Submitted:
04 June 2025
Posted:
04 June 2025
You are already at the latest version
Abstract
This work explores the application of yeast in beer brewing and winemaking, comparing traditional genetic manipulation methods with innovative techniques, both GMO-free and GMO-based. Traditional approaches, such as sexual breeding and random mutagenesis, are contrasted with modern methods like Adaptive Laboratory Evolution (ALE), the integration of big data, AI, and omics technologies, and synthetic microbial communities, which do not involve genetic modification. GMO-based techniques, including synthetic biology and CRISPR/Cas9, enable more precise and efficient genome modifications, making multiplex genome engineering scalable, thanks to the efficient recombination machinery of Saccharomyces cerevisiae. Despite the promise of these advanced techniques, the commercialization of GMO-based methods faces significant challenges.
Keywords:
brewing
; genome editing
; synthetic biology
; synthetic genomic
; machine-learning
; adaptive laboratory evolution
1. Introduction
Yeasts have played a crucial role in biotechnology for millennia. The first biotechnological process involving yeasts dates back to 6000 BCE during the ancient Egyptian period, specifically for beer production. At the same time, the Sumerians also employed yeasts for bread-making [1]. However, the role of yeasts in fermentation remained unknown until 1876, when Pasteur demonstrated that fermentation is driven by microorganisms. Another significant advancement occurred in 1881 when Hansen successfully isolated the first strain of brewing yeast [2]. Over the years, yeasts have been regarded as model organisms, with many Nobel laureates having worked with these microorganisms. Today, it is well established that yeasts can be utilized for various purposes, such as cellular factories for producing not only food, bioethanol, and chemicals but also many recombinant proteins, including roughly 15% of all protein-based biopharmaceuticals for humans on the market [3]. In the NGS era, yeast is popular model for population genetics, microbial ecology, and synthetic biology [3,4,5,6,7].
The use of yeast starter cultures in the brewing and winemaking industries has been a well-established practice since the middle of the 20th century. Beer is the third most consumed beverage worldwide, following by water and tea [8], and it holds the largest market segment within the global alcoholic beverage sector [9,10]. The global craft beer market is expanding, driven by diverse flavor profiles and appeal for local brands [9,11] Craft beer drinkers are highly engaged and willing to pay a premium for unique beers, presenting opportunities for further market growth [9,11].
In alcoholic fermentation, Saccharomyces species are widely employed, due to their superior resistance to high sugar concentrations (200-300 g/L), low pH (3-4), and elevated ethanol concentrations [12]. Alcoholic fermentation in beer, driven by yeast, can be categorized into two main types: ale yeast and lager yeast. Lager-style beer dominates the market, accounting for approximately 90% of global beer production. These two yeast groups primarily differ in their optimal growth temperatures. Ale yeasts, represented by Saccharomyces cerevisiae, are primarily used for ale beer production. They grow best within an optimal temperature range of 15–26 °C. In contrast, lager yeast, represented by Saccharomyces pastorianus, are used for cold-fermented beer production. These yeasts are cold-tolerant, thriving in temperatures ranging from 8–15 °C [2,13].
Lager beer yeasts, classified as S. pastorianus, are natural hybrids derived from the crossing of S. cerevisiae and Saccharomyces eubayanus [14]. The hybridization provides several competitive advantages, including cryotolerance, inherited from S. eubayanus, and efficient maltotriose utilization, a trait of S. cerevisiae, both of which are particularly beneficial for low-temperature brewing [15,16]. It is hypothesized that the hybridization between S. cerevisiae and S. eubayanus, leading to the formation of S. pastorianus, occurred in the Munich Hofbräuhaus between 1602 and 1615, during a period when both wheat beer and lager were being brewed simultaneously [17]. All S. pastorianus strains have an approximately diploid S. eubayanus subgenome, but some strains (referred to as Group 1 or Saaz strains) have a generally incomplete and haploid S. cerevisiae subgenome, while other strains (called Group 2 or Frohberg) have a diploid or higher S. cerevisiae subgenome. Other notable natural interspecific hybrids include those between S. cerevisiae and Saccharomyces kudriavzevii [2], which have been used in brewing of Belgian-style ales. These hybrids illustrate the diverse evolutionary trajectories within the Saccharomyces genus, highlighting their central role in beer innovation.
In addition to alcoholic fermentation, yeasts significantly contribute to the flavor and taste of beer and wine which are the major determinants of beer and wine quality and consumer preferences [18]. Beer flavor research focuses on specific metabolites and genes directly influencing quality and taste [19]. For instance, genes like EEB1, ETR1, and ATF1 affect ester production, while ARO9 and ARO10 are essential for higher alcohol synthesis [19,20]. However, the complete metabolic pathways and regulatory mechanisms of flavor compounds in beer and wine remain incompletely understood [18].
There is an increasing demand from the brewing and winemaking industry for novel and more diverse options for fermentation which can offer significant advantages in industrial demands. Current efforts focus on improving production processes to ensure high-quality beer [21], expanding flavor profiles, and developing beer with reduced sugar and alcohol content to promote human health. Despite innovative techniques, genetically modified (GM) yeasts are not yet permitted in many countries, including the European Union, for ethical reasons. Classical techniques that do not produce GMOs, thereby avoiding stringent regulations, include mutagenesis and hybridization [22]. These methods aim to enhance and combine traits under polygenic control, such as fermentation capacity, ethanol tolerance, or the absence of off-flavors. While these techniques do not produce GMOs, they pose challenges in adding or removing characteristics without altering strain performance [23]. When considering genetic improvement techniques, it is crucial to first define whether the target trait is monogenic or polygenic to select the optimal approach. Generally, the most important oenological traits, such as yield, ethanol tolerance, or temperature-dependent growth profiles, depend on multiple loci (QTLs) distributed across the genome and are thus poorly characterized [23].
Ongoing research aims to identify new natural yeast strains to enhance wine and beer quality or to use selected yeasts in place of relying on spontaneous fermentation with indigenous yeasts [24]. Yeasts found in Patagonia, for example, exhibit high ethanol tolerance [25]. A particular case is Lachancea cidri, which demonstrated a strong ability to ferment wine and hydromel, as well as to produce acetic and succinic acids [25]. Similarly, yeasts isolated from the American Midwest, such as Hanseniaspora vineae, Lachancea fermentati, Lachancea thermotolerans, Schizosaccharomyces japonicus, and Wickerhamomyces anomalus, are of interest because they can substitute lactic acid bacteria (LAB) in wort fermentation, owing to their ability to produce both ethanol and lactic acid. This is particularly relevant to produce sour beer [26]. However, further genomic studies are needed, as some strains of the same species contradict the hypothesis of a common metabolic process for heterolactic yeasts [27]. Clonal selection in this context involves isolating specific yeast strains from a natural population and propagating them while preserving distinctive traits that are useful for winemaking [28]. Yeasts from food fermentations (e.g., cacao, kombucha, sourdough) also hold potential for brewing applications [29].
In this work, we aim to outline and describe a range of both GMO and non-GMO techniques designed for the genetic optimization of yeast, ultimately enhancing the quality of the final product. There is no universal approach to yeast improvement—different techniques must be applied based on specific needs, depending on the desired phenotype and the available knowledge of its genetic basis (Table 1).
2. GMO-Free Genetic Improvement Techniques
2.1. Classical Genetic Improvement Techniques for Non-GMO Yeasts
2.1.1. Random Mutagenesis
One traditional genetic improvement technique is random mutagenesis, aimed at eliminating undesirable traits. Since the spontaneous mutation rate in S. cerevisiae is generally low, approximately 10-6 per generation, and even lower than that of meiotic genetic conversion [12], mutagens such as UV radiation or chemical agents, including ethyl methanesulfonate (EMS), methyl methanesulfonate (MMS), N-methyl-N’-nitro-N-nitrosoguanidine (MNNG), and nitrous acid (NA) [23], are employed. The resulting library of mutagenized strains are submitted to either positive or negative selection processes by replica plating, as shown in Figure 2 [12]. The main constraint of this technique is that mutants can only be isolated in phenotypes that in one way or another can be selected under specific conditions.
The technique is also constrained by the genomic structure of industrial yeasts, which can be diploid or polyploid. While mutations occur at similar frequencies in haploid, diploid, and polyploid cells, they are easier to identify in haploids because diploids and polyploids retain unmutated alleles, making dominant mutations with phenotypic effects more challenging to detect [23]. Additionally, while mutations can improve specific traits, they may simultaneously impair others [30].
Random mutagenesis has been applied to the improvement of wine and beer yeasts, particularly in S. pastorianus, whose genome is inherently unstable and thus more susceptible to mutagenic effects associated with chromosome copy number variation (CCNV). A summary of key mutagenesis-based yeast improvements is provided in Table 2.
2.1.2. Sexual Hybridization
Another traditional genetic improvement technique is sexual reproduction, which exploits genetic diversity generated by sexuality to develop new strains with desirable traits [23]. According to Giudici et al. [22], this technique can also be used to enhance enological traits, such as ethanol tolerance, which involves more than 250 genes [12]. Figure 2 shows the spore-to-spore method, which involves placing spores from two parental strains in proximity, allowing them to mate. It offers high hybridization frequencies and greater genetic stability but requires that the parental strains produce viable spores, and physiological traits may be lost during meiotic recombination [38]. To counteract this last inconvenience, monosporic cultures with the desired phenotype can be constructed by exploiting haplo-selfing mechanism in homothallic yeasts and then used as parental lineages, fixing the desired phenotype in zygote.
Mating is possible only between haploid cells; however, commercial yeasts often sporulate poorly or do not produce viable haploid gametes. Given the challenges associated with using spores, rare mating can also be employed, especially when spore viability is low or mating frequencies are insufficient. Rare mating relies on the occasional occurrence of an erroneous mating-type switch, where some individuals in a diploid or polyploid cell population become either MATa/MATa or MATα/MATα homozygous and can mate with a haploid cell of the opposite mating type or even with a homozygous diploid cell of the opposite mating type [12].
A summary of key hybridization-based yeast improvements in beer and winemaking is provided in Table 3.
While interspecific hybridization among Saccharomyces species, such as between Saccharomyces uvarum (cryotolerant) and S. cerevisiae (non-cryotolerant), has been successful, these hybrids are often sterile, limiting the selection process to the F1 generation and reducing the potential for further recombination [22]. The creation of innovative yeast hybrids can enhance fermentation, stress tolerance, sugar utilization, and aroma compound production. However, there are still many unexplored areas, such as antioxidant formation, flavor reduction, and increased glycerol production for low-alcohol beers. Research has predominantly focused on hybrids created with S. cerevisiae or S. eubayanus, but other Saccharomyces species could serve as valuable alternatives [43].
2.2. Innovative Genetic Improvement Techniques in Fermentation
2.2.1. Adaptive Laboratory Evolution - ALE
Among the innovative non-GMO techniques for strain improvement, Adaptive Laboratory Evolution (ALE) relies on genetic diversity and selective advantages under specific conditions. Figure 3 shows a schematic representation of ALE method, which enhances desirable beer characteristics and mitigates undesirable ones [15]. In brewing, ALE extends the process that led to the domestication of strains used in commercial breweries. Most domesticated beer strains exhibit traits such as ethanol tolerance and the absence of phenolic aromas, which have evolved through natural selection [44,45]. Early brewers realized that using a small portion of completed fermentation to initiate a new one (“backslopping”) led to faster and more reliable fermentations [46]. These wild yeasts may hold potential for beer production [47].
Advances in yeast biology now allow for the rational development of traits to meet specific brewers’ needs [14]. ALE is used to improve industrial microorganisms for biological processes, such as food, pharmaceutical, and chemical production. Short generation times, which can be increased with mutagens, transposons, or mutator strains [48], and easy culture conditions make yeasts ideal candidates for ALE [49] which relies on genetic diversity and selective advantages in specific conditions [15]. Selectable traits include growth rate, vitality, and substrate consumption, while complex traits require mating strategies [50]. ALE was first applied by Dallinger in the late 19th century in a high-temperature adaptation experiment and has since been used in various organisms, including E. coli, viruses, and microalgae [49]. During ALE, natural selection is directed toward a selected environment, aiming to improve the physical condition of populations in relation to the desired conditions, such as the production of high-value metabolites (ethanol, butanol, lactic acid, carotenoids, etc.) [51].
Organisms adapt to new environments through genetic modifications, such as single nucleotide mutations, frameshifts, insertions, or deletions, which can modify open reading frames, alter regulatory sequences or chromatin structure, and affect protein or gene expression [6]. Structural variations (SV) events, such as copy number variations (CNV), play a significant role in evolutionary adaptation by causing changes in gene dosages. For example, the increased copy number of genes related to glycolysis, nutrient transporters, and stress resistance is a common adaptation in Saccharomyces cerevisiae [52]. Batch and continuous cultures in bioreactors can be used to optimize evolution [53]. The selection pressure can be constant or dynamic, favoring constitutive adaptations. A summary of key yeast strain improvements achieved through ALE is provided in Table 4.
Despite its advantages, ALE requires significant time, intensive monitoring, and aseptic conditions to avoid contamination [64]. Recently, automation has been introduced to improve ALE experiment efficiency, overcoming difficulties associated with manual execution [51]. Automation tools like eVOLVER optimize multiple cultures management by enabling simultaneous execution of multiple independent experimental versions. ALEsim is a simulator which helps in designing ALE experiments with E. Coli, while VERT system uses fluorescent markers to monitor adaptation events in isogenic strains [64]. Evolved clones are sequenced to identify adaptive mutations, and backcrossing or QTL analysis can further support the identification of genetic bases underpinning complex traits [53].
These advancements, if implemented, expand the application of ALE, generating faster novel strains for diversification and innovation in the food and beverage industries.
2.2.2. Big Data, AI and Omics
Accurate phenotype prediction from genotypes is a cornerstone in biology. Omics technologies, including genomics, proteomics, metabolomics, and transcriptomics, are revolutionizing our understanding of complex biological systems. The integration of these multiple omics’ datasets offer unprecedented opportunities to reconstruct metabolic networks at the genome scale with information on stoichiometry, compartmentalization, biomass composition, thermodynamics, and genes responsible for each reaction [65]. Genomics and transcriptomics form the foundation of omics research, focusing on genome sequencing and RNA transcript analysis, respectively. While genomics provides a comprehensive blueprint of an organism’s genetic architecture, it does not inherently link genotype to phenotype, highlighting the need for complementary approaches [65].
Proteomics and metabolomics fill this gap by examining proteins and metabolites, which are directly connected to biological function and phenotype. Metabolomics captures rapid responses to genetic and environmental changes, offering a snapshot of a cell’s metabolic and physiological state. However, the chemical complexity and diversity of proteins and metabolites make proteomics and metabolomics more challenging to analyze compared to genomics and transcriptomics, emphasizing the importance of integrating these datasets for a comprehensive understanding of biological systems [66].
Extracting knowledge from large and multi-dimensional omics datasets is challenging. One approach, termed “knowledge-driven” analyzes omics datasets with constraints-based mathematical models. The resulting genome-scale-models (GEM) are mathematical representations of metabolic networks suitable to produce a computational prediction of biological functions and facilitate precise manipulation of biological systems [65]
There are more than 100 species for which GEMs are public and accessible for download (e.g. www.optflux.org/models and http://systemsbiology.ucsd.edu/InSilicoOrganisms/OtherOrganisms). As being the first eukaryote to be genome sequenced, S. cerevisiae has models which define the gene–protein–reaction associations of all metabolic genes and can assist researchers in metabolic engineering. Yeast-GEMs are continually released and improved by the community [66].
In parallel, machine learning is another complementary approach to predicting the phenotypic output of genetic manipulation. As sub-category of AI, machine learning basically seeks to improve automatically through the experience (learning). Machine learning develops algorithms which statistically link an input to its associated response (e.g. protein sequence to function or tertiary structure) without developing any mechanistic knowledge about the process. Even if no biological information is encoded in the algorithm, all the biological information must be carefully chosen as training data, so the algorithm can learn the desired relationship [67].
In this regard, the increasing amount of yeast omics data sets enables in silico screening of yeast. The 1,011 yeast genomes project [6] and derivatives, which include genomic, transcriptomic, and proteomic data, can be leveraged to identify genes responsible for desired phenotypes and predict strains with the desired phenotype. This approach, also called genome-wide association study (GWAS), evaluates the association between each genotyped marker and a phenotype of interest that has been scored across a large number of individuals. Genetics associated with brewing-related phenotypes was used to successfully predict strains from no brewing environment with brewing aptitude [68]. This in silico approach offers advantages over traditional in vitro experiments, such as faster results and lower costs.The application of multi-omics approaches in industrial biotechnology is exemplified by their role in optimizing fermentation processes, such as beer production. In particular, the MAL loci are positively correlated with maltose consumption [69] [43], while the STA1 gene is associated with maltotriose and dextrin consumption [70]. Additionally, the IRA2 and INA2 genes are linked to increased ethanol tolerance [71,72]. GWAS was successful in identifying key loci and genes, including LEU1, BAT1, EHT1, and IRA1, involved in flavor modulation in S. cerevisiae. These findings have been validated through CRISPR-Cas9 overexpression and knockout experiments, highlighting the potential of multi-omics to improve product quality [18]. Furthermore, studies on non-conventional yeasts like Pichia kluyveri and Hanseniaspora have uncovered unique aroma production pathways and gene regulation mechanisms, advancing the development of specialized fermentative strains [73,74]. Recently, a combination of GEM and machine learning have been recently proposed to enable the selection of mutant strains producing high titer of tryptophan [75]. In another study, fluxomic data obtained by GEM simulation and experimental transcriptomic data were used in combination to predict cell growth of numerous S. cerevisiae strains [76]. Recently, machine learning and probabilistic modeling techniques produced effective recommendations for bioengineer yeast to produce hoppy beer [77].
Despite these advances, the integration of multi-omics data remains challenging due to inherent differences in data types. Genomic data are typically qualitative, accurate, and reproducible, while proteomic and metabolomic data are often noisy, less reproducible, and both qualitative and quantitative [75]. Addressing these challenges requires careful experimental design, precise biological sampling, and rigorous data processing before integration. Expertise in individual omics fields and robust IT support are crucial for accurate data validation and interpretation [75].
Integrating multi-omics data with artificial intelligence and machine learning holds immense potential for driving innovation across various fields. Future research should focus on refining data integration methods, expanding datasets, and improving the interpretability of machine learning models to maximize the impact of these technologies. As omics technologies continue to evolve, their ability to unravel the complexities of cellular dynamics promises transformative applications in both biomedical and industrial fields, paving the way for sustainable and efficient biotechnological solutions [76].
2.2.3. Synthetic Microbial Communities
Humans have long utilized natural microbial consortia to produce fermented products, often without a complete understanding of microbial roles. More recently, advances in metagenomics have provided evidence that genotypically diverse microbial consortia are more robust to disturbance and can display improved functioning relative to isogenic cultures. Furthermore, new functionalities can arise from polymicrobial consortia, which are absent in isogenic cultures isolated from the consortium [78]. This body of knowledge has led to the development of artificial binary co-cultures and synthetic consortia composed of multiple cultivable strains in bioproduction, where well-defined microbial groups are optimized for specific purposes [79].
In natural microbial consortia cells need to engage in synergistic and antagonistic ecological interactions with other organisms. In laboratory conditions, simplified models have been used to explore these interactions, focusing on factors such as inoculum ratios, inoculation timing, and inhibitory metabolite production [80]. Despite these efforts, the full spectrum of interactions in multi-species fermentations remains unclear. To better understand these dynamics, synthetic microbial consortia mimicking natural wine yeast communities have been proposed as model systems [80]. Methods such as Automated Ribosomal Intergenic Spacer Analysis (ARISA) have been employed to monitor population dynamics in wine fermentations, offering valuable insights into yeast interactions, though limitations remain, such as the inability to distinguish between live and dead cells [80]. In another work a fluorescence-based method was developed for real-time tracking of population dynamics, physical interactions and cell sorting within a synthetic wine yeast ecosystem [81].
In a 2015 study, Bagheri et al. [82] investigated yeast-yeast interactions during wine fermentation using a multi-species yeast consortium. The study revealed that S. cerevisiae specifically suppressed certain non-Saccharomyces species while promoting the persistence of others. ALE approach was successfully used to develop evolved consortia instead of isogenic cultures. Co-evolution of wild-type lactic acid bacteria and engineered auxotrophic S. cerevisiae in a synthetic growth medium led to bacterial isolates with enhanced secretion of two B-group vitamins [83]. The potential of co-cultures extends beyond winemaking [80]. Co-cultures offer advantages over pure cultures by reducing metabolic burdens, preventing toxic by-product accumulation, and alleviating thermodynamic constraints. This division of labor results in higher productivity and yields [84]. Table 5 shows studies have explored how microbial consortia can be harnessed to improve the sensory characteristics of both beer [84] and wine [85].
In conclusion, microbial interactions are fundamental not only to fermentation processes but also to the sensory profiles and sustainability of fermented products. Advances in synthetic microbial consortia and the strategic use of non-Saccharomyces yeasts are enhancing our ability to control fermentation dynamics, creating new opportunities to improve the quality and uniqueness of fermented beverages. These consortia have also proved valuable in addressing environmental challenges, particularly in brewery effluent management [90]. By embracing these innovations, the beer and wine industries can achieve greater efficiency, aromatic complexity, and environmental sustainability.
2.3. GMO-Based Genetic Improvement Techniques
2.3.1. Synthetic Biology and CRISPR/Cas9
In recent years, synthetic biology has emerged as a key discipline combining biological research with engineering to design biological circuits for cellular control and understand biological system design [95]. It enables researchers to create new biomolecular components, pathways, and networks to reprogram organisms as engineered cellular factories [95]. In the food industry, synthetic biology addresses challenges related to food safety, nutrition, and sustainability [95].
Interest in improving the genetic makeup of wine yeast strains grew in the 1990s with genetic engineering [96] and accelerated with CRISPR/Cas-based genome editing [97] (Figure 4). These tools allow precise genetic modifications without introducing unwanted DNA sequences, enhancing process efficiency and substrate utilization [12]. Traditional genetic methods, such as mutagenesis and hybridization, often alter unintended traits, whereas genetic engineering enables targeted improvements by integrating heterologous genes [98]. Multiplex genome editing strategies, including delta and rDNA cluster integration with CRISPR-Cas tools, allow multi-copy gene insertions, significantly advancing strain development. Genome resequencing confirmed minimal off-target effects compared to wild-type strains, highlighting the approach’s specificity.
CRISPR/Cas9 efficiency declines as sgRNA numbers increase due to competition for Cas proteins. To address this, auxiliary elements like ribozymes, tRNA, and Csy4 optimize sgRNA levels [99]. High-copy gene integrations can impose metabolic burdens, mitigated by synthetic repetitive sequences and “landing pad” DNA cassettes, such as the “wicket” cassette, which optimize gene expression [95].
For example, Di-CRISPR, combining CRISPR/Cas9 with delta integration, facilitates large gene cassette insertions by leveraging delta sequences [95]. This method successfully integrated a 2,3-butanediol biosynthesis pathway into the S. cerevisiae genome [100]. Additionally, inserting genes near autonomously replicating sequences enhances both efficiency and activity [95].
Further research is needed to optimize gene expression, minimize off-target effects, and improve scalability for industrial applications. A deeper understanding of CRISPR components’ in vivo behavior will be essential for refining large-scale applications (101). Despite challenges, CRISPR/Cas9 holds vast potential to transform industrial processes and promote sustainable production [102].
Genetically modified yeasts are commercially available in North America; however, GMOs have not achieved much commercial success in wine production, likely due to regulatory and market constraints. Two recombinant S. cerevisiae strains were developed for winemaking, one to convert malic acid into lactic acid and the other to reduce urea release during arginine metabolism [93]. Despite their potential, both strains had limited success, as the problems they addressed became less problematic due to advancements in bacterial starters and more precise fermentation control. Furthermore, the limited genetic diversity of recombinant strains contributed to their poor adoption in the wine industry.
Although the study of wine yeast genetics began later than that of beer yeasts, commercial strains have been optimized for ethanol tolerance, metabolite production, and flocculation, with recombinant DNA techniques becoming essential [87]. Table 6 provides a detailed summary of the major genetic modifications in yeast biotechnology. In brewing CRISPR-Cas9 technology has been used to switch off the FDC1 gene responsible for 4-vinul guaiacol off-flavor production in S. eubayanus (103), as well as to increase ester production by esterase encoding gene deletion [104]. Krogerus et al. used CRISPR/Cas to generate DSB in MAT locus and enhance the hybrid strain construction [105].
2.3.2. Ethical and Commercial Challenges
GM foods, a key area of debate within biotechnology, have garnered significant attention due to concerns regarding safety, ethics, and environmental impacts. Proponents of GM technology argue that the benefits of GM far outweigh the risks [115]. Opponents, however, contend that genetic modifications could lead undesirable off-target or on-target effects which can affect the nutritional quality of food, as well as introduce health risks such as antibiotic resistance and allergenicity [115].Regulations on GMOs differ from country to country, and in the European Union, there are stricter rules, partly because the EU’s GMO regulations, which date back to the early days of recombinant DNA technology, have not kept pace with current scientific knowledge. Currently, European Union (EU) regulations consider organisms modified through either gene editing or genetic engineering as GM even if they are free of any transgene [116]. The EU’s approach focuses on the process itself, classifying something as a GMO simply because it has been genetically modified, without considering the final product [117]. Based on this assumption, products classified as GMO need to seek extensive regulatory and safety assessments before their approval for commercial use by the European food safety Authority (EFSA). This precautionary approach may have serious implications on research, development, and commercialization of CRISPR technology in Europe.Today, there are several proposals to update GMO regulations. One key suggestion is to shift the focus from regulating the process (how genetic modifications are made) to regulating the final product (the traits or phenotypes resulting from those modifications), with a more flexible and scalable regulation that matches the level of genomic alteration. There’s also a growing recognition of the need for international cooperation and harmonization of regulations, especially as different regions (like the EU, USA, and Canada) have varying approaches to genetic editing [117]. In conclusion, GM foods present both opportunities and challenges. While GM technology offers significant advantages, it requires careful management to mitigate risks. Adequate field testing, long-term assessments, and regulations are essential to monitor impacts on health, the environment, and pest populations [115].
3. Conclusion
Yeasts are one of the fundamental elements in wine and beer production, as they directly influence the fermentation process, the stability of the final product, and its organoleptic qualities [118]. The introduction of genetically modified (GM) yeasts could enable the development of highly specialized strains capable of better tolerating extreme conditions, such as high temperatures or high alcohol concentrations, while also reducing the number of unwanted by-products during fermentation.
However, the widespread adoption of these technologies still faces several challenges. Ethical concerns and safety issues surrounding GM products need to be carefully addressed. Public opinion, often shaped by negative perceptions and misinformation, may be reluctant to accept GM yeasts, despite the potential scientific and practical benefits. To overcome this, it’s essential to foster clear and transparent communication, dispelling myths and fears. This will help both the public and industry professionals make informed decisions, based on reliable, evidence-backed information.
It is also important to stress that GM techniques should not be seen as a cure-all that can completely replace traditional methods of selection and improvement. In fact, the true potential lies in combining innovative approaches with established practices. This collaboration could help achieve ambitious goals without losing the richness of winemaking tradition. By integrating both GM and non-GM techniques, there’s the potential for significant benefits, such as energy savings, a reduced environmental impact, and greater biodiversity, all while enhancing quality and production efficiency.
As research continues to advance, genetically modified wine yeasts are expected to become an increasingly valuable tool within the wine and beer industry’s technological toolkit. However, to fully unlock the potential of these innovations, it is essential to implement policies that encourage further research, prioritize consumer safety, and promote positive, evidence-based communication. A balanced approach that integrates science, ethics, and industry practices will be crucial in fully realizing the benefits of this new era of genetic improvements in wine yeasts, ultimately driving sustainable progress and innovation across the entire sector
Acknowledgments
We acknowledge financial support by the European Union – NextGenerationEUGrant, CN_00000033, Project “National Biodiversity Future Center – NBFC”. CUP E93C22001090001.The PhD grant of CN is supported by PNRR DM 352/2022, Mission 4, Component 2, Investment 3.3 aimed at supporting scholarships for PNRR research doctorates.
References
- Belderok, B. Developments in bread-making processes. Plant Foods Hum. Nutr. 2000, 55, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Saerens, S.M.G.; Duong, C.T.; Nevoigt, E. Genetic improvement of brewer’s yeast: current state, perspectives and limits. Appl Microbiol Biotechnol. 2010, 86, 1195–1212. [Google Scholar] [CrossRef]
- Nielsen, J. Production of biopharmaceutical proteins by yeast: Advances through metabolic engineering. Bioengineered 2013, 4, 207–211. [Google Scholar] [CrossRef]
- Botstein, D.; Chervitz, S.A.; Cherry, M. Yeast as a Model Organism. Science 1997, 277, 1259–1260. [Google Scholar] [CrossRef] [PubMed]
- Goddard, M.R.; Greig, D. Saccharomyces cerevisiae: a nomadic yeast with no niche? FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef]
- Peter, J.; De Chiara, M.; Friedrich, A.; Yue, J.X.; Pflieger, D.; Bergström, A.; Sigwalt, A.; Barre, B.; Freel, K.; Llored, A.; Cruaud, C. Genome evolution across 1,011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, I.S.; Boeke, J.D. Yeast 2.0—connecting the dots in the construction of the world’s first functional synthetic eukaryotic genome. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef]
- Melewar, T.; Skinner, H. Territorial brand management: Beer, authenticity, and sense of place. J. Bus. Res. 2020, 116, 680–689. [Google Scholar] [CrossRef]
- Bimbo, F.; De Meo, E.; Baiano, A.; Carlucci, D. The Value of Craft Beer Styles: Evidence from the Italian Market. Foods 2023, 12, 1328. [Google Scholar] [CrossRef]
- Organization WH. Global Status Report on Alcohol and Health 2018. World Health Organization; 2019. 472 p.
- Wheat Beers Market Research Report 2023.
- Pretorius, I.S. Tailoring wine yeast for the new millennium: novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Bonatto, D. The diversity of commercially available ale and lager yeast strains and the impact of brewer’s preferential yeast choice on the fermentative beer profiles. Food Res. Int. 2021, 141, 110125. [Google Scholar] [CrossRef] [PubMed]
- Libkind, D.; Hittinger, C.T.; Valério, E.; Gonçalves, C.; Dover, J.; Johnston, M.; Gonçalves, P.; Sampaio, J.P. Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 14539–14544. [Google Scholar] [CrossRef] [PubMed]
- Gibson, B.; Dahabieh, M.; Krogerus, K.; Jouhten, P.; Magalhães, F.; Pereira, R.; Siewers, V.; Vidgren, V. Adaptive Laboratory Evolution of Ale and Lager Yeasts for Improved Brewing Efficiency and Beer Quality. Annu. Rev. Food Sci. Technol. 2020, 11, 23–44. [Google Scholar] [CrossRef] [PubMed]
- Krogerus, K.; Magalhães, F.; Vidgren, V.; Gibson, B. New lager yeast strains generated by interspecific hybridization. J. Ind. Microbiol. Biotechnol. 2015, 42, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Hutzler, M.; Morrissey, J.P.; Laus, A.; Meussdoerffer, F.; Zarnkow, M. A new hypothesis for the origin of the lager yeast Saccharomyces pastorianus. FEMS Yeast Res. 2023, 23. [Google Scholar] [CrossRef]
- Zavaleta, V.; Pérez-Través, L.; Saona, L.A.; Villarroel, C.A.; Querol, A.; Cubillos, F.A.; Gibbons, J.G. Understanding brewing trait inheritance in de novo Lager yeast hybrids. mSystems 2024, 9, e0076224. [Google Scholar] [CrossRef]
- Li, C.; Zhang, S.; Dong, G.; Bian, M.; Liu, X.; Dong, X.; Xia, T. Multi-omics study revealed the genetic basis of beer flavor quality in yeast. LWT 2022, 168. [Google Scholar] [CrossRef]
- Steen, E.J.; Chan, R.; Prasad, N.; Myers, S.; Petzold, C.J.; Redding, A.; Ouellet, M.; Keasling, J.D. Metabolic engineering of Saccharomyces cerevisiae for the production of n-butanol. Microb. Cell Factories 2008, 7, 1–8. [Google Scholar] [CrossRef]
- Saerens, S.; Swiegers, J.H. Production of low-alcohol or alcohol-free beer with. Pichia kluyveri. 2014.
- Giudici, P.; Solieri, L.; Pulvirenti, A.M.; Cassanelli, S. Strategies and perspectives for genetic improvement of wine yeasts. Appl. Microbiol. Biotechnol. 2004, 66, 622–628. [Google Scholar] [CrossRef]
- Pérez-Torrado, R.; Querol, A.; Guillamón, J.M. Genetic improvement of non-GMO wine yeasts: Strategies, advantages and safety. Trends Food Sci. Technol. 2015, 45, 1–11. [Google Scholar] [CrossRef]
- Schuller, D.; Casal, M. The use of genetically modified Saccharomyces cerevisiae strains in the wine industry. Appl. Microbiol. Biotechnol. 2005, 68, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Molinet, J.; Navarrete, J.P.; Villarroel, C.A.; Villarreal, P.; Sandoval, F.I.; Nespolo, R.F.; Stelkens, R.; Cubillos, F.A.; Sampaio, J.P. Wild Patagonian yeast improve the evolutionary potential of novel interspecific hybrid strains for lager brewing. PLOS Genet. 2024, 20, e1011154. [Google Scholar] [CrossRef] [PubMed]
- Osburn, K.; Amaral, J.; Metcalf, S.R.; Nickens, D.M.; Rogers, C.M.; Sausen, C.; Caputo, R.; Miller, J.; Li, H.; Tennessen, J.M.; Bochman, M.L. Primary souring: a novel bacteria-free method for sour beer production. Food Microbiology 2018, 70, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; House, J.F.; Joseph, C.M.; Bisson, L.F.; Bamforth, C.W. Lachancea thermotolerans as an alternative yeast for the production of beer. Journal of the Institute of Brewing 2016, 122, 599–604. [Google Scholar] [CrossRef]
- Varela, C.; Borneman, A.R. Yeasts found in vineyards and wineries. Yeast 2016, 34, 111–128. [Google Scholar] [CrossRef]
- Molinet, J.; Navarrete, J.P.; Villarroel, C.A.; Villarreal, P.; Sandoval, F.I.; Nespolo, R.F.; Stelkens, R.; Cubillos, F.A.; Sampaio, J.P. Wild Patagonian yeast improve the evolutionary potential of novel interspecific hybrid strains for lager brewing. PLOS Genet. 2024, 20, e1011154. [Google Scholar] [CrossRef]
- Akada, R.; Hirosawa, I.; Kawahata, M.; Hoshida, H.; Nishizawa, Y. Sets of integrating plasmids and gene disruption cassettes containing improved counter-selection markers designed for repeated use in budding yeast. Yeast 2002, 19, 393–402. [Google Scholar] [CrossRef]
- de Vries, A.R.G.; Knibbe, E.; van Roosmalen, R.; Broek, M.v.D.; Cortés, P.d.l.T.; O’herne, S.F.; Vijverberg, P.A.; el Masoudi, A.; Brouwers, N.; Pronk, J.T.; et al. Improving Industrially Relevant Phenotypic Traits by Engineering Chromosome Copy Number in Saccharomyces pastorianus. Front. Genet. 2020, 11, 518. [Google Scholar] [CrossRef]
- Diderich, J.A.; Weening, S.M.; Broek, M.v.D.; Pronk, J.T.; Daran, J.-M.G. Selection of Pof-Saccharomyces eubayanus Variants for the Construction of S. cerevisiae × S. eubayanus Hybrids With Reduced 4-Vinyl Guaiacol Formation. Front. Microbiol. 2018, 9, 1640. [Google Scholar] [CrossRef]
- Salmon, J.-M.; Barre, P. Improvement of Nitrogen Assimilation and Fermentation Kinetics under Enological Conditions by Derepression of Alternative Nitrogen-Assimilatory Pathways in an Industrial Saccharomyces cerevisiae Strain. Appl. Environ. Microbiol. 1998, 64, 3831–3837. [Google Scholar] [CrossRef]
- Quirós, M.; Gonzalez-Ramos, D.; Tabera, L.; Gonzalez, R. A new methodology to obtain wine yeast strains overproducing mannoproteins. Int. J. Food Microbiol. 2010, 139, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Cordente, A.G.; Cordero-Bueso, G.; Pretorius, I.S.; Curtin, C.D. Novel wine yeast with mutations in YAP1 that produce less acetic acid during fermentation. FEMS Yeast Res. 2013, 13, 62–73. [Google Scholar] [CrossRef]
- Wöhrmann, K.; Lance, P. THE POLYMORPHISM OF ESTERASES IN YEAST (SACCHAROMYCES CEREVISIAE). J. Inst. Brew. 1980, 86, 174. [Google Scholar] [CrossRef]
- Giudici, P.; Zinnato, A. Influenza dell’uso di mutanti nutrizionali sulla produzione di alcooli superiori. Vignevini 1983, 10. [Google Scholar]
- Rous, C.V.; Snow, R.; Kunkee, R.E. REDUCTION OF HIGHER ALCOHOLS BY FERMENTATION WITH A LEUCINE-AUXOTROPHIC MUTANT OF WINE YEAST. J. Inst. Brew. 1983, 89, 274–278. [Google Scholar] [CrossRef]
- Krogerus, K.; Arvas, M.; De Chiara, M.; Magalhães, F.; Mattinen, L.; Oja, M.; Vidgren, V.; Yue, J.-X.; Liti, G.; Gibson, B. Ploidy influences the functional attributes of de novo lager yeast hybrids. Appl. Microbiol. Biotechnol. 2016, 100, 7203–7222. [Google Scholar] [CrossRef] [PubMed]
- Gyurchev, N.Y.; Coral-Medina, Á.; Weening, S.M.; Almayouf, S.; Kuijpers, N.G.A.; Nevoigt, E.; Louis, E.J. Beyond Saccharomyces pastorianus for modern lager brews: Exploring non-cerevisiae Saccharomyces hybrids with heterotic maltotriose consumption and novel aroma profile. Front. Microbiol. 2022, 13, 1025132. [Google Scholar] [CrossRef]
- Eschenbruch, R.; Cresswell, K.J.; Fisher, B.M.; Thornton, R.J. Selective hybridisation of pure culture wine yeasts. Eur. Appl. Microbiol. Biotechnol. 1982, 14, 155–158. [Google Scholar] [CrossRef]
- Shinohara, T.; Mamiya, S.; Yanagida, F. Introduction of flocculation property into wine yeasts (Saccharomyces cerevisiae) by hybridization. J. Ferment. Bioeng. 1997, 83, 96–101. [Google Scholar] [CrossRef]
- Nikulin, J.; Krogerus, K.; Gibson, B. Alternative Saccharomyces interspecies hybrid combinations and their potential for low-temperature wort fermentation. Yeast 2017, 35, 113–127. [Google Scholar] [CrossRef]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410.e16. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.; Pontes, A.; Almeida, P.; Barbosa, R.; Serra, M.; Libkind, D.; Hutzler, M.; Gonçalves, P.; Sampaio, J.P. Distinct Domestication Trajectories in Top-Fermenting Beer Yeasts and Wine Yeasts. Curr. Biol. 2016, 26, 2750–2761. [Google Scholar] [CrossRef]
- Whittington, H.D.; Dagher, S.F.; Bruno-Bárcena, J.M. Production and conservation of starter cultures: From “backslopping” to controlled fermentations. How fermented foods feed a healthy gut microbiota: A nutrition continuum. 2019:125-38. [CrossRef]
- Beato, F.B.; Bergdahl, B.; Rosa, C.A.; Forster, J.; Gombert, A.K.; Kang, H. Physiology of Saccharomyces cerevisiae strains isolated from Brazilian biomes: new insights into biodiversity and industrial applications. FEMS Yeast Res. 2016, 16, fow076. [Google Scholar] [CrossRef]
- Sharp, N.P.; Sandell, L.; James, C.G.; Otto, S.P. The genome-wide rate and spectrum of spontaneous mutations differ between haploid and diploid yeast. Proc. Natl. Acad. Sci. 2018, 115, E5046–E5055. [Google Scholar] [CrossRef]
- Lee, S.; Kim, P. Current Status and Applications of Adaptive Laboratory Evolution in Industrial Microorganisms. J. Microbiol. Biotechnol. 2020, 30, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Cauet, G.; Degryse, E.; Ledoux, C.; Spagnoli, R.; Achstetter, T. Pregnenolone esterification in Saccharomyces cerevisiae. Eur. J. Biochem. 1999, 261, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Mavrommati, M.; Daskalaki, A.; Papanikolaou, S.; Aggelis, G. Adaptive laboratory evolution principles and applications in industrial biotechnology. Biotechnol. Adv. 2022, 54, 107795. [Google Scholar] [CrossRef]
- Mattenberger, F.; Fares, M.A.; Toft, C.; Sabater-Muñoz, B. The Role of Ancestral Duplicated Genes in Adaptation to Growth on Lactate, a Non-Fermentable Carbon Source for the Yeast Saccharomyces cerevisiae. Int. J. Mol. Sci. 2021, 22, 12293. [Google Scholar] [CrossRef]
- Gonzalez, R.; Tronchoni, J.; Quirós, M.; Morales, P. Genetic improvement and genetically modified microorganisms. Wine Safety, Consumer Preference, and Human Health. 2016:71-96. [CrossRef]
- Blieck, L.; Toye, G.; Dumortier, F.; Verstrepen, K.J.; Delvaux, F.R.; Thevelein, J.M.; Van Dijck, P. Isolation and Characterization of Brewer’s Yeast Variants with Improved Fermentation Performance under High-Gravity Conditions. Appl. Environ. Microbiol. 2007, 73, 815–824. [Google Scholar] [CrossRef]
- Krogerus, K.; Holmström, S.; Gibson, B.; Schaffner, D.W. Enhanced Wort Fermentation with De Novo Lager Hybrids Adapted to High-Ethanol Environments. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef]
- Avrahami-Moyal, L.; Engelberg, D.; Wenger, J.W.; Sherlock, G.; Braun, S. Turbidostat culture of Saccharomyces cerevisiae W303-1A under selective pressure elicited by ethanol selects for mutations in SSD1 and UTH1. FEMS Yeast Res. 2012, 12, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.K.; Tremaine, M.; Parreiras, L.S.; Hebert, A.S.; Myers, K.S.; Higbee, A.J.; et al. Directed Evolution Reveals Unexpected Epistatic Interactions That Alter Metabolic Regulation and Enable Anaerobic Xylose Use by Saccharomyces cerevisiae. PLOS Genetics 2016, 12, e1006372. [Google Scholar]
- Castro, M.C. Improvement of Torulaspora delbrueckii’s sulphite resistance in winemaking through Adaptive Laboratory Evolution (Doctoral dissertation).
- Bartel, C.; Roach, M.; Onetto, C.; Curtin, C.; Varela, C.; Borneman, A. Adaptive evolution of sulfite tolerance in Brettanomyces bruxellensis. FEMS Yeast Res. 2021, 21. [Google Scholar] [CrossRef]
- He, Y.; Yin, H.; Dong, J.; Yu, J.; Zhang, L.; Yan, P.; Wan, X.; Hou, X.; Zhao, Y.; Chen, R.; et al. Reduced sensitivity of lager brewing yeast to premature yeast flocculation via adaptive evolution. Food Microbiol. 2022, 106, 104032. [Google Scholar] [CrossRef] [PubMed]
- Kutyna, D.R.; Varela, C.; Stanley, G.A.; Borneman, A.R.; Henschke, P.A.; Chambers, P.J. Adaptive evolution of Saccharomyces cerevisiae to generate strains with enhanced glycerol production. Appl. Microbiol. Biotechnol. 2011, 93, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Mezzetti, F.; De Vero, L.; Giudici, P. Evolved Saccharomyces cerevisiae wine strains with enhanced glutathione production obtained by an evolution-based strategy. FEMS Yeast Res. 2014, 14, 977–987. [Google Scholar] [CrossRef]
- Mohammadzadeh, A.; Kayacan, E. A novel fractional-order type-2 fuzzy control method for online frequency regulation in ac microgrid. Eng. Appl. Artif. Intell. 2020, 90. [Google Scholar] [CrossRef]
- Fernandes, T.; Osório, C.; Sousa, M.J.; Franco-Duarte, R. Contributions of Adaptive Laboratory Evolution towards the Enhancement of the Biotechnological Potential of Non-Conventional Yeast Species. J. Fungi 2023, 9, 186. [Google Scholar] [CrossRef]
- Baidoo, E.E.; Teixeira Benites, V. Mass spectrometry-based microbial metabolomics: Techniques, analysis, and applications. Microbial metabolomics: methods and protocols 2019, 11–69. [Google Scholar]
- Amer, B.; Baidoo, E.E.K. Omics-Driven Biotechnology for Industrial Applications. Front. Bioeng. Biotechnol. 2021, 9. [Google Scholar] [CrossRef]
- Zhang, C.; Sánchez, B.J.; Li, F.; Eiden, C.W.Q.; Scott, W.T.; Liebal, U.W.; Blank, L.M.; Mengers, H.G.; Anton, M.; Rangel, A.T.; et al. Yeast9: a consensus genome-scale metabolic model for S. cerevisiae curated by the community. Mol. Syst. Biol. 2024, 20, 1134–1150. [Google Scholar] [CrossRef]
- Krogerus, K.; Rettberg, N. Creating Better Brewing Yeast With the 1011 Yeast Genomes Data Sets. Yeast 2025, 42, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Duval, E.H.; Alves Jr, S.L.; Dunn, B.; Sherlock, G.; Stambuk, B.U. Microarray karyotyping of maltose-fermenting Saccharomyces yeasts with differing maltotriose utilization profiles reveals copy number variation in genes involved in maltose and maltotriose utilization. Journal of Applied Microbiology 2010, 109, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Krogerus, K.; Magalhães, F.; Kuivanen, J.; Gibson, B. A deletion in the STA1 promoter determines maltotriose and starch utilization in STA1+ Saccharomyces cerevisiae strains. Appl. Microbiol. Biotechnol. 2019, 103, 7597–7615. [Google Scholar] [CrossRef]
- Tristão, L.E.; de Sousa, L.I.S.; Vargas, B.d.O.; José, J.; Carazzolle, M.F.; Silva, E.M.; Galhardo, J.P.; Pereira, G.A.G.; Mello, F.d.S.B.d. Unveiling genetic anchors in Saccharomyces cerevisiae: QTL mapping identifies IRA2 as a key player in ethanol tolerance and beyond. Mol. Genet. Genom. 2024, 299, 1–13. [Google Scholar] [CrossRef]
- Albillos-Arenal, S.; Alonso-del-Real, J.; Lairón-Peris, M.; Barrio, E.; Querol, A. Identification of a crucial INO2 allele for enhancing ethanol resistance in an industrial fermentation strain of Saccharomyces cerevisiae. bioRxiv 2024. [Google Scholar]
- Wei, J.; Zhang, Y.; Zhang, X.; Guo, H.; Yuan, Y.; Yue, T. Multi-omics discovery of aroma-active compound formation by Pichia kluyveri during cider production. LWT 2022, 159. [Google Scholar] [CrossRef]
- van Wyk, N.; Badura, J.; von Wallbrunn, C.; Pretorius, I.S. Exploring future applications of the apiculate yeast Hanseniaspora. Crit. Rev. Biotechnol. 2023, 44, 100–119. [Google Scholar] [CrossRef]
- Zhang, J.; Petersen, S.D.; Radivojevic, T.; Ramirez, A.; Pérez-Manríquez, A.; Abeliuk, E.; Sánchez, B.J.; Costello, Z.; Chen, Y.; Fero, M.J.; et al. Combining mechanistic and machine learning models for predictive engineering and optimization of tryptophan metabolism. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Liu, X.; Peng, T.; Xu, M.; Lin, S.; Hu, B.; Chu, T.; Liu, B.; Xu, Y.; Ding, W.; Li, L.; et al. Spatial multi-omics: deciphering technological landscape of integration of multi-omics and its applications. J. Hematol. Oncol. 2024, 17, 1–24. [Google Scholar] [CrossRef]
- Radivojević, T.; Costello, Z.; Workman, K.; Martin, H.G. A machine learning Automated Recommendation Tool for synthetic biology. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.; Shitut, S.; Kost, C. Harnessing ecological and evolutionary principles to guide the design of microbial production consortia. Curr. Opin. Biotechnol. 2020, 62, 228–238. [Google Scholar] [CrossRef]
- Mittermeier, F.; Bäumler, M.; Arulrajah, P.; Lima, J.d.J.G.; Hauke, S.; Stock, A.; Weuster-Botz, D. Artificial microbial consortia for bioproduction processes. Eng. Life Sci. 2022, 23, e2100152. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, B.; Bauer, F.F.; Setati, M.E. The Impact of Saccharomyces cerevisiae on a Wine Yeast Consortium in Natural and Inoculated Fermentations. Front. Microbiol. 2017, 8, 1988. [Google Scholar] [CrossRef] [PubMed]
- Conacher, C.G.; Naidoo-Blassoples, R.K.; Rossouw, D.; Bauer, F.F. Real-time monitoring of population dynamics and physical interactions in a synthetic yeast ecosystem by use of multicolour flow cytometry. Appl. Microbiol. Biotechnol. 2020, 104, 5547–5562. [Google Scholar] [CrossRef]
- Bagheri, B.; Bauer, F.; Setati, M. The Diversity and Dynamics of Indigenous Yeast Communities in Grape Must from Vineyards Employing Different Agronomic Practices and their Influence on Wine Fermentation. South Afr. J. Enol. Vitic. 2016, 36, 243–251. [Google Scholar] [CrossRef]
- Konstantinidis, D.; Pereira, F.; Geissen, E.M.; Grkovska, K.; Kafkia, E.; Jouhten, P.; Kim, Y.; Devendran, S.; Zimmermann, M.; Patil, K.R. Adaptive laboratory evolution of microbial co-cultures for improved metabolite secretion. Molecular systems biology 2021, 17, e10189. [Google Scholar] [CrossRef]
- Roell, M.-S.; Zurbriggen, M.D. The impact of synthetic biology for future agriculture and nutrition. Curr. Opin. Biotechnol. 2020, 61, 102–109. [Google Scholar] [CrossRef]
- Griggs, R.G.; Steenwerth, K.L.; Mills, D.A.; Cantu, D.; Bokulich, N.A. Sources and assembly of microbial communities in vineyards as a functional component of winegrowing. Frontiers in Microbiology 2021, 12, 673810. [Google Scholar] [CrossRef]
- Motlhanka, K.; Lebani, K.; Boekhout, T.; Zhou, N. Fermentative Microbes of Khadi, a Traditional Alcoholic Beverage of Botswana. Fermentation 2020, 6, 51. [Google Scholar] [CrossRef]
- Vaughan, A.; O’SUllivan, T.; Sinderen, D. Enhancing the Microbiological Stability of Malt and Beer - A Review. J. Inst. Brew. 2005, 111, 355–371. [Google Scholar] [CrossRef]
- Mas, A.; Portillo, M.C. Strategies for microbiological control of the alcoholic fermentation in wines by exploiting the microbial terroir complexity: A mini-review. Int. J. Food Microbiol. 2022, 367, 109592. [Google Scholar] [CrossRef]
- Jolly, N.; Augustyn, O.; Pretorius, I. The Occurrence of Non-Saccharomyces cerevisiae Yeast Species Over Three Vintages in Four Vineyards and Grape Musts From Four Production Regions of the Western Cape, South Africa. South Afr. J. Enol. Vitic. 2003, 24, 35–42. [Google Scholar] [CrossRef]
- Sainz, J.; Pizarro, F.; Pérez-Correa, J.R.; Agosin, E. Modeling of yeast metabolism and process dynamics in batch fermentation. Biotechnol. Bioeng. 2003, 81, 818–828. [Google Scholar] [CrossRef]
- Mendoza, L.M.; de Nadra, M.C.M.; Bru, E.; Farías, M.E. Influence of wine-related physicochemical factors on the growth and metabolism of non-Saccharomyces and Saccharomyces yeasts in mixed culture. J. Ind. Microbiol. Biotechnol. 2008, 36, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Esteve-Zarzoso, B.; Cocolin, L.; Mas, A.; Rantsiou, K. Viable and culturable populations of Saccharomyces cerevisiae, Hanseniaspora uvarum and Starmerella bacillaris (synonym Candida zemplinina) during Barbera must fermentation. Food Research International 2015, 78, 195–200. [Google Scholar] [CrossRef]
- Albergaria, H.; Arneborg, N. Dominance of Saccharomyces cerevisiae in alcoholic fermentation processes: role of physiological fitness and microbial interactions. Appl. Microbiol. Biotechnol. 2016, 100, 2035–2046. [Google Scholar] [CrossRef] [PubMed]
- 94. Cambon, B.; Monteil, V.; Remize, F.; Camarasa, C.; Dequin, S. Effects of GPD1 Overexpression in Saccharomyces cerevisiae Commercial Wine Yeast Strains Lacking ALD6 Genes. Applied and Environmental Microbiology 2006, 72, 4688–4694. [Google Scholar] [CrossRef]
- Malcı, K.; Walls, L.E.; Rios-Solis, L. Multiplex Genome Engineering Methods for Yeast Cell Factory Development. Front. Bioeng. Biotechnol. 2020, 8, 589468. [Google Scholar] [CrossRef]
- Cebollero, E.; Gonzalez-Ramos, D.; Tabera, L.; Gonzalez, R. Transgenic wine yeast technology comes of age: is it time for transgenic wine? Biotechnol. Lett. 2006, 29, 191–200. [Google Scholar] [CrossRef]
- DiCarlo, J.E.; Norville, J.E.; Mali, P.; Rios, X.; Aach, J.; Church, G.M. Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucleic Acids Res. 2013, 41, 4336–4343. [Google Scholar] [CrossRef]
- Bidenne, C.; Blondin, B.; Dequin, S.; Vezinhet, F. Analysis of the chromosomal DNA polymorphism of wine strains of Saccharomyces cerevisiae. Curr. Genet. 1992, 22, 1–7. [Google Scholar] [CrossRef]
- Ren, Q.; Zhong, Z.; Wang, Y.; You, Q.; Li, Q.; Yuan, M.; He, Y.; Qi, C.; Tang, X.; Zheng, X.; et al. Bidirectional Promoter-Based CRISPR-Cas9 Systems for Plant Genome Editing. Front. Plant Sci. 2019, 10, 1173. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Geng, A. High-copy genome integration of 2,3-butanediol biosynthesis pathway in Saccharomyces cerevisiae via in vivo DNA assembly and replicative CRISPR-Cas9 mediated delta integration. J. Biotechnol. 2020, 310, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Shen, J.; Li, D.; Cheng, Y. Strategies in the delivery of Cas9 ribonucleoprotein for CRISPR/Cas9 genome editing. Theranostics 2021, 11, 614–648. [Google Scholar] [CrossRef] [PubMed]
- Dixit, Y.; Yadav, P.; Sharma, A.K.; Pandey, P.; Kuila, A. Multiplex genome editing to construct cellulase engineered Saccharomyces cerevisiae for ethanol production from cellulosic biomass. Renew. Sustain. Energy Rev. 2023, 187. [Google Scholar] [CrossRef]
- Mertens, S.; Gallone, B.; Steensels, J.; Herrera-Malaver, B.; Cortebeek, J.; Nolmans, R.; et al. Reducing phenolic off-flavors through CRISPR-based gene editing of the FDC1 gene in Saccharomyces cerevisiae x Saccharomyces eubayanus hybrid lager beer yeasts. PLOS ONE 2019, 14, e0209124. [Google Scholar]
- Dank, A.; Smid, E.J.; Notebaart, R.A. CRISPR-Cas genome engineering of esterase activity in Saccharomyces cerevisiae steers aroma formation. BMC Res. Notes 2018, 11, 1–6. [Google Scholar] [CrossRef]
- Krogerus, K.; Fletcher, E.; Rettberg, N.; Gibson, B.; Preiss, R. Efficient breeding of industrial brewing yeast strains using CRISPR/Cas9-aided mating-type switching. Appl. Microbiol. Biotechnol. 2021, 105, 8359–8376. [Google Scholar] [CrossRef]
- González, J.P.; González, R.; Querol, A.; Sendra, J.; Ramón, D. Construction of a recombinant wine yeast strain expressing β-(1, 4)-endoglucanase and its use in microvinification process. Applied and Environmental Microbiology 1993, 59, 2801–2806. [Google Scholar] [CrossRef]
- Vigentini, I.; Gebbia, M.; Belotti, A.; Foschino, R.; Roth, F.P. The CRISPR/Cas9 system as a molecular strategy to decrease urea production in wine yeasts (2017).
- Hirata, D.; Aoki, S.; Watanabe, K.-I.; Tsukioka, M.; Suzuki, T. Stable Overproduction of Isoamyl Alcohol by Saccharomyces cerevisiae with Chromosome-integrated Multicopy LEU4 Genes. Biosci. Biotechnol. Biochem. 1992, 56, 1682–1683. [Google Scholar] [CrossRef]
- Kitamoto, K.; Oda, K.; Gomi, K.; Takahashi, K. Genetic engineering of a sake yeast producing no urea by successive disruption of arginase gene. Appl. Environ. Microbiol. 1991, 57, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Schoeman, H.; Vivier, M.A.; du Toit, M.; Dicks, L.M.T.; Pretorius, I.S. The development of bactericidal yeast strains by expressing thePediococcus acidilactici pediocin gene (pedA) inSaccharomyces cerevisiae. Yeast 1999, 15, 647–656. [Google Scholar] [CrossRef]
- Sone, H.; Fujii, T.; Kondo, K.; Shimizu, F.; Tanaka, J.; Inoue, T. Nucleotide sequence and expression of the Enterobacter aerogenes alpha-acetolactate decarboxylase gene in brewer’s yeast. Appl. Environ. Microbiol. 1988, 54, 38–42. [Google Scholar] [CrossRef]
- Volschenk, H.; Viljoen, M.; Grobler, J.; Bauer, F.; Lonvaud-Funel, A.; Denayrolles, M.; Subden, R.E.; Van Vuuren, H.J.J. Malolactic Fermentation in Grape Musts by a Genetically Engineered Strain of Saccharomyces cerevisiae. Am. J. Enol. Vitic. 1997, 48, 193–197. [Google Scholar] [CrossRef]
- Denby, C.M.; Li, R.A.; Vu, V.T.; Costello, Z.; Lin, W.; Chan, L.J.G.; Williams, J.; Donaldson, B.; Bamforth, C.W.; Petzold, C.J.; et al. Industrial brewing yeast engineered for the production of primary flavor determinants in hopped beer. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- de Ruijter, J.C.; Aisala, H.; Jokinen, I.; Krogerus, K.; Rischer, H.; Toivari, M. Production and sensory analysis of grape flavoured beer by co-fermentation of an industrial and a genetically modified laboratory yeast strain. Eur. Food Res. Technol. 2023, 249, 1991–2000. [Google Scholar] [CrossRef]
- Uzogara, S.G. The impact of genetic modification of human foods in the 21st century. Biotechnol. Adv. 2000, 18, 179–206. [Google Scholar] [CrossRef]
- Directive 2001/18/EC of the European Parliament and of the Council of 12 March 2001 on the deliberate release into the environment of genetically modified organisms and repealing Council Directive 90/220/EEC - Commission Declaration.
- Davison, J.; Ammann, K. New GMO regulations for old: Determining a new future for EU crop biotechnology. GM Crop. Food 2017, 8, 13–34. [Google Scholar] [CrossRef]
- Nasuti, C.; Solieri, L. Yeast Bioflavoring in Beer: Complexity Decoded and Built up Again. Fermentation 2024, 10, 183. [Google Scholar] [CrossRef]
Figure 2.
Schematic representation of the mutagenesis process. The process begins with induced mutagenesis, either through UV irradiation or exposure to chemical mutagenic agents. The mutagenized culture is then plated on a selective medium, making the presence of selection markers or easily distinguishable phenotypic traits crucial. Based on the desired phenotype, mutant clones can be identified and selected. If the desired phenotype is negative, replica plating is required.
Figure 2.
Schematic representation of the mutagenesis process. The process begins with induced mutagenesis, either through UV irradiation or exposure to chemical mutagenic agents. The mutagenized culture is then plated on a selective medium, making the presence of selection markers or easily distinguishable phenotypic traits crucial. Based on the desired phenotype, mutant clones can be identified and selected. If the desired phenotype is negative, replica plating is required.
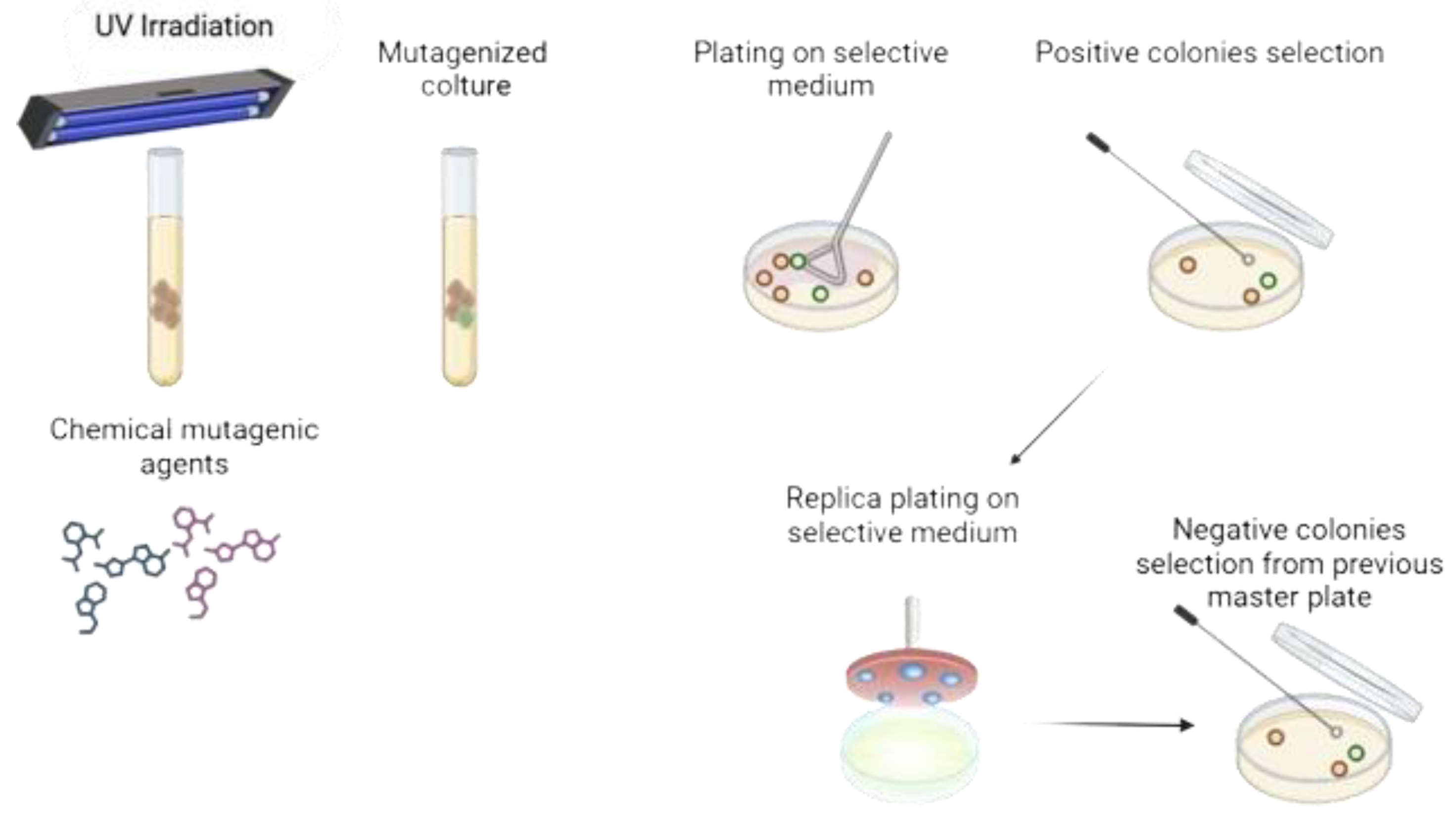
Figure 2.
Graphical representation of spore-to-spore mating hybridization. Two meiosis-competent cells undergo sporulation under stress conditions, such as carbon and nitrogen starvation. The ascus containing the haploid gametes as spores is then broken to obtain monosporic cultures. The monosporic cells from the parental strains are subsequently placed next to each other to induce mating, leveraging the opposite mating types. As a result, a diploid (2n) MATa/MATα daughter cell is formed.
Figure 2.
Graphical representation of spore-to-spore mating hybridization. Two meiosis-competent cells undergo sporulation under stress conditions, such as carbon and nitrogen starvation. The ascus containing the haploid gametes as spores is then broken to obtain monosporic cultures. The monosporic cells from the parental strains are subsequently placed next to each other to induce mating, leveraging the opposite mating types. As a result, a diploid (2n) MATa/MATα daughter cell is formed.
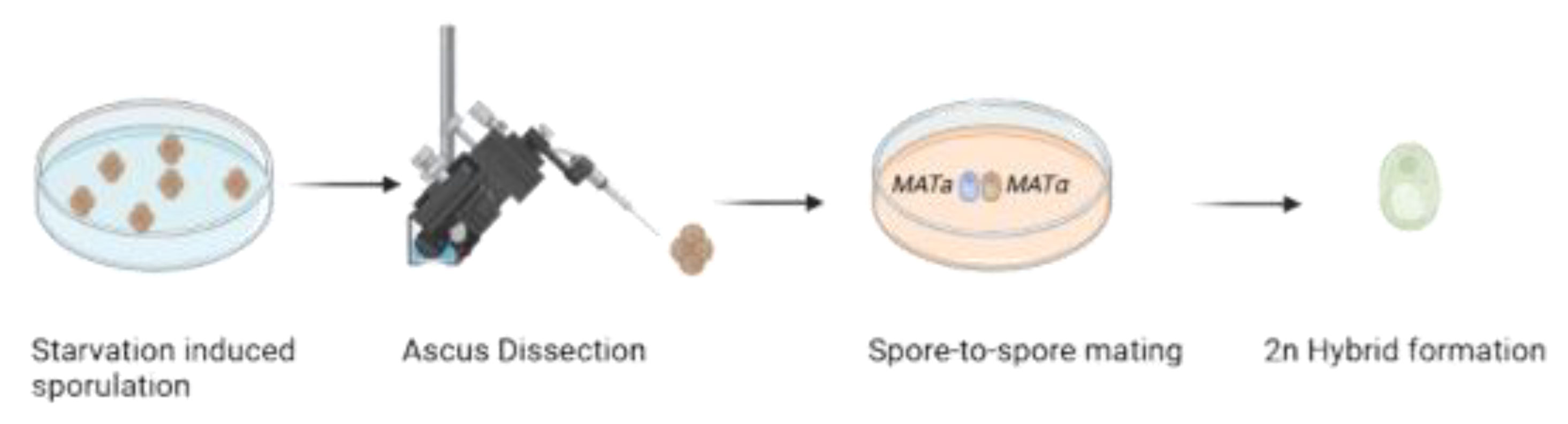
Figure 3.
Graphical representation of Adaptive Laboratory Evolution (ALE). The ancestral strain is exposed to a selective pressure environment, either constant or gradually increasing over time. This process allows for the selection of evolved populations with phenotypic traits that enhance survival under selective pressure. After a certain number of generations, evolved clones can be isolated to identify beneficial mutations responsible for the adaptive phenotype.
Figure 3.
Graphical representation of Adaptive Laboratory Evolution (ALE). The ancestral strain is exposed to a selective pressure environment, either constant or gradually increasing over time. This process allows for the selection of evolved populations with phenotypic traits that enhance survival under selective pressure. After a certain number of generations, evolved clones can be isolated to identify beneficial mutations responsible for the adaptive phenotype.
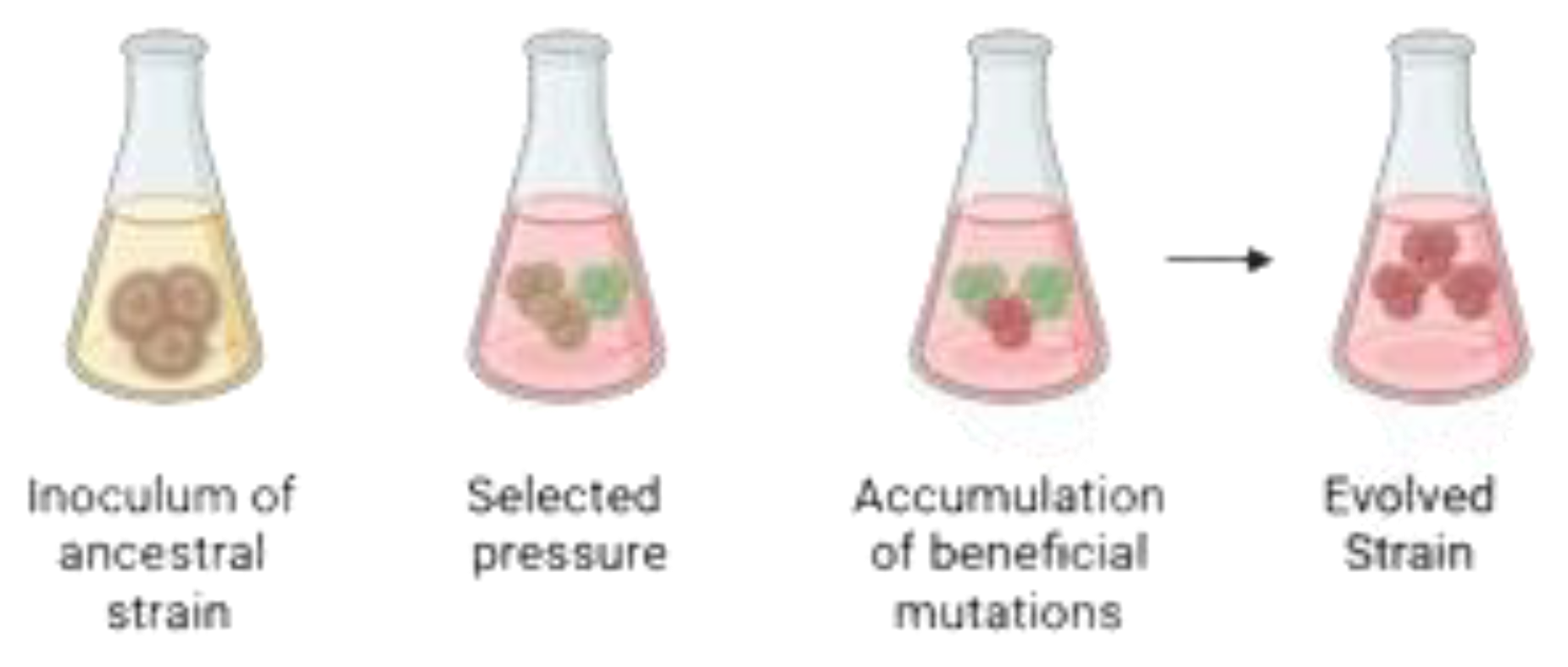
Figure 4.
Schematic representation of the CRISPR-Cas9 genome editing process. The process begins with the design of a guide RNA (gRNA) complementary to the target DNA sequence. The gRNA binds to the Cas9 protein, forming the active CRISPR-Cas9 complex. This complex recognizes and binds to the target DNA site, where Cas9 induces a double-strand break (DSB). The DNA break is then repaired through either Non-Homologous End Joining (NHEJ), which can introduce mutations or result in gene knockout, or Homology-Directed Repair (HDR), which enables precise gene editing if a repair template is provided.
Figure 4.
Schematic representation of the CRISPR-Cas9 genome editing process. The process begins with the design of a guide RNA (gRNA) complementary to the target DNA sequence. The gRNA binds to the Cas9 protein, forming the active CRISPR-Cas9 complex. This complex recognizes and binds to the target DNA site, where Cas9 induces a double-strand break (DSB). The DNA break is then repaired through either Non-Homologous End Joining (NHEJ), which can introduce mutations or result in gene knockout, or Homology-Directed Repair (HDR), which enables precise gene editing if a repair template is provided.
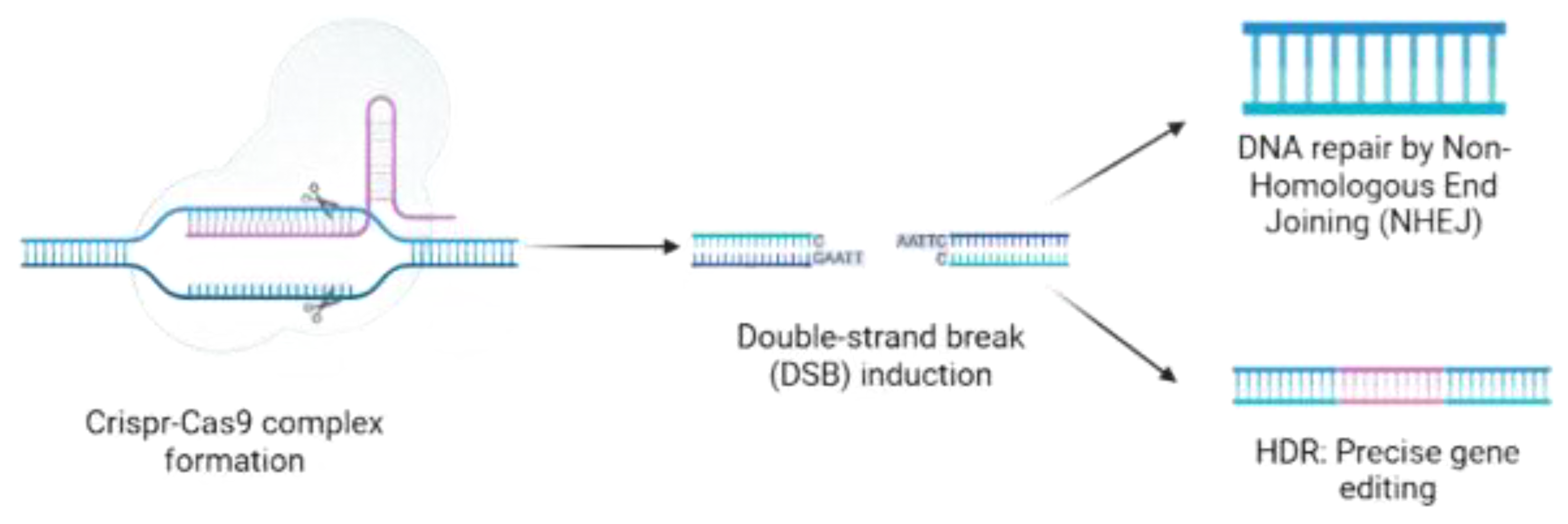

Table 1.
Overview of different genetic techniques for yeasts improvement.
| Technique | Pros | Cons |
|---|---|---|
|
CRISPR-Cas9 GMO |
Precise and efficient genome editing; Allows for modifications of multiple genes at once; highly versatile across different yeast strains. |
Ethical concerns around gene editing; Requires optimization for different yeast species; Requires advanced understanding of yeast genetics |
|
Synthetic Microbial Communities (SMC) GMO or Non-GMO (depending on strains used) |
Enables creation of yeast strains consortia with complementary traits that work together; Can improve metabolic networks. |
Requires compatibility between strains, including nutrient requirements; If OGM strains are used, there are ethical concerns around genetic modifications. |
|
Hybridization Non-GMO |
Simple method to combine beneficial traits from different yeast strains; Heterosis compared with parents; Well-established and cost-effective. |
May result in sterility or instability of the hybrid offspring; Difficult to obtain for poorly sporulating strains; undesirable characteristics may emerge. |
|
Mutagenesis Non-GMO |
Generates a wide variety of potential phenotypes; Relatively inexpensive and straightforward; Suitable for monogenic phenotypes. |
Random outcomes make it difficult to predict results; Can introduce harmful mutations or undesired traits; Not suitable for polygenic or complex phenotypes. |
|
Adaptive Laboratory Evolution (ALE) Non-GMO |
Mimics natural selection, leading to improved fitness-related traits over time; Suitable for phenotypes that can be directly selected under controlled environments. |
Time-consuming and labor-intensive; Unintended side effects may occur, as it is difficult to control specific outcomes; May not be suitable for traits without clear selection markers. |
|
Multi-Omics and AI Integration Non-GMO |
Provides a comprehensive view of yeast metabolism and gene expression; Unintended side effects may occur, as it is difficult to control specific outcomes. |
Requires large data sets and significant computational resources; Interpreting the data can be complex and requires expert knowledge. |
Table 2.
Summary of yeast strain improvements through mutagenesis.
| Objective | Work |
|---|---|
| Increased ethanol tolerance | [31] |
| Elimination of phenolic off-flavors (POF) | [32] |
| Enhanced nitrogen source utilization | [33] |
| Increased mannoprotein release | [34] |
| Reduced volatile acidity | [35] |
| Increased aroma compound concentration (esters) | [36] |
| Reduction of isoamyl alcohol (3-methylbutanol) production | [37] |
Table 3.
Summary of yeast strain improvements through hybridization.
| Objective | Work |
|---|---|
| Improved fermentation and aroma production in lager hybrids | [19,30,39,40] |
| Elimination of undesirable traits (e.g., SO₂ formation, excessive foam production) | [41] |
| Enhanced wine quality and fermentation rates via interspecific hybridization | [23,42] |
Table 4.
Summary of yeast strain improvements through adaptive laboratory evolution (ALE).
| Objective | Work |
|---|---|
| Increased tolerance to acetic acid | [53] |
| Selection of Atf2-overexpressing strains for ester formation and pregnenolone detoxification | [50] |
| Improved beer yeast performance via UV mutagenesis and high-gravity wort fermentations | [54] |
| Enhanced ethanol tolerance in hybrid yeasts | [55] |
| Genomic adaptations linked to chromosomal duplications and mutations in IRA2 and UTH1 | [56,57] |
| Increased sulfite tolerance in T. delbrueckii | [58] |
| Enhanced sulfite resistance in B. bruxellensis | [59] |
| Adaptation of Saccharomyces variants to overcome premature yeast flocculation (PYF) | [60] |
| Reduction of phenolic off-flavors via PAD1 and FDC1 mutations | [44,45] |
| Increased glycerol production for lower ethanol wines | [61] |
| Development of yeast strains producing higher levels of glutathione (GSH) | [62] |
| Enhanced yeast flocculation for easier removal after fermentation | [63] |
Table 5.
Summary of microbial interactions in beer and wine fermentation and bioprocesses.
| Application | Involved Species | Work |
|---|---|---|
| Alcoholic fermentation in beer production | Saccharomyces cerevisiae | [86] |
| Bio-acidification and microbial control in beer | Lactic acid bacteria (LAB) | [87] |
| Malolactic fermentation for flavor complexity in wine | Oenococcus oeni | [86] |
| Adaptation to vineyard microbial terroir | Various LAB and yeast species | [88] |
| Yeast community shifts during grape must fermentation | K. apiculate, C. stellata, C. pulcherrima | [89] |
| Influence of nutrient scarcity, oxygen availability, and ethanol on fermentation | Saccharomyces cerevisiae and other yeasts | [90,91] |
| Ecological interactions driving fermentation outcomes | Multiple yeast species | [92] |
| Yeast-yeast interactions, including S. cerevisiae with non-Saccharomyces species | Wickerhamomyces anomalus, Hanseniaspora vineae | [93] |
| Persistence of certain non-Saccharomyces yeasts in vineyard ecosystems | Starmerella bacillaris, Lachancea thermotolerans | [94] |
| Yeast ecosystem modulation by S. cerevisiae | Various non-Saccharomyces species | [80] |
| Optimization of yeast interactions for improved fermentation | Selected co-cultures of Saccharomyces and non-Saccharomyces | [89] |
Table 6.
Key Genetic Modifications in wine and beer yeasts.
| Process | Technique | References |
|---|---|---|
| GPD1 overexpression and ALD6 delation to reduce reduce alcohol yield in wine yeast | Episomal vector; KanMX deletion cassette | [94] |
| Expression of extracellular hydrolytic enzymes to improve juice extraction and release primary aromas | Episomal vector constructed by restriction cloning | [106] |
| Reduction of urea and ethyl carbamate formation | CRISPR/Cas9 | [107] |
| Overexpression of LEU4 (α-isopropylmalate synthase) in sake yeast to increase isoamyl alcohol and esters | Plasmid multicopy integration at chromosome level | [108] |
| Reduction of ethyl carbamate production | Plasmid distruption | [109] |
| Heterologous expression of pediocin to increase resistance to wild yeasts and bacteria | Episomal vector constructed by restriction cloning | [110] |
| Expression of acetolactate decarboxylase (ALDC) to reduce diacetyl formation | Episomal vector constructed by restriction cloning | [111] |
| Expression of malolactic enzymes to degrade malate and integrate malolactic fermentation | Episomal vector constructed by restriction cloning | [112] |
| Engineering yeast strains to produce hop monoterpenes | Plasmids obtained by Golden Gate assembly | [113] |
| Engineering yeast strains to produce methyl anthranilate with grape aroma | CRISPR/Cas9 | [114] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



