Submitted:
14 May 2025
Posted:
15 May 2025
You are already at the latest version
Abstract
Infectious plant diseases impair ecosystems, disrupting global food production and risking human existence. Puccinia striiformis f. sp. tritici causes yellow rust disease, impacting wheat production. Puccinia striiformis f. sp. tritici (Pst)-caused yellow rust is one of the most devastating diseases of this crucial global cereal crop. Yellow rust severely impacts wheat production by tremendous yield loss threats to global food security. Yellow pustules on plant leaves evolve into big lesions that interfere with photosynthesis, resulting in stunted growth and decreased grain yield and quality. Yellow rust becomes harder to control because climate change modifies the pathogen's behavior and makes control methods previously used ineffective. Climate change conditions, such as increased temperatures and changed rainfall patterns, in addition to the escalation of extreme weather events, aid yellow rust spread and establishment, making predicting and controlling outbreaks challenging. This chapter discusses how the resistance of the host provides an effective and sustainable way of controlling wheat yellow rust under conditions of a changing climate.
Keywords:
wheat
; yellow rust
; climate change
; host resistance
; management
1. Introduction
Climate change is a global challenge, with the current global temperatures being approximately 1.1°C above the pre-industrial temperature era (WMO, 2022; 2023). This thermal escalation is predominantly driven by manmade greenhouse gas releases and alterations in the land-use will persist provided the net greenhouse gas releases continue (IPCC, 2023). The impacts of climate transformation are already evident, with harmful effects to ecological systems, human populations, and structural systems. Persistent global warming is expected to intensify these dangers and associated uncertainties (IPCC, 2022). Both food supply and food security are particularly vulnerable, facing direct threats from droughts and flooding and indirect challenges from harmful insects and plant diseases (Bezner Kerr et al., 2022). Studies have indicated that climate change has negatively affected agricultural productivity around the world (Ortiz-Bobea et al., 2021). Wheat, a major food crop cultivated on 222 million hectares around the world (USDA, 2023) which is vulnerable and at risk to various diseases as a result of a changing climate. It is well-documented that climatic change patterns influence the occurrence and distribution of pathogens of economic significance (Juroszek and Tiedemann, 2013). Notably, alterations in climate profoundly impacted the epidemiology of wheat rust diseases, creating major obstacles for global wheat production (see Figure 1).
More frequent occurrences of extreme weather events, higher temperatures, and changing precipitation patterns are modifying the intensity, seasonality, and distribution of wheat rust epidemics (Nnadi & Carter, 2021). As much as it may extend the wheat growing season, warmer climates also promote the conditions for the propagation and spread of rust pathogens, leading to increased and intense occurrences (Miedaner & Juroszek, 2021). Moisture regimes-most notably high humidity and changing patterns of rainfall-play a decisive role in germination of spores, infection processes, and aerial dispersal; for example, high humidity enhances rust progression, and varying wind directions can disseminate spores over great distances (Sánchez et al., 2023). Further, climate-based stresses have been found to break down wheat’s natural resistance systems, making crops more susceptible to rust infections (Waheed et al., 2023). Together, these climatic changes favor the survival, growth, and transmission of pathogens, thus escalating the occurrence and severity of wheat rust diseases.
Yellow rust, caused by a fungus called Pst, is a big problem for wheat farmers. It usually shows up in cooler places, like northern areas or higher altitudes, especially where wheat starts to grow when it’s still a bit chilly. Table 1 shows the best conditions for this yellow rust fungus, based on past research (Roelfs et al., 1992). However, recent large-scale epidemics have emerged in warmer wheat-growing regions, driven by the evolution of high-temperature-adapted strains of Pst identified across different regions globally (Hovmøller et al., 2008). The capacity of wheat rusts for long-distance dispersal, rapid virulence development, and adaptation to diverse climates establishes them as a critical global threat to wheat production (Ali and Hodson, 2017).
Considering the global prevalence and economic significance of yellow rust, this review aims to synthesize and update the current knowledge on the disease with climate change. The primary objective is to inform the development of sustainable and effective management approaches, with a particular emphasis on utilizing host resistance to mitigate the challenges arising from climate change.
2. The Global Economic Impact of Wheat and Yellow Rust
Global wheat acreage spans approximately 222 million hectares, making it one of the most significant cereal crops worldwide in terms of both production and consumption (He et al., 2022). In the 2023-24 cropping cycle, global-scale wheat production was 784.91 million metric tons, while consumption totaled 796.44 million metric tons. The top ten wheat-producing countries include China, India, Russia, the United States, France, Canada, Germany, Pakistan, Turkey, and Ukraine. A summary of global production and consumption trends over the past seven years is presented in Table 2. Wheat is a dietary staple and the primary source of plant-derived protein for human consumption, thereby making it integral to global food security. To meet the needs of a forecasted global population size of 9.6 billion by the year 2050, wheat cultivation output must rise by 60%, despite challenges such as constrained availability of land and water and the effects of a changing climate (Savadi, 2018). Under these changing climatic conditions, wheat production is increasingly threatened by a range of biotic and abiotic factors, with the yellow rust pathogen among the most severe biotic challenges.
Yellow rust, also referred to as “Stripe rust” due to the yellow-to-orange coloration of the uredinia which forms characteristic “yellow stripes” on infected leaves (Figure 2), spores freed from the uredinia are capable of reinfecting the same plant, and other surrounding plants within that field, or spreading to host plants situated hundreds of kilometers from the source. Yellow rust has been documented in more than sixty countries on all continents except Antarctica (Chen, 2005). An international survey conducted between 2000 and 2009 on the severity and frequency of yellow rust outbreaks included data from more than 60 countries (Wellings, 2011; Boshoff et al., 2019).
The survey revealed that large-scale yellow rust outbreaks are common two to three times every five years across many wheat-growing regions worldwide, and the disease can cause crop losses of 5% to 10%. About 88% of wheat produced worldwide has been estimated to be threatened by rust diseases, which cause a yearly economic loss of about $1 billion (Wellings, 2011; Beddow, 2015; Schwessinger, 2017). Further, a recent world survey of pathogens and pests causing damage to prominent food crops indicated yellow rust among the major pathogens, responsible for yield losses above 1% globally (Savary et al., 2019).
Major epidemics of yellow rust have occurred during the last twenty years in major wheat-growing areas of the world. Prominent outbreaks have been reported in China (Li & Zeng, 2002), South Asia (Hussain et al., 2004; Duveiller et al., 2007; Aggarwal et al., 2018; Tariq-Khan et al., 2020), the Middle East and Central Asia (Solh et al., 2012), Central and West Asia (Chen & Kang, 2017), Europe (Solh et al., 2012), Africa (Pretorius, 2004; ICARDA, 2011; Solh et al., 2012), Australasia (Murray & Brennan, 2009; Solh et al., 2012), South America (Ochoa et al., 2007; Wellings, 2011), and North America (Line, 2002; Chen, 2005, 2007; Chen & Kang, 2017; Brar et al., 2019).
3. Climate Change and Wheat Yellow Rust
3.1. Emergence of High-Temperature-Adapted Pst Strains and Rust Expansion
Historically, Pst is viewed as a kind of pathogen that can amoeba in mild to cooler climates and thereby restrict its proliferation by increased temperatures (Dennis, 1987). So, its adaptability to warmer conditions increases the chances of its spread into new places considerably (Vidal et al., 2022). Culminating that, the spreading of a single strain of Pst, PstS1, in North America in 2000 and in Australia in 2002, as well as the following spread by a genetically near identical strain PstS2 into Europe and Asia, is said to be one of the fastest and largest dispersals of a crop pathogen in history (Hovmøller et al., 2023). They both have adapted to the causes of increased temperatures and have been mainstream in the southern central region of U. S States since 2000, a period that did not flag the disease as a significant concern (Milus et al., 2009; Chen, 2005). Research, both experimental and field studies, thus, has evidenced that infectious plant diseases may be very responsive to temperature fluctuations (Chaloner et al., 2020), and much evidence exists on their predicted range extension under future climate change scenarios (Lafferty, 2009; Sturrock et al., 2011; Ghelardini et al., 2016; Wyka et al., 2017; Pathak, 2018;).
Traditionally, before 2000, yellow rust was mainly confined in the U.S. to the Pacific Northwest and California. However, after 2000, the disease gradually spread into southern-central states and across the central Great Plains (Chen, 2005). First, this debate must assume that Pst is an important pathogen of cereal crops in the temperate regions, where it was considered to be cool-weather-loving, but with recent initiatives, Pst may have been adapted to heating climates (Milus et al., 2006). The comparison between 1970s isolates and 2003 isolates showed that post-2000 isolates were found to germinate significantly faster and had shorter latent periods at 18 degrees Celsius, while not much difference was seen at 12 degrees Celsius. A recent study has also shown that Pst recovers from heat stress more rapidly than previously thought (Gardner et al., 2023).
Yellow rust has been the dominant wheat disease in Serbia since 2014, a trend linked to increasing winter temperatures—namely, increases to 4.2°C in January and 6.1°C in February compared to the long-term averages of -0.1°C and 1.8°C, respectively, since 1964 (Jevtić et al., 2017). The disease has also been more commonly reported in Northwestern Russia (Gultyaeva et al., 2021), West Siberia (Ivanova et al., 2021), the Volga area, and the Central Chernozem region (Druzhin, 2010; Zeleneva et al., 2022), spreading beyond its previously predominant status in the North Caucasus. These transformations have most probably been brought on by global climate change and its effects on the distribution of the disease. Warrior race wheat yellow rust, which is the most dominant strain in Serbia, is shown to have profuse adaptability across ambient temperatures, thriving in both warmth, such as in Spain, and coolness like Sweden (Jevtić et al., 2017; Hovmøller et al., 2016). These are said to be categorized under the thermal generalists, showing a peak in infection efficiency at 10°C and 15°C (de Vallavielle-Pope et al., 2018). Before 2004 in France, the Pst population adapted to local temperature variations, resulting in notable spatial differences in the interactions between pathotype and temperature that influenced urediniospore germination and infection efficiency, especially between the southern and northern regions of France (Mboup et al., 2012). Similar adaptations to temperature ranges have been observed in Pst isolates from the US and Australia (Loladze et al., 2014; Sharma-Poudyal et al., 2014). Recently, Pst has demonstrated diminished importance in the wheat growing seasons throughout the Nordic and Baltic areas (Strandberg et al., 2024).
Historically, wheat yellow rust was regarded as a minor concern along the eastern shores of Zhejiang and Jiangsu Provinces in China, with its progression being slow and often interrupted by early April. However, by early May in 2019, the severity of the disease had not increased, probably due to the warmer climatic conditions favoring the high-temperature-adapted isolates (Ju et al., 2022; Zhao & Kang, 2023). Since 2012, there have been some scattered incidences of different Pst pathotypes in China, where 126 Pst isolates from 12 provinces have exhibited high-temperature tolerance. This pathogen population has shown significant adaptation to high temperature levels, thriving well at temperatures above the critical 23°C threshold (Zhang et al., 2013).
The dissemination of rust pathogens has grown more alarming, with reports indicating that Pst has extended to South Africa, a phenomenon most likely associated with changes in rainfall patterns (Boshoff et al., 2002). To this, both stem rust and yellow rust are now threatening to set up homes in previously uninfected regions as milder winters enable these pathogens to make it through the harsher periods (Ma et al., 2015; Novotná et al., 2017; Prank et al., 2019). This indicates an increasing susceptibility of wheat crops while climate conditions keep evolving (Juroszek et al., 2020). Predictions for 2050 suggest that climate change may significantly redefine major wheat-producing areas, allowing rust fungi to grow and expand into new regions (Ortiz et al., 2008)..
In north of India, the current epidemic of yellow rust in Punjab has been associated with the break of the new pathotype (78S84), which can infect wheat at elevated temperatures. The disease initially appeared during late December, as a result of favorable climatic conditions and rising temperatures (Prashar et al., 2007; Jindal et al., 2012). Severe weather occurrences, including heatwaves, have also been speculated to prefer particular Pst lineages, making pathogen populations change temperature optima (Strandberg et al., 2024).
3.2. Pathogen Survival, Reproduction, and Increased Aggressiveness
Rain and temperature are critical environmental elements that impact Pst germination, infection, and survival (Lyon & Broders, 2017). Increased winter temperature is known to support overwinter survival of pathogens in the growing season (Gladders et al., 2007). In Canada, rust pathogens do not generally survive the winter; however, moderate winters will allow for winter survival and/or the advanced initiation of rust and perhaps enhanced severity of disease (Boland et al., 2004). In a similar vein, enhanced off-season rainfall can contribute to more suitable conditions for the persistence of pathogens during summer in some parts of Australia. Temperature’s effect on Pst’s reproductive cycle is well established, with both sexual and asexual phases being sensitive to temperature. Research has established that telia and teliospore production is enhanced under high-temperature conditions (McDonald & Linde, 2002; Chen et al., 2021).
Aggressiveness, one of the measurable elements of pathogenicity, is the level of injury caused to host plants (Andrivon et al., 2007; Pariaud et al., 2009). It is an important biological characteristic that indicates the degree to which a pathogen can develop, become established, and reproduce in a host plant. It covers several infection-related traits associated with the fitness of the pathogen as a whole, such as how well it infects, when it will produce symptoms (latency period), how quickly it produces spores, how long it can continue to infect, and the lesion size it produces (Pariaud et al., 2009). Due to this, aggressiveness is central in the constant evolutionary struggle between hosts and pathogens. Scientists can measure this characteristic at different developmental stages of the plant, both during seedling development (Milus et al., 2006; de Vallavieille-Pope et al., 2018) and in adult plants (Pariaud et al., 2009b; Azzimonti et al., 2013).
Temperature is crucial in the way pathogen evolution occurs, especially in determining their capacity to adapt to different temperature environments (Zhan & McDonald, 2011; Yang et al., 2016). It also determines how virulent these pathogens can be (Schade et al., 2014). The relationship between temperature and virulence is well established, particularly with wheat yellow rust. Warmer temperatures have been found to enhance the virulence of Pst, highlighting the fungus’s ability to adapt to increasing temperatures (Mboup et al., 2012). New and highly virulent Pst pathotypes, with wider virulence and greater adaptation to higher temperatures, have appeared in recent years (Milus et al., 2009; Walter et al., 2016). For example, in the eastern United States, Pst populations are still adapting under conditions of warming, demonstrating enhanced fitness and virulence, even when certain resistance genes are present (Lyon & Broders, 2017). Likewise, Latvia has indicated the appearance of more virulent Pst races like ‘Warrior,’ ‘Kranich,’ and ‘Triticale’ (Feodorova Fedotova & Bankina, 2018).
Some Pst strains, especially those in the PstS7 lineage, are considered thermal generalists with high performance at a wide temperature regimen-instead of being thermal specialists like older isolates (de Vallavieille-Pope et al., 2018). In Serbia, there was a remarkable rise in the incidence of highly pathogenic Pst races in 2023 (Župunski et al., 2024).
In China, the dissemination of G22-virulent pathotypes has proceeded rapidly, their spread to new areas being due to boosted infectivity and enhanced virulence (Liu et al., 2012; Li et al., 2016; Wang et al., 2017). Among the most prevalent and virulent races detected in major wheat-producing regions are G22-9 (also referred to as CYR34), CYR32, and CYR33, whose occurrence has been reported several times in major regions (Liu et al., 2017; Bai et al., 2018; Huang et al., 2020). Moreover, the Su11 race group remains dominant in Xinjiang, showing excellent fitness and good adaptability in a wide range of wheat cultivars and environments (Ma et al., 2023).
New five Pst races have been identified by recent reports from India, including 46S117, 110S119, 238S119, 110S247, and 110S84, of which the most widespread and virulent is race 110S119, characterized by its high rate of growth (Gangwar et al., 2016). Research conducted to analyze spore germination at different temperatures, 5°C, 10°C, 15°C, and 20°C, has found that the germination reached a peak at 15°C, while at other temperature levels, germination sharply decreased (Anand et al., 2023).
It was recently discovered that Pst aggressiveness in the Middle East and Mediterranean is temperature-dependent, with no variation between isolates from low- and high-altitude origins (El Amil et al., 2022). Isolates LB6 from Lebanon, TK3 from Turkey, and SY2 from Syria showed high aggressiveness at high temperatures regardless of their native altitudinal origins (El Amil et al., 2022).
There has been a sharp rise in aggressiveness of yellow rust in Egypt in recent times, with the disease spreading to all parts of the country, including areas of hotter and drier climates like Upper Egypt (Esmail et al., 2023). Between 2018 and 2020, 42 Pst races were reported, with six novel aggressive strains (72E8, 135E16, 151E80, 160E173, 224E191and 238E143) emerging for the first time (Esmail et al., 2021).
3.3. New Strains and Pathotypes
Rising temperatures have facilitated the development of more virulent Pst races globally, significantly affecting wheat crop productivity (Milus et al., 2009). Earlier to2000, virulence and molecular investigations regrading isolates from major wheat growing regions of Europe, Australia, and North America consistently documented clonal Pst populations, characterized by closely related pathotypes and low genetic diversity, primarily driven through single-step mutations (Steele et al., 2001; Hovmøller et al., 2002; Chen, 2005; Enjalbert et al., 2005; Chen et al., 2010; Ali et al., 2014a; Hubbard et al., 2015; Hovmøller et al.,2016). However, in the past two decades, novel virulence profiles have notably emerged, and more aggressive strains have emerged worldwide. Notably, the PstS1 and PstS2 strains emerged rapidly throughout the U.S. Chen et al., 2002; Markell and Milus, 2008), Australia (Wellings, 2007), and Europe (Hovmøller and Justesen, 2007) within a short timeframe of three years at the beginning of the 2000s. These strains are distinguished by their adaptation to high temperatures, which is atypical for this pathogen (Markell and Milus, 2008; Milus et al., 2006, 2009).
The appearance of Pst pathotypes PstS1 and PstS2 has allowed the yellow rust pathogen to become established in areas previously deemed unsuitable for its growth (Milus et al., 2006). These pathotypes have also greatly widened their host range and geographic limits, now being a serious threat to wheat cultivation across the US and regions in the African and Australian continents (Bahri et al., 2009; Wellings, 2011; Chen et al., 2014; Walter et al., 2016). Another analysis (Yahyaoui et al., 2002), considering Pst populations in Syria and Lebanon between the years 1994 and 1999, showed an abundant pathotypic diversity, where 25 pathotypes were found to exist in Syria and 11 in Lebanon. Various new versions also appeared throughout this time with virulence attributes like v2, v6, v7, v8, and v9-signifiers later ascribed to the PstS2 pathotype (Hovmøller et al., 2011). PstS2 first emerged in the East African region during the early 1980s and is now among the most global and long-standing yellow rust pathotypes (Ali et al., 2014a; Walter et al., 2016).
During the period 2005-2006, 268 Pst isolates from 12 countries located in the western and eastern Mediterranean regions were studied. This analysis resulted in the characterization of twelve pathotypes, with the interesting “PstS2-v27” among them (Bahri et al., 2009). A point of note was that eight pathotypes occurred solely in the eastern Mediterranean, emphasizing the rich genetic diversity of the region and its status as a hotspot for new Pst pathotypes to emerge.
A set of 268 Pst isolates gathered from 12 nations in the western and eastern Mediterranean were tested between 2005 and 2006, leading to the detection of twelve different pathotypes, such as the significant “PstS2-v27” (Bahri et al., 2009). Eight of these 12 pathotypes were confined to the eastern Mediterranean region, highlighting high Pst diversity of the region and its role as a key region for the origin of novel strains. One Israeli sample contained virulence genes (i.e v2, v6, v7, v8, v9, v25, and v27), corresponding to the PstS2-v27 pathotype.
New variants and pathotypes of PstS1/PstS2 have been established in North African and West Asian regions, acquiring additional virulence to Yr resistance genes, including Yr1, Yr3, Yr10, and Yr27, few of these are extensively used in wheat breeding programs (Singh et al., 2004; El Amil, 2015). Current variants of PstS1/PstS2 include strains such as PstS1/2b (North America, Australia), PstS1/2-v1 (East Africa), PstS1/2-v3 (East Africa), PstS1/2-v27 (East Africa, West Asia, North Africa), along with several other combinations involving v1, v3, v10, and v27 across various regions (GRRC, 2022). These emerging pathotypes highlight the importance of the ongoing adaptation of Pst to changing environmental conditions and its increasing influence on wheat production around the world.
3.4. Regional Impact of Yellow Rust
Reports indicate that yellow rust epidemics have originated in distant areas, appearing either in regions where they were previously absent or reappearing in more virulent forms. (Bahri et al. 2009; Hovmøller et al. 2023b). The emergence of new strains such as PstS1 and PstS2 has enabled Pst to spread into warmer regions previously considered unsuitable for the progression of the disease (Milus et al., 2006). In recent decades, numerous yellow rust epidemics have occurred globally, with significant impacts on wheat production. In North America, yellow rust has increasingly become a significant issue in wheat production (Chen and Kang 2017). From 2000 to 2007, yellow rust was reported annually in at least 15 U.S. states, resulting in estimated yield losses surpassing 6.5 million tonnes (Chen et al., 2010). In the 2010 epidemic, however, yield losses were estimated at 2.2 million tonnes, with additional fungicide costs totaling around $30 million solely in Washington State (X.M. Chen, pers. comm.). Although yellow rust posed a lesser threat in the Great Plains in 2011, largely due to extensive drought conditions, the Pacific Northwest had a severe impact than in 2010. Yield losses in susceptible varieties were projected to surpass 70%, according to data from experimental plots and growth stage assessments. In the 2012 growing season, yield reductions in highly susceptible wheat varieties were forecasted to reach 50%. In Canada, the most widespread and severe outbreaks were reported in western Canada during 2010 and 2011 (Brar et al. 2019). In South America, Yellow rust has posed a substantial challenge in several countries (Wellings 2011). While Pst occurred sporadically in Uruguay from its initial detection in 1929 until 2016, it has since caused widespread epidemics in Uruguay and Argentina starting in 2017 (Carmona and Sautua 2018; German et al. 2018, 2021).
Europe is well-known for its extensive research on yellow rust and its recurring epidemics, particularly in the northwestern region. The Warrior/Ambition race was first identified during the 2009/2010 season in the UK, Germany, Denmark, France, and Scandinavia, and into Spain, leading to widespread epidemics and severe crop damage (Vergara-Diaz et al., 2015; Hovmøller et al., 2016). The Kranish race, identified in 2011, contributed to disease outbreaks across many European countries (Solh et al. 2012; Gomes et al., 2018). In Portugal, Pst had been absent for two decades but has since become a major threat to wheat and triticale production beginning in 2013 (Gomes et al., 2018). Italy has experienced at least three significant outbreaks of yellow rust over the past decade (M. Maccaferri, personal communication). In 2019, severe yellow rust epidemics were reported across several European regions (Anonymous 2019). Although less frequent in Eastern Europe, yellow rust has caused notable damage in countries. In Serbia, yellow rust predominated over leaf rust following a shift in climate conditions in 2014, with disease severity reaching up to 90% in field trials (Jevtić et al., 2017; Župunski et al., 2016).
Australian farmers spent an estimated $40-90 million annually between 2003 and 2006 on fungicides (Wellings 2007) to manage yellow rust. Pathotypes carrying virulences v17 and v27 were identified as serious threats to wheat-producing regions in Australia. Despite recurring outbreaks and occasional appearance of nonnative pathotypes, Australia’s countrywide breeding program focused on rust resistance has been largely effective in mitigating the most severe effects of yellow rust epidemics.
In Africa, yellow rust remains a significant problem in many countries located in the northeast, northwest, and South Africa, and in the south. A major outbreak occurred in Morocco in 2009, and since 2010, the disease has spread widely throughout the East African region, causing considerable financial losses, particularly in low-input subsistence farming systems (Singh et al., 2016). The 2010 outbreak in Ethiopia caused production losses ranging from 67-100% on commercial wheat varieties, costing over $3.2 million on fungicide treatments (ICARDA 2011; Solh et al. 2012).
Facultative winter wheat in the CWANA (Central and West Asia and North Africa) region of southern Kazakhstan and Uzbekistan has regularly experienced yellow rust outbreaks, with particularly intense outbreaks during 2009 and 2010. The loss of resistance to Yr27 in wheat cultivars such as PBW343(India), Inquilab 91(Pakistan), and Chamran (Iran)was documented from 2002 and 2004. While yellow rust epidemics were sporadic in certain zones, adverse climatic conditions likely limited the speedy increase of pathotypes carrying v27virlence until 2009, when favorable climate led to widespread outbreaks in at least 11 countries in the CWANA region. These favorable conditions persisted into 2010, characterized by less severe winters and sufficient rainfall across CWANA countries, leading to early yellow rust outbreaks. The 2010 yellow rust pandemic led to significant yield losses throughout key wheat-producing regions in CWANA and the Caucasus, with Syria experiencing particularly severe losses due to the widespread cultivation of Cham 8 (carrying Yr27), which covered more than 70% of the wheat acreage. Even with conducive conditions in 2011 and 2012, significant yellow rust outbreaks did not materialize, highlighting the interannual variation of crop diseases and their effects. Key varieties were susceptible in the major wheat-producing countries of the Central Asian and Middle Eastern region. A catastrophic epidemic broke out in Turkey’s Central Plateau, where Gerek 79, a susceptible cultivar, was the most common. Identified in East Africa as early as the 1980s (Walter et al., 2016), the closely related clonal lineages PstS1 and PstS2 have driven significant epidemics around the world from the year 2000 onward (Hovmøller et al., 2011).
In China, multiple large-scale yellow rust outbreaks have been documented (Li and Zeng 2002), including a severe outbreak in 2002 that affected 6.6 million hectares across 11 provinces (Chen 2020). In South Asia, yellow rust is a significant challenge to wheat farming in India, Pakistan, Nepal, and southeastern Afghanistan.
Yellow rust has grown to become more widespread in the cooler northern parts of India, infesting around 10 million hectares of wheat (Bhardwaj et al., 2019). Punjab suffered from a major yellow rust epidemic in 2008, and the disease persisted to spread over different regions of the country throughout the 2010–2011 wheat growing seasons (Aggarwal et al., 2018). A similar epidemic had already been seen in Pakistan in 2005. Yellow rust epidemics of different magnitudes were also noted in several countries in Central and West Asia in 2010 (Rahmatov et al., 2012; Chen & Kang, 2017), with especially severe instances in Syria and Lebanon (El Amil, 2015). Turkey was also severely affected, with substantial epidemics in 2007, 2009, and 2010 that resulted in huge wheat yield losses (Solh et al., 2012).
3.5. Effects of Elevated GHGs and Abiotic Factors on Host-Pathogen Dynamics
Increased global temperatures, elevated levels of atmospheric carbon dioxide, and the occurrence of more intense and frequent weather events such as droughts and floods have the potential to greatly affect how plants defend against disease caused by pathogens (Dossa et al., 2015). These climatic changes can also have the potential to change the dynamics of plants and pathogens, how and where disease occurs, and how disease range increases (Chakraborty, 2005; Burdon et al., 2006; Garrett et al., 2006; Crowl et al., 2008; Eastburn et al., 2011).
Between 2016 and 2019, conditions of severe winter temperatures exposed discernible variation in the susceptibility of wheat genotypes to obligate pathogens, with implications for the disease-susceptible role of climate (Jevtić et al., 2020). More extensive climate change patterns, including increased temperatures, elevated atmospheric CO₂ concentrations, and enhanced frequency of extreme weather events such as flooding and drought, also have the potential to impact the ability of plants to resist pathogens (Dossa et al., 2015). These climatic shifts are expected to alter host-pathogen interactions and shape the temporal and spatial dynamics of disease outbreaks (Chakraborty, 2005; Burdon et al., 2006; Garrett et al., 2006; Crowl et al., 2008; Eastburn et al., 2011). Worldwide epidemic spreads of rust diseases under regions having elevated levels of ambient ozone (O₃) levels confirm prospective interactions of the mentioned factors and justify parallel breeding efforts focusing on increasing resistance against rusts, as well as tolerance towards ozone, in wheat (Mashaheet et al., 2020). Ethiopian studies proved that wheat lines showing yellow rust resistance had a greater production under conditions of drought than vulnerable varieties (Abro et al., 2017). Moreover, differences in bread wheat response to heat stress and susceptibility to yellow rust have been noted in Egypt, which means that it is feasible to breed for heat stress tolerance in combination with rust resistance without negatively impacting yield (Megahed et al., 2022.
4. Host Resistance as a Primary Control Strategy
The warming global climate has created conditions increasingly conducive to the establishment and spread of the wheat yellow rust pathogen (Chakraborty et al., 2010). While various control strategies exist, among these, developing and promoting resistant wheat varieties remains the most efficient and sustainable method from both a cost and environmental perspective (Chen, 2013). Wheat resistance to yellow rust is of two categories: All-Stage Resistance (ASR) and Adult-Plant Resistance (APR). To date, over 300 resistance-associated specific genes or associated trait loci (QTL) have been identified, with 86 genes (Yr1–Yr86) officially named (Zhu et al., 2023).
ASR genes, including Yr1, Yr2, Yr3a/b/c, Yr4a/b, Yr6, Yr7, Yr9, Yr10, Yr17, Yr24/Yr26, and Yr27, were extensively utilized in breeding programs worldwide. However, ASR genes are highly vulnerable to becoming compromised by the rapid adoption of Pst populations (Hulbert and Pumphrey, 2014). In many regions, the development of virulent Pst pathotypes has rendered most ASR genes ineffective (Wang and Chen, 2017; Chen, 2020). Conversely, APR genes, which generally confer partial resistance, have demonstrated greater durability compared to ASR genes. A potential solution to counteract the loss of rust resistance is the deployment of APR genes, active at the later growth stages and providing broader, more durable resistance (Brown, 2015; Niks et al., 2015). Among APR types, high-temperature-triggered resistance in mature plants (HTAP) is particularly significant. HTAP resistance is durable with non-race-specificity, and the resistance levels increase as plants mature and environmental temperatures rise (Chen, 2005, 2013). Wheat varieties carrying HTAP resistance display susceptibility to Pst at the juvenile plant stage at low temperatures during the adult-plant stage. However, they exhibit strong resistance at higher temperatures during the post-seeding adult stage (Chen, 2007, 2013).
Currently, twenty-five Yr resistance genes along with multiple QTLs have been identified as associated with APR. Among these genes, Yr18, Yr36, Yr52, Yr59, Yr62, Yr78, and Yr79 are reported to be effective in conferring HTAP resistance (Santra et al., 2008; Fu et al., 2009; Krattinger et al., 2009; Ren et al., 2012; Lu et al., 2014; Zhou et al., 2014; Dong et al., 2017; Wang and Chen, 2017; Feng et al., 2018; Liu et al., 2018, 2019, 2020; Mu et al., 2020). Rising global temperatures are expected to impact wheat growth stages, accelerate leaf senescence, and potentially disrupt yellow rust development. However, the pathogen itself has demonstrated an ability to adapt to higher temperatures, emphasizing the importance of incorporating HTAP resistance genes into wheat breeding programs to enhance resilience (Chakraborty and Newton, 2011; Asseng et al., 2011; Juroszek and von Tiedemann, 2013; Khanfri et al., 2018). The combination of all-stage resistance (ASR) and high-temperature adult-plant (HTAP) resistance presents a compelling method of acquiring durable, long-term protection against yellow rust in wheat, a more critical objective as climate change continues to compromise crop defense mechanisms (Bariana et al., 2022).
5. Conclusion and Way Forward
The increasing number of yellow rust epidemics, induced by an airborne fungal pathogen, emphasizes the imperative for new and sustainable control measures to safeguard world wheat production and global food security. This yield-limiting disease causes serious damage and is becoming increasingly difficult to control as climate change modifies pathogen behavior and diminishes the effectiveness of conventional control measures. A holistic solution is needed to solve this multifaceted problem. One of the most powerful and enduring solutions is to build host resistance by developing rust-resistant wheat varieties with a diverse array of resistance genes. Moreover, incorporating climate-adaptive disease management practices, such as enhancing forecasting systems and synchronizing breeding programs with climate forecasts, will be critical to keeping up with the changing patterns of disease caused by global warming.
Developing robust disease surveillance systems and employing sophisticated diagnostic technologies are key to detecting outbreaks early and acting swiftly to new threats. Adopting sustainable agricultural practices, such as crop rotation and effective irrigation, can also serve to contain the spread of pathogens while enhancing the resilience of agricultural systems. Encouraging collaboration between disciplines and establishing healthy policy frameworks are as critical as bridging knowledge gaps, boosting innovation, and implementing control approaches efficiently. Putting these efforts as priorities will help stakeholders make their wheat production systems more resilient, as well as play a significant role in global food security.
References
- Abou-Zeid, M. A., & Mourad, A. M. (2021). Genomic regions associated with yellow rust resistance against the Egyptian race revealed by genome-wide association study. BMC Plant Biology, 21, 1-14. [CrossRef]
- Abro, Z. A., Jaleta, M., & Qaim, M. (2017). Yield effects of rust-resistant wheat varieties in Ethiopia. Food security, 9(6), 1343-1357. [CrossRef]
- Aggarwal, R., Kulshreshtha, D., Sharma, S., Singh, V. K., Manjunatha, C., Bhardwaj, S. C., & Saharan, M. S. (2018). Molecular characterization of Indian pathotypes of Puccinia striiformis f. sp. tritici and multigene phylogenetic analysis to establish inter-and intraspecific relationships. Genetics and Molecular Biology, 41, 834-842.
- Ali, S., & Hodson, D. (2017). Wheat rust surveillance: field disease scoring and sample collection for phenotyping and molecular genotyping. Wheat Rust Diseases: Methods and Protocols, 3-11.
- Ali, S., Gladieux, P., Leconte, M., Gautier, A., Justesen, A. F., Hovmøller, M. S., & de Vallavieille-Pope, C. (2014). Origin, migration routes and worldwide population genetic structure of the wheat yellow rust pathogen Puccinia striiformis f. sp. tritici. PLoS pathogens, 10(1), e1003903. [CrossRef]
- Ali, S., Rodriguez-Algaba, J., Thach, T., Sørensen, C. K., Hansen, J. G., Lassen, P., & Hovmøller, M. S. (2017). Yellow rust epidemics worldwide were caused by pathogen races from divergent genetic lineages. Frontiers in Plant Science, 8, 1057. [CrossRef]
- Anand, S., Sandhu, S. K., & Tak, P. S. (2023). Effect of abiotic factors on pathotypes causing yellow and brown rust in wheat. Journal of Agrometeorology, 25(3), 462-465. [CrossRef]
- Andrivon, D., Pilet, F., Montarry, J., Hafidi, M., Corbière, R., Achbani, E. H., & Ellisseche, D. (2007). Adaptation of Phytophthora infestans to partial resistance in potato: evidence from French and Moroccan populations. Phytopathology, 97(3), 338-343.
- Anonymous (2019) Rust Watch Report 34 PP.
- Asseng, S., Foster, I. A. N., & Turner, N. C. (2011). The impact of temperature variability on wheat yields. Global change biology, 17(2), 997-1012. [CrossRef]
- Azzimonti, G., Lannou, C., Sache, I., & Goyeau, H. (2013). Components of quantitative resistance to leaf rust in wheat cultivars: diversity, variability and specificity. Plant Pathology, 62(5), 970-981. [CrossRef]
- Bahri, B., Leconte, M., Ouffroukh, A., DE VALLAVIEILLE-POPE, C., & Enjalbert, J. (2009). Geographic limits of a clonal population of wheat yellow rust in the Mediterranean region. Molecular Ecology, 18(20), 4165-4179. [CrossRef]
- Bariana, H., Kant, L., Qureshi, N., Forrest, K., Miah, H., & Bansal, U. (2022). Identification and characterisation of yellow rust resistance genes Yr66 and Yr67 in wheat cultivar VL Gehun 892. Agronomy, 12(2), 318.
- Beddow, J. M., Pardey, P. G., Chai, Y., Hurley, T. M., Kriticos, D. J., Braun, H. J., & Yonow, T. (2015). Research investment implications of shifts in the global geography of wheat Yellow rust. Nature Plants, 1(10), 1-5. [CrossRef]
- Beresford, R. M. (1982). Yellow rust (Puccinia striiformis), a new disease of wheat in New Zealand. Cereal Rusts Bulletin, 10, 35-41.
- Bezner Kerr R et al 2022 Food, Fibre, and Other Ecosystem Products Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change ed H-O Pörtner et al(Cambridge University Press) pp 713–906.
- Bhardwaj, Subhash C., Gyanendra P. Singh, Om P. Gangwar, Pramod Prasad, and Subodh Kumar. “Status of wheat rust research and progress in rust management-Indian context.” Agronomy 9, no. 12 (2019): 892. [CrossRef]
- Boland, G. J., Melzer, M. S., Hopkin, A., Higgins, V., & Nassuth, A. (2004). Climate change and plant diseases in Ontario. Canadian Journal of Plant Pathology, 26(3), 335-350. [CrossRef]
- Boshoff, W. H. P., Pretorius, Z. A., & Van Niekerk, B. D. (2002). Establishment, distribution, and pathogenicity of Puccinia striiformis f. sp. tritici in South Africa. Plant Disease, 86(5), 485-492.
- Boshoff, W. H. P., Visser, B., Lewis, C. M., Adams, T. M., Saunders, D. G. O., Terefe, T., & Pretorius, Z. A. (2020). First report of Puccinia striiformis f. sp. tritici, causing yellow rust of wheat, in Zimbabwe. Plant Disease, 104(1), 290-290.
- Brar, G. S., Fetch, T., McCallum, B. D., Hucl, P. J., & Kutcher, H. R. (2019). Virulence dynamics and breeding for resistance to Yellow, stem, and leaf rust in Canada since 2000. Plant disease, 103(12), 2981-2995.
- Brown, J. K. (2015). Durable resistance of crops to disease: a Darwinian perspective. Annual review of phytopathology, 53(1), 513-539. [CrossRef]
- Brown, J. K., & Hovmøller, M. S. (2002). Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science, 297(5581), 537-541. [CrossRef]
- Burdon, J. J., Thrall, P. H., & Ericson, A. L. (2006). The current and future dynamics of disease in plant communities. Annu. Rev. Phytopathol., 44(1), 19-39. [CrossRef]
- Chakraborty, S. (2005). Potential impact of climate change on plant-pathogen interactions: Presented as a Keynote Address at the 15th Biennial Conference of the Australasian Plant Pathology Society, 26–29 September 2005, Geelong. Australasian Plant Pathology, 34(4), 443-448.
- Chakraborty, S., & Datta, S. (2003). How will plant pathogens adapt to host plant resistance at elevated CO2 under a changing climate?. New Phytologist, 159(3), 733-742. [CrossRef]
- Chakraborty, S., & Newton, A. C. (2011). Climate change, plant diseases and food security: an overview. Plant pathology, 60(1), 2-14. [CrossRef]
- Chakraborty, S., Luck, J., Hollaway, G., Fitzgerald, G., & White, N. (2011). Rust-proofing wheat for a changing climate. Euphytica, 179, 19-32. [CrossRef]
- Chakraborty, S., Luck, J., Hollaway, G., Fitzgerald, G., & White, N. (2011). Rust-proofing wheat for a changing climate. Euphytica, 179, 19-32. [CrossRef]
- Chakraborty, S., Tiedemann, A. V., & Teng, P. S. (2000). Climate change: potential impact on plant diseases. Environmental pollution, 108(3), 317-326. [CrossRef]
- Chaloner, T. M., Gurr, S. J., & Bebber, D. P. (2020). Geometry and evolution of the ecological niche in plant-associated microbes. Nature Communications, 11(1), 2955. [CrossRef]
- Chen, F., Duan, G. H., Li, D. L., & Zhan, J. (2017). Host resistance and temperature-dependent evolution of aggressiveness in the plant pathogen Zymoseptoria tritici. Frontiers in Microbiology, 8, 1217. [CrossRef]
- Chen, W., Wellings, C., Chen, X., Kang, Z., & Liu, T. (2014). Wheat Yellow (yellow) rust caused by Puccinia striiformis f. sp. tritici. Molecular plant pathology, 15(5), 433-446.
- Chen, W., Zhang, Z., Chen, X., Meng, Y., Huang, L., Kang, Z., & Zhao, J. (2021). Field production, germinability, and survival of Puccinia striiformis f. sp. tritici teliospores in China. Plant Disease, 105(8), 2122-2128.
- Chen, X. (2020). Pathogens which threaten food security: Puccinia striiformis, the wheat yellow rust pathogen. Food Security, 12(2), 239-251. [CrossRef]
- Chen, X. M. (2005). Epidemiology and control of yellow rust [Puccinia striiformis f. sp. tritici] on wheat. Canadian journal of plant pathology, 27(3), 314-337.
- Chen, X. M. (2007). Challenges and solutions for yellow rust control in the United States. Australian Journal of Agricultural Research, 58(6), 648-655.
- Chen, X. M. (2007). Challenges and solutions for yellow rust control in the United States. Australian Journal of Agricultural Research, 58(6), 648-655. [CrossRef]
- Chen, X. M., & Kang, Z. S. (Eds.). (2017). Yellow Rust (719 pages). Dordrecht: Springer.
- Chen, X., Wang, M., Wan, A., Bai, Q., Li, M., López, P. F., & Abdelrhim, A. S. (2021). Virulence characterization of Puccinia striiformis f. sp. tritici collections from six countries in 2013 to 2020. Canadian Journal of Plant Pathology, 43(sup2), S308-S322.
- Chen, X.M. (2013) Review article: High-temperature adult-plant resistance, key for sustainable control of yellow rust. Am. J. Plant Sci. 4, 608–627.
- Clifford, B. C., & Harris, R. G. (1981). Controlled environment studies of the epidemic potential of Puccinia recondita f. sp. tritici on wheat in Britain. Transactions of the British mycological Society, 77(2), 351-358. [CrossRef]
- Coakley, S. M. (1979). Climate variability in the Pacific Northwest and its effect on yellow rust disease of winter wheat. Climatic Change, 2(1), 33-51. [CrossRef]
- Coakley, S. M., Scherm, H., & Chakraborty, S. (1999). Climate change and plant disease management. Annual review of phytopathology, 37(1), 399-426. [CrossRef]
- Crowl, T. A., Crist, T. O., Parmenter, R. R., Belovsky, G., & Lugo, A. E. (2008). The spread of invasive species and infectious disease as drivers of ecosystem change. Frontiers in Ecology and the Environment, 6(5), 238-246.
- de Vallavieille-Pope, C., Ali, S., Leconte, M., Enjalbert, J., Delos, M., & Rouzet, J. (2012). Virulence dynamics and regional structuring of Puccinia striiformis f. sp. tritici in France between 1984 and 2009. Plant Disease, 96(1), 131-140.
- de Vallavieille-Pope, C., Bahri, B., Leconte, M., Zurfluh, O., Belaid, Y., Maghrebi, E., & Bancal, M. O. (2018). Thermal generalist behaviour of invasive Puccinia striiformis f. sp. tritici strains under current and future climate conditions. Plant Pathology, 67(6), 1307-1320.
- Dean, R., Van Kan, J. A., Pretorius, Z. A., Hammond-Kosack, K. E., Di Pietro, A., Spanu, P. D., & Foster, G. D. (2012). The Top 10 fungal pathogens in molecular plant pathology. Molecular plant pathology, 13(4), 414-430.
- Dennis, J. I. (1987). Effect of high temperatures on survival and development of Puccinia striiformis on wheat. Transactions of the British Mycological Society, 88(1), 91-96. [CrossRef]
- Dennis, J.I. (1987) Temperature and wet-period conditions for infection by Puccinia striiformis f. sp. tritici race 104e137a+. Transactions of the British Mycological Society, 88: 119-121.
- Donald, F., Green, S., Searle, K., Cunniffe, N. J., & Purse, B. V. (2020). Small scale variability in soil moisture drives infection of vulnerable juniper populations by invasive forest pathogen. Forest Ecology and Management, 473, 118324. [CrossRef]
- Dong, Z., Hegarty, J. M., Zhang, J., Zhang, W., Chao, S., Chen, X., & Dubcovsky, J. (2017). Validation and characterization of a QTL for adult plant resistance to yellow rust on wheat chromosome arm 6BS (Yr78). Theoretical and Applied Genetics, 130, 2127-2137.
- Dossa, G. S., Oliva, R., Maiss, E., Vera Cruz, C., & Wydra, K. (2016). High temperature enhances the resistance of cultivated African rice, Oryza glaberrima, to bacterial blight. Plant Disease, 100(2), 380-387. [CrossRef]
- Draz, I. S. (2019a). Common Ancestry of Egyptian Puccinia striiformis population along with effective and ineffective resistance genes. Asian Journal of Biological Sciences, 12, 217-221. [CrossRef]
- Draz, I. S. (2019b). Pathotypic and molecular evolution of contemporary population of Puccinia striiformis f. sp. tritici in Egypt during 2016–2018. Journal of Phytopathology, 167(1), 26-34.
- Druzhin, A.E. Effect of climate change on the structure of populations of spring wheat pathogens in the Volga region. Agrar. Report. South-East 2010, 1, 31-36. (In Russian).
- Duveiller, E., Singh, R. P., & Nicol, J. M. (2007). The challenges of maintaining wheat productivity: pests, diseases, and potential epidemics. Euphytica, 157(3), 417-430. [CrossRef]
- Dyck, P. L., & Johnson, R. (1983). Temperature sensitivity of genes for resistance in wheat to Puccinia recondita. Canadian Journal of Plant Pathology, 5(4), 229-234. [CrossRef]
- Eastburn, D. M., McElrone, A. J., & Bilgin, D. D. (2011). Influence of atmospheric and climatic change on plant–pathogen interactions. Plant pathology, 60(1), 54-69. [CrossRef]
- Eizenberg, H., Plakhine, D., Hershenhorn, J., Kleifeld, Y., & Rubin, B. (2003). Resistance to broomrape (Orobanche spp.) in sunflower (Helianthus annuus L.) is temperature dependent. Journal of Experimental Botany, 54(385), 1305-1311. [CrossRef]
- El Amil, R., Shykoff, J. A., Vidal, T., Boixel, A. L., Leconte, M., Hovmøller, M. S., ... & de Vallavieille-Pope, C. (2022). Diversity of thermal aptitude of Middle Eastern and Mediterranean Puccinia striiformis f. sp. tritici isolates from different altitude zones. Plant Pathology, 71(8), 1674-1687.
- Elbasyoni, I. S., El-Orabey, W. M., Morsy, S., Baenziger, P. S., Al Ajlouni, Z., & Dowikat, I. (2019). Evaluation of a global spring wheat panel for yellow rust: Resistance loci validation and novel resources identification. PLoS One, 14(11), e0222755.
- El-Orabey, W. M., Elbasyoni, I. S., El-Moghazy, S. M., & Ashmawy, M. A. (2019). Effective and ineffective of some resistance genes to wheat leaf, stem and yellow rust diseases in Egypt. Journal of Plant Production, 10(4), 361-371. [CrossRef]
- Eriksson, J. (1894). Uber die Spezialisierung des Parasitismus bei den Getreiderostpilzen. Berichte der Deutschen Botanischen Gesellschaft, 12, 292–331.
- Esmail, S. M., Draz, I. S., Ashmawy, M. A., & El-Orabey, W. M. (2021). Emergence of new aggressive races of Puccinia striiformis f. sp. tritici causing yellow rust epiphytotic in Egypt. Physiological and Molecular Plant Pathology, 114, 101612. [CrossRef]
- Esmail, S. M., Omar, G. E., El-Orabey, W. M., Börner, A., & Mourad, A. M. (2023). Exploring the genetic variation of yellow rust foliar and head infection in Egyptian wheat as an effect of climate change. Agronomy, 13(6), 1509. [CrossRef]
- Ezzahiri B, Yahyaoui A, Hovmøller MS (2009) An analysis of the 2009 Epidemic of yellow rust on wheat in Morocco. In The 4th Regional Yellow Rust Conference for Central and West Asia and North Africa (Antalya: Turkish Ministry of Agriculture and Rural Affairs, ICARDA, CIMMYT, FAO of the United Nations).
- Feng, J., Wang, M., See, D. R., Chao, S., Zheng, Y., & Chen, X. (2018). Characterization of novel gene Yr79 and four additional quantitative trait loci for all-stage and high-temperature adult-plant resistance to yellow rust in spring wheat PI 182103. Phytopathology, 108(6), 737-747. [CrossRef]
- Feodorova-Fedotova, L., & Bankina, B. (2018). Characterization of yellow rust (Puccinia striiformis westend). Research for Rural Development, 2.
- Fu, D., Uauy, C., Distelfeld, A., Blechl, A., Epstein, L., Chen, X., & Dubcovsky, J. (2009). A kinase-START gene confers temperature-dependent resistance to wheat yellow rust. science, 323(5919), 1357-1360. [CrossRef]
- Gangwar, O. P., Kumar, S., Prasad, P., Bhardwaj, S. C., Khan, H., & Verma, H. (2016). Virulence pattern and emergence of new pathotypes in Puccinia striiformis f. sp. tritici during 2011-15 in India. Indian Phytopathol, 69(4s), 178-185.
- Gardner, H., Onofre, K. F. A., & De Wolf, E. D. (2023). Characterizing the response of Puccinia striiformis f. sp. tritici to periods of heat stress that are common in Kansas and the great plains region of North America. Phytopathology®, 113(8), 1457-1464.
- Garrett, K. A., Bebber, D. P., Etherton, B. A., Gold, K. M., Plex Sulá, A. I., & Selvaraj, M. G. (2022). Climate change effects on pathogen emergence: Artificial intelligence to translate big data for mitigation. Annual Review of Phytopathology, 60(1), 357-378. [CrossRef]
- Garrett, K. A., Dendy, S. P., Frank, E. E., Rouse, M. N., & Travers, S. E. (2006). Climate change effects on plant disease: genomes to ecosystems. Annual Review of Phytopathology 44(1), 489-509.
- Ghelardini, L., Pepori, A. L., Luchi, N., Capretti, P., & Santini, A. (2016). Drivers of emerging fungal diseases of forest trees. Forest Ecology and Management, 381, 235-246. [CrossRef]
- Gladders, P., Langton, S. D., Barrie, I. A., Hardwick, N. V., Taylor, M. C., & Paveley, N. D. (2007). The importance of weather and agronomic factors for the overwinter survival of yellow rust (Puccinia striiformis) and subsequent disease risk in commercial wheat crops in England. Annals of applied Biology, 150(3), 371-382. [CrossRef]
- Gomes, C., Almeida, A. S., Coutinho, J., Costa, R., Pinheiro, N., Coco, J., & Maçãs, B. (2018). Foliar fungicide application as management strategy to minimize the growing threat of yellow rust on wheat in Portugal. Emirates Journal of Food and Agriculture, 30(9), 715-724.
- GRRC report of yellow and stem rust races 2022: GRRC, Aarhus University, Denmark www.wheatrust.org.
- Gulev SK, Thorne PW, Ahn J, Dentener FJ, Domingues CM, Gerland S, Gong D, Kaufman DS, Nnamchi HC, Quaas J, Rivera JA, Sathyendranath S, Smith SL, Trewin B, von Schuckmann K, Vose RS (2021) Changing state of the climate system. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, Chapter 2. [CrossRef]
- Gultyaeva, E., Shaydayuk, E., Gannibal, P., & Kosman, E. (2021). Analysis of host-specific differentiation of Puccinia striiformis in the South and North-West of the European Part of Russia. Plants, 10(11), 2497.
- He, X., Gahtyari, N. C., Roy, C., Dababat, A. A., Brar, G. S., & Singh, P. K. (2022). Globally important non-rust diseases of wheat. In Wheat Improvement: Food Security in a Changing Climate (pp. 143-158). Cham: Springer International Publishing.
- Heagle, A. S., Spencer, S., & Letchworth, M. B. (1979). Yield response of winter wheat to chronic doses of ozone. Canadian Journal of Botany, 57(19), 1999-2005. [CrossRef]
- Helfer, S. (2014). Rust fungi and global change. New phytologist, 201(3), 770-780. [CrossRef]
- Helguera, M., Khan, I. A., Kolmer, J., Lijavetzky, D., Zhong-Qi, L., & Dubcovsky, J. (2003). PCR assays for the Lr37-Yr17-Sr38 cluster of rust resistance genes and their use to develop isogenic hard red spring wheat lines. Crop science, 43(5), 1839-1847. [CrossRef]
- Hovmøller, M. S., & Justesen, A. F. (2007). Appearance of atypical Puccinia striiformis f. sp. tritici phenotypes in north-western Europe. Australian Journal of Agricultural Research, 58(6), 518-524. [CrossRef]
- Hovmøller, M. S., Thach, T., & Justesen, A. F. (2023). Global dispersal and diversity of rust fungi in the context of plant health. Current Opinion in Microbiology, 71, 102243. [CrossRef]
- Hovmøller, M. S., Walter, S., Bayles, R. A., Hubbard, A., Flath, K., Sommerfeldt, N., ... & de Vallavieille-Pope, C. (2016). Replacement of the European wheat yellow rust population by new races from the centre of diversity in the near-Himalayan region. Plant Pathology, 65(3), 402-411. [CrossRef]
- Hovmøller, M. S., Yahyaoui, A. H., Milus, E. A., & Justesen, A. F. (2008). Rapid global spread of two aggressive strains of a wheat rust fungus. Molecular Ecology, 17(17), 3818-3826. [CrossRef]
- Hubbard, A., Lewis, C. M., Yoshida, K., Ramirez-Gonzalez, R. H., de Vallavieille-Pope, C., Thomas, J., ... & Saunders, D. G. (2015). Field pathogenomics reveals the emergence of a diverse wheat yellow rust population. Genome biology, 16, 1-15. [CrossRef]
- Hulbert, S., & Pumphrey, M. (2014). A time for more booms and fewer busts? Unraveling cereal–rust interactions. Molecular Plant-Microbe Interactions, 27(3), 207-214. [CrossRef]
- Hussain M, Kirmani MAS, Haque E, 2004. Pathotypes and man guided evolution of Puccinia striiformis West sp. tritici in Pakistan. Abstracts, Second Regional Yellow Rust Conference for Central & West Asia and North Africa, 22–26 March 2004, Islamabad, Pakistan. 21.
- ICARDA. (2011). Strategies to reduce the emerging wheat yellow rust disease. Synthesis of a dialog between policy makers and scientists from 31 countries at: International Wheat Yellow Rust Symposium, Aleppo, Syria, April 2011. International Center for Agriculture Research in the Dry Areas (ICARDA) http://www.fao.org/familyfarming/detail/en/c/325927/. Accessed 8 Nov 2019.
- Indu, S., & Saharan, M. S. (2011). Status of wheat diseases in India with a special reference to yellow rust. Plant Dis. Res, 26, 156.
- IPCC 2022 Summary for Policymakers Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change ed H O Pörtner et al (Cambridge University Press) pp 3–33.
- IPCC 2023 Summary for policymakers Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change Core Writing Team ed H Lee and J Romero (IPCC, Geneva) pp 1–34.
- IPCC Secretariat. (2021). Scientific review of the impact of climate change on plant pests-A global challenge to prevent and mitigate plant pest risks in agriculture, forestry and ecosystems. FAO on behalf of the IPCC Secretariat. https://doir.org/10.4060/cb4769en.
- Ivanova, Y. N., Rosenfread, K. K., Stasyuk, A. I., Skolotneva, E. S., & Silkova, O. G. (2021). Raise and characterization of a bread wheat hybrid line (Tulaykovskaya 10× Saratovskaya 29) with chromosome 6Agi2 introgressed from Thinopyrum intermedium. Vavilov Journal of Genetics and Breeding, 25(7), 701-712. [CrossRef]
- Jevtić, R., Župunski, V., Lalošević, M., & Župunski, L. (2017). Predicting potential winter wheat yield losses caused by multiple disease systems and climatic conditions. Crop Protection, 99, 17-25. [CrossRef]
- Jevtić, R., Župunski, V., Lalošević, M., Jocković, B., Orbović, B., & Ilin, S. (2020). Diversity in susceptibility reactions of winter wheat genotypes to obligate pathogens under fluctuating climatic conditions. Scientific Reports, 10(1), 19608. [CrossRef]
- Jin, Y., Szabo, L. J., & Carson, M. (2010). Century-old mystery of Puccinia striiformis life history solved with the identification of Berberis as an alternate host. Phytopathology, 100(5), 432-435. [CrossRef]
- Jindal MM, Mohan C, Pannu PPS (2012) Status of Yellow rust of wheat in Punjab during 2011-12 season. In: Proceedings of brain storming session. Department of Plant Pathology PAU, Ludhian, p 56.
- Juroszek, P., & von Tiedemann, A. (2013). Climate change and potential future risks through wheat diseases: a review. European Journal of Plant Pathology, 136, 21-33.
- Juroszek, P., Racca, P., Link, S., Farhumand, J., & Kleinhenz, B. (2020). Overview on the review articles published during the past 30 years relating to the potential climate change effects on plant pathogens and crop disease risks. Plant pathology, 69(2), 179-193. [CrossRef]
- Khanfri, S., Boulif, M., & Lahlali, R. (2018). Yellow rust (Puccinia striiformis): a serious threat to wheat production worldwide. Notulae Scientia Biologicae, 10(3), 410-423. [CrossRef]
- Kokhmetova, A., Sharma, R. C., Rsaliyev, S., Galymbek, K., Baymagambetova, K., Ziyaev, Z., & Morgounov, A. (2018). Evaluation of Central Asian wheat germplasm for yellow rust resistance. Plant Genetic Resources, 16(2), 178-184. [CrossRef]
- Krattinger, S.G., Lagudah, E.S., Spielmeyer, W., Singh, R.P., Huerta-Espino, J., McFadden, H., Bossolini, E., Selter, L.L., Keller, B. (2009). A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 323, 1360–1363. [CrossRef]
- Lafferty, K. D. (2009). The ecology of climate change and infectious diseases. Ecology, 90(4), 888-900.
- Lan, C., Randhawa, M. S., Huerta-Espino, J., & Singh, R. P. (2017). Genetic analysis of resistance to wheat rusts. Wheat Rust Diseases: Methods and Protocols, 137-149.
- Lee JY, Marotzke J, Bala G, Cao L, Corti S, Dunne JP, Engelbrecht F, Fischer E, Fyfe JC, Jones C, Maycock A, Mutemi J, Ndiaye O, Panickal S, Zhou T (2021) Future global climate: scenario-based projections and near-term information. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, Chapter 4. [CrossRef]
- Li, Z. Q., & Zeng, S. M. (2002). Wheat rusts in China (pp. 379). Beijing: Chinese Agricultural Press (In Chinese).
- Line, R. F. (2002). Yellow rust of wheat and barley in North America: a retrospective historical review. Annual review of phytopathology, 40(1), 75-118.
- Liu, L., Wang, M. N., Feng, J. Y., See, D. R., Chao, S. M., & Chen, X. M. (2018). Combination of all-stage and high-temperature adult-plant resistance QTL confers high-level, durable resistance to yellow rust in winter wheat cultivar Madsen. Theoretical and Applied Genetics, 131, 1835-1849. [CrossRef]
- Liu, L., Wang, M., Zhang, Z., See, D. R., & Chen, X. (2020). Identification of Yellow rust resistance loci in US spring wheat cultivars and breeding lines using genome-wide association mapping and Yr gene markers. Plant Disease, 104(8), 2181-2192.
- Liu, L., Yuan, C. Y., Wang, M. N., See, D. R., Zemetra, R. S., & Chen, X. M. (2019). QTL analysis of durable yellow rust resistance in the North American winter wheat cultivar Skiles. Theoretical and Applied Genetics, 132, 1677-1691.
- Loladze, A., Druml, T., & Wellings, C. R. (2014). Temperature adaptation in Australasian populations of Puccinia striiformis f. sp. tritici. Plant pathology, 63(3), 572-580.
- Lu, Y., Wang, M., Chen, X., See, D., Chao, S., & Jing, J. (2014). Mapping of Yr62 and a small-effect QTL for high-temperature adult-plant resistance to yellow rust in spring wheat PI 192252. Theoretical and Applied Genetics, 127, 1449-1459. [CrossRef]
- Lyon, B., & Broders, K. (2017). Impact of climate change and race evolution on the epidemiology and ecology of yellow rust in central and eastern USA and Canada. Canadian Journal of Plant Pathology, 39(4), 385-392.
- Ma, L., Qiao, J., Kong, X., Zou, Y., Xu, X., Chen, X., & Hu, X. (2015). Effect of low temperature and wheat winter-hardiness on survival of Puccinia striiformis f. sp. tritici under controlled conditions. PLoS One, 10(6), e0130691. [CrossRef]
- Mashaheet, A. M., Burkey, K. O., Saitanis, C. J., Abdelrhim, A. S., Rafiullah, & Marshall, D. S. (2020). Differential ozone responses identified among key rust-susceptible wheat genotypes. Agronomy, 10(12), 1853.
- Mboup, M., Bahri, B., Leconte, M., De Vallavieille-Pope, C., Kaltz, O., & Enjalbert, J. (2012). Genetic structure and local adaptation of European wheat yellow rust populations: the role of temperature-specific adaptation. Evolutionary applications, 5(4), 341-352. [CrossRef]
- McDonald, B. A., & Linde, C. (2002). Pathogen population genetics, evolutionary potential, and durable resistance. Annual review of phytopathology, 40(1), 349-379.
- Mcelrone, A. J., Reid, C. D., Hoye, K. A., Hart, E., & Jackson, R. B. (2005). Elevated CO2 reduces disease incidence and severity of a red maple fungal pathogen via changes in host physiology and leaf chemistry. Global Change Biology, 11(10), 1828-1836. [CrossRef]
- Megahed, E. M., Awaad, H. A., Ramadan, I. E., Abdul-Hamid, M. I., Sweelam, A. A., El-Naggar, D. R., & Mansour, E. (2022). Assessing performance and stability of yellow rust resistance, heat tolerance, and agronomic performance in diverse bread wheat genotypes for enhancing resilience to climate change under Egyptian conditions. Frontiers in Plant Science, 13, 1014824.
- Miedaner, T., & Juroszek, P. (2021). Climate change will influence disease resistance breeding in wheat in Northwestern Europe. Theoretical and Applied Genetics, 134(6), 1771-1785. [CrossRef]
- Mills, G., Sharps, K., Simpson, D., Pleijel, H., Broberg, M., Uddling, J., ... & Van Dingenen, R. (2018). Ozone pollution will compromise efforts to increase global wheat production. Global change biology, 24(8), 3560-3574. [CrossRef]
- Milus, E. A., Kristensen, K., & Hovmøller, M. S. (2009). Evidence for increased aggressiveness in a recent widespread strain of Puccinia striiformis f. sp. tritici causing yellow rust of wheat. Phytopathology, 99(1), 89-94.
- Milus, E. A., Seyran, E., & McNew, R. (2006). Aggressiveness of Puccinia striiformis f. sp. tritici isolates in the south-central United States. Plant Disease, 90(7), 847-852.
- Mourad, A. M., Abou-Zeid, M. A., Eltaher, S., Baenziger, P. S., & Börner, A. (2021). Identification of candidate genes and genomic regions associated with adult plant resistance to yellow rust in spring wheat. Agronomy, 11(12), 2585. [CrossRef]
- Mu, J., Liu, L., Liu, Y., Wang, M., See, D. R., Han, D., & Chen, X. (2020). Genome-wide association study and gene specific markers identified 51 genes or QTL for resistance to yellow rust in US winter wheat cultivars and breeding lines. Frontiers in Plant Science, 11, 998. [CrossRef]
- Murray, G. M., & Brennan, J. P. (2009). Estimating disease losses to the Australian wheat industry. Australasian Plant Pathology, 38(6), 558-570.
- Niks, R. E., Qi, X., & Marcel, T. C. (2015). Quantitative resistance to biotrophic filamentous plant pathogens: concepts, misconceptions, and mechanisms. Annual review of phytopathology, 53(1), 445-470. [CrossRef]
- Nnadi, N. E., & Carter, D. A. (2021). Climate change and the emergence of fungal pathogens. PLoS pathogens, 17(4), e1009503.
- Novotná M, Hloucalová P, Skládanka J, Pokorný R (2017) Effect of weather on the occurrence of Puccinia graminis subsp. graminicola and Puccinia coronata f. sp. lolii at Lolium perenne L. and Deschampsia caespitosa (L.). Acta Universitatis Agriculturae et Silviculturae Mendelianae Brunensis, 65: 125-134. [CrossRef]
- Ochoa, J. B., Danial, D. L., & Paucar, B. (2007). Virulence of wheat yellow rust races and resistance genes of wheat cultivars in Ecuador. Euphytica, 153(3), 287-293. [CrossRef]
- Ortiz, R., Sayre, K. D., Govaerts, B., Gupta, R., Subbarao, G. V., Ban, T., ... & Reynolds, M. (2008). Climate change: can wheat beat the heat?. Agriculture, Ecosystems & Environment, 126(1-2), 46-58. [CrossRef]
- Ortiz-Bobea, A., Ault, T. R., Carrillo, C. M., Chambers, R. G., & Lobell, D. B. (2021). Anthropogenic climate change has slowed global agricultural productivity growth. Nature Climate Change, 11(4), 306-312.
- Pariaud, B., Ravigné, V., Halkett, F., Goyeau, H., Carlier, J., & Lannou, C. (2009). Aggressiveness and its role in the adaptation of plant pathogens. Plant Pathology, 58(3), 409-424. [CrossRef]
- Pariaud, B., Robert, C., Goyeau, H., & Lannou, C. (2009b). Aggressiveness components and adaptation to a host cultivar in wheat leaf rust. Phytopathology, 99(7), 869-878.
- Pathak, R., Singh, S. K., Tak, A., & Gehlot, P. (2018). Impact of climate change on host, pathogen and plant disease adaptation regime: a review. Biosciences Biotechnology Research Asia, 15(3), 529-540. [CrossRef]
- Pautasso, M., Döring, T. F., Garbelotto, M., Pellis, L., & Jeger, M. J. (2012). Impacts of climate change on plant diseases-opinions and trends. European journal of plant pathology, 133, 295-313.
- Prank, M., Kenaley, S. C., Bergstrom, G. C., Acevedo, M., & Mahowald, N. M. (2019). Climate change impacts the spread potential of wheat stem rust, a significant crop disease. Environmental Research Letters, 14(12), 124053. [CrossRef]
- Prashar, M., Bhardwaj, S. C., Jain, S. K., & Datta, D. (2007). Pathotypic evolution in Puccinia striiformis in India during 1995–2004. Australian Journal of Agricultural Research, 58(6), 602-604. [CrossRef]
- Prashar, M., Bhardwaj, S. C., Jain, S. K., & Datta, D. (2007). Pathotypic evolution in Puccinia striiformis in India during 1995–2004. Australian Journal of Agricultural Research, 58(6), 602-604. [CrossRef]
- Pretorius, Z. A. (2004). The impact of wheat yellow rust in South Africa. In Proc 11th Intl Cereal Rusts and Powdery Mildews Conf, 22–27 Aug 2004, John Innes Centre, Norwich, UK, European and Mediterranean Cereal Rust Foundation, Wageningen, Netherland, Cereal Rusts and Powdery Mildews Bulletin, Abstract A1.29.
- Priestley, R. H., & Bayles, R. A. (1988). The contribution and value of resistant cultivars to disease control in cereals. In B. C. Clifford & E. Lester (Eds.), Control of plant diseases, Costs and Benefits (pp. 53–65). Oxford: Blackwell Scientific Publications.
- Rahmatov M, Eshonova Z, Ibrogimov A, Otambekova M, Khuseinov B, Muminjanov H et al (2012) Monitoring and evaluation of yellow rust for breeding resistant varieties of wheat in Tajikistan. In Meeting the Challenge of Yellow Rust in Cereal Crops Proceedings of the 2nd, 3rd and 4th Regional Conferences on Yellow Rust in Central and West Asia and North Africa (CWANA) Region, Yahyaoui A, Rajaram S (eds) International Center for Agricultural Research in the Dry Areas, Alnarp.
- Ren, R. S., Wang, M. N., Chen, X. M., & Zhang, Z. J. (2012). Characterization and molecular mapping of Yr52 for high-temperature adult-plant resistance to yellow rust in spring wheat germplasm PI 183527. Theoretical and Applied Genetics, 125, 847-857. [CrossRef]
- Roelfs, A. P., R. P. Singh and E. E. Saari. 1992. Rust Diseases of Wheat: Concepts and.
- methods of disease management. Mexico, D.F.: CIMMYT. 81 pages.
- Rodriguez-Algaba, J., Sørensen, C. K., Labouriau, R., Justesen, A. F., & Hovmøller, M. S. (2019). Susceptibility of winter wheat and triticale to yellow rust influenced by complex interactions between vernalisation, temperature, plant growth stage and pathogen race. Agronomy, 10(1), 13. [CrossRef]
- Saari, E. E., & Prescott, J. M. (1985). World distribution in relation to economic losses. In A. P. Roelfs & W. R. Bushnell (Eds.), The cereal rusts (Vol. II, pp. 259–298). Orlando: Academic Press.
- Sánchez Espinosa, K. C., Fernández-González, M., Almaguer, M., Guada, G., & Rodríguez-Rajo, F. J. (2023). Puccinia Spore Concentrations in Relation to Weather Factors and Phenological Development of a Wheat Crop in Northwestern Spain. Agriculture, 13(8), 1637. [CrossRef]
- Sanin, S.S. Agricultural plant disease control-The main factor of the crop production intensification. Plant Prot. News. 2010, 1,3–14. (In Russian).
- Santra, D. K., Chen, X. M., Santra, M., Campbell, K. G., & Kidwell, K. K. (2008). Identification and mapping QTL for high-temperature adult-plant resistance to Yellow rust in winter wheat (Triticum aestivum L.) cultivar ‘Stephens’. Theoretical and Applied Genetics, 117, 793-802.
- Savadi, S. (2018). Molecular regulation of seed development and strategies for engineering seed size in crop plants. Plant Growth Regulation, 84(3), 401-422. [CrossRef]
- Savary, S., Willocquet, L., Pethybridge, S. J., Esker, P., McRoberts, N., & Nelson, A. (2019). The global burden of pathogens and pests on major food crops. Nature ecology & evolution, 3(3), 430-439.
- Schade, F. M., Shama, L. N., & Wegner, K. M. (2014). Impact of thermal stress on evolutionary trajectories of pathogen resistance in three-spined stickleback (Gasterosteus aculeatus). BMC evolutionary biology, 14, 1-12. [CrossRef]
- Scherm H, Yang, X. B. (1995) Interannual variations in wheat rust development in China and the United States in relation to the El Niño/Southern Oscillation. Phytopathology 85:970-976.
- Scherm, H., & Yang, X. B. (1998). Atmospheric teleconnection patterns associated with wheat Yellow rust disease in North China. International Journal of Biometeorology, 42, 28-33. [CrossRef]
- Schimanke, S., Joelsson, M., Andersson, S., Carlund, T., Wern, L., Hellström, S., & Kjellström, E. (2022). Observerad klimatförändring i Sverige 1860–2021.
- Scholthof, K. B. G. (2007). The disease triangle: pathogens, the environment and society. Nature Reviews Microbiology, 5(2), 152-156.
- Schwessinger, B. (2017). Fundamental wheat Yellow rust research in the 21st century. New Phytologist, 213(4), 1625-1631.
- Shahin, A. A. (2020). Occurrence of new races and virulence changes of the wheat Yellow rust pathogen (Puccinia striiformis f. sp. tritici) in Egypt. Archives of Phytopathology and Plant Protection, 53(11-12), 552-569.
- Shahin, A., Draz, I., & Esmail, S. (2020). Race specificity of Yellow rust resistance in relation to susceptibility of Egyptian wheat cultivars. Egyptian Journal of Phytopathology, 48(1), 1-13. [CrossRef]
- Shahin, A.; Shaheen, S.; Abu, A. A. A. (2015) Virulence and Diversity of Wheat Yellow Rust Pathogen in Egypt. J. Am. Sci., 11, 47–52.
- Sharma-Poudyal, D., Chen, X. M., Wan, A. M., Zhan, G. M., Kang, Z. S., Cao, S. Q., ... & Patzek, L. J. (2013). Virulence characterization of international collections of the wheat Yellow rust pathogen, Puccinia striiformis f. sp. tritici. Plant Disease, 97(3), 379-386.
- Sharma-Poudyal, D., Chen, X., & Rupp, R. A. (2014). Potential over summering and overwintering regions for the wheat yellow rust pathogen in the contiguous United States. International journal of biometeorology, 58, 987-997.
- Shaw, M. W., Bearchell, S. J., Fitt, B. D., & Fraaije, B. A. (2008). Long-term relationships between environment and abundance in wheat of Phaeosphaeria nodorum and Mycosphaerella graminicola. New Phytologist, 177(1), 229-238. [CrossRef]
- Singh, V. K., Mathuria, R. C., Gogoi, R. O. B. I. N., & Aggarwal, R. A. S. H. M. I. (2016). Impact of different fungicides and bioagents, and fungicidal spray timing on wheat Yellow rust development and grain yield. Indian Phytopath, 69(4), 357-362.
- Solh, M., Nazari, K., Tadesse, W., & Wellings, C. R. (2012). The growing threat of Yellow rust worldwide. BGRI 2012 Technical workshop, 1–4 September 2012, Beijing, China. Borlaug Global Rust Initiative.
- Strandberg, G., Andersson, B., & Berlin, A. (2024). Plant pathogen infection risk and climate change in the Nordic and Baltic countries. Environmental research communications, 6(3), 031008. [CrossRef]
- Stubbs, R. W. (1985). Yellow rust. In A. P. Roelfs & W. R. Bushnell (Eds.), The cereal rusts (Vol. II, pp. 61–101). Orlando: Academic Press.
- Sturrock, R. N., Frankel, S. J., Brown, A. V., Hennon, P. E., Kliejunas, J. T., Lewis, K. J., ... & Woods, A. J. (2011). Climate change and forest diseases. Plant pathology, 60(1), 133-149.
- Sukumar Chakraborty, S. C., Luck, J., Hollaway, G., Freeman, A., Norton, R., Garrett, K. A., & Karnosky, D. F. (2008). Impacts of global change on diseases of agricultural crops and forest trees. CABI Reviews, (2008), 1-15.
- Tariq-Khan, M., Younas, M. T., Mirza, J. I., Awan, S. I., Jameel, M., Saeed, M., & Mahmood, B. (2020). Evaluation of major and environmentally driven genes for resistance in Pakistani wheat landraces and their prospected potential against yellow rust. International Journal of Phytopathology, 9(3), 145-156. [CrossRef]
- Tiedemann, A. V., & Firsching, K. H. (2000). Interactive effects of elevated ozone and carbon dioxide on growth and yield of leaf rust-infected versus non-infected wheat. Environmental Pollution, 108(3), 357-363. [CrossRef]
- Uauy, C., Brevis, J. C., Chen, X., Khan, I., Jackson, L., Chicaiza, O., ... & Dubcovsky, J. (2005). High-temperature adult-plant (HTAP) Yellow rust resistance gene Yr36 from Triticum turgidum ssp. dicoccoides is closely linked to the grain protein content locus Gpc-B1. Theoretical and Applied Genetics, 112, 97-105.
- Vergara-Diaz, O., Kefauver, S. C., Elazab, A., Nieto-Taladriz, M. T., & Araus, J. L. (2015). Grain yield losses in yellow-rusted durum wheat estimated using digital and conventional parameters under field conditions. The Crop Journal, 3(3), 200-210.
- Vidal, T., Boixel, A. L., Maghrebi, E., Perronne, R., du Cheyron, P., Enjalbert, J., ... & de Vallavieille-Pope, C. (2022). Success and failure of invasive races of plant pathogens: the case of Puccinia striiformis f. sp. tritici in France. Plant pathology, 71(7), 1525-1536.
- Waheed, A., Haxim, Y., Islam, W., Ahmad, M., Muhammad, M., Alqahtani, F. M., ... & Zhang, D. (2023). Climate change reshaping plant-fungal interaction. Environmental Research, 117282. [CrossRef]
- Walter, S., Ali, S., Kemen, E., Nazari, K., Bahri, B. A., Enjalbert, J., ... & Justesen, A. F. (2016). Molecular markers for tracking the origin and worldwide distribution of invasive strains of Puccinia striiformis. Ecology and Evolution, 6(9), 2790-2804. [CrossRef]
- Wan, A., Zhao, Z., Chen, X., He, Z., Jin, S., Jia, Q., ... & Yuan, Z. (2004). Wheat Yellow rust epidemic and virulence of Puccinia striiformis f. sp. tritici in China in 2002. Plant Disease, 88(8), 896-904.
- Wang, M.N., Chen, X.M., 2017. Yellow rust resistance. In: Kang, Z.S. (Ed.), Yellow Rust. X. M. Chen. Springer, Dordrecht, pp. 353–558.
- Webb, K. M., Ona, I., Bai, J., Garrett, K. A., Mew, T., Vera Cruz, C. M., & Leach, J. E. (2010). A benefit of high temperature: increased effectiveness of a rice bacterial blight disease resistance gene. New Phytologist, 185(2), 568-576. [CrossRef]
- Wellings, C. R. (2007). Puccinia striiformis in Australia: a review of the incursion, evolution, and adaptation of Yellow rust in the period 1979–2006. Australian Journal of Agricultural Research, 58(6), 567-575. [CrossRef]
- Wellings, C. R. (2011). Global status of Yellow rust: a review of historical and current threats. Euphytica, 179(1), 129-141.
- Weng, Y., Azhaguvel, P., Devkota, R. N., & Rudd, J. C. (2007). PCR-based markers for detection of different sources of 1AL. 1RS and 1BL. 1RS wheat–rye translocations in wheat background. Plant Breeding, 126(5), 482-486.
- Wiik, L., & Ewaldz, T. (2009). Impact of temperature and precipitation on yield and plant diseases of winter wheat in southern Sweden 1983–2007. Crop Protection, 28(11), 952-962. [CrossRef]
- WMO 2022 State of the global climate 2021 WMO-No. 1290 57 ISBN 978-92-63-11290-3 https://library.wmo.int/records/item/56300-state-of-the-global-climate-2021 last access Aug 31, 2023.
- WMO 2023 State of the global climate 2022, WMO-No 1316 55 ISBN 978-92-63-11316-0 https://library.wmo.int/records/item/66214-state-of-the-global-climate-2022 last access Aug 31, 2023.
- World agricultural production. USDA https://apps.fas.usda.gov/psdonline/circulars/production.pdf (2022).
- Wyka, S. A., Smith, C., Munck, I. A., Rock, B. N., Ziniti, B. L., & Broders, K. (2017). Emergence of white pine needle damage in the northeastern United States is associated with changes in pathogen pressure in response to climate change. Global Change Biology, 23(1), 394-405. [CrossRef]
- Yang, L. N., Zhu, W., Wu, E. J., Yang, C., Thrall, P. H., Burdon, J. J., ... & Zhan, J. (2016). Trade-offs and evolution of thermal adaptation in the Irish potato famine pathogen Phytophthora infestans. Molecular Ecology, 25(16), 4047-4058. [CrossRef]
- Zadoks, J. C., & Rijsdijk, F. H. (1984). Atlas of cereal diseases and pests in Europe. Agro-ecological atlas of cereal growing in Europe Vol III. Pudoc, Wageningen, the Netherlands.
- Zeleneva, Y. V., Sudnikova, V. P., & Buchneva, G. N. (2022). Immunological characteristics of soft winter wheat varieties in conditions of the CBR. Proc. Kuban State Agrar. Univ, 96, 95-99. [CrossRef]
- Zhan, J., & McDONALD, B. A. (2011). Thermal adaptation in the fungal pathogen Mycosphaerella graminicola. Molecular Ecology, 20(8), 1689-1701. [CrossRef]
- Zhang, J. Q., Liu, B., Chen, W. Q., Liu, T. G., & Gao, L. (2013). Temperature-sensitivity of population of Puccinia striiformis Westend. Acta Phytopathologica Sinnica, 43(1), 88-90.
- Zhao, J., & Kang, Z. (2023). Fighting wheat rusts in China: a look back and into the future. Phytopathology Research, 5(1), 6.
- Zhao, J., Wang, M., Chen, X., & Kang, Z. (2016). Role of alternate hosts in epidemiology and pathogen variation of cereal rusts. Annual Review of Phytopathology, 54(1), 207-228. [CrossRef]
- Zhou, X. L., Wang, M. N., Chen, X. M., Lu, Y., Kang, Z. S., & Jing, J. X. (2014). Identification of Yr59 conferring high-temperature adult-plant resistance to Yellow rust in wheat germplasm PI 178759. Theoretical and Applied Genetics, 127, 935-945. [CrossRef]
- Zhu, Z., Cao, Q., Han, D., Wu, J., Wu, L., Tong, J., ... & Hao, Y. (2023). Molecular characterization and validation of adult-plant Yellow rust resistance gene Yr86 in Chinese wheat cultivar Zhongmai 895. Theoretical and Applied Genetics, 136(6), 142. [CrossRef]
- Župunski, V., Jevtić, R. & Lalošević, M. Te occurrence of yellow rust of winter wheat in relation to year. Book of abstracts of XV Symposium on plant protection, Zlatibor, Serbia, 28 November–02 December, pp. 89–90 (2016) (in Serbian).
Figure 1.
Changing climate impact on host and epidemiology of pathogens.
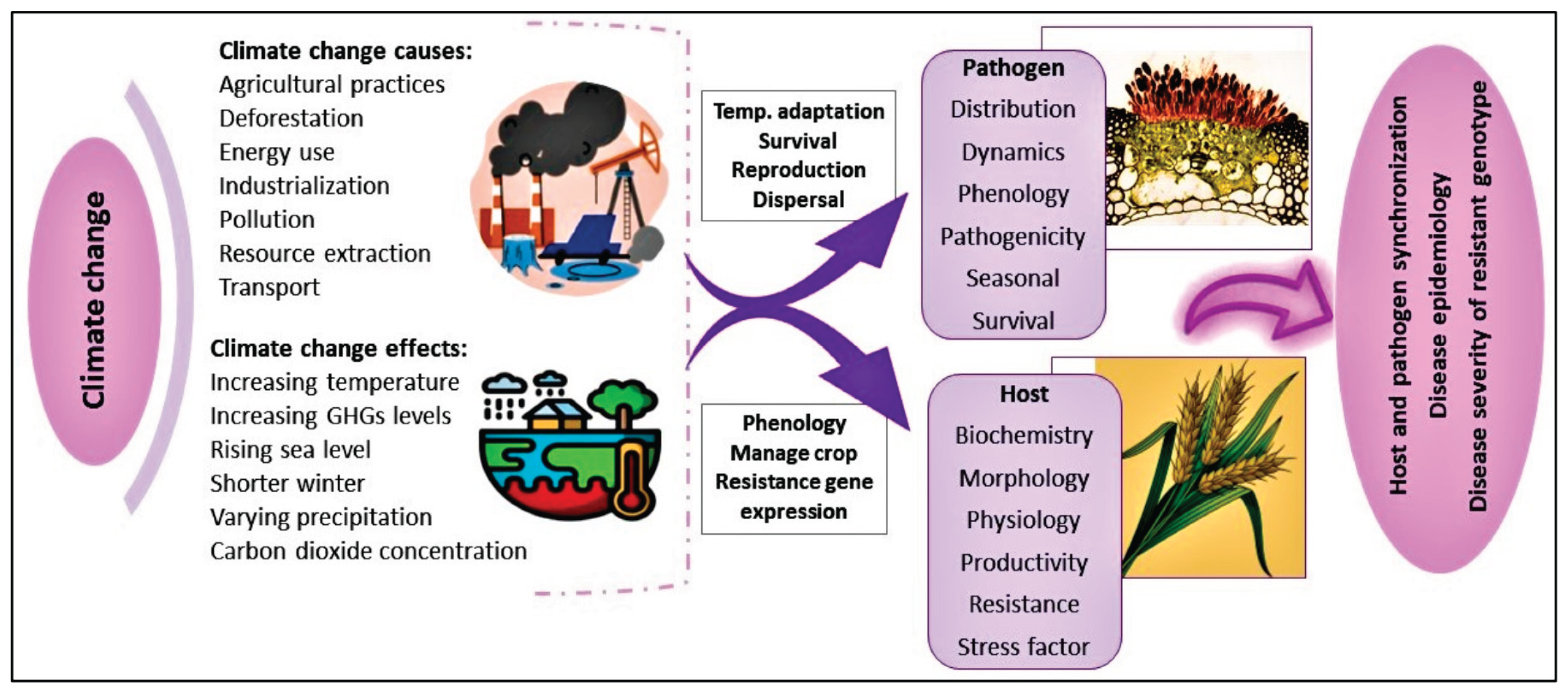
Figure 2.
source Wiik, L. (2009). Control of fungal diseases in winter wheat (Vol. 2009, No. 2009: 97).
Figure 2.
source Wiik, L. (2009). Control of fungal diseases in winter wheat (Vol. 2009, No. 2009: 97).


Table 1.
Climatic factors governing the progression of the yellow rust pathogen at various developmental stages.
Table 1.
Climatic factors governing the progression of the yellow rust pathogen at various developmental stages.
| Developmental Stage |
Temperature (°C) | Light | Free water | ||
|---|---|---|---|---|---|
|
Lowest |
Optimum |
Highest |
|||
| Germination | 0 | 9-13 | 23 | Low | Required |
| Germling | - | 10-15 | - | Low | Required |
| Appressorium | - | - | (not formed) | None | Required |
| Penetration | 2 | 9-13 | 23 | Low | Required |
| Growth | 3 | 12-15 | 20 | High | Not required |
| Sporulation | 5 | 12-15 | 20 | High | Not required |
Table 2.
Global wheat production and consumption during 2017-18 to 2023-24.
| Years | Production (Million metric tons) |
Consumption (Million metric tons) |
|---|---|---|
| 2017-18 | 761.54 | 742.37 |
| 2018-19 | 730.92 | 735.31 |
| 2019-20 | 759.39 | 745.71 |
| 2020-21 | 773.09 | 786.57 |
| 2021-22 | 780.05 | 791.16 |
| 2022-23 | 789.17 | 790.93 |
| 2023-24 | 784.91 | 796.44 |
Source: (Production of wheat worldwide 2023/24 | Statista; Total wheat consumption worldwide 2023/24 | Statista).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



