholar] [CrossRef]
- Saha, S.; Gonzalez, J.; Rosenfeld, G.; Keiser, H.; Peeva, E. Prolactin alters the mechanisms of B cell tolerance induction. Arthritis Rheum. 2009, 60(6), 1743–52. [Google Scholar] [CrossRef]
- Richards, S.M.; Garman, R.D.; Keyes, L.; Kavanagh, B.; McPherson, J.M. Prolactin is an antagonist of TGF-beta activity and promotes proliferation of murine B cell hybridomas. Cell Immunol. 1998, 184(2), 85–91. [Google Scholar] [CrossRef]
- Peeva, E.; Zouali, M. Spotlight on the role of hormonal factors in the emergence of autoreactive B-lymphocytes. Immunol Lett. 2005, 101(2), 123–43. [Google Scholar] [CrossRef]
- Athreya, B.H.; Pletcher, J.; Zulian, F.; Weiner, D.B.; Williams, W.V. Subset-specific effects of sex hormones and pituitary gonadotropins on human lymphocyte proliferation in vitro. Clin Immunol Immunopathol. 1993, 66(3), 201–11. [Google Scholar] [CrossRef]
- Matera, L.; Galetto, A.; Geuna, M.; Vekemans, K.; Ricotti, E.; Contarini, M.; Moro, F.; Basso, G. Individual and combined effect of granulocyte–macrophage colony-stimulating factor and prolactin on maturation of dendritic cells from blood monocytes under serum-free conditions. Immunology. 2000, 100(1), 29–36. [Google Scholar] [CrossRef]
- Matera, L.; Mori, M.; Galetto, A. Effect of prolactin on the antigen presenting function of monocyte-derived dendritic cells. Lupus. 2001, 10(10), 728–34. [Google Scholar] [CrossRef]
- McMurray, R.W. Estrogen, prolactin, and autoimmunity: actions and interactions. Int Immunopharmacol. 2001, 1(6), 995–1008. [Google Scholar] [CrossRef]
- Vera-Lastra, O.; Jara, L.J.; Espinoza, L.R. Prolactin and autoimmunity. Autoimmun Rev. Autoimmun Rev. 2002, 1(6), 360–4. [Google Scholar] [CrossRef]
- Yu-Lee, L.Y. Prolactin modulation of immune and inflammatory responses. Recent Prog Horm Res. 2002, 57, 435–55. [Google Scholar] [CrossRef]
- Yang, L.; Hu, Y.; Li, X.; Zhao, J.; Hou, Y. Prolactin modulates the functions of murine spleen CD11c-positive dendritic cells. Int Immunopharmacol. 2006, 6(9), 1478–86. [Google Scholar] [CrossRef] [PubMed]
- Jara, L.J.; Benitez, G.; Medina, G. Prolactin, dendritic cells, and systemic lupus erythematosus. Autoimmun Rev. 2008, 7(3), 251–5. [Google Scholar] [CrossRef] [PubMed]
- Chikanza, I.C. Prolactin and neuroimmunomodulation: in vitro and in vivo observations. Ann N Y Acad Sci. 1999, 876, 119–30. [Google Scholar] [CrossRef] [PubMed]
- Walton, P.E.; Cronin, M.J. Tumor necrosis factor-alpha and interferon-gamma reduce prolactin release in vitro. Am J Physiol. 1990, 259(5), E672–6. [Google Scholar] [CrossRef]
- Koike, K.; Masumoto, N.; Kasahara, K.; Yamaguchi, M.; Tasaka, K.; Hirota, K.; Miyake, A.; Tanizawa, O. Tumor necrosis factor-alpha stimulates prolactin release from anterior pituitary cells: a possible involvement of intracellular calcium mobilization. Endocrinology. 1991, 128(6), 2785–90. [Google Scholar] [CrossRef]
- Theas, S.; Pisera, D.; Duvilanski, B.; De Laurentiis, A.; Pampillo, M.; Lasaga, M.; Seilicovich, A. Estrogens modulate the inhibitory effect of tumor necrosis factor-alpha on anterior pituitary cell proliferation and prolactin release. Endocrine. 2000, 12(3), 249–55. [Google Scholar] [CrossRef] [PubMed]
- Majumder, R.; Nguyen, T. Protein S: function, regulation, and clinical perspectives. Curr Opin Hematol. 2021, 28(5), 339–44. [Google Scholar] [CrossRef]
- Matera, L.; Mori, M. Cooperation of Pituitary Hormone Prolactin with Interleukin-2 and Interleukin-12 on Production of Interferon-γ by Natural Killer and T Cells. Ann N Y Acad Sci. 2000, 917(1), 505–13. [Google Scholar] [CrossRef]
- Kumar, A;. Singh, S.M.; Sodhi, A. Effect of prolactin on nitric oxide and interleukin-1 production of murine peritoneal macrophages: role of Ca2+ and protein kinase C. Int J Immunopharmacol 1997, 19(3), 129–33. [CrossRef] [PubMed]
- Tseng, Y.H.; Kessler, M.A.; Schuler, L.A. Regulation of interleukin (IL)-1alpha, IL-1beta, and IL-6 expression by growth hormone and prolactin in bovine thymic stromal cells. Mol Cell Endocrinol 1997, 128(1-2), 117–27. [Google Scholar] [CrossRef]
- Song, G.G.; Lee, Y.H. Circulating prolactin level in systemic lupus erythematosus and its correlation with disease activity: a meta-analysis. Lupus. 2017, 26(12), 1260–8. [Google Scholar] [CrossRef] [PubMed]
- Praprotnik, S.; Agmon-Levin, N.; Porat-Katz, B.S.; Blank, M.; Meroni, P.L.; Cervera, R.; Miesbach, W.; Stojanovich, L.; Szyper-Kravitz, M.; Rozman, B.; et al. Prolactin's role in the pathogenesis of the antiphospholipid syndrome. Lupus. 2010, 19(13), 1515–9. [Google Scholar] [CrossRef]
- Walker, S.E.; Allen, S.H.; Hoffman, R.W.; McMurray, R.W. Prolactin: a stimulator of disease activity in systemic lupus erythematosus. Lupus. 1995, 4(1), 3–9. [Google Scholar] [CrossRef]
- Leaños-Miranda, A.; Cárdenas-Mondragón, G. Serum free prolactin concentrations in patients with systemic lupus erythematosus are associated with lupus activity. Rheumatology. 2006, 45(1), 97–101. [Google Scholar] [CrossRef]
- McMurray, R.; Keisler, D.; Kanuckel, K.; Izui, S.; Walker, S.E. Prolactin influences autoimmune disease activity in the female B/W mouse. J Immunol. 1991, 147(11), 3780–7. [Google Scholar] [CrossRef]
- McMurray, R.W.; Weidensaul, D.; Allen, S.H.; Walker, S.E. Efficacy of bromocriptine in an open label therapeutic trial for systemic lupus erythematosus. J Rheumatol. 1995, 22(11), 2084–91. [Google Scholar]
- Ochoa-Amaya, J.E.; Marino, L.P.; Tobaruela, C.N.; Namazu, L.B.; Calefi, A.S.; Margatho, R.; Gonçalves, V., Jr.; Queiroz-Hazarbassanov, N.; Klein, M.O.; Palermo-Neto, J.; et al. Attenuated allergic inflammatory response in the lungs during lactation. Life Sci. 2016, 151, 281–7. [Google Scholar] [CrossRef] [PubMed]
- Semik-Orzech, A.; Skoczyński, S.; Pierzchała, W. Serum estradiol concentration, estradiol-toprogesterone ratio and sputum IL-5 and IL-8 concentrations are increased in luteal phase of the menstrual cycle in perimenstrual asthma patients. Eur Ann Allergy Clin Immunol. 2017, 49(04), 161. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lai, Q.; Wang, C.; Zhou, G. Protective Effects of Herba Houttuyniae Aqueous Extract against OVA-Induced Airway Hyperresponsiveness and Inflammation in Asthmatic Mice. Evid Based Complement Alternat Med. 2022, 2022, 1–11. [Google Scholar] [CrossRef]
- Arnold, E.; Thébault, S.; Aroña, R.; Martínez de la Escalera, G.; Clapp, C. Prolactin mitigates deficiencies of retinal function associated with aging. Neurobiol Aging. 2020, 85, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Leung, Y.Y.; Toledo, J.B.; Nefedov, A.; Polikar, R.; Raghavan, N.; Xie, S.X.; Farnum, M.; Schultz, T.; Baek, Y.; Deerlin, V.V.; et al. Identifying amyloid pathology–related cerebrospinal fluid biomarkers for Alzheimer's disease in a multicohort study. Alzheimers Dement. 2015, 1(3), 339–48. [Google Scholar] [CrossRef]
- Zverova, M.; Kitzlerova, E.; Fisar, Z.; Jirak, R.; Hroudova, J.; Benakova, H.; Lelkova, P.; Martasek, P.; Raboch, J. Interplay between the APOE Genotype and Possible Plasma Biomarkers in Alzheimer's Disease. Curr Alzheimer Res. 2018, 15(10), 938–50. [Google Scholar] [CrossRef]
- Winkler, A.S.; Landau, S.; Chaudhuri, K.R. Serum prolactin levels in Parkinson's disease and multiple system atrophy. Clin Auton Res. 2002, 12(5), 393–8. [Google Scholar] [CrossRef]
- Nitkowska, M.; Tomasiuk, R.; Czyżyk, M.; Friedman, A. Prolactin and sex hormones levels in males with Parkinson's disease. Acta Neurol Scand. 2015, 131(6), 411–6. [Google Scholar] [CrossRef]
- Chiloiro, S.; Giampietro, A.; Bianchi, A.; De Marinis, L. Prolactinoma and Bone. Curr Opin Endocr Metab Res. 2018, 3, 21–4. [Google Scholar] [CrossRef]
- Yun, S.J.; Sang, H.; Park, S.Y.; Chin, S.O. Effect of Hyperprolactinemia on Bone Metabolism: Focusing on Osteopenia/Osteoporosis. Int J Mol Sci. 2024, 25(3), 1474. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PRL and PRLR structures. a) PRL structure according to PDB 1RW5. b) PRLR structure according to PDB 1BP3.
Figure 1.
PRL and PRLR structures. a) PRL structure according to PDB 1RW5. b) PRLR structure according to PDB 1BP3.
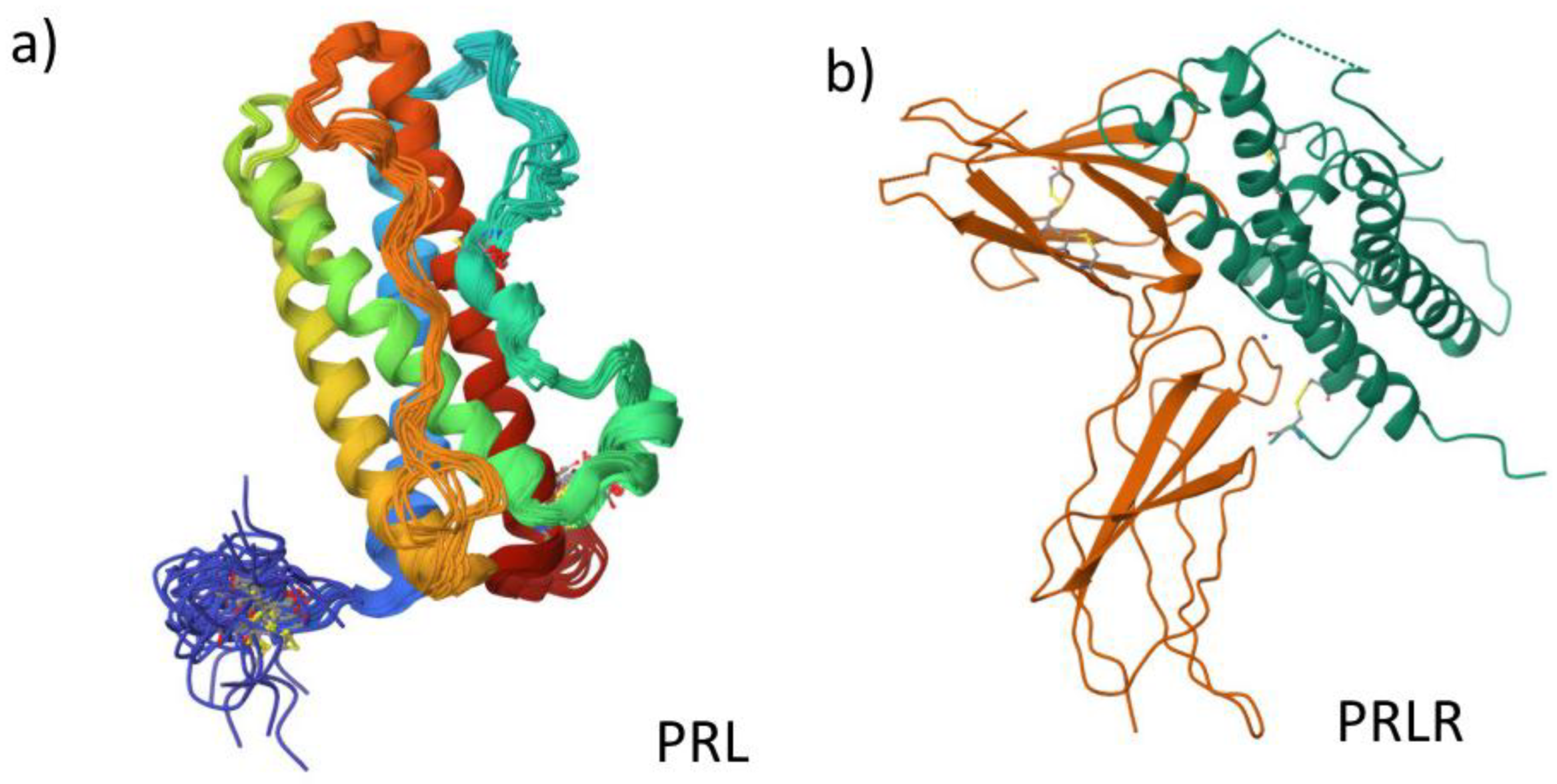
Figure 2.
PRL interactions. This figure indicates the network biology of PRL according to the STRING database. On the right side of the figure, we can see the types of interactions, network properties, and enrichment analysis of the main pathways involved in the PRL network.
Figure 2.
PRL interactions. This figure indicates the network biology of PRL according to the STRING database. On the right side of the figure, we can see the types of interactions, network properties, and enrichment analysis of the main pathways involved in the PRL network.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



