
Submitted:
14 May 2025
Posted:
16 May 2025
You are already at the latest version
Abstract
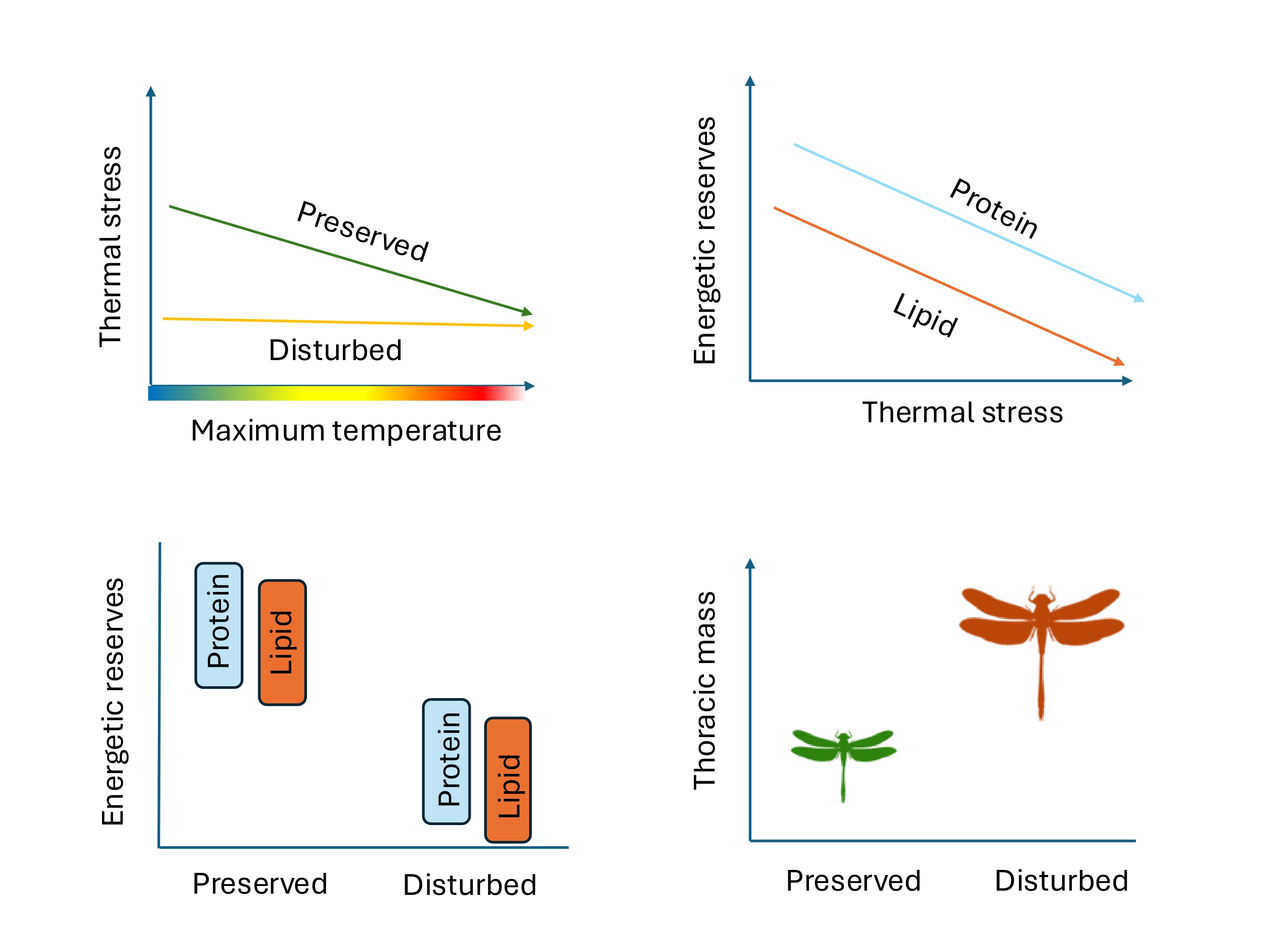
Anthropogenic disturbance alters macro and microclimatic conditions, often increasing ambient temperatures. These changes can strongly affect insects, particularly those experiencing high thermal stress (i.e, large differences between body and environmental temperature), as prolonged exposure to elevated temperatures can reduce their energetic reserves due to increased metabolic demands and physiological stress. We evaluated thermal stress in 16 insect dragonfly species during two sampling periods (2019 and 2022) in preserved and disturbed sites within a tropical dry forest in western Mexico. Also, we compared energetic condition (lipid and protein content) and thoracic mass for the seven most abundant species between both habitat types. In preserved sites, insects showed higher thermal stress at lower maximum temperatures, which decreased as temperatures increased. Dragonflies in disturbed sites maintained consistent levels of thermal stress across the temperature gradient. Thermal stress and body size were positively associated, and both were linked to lower lipid and protein content. Despite this, individuals from disturbed sites were larger but had lower energy reserves than those from preserved sites. Our findings suggest that although species can persist in disturbed environments, their energetic condition may be compromised, especially in individuals experiencing high thermal stress — potentially affecting their performance and fitness. Preserving suitable habitats is essential not only for maintaining species diversity but also ensuring ecological functions, particularly those performed by flying insects with high thermal stress such as dragonflies.
Keywords:
conservation
; dragonflies
; tropics
; disturbance
; body size
; thermal stress
; protein
; lipid
1. Introduction
Anthropogenic disturbances, such as land-use changes for livestock farming, agriculture, and urban expansion, have altered environmental conditions, leading to water and air pollution and deforestation [1,2,3]. Reduction in vegetation cover modifies microclimatic conditions, increasing solar radiation exposure and altering the relative humidity [4,5,6]. These changes can be detrimental, or even lethal to ectothermic insects since they depend on environmental temperature to perform essential functions such as foraging, development, and reproduction[7].
Temperature increase and habitat disturbance have been linked to declines in insect abundance and diversity [3,8,9]. However, some species persist in disturbed environments, suggesting that certain functional traits enhance their thermal tolerance. For example, Giménez Gómez et al. [10] showed that smaller dung beetles with lower thermal stress (i.e., the difference between body temperature and ambient temperature) could exploit disturbed open-canopy habitats more effectively. Similarly, Rocha et al. [11] reported that heliothermic odonates, which tolerate higher levels of solar exposure perform better in disturbed habitats, whereas thermoconforming species tend to favor preserved habitats.
Species from the suborder Anisoptera (also called dragonflies, order Odonata) have been described as capable of tolerating extreme temperature conditions and highly disturbed habitats. For instance, Castillo-Pérez et al. [12] found a positive correlation between dragonfly taxonomic diversity and higher temperatures, while Suárez-Tovar et al. [13] reported that dragonfly communities were relatively unaffected by urbanization. This tolerance has been associated with dragonfly high dispersal ability and their thermoregulatory capacity, which relies on both behavioral and physiological mechanisms [14,15,16]. However, thermoregulatory strategies, such as seeking shade to avoid overheating or adjusting flight patterns, can be energetically costly [17,18]. These energetic costs may be further exacerbated by reduced foraging and mating opportunities, as individuals potentially invest more time to behavioral thermoregulation [19]. Prolonged thermal stress may also lead to a depletion of energy reserves [17]. In relation to this, a substantial body fat reduction can be caused by the dead of fat body cells, as found in Drosophila [20]. The dead of such cells reduce the ability to store more lipids.
Dragonflies inhabiting tropical dry forests can experience high thermal stress in disturbed sites, due to high temperatures, reduced vegetation cover and periods of water scarcity [21,22] . This can be quantified as the difference between body and environmental temperature and is primarily influenced by muscular activity before and during flight [10,23,24]. Body size has also been identified as a trait influencing thermal stress, as smaller odonates are capable of maintaining lower basal metabolic rates (BMR) under high-temperature stress compared to larger individuals [25]. As indicated before, high temperatures and thermal stress may lead to a decline in energy reserves [20,26]. Therefore, the lower BMR in small dragonflies potentially reduces their energetic demands in extreme heat conditions, providing an advantage in disturbed environments where resource availability is limited. The extent to which dragonflies can modulate thermal stress may depend on habitat conditions, with individuals in preserved and disturbed environments showing different behavioral thermoregulatory responses. Besides modifying environmental temperature, habitat disturbance can also influence prey availability and nutritional quality [27], further affecting energy reserves and body size of individuals in disturbed environments. This reduction in energy availability may, in turn, increase the energy costs of thermoregulation, although the underlying mechanisms are not yet fully understood.
This study investigated the relationship between thermal stress and the energetic condition of dragonflies in preserved and disturbed sites in a highly degraded and fragile tropical dry forest with high temperatures and a pronounced dry season in western Mexico. First, we predict that individuals who inhabit preserved sites with lower temperatures will show lower levels of thermal stress, while individuals who inhabit disturbed sites with higher temperatures will exhibit higher thermal stress. Second, we expect that species with higher levels of thermal stress will show poorer energetic reserves (i.e., lipid or protein reserves) in disturbed (hotter) sites. Third, we predict that individuals from disturbed sites will have a smaller thoracic mass.
2. Materials and Methods
2.1. Study Site
We carried out this study in the deciduous tropical forest of the Chamela-Cuixmala Reserve and its surroundings, located in the municipality of La Huerta, Jalisco, Mexico (19°29'39.8 "N, 105°02'48.4 "W). The area features a warm, subhumid climate with a pronounced dry season from November to June [28]. The average annual rainfall is 832 ± 277 mm, though there is wide variation between years, ranging from 340 to 1394 mm [29]. The maximum temperature in the region is 37 °C[30,31]. The area comprises a mosaic of intact tropical forests, agricultural and livestock fields, and second-growth forests in multiple successional stages [32,33].
Data were collected in 14 sites: night were anthropogenically disturbed, and five were preserved (Figure 1). Disturbed sites consisted of ponds, rivers or streams outside the Chamela-Cuixmala Reserve, on non-conservation land, near houses, hotel complexes, agricultural plots, and cattle pastures [30]. In contrast, preserved sites included similar water bodies — ponds, rivers or streams located whitin the Chamela-Cuixmala Reserve, where forests have been under protection for over 50 years and represent an important remnant of native old-growth vegetation [29]. Furthermore, preserved sites in this study have a higher percentage of vegetation cover compared to the disturbed sites [30,32].
2.2. Environmental and Body Temperature Recording
Between September 6 and October 5, 2019, and between August 31 and September 30, 2022, we recorded ambient and maximum temperatures at the 14 sampling sites using a HOBO® data logger (MX 2201, ±0.5°C) positioned 1.5 meters above the ground. Measurements were taken every five minutes for a minimum of two days, allowing us to obtain both the maximum and average temperature values for each sampling point. The body temperature of adult male anisopterans was recorded between 10:00 and 15:00 h (UTC-6), which corresponds to the hours of maximum activity in odonates. To determine the individual's body temperature, we captured them using an entomological net while they were flying. After capture, we held individuals by the wings with a set of metal forceps and took three thermographic images at 3-second intervals using a FLIR® model E6 camera (resolution of 160 x 120 pixels with a spectral range of 7.5- 13 µm and a thermal sensitivity of < 60 mK at 30°C). The camera was calibrated using the standard calibration service provided by FLIR® to guarantee accuracy. Heat exchange between the observer and the experimental individual was reduced by taking the thermographic images within five seconds of gripping the wings with the forceps, avoiding exposing individuals to direct solar radiation when the thermographic image was taken. Once the thermographic images were stored in the memory of the thermographic camera, the right hindwing of each individual was marked with a fine-tipped black marker to prevent recaptures. Additionally, we measured total body length, extending from the head to the last abdominal segment, as a measure of body size [34] using a Lion Tools digital caliper (± 0.05 mm). Individuals were released at the same site where they were captured.
2.3. Thermographic Image Analysis and Thermal Stress Analysis
We used FLIR Tools® software to obtain temperatures of synthorax (1mm of perimeter), which is the warmest tagma of odonates[16]. The average of the three thermographic measurements taken for each individual was considered the body temperature (Tb). To estimate thermal stress, we calculated the difference between the body temperature and the ambient temperature (Tb-Ta) at the time of capture, following the method described by Giménez Gómez et al. [10].
2.4. Energy Reserves Calculation
We captured males of the seven most common anisopteran species — occurring in both preserved and disturbed sites — using an entomological net between August 31 and September 30, 2022, from 10:00 to 150:0 h (UTC-6). Only individuals from these seven species were used for body condition analyses (Table S1). After capture, individuals were placed in centrifuge tubes and stored in a cooler at 5°C. Subsequently, they were sacrificed in a freezer at -20°C, where they remained until we performed the individual condition analyses.
Protein and lipid content (µg/mg) were quantified for each individual using the protocol of Foray et al.[35]. The thorax of individuals — which contains the flight muscles and lipid reserves used in energetically demanding activities such as flying [36]— was isolated and its mass measured using a Velab VE-210 analytical balance (sensitivity: 0.1 mg; maximum capacity: 210 g). To prepare the samples, thoraxes were placed in 2 ml centrifuge tubes with steel beads and 180 µL of phosphate lysis buffer (100 mM KH₂PO₄, 1 mM DTT, 1 mM EDTA, pH 7.4). The tissues were homogenized into a hyaline solution using a TissueLyser II bead mill (Qiagen, Valencia, CA, U.S.A.). For protein quantification, the homogenized thoracic tissue was centrifuged at 180 g relative centrifugal force (RCF) for 5 min at 4°C. Then, 2.5 μL of the supernatant were taken by duplicate and placed into a 96-well microplate. Bradford reagent (250μL) was added, and the plate was incubated for 20 minutes. Absorbance was measured spectrophotometrically at 595 nm using an EPOCH spectrophotometer (200-900nm range; BioTek, Winooski, VT, USA). Protein content in the tissue was determined by comparing absorbance values with a calibration curve prepared using a dilution series of bovine serum albumin.
For lipid measurement, 180 μL of the thorax tissue homogenate were mixed with 20μL of Na2SO4, 5μL of phosphate lysis buffer, and 1500 μL of methanol: chloroform solution (2:1). The mixture was vortexed for 2 minutes and centrifuged twice for 15 minutes at 180 g relative centrifugal force (RCF). Then, 100 μL of the supernatant were transferred in duplicate to a U-bottom plate and heated at 90°C in a water bath until complete evaporation. After evaporation, 10μL of 98% sulfuric acid was added to each well, and the plate was incubated at 90°C for 2 min. The reaction was cooled, followed by the addition of 190 μL vanillin reagent. After a 15-minute incubation, absorbance was measured at 525 nm. Lipid content was calculated using a calibration curve based on a glyceryl trioleate lipid dilution series.
2.5. Statistical Analysis
We employed linear-mixed models (LMMs) to assess the effects of maximum temperature, site condition (preserved/disturbed), and body size on thermal stress. Fixed effects included site condition (preserved/disturbed), body size, maximum temperature and the interaction between site condition and maximum temperature. Collection year, sampling site, and species were added as random effects. Multiple candidate models were evaluated, and the most parsimonious model was selected based on the Akaike Information Criterion (AIC). No collinearity was found among predictor variables (VIF <5).
To analyze the relationship between energetic reserves (protein and lipid content) and body size with mean thermal stress, site condition, and mean site temperature, we constructed generalized linear mixed models (GLMMs) with a Gaussian distribution using the "glmmTMB" package [37]. Separate models were built for each response variable (protein content, lipid content, and body size) and sampling site and species were included as random effects. All response variables were log-transformed to meet model assumptions of normality and homogeneity of variance. Prior to model selection, we assessed multicollinearity among predictors using the variance inflation factor (VIF), and found no indication of collinearity (VIF <5). Candidate models were compared using the Akaike Information Criterion (AIC), and the most parsimonious model was retained for interpretation [38].
Model validation for the linear mixed analysis was conducted visually through diagnostic plots, including theoretical quantiles versus standardized residuals (Normal Q-Q plot) and residuals versus fitted values. Additionally, Shapiro-Wilk tests were performed to assess residual normality, and Breusch-Pagan tests were used to check for homogeneity of variance using the "performance" package [39]. The generalized mixed models were validated using the "DHARMa" package [40]. Specifically, we used the "simulateResiduals" function to simulate residuals and conducted a uniformity test to verify residual normality. We also applied the "testDispersion" function to detect potential dispersion issues. All analyses were performed using R version 4.3.2 [41].
3. Results
3.1. Environmental Temperatures
Disturbed sites showed significantly higher air temperatures (35.71 °C [± 4.02 °C SD]) compared to preserved sites (32.60 °C [± 4.35 °C SD]; t = -7.189, p <0.001).
3.2. Thermal Stress
We analyzed thermal stress in 393 adult individuals belonging to 16 species (12 species in preserved sites and 14 in disturbed sites; Table S1) during two sampling periods (2019 and 2022). The species with the highest average thermal stress were Tramea onusta and Dythemis nigrescens, while those with the lowest were Perithemis intensa and Orthemis levis (Figure S1). According to the best linear mixed-effects model (Table S3), there was a significant interaction between maximum environmental temperature and site condition (t=2.05, p=0.041; Figure 2 A). Dragonflies from preserved sites showed higher thermal stress at lower maximum temperatures (t=-2.33, p=0.021); however, they reduced their thermal stress as maximum temperatures increased. In contrast, individuals from disturbed sites maintained a constant level of thermal stress at all maximum temperatures. Dragonflies in disturbed sites consistently exhibited lower overall thermal stress compared to those in preserved sites, but this difference was only significant at lower maximum temperatures (t=-2.33, p=0.021). Maximum environmental temperatures were negatively associated with thermal stress (t=-3.80, p<0.001), and larger individuals showed higher thermal stress (t=2.31, p=0.021; Figure 2 B).
3.3. Energy Reserves
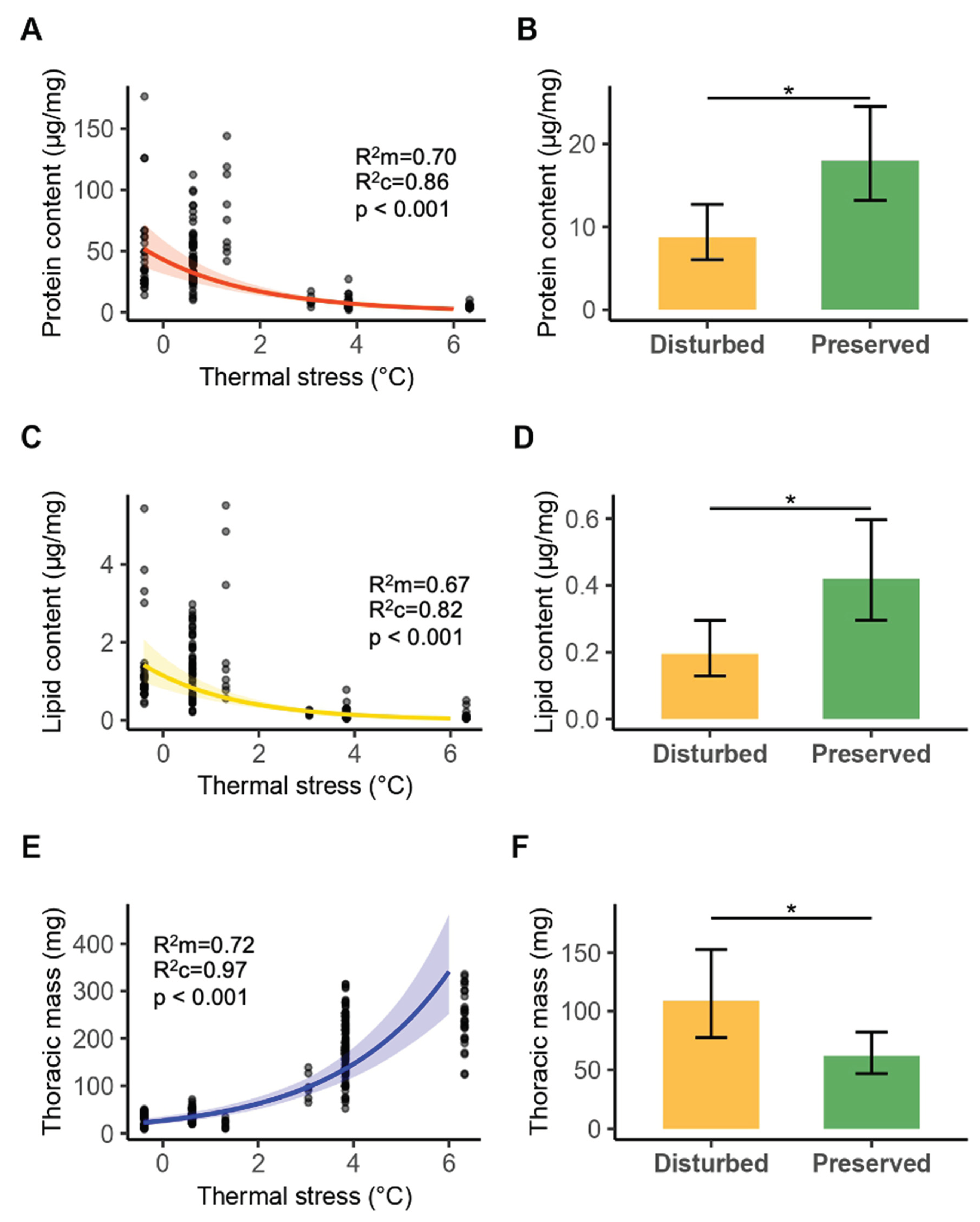
We assessed the energetic reserves (lipids and proteins) and thoracic mass exclusively in males of the seven most common anisopteran species, collected in 2022 from preserved (114 individuals) and disturbed (126 individuals) sites (Table S1). According to the best-fitting generalized mixed models (Table S2), individuals experiencing higher thermal stress had significantly lower protein content (z= -10.655, p <0.001; Figure 3A) and lipid content (z=-10.658, p< 0.001; Figure 3C). Additionally, individuals from disturbed sites showed lower protein content (z=-2.511, p= 0.012; Figure 3B) and lipid content (z= -2.382, p=0.017; Figure 3D). However, individuals under higher thermal stress displayed a larger body size (z=10.680, p < 0.001). Similarly, individuals from disturbed sites had a significantly larger thoracic mass (z= 2.165, p= 0.030).
4. Discussion
The main salient results of our research are as follow. First, based on data from 16 dragonfly species collected in two periods (2019 and 2022), individuals from preserved sites experience higher thermal stress at lower maximum temperatures but can reduce their thermal stress at higher temperatures, whereas dragonflies from disturbed sites maintain a consistent level of thermal stress across different maximum temperatures. Second, using data exclusively from the seven most common anisopteran species collected in 2022, we found that individuals experiencing higher thermal stress and greater thoracic mass had lower protein and lipid content. Third, again focusing only on these seven most common species present in both preserved and disturbed sites, individuals from preserved sites exhibited smaller body size but higher protein and lipid content than those from disturbed sites.
Contrary to our predictions, individuals from preserved sites showed higher thermal stress at lower maximum temperatures; however, they reduced their thermal stress as maximum temperatures increased. In contrast, individuals from disturbed sites maintained a constant level of thermal stress at all maximum temperatures. We hypothesize that individuals from disturbed sites may be engaging in active thermoregulation to maintain a stable level of thermal stress since species experiencing high thermal stress should rely on active thermoregulatory mechanisms to diminish thermal stress and prevent reaching a critical heat shock temperature during flight [10]. This pattern may also be associated with body size, as larger odonates generally have greater thermal inertia, which could help buffer temperature fluctuations [18,25,42]. In accordance with this pattern, Rocha et al.[11] described how medium and large-sized Odonata species were favored in disturbed environments due to their more efficient thermoregulatory mechanisms. Similarly, Castillo-Pérez et al. [12] found that the most abundant species in disturbed sites within our study area exhibited a larger body size, suggesting that these species may also rely on more effective thermoregulatory mechanisms — either behavioral, such as selecting cooler microhabitats or adopting obelisk posture, or physiological, including hemolymph circulation [14,25,43,44]
While species from preserved sites experience higher thermal stress at lower maximum temperatures, their smaller body size also implies a higher rate of heat exchange through convection due to their higher surface-to-volume ratio [15]. Additionally, preserved sites have been shown to exhibit greater habitat heterogeneity and vegetation cover in previous studies[12,22], which likely provides more perching opportunities and greater access to shade. Therefore, if temperatures become extremely high, intensifying thermal stress, these species may mitigate it by seeking cooler microhabitats[15,44]. However, the thermoregulatory mechanisms of species inhabiting disturbed and preserved sites require further investigation to understand their role in thermal adaptation.
Thermoregulatory mechanisms, whether physiological or behavioral, and constant exposure to high temperatures, can be energetically costly [17,18,45]. While our models did not show a significant effect of either average or maximum temperature on energy reserves, disturbed sites that have greater sun exposure and higher average and maximum temperatures [12,22], may impose an additional energetic cost on both adults and larvae. For example, Córdoba-Aguilar and Rocha-Ortega [46] found that increased sun exposure due to vegetation loss forced the damselfly Hetaerina americana to spend more time in sunny areas, likely increasing energetic costs. Similarly, Tüzün and Stoks [47] found that larvae of the damselfly Coenagrion puella inhabiting rural areas experienced a decrease in energy availability under experimentally induced heatwaves. Therefore, exposure to high temperatures in disturbed sites may still reduce energetic reserves of individuals. Further observational studies are needed to better assess the energetic costs of high temperatures under natural conditions.
According to our findings, higher thermal stress reduced lower energy reserves (proteins and lipids), regardless of habitat type. This could be related to their higher flight frequency [15,48]. Flight is one of the most metabolically demanding activities in insects, relying primarily on lipids — and, to a lesser extent, proteins — as fuel [35,49,50,51]. For instance, in the anisopteran Pantala flavescens, lipid reserves were observed to decrease after prolonged flight periods [52]. Consistent with these results, we found that individuals from disturbed sites had lower energy reserves (proteins and lipids), regardless of their thermal stress levels. Environmental factors such as wastewater contamination and loss of vegetation cover in the study area may further contribute to this decline in energy reserves. For example, Hetaerina americana exhibited lower energy reserves due to increased water pH in urbanized areas [46,53]. Similarly, in dung beetles, Dichotomius guaribensis showed reduced energetic condition as urbanization increased [54]. Therefore, species experiencing higher thermal stress may be more vulnerable to habitat disturbance, as they face the combined effect of environmental disturbance and the high energetic demands associated with coping with high thermal stress. Another consequence of a reduced lipid and protein content is that fitness can be impaired as these triats are linked to key life history traits. For example, while lipids are used to sustain flight and repell intruders during male competition over mating territories[55], proteins are used to produce eggs [56]. We hypothesize that this fitness impairment can lead to an eventual population collapse.
Contrary to our expectations, thoracic mass was significantly larger in disturbed sites across the seven species that inhabit both habitat types (disturbed/preserved) despite their lower energetic reserves. This results contrast with previous studies on damselflies [46,57], butterflies [58] and bumblebees [59], where body size was smaller in disturbed sites. One hypotesis is that larger body size could confer greater thermal tolerance in disturbed sites, as suggested by previous studies in odonates [22]. This advantage may be related to a higher capacity for water storage in larger individuals [60]. Future studies should further investigate this relationship.
Odonates are considered highly resilient to habitat modification although the supporting evidence has used presence/absence of individuals and/or species for this claim [61,62]. Despite this, our results suggest that although species persist in disturbed environments, their overall energy condition is compromised, . This pattern could also extend to other flying insect groups that experience high thermal stress, such as moths and bumblebees [23,63]. Furthermore, if maximum temperatures continue to rise at micro and macro scales, species with high thermal stress may face additional challenges. Our findings indicate that these species likely engage in behavioral or physiological thermoregulatory strategies to keep their thermal stress (i.e, the difference between body and ambient temperature) within functional limits in disturbed sites, leading to higher energy costs. Thermal ecology research is essential for developing conservation strategies tailored to highly mobile insect species such as dragonflies, which play crucial roles in ecosystem functions such as transferring materials and energy from aquatic to terrestrial ecosystems [64,65] and by regulating populations of other insects, including human disease vectors such as mosquitoes [66].
5. Conclusions
Our study highlights how thermal stress, energy reserves, and body size vary in dragonflies inhabiting preserved and disturbed habitats within a tropical dry forest characterized by high temperatures. We found that in disturbed habitats, dragonflies maintain a stable thermal stress across different temperatures, while in preserved habitats, species experience higher thermal stress at lower maximum temperatures but reduce it as temperatures increase. The energetic costs of thermoregulation appear to be higher in disturbed habitats, as individuals experiencing high thermal stress exhibited lower lipid and protein reserves. Finally, odonate individuals in disturbed sites exhibited larger body sizes despite having lower energy reserves. This pattern may be explained by a higher thermal tolerance associated with larger body size, which has been suggested as an adaptive advantage under thermally challenging conditions. These findings emphasize the importance of studying thermal ecology in insect species living in disturbed environments to inform conservation efforts.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, E.U.C.-P., A.E.-C., C.M.S.-T., J.D.R.-D., D.G.-T. and A.C.-A.; methodology, E.U.C.-P., A.E.-C., C.M.S.-T. and A.C.-A.; data curation, E.U.C.-P., A.E.-C., C.M.S.-T. and J.D.R.-D.; formal analysis, E.U.C.-P. and J.D.R.-D.; investigation, E.U.C.-P.; visualization, E.U.C.-P., J.D.R.-D., D.G.-T. and A.C.-A.; validation, J.D.R.-D., D.G.-T. and A.C.-A.; writing—original draft preparation, E.U.C.-P., A.E.-C., C.M.S.-T., J.D.R.-D., D.G.-T. and A.C.-A.; writing—review and editing, J.D.R.-D., D.G.-T. and A.C.-A.; supervision, D.G.-T. and A.C.-A.; project administration, A.C.-A.; funding acquisition, A.C.-A.; resources, A.C.-A.
Funding
This work was made possible thanks to financing from the PAPIIT grants IN206618 and IN204921. E.U.C.-P, is supported by a scholarship awarded by the Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI).
Institutional Review Board Statement
The collection of specimens was approved by the Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT) of Mexico, under licence number 09/K4-0129/12/24.
Data Availability Statement
The original data presented in the study are openly available in FigShare at: https://doi.org/10.6084/m9.figshare.28953008.v1.
Conflicts of Interest
Authors declare not to have any conflict of interests.
References
- Püttker, T.; Crouzeilles, R.; Almeida-Gomes, M.; Schmoeller, M.; Maurenza, D.; Alves-Pinto, H.; Pardini, R.; Vieira, M. V; Banks-Leite, C.; Fonseca, C.R.; et al. Indirect Effects of Habitat Loss via Habitat Fragmentation: A Cross-Taxa Analysis of Forest-Dependent Species. Biol Conserv 2020, 241, 108368. [Google Scholar] [CrossRef]
- Piessens, K.; Adriaens, D.; Jacquemyn, H.; Honnay, O. Synergistic Effects of an Extreme Weather Event and Habitat Fragmentation on a Specialised Insect Herbivore. Oecologia 2009, 159, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Schowalter, T.D. Insect Responses to Major Landscape-Level Disturbance. Annu Rev Entomol 2012, 57, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Royer, P.D.; Cobb, N.S.; Clifford, M.J.; Huang, C.-Y.; Breshears, D.D.; Adams, H.D.; Villegas, J.C. Extreme Climatic Event-Triggered Overstorey Vegetation Loss Increases Understorey Solar Input Regionally: Primary and Secondary Ecological Implications. Journal of Ecology 2011, 99, 714–723. [Google Scholar] [CrossRef]
- Tuff, K.T.; Tuff, T.; Davies, K.F. A Framework for Integrating Thermal Biology into Fragmentation Research. Ecol Lett 2016, 19, 361–374. [Google Scholar] [CrossRef]
- Zlotnick, O.B.; Musselman, K.N.; Levy, O. Deforestation Poses Deleterious Effects to Tree-Climbing Species under Climate Change. Nat Clim Chang 2024, 14, 289–295. [Google Scholar] [CrossRef]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in Fluctuating Thermal Environments. Annu Rev Entomol 2015, 60, 123–140. [Google Scholar] [CrossRef]
- Outhwaite, C.L.; McCann, P.; Newbold, T. Agriculture and Climate Change Are Reshaping Insect Biodiversity Worldwide. Nature 2022, 605, 97–102. [Google Scholar] [CrossRef]
- Wagner, D.L. Insect Declines in the Anthropocene. Annu Rev Entomol 2020, 65, 457–480. [Google Scholar] [CrossRef]
- Giménez Gómez, V.C.; Verdú, J.R.; Zurita, G.A. Thermal Niche Helps to Explain the Ability of Dung Beetles to Exploit Disturbed Habitats. Sci Rep 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Rocha, T.S.; Calvão, L.B.; Juen, L.; Oliveira-Junior, J.M.B. Effect of Environmental Integrity on the Functional Composition of the Odonata (Insecta) Community in Streams in the Eastern Amazon. Front Ecol Evol 2023, 11. [Google Scholar] [CrossRef]
- Castillo-Pérez, E.U.; Rivera-Duarte, J.D.; Abellán, P.; del-Val, E.; González-Tokman, D.; Córdoba-Aguilar, A. Thriving in the Heat: How High Temperatures and Habitat Disturbance Shape Odonate Taxonomic and Functional Diversity in the Tropics. Insect Conserv Divers 2024. [Google Scholar] [CrossRef]
- Suárez-Tovar, C.M.; Ulises Castillo-Pérez, E.; Antonio Sandoval-García, I.; Schondube, J.E.; Cano-Santana, Z.; Córdoba-Aguilar, A. Resilient Dragons: Exploring Odonata Communities in an Urbanization Gradient. Ecol Indic 2022, 141, 109134. [Google Scholar] [CrossRef]
- Castillo-Pérez, U.; May, M.L.; Córdoba-Aguilar, A. Thermoregulation in Odonata. Dragonflies and Damselflies 2022, 101–112. [Google Scholar]
- May, M.L. Thermoregulation and Adaptation to Temperature in Dragonflies (Odonata : Anisoptera). Ecol Monogr 1976, 46, 1–32. [Google Scholar] [CrossRef]
- Polcyn, D.M. Thermoregulation During Summer Activity in Mojave Desert Dragonflies (Odonata: Anisoptera). Funct Ecol 1994, 8, 441–449. [Google Scholar] [CrossRef]
- Lahondère, C. Recent Advances in Insect Thermoregulation. Journal of Experimental Biology 2023, 226. [Google Scholar] [CrossRef]
- May, M. Insect Thermoregulation. Annu Rev Entomol 1979, 24, 313–349. [Google Scholar] [CrossRef]
- Ma, G.; Ma, C. Sen Effect of Acclimation on Heat-Escape Temperatures of Two Aphid Species: Implications for Estimating Behavioral Response of Insects to Climate Warming. J Insect Physiol 2012, 58, 303–309. [Google Scholar] [CrossRef]
- Klepsatel, P.; Gáliková, M.; Xu, Y.; Kühnlein, R.P. Thermal Stress Depletes Energy Reserves in Drosophila. Sci Rep 2016, 6, 33667. [Google Scholar] [CrossRef]
- García-Oliva, F.; Camou, A.; Maass, J.M. El Clima de La Región Central de La Costa Del Pacífico Mexicano. In Historia Natural de Chamela; Noguera-Alderte, A.N., Vega-Rivera, J.H., García-Aldrete, A.N., Quesada, M., Eds.; Instituto de Biología, UNAM: México, 2002; pp. 3–10. [Google Scholar]
- Castillo-Pérez, E.U.; Suárez-Tovar, C.M.; González-Tokman, D.; Schondube, J.E.; Córdoba-Aguilar, A. Insect Thermal Limits in Warm and Perturbed Habitats: Dragonflies and Damselflies as Study Cases. J Therm Biol 2022, 103, 103164. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, B. Insect Thermoregulation. Endeavour 1995, 19, 28–33. [Google Scholar] [CrossRef]
- Verdú, J.R.; Arellano, L.; Numa, C. Thermoregulation in Endothermic Dung Beetles (Coleoptera: Scarabaeidae): Effect of Body Size and Ecophysiological Constraints in Flight. J Insect Physiol 2006, 52, 854–860. [Google Scholar] [CrossRef]
- Bota-Sierra, C.A.; Cordero-Rivera, A.; Novelo-Gutiérrez, R.; Sánchez-Herrera, M.; Londoño, G.A. Can High Temperatures Affect Body Size in Insects? The Case of Rubyspot Damselflies in the Colombian Western Andes. Diversity (Basel) 2024, 16, 743. [Google Scholar] [CrossRef]
- Klepsatel, P.; Wildridge, D.; Gáliková, M. Temperature Induces Changes in Drosophila Energy Stores. Sci Rep 2019, 9, 5239. [Google Scholar] [CrossRef]
- Portela Salomão, R.; González-Tokman, D.; Dáttilo, W.; López-Acosta, J.C.; Favila, M.E. Landscape Structure and Composition Define the Body Condition of Dung Beetles (Coleoptera: Scarabaeinae) in a Fragmented Tropical Rainforest. Ecol Indic 2018, 88, 144–151. [Google Scholar] [CrossRef]
- Martínez-Ibarra, J.A.; Martínez-Hernández, F.; Villalobos, G.; Vences-Blanco, M.O.; Salazar-Schettino, P.M. Update on the Distribution of Triatoma Bolivari and Triatoma Brailovskyi (Hemiptera: Reduviidae: Triatominae) in Western Mexico. Journal of Vector Ecology 2010, 35, 432–434. [Google Scholar] [CrossRef]
- Takano-Rojas, H.; Murray-Tortarolo, G.; Maass, M.; Castillo, A. Characterization, Variability and Long-Term Trends on Local Climate in a Mexican Tropical Dry Forest. International Journal of Climatology 2023, 43, 5077–5091. [Google Scholar] [CrossRef]
- Flores-Casas, R.; Ortega-Huerta, M.A. Modelling Land Cover Changes in the Tropical Dry Forest Surrounding the Chamela-Cuixmala Biosphere Reserve, Mexico. Int J Remote Sens 2019, 40, 6948–6974. [Google Scholar] [CrossRef]
- IBUNAM Datos Climáticos Estación Chamela Available online:. Available online: http://www.ibiologia.unam.mx/ebchamela/www/clima.html (accessed on 1 November 2019).
- Sánchez-Azofeifa, G.A.; Quesada, M.; Cuevas-Reyes, P.; Castillo, A.; Sánchez-Montoya, G. Land Cover and Conservation in the Area of Influence of the Chamela-Cuixmala Biosphere Reserve, Mexico. For Ecol Manage 2009, 258, 907–912. [Google Scholar] [CrossRef]
- Villa-Galaviz, E.; Boege, K.; Del-Val, E. Resilience in Plant-Herbivore Networks during Secondary Succession. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Johansson, F.; Crowley, P.H.; Brodin, T. Sexual Size Dimorphism and Sex Ratios in Dragonflies (Odonata). Biological Journal of the Linnean Society 2005, 86, 507–513. [Google Scholar] [CrossRef]
- Foray, V.; Pelisson, P.-F.; Bel-Venner, M.-C.; Desouhant, E.; Venner, S.; Menu, F.; Giron, D. ; Rey Benjamin A Handbook for Uncovering the Complete Energetic Budget in Insects: The van Handel’s Method (1985) Revisited. Physiol Entomol 2012, 37, 295–302. [Google Scholar] [CrossRef]
- Marden, J.H. Dragonfly Flight Performance: A Model System for Biomechanics, Physiological Genetics, and Animal Competitive Behaviour. Dragonflies and Damselflies. Model Organisms for Ecological and Evolutionary Research. Oxford University Press, Oxford 2008, 249–261.
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. GlmmTMB Balances Speed and Flexibility Among Packages for Zero-Inflated Generalized Linear Mixed Modeling. R J 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Wagenmakers, E.-J.; Farrell, S.; Wagenmakers, -J AIC Model Selection Using Akaike Weights; 2004.
- Lüdecke, D.; Ben-Shachar, M.; Patil, I.; Waggoner, P.; Makowski, D. Performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J Open Source Softw 2021, 6, 3139. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models 2024.
- R Core Team R: A Language and Environment for Statistical Computing 2023.
- Chown, S.L.; Nicolson, S.W. Thermoregulation. In Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press: Oxford, 2004. [Google Scholar]
- Corbet, P.S. Dragonflies: Behaviour and Ecology of Odonata; Colchester,UK, 1999; ISBN 0-8014-2592-1.
- Heinrich, B.C.; Casey, T.M. Heat Transfer in Dragonflies: ‘Fliers’ and ‘Perchers. ’ Journal of Experimental Biology 1978, 74, 17–36. [Google Scholar] [CrossRef]
- González-Tokman, D.; Córdoba-Aguilar, A.; Dáttilo, W.; Lira-Noriega, A.; Sánchez-Guillén, R.A.; Villalobos, F. Insect Responses to Heat: Physiological Mechanisms, Evolution and Ecological Implications in a Warming World. Biological Reviews 2020, 95, 802–821. [Google Scholar] [CrossRef]
- Córdoba-Aguilar, A.; Rocha-Ortega, M. Damselfly (Odonata: Calopterygidae) Population Decline in an Urbanizing Watershed. Journal of Insect Science 2019, 19, 30. [Google Scholar] [CrossRef]
- Tüzün, N.; Stoks, R. Lower Bioenergetic Costs but Similar Immune Responsiveness under a Heat Wave in Urban Compared to Rural Damselflies. Evol Appl 2021, 14, 24–35. [Google Scholar] [CrossRef]
- Corbet, P.S.; May, M.L. Fliers and Perchers among Odonata: Dichotomy or Multidimensional Continuum? A Provisional Reappraisal. International Journal of Odonatology 2008, 11, 155–171. [Google Scholar] [CrossRef]
- Candy, D.J.; Becker, A.; Wegener, G. Coordination and Integration of Metabolism in Insect Flight*. Comp Biochem Physiol B Biochem Mol Biol 1997, 117, 497–512. [Google Scholar] [CrossRef]
- Janssens, M. Hormonal Control of Flight Metabolism in Odonata?, 1995.
- Sacktor, B. Biochemistry of Insect Flight. In Insect Biochemistry and Function; Candy, D.J., Kilby, B.A., Eds.; Springer US: Boston, MA, 1975; ISBN 978-1-4899-3204-4. [Google Scholar]
- Kallapur, V.L.; George, C.J. Fatty Acid Oxidation by the Flight Muscles of the Dragonfly, Pantala Flavescens. J Insect Physiol 1973, 19, 1035–1040. [Google Scholar] [CrossRef]
- Suárez-Tovar, C.M.; Rocha-Ortega, M.; Córdoba-Aguilar, A. Is Body Condition of Mexican Rubyspot (Odonata:Zygoptera) Associated with Urbanization? J Insect Conserv 2023, 27, 961–969. [Google Scholar] [CrossRef]
- Salomão, R.P.; Alvarado, F.; Baena-Díaz, F.; Favila, M.E.; Iannuzzi, L.; Liberal, C.N.; Santos, B.A.; Villegas-Guzmán, G.A.; González-Tokman, D. Negative Effects of Urbanisation on the Physical Condition of an Endemic Dung Beetle from a Neotropical Hotspot. Ecol Entomol 2020, 45, 886–895. [Google Scholar] [CrossRef]
- Córdoba-Aguilar, A.; González-Tokman, D.M. The Behavioral and Physiological Ecology of Adult Rubyspot Damselflies (Hetaerina, Calopterygidae, Odonata). Adv Study Behav 2014, 46, 311–341. [Google Scholar] [CrossRef]
- Roeder, K.A.; Behmer, S.T. Lifetime Consequences of Food Protein-Carbohydrate Content for an Insect Herbivore. Funct Ecol 2014, 28, 1135–1143. [Google Scholar] [CrossRef]
- Suárez-Tovar, C.M.; Rocha-Ortega, M.; González-Voyer, A.; González-Tokman, D.; Córdoba-Aguilar, A. The Larger the Damselfly, the More Likely to Be Threatened: A Sexual Selection Approach. J Insect Conserv 2019, 23, 535–545. [Google Scholar] [CrossRef]
- Attiwilli, S.; Karmakar, T.; Isvaran, K.; Kunte, K. Habitat Preference and Functional Traits Influence Responses of Tropical Butterflies to Varied Habitat Disturbance. Int J Trop Insect Sci 2022, 42, 855–864. [Google Scholar] [CrossRef]
- Eggenberger, H.; Frey, D.; Pellissier, L.; Ghazoul, J.; Fontana, S.; Moretti, M. Urban Bumblebees Are Smaller and More Phenotypically Diverse than Their Rural Counterparts. Journal of Animal Ecology 2019, 88, 1522–1533. [Google Scholar] [CrossRef]
- O’Donnell, M.J. A Perspective on Insect Water Balance. Journal of Experimental Biology 2022, 225, jeb242358. [Google Scholar] [CrossRef]
- Rocha-Ortega, M.; Rodríguez, P.; Córdoba-Aguilar, A. Spatial and Temporal Effects of Land Use Change as Potential Drivers of Odonate Community Composition but Not Species Richness. Biodivers Conserv 2019, 28, 451–466. [Google Scholar] [CrossRef]
- Rocha-Ortega, M.; Rodríguez, P.; Córdoba-Aguilar, A. Can Dragonfly and Damselfly Communities Be Used as Bioindicators of Land Use Intensification? Ecol Indic 2019, 107. [Google Scholar] [CrossRef]
- Heinrich, B. Thermoregulation in Endothermic Insects. Science (1979) 1974, 185, 747–756. [Google Scholar] [CrossRef] [PubMed]
- May, M.L. Odonata: Who They Are and What They Have Done for Us Lately: Classification and Ecosystem Services of Dragonflies. Insects 2019, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.B.; Chumchal, M.M.; Drenner, R.W.; Kennedy, J.H. Seasonality of Odonate-mediated Methylmercury Flux from Permanent and Semipermanent Ponds and Potential Risk to Red-winged Blackbirds (Agelaius Phoeniceus). Environ Toxicol Chem 2017, 36, 2833–2837. [Google Scholar] [CrossRef]
- Córdoba-Aguilar, A.; San Miguel-Rodríguez, M.; Rocha-Ortega, M.; Lanz-Mendoza, H.; Cime-Castillo, J.; Benelli, G. Adult Damselflies as Possible Regulators of Mosquito Populations in Urban Areas. Pest Manag Sci 2021, 77, 4274–4287. [Google Scholar] [CrossRef]
Figure 1.
A) Map of sampling sites in municipality of La Huerta, Jalisco, Mexico; B) State of Jalisco within the Mexican Republic; C) Chamela-Cuixmala Biosphere Reserve. Symbols correspond to sample sites.
Figure 1.
A) Map of sampling sites in municipality of La Huerta, Jalisco, Mexico; B) State of Jalisco within the Mexican Republic; C) Chamela-Cuixmala Biosphere Reserve. Symbols correspond to sample sites.
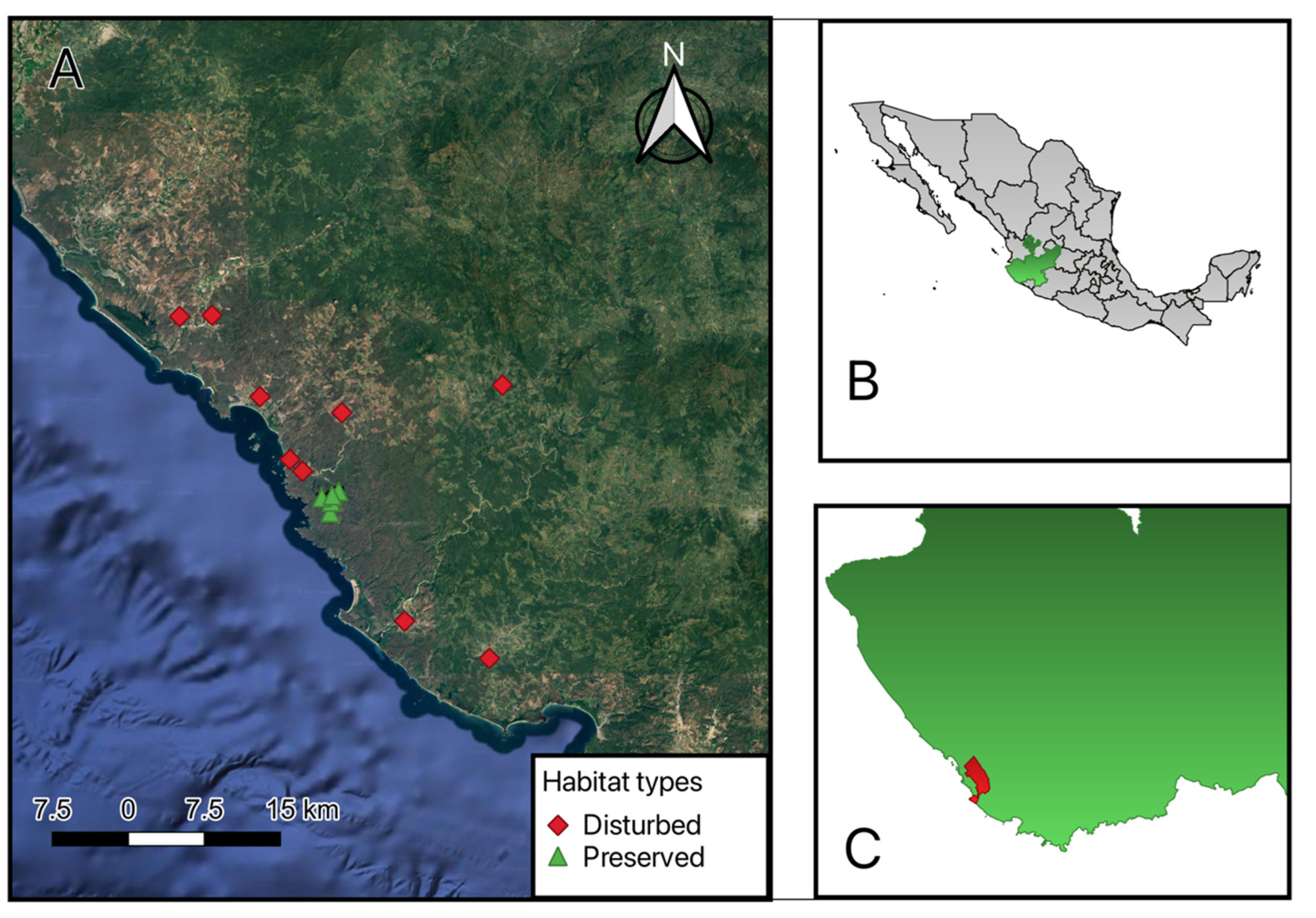
Figure 2.
A) Relationship between thermal stress and maximum environmental temperature in preserved and disturbed sites in 18 species of odonates. B) Relationship between thermal stress and total body size. Shaded areas represent 95% confidence intervals. Maginal (R2m) and conditional (R2c) coefficients of determination are reported to indicate the proportion of variance explained by fixed effects alone and by both fixed and random effects, respectively.
Figure 2.
A) Relationship between thermal stress and maximum environmental temperature in preserved and disturbed sites in 18 species of odonates. B) Relationship between thermal stress and total body size. Shaded areas represent 95% confidence intervals. Maginal (R2m) and conditional (R2c) coefficients of determination are reported to indicate the proportion of variance explained by fixed effects alone and by both fixed and random effects, respectively.
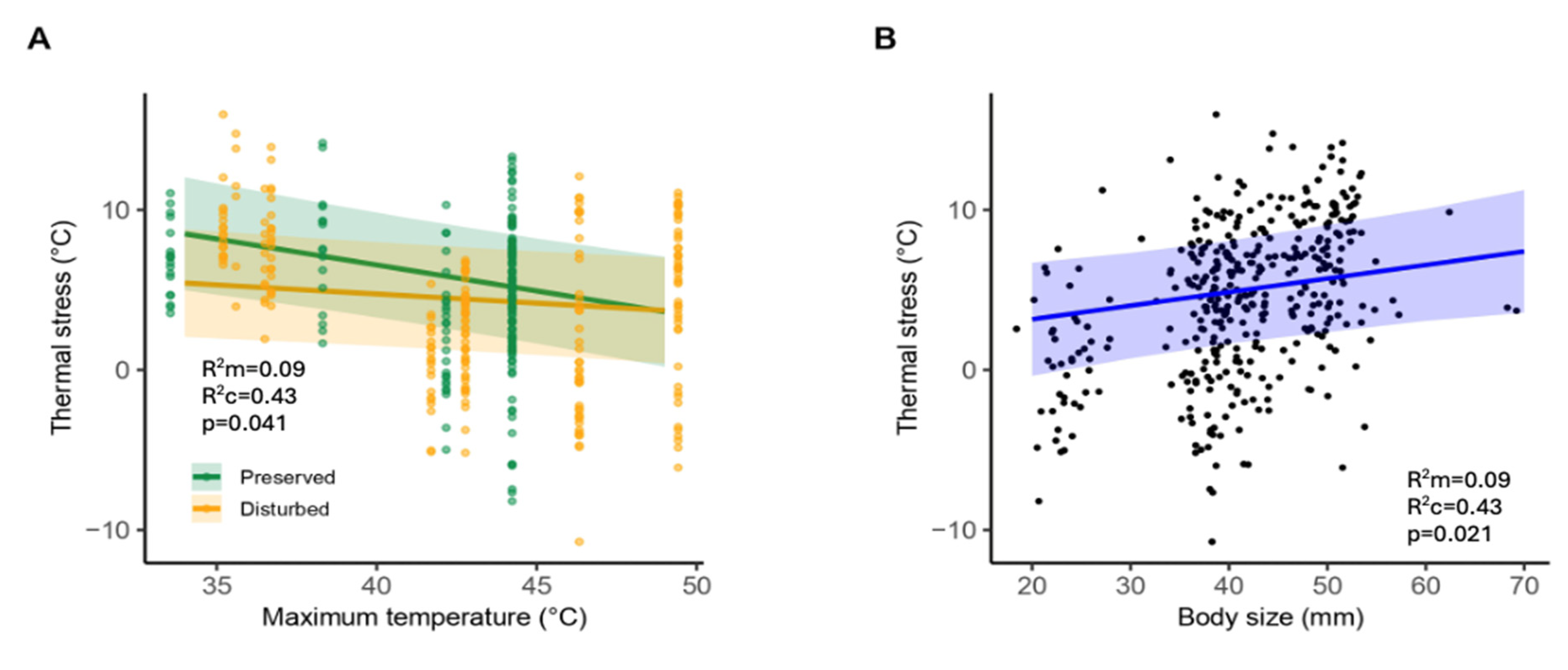
Figure 3.
A) Relationship between protein content (µg/mg) and thermal stress (°C). B) Protein content (µg/mg) in disturbed and preserved sites. Error bars show 95% confidence intervals. C) Relationship between lipid content (µg/mg) and thermal stress (°C). D) Lipid content (µg/mg) in disturbed and preserved sites. Error bars show 95% confidence intervals. E) Relationship between thoracic mass (g) and thermal stress (°C). F) Thoracic mass (g) in disturbed and preserved sites. Error bars show 95% confidence intervals. Predicted protein, lipid, and body size values were back-transformed from the logarithmic scale using the exponential function for biological interpretability and visualization. Maginal (R2m) and conditional (R2c) coefficients of determination are reported to indicate the proportion of variance explained by fixed effects alone and by both fixed and random effects, respectively.
Figure 3.
A) Relationship between protein content (µg/mg) and thermal stress (°C). B) Protein content (µg/mg) in disturbed and preserved sites. Error bars show 95% confidence intervals. C) Relationship between lipid content (µg/mg) and thermal stress (°C). D) Lipid content (µg/mg) in disturbed and preserved sites. Error bars show 95% confidence intervals. E) Relationship between thoracic mass (g) and thermal stress (°C). F) Thoracic mass (g) in disturbed and preserved sites. Error bars show 95% confidence intervals. Predicted protein, lipid, and body size values were back-transformed from the logarithmic scale using the exponential function for biological interpretability and visualization. Maginal (R2m) and conditional (R2c) coefficients of determination are reported to indicate the proportion of variance explained by fixed effects alone and by both fixed and random effects, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



