
Submitted:
12 May 2025
Posted:
13 May 2025
You are already at the latest version
Abstract
The tumor microenvironment (TME) plays a pivotal role in the initiation, progression, and therapeutic resistance of lung cancer. Comprising a complex network of immune cells, fibroblasts, endothelial cells, extracellular matrix components, and signaling molecules, the TME supports tumor survival and metastasis while suppressing anti-tumor immunity. In lung cancer, tumor-associated macrophages, cancer-associated fibroblasts, mast cells, and dendritic cells interact through cytokines, chemokines, growth factors, and matrix metalloproteinases to shape an immunosuppressive and pro-angiogenic milieu. Hypoxic conditions within the TME further enhance cancer cell adaptability through hypoxia-inducible factors (HIFs), promoting epithelial-mesenchymal transition, immune evasion, and metastatic potential. Moreover, microRNAs have emerged as key regulators of gene expression within the TME, offering novel insights into tumor behavior and potential targets for therapy. Targeting the dynamic interactions within the TME—especially through modulation of immune responses, angiogenesis, and stromal remodeling—offers promising avenues for precision pharmacology. This review summarizes current knowledge of the lung TME, highlighting its impact on cancer patho-physiology and treatment strategies. Understanding and therapeutically reprogramming the TME may pave the way for personalized and more effective interventions in lung cancer management.
Keywords:
tumor microenvironment
; lung cancer
; matrix metalloproteinase
; hypoxia
1. Introduction
The cellular environment in which the tumor exists is called the tumor microenvironment (TME) [1]. The tumor microenvironment (TME) comprises various cell types, including stromal, mesenchymal, endothelial, cancer-associated fibroblasts, and cancer cells with specific phenotypic and genetic characteristics [2]. In the lung tumor microenvironment, cellular elements (inflammatory cells, tumor cells, fibroblasts, vascular, structural, and vascular smooth muscle cells, and pericytic cells, among others), soluble (proteases, cytokines, and hormones, among others), are the main components [1]. Genetic alteration shifts in the tumour in oncogenes that modulate tumor growth and invasion into the surrounding tissue orchestrate the entity of anti-inflammatory infiltrates, which vary by size and form in diverse tumour types and different stages of tumor development. Within the tumors, features of the complex inflammatory cells have revealed cells involved in both the intrinsic and adaptive branches of the immunologic response among the infiltrate of lung cancer will help to find their role in tumour progression. This understanding will be vital to finding novel anticancer therapies, though the data gathered will take us nearer to individualized medicine, and give particular treatment to reprogram the tumor microenvironment to control disease [3]. Lung cancer remains the leading cause of cancer-related fatalities worldwide [4]. NSCLC accounts for approximately 85% of lung cancer cases [5]. Lung cancer is divided into two types: non-small cell lung cancer (NSCLC, 80-85%) and small cell lung cancer (10-15%). Adenocarcinoma (45-60%), squamous cell carcinoma (20-25%), and neuroendocrine carcinoma (10-15%) are all types of non-small cell lung cancer. These subgroups differ in treatment and prognosis: NSCLC is treated with targeted medicines and has a 5-year overall survival rate of 23%, whereas SCLC is treated mostly with platinum-based chemotherapy and has a median survival of less than one year [6]. Immune checkpoint blockade (ICB) immunotherapy is the first-line treatment for advanced non-small cell lung cancer (NSCLC) with no known driver-gene mutation [7]. Chemokines have emerged as important participants in cancer immunotherapy, coordinating immune cell signaling inside the TME and aiding their recruitment to cancer cells [8]. Coherent with their capability to react to local environmental cues, proinflammatory interleukins, and chemokines are present at high levels in the microenvironment of epithelial tumors [9,10]. The tumor microenvironment also contains T and B lymphocytes of adaptive immunity, and these phenotypes of the T and B subsets are regulatory and weaken the immunologic response against the tumor and evoke a chronic inflammatory state of the tumor microenvironment [11]. Several stimuli-responsive materials that degrade in the pathological tumor microenvironment (TME) have been produced and investigated for drug delivery applications employing nanotechnological methods [12]. In the setting of lung cancer and its complex tumor microenvironment (TME), where conventional treatments frequently confront problems such as drug resistance and immune evasion, traditional plant-based medicines are gaining popularity. Artocarpus chaplasha, a traditional medicine herb, has demonstrated promising antioxidant and cytotoxic activities [13]. Despite substantial research, the role of the tumor microenvironment (TME) in immunotherapy resistance is still unknown. Variable responses in NSCLC can be attributed to variables such as low MHC-I expression, inadequate neoantigen release, and restricted CD8+ T cell infiltration. Understanding these mechanisms is crucial for overcoming resistance [14]. In recent years, a growing study has highlighted the tumor microenvironment (TME), Riera-Domingo et al. offering a complete overview of its metabolic features and hypoxia, as well as their impact on immune function and response to immunotherapies [15]. Understanding the complicated interactions between tumor cells and TME components, such as immune and stromal cells, may lead to novel ways for better lung cancer management and therapy [16]. Therefore, it is crucial to understand the cellular and molecular interplay in the tumor microenvironment to develop therapeutic strategies. In this review, we focused on the cellular and molecular interplay in the lung tumor-microenvironment and discussed several cellular and molecular components that can be focused on while developing any therapeutics.
2. Tumor-Infiltrating Immune Cells
Tumor-infiltrating immune cells are surrounded by infiltrating inflammatory cells, especially macrophages and lymphocytes [17]. Evidence suggests that cancer malignancy is caused by both tumor-intrinsic characteristics and TME factors, particularly invading immune cells. Lung cancer avoids immunosurveillance by low antigenicity, decreased MHC I/II/non-classical expression, a lack of costimulatory signals, and dysregulated immune cell infiltration [18]. The development of tumor-specific adaptive immune responses is particularly driven by tumor antigens [19]. The main components of tumor-specific cellular adaptive immunity are two types of T lymphocytes (CD4+ and CD8+). In which tumor cells are attacked by the CD8+ T lymphocytes, introducing tumor-associated antigen peptide with histocompatibility complex class I (MHC I) by producing interferon-g on their surface. Mechanisms of Interferon-g-dependent tumor cell cytostasis and killing constitute cell cycle prohibition, angiostasis, apoptosis, and also induction of antitumorigenic activity of macrophages [20].

Table 1.
Major Tumor-infiltrating immune cells observed in the lung cancer tumor microenvironment, their main roles, and their relationships with clinical outcomes.
Table 1.
Major Tumor-infiltrating immune cells observed in the lung cancer tumor microenvironment, their main roles, and their relationships with clinical outcomes.
| Immune cell type | Main Function in TME | Clinical/Prognostic Association |
| CD8+ T cells | Cytotoxic killing of tumor cells | Improved survival, better ICI response[21],[22] |
| CD4+ T cells | Helper/regulatory roles; coordinate immune responses | Variable; subset-dependent [22,23] |
| Regulatory T cells (Tregs) | Suppress anti-tumor immunity | Poorer prognosis [22,24] |
| B cells | Antibody production, antigen presentation | Mixed; high density may predict HPD [21,25] |
| Macrophages (M1/M2) | M1: pro-inflammatory/anti-tumor; M2: immunosuppressive | M1: favorable; M2: poor prognosis [22,23,24] |
| Myeloid-derived suppressor cells (MDSCs) | Suppress T cell function, promote tumor growth | Poorer prognosis [22] |
| Natural Killer (NK) cells | Direct killing of tumor cells (innate immunity) | Generally favorable [23] |
| Dendritic cells (DCs) | Antigen presentation, T cell activation | It can be immunosuppressive in TME [24] |
| Mast cells | Modulate inflammation, angiogenesis | Prognostic value in LUAD [23] |
2.1. Tumor-Infiltrating Lymphocytes (TIL)
TIL density, distribution, and phenotypic characteristics are important indicators of responsiveness to immune checkpoint inhibitors in lung cancer. TIL subsets (CD4⁺, CD8⁺, and CD19/20⁺) have both effector (anti-tumor) and suppressive (pro-tumor) roles that are influenced by the tumor environment. Their balance ultimately determines disease progression and immunological status.
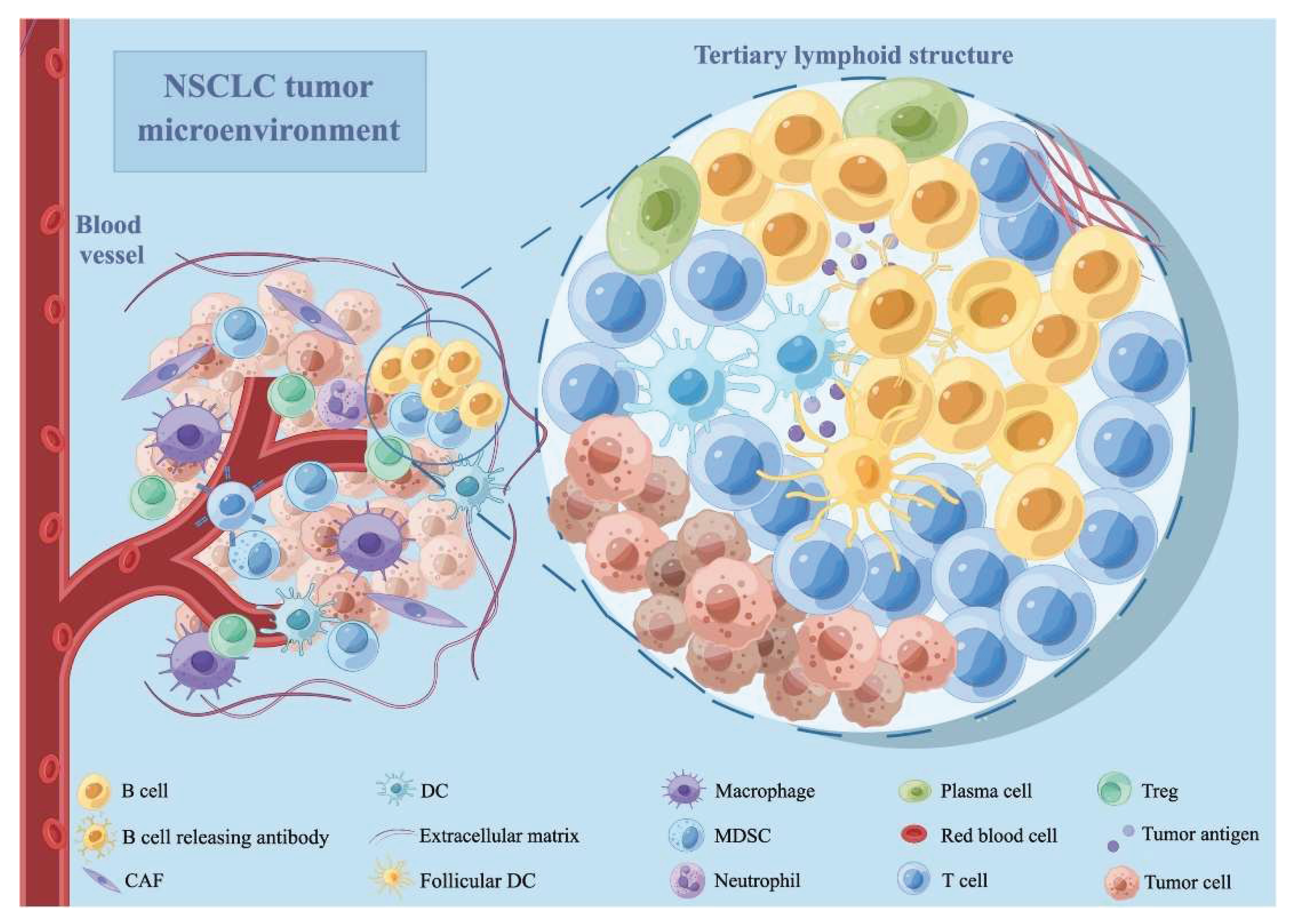
Figure 1.
The tumor microenvironment of NSCLC. CAF stands for cancer-associated fibroblast; DC is a dendritic cell; MDSC is a myeloid-derived suppressor cell; NSCLC is a non-small cell lung cancer; and Treg is a regulatory T cell. The figure is reprinted from Wang, Fen, et al. 2022 with free reuse permission [26].
Figure 1.
The tumor microenvironment of NSCLC. CAF stands for cancer-associated fibroblast; DC is a dendritic cell; MDSC is a myeloid-derived suppressor cell; NSCLC is a non-small cell lung cancer; and Treg is a regulatory T cell. The figure is reprinted from Wang, Fen, et al. 2022 with free reuse permission [26].
2.1.1. Cytotoxic CD8+ T Lymphocytes
CD8+ T cell density is associated with increased overall survival (OS) in lung cancer. CD8⁺ T cells play a crucial role in lung cancer immunology, and increased intertumoral concentrations correspond with improved survival rates [27]. However, cancers avoid these cytotoxic cells by:
Exhausted CD8⁺ TILs expressing multiple checkpoints resist ICI therapy. However, tumors rich in PD-1⁺CD8⁺ cells ("hot" TMEs) frequently respond better to PD-1 blocking [32]. Responders have enhanced gene expression profiles with memory/effector signatures, while non-responders have dysfunction-related genes.
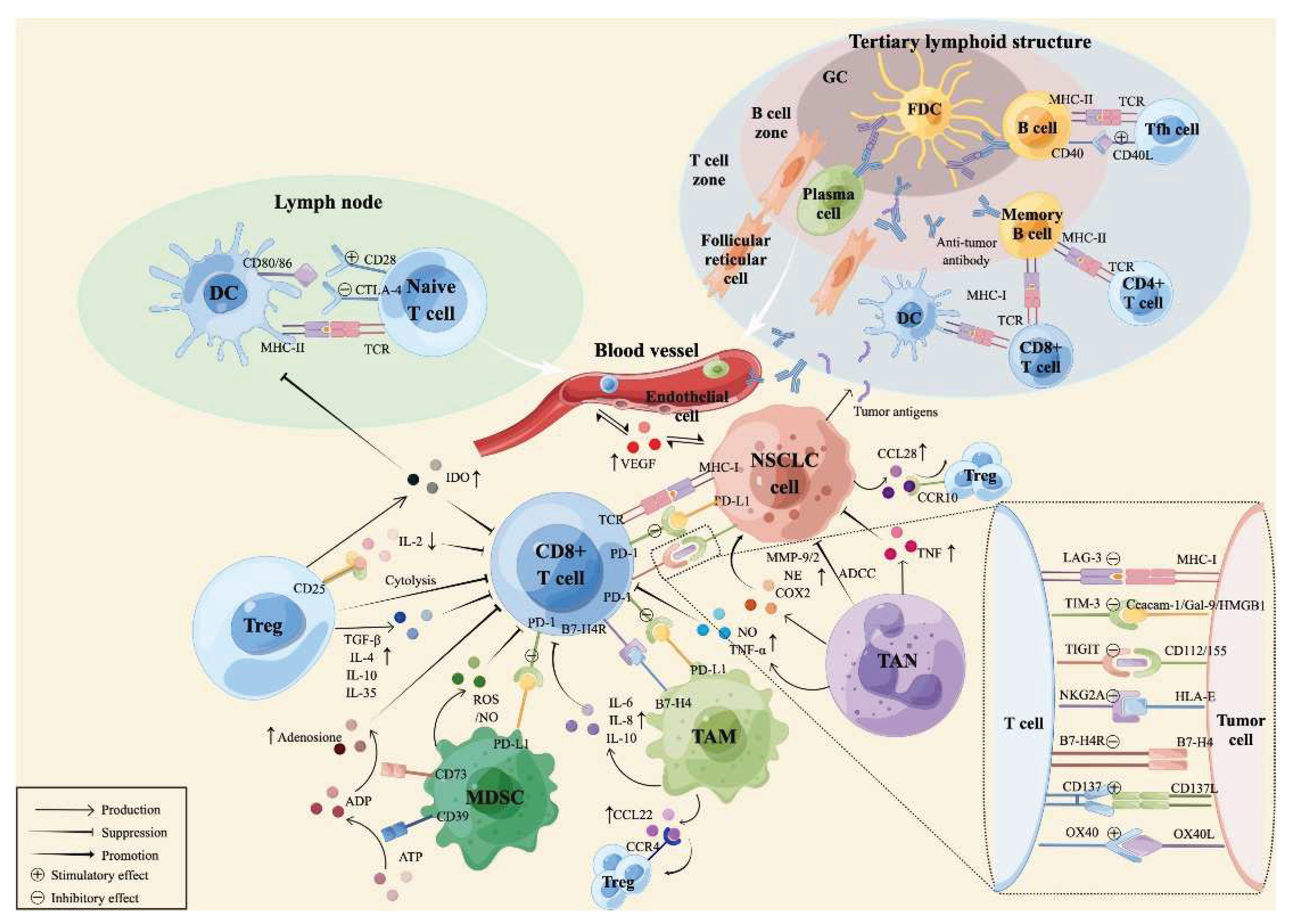
Figure 2.
The tumor microenvironment of NSCLC. CAF stands for cancer-associated fibroblast; DC is a dendritic cell; MDSC is a myeloid-derived suppressor cell; NSCLC is a non-small cell lung cancer; and Treg is a regulatory T cell. The figure is reprinted from Wang, Fen, et al. 2022 with free reuse permission.
Figure 2.
The tumor microenvironment of NSCLC. CAF stands for cancer-associated fibroblast; DC is a dendritic cell; MDSC is a myeloid-derived suppressor cell; NSCLC is a non-small cell lung cancer; and Treg is a regulatory T cell. The figure is reprinted from Wang, Fen, et al. 2022 with free reuse permission.
3. Tumor Expressing Cytokines
Cytokines—membrane-bound or released proteins produced by innate and adaptive immune cells in response to tumor antigens and pathogens—are responsible for immunological homeostasis. Their activities are dependent on local concentration, receptor expression, and pathway integration. Cytokines have an important role in tumor immunity, as evidenced by higher tumor incidence in animals lacking type I or II interferon receptors or downstream signaling components [33]. Cytokines signal through a series of common and shared receptors, there are seven types of Cytokines signal through a series of common and shared receptors, of which Type I and II, Type III cytokine receptor families have the most promising clinical potential. Type I includes IL-2, IL-4, IL-7, IL-9, IL-15, and IL-21; they initiate intracellular signals through the coordinated activity of signal transducers of activated T molecule and Janus kinases 1 and 3 [34].
| Cytokine | Primary Source | Target Cells | Primary Function |
| IL-1 | Monocytes, macrophages, fibroblasts | T/B cells, endothelium, hypothalamus | Co-stimulation, inflammation, fever |
| IL-2 | T cells, NK cells | T/B/NK cells, monocytes | Growth and activation of immune cells |
| IL-4 | T cells | T/B cells | Th2 differentiation, IgE switching |
| IL-6 | T cells, macrophages | T/B cells, liver | Acute phase response, inflammation |
| IL-10 | Th2 cells | Macrophages, T cells | Anti-inflammatory, suppresses APCs |
| IL-12 | Macrophages, NK cells | T cells | Promotes Th1 differentiation |
| IL-17 | NKT cells, ILCs | Epithelial, endothelial cells | Inflammation, infection control |
| IL-21 | CD4+ T cells, NKT cells | T/B/NK cells | Enhances immune responses |
| IL-23 | APCs | T cells, NK cells | Promotes chronic inflammation via Th17 |
| IFN-γ | T, NK, NKT cells | Monocytes, endothelial cells | MHC upregulation, macrophage activation |
| TNF-α | Macrophages, T cells | Immune, endothelial, liver cells | Inflammation, fever, acute-phase response |
| TGF-β | T cells, macrophages | T cells | Suppresses immune activation |
| IL-35 | Tregs | T cells | Immunosuppressive, induces iTr35 |
| IL-37 | Monocytes, DCs | Macrophages, B cells | Dampens excessive inflammation |
Contributions of oncostatin receptor and leukemia inhibitory factor receptor to signal transduction in heterodimeric complexes with glycoprotein 130. Type II Cytokine Receptors include IFN-α/β, IFN-γ, IL-10, IL-20, IL-22, IL-28; they mediate a signaling chain and a ligand binding chain[36]. Recently-discovered type III family contains IFN-1, IFN-2, and IFN-3 that activate an IL-10 receptor 2 and IL-28 receptor subunit complex[37]. This may be a vital subgroup in the future, but at this time, there are no established therapeutic uses for Type III IFNs. JAK-STAT pathways lead to the development of malignant cell growth, survival, and death and occur by different combinations of JAK kinases and their substrate [38].
4. Lung Tumor Microenvironment
Lung cancer is identified and characterized by uncontrolled proliferation of cells inside the lung; most commonly, epithelial cells are known as carcinomas. Metastasis is responsible for >70% of deaths in Europe and the USA, whereas lung cancer is placed in the leading position of cancer-related death. Most patients with last-stage lung cancer die within 18 months of diagnosis [39]. The lung tumor microenvironment is characterized by vascularization and oxygenation. Cigarette smoking is treated as a significant risk factor for the growth of lung cancer. Cigarette smoke results in an inflammatory reaction within the alveolar inflammatory cells of the lungs, and the changed secretion of cytokines predisposes towards lung cancer [40]. The development of lung cancer and tissue preferences for metastasis result from the interaction between tumor cells and stroma and reflect the migration of cancer cells to release chemoattractants instead of propagating cancer cells that may have the power to last in specific tissue microenvironments. Several stages are involved in the metastasis of epithelial cancers [41], First local tumor invasion through the cellar membrane and stroma [42], Intravasation into the lymphatic system [43], then the tumor cells survive within the circulation [44], Grab at the detached tumor site [45], speed up the development of circulating tumor cell microemboli [46], extravasation into the detached tissue microenvironment which is facilitated by tumor-cell secreted factors [47], tumor cells initially survive in the detached tumor stromal environment, responsible for the formation of a ‘‘pre-metastatic niche’’[48,49], modifying growth of cancer-cell intrinsic events at distant sites, and the development of macro metastases [32,50]. The above steps exist as a barrier to dissemination, making the overall metastatic process inefficient. The substantial latency period between primary diagnosis and subsequent formation of distant metastasis is a general observation within epithelial carcinomas. It is proposed that cells that depart the primary tumor are sufficiently well adapted to survive at distinct metastatic sites so that the latency period of the lung does not exhibit [51]. This may also be considered the relatively late stage of almost all types of lung cancer diagnosis.
5. Cells of the Stroma
5.1. Fibroblast Cells
In the late 19th century, based on the location and the microscopic appearance, fibroblasts were first described [52]. Fibroblasts are prolonged cells with extensive cell processes that show a fusiform or spindle-like shape in visibility. Deposition of extracellular matrix (ECM), regulation of epithelial differentiation, and regulation of inflammation are the crucial functions of fibroblasts [53,54]. Many of the constituents of the fibrillar ECM, such as type I, type III, and type V collagen, and fibronectin synthesized by fibroblasts [55]. These cells have a well-perceived role in the carcinogenic process. Stromal fibroblasts dominate tumor biology, but the exact mechanisms involved are still unclear. Fibroblasts in mammals are highly disparate, different sites reflect a substantial topographic diversity disjunct from those cells [56]. A particular type of stromal cells known as cancer-associated fibroblasts (CAFs) are myofibroblasts that express α-smooth muscle actin (α-SMA) and produce collagen and other ECM proteins. CAFs are now recognized for their function in epithelial tumor progression, growth, and metastasis by secreting substances that promote angiogenesis, cancer cell proliferation and invasion, macrophage recruitment, and T-cell-mediated immune suppression [57,58]. The functions of immune cells, tumor cells, and endothelial cells are regulated by CAFs, a poly-functionality that could be affected by the entity of multiple CAF lineages with distinguished intra-tumoral functions. In support of this decision, lung cancer patient prognosis is improved by the presence of PDGFR-α/β+ CAFs and aggravated by extracellular matrix (ECM) proteins commonly released by myofibroblastic CAFs. Because elements released by CAFs can increase tumor cell proliferation and invasion, encourage angiogenesis, and inhibit anti-tumor immunity, CAFs are a promising target for cancer prevention and treatment [59]. Immunotherapy is typically ineffective in some NSCLC patients, owing to a lack of CD8+ T cell penetration into tumors. CAF-rich tumors contribute to this by forming a physical barrier and promoting an immunologically cool milieu. CAFs also decrease CD8+ and CD4+ T cell activity, promote Treg differentiation, and recruit MDSCs, all of which aid in tumor growth and resistance to treatment. Thus, targeting CAFs has potential in improving immunotherapy responses in solid malignancies [60].
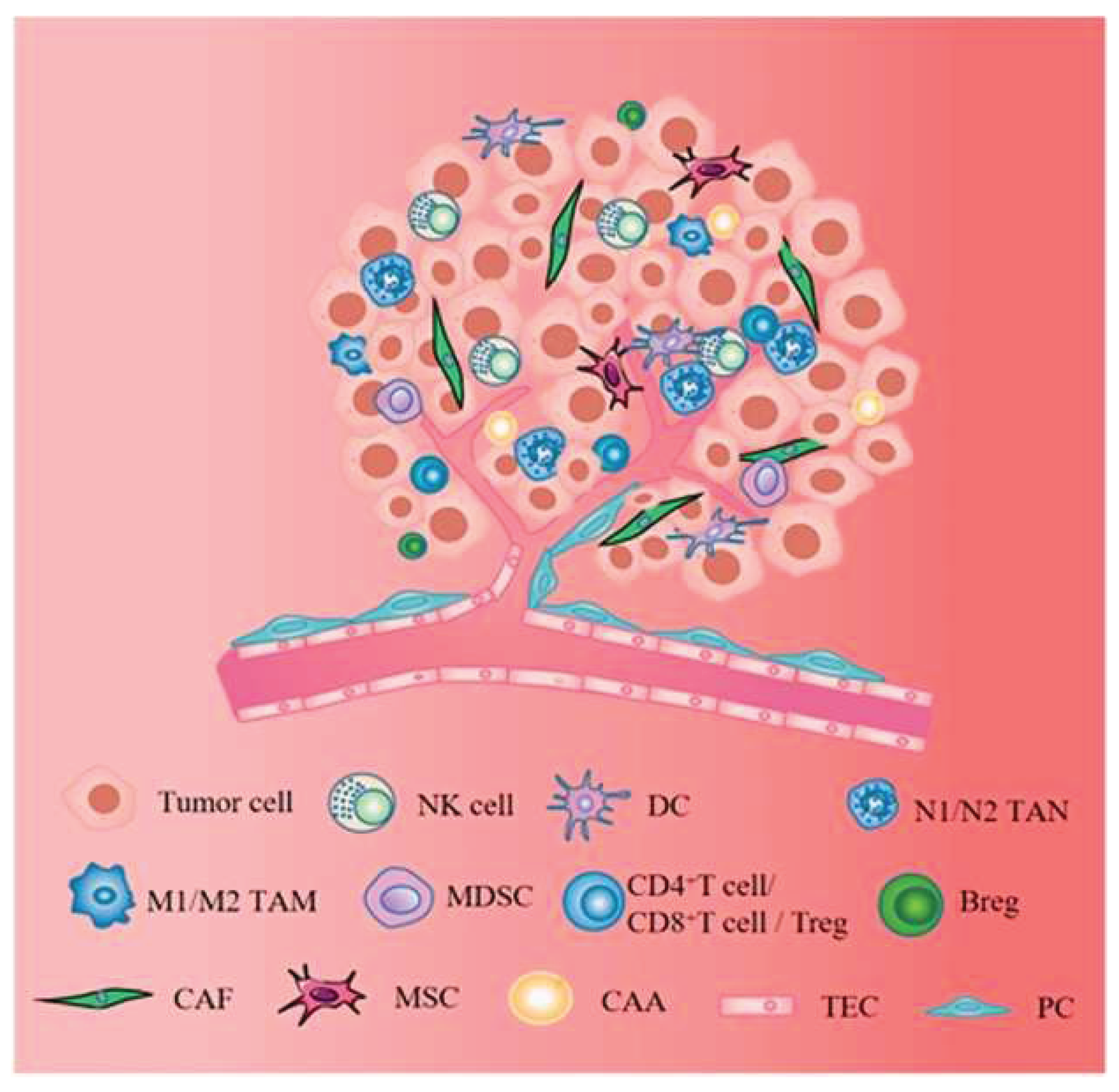
Figure 3.
Stromal cells, including CAFs, MSCs, CAAs, TECs, and PCs, establish complex signaling networks that regulate tumor development, growth, and resistance to treatment. The figure is reprinted from Zhao, Yan, et al. 2023 with free reuse permission [61].
Figure 3.
Stromal cells, including CAFs, MSCs, CAAs, TECs, and PCs, establish complex signaling networks that regulate tumor development, growth, and resistance to treatment. The figure is reprinted from Zhao, Yan, et al. 2023 with free reuse permission [61].
5.2. Immune Cells
Tumor cells induce angiogenesis and can increase endothelial cell proliferation, supporting tumor growth. Immune cells play a dual role in tumor progression, initially attacking tumor cells via cytokine production before being co-opted by tumors to boost growth and spread. Infiltrating immune cells are essential components of the tumor microenvironment, which tumor cells can use to cause immunological dysfunction and pro-inflammatory cytokine production. Exosomal miRNAs facilitate intercellular communication, which contributes to these activities [62]. Immune cells play a role in cancer dissemination that was predicted by Ehrlich in 1909; significant research has been conducted to question, re-judge, or approve this theory. In the past century, in vivo and in vitro work has shown that several types of cancer tissues, along with breast cancer, lung cancer, and colorectal cancer tissue, hold higher divides of unmarked immune cells than the normal tissue likeness, suggesting that the immune system acts very closely with the tumor microenvironment [63].
5.2.1. T-Cells
The class of immune cells, which represent the key actors of the adaptive immune response, is the T-cells. The tumour microenvironment (TME) can bind antitumoural T cell reactions in respective ways, including immunomodulation and impaired antigen presentation. Several cancer models in mice, antitumoural T cell responses, and T reg cell depletion have been shown to restrain tumour growth can be inhibited by T reg cells. T cell–attained Areg, likely by acting on normal cells in the TME, aided the increase of arising tumors in the lungs. The detected effect on tumor growth was not related to alterations in the number of intratumoral T cells or their capability to produce proinflammatory cytokines, proposing that neither pan–T cell lack in Areg nor its selective loss in T reg cells had immunomodulatory effects on the TME [64]. Regulatory T cells (Tregs) can infiltrate the tumor site and inhibit the function of effector T cells, which identify and destroy cancer cells. Tregs contribute to the formation of an immunosuppressive environment, allowing tumor cells to elude immune surveillance and accelerate tumor development. Inflammatory factors serve crucial roles in disease [65,66].
5.2.2. Macrophages
Macrophages account for the majority of the inflammatory infiltration in malignancies [67]. In inflammatory cells and the wound healing process, macrophages are the main component, which are derived from the monocytes later on, and they go into the tissue from the circulation. Macrophages are a significant part of the body’s primary immune response to infection and have also manifested antitumour action under normal conditions. TAM signaling promotes vascularization, invasiveness, growth, cell survival, and immunosuppression, all resulting in continued tumor progression, when an immune injury-healing response is misplaced [68]. The macrophages that infiltrate into the TME are known as TAMs. TAMs are often observed in close proximity to CAFs in various tumor types, proposing interactions between these two cell types. The cleavage of type I collagen by FAP derived from CAFs can activate macrophages [69]. Most malignancies have a high proportion of TAMs with the M2 phenotype, which is associated with a poor prognosis. These M2-like TAMs help tumors survive, grow, and spread by encouraging angiogenesis, epithelial-to-mesenchymal transition, and immune suppression [70,71]. Although M1 macrophage infiltration is uncommon in malignancies, it has been observed in colorectal cancer, where their presence is associated with a better prognosis, even when M2 macrophages are more plentiful. (TAMs) is modified by the tumor's developmental stage as well as the microenvironment. Tumor advancement may result from the interaction of intrinsic (genetic) and extrinsic (microenvironmental) mechanisms, in which one initiates tumorigenesis and then drives the other to advance malignancy [72,73].
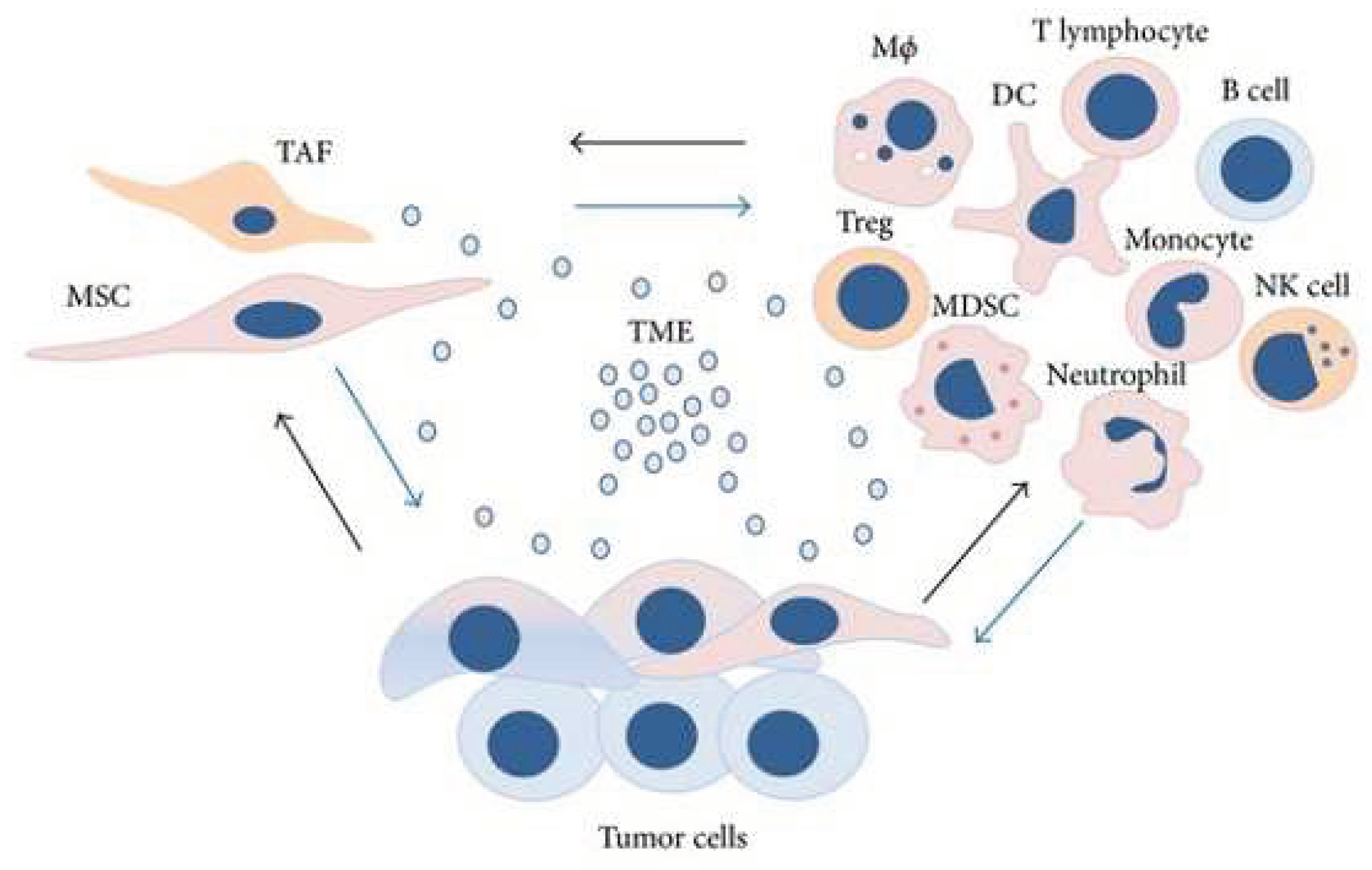
Figure 4.
The tumor microenvironment (TME) provides a dynamic and supportive environment in which various cell types constantly modify their phenotypic and functions. It is made up of a complex mix of mesenchymal stromal/stem cells (MSCs), tumor-associated fibroblasts (TAFs), and a variety of immune cells, including macrophages, regulatory T cells (Tregs), myeloid-derived suppressor cells (MDSCs), natural killer (NK) cells, dendritic cells (DCs), monocytes, neutrophils, T lymphocytes, and B cells, as well as a diverse population of tumor cells. The figure is reprinted from Trivanović, Drenka, et al. 2016 with free reuse permission [74].
Figure 4.
The tumor microenvironment (TME) provides a dynamic and supportive environment in which various cell types constantly modify their phenotypic and functions. It is made up of a complex mix of mesenchymal stromal/stem cells (MSCs), tumor-associated fibroblasts (TAFs), and a variety of immune cells, including macrophages, regulatory T cells (Tregs), myeloid-derived suppressor cells (MDSCs), natural killer (NK) cells, dendritic cells (DCs), monocytes, neutrophils, T lymphocytes, and B cells, as well as a diverse population of tumor cells. The figure is reprinted from Trivanović, Drenka, et al. 2016 with free reuse permission [74].
5.2.3. Mast Cells
Paul Ehrlich first described mast cells (MCs) as versatile, tissue-homing secretory cells. He reported that MC was enhanced in chronically inflamed tissues, assuming they were providing nutritional aid to damaged cells [75]. Their distribution all over vascularized tissues, revealed to the exterior environment, including the lungs, provides interaction with environmental antigens, invading pathogens, or toxins [76]. Mast cells (MCs) are innate immune cells that reside at tumor edges and in the tumor microenvironment (TME), frequently near blood vessels. MCs, which differ from bone marrow progenitors, circulate in the blood and are guided to certain organs by chemoattractive signals. Several cytokines, including stem cell factor (SCF), CXCL12, IL-3, IL-4, IL-9, IL-10, IL-33, and TGF-β, influence their survival and proliferation. While MCs have generally been examined in the context of allergy reactions, current research has revealed that they can operate as proinflammatory and angiogenic mediators in malignancies. When activated, MCs produce mediators that attract immune cells such as neutrophils, macrophages, eosinophils, and B and T cells, thereby contributing to antitumor immune responses [77]. Mast cells (MCs), with their dual involvement in inducing inflammation and angiogenesis inside the tumor microenvironment (TME), are intriguing targets for adjuvant cancer therapy. Strategies may include suppressing angiogenesis and tissue remodeling, restricting the production of tumor-promoting proteins, and reversing MC-driven immune suppression—all while increasing their ability to produce cytotoxic cytokines, which boosts anticancer effectiveness [78].
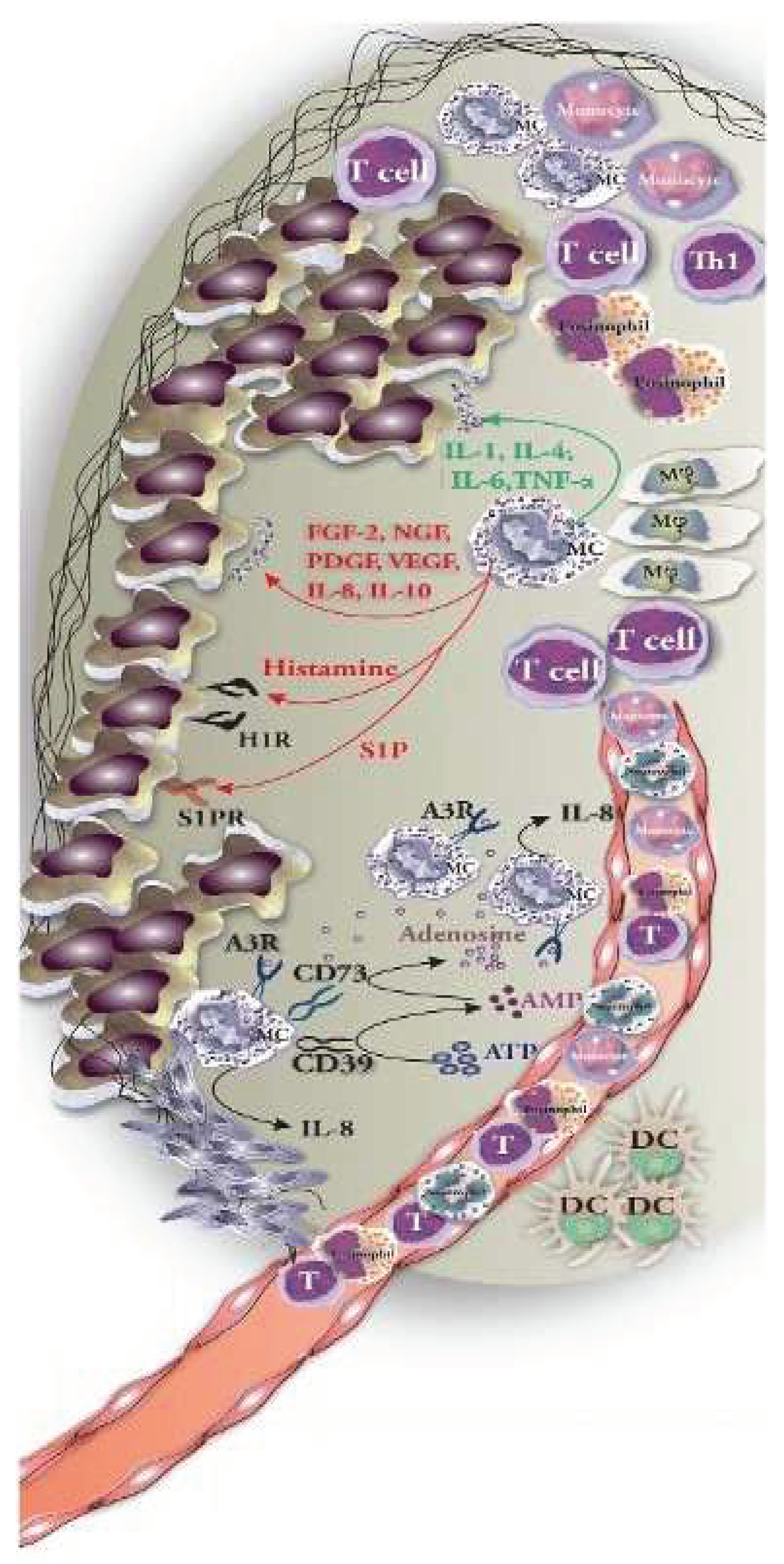
Figure 5.
Mast cells (MCs) engage with tumor cells by releasing mediators and making direct cell-to-cell contact. MC-derived mediators can have dual effects: some promote tumor growth (e.g., histamine, VEGF, IL-10—shown in red) while others repress tumors (e.g., TNF-α, IL-1, granzyme B—shown in green). Furthermore, direct interactions with immune and tumor cells can either activate or repress cellular responses, impacting tumor development or immunological activation, depending on the situation. The figure is reprinted from Komi & Redegeld et al. 2020 with free reuse permission [77].
Figure 5.
Mast cells (MCs) engage with tumor cells by releasing mediators and making direct cell-to-cell contact. MC-derived mediators can have dual effects: some promote tumor growth (e.g., histamine, VEGF, IL-10—shown in red) while others repress tumors (e.g., TNF-α, IL-1, granzyme B—shown in green). Furthermore, direct interactions with immune and tumor cells can either activate or repress cellular responses, impacting tumor development or immunological activation, depending on the situation. The figure is reprinted from Komi & Redegeld et al. 2020 with free reuse permission [77].
5.2.4. Dendritic Cells
Dendritic cells (DCs) are responsible for starting, maintaining, and directing anti-tumor immune responses. They detect DAMPs generated by dying tumor cells, which cause cytokine production and T-cell activation [79]. Dendritic cells (DCs) take up antigens and move to lymph nodes to deliver them to CD8⁺ T lymphocytes. DCs are divided into two types: cDC1s, which deliver antigens to CD8⁺ T cells, and cDC2s, which trigger CD4⁺ T cell responses [80,81]. Dendritic cells (DCs) are classified as conventional type 1 (cDC1s) or type 2 (cDC2s). cDC1s promote anti-tumor immunity by delivering antigens to CD8⁺ T cells, while cDC2s focus on priming CD4⁺ T cell responses. cDC2s are classed as anti-inflammatory cDC2A (T-bet⁺) or pro-inflammatory cDC2B (T-bet⁻) based on their transcriptional and chromatin patterns. High cDC1 infiltration correlates with increased T cell presence and better patient outcomes. Tumors may avoid immune recognition by preventing cDC1 penetration into the tumor microenvironment [82,83]. cDC1s use the C-type lectin receptor DNGR-1 to identify F-actin on necrotic cells, which activates CD8⁺ T lymphocytes by antigen absorption and cross-presentation. However, tumor-secreted gelsolin (sGSN) can disrupt this mechanism by preventing DNGR-1-dependent cross-presentation, reducing the immune response [84,85]. Plasmacytoid dendritic cells (pDCs) produce significant quantities of interferon-α (IFN-α), similar to plasma cells. In breast and ovarian cancer patients, malfunctioning pDCs produce less type I IFN and enhance regulatory T cell (Treg) development, which contributes to tumor progression. Understanding how DCs differentiate in the tumor microenvironment is still an active topic of research, with important implications for future immunotherapies [86].
5.2.5. Vascular Cells
To meet the metabolic and nutritional demands for growth, Tumors require the formation of a complex vascular network. VEGF is the main component involved in the formation of tumor vessels. It is released by the tumor cells directly and by fibroblasts and inflammatory cells in the stroma and is responsible for the ‘‘angiogenic switch’’ where new vasculature is shaped to provide the tumor with nutrients. Tumor vessels formed as a result of VEGF are abnormal; they are non-uniformly distributed and irregularly formed, unsuitably branched and tortuous, often terminating blindly. They do not have the classic hierarchical placement of arterioles, venules, and capillaries and often form arteriovenous shunts. These vessels are variably fenestrated and leaky, leading to high interstitial pressures, further exacerbating tissue hypoxia and stimulating additional VEGF production [87,88]. Under the dominance of VEGF, tumour vessels are shaped by one of various mechanisms, including the beginning of being vascular networks, recruitment of vascular progenitor cells to form new vascular channels, or ‘‘vascular mimicry’’—a process by which tumor cell-lined channels contribute to the blood tributaries supplying the tumor [89].
6. Extracellular Molecules
6.1. Cytokines
Given the limits of current lung cancer treatments, developing immune-based alternatives is critical. TILs in the tumor microenvironment (TME), including lymphocytes and macrophages, release low-molecular-weight cytokines and chemokines (<30 kDa) that govern cellular activities like metabolism, proliferation, tissue repair, and chemotaxis. These chemicals act on specific cell receptors to facilitate intercellular communication via autocrine, paracrine, and endocrine signaling [90,91]. Not only do cytokines and chemokines orchestrate immune responses, but they also drive local and systemic inflammation. They have a substantial impact on tumor growth, metastasis, and therapeutic resistance in the tumor microenvironment (TME). Several cytokines, including IL-6, IL-10, IL-17, IL-27, IL-35, TNF-α, IFN-γ, and TGF-β, and chemokines, such as CCL-2, CCL-5, CCL-18, CCR4, CXCR4, CX3CL1, CXCL-1, CXCL-5, CXCL-8, and CXCL-13, are extensively explored as therapeutic targets and biomarkers in lung cancer therapy techniques [92]. Lung tumor-derived prostaglandin E2 (PGE2) regulates immunological responses by increasing IL-10 synthesis from lymphocytes and macrophages while decreasing IL-12 production by macrophages. An immunohistochemical examination of human non-small cell lung cancer (NSCLC) tissues showed cytoplasmic COX-2 expression within tumor cells. This is the first report of functional COX-2 expression by NSCLC cells, and it identifies a method via which high COX-2 and PGE2 levels affect cytokine balance, changing the immunological landscape of the lung cancer microenvironment [93].
Table 3.
Cytokines with their source and functions [94].
Table 3.
Cytokines with their source and functions [94].
| Cytokine | Source | Functions |
| IL-6 | T-cells, macrophages, adipocytes | Proinflammatory action, promotes differentiation and cytokine production |
| IL-8 | Epithelial cells, macrophages, endothelial cells | Proinflammatory action, promotes angiogenesis and chemotaxis |
| IL-10 | Monocytes, B-cells, T-cells | Anti-inflammatory action, inhibits proinflammatory cytokines |
| IL-17 | Th17 cells | Proinflammatory action, enhances cytokine and chemokine production, contributes to antitumor immunity |
| IL-27 | Antigen-presenting cells (APCs) | Anti-inflammatory action, induces IL-10 production |
| IL-35 | Regulatory T-cells (Tregs) | Anti-inflammatory action, promotes Treg proliferation, suppresses Th17 cells |
| IL-37 | NK cells, monocytes, epithelial cells, B-cells | Anti-inflammatory, antimicrobial, and contributes to antitumor immunity |
| TNF-α | Macrophages, CD4+ lymphocytes, adipocytes, NK cells | Proinflammatory action, induces cell proliferation, cytokine production, and apoptosis |
| IFN-γ | NK cells, T-cells | Antiviral and proinflammatory action |
| TGF-β | T-cells, macrophages | Anti-inflammatory action, suppresses proinflammatory cytokine production |
| Granulocyte-macrophage colony-stimulating factor (GM-CSF) | T-cells, macrophages, fibroblasts | Proinflammatory action, enhances neutrophil and monocyte function, activates macrophages |
| Vascular endothelial growth factor (VEGF) | Macrophages, endothelial cells, platelets | Promotes vasculogenesis, angiogenesis, endothelial chemotaxis, and migration |
6.2. Growth Factors
Epidermal growth factor is a transmembrane glycoprotein. The growth of cultured benign and malignant cells is promoted by the epidermal growth factor (EGF). Recent studies have manifested that the amount of EGF receptor, when compared with normal epidermal cells, is increased in squamous cell carcinoma cells in tissue culture [95]. Epidermal growth factor (EGFR) is a receptor tyrosine kinase recognized as being highly expressed in cancer cells, including lung cancers. These trans-membrane proteins are activated complying binding with peptide growth factors of the EGF-family of proteins. It has been distinguished that mechanisms might contribute to amplifying the signal obstructed by growth factors, as expression of a high number of receptors on the surface of tumour cells can increase their sensitivity to low concentrations of host or tumour-derived growth factors. A direct correlation also exists between growth factors and cellular proto-oncogenes [96]. Many lines of manifest suggest that EGFR has relevance to patients with NSCLC and thus may serve as a potential therapeutic target. Expression of EGFR has been observed by immunohistochemistry testing in from 62% to 93% of resected primary tumors, and EGFR mRNA has been found in 100%. The overexpression of EGFR has been variably correlated with clinical outcomes [97].
6.3. Matrix Metalloproteinase
Growing evidence indicates that extracellular proteinases, notably matrix metalloproteinases (MMPs), play an important role in mediating microenvironmental alterations during tumor growth. These enzymes govern a wide range of physiological activities and signaling pathways, making them critical to the molecular interactions that occur between tumor cells and the stroma [42]. Matrix metalloproteinases (MMPs) are a class of zinc-dependent endopeptidases discovered about 50 years ago. They are required for a variety of physiological activities, including tissue remodeling, wound healing, and organ development, as they degrade extracellular matrix components and modulate cell activity [98,99]. Certain MMPs have tumor-suppressive roles, explaining the limited success of broad-spectrum MMP inhibitors (MPIs) in cancer therapy. For example, MMP-8 deficiency increases cancer risk, and macrophage-derived MMP-12 suppresses lung metastases by modulating tumor vasculature. Additionally, MMPs can exert non-proteolytic effects via domains like hemopexin, which are not targeted by typical MPIs, reducing their therapeutic efficacy [42].
7. Immune Regulation by Stroma
Immune system dysfunction is strongly associated with NSCLC, and immune checkpoint inhibitors are currently important second-line treatments following chemotherapy failure. Treatments for lung cancer that target CTLA-4, PD-1, and PD-L1 have demonstrated efficacy. Pembrolizumab increased progression-free and overall survival in NSCLC patients, but TG4010 with chemotherapy improved PFS in advanced cases [100]. Immune cells of a stoma include monocytes/macrophages, neutrophils, and lymphocytes, which enter into and occupy the tumor stroma. Monocytes are actively recruited into tumors along defined chemotactic gradients. Once in the tumor, they differentiate into tumor-associated macrophages (TAMs). TAMs seem to be preferentially pulled to and continued in areas of necrosis and hypoxia, where they become phenotypically changed and upregulate hypoxia-induced transcription factors. Macrophages also release several factors that influence endothelial cell behavior, including VEGF, HGF, MMP2, and IL-8. Neutrophils are distinguished as angiogenesis stimulators by releasing VEGF, HGF, MMP2, and IL-8. Additional immune cell populations have a less well-authenticated part in carcinogenesis and are not consistent occupants of stroma, with their presence limited to specific types of tumors. These include myeloid suppressor cells, which have the phenotypic features of some macrophages and granulocytes and various consequences of immune suppression, a product of MMP9 and VEGF, and the extra power to immediately incorporate into vessel walls [101,102].
8. Hypoxia and Tumour Microenvironment
Hypoxia is a common and significant characteristic of solid tumors. Hypoxic tissue is defined as having an oxygen tension of less than 10 mmHg, as opposed to 40- 60 mmHg in most normal tissue [103]. Cancer-related inflammation contributes to tumor initiation and progression by boosting genomic instability, cell proliferation, angiogenesis, apoptosis resistance, and metastasis. Hypoxia also contributes by allowing tumor cells to evade immune attack and avoid immunosurveillance. Hypoxia-inducible factors (HIFs), which are essential for hypoxic signaling, control genes implicated in tumor immune responses in low-oxygen environments [104]. Often achieved by induction of the hypoxia-inducible factor (HIF) family of transcription factors is the mechanism behind these effects. This family includes three members, HIF-1, -2, and -3, which act to govern cellular processes involved in glucose metabolism, angiogenesis, cell proliferation, and tissue remodeling in response to low oxygen levels. A group of prolyl-4-hydroxylases (PHDs) hydroxylate HIF-1α on two maintained residues, proline 402 and proline 564, under normal oxygen conditions [105,106,107]. One strategy is to ameliorate drug delivery, thus enhancing drug accumulation within the tumor. Several treatment techniques for hypoxia have been developed. One technique is to directly provide oxygen to the tumor using technologies such as the catalytic breakdown of endogenous hydrogen peroxide (H2O2) and light-triggered water splitting [108]. Several ongoing clinical trials are investigating hypoxia-targeted therapeutics employing a variety of medicines with distinct mechanisms. HIF inhibitors, including PX-478 and LW6, constitute an important class of therapeutic medicines. Studies have revealed that HIF inhibitors, either alone or in conjunction with other therapies, offer promising anti-tumor properties [104].
9. Role of microRNAs in Regulating Tumor Microenvironment
MicroRNAs (miRNAs) are tiny, noncoding RNAs that control gene expression by degrading mRNA or impeding translation. Dysregulated miRNA expression is associated with cancer initiation, progression, and metastasis. Some upregulated miRNAs serve as oncogenic drivers (oncomiRs), whereas downregulated miRNAs operate as tumor suppressors. Aberrant miRNA expression can interfere with cellular processes such as apoptosis, cell proliferation, invasion, metastasis, and treatment sensitivity, all of which contribute to tumor development [109]. According to studies, a single miRNA can bind to over 200 target genes, altering processes such as transcription, receptor activation, and transport. This intricacy makes it difficult to determine the exact transcripts and pathways controlled by individual miRNAs [110]. MicroRNAs have been depicted to be involved in several cellular processes, including proliferation, development, metabolism, differentiation, and apoptosis. Most importantly, pathological conditions involving cancer have been associated with deregulation of microRNA expression and role [111,112]. In cancer, microRNAs have been separated into oncogenic or tumor suppressor miRNAs. In the first instance, microRNAs assume an oncogenic function by post-transcriptionally lowering mRNA and, subsequently, protein levels of molecules with tumor suppressor functions. Despite the enormous quantity of knowledge we already have on microRNAs, new publications continue appearing, depicting yet other novel purposes, making us wonder about the wide-reaching possibility of these small molecules. In summary, microRNAs seem to be a very promising research target; however, the complexity of their activities makes keeping this promise challenging [113].
Table 4.
microRNA expression status in lung cancer.
| microRNA | Targets | Expression status (Under-expressed/Over-expressed/ Unchanged) |
Comments (if any) |
Reference |
|---|---|---|---|---|
| miR-487b | SUZ12,BM11, MYC | Over-expressed | Tumour suppressor | Xi et al, 2013 [114] |
| miR-449 | HDAC1 | Over-expressed | AM Rusek et al;2015 [113] | |
| miR-101 | EZH2 | Over-expressed | AM Rusek et al;2015 [113] | |
| miR-486 | IGF1R | Under-expressed | NSCLC-Tumour suppressor | C M. Croce et al;2013 [115] |
| miR-9 | MHC 1 gene | Over-expressed | AM Rusek et al;2015 [113] | |
| miR-124a | CDK6 | Over-expressed | Tumour suppressor | A Lujambio et al;2007 [116] |
| miR-221 | TIMP3 | Over-expressed | AM Rusek et al;2015 [113] | |
| miR-222 | TIMP3 | Over-expressed | AM Rusek et al;2015 [113] | |
| miR-429 | ZEB1/2 | Over-expressed | NSCLC- Oncogenic | Wu Cl et al;2018 [117] |
| miR-128b | EGFR in NSCLC | Under-expressed | Tumour Suppressor | Becker-Santos DD et al 2012 [118] |
| miR-1827 | SK-LU-1, RBX1 in NSCLC | Under-expressed | NSCLC- Tumour suppressor | SM Noor et al;2018 [119] |
| miR-378 | RBX1, CRKL in NSCLC | Over-expressed | NSCLC- Tumour suppressor | SM Noor et al;2018 [119] |
| miR-630 | Mut-Bcl-2-3ʹ-UTR | Unchanged | NSCLC- Tumour suppressor | Huei Lee et al;2018 [120] |
| miR-31 | LATS2/PPP2R2A | Overexpressed | NSCLC -Oncogenic | Liu et al;2010 [121] |
| miR-221/222 | PUMA | Overexpressed | NSCLC -Oncogenic | Zhang et al;2014 [122] |
| miR-197 | PD-L1 | Overexpressed | NSCLC -Oncogenic | Fujita et al; 2015 [123] |
| microRNA-146a | EGFR | Overexpressed | NSCLC- Tumour suppressor | Chen et al; 2013 [124] |
10. Targeting the Tumor Microenvironment for Cancer Therapy
The malignant features of cancer cells cannot be proved without a crucial interplay between cancer cells and their local environment. Angiogenic vascular cells, lymphatic endothelial cells, immune cells, and cancer-associated fibroblastic cells are the composition of the tumor infiltrate, which actively leads to cancer progression. The efficiency to alter these surroundings is a crucial feature by which tumor cells are capable of gaining some of the hallmark functions necessary for tumor development and metastatic dissemination. Targeting the tumor microenvironment to encapsulate or destroy cancer cells in their local environment has become compulsory. The difference in stromal cells, the complexity of the molecular elements of the tumor stroma, and the resemblance with normal tissue present huge challenges for therapies targeting the tumor microenvironment. Most recent investigations have shed light on the significant role in cancer progression played by the noncellular stromal compartment composed of the extracellular matrix [125].
This TME will influence the therapeutic effect/response and has an impact on the explicit surface receptors and activated or silenced signaling pathways. To deal with troubles such as no response to therapy or tumor resistance, and aiming to attain a personalized medicine in oncology, each tumor must be considered as a complex disease, different in each patient, and thus demanding a different strategy concerning therapeutics, especially centered on combinations. Hence, to formulate new therapeutic strategies towards a more effective targeting of TME, a big attempt has been made, which centers on (i) therapeutic strategies that target TME components and (ii) the development of models that exactly correspond to the TME for bench investigations [126]. It is now progressively accepted that cancer cells interact intimately with the extracellular matrix (ECM) and stromal cells, which together form the major construct of TME, instead of working alone [127].
Author Contributions
Conceptualization- I.M. and M.M.H; writing—original draft preparation- S.M.S, S.N.T and A.I.I; Review and editing- S.M.S, S.N.T, A.I.I, M.M, F.A, M.A.M, I.M, and M.M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| NSCLC | Non-small cell lung cancer |
| SCLC | Small cell lung cancer |
| TME | Tumor-microenvironment |
| ICB | Immune checkpoint blockade |
| MSCs | Mesenchymal stromal/stem cells |
| TAFs | Tumor-associated fibroblasts |
| Tregs | Regulatory T-cells |
| MDSCs | Myeloid-derived suppressor cells |
| NK | Natural killer cells |
| DCs | Dendritic cells |
| ECM | Extracellular matrix |
| HIFs | Hypoxia-inducible factors |
| MMPs | Matrix metalloproteinases |
References
- Sica, A.; Bronte, V. Altered macrophage differentiation and immune dysfunction in tumor development. J. Clin. Investig. 2007, 117, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Mimi, M.A.; Hasan, M.; Takanashi, Y.; Waliullah, A.; Mamun, A.; Chi, Z.; Kahyo, T.; Aramaki, S.; Takatsuka, D.; Koizumi, K.; et al. UBL3 overexpression enhances EV-mediated Achilles protein secretion in conditioned media of MDA-MB-231 cells. Biochem. Biophys. Res. Commun. 2024, 738, 150559. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S. Tumor Microenvironment and Myelomonocytic Cells. BoD–Books on Demand. 2012. [Google Scholar]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Goldstraw P, Ball D, Jett JR, et al. Non-small-cell lung cancer. Lancet 2011, 378, 1727–1740. [CrossRef] [PubMed]
- Pirlog, R.; Chiroi, P.; Rusu, I.; Jurj, A.M.; Budisan, L.; Pop-Bica, C.; Braicu, C.; Crisan, D.; Sabourin, J.-C.; Berindan-Neagoe, I. Cellular and Molecular Profiling of Tumor Microenvironment and Early-Stage Lung Cancer. Int. J. Mol. Sci. 2022, 23, 5346. [Google Scholar] [CrossRef]
- Mok, T.S.K.; Wu, Y.-L.; Kudaba, I.; Kowalski, D.M.; Cho, B.C.; Turna, H.Z.; Castro, G., Jr.; Srimuninnimit, V.; Laktionov, K.K.; Bondarenko, I.; et al. Pembrolizumab versus chemotherapy for previously untreated, PD-L1-expressing, locally advanced or metastatic non-small-cell lung cancer (KEYNOTE-042): a randomised, open-label, controlled, phase 3 trial. Lancet 2019, 393, 1819–1830. [Google Scholar] [CrossRef]
- Jung, H.; Paust, S. Chemokines in the tumor microenvironment: implications for lung cancer and immunotherapy. Front. Immunol. 2024, 15, 1443366. [Google Scholar] [CrossRef]
- A Arenberg, D.; Keane, M.P.; DiGiovine, B.; Kunkel, S.L.; Morris, S.B.; Xue, Y.Y.; Burdick, M.D.; Glass, M.C.; Iannettoni, M.D.; Strieter, R.M. Epithelial-neutrophil activating peptide (ENA-78) is an important angiogenic factor in non-small cell lung cancer. J. Clin. Investig. 1998, 102, 465–472. [Google Scholar] [CrossRef]
- Huang, M.; Wang, J.; Lee, P.; Sharma, S.; Mao, J.T.; Meissner, H.; Uyemura, K.; Modlin, R.; Wollman, J.; Dubinett, S.M. Human non-small cell lung cancer cells express a type 2 cytokine pattern. . 1995, 55, 3847–53. [Google Scholar]
- Jover, R.; Nguyen, T.; Pérez–Carbonell, L.; Zapater, P.; Payá, A.; Alenda, C.; Rojas, E.; Cubiella, J.; Balaguer, F.; Morillas, J.D.; et al. 5-Fluorouracil Adjuvant Chemotherapy Does Not Increase Survival in Patients With CpG Island Methylator Phenotype Colorectal Cancer. Gastroenterology 2011, 140, 1174–1181. [Google Scholar] [CrossRef]
- Srinivasarao, D.A.; Shah, S.; Famta, P.; Vambhurkar, G.; Jain, N.; Pindiprolu, S.K.S.; Sharma, A.; Kumar, R.; Padhy, H.P.; Kumari, M.; et al. Unravelling the role of tumor microenvironment responsive nanobiomaterials in spatiotemporal controlled drug delivery for lung cancer therapy. Drug Deliv. Transl. Res. 2024, 15, 407–435. [Google Scholar] [CrossRef]
- Sohag, S.M.; Toma, S.N.; Morshed, N.; Imon, A.I.; Islam, M.; Piash, I.; Shahria, N.; Mahmud, I. Exploration of analgesic and anthelmintic activities of Artocarpus chaplasha ROXB. leaves supported by in silico molecular docking. Phytomedicine Plus 2025, 5. [Google Scholar] [CrossRef]
- Huang S, Chung JYF, Li C, et al. Cellular dynamics of tumor microenvironment driving immunotherapy resistance in non-small-cell lung carcinoma. Cancer Lett. Published online 2024, 217272. [Google Scholar]
- Riera-Domingo C, Audigé A, Granja S, et al. Immunity, hypoxia, and metabolism–the Ménage à Trois of cancer: implications for immunotherapy. Physiol Rev. 2020, 100, 1–102. [Google Scholar] [CrossRef] [PubMed]
- Saeed Issa B, Adhab AH, Salih Mahdi M, et al. Decoding the complex web: cellular and molecular interactions in the lung tumour microenvironment. J Drug Target. 2025, 33, 666–690. [Google Scholar] [CrossRef] [PubMed]
- Shackleton, M.; Vaillant, F.; Simpson, K.J.; Stingl, J.; Smyth, G.K.; Asselin-Labat, M.-L.; Wu, L.; Lindeman, G.J.; Visvader, J.E. Generation of a functional mammary gland from a single stem cell. Nature 2006, 439, 84–88. [Google Scholar] [CrossRef]
- Liu, X.; Wu, S.; Yang, Y.; Zhao, M.; Zhu, G.; Hou, Z. The prognostic landscape of tumor-infiltrating immune cell and immunomodulators in lung cancer. Biomed. Pharmacother. 2017, 95, 55–61. [Google Scholar] [CrossRef]
- van der Bruggen P, Van Pel A. Tumor antigens recognized by T lymphocytes. Annu Rev, Immunol. 1994, 12, 337–365. [Google Scholar] [CrossRef]
- Dunn, G.P.; Old, L.J.; Schreiber, R.D. The Immunobiology of Cancer Immunosurveillance and Immunoediting. Immunity 2004, 21, 137–148. [Google Scholar] [CrossRef]
- Cheng, C.; Nguyen, T.T.; Tang, M.; Wang, X.; Jiang, C.; Liu, Y.; Gorlov, I.; Gorlova, O.; Iafrate, J.; Lanuti, M.; et al. Immune Infiltration in Tumor and Adjacent Non-Neoplastic Regions Codetermines Patient Clinical Outcomes in Early-Stage Lung Cancer. J. Thorac. Oncol. 2023, 18, 1184–1198. [Google Scholar] [CrossRef]
- A Barnes, T.; Amir, E. HYPE or HOPE: the prognostic value of infiltrating immune cells in cancer. Br. J. Cancer 2017, 117, 451–460. [Google Scholar] [CrossRef]
- Tong, Z.; Wang, X.; Liu, H.; Ding, J.; Chu, Y.; Zhou, X. The relationship between tumor infiltrating immune cells and the prognosis of patients with lung adenocarcinoma. J. Thorac. Dis. 2023, 15, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Steffens, S.; Kayser, C.; Roesner, A.; Rawluk, J.; Schmid, S.; Gkika, E.; Kayser, G. Low densities of immune cells indicate unfavourable overall survival in patients suffering from squamous cell carcinoma of the lung. Sci. Rep. 2024, 14, 1–11. [Google Scholar] [CrossRef]
- Ku, B.M.; Kim, Y.; Lee, K.Y.; Kim, S.; Sun, J.; Lee, S.; Ahn, J.S.; Park, K.; Ahn, M. Tumor infiltrated immune cell types support distinct immune checkpoint inhibitor outcomes in patients with advanced non-small cell lung cancer. Eur. J. Immunol. 2021, 51, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yang, M.; Luo, W.; Zhou, Q. Characteristics of tumor microenvironment and novel immunotherapeutic strategies for non-small cell lung cancer. J. Natl. Cancer Cent. 2022, 2, 243–262. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Shao, Y.; He, W.; Hu, W.; Xu, Y.; Chen, J.; Wu, C.; Jiang, J. Prognostic Role of Tumor-Infiltrating Lymphocytes in Lung Cancer: a Meta-Analysis. Cell. Physiol. Biochem. 2015, 37, 1560–1571. [Google Scholar] [CrossRef]
- Vinay, D.S.; Ryan, E.P.; Pawelec, G.; Talib, W.H.; Stagg, J.; Elkord, E.; Lichtor, T.; Decker, W.K.; Whelan, R.L.; Kumara, H.M.C.S.; et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin. Cancer Biol. 2015, 35, S185–S198. [Google Scholar] [CrossRef]
- Ye, B.; Stary, C.M.; Gao, Q.; Wang, Q.; Zeng, Z.; Jian, Z.; Gu, L.; Xiong, X. Genetically Modified T-Cell-Based Adoptive Immunotherapy in Hematological Malignancies. J. Immunol. Res. 2017, 2017, 1–13. [Google Scholar] [CrossRef]
- Keenan, T.E.; Burke, K.P.; Van Allen, E.M. Genomic correlates of response to immune checkpoint blockade. Nat. Med. 2019, 25, 389–402. [Google Scholar] [CrossRef]
- Walunas, T.L.; Lenschow, D.J.; Bakker, C.Y.; Linsley, P.S.; Freeman, G.J.; Green, J.M.; Thompson, C.B.; Bluestone, J.A. CTLA-4 can function as a negative regulator of T cell activation. Immunity 1994, 1, 405–413. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, Y.; Wang, H.; Xu, Z.; Wang, Y.; Li, S.; Liu, J.; Chen, Y.; Luo, H.; Wu, L.; et al. Massive PD-L1 and CD8 double positive TILs characterize an immunosuppressive microenvironment with high mutational burden in lung cancer. J. Immunother. Cancer 2021, 9, e002356. [Google Scholar] [CrossRef] [PubMed]
- Lee S, Margolin K. Cytokines in cancer immunotherapy. Cancers (Basel) 2011, 3, 3856–3893. [Google Scholar] [CrossRef] [PubMed]
- Rochman, Y.; Spolski, R.; Leonard, W.J. New insights into the regulation of T cells by γc family cytokines. Nat. Rev. Immunol. 2009, 9, 480–490. [Google Scholar] [CrossRef]
- Essogmo, F.E.; Zhilenkova, A.V.; Tchawe, Y.S.N.; Owoicho, A.M.; Rusanov, A.S.; Boroda, A.; Pirogova, Y.N.; Sangadzhieva, Z.D.; Sanikovich, V.D.; Bagmet, N.N.; et al. Cytokine Profile in Lung Cancer Patients: Anti-Tumor and Oncogenic Cytokines. Cancers 2023, 15, 5383. [Google Scholar] [CrossRef] [PubMed]
- Kotenko, S.V.; Pestka, S. Jak-Stat signal transduction pathway through the eyes of cytokine class II receptor complexes. Oncogene 2000, 19, 2557–2565. [Google Scholar] [CrossRef]
- Vilcek, J. Novel interferons. Nat. Immunol. 2003, 4, 8–9. [Google Scholar] [CrossRef]
- Steen, H.C.; Gamero, A.M. Interferon-Lambda as a Potential Therapeutic Agent in Cancer Treatment. J. Interf. Cytokine Res. 2010, 30, 597–602. [Google Scholar] [CrossRef]
- Langer, C.J.; Besse, B.; Gualberto, A.; Brambilla, E.; Soria, J.-C. The Evolving Role of Histology in the Management of Advanced Non–Small-Cell Lung Cancer. J. Clin. Oncol. 2010, 28, 5311–5320. [Google Scholar] [CrossRef]
- I Grivennikov, S.; Karin, M. Inflammation and oncogenesis: a vicious connection. Curr. Opin. Genet. Dev. 2010, 20, 65–71. [Google Scholar] [CrossRef]
- Valastyan, S.; Weinberg, R.A. Tumor Metastasis: Molecular Insights and Evolving Paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix Metalloproteinases: Regulators of the Tumor Microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Azzali, G. The modality of transendothelial passage of lymphocytes and tumor cells in the absorbing lymphatic vessel. . 2007, 73–7. [Google Scholar]
- Chunhacha, P.; Chanvorachote, P. Roles of caveolin-1 on anoikis resistance in non small cell lung cancer. . 2012, 4, 149–55. [Google Scholar] [PubMed]
- Al-Mehdi, A.B.; Tozawa, K.; Fisher, A.B.; Shientag, L.; Lee, A.; Muschel, R.J. Intravascular origin of metastasis from the proliferation of endothelium-attached tumor cells: A new model for metastasis. Nat. Med. 2000, 6, 100–102. [Google Scholar] [CrossRef]
- Chambers, A.F.; Groom, A.C.; MacDonald, I.C. Dissemination and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2002, 2, 563–572. [Google Scholar] [CrossRef]
- Gupta, G.P.; Nguyen, D.X.; Chiang, A.C.; Bos, P.D.; Kim, J.Y.; Nadal, C.; Gomis, R.R.; Manova-Todorova, K.; Massagué, J. Mediators of vascular remodelling co-opted for sequential steps in lung metastasis. Nature 2007, 446, 765–770. [Google Scholar] [CrossRef]
- Psaila, B.; Lyden, D. The metastatic niche: adapting the foreign soil. Nat. Rev. Cancer 2009, 9, 285–293. [Google Scholar] [CrossRef]
- Erler, J.T.; Bennewith, K.L.; Cox, T.R.; Lang, G.; Bird, D.; Koong, A.; Le, Q.-T.; Giaccia, A.J. Hypoxia-Induced Lysyl Oxidase Is a Critical Mediator of Bone Marrow Cell Recruitment to Form the Premetastatic Niche. Cancer Cell 2009, 15, 35–44. [Google Scholar] [CrossRef]
- Kang, Y.; Siegel, P.M.; Shu, W.; Drobnjak, M.; Kakonen, S.M.; Cordón-Cardo, C.; Guise, T.A.; Massagué, J. A multigenic program mediating breast cancer metastasis to bone. Cancer Cell 2003, 3, 537–549. [Google Scholar] [CrossRef]
- Nguyen, D.X.; Bos, P.D.; Massague, J. Metastasis: from dissemination to organ-specific colonization. Nat. Rev. Cancer 2009, 9, 274–284. [Google Scholar] [CrossRef]
- Bozkurt, A.S. Publication Status of Mouse Embryonic Fibroblast Cells in Scientific Journals. Eur. J. Ther. 2021, 27, 135–141. [Google Scholar] [CrossRef]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Parsonage, G.; Filer, A.D.; Haworth, O.; Nash, G.B.; Rainger, G.E.; Salmon, M.; Buckley, C.D. A stromal address code defined by fibroblasts. Trends Immunol. 2005, 26, 150–156. [Google Scholar] [CrossRef]
- Rodemann, H.P.; Müller, G.A. Characterization of Human Renal Fibroblasts in Health and Disease: II. In Vitro Growth, Differentiation, and Collagen Synthesis of Fibroblasts From Kidneys With Interstitial Fibrosis. Am. J. Kidney Dis. 1991, 17, 684–686. [Google Scholar] [CrossRef]
- Chang, H.Y.; Chi, J.-T.; Dudoit, S.; Bondre, C.; van de Rijn, M.; Botstein, D.; Brown, P.O. Diversity, topographic differentiation, and positional memory in human fibroblasts. Proc. Natl. Acad. Sci. 2002, 99, 12877–12882. [Google Scholar] [CrossRef]
- Desmouliere, A.; Guyot, C.; Gabbiani, G. The stroma reaction myofibroblast: a key player in the control of tumor cell behavior. Int. J. Dev. Biol. 2004, 48, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Erez, N.; Truitt, M.; Olson, P.; Hanahan, D. Cancer-Associated Fibroblasts Are Activated in Incipient Neoplasia to Orchestrate Tumor-Promoting Inflammation in an NF-κB-Dependent Manner. Cancer Cell 2010, 17, 135–147. [Google Scholar] [CrossRef]
- Schliekelman, M.J.; Creighton, C.J.; Baird, B.N.; Chen, Y.; Banerjee, P.; Bota-Rabassedas, N.; Ahn, Y.-H.; Roybal, J.D.; Chen, F.; Zhang, Y.; et al. Thy-1+ Cancer-associated Fibroblasts Adversely Impact Lung Cancer Prognosis. Sci. Rep. 2017, 7, 6478–6478. [Google Scholar] [CrossRef]
- Shintani, Y.; Kimura, T.; Funaki, S.; Ose, N.; Kanou, T.; Fukui, E. Therapeutic Targeting of Cancer-Associated Fibroblasts in the Non-Small Cell Lung Cancer Tumor Microenvironment. Cancers 2023, 15, 335. [Google Scholar] [CrossRef]
- Zhao Y, Shen M, Wu L, et al. Stromal cells in the tumor microenvironment: accomplices of tumor progression? Cell Death Dis. 2023, 14, 587. [Google Scholar] [CrossRef]
- Jiang, X.; Hu, S.; Liu, Q.; Qian, C.; Liu, Z.; Luo, D. Exosomal microRNA remodels the tumor microenvironment. PeerJ 2017, 5, e4196. [Google Scholar] [CrossRef] [PubMed]
- Seager, R.J.; Hajal, C.; Spill, F.; Kamm, R.D.; Zaman, M.H. Dynamic interplay between tumour, stroma and immune system can drive or prevent tumour progression. Converg. Sci. Phys. Oncol. 2017, 3, 034002. [Google Scholar] [CrossRef]
- Green, J.A.; Arpaia, N.; Schizas, M.; Dobrin, A.; Rudensky, A.Y. A nonimmune function of T cells in promoting lung tumor progression. J. Exp. Med. 2017, 214, 3565–3575. [Google Scholar] [CrossRef]
- Xiao, J.; Huang, K.; Lin, H.; Xia, Z.; Zhang, J.; Li, D.; Jin, J. Mogroside IIE Inhibits Digestive Enzymes via Suppression of Interleukin 9/Interleukin 9 Receptor Signalling in Acute Pancreatitis. Front. Pharmacol. 2020, 11, 859. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.; Lin, Y.; Navin, N. Advancing Cancer Research and Medicine with Single-Cell Genomics. Cancer Cell 2020, 37, 456–470. [Google Scholar] [CrossRef]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Ostuni, R.; Kratochvill, F.; Murray, P.J.; Natoli, G. Macrophages and cancer: from mechanisms to therapeutic implications. Trends Immunol. 2015, 36, 229–239. [Google Scholar] [CrossRef]
- Mazur, A.; Holthoff, E.; Vadali, S.; Kelly, T.; Post, S.R. Cleavage of Type I Collagen by Fibroblast Activation Protein-α Enhances Class A Scavenger Receptor Mediated Macrophage Adhesion. PLOS ONE 2016, 11, e0150287. [Google Scholar] [CrossRef]
- Lewis, C.E.; Pollard, J.W. Distinct Role of Macrophages in Different Tumor Microenvironments. Cancer Res. 2006, 66, 605–612. [Google Scholar] [CrossRef]
- Bingle, L.; Brown, N.; Lewis, C.E. The role of tumour-associated macrophages in tumour progression: implications for new anticancer therapies. J. Pathol. 2002, 196, 254–265. [Google Scholar] [CrossRef]
- Conway EM, Pikor LA, Kung SHY, et al. Macrophages, inflammation, and lung cancer. Am J Respir Crit Care Med. 2016, 193, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Edin S, Wikberg ML, Oldenborg PA, Palmqvist R. Macrophages: Good guys in colorectal cancer. Oncoimmunology 2013, 2, e23038. [Google Scholar] [CrossRef] [PubMed]
- Trivanović, D.; Krstić, J.; Djordjević, I.O.; Mojsilović, S.; Santibanez, J.F.; Bugarski, D.; Jauković, A. The Roles of Mesenchymal Stromal/Stem Cells in Tumor Microenvironment Associated with Inflammation. Mediat. Inflamm. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M. Mast cells in allergy and infection: Versatile effector and regulatory cells in innate and adaptive immunity. Eur. J. Immunol. 2010, 40, 1843–1851. [Google Scholar] [CrossRef]
- Boldrini, L.; Gisfredi, S.; Ursino, S.; Lucchi, M.; Melfi, F.; Mussi, A.; Basolo, F.; Fontanini, G. Tumour necrosis factor-α: prognostic role and relationship with interleukin-8 and endothelin-1 in non-small cell lung cancer. Int. J. Mol. Med. 2006, 17, 887–892. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Redegeld, F.A. Role of Mast Cells in Shaping the Tumor Microenvironment. Clin. Rev. Allergy Immunol. 2020, 58, 313–325. [Google Scholar] [CrossRef]
- Cimpean, A.M.; Tamma, R.; Ruggieri, S.; Nico, B.; Toma, A.; Ribatti, D. Mast cells in breast cancer angiogenesis. Crit. Rev. Oncol. 2017, 115, 23–26. [Google Scholar] [CrossRef]
- Jang, G.-Y.; Lee, J.W.; Kim, Y.S.; Lee, S.E.; Han, H.D.; Hong, K.-J.; Kang, T.H.; Park, Y.-M. Interactions between tumor-derived proteins and Toll-like receptors. Exp. Mol. Med. 2020, 52, 1926–1935. [Google Scholar] [CrossRef]
- Dudziak, D.; Kamphorst, A.O.; Heidkamp, G.F.; Buchholz, V.R.; Trumpfheller, C.; Yamazaki, S.; Cheong, C.; Liu, K.; Lee, H.-W.; Park, C.G.; et al. Differential Antigen Processing by Dendritic Cell Subsets in Vivo. Science 2007, 315, 107–111. [Google Scholar] [CrossRef]
- Roberts EW, Broz ML, Binnewies M, et al. Critical role for CD103+/CD141+ dendritic cells bearing CCR7 for tumor antigen trafficking and priming of T cell immunity in melanoma. Cancer Cell. 2016, 30, 324–336. [Google Scholar] [CrossRef]
- Böttcher, J.P.; Bonavita, E.; Chakravarty, P.; Blees, H.; Cabeza-Cabrerizo, M.; Sammicheli, S.; Rogers, N.C.; Sahai, E.; Zelenay, S.; e Sousa, C.R. NK Cells Stimulate Recruitment of cDC1 into the Tumor Microenvironment Promoting Cancer Immune Control. Cell 2018, 172, 1022–1037.e14. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.C.; Gudjonson, H.; Pritykin, Y.; Deep, D.; Lavallée, V.-P.; Mendoza, A.; Fromme, R.; Mazutis, L.; Ariyan, C.; Leslie, C.; et al. Transcriptional Basis of Mouse and Human Dendritic Cell Heterogeneity. Cell 2019, 179, 846–863.e24. [Google Scholar] [CrossRef] [PubMed]
- Canton, J.; Blees, H.; Henry, C.M.; Buck, M.D.; Schulz, O.; Rogers, N.C.; Childs, E.; Zelenay, S.; Rhys, H.; Domart, M.-C.; et al. The receptor DNGR-1 signals for phagosomal rupture to promote cross-presentation of dead-cell-associated antigens. Nat. Immunol. 2020, 22, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Giampazolias, E.; Schulz, O.; Lim, K.H.J.; Rogers, N.C.; Chakravarty, P.; Srinivasan, N.; Gordon, O.; Cardoso, A.; Buck, M.D.; Poirier, E.Z.; et al. Secreted gelsolin inhibits DNGR-1-dependent cross-presentation and cancer immunity. Cell 2021, 184, 4016–4031.e22. [Google Scholar] [CrossRef]
- Ahluwalia, P.; Ahluwalia, M.; Mondal, A.K.; Sahajpal, N.S.; Kota, V.; Rojiani, M.V.; Kolhe, R. Natural Killer Cells and Dendritic Cells: Expanding Clinical Relevance in the Non-Small Cell Lung Cancer (NSCLC) Tumor Microenvironment. Cancers 2021, 13, 4037. [Google Scholar] [CrossRef]
- Carmeliet, P. Angiogenesis in life, disease and medicine. Nature 2005, 438, 932–936. [Google Scholar] [CrossRef]
- Dvorak, H.F. Angiogenesis: update 2005. J. Thromb. Haemost. 2005, 3, 1835–1842. [Google Scholar] [CrossRef]
- Ribatti, D.; Ennas, M.G.; Vacca, A.; Ferreli, F.; Nico, B.; Orru, S.; Sirigu, P. Tumor vascularity and tryptase-positive mast cells correlate with a poor prognosis in melanoma. Eur. J. Clin. Investig. 2003, 33, 420–425. [Google Scholar] [CrossRef]
- Hogan, K.A.; Cho, D.S.; Arneson, P.C.; Samani, A.; Palines, P.; Yang, Y.; Doles, J.D. Tumor-derived cytokines impair myogenesis and alter the skeletal muscle immune microenvironment. Cytokine 2018, 107, 9–17. [Google Scholar] [CrossRef]
- Poeta, V.M.; Massara, M.; Capucetti, A.; Bonecchi, R. Chemokines and Chemokine Receptors: New Targets for Cancer Immunotherapy. Front. Immunol. 2019, 10, 379. [Google Scholar] [CrossRef]
- Sarode, P.; Schaefer, M.B.; Grimminger, F.; Seeger, W.; Savai, R. Macrophage and Tumor Cell Cross-Talk Is Fundamental for Lung Tumor Progression: We Need to Talk. Front. Oncol. 2020, 10, 324. [Google Scholar] [CrossRef]
- Huang, M.; Stolina, M.; Sharma, S.; Mao, J.T.; Zhu, L.; Miller, P.W.; Wollman, J.; Herschman, H.; Dubinett, S.M. Non-small cell lung cancer cyclooxygenase-2-dependent regulation of cytokine balance in lymphocytes and macrophages: up-regulation of interleukin 10 and down-regulation of interleukin 12 production. . 1998, 58, 1208–16. [Google Scholar] [PubMed]
- Ramachandran, S.; Verma, A.K.; Dev, K.; Goyal, Y.; Bhatt, D.; Alsahli, M.A.; Rahmani, A.H.; Almatroudi, A.; Almatroodi, S.A.; Alrumaihi, F.; et al. Role of Cytokines and Chemokines in NSCLC Immune Navigation and Proliferation. Oxidative Med. Cell. Longev. 2021, 2021, 5563746. [Google Scholar] [CrossRef] [PubMed]
- Hendler, F.J.; Ozanne, B.W. Human squamous cell lung cancers express increased epidermal growth factor receptors. J. Clin. Investig. 1984, 74, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Normanno, N.; De Luca, A.; Bianco, C.; Strizzi, L.; Mancino, M.; Maiello, M.R.; Carotenuto, A.; De Feo, G.; Caponigro, F.; Salomon, D.S. Epidermal growth factor receptor (EGFR) signaling in cancer. Gene 2006, 366, 2–16. [Google Scholar] [CrossRef]
- Kris MG, Natale RB, Herbst RS, et al. Efficacy of gefitinib, an inhibitor of the epidermal growth factor receptor tyrosine kinase, in symptomatic patients with non–small cell lung cancer: a randomized trial. Jama 2003, 290, 2149–2158. [Google Scholar] [CrossRef]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef]
- Gross, J.; Lapiere, C.M. COLLAGENOLYTIC ACTIVITY IN AMPHIBIAN TISSUES: A TISSUE CULTURE ASSAY. Proc. Natl. Acad. Sci. 1962, 48, 1014–1022. [Google Scholar] [CrossRef]
- Liu, L.; Xu, S.; Huang, L.; He, J.; Liu, G.; Ma, S.; Weng, Y.; Huang, S. Systemic immune microenvironment and regulatory network analysis in patients with lung adenocarcinoma. Transl. Cancer Res. 2021, 10, 2859–2872. [Google Scholar] [CrossRef]
- Serafini, B.; Rosicarelli, B.; Magliozzi, R.; Stigliano, E.; Aloisi, F. Detection of Ectopic B-cell Follicles with Germinal Centers in the Meninges of Patients with Secondary Progressive Multiple Sclerosis. Brain Pathol. 2004, 14, 164–174. [Google Scholar] [CrossRef]
- A Rosenberg, S.; Yang, J.C.; Restifo, N.P. Cancer immunotherapy: moving beyond current vaccines. Nat. Med. 2004, 10, 909–915. [Google Scholar] [CrossRef]
- Brown, J.M.; Wilson, W.R. Exploiting tumour hypoxia in cancer treatment. Nat. Rev. Cancer 2004, 4, 437–447. [Google Scholar] [CrossRef]
- Li Y, Zhao L, Li XF. Hypoxia and the tumor microenvironment. Technol Cancer Res Treat. 2021, 20, 15330338211036304. [Google Scholar] [CrossRef]
- Bedogni, B.; Powell, M.B. Hypoxia, melanocytes and melanoma – survival and tumor development in the permissive microenvironment of the skin. Pigment. Cell Melanoma Res. 2009, 22, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.A.; Sutphin, P.D.; Denko, N.C.; Giaccia, A.J. Role of Prolyl Hydroxylation in Oncogenically Stabilized Hypoxia-inducible Factor-1α. J. Biol. Chem. 2002, 277, 40112–40117. [Google Scholar] [CrossRef] [PubMed]
- Masson, N.; Willam, C.; Maxwell, P.H.; Pugh, C.W.; Ratcliffe, P.J. Independent function of two destruction domains in hypoxia-inducible factor-α chains activated by prolyl hydroxylation. EMBO J. 2001, 20, 5197–5206. [Google Scholar] [CrossRef] [PubMed]
- Sahu, A.; Kwon, I.; Tae, G. Improving cancer therapy through the nanomaterials-assisted alleviation of hypoxia. Biomaterials 2020, 228, 119578. [Google Scholar] [CrossRef]
- Otmani, K.; Lewalle, P. Tumor Suppressor miRNA in Cancer Cells and the Tumor Microenvironment: Mechanism of Deregulation and Clinical Implications. Front. Oncol. 2021, 11. [Google Scholar] [CrossRef]
- Palmero, E.I.; de Campos, S.G.P.; Campos, M.; de Souza, N.C.N.; Guerreiro, I.D.C.; Carvalho, A.L.; Marques, M.M.C. Mechanisms and role of microRNA deregulation in cancer onset and progression. Genet. Mol. Biol. 2011, 34, 363–370. [Google Scholar] [CrossRef]
- Lee YS, Dutta A. MicroRNAs in cancer. Annu Rev Pathol Mech Dis. 2009, 4, 199–227. [Google Scholar] [CrossRef]
- Garzon R, Calin GA, Croce CM. MicroRNAs in cancer. Annu Rev Med. 2009, 60, 167–179. [Google Scholar] [CrossRef]
- Rusek, A.M.; Abba, M.; Eljaszewicz, A.; Moniuszko, M.; Niklinski, J.; Allgayer, H. MicroRNA modulators of epigenetic regulation, the tumor microenvironment and the immune system in lung cancer. Mol. Cancer 2015, 14, 34–34. [Google Scholar] [CrossRef]
- Xi, S.; Xu, H.; Shan, J.; Tao, Y.; Hong, J.A.; Inchauste, S.; Zhang, M.; Kunst, T.F.; Mercedes, L.; Schrump, D.S. Cigarette smoke mediates epigenetic repression of miR-487b during pulmonary carcinogenesis. J. Clin. Investig. 2013, 123, 1241–1261. [Google Scholar] [CrossRef]
- Peng, Y.; Dai, Y.; Hitchcock, C.; Yang, X.; Kassis, E.S.; Liu, L.; Luo, Z.; Sun, H.-L.; Cui, R.; Wei, H.; et al. Insulin growth factor signaling is regulated by microRNA-486, an underexpressed microRNA in lung cancer. Proc. Natl. Acad. Sci. 2013, 110, 15043–15048. [Google Scholar] [CrossRef]
- Lujambio, A.; Ropero, S.; Ballestar, E.; Fraga, M.F.; Cerrato, C.; Setién, F.; Casado, S.; Suarez-Gauthier, A.; Sanchez-Cespedes, M.; Gitt, A.; et al. Genetic Unmasking of an Epigenetically Silenced microRNA in Human Cancer Cells. Cancer Res. 2007, 67, 1424–1429. [Google Scholar] [CrossRef]
- Wu, C.; Ho, J.; Hung, S.; Yu, D. miR-429 expression in bladder cancer and its correlation with tumor behavior and clinical outcome. Kaohsiung J. Med Sci. 2018, 34, 335–340. [Google Scholar] [CrossRef]
- Hubaux R, Becker-Santos DD, Enfield KSS, Lam S, Lam WL, Martinez VD. MicroRNAs As Biomarkers For Clinical Features Of Lung Cancer. Metabolomics open access. 2012, 2, 1000108. [Google Scholar] [CrossRef]
- San Ho C, Noor SM, Nagoor NH. MiR-378 and MiR-1827 regulate tumor invasion, migration and angiogenesis in human lung adenocarcinoma by targeting RBX1 and CRKL, respectively. J Cancer 2018, 9, 331. [Google Scholar] [CrossRef]
- Chen, M.-J.; Wu, D.-W.; Wang, G.-C.; Wang, Y.-C.; Chen, C.-Y.; Lee, H. MicroRNA-630 may confer favorable cisplatin-based chemotherapy and clinical outcomes in non-small cell lung cancer by targeting Bcl-2. Oncotarget 2018, 9, 13758–13767. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sempere, L.F.; Ouyang, H.; Memoli, V.A.; Andrew, A.S.; Luo, Y.; Demidenko, E.; Korc, M.; Shi, W.; Preis, M.; et al. MicroRNA-31 functions as an oncogenic microRNA in mouse and human lung cancer cells by repressing specific tumor suppressors. J. Clin. Investig. 2010, 120, 1298–1309. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Zhang, C.; Zhang, J.; Zhang, A.; Wang, Y.; Han, L.; You, Y.; Pu, P. PUMA is a novel target of miR-221/222 in human epithelial cancers. Int. J. Oncol. 2010, 37, 1621–1626. [Google Scholar] [CrossRef]
- Fujita, Y.; Yagishita, S.; Hagiwara, K.; Yoshioka, Y.; Kosaka, N.; Takeshita, F.; Fujiwara, T.; Tsuta, K.; Nokihara, H.; Tamura, T.; et al. The Clinical Relevance of the miR-197/CKS1B/STAT3-mediated PD-L1 Network in Chemoresistant Non-small-cell Lung Cancer. Mol. Ther. 2015, 23, 717–727. [Google Scholar] [CrossRef]
- Chen, G.; Umelo, I.A.; Lv, S.; Teugels, E.; Fostier, K.; Kronenberger, P.; Dewaele, A.; Sadones, J.; Geers, C.; De Grève, J. miR-146a Inhibits Cell Growth, Cell Migration and Induces Apoptosis in Non-Small Cell Lung Cancer Cells. PLOS ONE 2013, 8, e60317. [Google Scholar] [CrossRef]
- Sounni NE, Noel A. Targeting the tumor microenvironment for cancer therapy. Clin Chem. 2013, 59, 85–93. [Google Scholar] [CrossRef]
- Roma-Rodrigues C, Mendes R, Baptista P V, Fernandes AR. Targeting tumor microenvironment for cancer therapy. Int J Mol Sci. 2019, 20, 840. [Google Scholar] [CrossRef]
- Pitt, J.M.; Marabelle, A.; Eggermont, A.; Soria, J.-C.; Kroemer, G.; Zitvogel, L. Targeting the tumor microenvironment: removing obstruction to anticancer immune responses and immunotherapy. Ann. Oncol. 2016, 27, 1482–1492. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



