
Submitted:
25 April 2025
Posted:
28 April 2025
You are already at the latest version
Abstract
Background/Objectives: Liver disease is one of the most common medical problems in the world. Pharmacocorrection of these pathologies includes the use of drugs with anti-oxidant and hepatoprotective action, among which there are natural and synthetic sul-fur-containing compounds. However, many of the drugs have side effects and their ap-plication does not always correspond to the approaches of evidence-based medicine. Therefore, today the urgent problem is the search for new effective substances with high metabolitotropic properties and high safety criteria. The aim of the work was in-depth study of the hepatoprotective and antioxidant action of the new investigational "lead-compound" among pteridine S-substituted (DCTP) under conditions of experi-mental tetrachloromethane hepatitis in rats in comparison with the reference drug "Thio-triazoline". Methods: The hepatoprotective effect of the compound was studied using the model of acute tetrachloromethane (CCl4) hepatitis in adult male Wistar rats. The levels of biochemical liver damage markers were estimated with spectrophotometric methods. Histological and immunohistochemical methods were used for determination of hepato-cytes damage. Statistical processing of data was performed using the nonparametric Wil-cox-on-Mann-Whitney method.
Results: The results of the studies showed that DCTP was superior to the reference drug Thiotriazoline in its effect on the level of AST, DC, Schiff bases and carbonylated proteins, markers of oxidative (Nrf2) and inflammatory (Lipocalin-2) stress, as well as on animal survival. The results were confirmed by histological examination data, which showed re-generation of the hepatocyte membrane structure, reduction of the infiltrative, destructive and inflammatory process in the liver, reduction of the cytolytic process, stabilization and increase in the functional activity of the liver due to the administration of the study drug. The pharmacological effects of lead-compound (DCTP) are probably associated with its structural similarity to tetrahydrofolic acid, which is an integral component of oxida-tion-reduction processes and a participant in the biosynthesis of nitrogenous bases of nu-cleotides, amino acids. The obtained data indicate antioxidant and hepatoprotective properties of the studied “lead-compound” from the pteridinethione group (DCTP). Con-clusions: It was shown that the studied substance DCTP significantly reduces acute hepa-totoxic effects caused by CCl4, as evidenced by a decrease in the level of lipid peroxi-da-tion and prooxidant markers, normalization of liver biochemical markers, as well as regeneration of liver architecture, limitation of inflammatory effects, a decrease in Nrf2 and Lipocalin-2 markers, and induction of liver antioxidant enzymes.
Keywords:
acute hepatitis
; S-substituted pteridine
; Lipocalin-2
; Nrf2
; hepatoprotectors
; antioxidants
; oxidative stress
1. Introduction
Liver pathologies of various etiologies are increasing in all countries [1]. This can be explained by the fact that liver diseases are induced by an extremely wide range of factors, from viruses to chemicals, drugs [2] and even food and beverages [3]. The problem is that liver diseases go through successive stages in their development: hepatitis, fibrosis, cirrhosis and/or hepatocellular cancer [4] and this process can take several years. At the initial stages, it is almost painless. The treatment is very ineffective when its detection occurs at later stages, when the pathology becomes chronic. In this regard, the search for ways to prevent and treat liver pathologies and the development of methods for testing new substances with hepatotropic action remain relevant.
As is known, one of the primary responses to liver dysfunction is a shift in the balance in the redox system towards an increase in prooxidants and the development of oxidative stress [5]. It is important to consider that changes in the redox system parameters are accompanied not only, and perhaps not so much, by the formation of negative hepatoxic compounds, but by the restructuring of regulatory systems in the liver and in the body as a whole [6]. Based on this, we believe that the assessment of the state of the redox system can be a marker in primary tests and assessment of the action of new hepatopropionic substances.
When selecting hepatoprotective substances, we proceeded from the fact that they should ensure the correction of the redox system. It is known that α-lipoic acid, methionine, S-adenosylmethionine, L-glutathione can stabilize and inhibit free radical oxidation in hepatocytes [7]. Thus, it has been shown that methionine helps maintain the oxidation-reduction status of cells by forming homocysteine, which is a substrate in the transsulfuration of glutathione pathway [8]. It also acts as a source of sulfur in the process of synthesis of the critical signaling molecule hydrogen sulfide [9]. S-adenosylmethionine is a universal donor of a methyl group in the methionine cycle in the presence of folates (THF, 5-MTHF, 5,10-MTHF) [10]. We believe that a promising approach to the creation of new effective hepatoprotectors may be the modification of the natural heterocyclic matrix - pteridine. This is supported by data showing that the drug Vesatolimod (an aminopteridine derivative) is undergoing clinical trials for the treatment of acute and chronic forms of hepatitis B [11], and S-substituted pteridines are effective inhibitors of dehydrofolate reductase and free radical scavengers [12], exhibit antioxidant activity in in vitro experiments and in toxic liver damage [13]. In addition, pteridines such as tetrahydrofolic acid and tetrahydrobiopterin are coenzymes in the synthesis of nucleotides, amino acids, and phospholipids – molecules necessary for regeneration processes [14].
To test this working hypothesis, an in-depth study of the hepatoprotective and antioxidant effects of the new investigational “lead-compound” among S-substituted pteridine (DCTP) [15] was conducted under conditions of experimental tetrachloromethane hepatitis in rats in comparison with the reference drug “Thiotriazoline”.
2. Materials and Methods
The hepatoprotective effect of the compound was studied using the model of acute tetrachloromethane (CCl4) hepatitis in adult male Wistar rats (6-8 months) weighing 220-350 grams, which were kept under standard vivarium conditions (temperature 20 ± 5 ° C, humidity 65 ± 5%). The rats were kept on a standard diet with free access to water and food, under conditions of a natural alternation of day and night [16].
Animal care and experimental protocols were carried out in accordance with the requirements of the Council Directive 86/609/EEC of 24 November 1986 on the care and use of laboratory animals, the ethical principles for animal experiments approved by the First National Congress of Ukraine on Bioethics (2001), international agreements and Ukrainian legislation in this area, were approved by the Ethics Committee, and in accordance with Directive 2010/63/EU of the European Parliament [17].
To assess the hepatoprotective activity, rats were divided into the following groups of 10 animals each.
Group I: intact animals, which were intraperitoneally administered with an appropriate volume of 0.9% sodium chloride solution for 14 days.
In rats of groups II-IV, experimental hepatitis was induced by subcutaneous administration of carbon tetrachloride (CCl4) at a dose of 0.8 ml/100 g of weight in the form of a 50% oil solution once a day for 2 days [18,19].
Group II: control (pathology), animals were administered only CCl4.
Group III: rats in this group were given intraperitoneal administration of a 2.5% aqueous solution of Thiotriazoline (TTZ) at a rate of 10 mg/100 g (Arterium Ukraine, Series: UA/2931/01/02, No.: LSR-0052882 dated 18.02.2015) once a day against the background of CCl4 hepatitis from the 1st to the 14th day [20].
Group IV: rats in this group were given intraperitoneal administration of an aqueous solution of “lead-compound” (DCTP) [15] at a rate of 1/50 LD50, at a dose of 6 mg/100 g [19] once a day against the background of CCl4 hepatitis from the 1st to the 14th day.
Decapitation of animals using ether anesthesia was performed on the 15th day after the termination of the experiment from 9:00 to 11:00.
The liver was divided into three fragments, the first part was immediately removed and frozen in liquid nitrogen. For extraction of diene conjugates, frozen liver fragments were homogenized with a mixture of heptane:propan-2-ol (1:1).
The concentration of diene conjugates (DC) was determined on a ULAB 108UV spectrophotometer at 232 nm using the method of Recknagel and Goshal [21] and expressed as nmol/g liver tissue.
The concentration of Schiff bases in liver extracts was determined on a Hitachi MPF-4 spectrofluorimeter at an adsorption wavelength of 360 nm and an emission wavelength of 430 nm using the method of C.A. Rice-Evans [22] and expressed as nmol/g liver.
Determination of Superoxide dismutase (SOD) concentration (EC 1.15.1.1). Liver homogenate was obtained by grinding liver tissue (homogenization) in PBS buffer (pH = 7.4) using a Potter-Elvehjem homogenizer in a 10-fold volume of 0.25 M sucrose. Homogenates were filtered through 2 layers of gauze and centrifuged (CL-310b, Poland) at 1000×g for 10 min. Then the supernatant was centrifuged at 10,000×g for 20 min. Subsequently, the pellet was suspended in 5 ml of 0.25 M sucrose (pH 7.4) and washed twice at 10,000×g for 20 min. All procedures were performed at 4°C [23]. The sediment of purified mitochondria was suspended in 1 ml of 0.25 M sucrose.
The concentration of carbonylated proteins in the samples was determined using the method of R.L. Levine [24] as modified by E. Dubinina. The method is based on the interaction of carbonylated groups and imino groups of oxidized amino acid residues of proteins with 2,4-dinitrophenolhydrazone (2,4-DNPH) to form 2,4-dinitrophenolhydrazones, which have a specific absorption spectrum in the ultraviolet and visible regions of the spectrum. The level of spontaneous oxidative modification of proteins reflects the amount of carbonyl derivatives of proteins present in the samples (formed in vivo). For this purpose, 0.1 ml of the mitochondrial fraction suspension was mixed with an equal volume of 20% trichloroacetic acid (TCAA). The samples were vigorously shaken, after which 1 ml of 5% TCAA was added to them and they were centrifuged at 3000 rpm for 15 minutes. The formed supernatant was removed, and the sediment was mixed with 1 ml of 1 M 2,4-dinitrophenylhydrazine solution dissolved in 2 M hydrochloric acid. The samples were mixed and incubated for an hour at room temperature (20–22°C) and centrifuged at 3000 rpm for 15 minutes. The supernatant was removed, and the sediment was mixed with 3 ml of ethanol-ethyl ether mixture (1:1) and centrifuged at 3000 rpm for 5 minutes. The procedure involving sediment washing was repeated three times. The washed sediment was dried at room temperature. After that, it was dissolved in 3 ml of 8 M urea solution. To determine the content of carbonated proteins, the optical density of the resulting solution was measured on a spectrophotometer at 363 nm. The results are presented as nmol per mg of mitochondrial protein.
To obtain serum, blood was collected in glass tubes without anticoagulant. The serum was separated from the blood clot, centrifuged for 15-20 min at 1000 g, and then stored at (-20° C) for evaluation of biochemical parameters [25].
The study of biochemical parameters was carried out on a semi-automatic open-type biochemical analyzer BTS 330 (BioSystems, Spain) with reagent kits manufactured by BioSystems (Spain), and the thymol test was carried out with reagents manufactured by Lachema (Czech Republic).
The activity of alanine aminotransferase (ALAT) (EC 2.6.1.2) and aspartate aminotransferase (ASAT) (EC 2.6.1.1) was determined in blood serum by the UV-kinetics method (IFCC), alkaline phosphatase (ALP) (EC 3.1.3.1) - by the UV-kinetics method (IFCC) with 2-amino-2-methyl-1-propanol buffer (AMP) [26] and the ASAT/ALAT ratio was calculated (De Ritis coefficient) [27].
The amount of total and direct bilirubin was determined by the method of Jendrassik and Grof [28].
The thymol test was determined by the intensity of turbidity due to the formation of a globulin-thymol-lipid complex [29].
Histology. For histological studies, liver pieces were taken, fixed in 10% neutral formalin solution and embedded in paraffin. The preparations were stained with hematoxylin and eosin, which were used to study the normal liver structure, as well as the nature and depth of morphological changes after tetrachloromethane intoxication and its complex correction with the studied drugs. Hematoxylin and eosin staining is the most common method of section staining. This method allows you to establish the relationship between the parts of the organ, perfectly revealing all cellular elements and some non-cellular structures. This staining is double: hematoxylin - the main dye - stains the cell nuclei, eosin - an acidic dye - stains the cell protoplasm and, to a lesser extent, various non-cellular structures. Paraffin sections were passed through a panel of alcohols, stained with Mayer's Hematoxylin, then with Eosin and placed under glass for further tissue study. For coloring, ready-made dyes produced by BioPrime, Kronshagen, Germany were used.
Immunohistochemistry. Paraffin blocks with liver tissue were processed as described [30]. Non-specific staining of sections was blocked with 50% FCS and 0.3% Triton X-100 in PBS for 30 min at 37°C followed by incubation with peroxidase, avidin and biotin. Sections were incubated with primary antibodies at 4°C overnight and then incubated with biotinylated secondary antibodies (BioPrime, Kronshagen, Germany), avidin-conjugated peroxidase (BioPrime, Kronshagen, Germany) and processed using 3,3'-diaminobenzidine substrate (BioPrime, Kronshagen, Germany).
Statistical processing of data was performed using the nonparametric Wilcoxon-Mann-Whitney method using the GraphPad Prism 6.0 software package.
3. Results
3.1. Hepatoprotective Effect of DCTP on the Model of Toxic Liver Damage by Carbon Tetrachloride.
The effect of carbon tetrachloride was manifested by an increase in the activity of ALT - by 3.47, AST - by 2.08, ALP - by 2.55, total bilirubin - by 2.06, direct - by 1.69, thymol test by 5.13 times in the blood serum compared with these indicators of the control group (Table 1). The activity of ALT after the administration of DCTP to animals with hepatitis (IV) decreased by 59.1% compared with the control (II) and corresponded to the activity of the comparison group (III). If we compare the activity of ALT in animals with hepatitis receiving DCTP (IV), it remained slightly increased (Table 1). It should be noted that the administration of TTZ to animals with hepatitis (IIІ) caused a similar effect, as did DCTP (Table 1).
The AST activity in rats with experimental hepatitis after administration of TTZ (III) and DCTP (IV) to animals significantly decreased by 20.4% and 43.0%, respectively, compared to the control (II), which may indicate the regeneration of liver mitochondria (Table 1). Administration of DCTP to animals (V) led to positive changes in AST compared to group III, the difference between the indicators was 28.4% (Figure 2). Bulleted lists look like this:
The value of the De Ritis coefficient after tetrachloromethane administration to experimental animals increased significantly in group III – by 91.8%, in group IV – by 40.9% compared to the control (II) (Table 1). It should be noted that the effect of TTZ and DCTP on animals with hepatitis was different in this indicator. In the experimental group III, it increased by 26.5% (P ≤ 0.05) compared to group IV (Table 1).
As is known, an increase in the activity of alkaline phosphatase in the blood serum is used as a marker of cholestasis. The activity of this enzyme in animals with hepatitis treated with TTZ (Group III) and DCTP (Group IV) decreased compared to animals with hepatitis (Group II) by 55.5% and 60.8%, respectively, and did not differ from the intact control (Table 1).
It was also shown that the level of both total and direct bilirubin increased in the blood of animals in the control group. The total bilirubin content in the groups of animals that received TTZ (Group III) and DCTP (Group IV) significantly decreased relative to the control by 46.0% and 32.7%, respectively. It was found that after the administration of TTZ (Group III) and DCTP (Group IV) to animals, a decrease in the direct bilirubin content in the serum was observed by 40.9% and 36.4%, respectively (Figure 2). In the groups of animals with hepatitis that received TTZ (Group III) and DCTP (Group IV), there was a decrease in the thymol test index by 58.3% and 58.8%, respectively, compared to the control (II).
3.2. Some Indicators of the Prooxidant-Antioxidant System in the Studied Groups of Animals
The results of the study showed that against the background of the action of CCl4, the content of diene conjugates in the liver increased by 2 times compared to intact animals, and Schiff bases by 3 times (Table 2).
In the case where animals with hepatitis received DCTP (Group IV) at a dose of 6 mg/100 g and TTZ (Group III) at a dose of 10 mg/100 g, there was a decrease in the amount of DC in the liver by 43.2% and 29.2%, respectively, compared to the control (Group II). The content of Schiff bases in the liver in these groups of animals decreased by 59% and 28.4%, respectively, compared to the control (Group II) (Figure 3).
It should be noted that the effects of the studied compounds (TTZ and DCTP) did not lead to the restoration of the Schiff base content to the level of intact animals (Table 2). These indicators remained higher by 62.5% and 22.6%, respectively, compared to the intact (Group I). Consequently, the action of DCTP provided a more pronounced effect on this indicator compared to Thiotriazoline.
The results of the conducted studies on the content of carbonylated proteins showed that their amount in the liver with hepatitis (Group II) was increased compared to the intact control (Group I) by 2.3 times (Figure 4). This indicates an increase in the rate of free radical oxidation of proteins and disruption of the antioxidant system. In the case when TTZ was administered to animals with hepatitis (Group III), this was accompanied by a decrease in carbonylated proteins in the liver by 35%, and if they received DCTP (Group IV), the amount of oxidized proteins decreased by 50.8% (Figure 4). Consequently, DCTP had a more pronounced effect in reducing the concentration of carbonylated proteins in the liver with hepatitis compared to TTZ (Group III), and this was 24.3% (Figure 4).
These results correlate with the increase in the activity of antioxidant enzymes in animals receiving the studied compounds. It was found that the SOD activity in animals with hepatitis (Group II) was lower than the intact control (Group I) by 53.3% (Figure 5). The administration of TTZ (Group III) and DCTP (Group IV) to animals with experimental hepatitis was accompanied by the restoration of SOD activity to the level of intact rats (Figure 5).
3.3. Immunohistological and Cytological Indices in the Studied Groups of Animals.
Immunohistochemical determination of bilirubin 2 protein in rats with hepatitis (group II) showed that its amount was increased compared to the intact group (group I, Figure 6B), which may indicate the manifestation of oxidative stress in the liver tissue. When TTZ was administered (group III), Nrf2 protein remained elevated compared to the intact group of animals (Figure 6C). At the same time, in animals receiving DCTP (group IV), the content of Nrf2 protein in the liver was reduced compared to group III receiving TTZ and did not differ from the intact control (Figure 6D). It should be noted that rats receiving an aqueous solution of DCTP to correct toxic damage, had not completely restored structure of the liver lobule, however, compared to the control and the group of rats receiving a TTZ solution to correct toxic changes, regeneration of the structure of hepatocytes and tissues is determined (Figure 6D).
It was found that the LCN2 content in rats increased after CCl4 injection (Figure 9B) and correlated with increased AST and ALT activity in the blood serum (Table 1). In addition, toxic damage caused the appearance of severe degenerative changes (macrovesicular hepatocytes) and/or necrosis, as well as fatty degeneration (Figure 7B) accompanied by minor reversible changes (vacuolar hepatocytes) or severe degenerative changes (micro- and macrovesicular).
The toxic effects of CCl4 were manifested in massive central, perivenular and intermediate hemorrhagic coagulative necrosis of the liver lobules compared to intact animals (Figure 7B). They developed large quantities of fat droplets, which also indicates the development of acute toxic liver damage.
The administration of DCTP and the comparison drug, Thiotriazoline, had a significant hepatoprotective effect in acute liver injury, as evidenced by the analysis of biochemical and histological studies (Figure 7C, D)
Histological examination of the livers of rats of the intact group showed the presence of partial, weakly expressed autolysis of hepatocytes while maintaining the structural organization of the liver lobules, blood filling is normal (Figure 8A). Such variations in structural changes in the liver reflect its reactions to the conditions of keeping animals and many unaccounted factors that always occur.
At the initial stages of development of toxic hepatitis induced by CCl4, an increase in the thickness of the Glisson’s capsule, into which immunocompetent cells have been incorporated, is observed (Figure 8B). In the liver of rats, partial autolysis of hepatocytes takes place (1), discomplexation of the liver beams is often encountered (2). The vessels are filled with blood (3), but the Disse space is reduced (4), in which Ito cells are often found. There are few fibroblasts, endothelial cells are found in moderate quantities, most often near the blood vessels. These minor structural changes in the liver, compared with the intact control, were accompanied by minor changes in the activity of specific liver enzymes. In such animals, there is no blood filling in the veins and arteries (3), in some cases hemorrhage in the central vein was observed. The integrity of the endothelium is impaired. The liver lobules are damaged.
In the group of rats given the Thiotriazoline solution (Figure 8C), the vessels are filled with blood (8). The Disse space is enlarged (3), Ito cells are quite common (4). Endothelial cells (5), small amounts of lymphocytes (6) and fibroblasts (7) are visible around the blood vessels. (Figure 8D).
In animals with toxic hepatitis treated with DCTP (Figure 10D), pronounced discomplexation of the liver trabeculum (1) and complete autolysis of hepatocytes (2) are observed. The nuclei are approximately the same shape and size, round. The central vein and blood artery (8), a moderate number of lymphocytes can be observed (6). The Disse space is reduced (3), Ito cells (4) are seen most often in the area of the liver lobules. Numerous fibroblasts and endotheliocytes (5) are found in the preparation, most often located near the vessels. Multiple ruptures are observed throughout the preparation. (Figure 18D).
3.4. Some Physiological Indicators in the Studied Groups of Animals
The conducted studies showed that when the hepatotoxic agent CCl4 was introduced, the lethality of animals increased depending on the chemical damage to liver cells (hepatocytes). The survival rate of animals was: intact (I) – 10 rats, control (II) – 7 rats, TTZ (III) – 8, the test compound (IV) – 10 animals. The highest survival rate (100%) was noted after the administration of the DCTP compound in group (IV).
Also, during the experiment, a change in the weight of the experimental animals was observed, especially an increase in the average weight was observed in the control group (II). A change in weight was observed in all groups in the first days after the administration of CCl4 (Figure 2), and after 5-6 days the weight began to recover. Weight growth in the control group (II) was observed due to the formation of adhesions between liver particles, the formation of a capsule around the liver (Figure 3B), as well as a general increase in liver weight by 30%.
The liver of rats from groups III and IV, which received TTZ and DCTP substance, was anatomically no different from the intact group (I).
4. Discussion
The liver plays an important role in detoxification and metabolic processes, but despite its strong regenerative capacity, it is also responsible for damage caused by chemicals, drugs and environmental toxicants [31]. It is known that CCl4 is often used to model acute hepatitis in experimental animals due to the induction of oxidative stress, lipid peroxidation, formation of carbonylated proteins and activation of inflammatory processes [32].
During oxidative stress, toxic metabolites of CCl4 cause a decrease in enzyme activity and a disruption of antioxidant protection (SOD, ALAT, ASAT, ALP, BT, BD, thymol test), while simultaneously enhancing the manifestation of prooxidant markers (carbonyl proteins, Schiff bases, diene conjugates, O2-). This leads to the development of oxidative stress and damage to liver cells [15,33].
The study revealed a high content of carbonylated proteins in the liver mitochondria of animals under oxidative stress, which leads to the loss of their biological activity. In the scientific literature, much attention has been paid to the study of prooxidant changes, where the significant role of oxidative modification of proteins (OMP) in the development of liver disorders is emphasized. OMP is considered one of the first and most reliable markers of tissue damage in pathologies associated with free radicals [34]. Carbonylated proteins are relatively stable compounds that are formed because of metal-catalyzed oxidation of proline, arginine, lysine or threonine residues, leading to the formation of Michael adducts [35].
There are many pathways for the formation of carbonylated proteins in the cell. They can be formed not only through direct oxidation, but also with the participation of lipid peroxidation products, as well as in the process of glycation or glycoxidation of lysine amino groups [36]. An increased level of carbonylated proteins (CP) is traditionally associated with carbonyl stress [37], which is a significant pathogenetic factor. This process leads to the suppression of enzymatic activity, changes in the structural organization of membrane proteins, and disruption of protein folding, which is especially noticeable in the carbonylation of Hsp90 chaperones [38,39].
However, studies have shown that in addition to pathological effects, carbonylated proteins also perform important physiological functions. They participate in the regulation of gene expression associated with antioxidant protection (via the NF-E2-related factor 2) [40], and also play a role in signaling mechanisms regulating various cellular processes [41].
During the lipid peroxidation phase, unneutralized toxic CCl4· radicals form covalent bonds with proteins and lipids of hepatocyte membranes, as well as with membranes of mitochondria and the endoplasmic reticulum. The reactive CCl3O2· radical then initiates the lipid peroxidation process, which leads to morphological and functional damage to liver cells [42].
In the inflammatory phase, CCl4· free radicals cause hypertrophy and hyperplasia of Kupffer cells, which in turn begin to produce and secrete many toxic and proinflammatory compounds. This increases damage to liver parenchymal cells [43]. During oxidative stress, toxic CCl4 metabolites contribute to an increase in the activity of liver damage markers (Nrf2 and Lipocalin-2) by triggering lipid peroxidation processes and the destruction of polyunsaturated fatty acids and phospholipids [33].
Lipocalin-2 (LCN2) acts as an early biomarker of liver inflammation, the level of which correlates with the severity of organ damage. Acute inflammatory and toxic liver damage, as well as secreted proinflammatory cytokines (IL-1β, IL-6, TNF-α), synthesized by activated Kupffer cells, are powerful stimulators of LCN2 expression in damaged hepatocytes. The highest immunohistochemical detection of LCN2 is observed in the centrilobular zone, which coincides with the morphological distribution of damaged hepatocytes in the liver acinus.
At the same time, released LCN2 stimulates Kupffer cells to release other chemokines, which attract neutrophils and monocytes to the area of toxic damage and inflammation [44,45].
The presented results demonstrate that acute toxic effects of CCl4 caused a significant increase in the activity of enzymes (ALT, AST, alkaline phosphatase, SOD), prooxidant markers of LPO (diene conjugates and Schiff bases), carbonylated proteins, as well as the level of direct and total bilirubin. These indicators significantly exceeded the values of the control group without treatment, which is consistent with the data of other studies [46,47].
The resulting shifts in liver metabolism were normalized using the reference drug Thiotriazoline (a hepatoprotective agent from the triazole group with antioxidant action) [48]. The administration of DCTP to rats with experimental acute hepatitis led to a decrease in the cytolytic process, regeneration of the functional activity of the liver and was not inferior in these indicators to the well-known drug TTZ, and in some indicators (the level of ASAT, diene conjugates, Schiff bases and carbonylated proteins) even exceeded TTZ.
The occurrence and development of toxic effects in the liver is caused by the activation of lipid peroxidation processes, oxidative modification of proteins, nucleic acids, oxidative damage to cell membranes, which leads to liver dysfunction [33] and is the main factor in the formation of oxidative stress [49].
At the next stage of the work, the influence of CCl4 on some indices of the prooxidant-antioxidant system in the liver [15] in the studied groups of animals was determined. For this purpose, the concentrations of primary (diene conjugates) and final (Schiff bases) were determined.
Under the influence of oxidative stress, the administration of the DCTP compound leads to a significant decrease in biochemical parameters, has a powerful antioxidant activity, blocks oxidative stress, prevents cell death by preventing OMP, can activate antioxidant enzyme systems by activating SOD and inhibiting active oxygen species (AOS).
Studies conducted on an experimental model of CCl4-induced hepatitis have proven the ability of the synthesized substance to reduce the concentration of LPO products. Data have been obtained on the ability of the DCTP substance to stimulate the protein-synthesizing function of the liver and prevent protein catabolism, inhibit inflammatory reactions and cytolysis of hepatocytes, because of which the bile secretory and detoxifying function of the liver is restored.
Consequently, the studied drugs reduced the manifestation of oxidative stress in animals with hepatitis and this was expressed to a greater extent for DCTP compared to Thiotriazoline.
Restoration of SOD activity may be one of the reasons for the decrease in antioxidant activity. In the 3rd and 4th groups of rats with experimental hepatitis, an increase in the SOD level was noted, indicating an increase in the compensatory mechanisms of antioxidant systems by 2.2 and 2.3 times compared to the control. With the administration of TTZ (III) and compound (IV), the SOD indicator was restored to 13.56 ± 1.01 and 14.19 ± 0.96, respectively, which is equal to the value of intact rats (group I). (Figure 5).
Oxidative stress underlies most liver diseases, including pharmacological liver injury, viral hepatitis, and alcoholic hepatitis. The Kerch system, ECH-associated protein 1-NFE2-associated factor 2 (Keap1-C), is an important protective mechanism of cells and organisms against oxidative stress. Its regulation reduces drug-induced liver injury in rats. In addition, many natural Nrf2 activators regulate lipid metabolism and oxidative stress in hepatocytes, thereby reducing mice fatty liver disease [50]. Increased Nrf2 is observed in acute hepatitis, fatty liver disease, as well as viral hepatitis and apoptosis [51]. Nrf2 also plays an important role in the activation of antioxidant enzymes, regulating their transcription [50].
Lipocalin-2 (LCN2) is expressed in pathological conditions such as intoxication, infection, inflammation and other forms of cellular stress. Experimental liver injury causes rapid and sustained formation of LCN2 by damaged hepatocytes. However, the exact biological function of LCN2 in the liver is still unknown. In this work, the content of LCN2 was determined on histological preparations in the studied liver samples [52].
The administration of the test substance significantly reduced the activity of specific damage biomarkers (Nrf2 and Lipocalin-2), which indicates its ability to protect hepatocyte membranes and organelles from the toxic effects of CCl4.
According to the literature, fibrotic changes involve a larger number of cells (fibroblasts, vascular cells, infiltrating immune cells, and biliary epithelial cells) [53]. Glisson's capsule is a layer of interstitial tissue that can also be classified as visceral fascia [54,55,56]. It surrounds the liver and is continuous with the interstitial spaces and matrix surrounding the portal triads, likely playing a role in fluid exchange and cell migration in the hepatobiliary system [56,58]. The capsule is known to be constructed in such a way that it allows for diurnal variations in size due to changes in hepatocyte size after meals, as well as circadian regulation by 34% in mice and 10-15% in humans [59]. To allow for this size variability, the normal liver capsule is thin, well innervated, and contains many elastic fibers, as is characteristic of the visceral fascia [60]. These characteristics create a unique niche in which different populations of both macrophages and fibroblasts coexist under normal physiological conditions [61]. As described, visualization of the surface of fibrous rat liver reveals altered collagen organization with a denser collagen network and loss of the characteristic fiber waviness. These changes are highly dependent on disease severity [62].
Therefore, to predict the degree of liver fibrosis in toxic hepatitis, it is possible to analyze the presence of changes, namely thickening of the Glisson’s capsule as a marker of fibrosis, which indicates changes in the capsular matrix.
Other rat studies have shown that in addition to changes in the matrix, the capsular cell population also changed, with mesothelial cells and fibroblasts migrating into the liver parenchyma and contributing to the formation of a myofibroblast population [61,63,64]. The capsule is an active site of pathology in rats’ liver fibrosis, with mesothelial cells, fibroblasts, and macrophages embedded in a thin layer of matrix.
Our study showed that animals treated with DCTP at the initial stages of fibrosis development had minor histological changes, indicating its pronounced hepatoprotective effect. The most pronounced deviations in this group were thickening of the Glisson’s capsule.
The dynamics of the relative liver mass on the 3rd-5th day after the administration of tetrachloromethane in all groups of animals were also observed. Changes were observed in the control group of animals, which were not subject to correction of pathological conditions. In the groups of animals subjected to treatment, the mass indices were at the intact level, coinciding with the studies of other authors [65].
The hepatoprotective effect of the studied substance is probably due to its structural similarity with tetrahydrofolic acid, an integral component of oxidation-reduction processes and a participant in the biosynthesis of nitrogenous bases of nucleotides and amino acids.
5. Conclusions
Experimental animals’ liver damage by CCl4 resulted cell membrane damage, increased LPO, increased AST, ALT, ALP, total and direct bilirubin, and thymol test values. The resulting shifts in liver metabolism were normalized by the administration of the studied pteridine derivative, which exceeded the effect of the TTZ reference solution in terms of its effect on the level of AST, DC, Schiff bases, and carbonylated proteins.
It was shown that the studied substance DCTP significantly reduces acute hepatotoxic effects caused by CCl4, as evidenced by a decrease in the level of lipid peroxidation and prooxidant markers, normalization of liver biochemical markers, as well as regeneration of liver architecture, limitation of inflammatory effects, a decrease in Nrf2 and Lipocalin-2 markers, and induction of liver antioxidant enzymes.
Our results indicate antioxidant and hepatoprotective properties of the studied S-substituted pteridine. Detailed data of the studied substance and the mechanisms underlying them require further research.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, V.S. and S.K.; methodology, A.B. and H.M.; software, O.A and O.V.; validation, A.B and H.M.; formal analysis, A.B. and H.M.; investigation, N.L., O.A. and V.S.; resources, O.V., V.O and O.K.; data curation, S.O. and S.K.; writing—original draft preparation, N.L. and O.A.; writing—review and editing, V.S., S.K. and O.A.; visualization, N.L. and O.V.; supervision, V.S.; project administration, S.K. and S.O.; funding acquisition, V.O. and O.K.
Funding
This work was funded by the Ministry of Education and Science of Ukraine, № 0125U001851, the name of the project "New azaheterocycles: molecular design, synthesis, prospects for use for pharmacocorrection of post-traumatic stressful disorders and metabolic syndrome".
Institutional Review Board Statement
The animal study protocol was approved by Ethics Committee of Zaporizhzhia National University (protocol №6, date of approval 27 January 2025).
Informed Consent Statement
Not applicable.
Acknowledgments
Authors gratefully acknowledge Armed Forces of Ukraine with Territorial Defense Forces of the Armed Forces of Ukraine for preparing this paper in the safe conditions of Zaporizhzhia and Dnipro, Ukraine.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of Liver Diseases in the World. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Conway, R.; Carey, J.J. Risk of Liver Disease in Methotrexate Treated Patients. World J. Hepatol. 2017, 9, 1092. [Google Scholar] [CrossRef] [PubMed]
- O'Shea, R.S.; Dasarathy, S.; McCullough, A.J. Alcoholic Liver Disease. Hepatology 2009, 51, 307–328. [Google Scholar] [CrossRef] [PubMed]
- Saeter, G.; Lee, C.Z.; Schwarze, P.K.; Ous, S.; Chen, D.S.; et al. Changes in Ploidy Distributions in Human Liver Carcinogenesis. J. Natl. Cancer Inst. 1988, 80, 1480–1484. [Google Scholar] [CrossRef]
- Reyes-Gordillo, K.; Shah, R.; Muriel, P. Oxidative Stress and Inflammation in Hepatic Diseases: Current and Future Therapy. Oxid. Med. Cell. Longev. 3140673. [CrossRef]
- Nolan, J.P. The Role of Intestinal Endotoxin in Liver Injury: A Long and Evolving History. Hepatology 2010, 52, 1829–1835. [Google Scholar] [CrossRef]
- Hoffman, D.R.; Marion, D.W.; Cornatzer, W.E.; Duerre, J.A. S-Adenosylmethionine and S-Adenosylhomocysteine Metabolism in Isolated Rat Liver. J. Biol. Chem. 1980, 255, 10822–10827. [Google Scholar] [CrossRef]
- Lan, X.; Field, M.S.; Stover, P.J. Cell Cycle Regulation of Folate-Mediated One-Carbon Metabolism. Wiley Interdiscip. Rev. Syst. Biol. Med. 2018, 10, e1426. [Google Scholar] [CrossRef]
- Hine, C.; Harputlugil, E.; Zhang, Y.; Ruckenstuhl, C.; Lee, B.C.; et al. Endogenous Hydrogen Sulfide Production is Essential for Dietary Restriction Benefits. Cell 2015, 160, 132–144. [Google Scholar] [CrossRef]
- Sanderson, S.M.; Gao, X.; Dai, Z.; Locasale, J.W. Methionine Metabolism in Health and Cancer: A Nexus of Diet and Precision Medicine. Nat. Rev. Cancer 2019, 19, 625–637. [Google Scholar] [CrossRef]
- DrugBank. DB12687. Available online: https://go.drugbank.com/drugs/DB12687 (accessed on 12 of December 2024).
- Kazunin, M.S.; Groma, N.V.; Nosulenko, I.S.; Kinichenko, A.O.; Antypenko, O.M.; Shvets, V.M.; Voskoboinik, O.Y.; Kovalenko, S.I. Thio-Containing Pteridines: Synthesis, Modification, and Biological Activity. Arch. Pharm. 2022, 355, 2200252. [Google Scholar] [CrossRef] [PubMed]
- Lohvinenko, N.; Shvets, V.; Berest, G.; Nosulenko, I.; Voskoboinik, O.; Severina, H.; Okovytyy, S.; Kovalenko, S. Prospects for the Use of Sulfur-Containing Pteridines in Toxic Liver Damage. Regul. Mech. Biosyst. 2024, 15, 374–381. [Google Scholar] [CrossRef]
- Milstien, S.; Kapatos, G.; Levine, R.A.; Shane, B. Chemistry and Biology of Pteridines and Folates; Springer US: Boston, MA, USA, 2002. [Google Scholar] [CrossRef]
- Groma, N.; Berest, G.; Antypenko, O.; Voskoboinik, O.; Kopiika, V.; et al. Evaluation of the Toxicity and Hepatoprotective Properties of New S-Substituted Pteridins. Curr. Issues Pharm. Med. Sci. 2023, 36(1), 21–26. [Google Scholar] [CrossRef]
- Kozhem'yakin, Yu.M.; Khromov, O.S.; Filonenko, M.A. Scientific and practical recommendations for the development of laboratory creatures and robots. K.: Avitsena; 2002; 156.
- European Convention for the Protection of Vertebrate Animals Used for Experimental and Other Scientific Purposes. European Treaty Series - No. 123. Strasbourg: Council of Europe; 1986.
- Stefanov, O.V. Preclinical Study of Drugs (Methodical Recommendation). Kyiv: Avitsena; 2001; 527.
- Bhakuni, G.S.; Bedi, O.; Bariwal, J.; Deshmukh, R.; Kumar, P. Animal Models of Hepatotoxicity. Inflammation Research 2015, 65, 13–24. [Google Scholar] [CrossRef]
- Mazur, I.A.; Voloshin, N.A.; Chekman, I.S. Thiotriazolin: Pharmacological Aspects and Clinical Application. Lvov: Zaporizhye; 2005; 160.
- Recknagel, R.O.; Ghoshal, A.K. Quantitative Estimation of Peroxidative Degeneration of Rat Liver Microsomal and Mitochondrial Lipids After Carbon Tetrachloride Poisoning. Exp. Mol. Pathol. 1966, 5, 413–426. [Google Scholar] [CrossRef]
- Symons, M.C.R.; Diplock, A.T.; Rice-Evans, C.A. Techniques in Free Radical Research; Elsevier Science & Technology Books: 1991. ISBN 978008085 8913.
- Kostyuk, V.A. A Simple and Sensitive Method for Determining Superoxide Dismutase Activity Based on the Oxidation Reaction of Quercetin. Questions Med. Chem. 1990, 36, 88–91. [Google Scholar]
- Levine, R.L.; Stadtman, E.R. Oxidative Modification of Proteins During Aging. Exp. Gerontol. 2001, 36, 1495–1502. [Google Scholar] [CrossRef]
- Vlizlo, V.V.; Fedoruk, R.S.; Ratych, I.B.; Vishurt, O.I.; Sharan, M.M.; Vudmaska, I.V.; Fedorovych, E.I. Laboratory Research Methods in Biology, Animal Husbandry and Veterinary Medicine: A Guide; Spolom: Lviv, 2012; 764p. [Google Scholar]
- Schumann, G.; Klauke, R.; Canalias, F.; Bossert-Reuther, S.; Franck, P.F.H.; et al. IFCC Primary Reference Procedures for the Measurement of Catalytic Activity Concentrations of Enzymes at 37 °C. Part 9: Reference Procedure for the Measurement of Catalytic Concentration of Alkaline Phosphatase. Clin. Chem. Lab. Med. 2011, 49, 9. [Google Scholar] [CrossRef]
- Davydov, V.V.; Shvets, V.N. Guide to Practical Classes in Biological Chemistry (for Students of Medical Schools III-IV Level of Accreditation); KhNU Named after V.N. Karazin: Kharkiv, 2011; 316p, ISBN 978-966-623-774-6. [Google Scholar]
- Jendrassik, L.; Grof, P. Colorimetric Method of Determination of Bilirubin. Biochem. Z. 1938, 297, 81–82. [Google Scholar]
- Menshikov, V.V. Laboratory Research Methods in Clinic. Meditsina 1987, 174–177. [Google Scholar]
- Borkham-Kamphorst, E.; Kovalenko, E.; Van Roeyen, C.R.C.; Gassler, N.; Bomble, M.; et al. Platelet-Derived Growth Factor Isoform Expression in Carbon Tetrachloride-Induced Chronic Liver Injury. Lab. Invest. 2008, 88, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-Y.; Yuan, W.-G.; He, P.; Lei, J.-H.; Wang, C.-X. Liver Fibrosis and Hepatic Stellate Cells: Etiology, Pathological Hallmarks and Therapeutic Targets. World J. Gastroenterol. 2016, 22, 10512. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; et al. Dose and Time Dependent Effects of Intraperitoneal Administration of Carbon Tetrachloride (CCl4) on Blood Lipid Profile in Wistar Rats. EC Pharmacol. Toxicol. 2020, 8, 21–25. [Google Scholar]
- Manhar, N.; Singh, S.K.; Yadav, P.; Bishnolia, M.; Khurana, A.; et al. Methyl Donor Ameliorates CCl4-Induced Nephrotoxicity by Inhibiting Oxidative Stress, Inflammation, and Fibrosis Through the Attenuation of Kidney Injury Molecule 1 and Neutrophil Gelatinase-Associated Lipocalin Expression. J. Biochem. Mol. Toxicol. 2025, 39, 3. [Google Scholar] [CrossRef]
- Stefan, N.; Kantartzis, K.; Häring, H.-U. Causes and Metabolic Consequences of Fatty Liver. Endocr. Rev. 2008, 29, 939–960. [Google Scholar] [CrossRef]
- Akagawa, M. Protein Carbonylation: Molecular Mechanisms, Biological Implications, and Analytical Approaches. Free Radic. Res. 2020, 1–37. [Google Scholar] [CrossRef]
- Friguet, B.; Bulteau, A.-L.; Chondrogianni, N.; Conconi, M.; Petropoulos, I.-B. Protein Degradation by the Proteasome and Its Implications in Aging. Ann. N. Y. Acad. Sci. 2006, 908, 143–154. [Google Scholar] [CrossRef]
- Davydov, V.V.; Bozhkov, A.I.; Kulchitsky, O.K. Physiological and Pathophysiological Role of Endogenous Aldehydes; Palmarium Academic Publishing: Saarbrucken, 2012; 240p. [Google Scholar]
- Carbone, D.L.; Doorn, J.A.; Kiebler, Z.; Ickes, B.R.; Petersen, D.R. Modification of Heat Shock Protein 90 by 4-Hydroxynonenal in a Rat Model of Chronic Alcoholic Liver Disease. J. Pharmacol. Exp. Ther. 2005, 315, 8–15. [Google Scholar] [CrossRef]
- Carbone, D.L.; Doorn, J.A.; Kiebler, Z.; Petersen, D.R. Cysteine Modification by Lipid Peroxidation Products Inhibits Protein Disulfide Isomerase. Chem. Res. Toxicol. 2005, 18, 1324–1331. [Google Scholar] [CrossRef]
- Walters, D.M.; Cho, H.-Y.; Kleeberger, S.R. Oxidative Stress and Antioxidants in the Pathogenesis of Pulmonary Fibrosis: A Potential Role for Nrf2. Antioxid. Redox Signal. 2008, 10, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.M.; Cheema, A.K.; Zhang, L.; Suzuki, Y.J. Protein Carbonylation as a Novel Mechanism in Redox Signaling. Circ. Res. 2008, 102, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Knockaert, L.; Berson, A.; Ribault, C.; Prost, P.-E.; Fautrel, A.; et al. Carbon Tetrachloride-Mediated Lipid Peroxidation Induces Early Mitochondrial Alterations in Mouse Liver. Lab. Invest. 2011, 92, 396–410. [Google Scholar] [CrossRef] [PubMed]
- VI International Scientific and Theoretical Conference «Sectoral Research XXI: Characteristics and Features»; Primedia eLaunch LLC, 2023; ISBN 9798889557678. [CrossRef]
- Borkham-Kamphorst, E.; Drews, F.; Weiskirchen, R. Induction of Lipocalin-2 Expression in Acute and Chronic Experimental Liver Injury Moderated by Pro-Inflammatory Cytokines Interleukin-1β Through Nuclear Factor-κB Activation. Liver Int. 2011, 31, 656–665. [Google Scholar] [CrossRef]
- Dahl, S.L.; Woodworth, J.S.; Lerche, C.J.; Cramer, E.P.; Nielsen, P.R.; et al. Lipocalin-2 Functions as Inhibitor of Innate Resistance to Mycobacterium tuberculosis. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- DeCicco, L.A.; Rikans, L.E.; Tutor, C.G.; Hornbrook, K.R. Serum and Liver Concentrations of Tumor Necrosis Factor α and Interleukin-1β Following Administration of Carbon Tetrachloride to Male Rats. Toxicol. Lett. 1998, 98, 115–121. [Google Scholar] [CrossRef]
- Teschke, R.; Vierke, W.; Goldermann, L. Carbon Tetrachloride (CCl4) Levels and Serum Activities of Liver Enzymes Following Acute CCl4 Intoxication. Toxicol. Lett. 1983, 17, 175–180. [Google Scholar] [CrossRef]
- Stepanov, Y.M.; Kosynskaya, S.I. The Use of Thiotriazoline in Patients with Chronic Liver Diseases. Zaporizhzhya Med. J. 2010, 5, 69–70. [Google Scholar]
- Bernabeu-Wittel, M.; Gómez-Díaz, R.; González-Molina, Á.; Vidal-Serrano, S.; Díez-Manglano, J.; et al. Oxidative Stress, Telomere Shortening, and Apoptosis Associated to Sarcopenia and Frailty in Patients with Multimorbidity. J. Clin. Med. 2020, 9, 2669. [Google Scholar] [CrossRef]
- Zhou, J.; Zheng, Q.; Chen, Z. The Nrf2 Pathway in Liver Diseases. Front. Cell Dev. Biol. 2022, 10, 826204. [Google Scholar] [CrossRef]
- Yamashita, Y.; Ueyama, T.; Nishi, T.; Yamamoto, Y.; Kawakoshi, A.; et al. Nrf2-Inducing Anti-Oxidation Stress Response in the Rat Liver - New Beneficial Effect of Lansoprazole. PLoS ONE 2014, 9(5), e97419. [Google Scholar] [CrossRef]
- Asimakopoulou, A.; Weiskirchen, S.; Weiskirchen, R. Lipocalin 2 (LCN2) Expression in Hepatic Malfunction and Therapy. Front. Physiol. 2016, 7, 430. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, J.; Fede, C.; Loneker, A.E.; Friday, C.S.; Hast, M.W.; et al. Glisson’s Capsule Matrix Structure and Function Is Altered in Patients with Cirrhosis Irrespective of Etiology. JHEP Rep. 2023, 100760. [Google Scholar] [CrossRef] [PubMed]
- Stecco, C.; Sfriso, M.M.; Porzionato, A.; Rambaldo, A.; Albertin, G.; et al. Microscopic Anatomy of the Visceral Fasciae. J. Anat. 2017, 231(1), 121–128. [Google Scholar] [CrossRef] [PubMed]
- Allen, W.E. Terminologia Anatomica: International Anatomical Terminology and Terminologia Histologica: International Terms for Human Cytology and Histology. J. Anat. 2009, 215(2), 221. [Google Scholar] [CrossRef]
- Benias, P.C.; Wells, R.G.; Sackey-Aboagye, B.; Klavan, H.; Reidy, J.; et al. Structure and Distribution of an Unrecognized Interstitium in Human Tissues. Sci. Rep. 2018, 8, 23062. [Google Scholar] [CrossRef]
- Hayashi, S.; Murakami, G.; Ohtsuka, A.; Itoh, M.; Nakano, T.; et al. Connective Tissue Configuration in the Human Liver Hilar Region with Special Reference to the Liver Capsule and Vascular Sheath. J. Hepatobiliary-Pancreatic Surg. 2008, 15(6), 640–647. [Google Scholar] [CrossRef]
- Wang, J.; Kubes, P. A Reservoir of Mature Cavity Macrophages That Can Rapidly Invade Visceral Organs to Affect Tissue Repair. Cell 2016, 165(3), 668–678. [Google Scholar] [CrossRef]
- Wagner, B.A.; Venkataraman, S.; Buettner, G.R. The Rate of Oxygen Utilization by Cells. Free Radic. Biol. Med. 2011, 51(3), 700–712. [Google Scholar] [CrossRef]
- Acharya, P.; Chouhan, K.; Weiskirchen, S.; Weiskirchen, R. Cellular Mechanisms of Liver Fibrosis. Front. Pharmacol. 2021, 12, 671640. [Google Scholar] [CrossRef]
- Guilliams, M.; Bonnardel, J.; Haest, B.; Vanderborght, B.; Wagner, C.; et al. Spatial Proteogenomics Reveals Distinct and Evolutionarily Conserved Hepatic Macrophage Niches. Cell 2022, 185, 379–396.e38. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Kang, C.H.; Gou, X.; Peng, Q.; Yan, J.; et al. Quantification of Liver Fibrosis via Second Harmonic Imaging of the Glisson's Capsule from Liver Surface. J. Biophotonics 2015, 9(4), 351–363. [Google Scholar] [CrossRef] [PubMed]
- Balog, S.; Li, Y.; Ogawa, T.; Miki, T.; Saito, T.; et al. Development of Capsular Fibrosis Beneath the Liver Surface in Humans and Mice. Hepatology 2020, 71(1), 291–305. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, J.; Asahina, K. Mesothelial Cells Give Rise to Hepatic Stellate Cells and Myofibroblasts via Mesothelial-Mesenchymal Transition in Liver Injury. Proc. Natl. Acad. Sci. 2013, 110(6), 2324–2329. [Google Scholar] [CrossRef]
- Bozhkov, A.; Ionov, I.; Kurhuzova, N.; Novikova, A.; Katerynych, O.; et al. Vitamin A Intake Forms Resistance to Hypervitaminosis A and Affects the Functional Activity of the Liver. Clin. Nutr. Open Sci. 2022, 41, 82–97. [Google Scholar] [CrossRef]
Figure 1.
Research design (distribution scheme of materials and research methods).
Figure 2.
Differences in the studied biochemical parameters in experimental animals that received TTZ (Thiotriazoline) and DCTP in percentages compared to the control.
Figure 2.
Differences in the studied biochemical parameters in experimental animals that received TTZ (Thiotriazoline) and DCTP in percentages compared to the control.
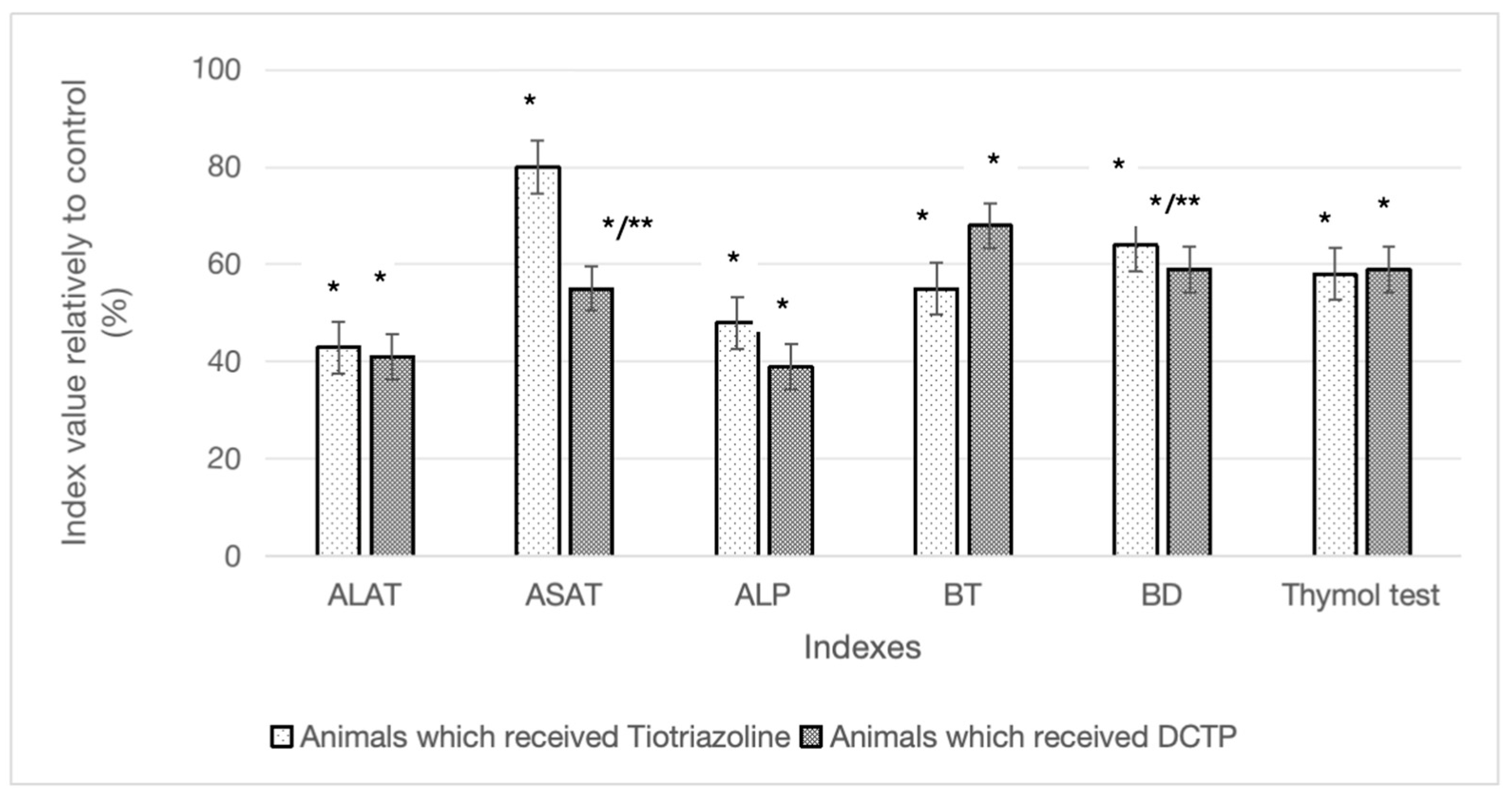
Figure 3.
Differences in the content of diene conjugates and Schiff bases in animals with hepatitis that received TTZ (Thiotriazoline) and DCTP, as a percentage compared to the control (Group II).
Figure 3.
Differences in the content of diene conjugates and Schiff bases in animals with hepatitis that received TTZ (Thiotriazoline) and DCTP, as a percentage compared to the control (Group II).
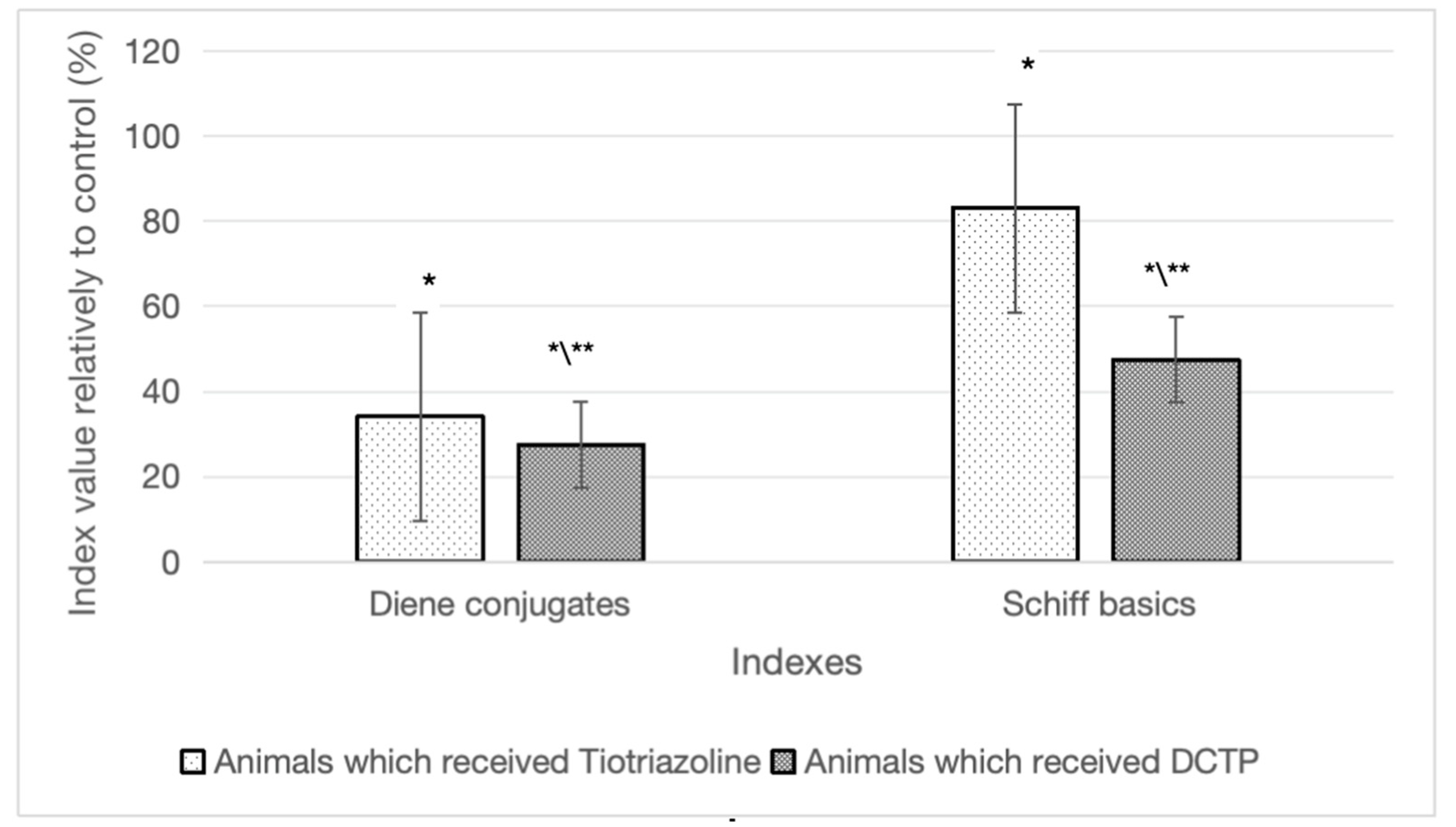
Figure 4.
Carbonyl protein content in liver homogenate of intact rats (intact), rats with hepatitis (control), rats with hepatitis that received Thiotriazoline (TTZ) and DCTP. Mean values and standard errors are shown. There were 10 surviving rats in the intact group, 7 surviving rats in the control group (tetrachloromethane intoxication), 8 surviving rats in the group receiving TTZ (Thiotriazoline) at a dose of 10 mg/100 g, and 10 surviving rats in the group receiving DCTP at a dose of 6 mg/100 g. *Significant difference from intact (P<0.05). **Significant difference from rats with hepatitis (P<0.05).
Figure 4.
Carbonyl protein content in liver homogenate of intact rats (intact), rats with hepatitis (control), rats with hepatitis that received Thiotriazoline (TTZ) and DCTP. Mean values and standard errors are shown. There were 10 surviving rats in the intact group, 7 surviving rats in the control group (tetrachloromethane intoxication), 8 surviving rats in the group receiving TTZ (Thiotriazoline) at a dose of 10 mg/100 g, and 10 surviving rats in the group receiving DCTP at a dose of 6 mg/100 g. *Significant difference from intact (P<0.05). **Significant difference from rats with hepatitis (P<0.05).
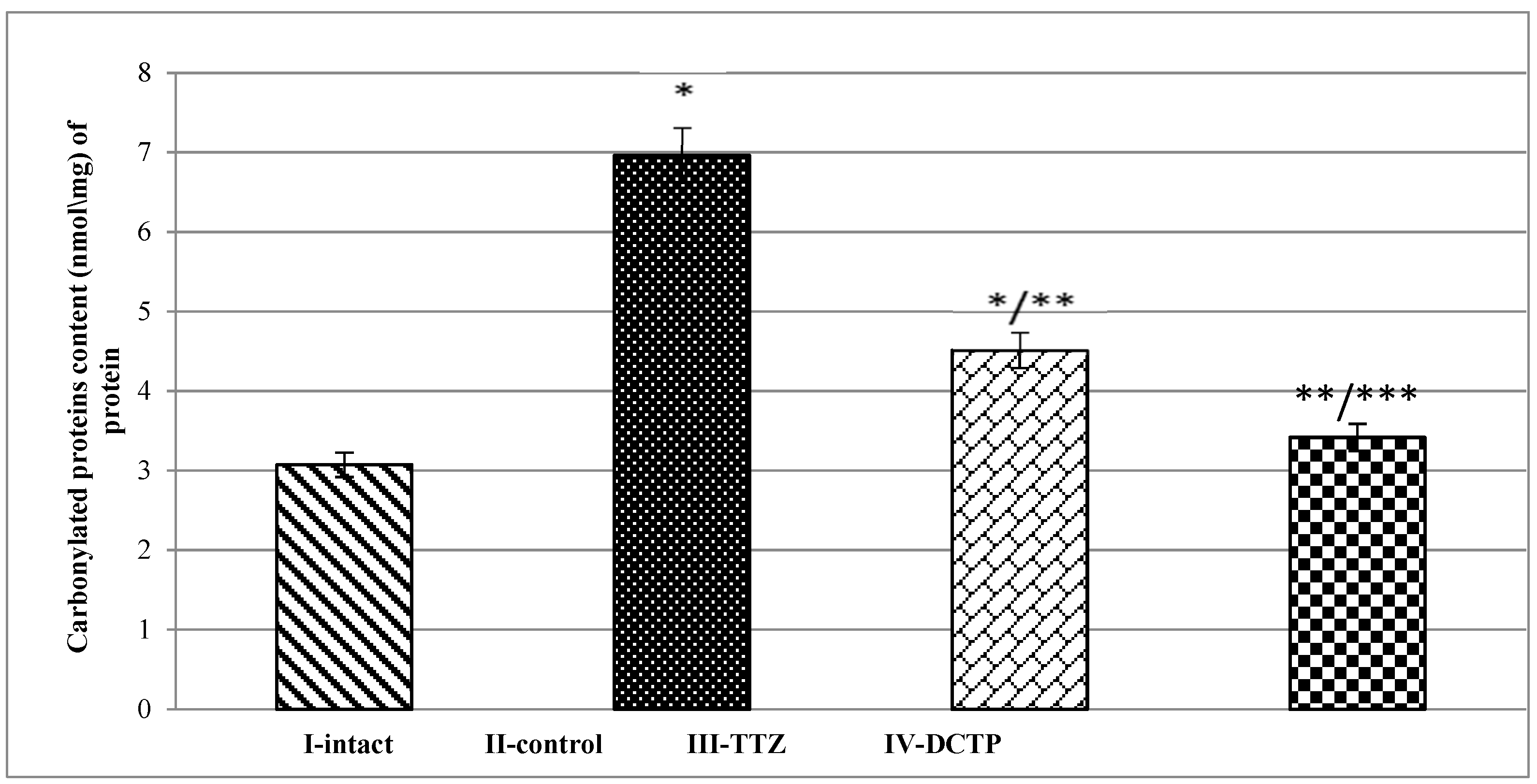
Figure 5.
Superoxide dismutase activity in liver homogenate of intact rats (intact), rats with hepatitis (control), rats with hepatitis that received TTZ (Thiotriazoline) and DCTP. Mean values and standard errors are shown. There were 10 surviving rats in the intact group, 7 surviving rats in the control group (tetrachloromethane intoxication), 8 surviving rats in the group receiving Thiotriazoline at a dose of 10 mg/100 g, and 10 surviving rats in the group receiving DCTP at a dose of 6 mg/100 g. *Significant difference from intact (P<0.05). **Significant difference from rats with hepatitis (P<0.05).
Figure 5.
Superoxide dismutase activity in liver homogenate of intact rats (intact), rats with hepatitis (control), rats with hepatitis that received TTZ (Thiotriazoline) and DCTP. Mean values and standard errors are shown. There were 10 surviving rats in the intact group, 7 surviving rats in the control group (tetrachloromethane intoxication), 8 surviving rats in the group receiving Thiotriazoline at a dose of 10 mg/100 g, and 10 surviving rats in the group receiving DCTP at a dose of 6 mg/100 g. *Significant difference from intact (P<0.05). **Significant difference from rats with hepatitis (P<0.05).
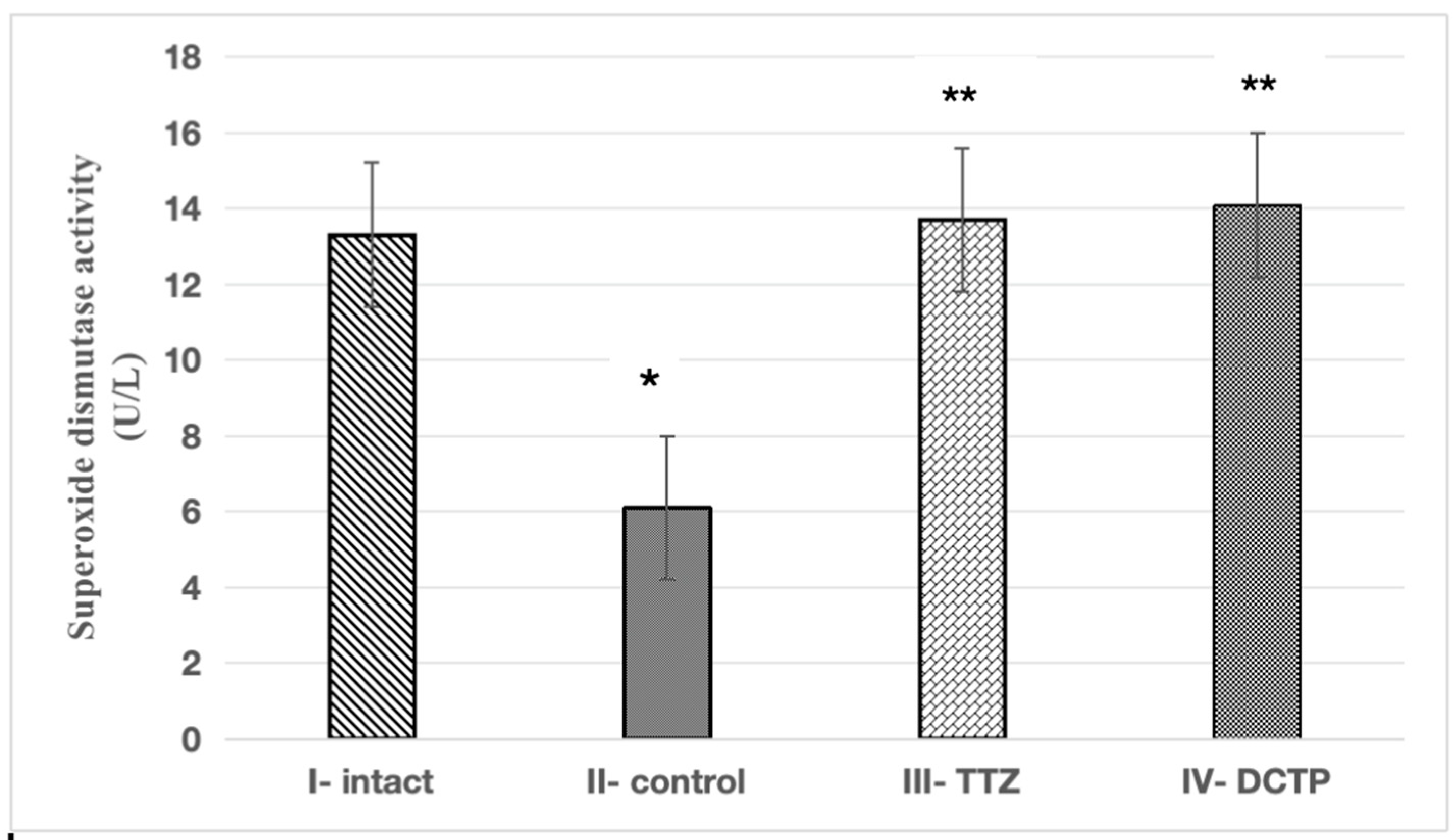
Figure 6.
Rat liver: A – intact group, B – after administration of tetrachloromethane, C – after administration of tetrachloromethane and correction with 2.5% TTZ (Thiotriazoline) solution, D – after administration of tetrachloromethane and correction with aqueous DCTP solution. The prepared preparations were scanned with a ZEISS Axioscan 7 scanner, the images were enlarged at 400-600X.
Figure 6.
Rat liver: A – intact group, B – after administration of tetrachloromethane, C – after administration of tetrachloromethane and correction with 2.5% TTZ (Thiotriazoline) solution, D – after administration of tetrachloromethane and correction with aqueous DCTP solution. The prepared preparations were scanned with a ZEISS Axioscan 7 scanner, the images were enlarged at 400-600X.
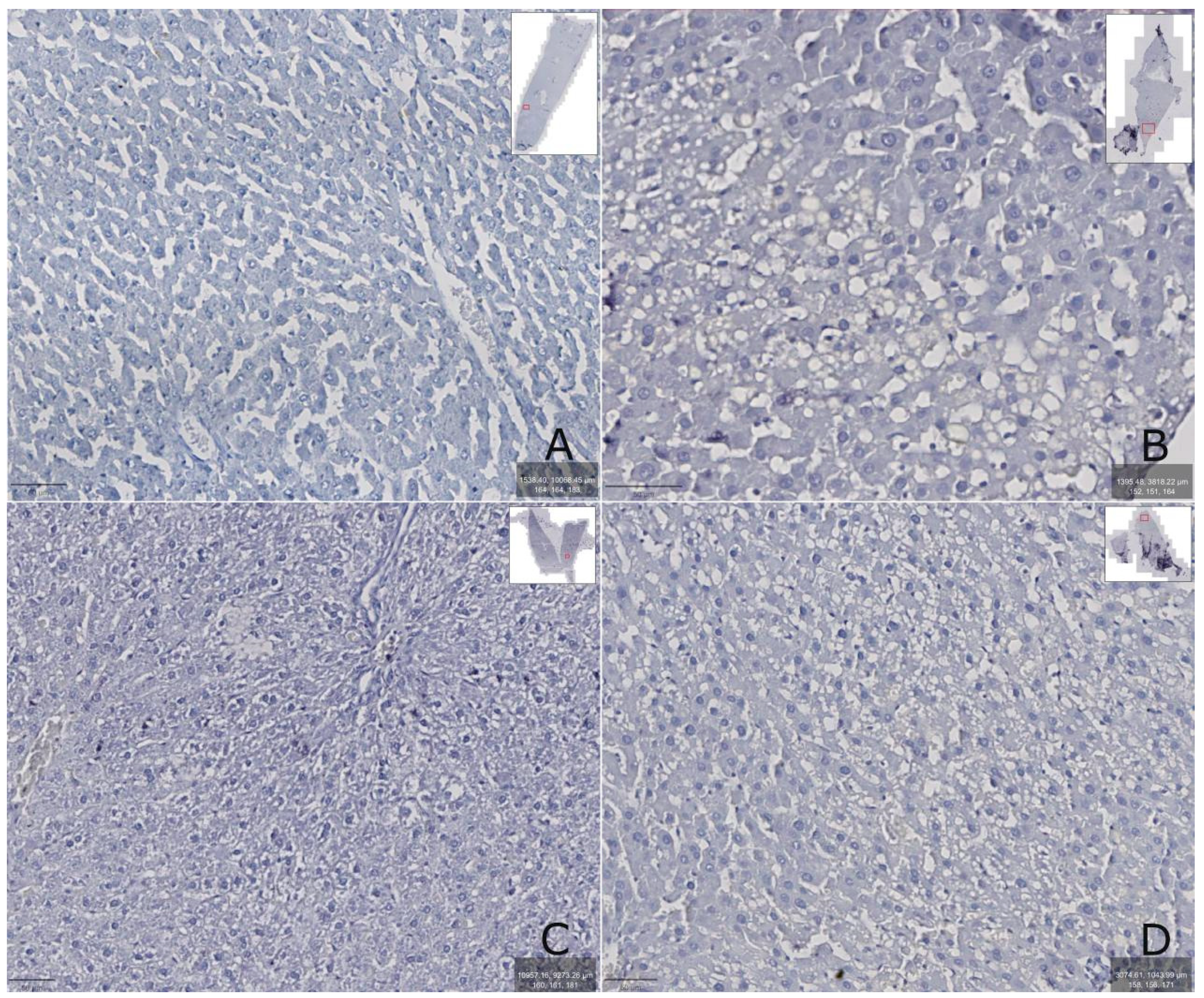
Figure 7.
Rat liver: A – intact group, B – after administration of carbon tetrachloride, C – after administration of carbon tetrachloride and correction with 2.5% TTZ (Thiotriazoline) solution, D – after administration of carbon tetrachloride and correction with aqueous DCTP solution. The prepared preparations were scanned with a ZEISS Axioscan 7 scanner, the images were enlarged at 400-600X.
Figure 7.
Rat liver: A – intact group, B – after administration of carbon tetrachloride, C – after administration of carbon tetrachloride and correction with 2.5% TTZ (Thiotriazoline) solution, D – after administration of carbon tetrachloride and correction with aqueous DCTP solution. The prepared preparations were scanned with a ZEISS Axioscan 7 scanner, the images were enlarged at 400-600X.
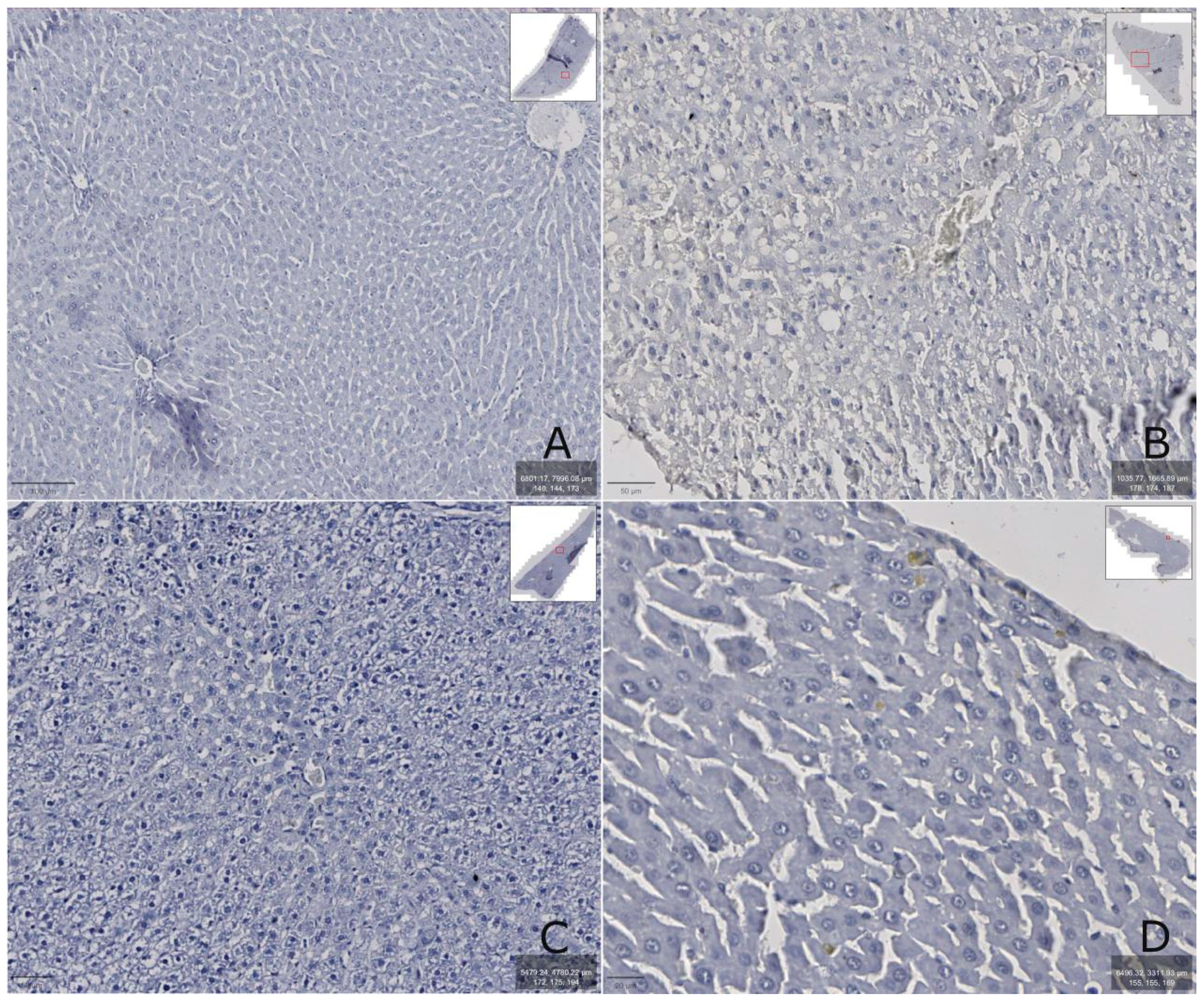
Figure 8.
Liver of rats: A – intact (group I), B – rats with tetrachloromethane hepatitis (group II), C – animals with tetrachloromethane hepatitis and administration of 2.5% Thiotriazoline solution (group III), D – rats with tetrachloromethane hepatitis corrected with an aqueous solution of DCTP (group IV). Hematoxylin-Eosin staining. X200.
Figure 8.
Liver of rats: A – intact (group I), B – rats with tetrachloromethane hepatitis (group II), C – animals with tetrachloromethane hepatitis and administration of 2.5% Thiotriazoline solution (group III), D – rats with tetrachloromethane hepatitis corrected with an aqueous solution of DCTP (group IV). Hematoxylin-Eosin staining. X200.
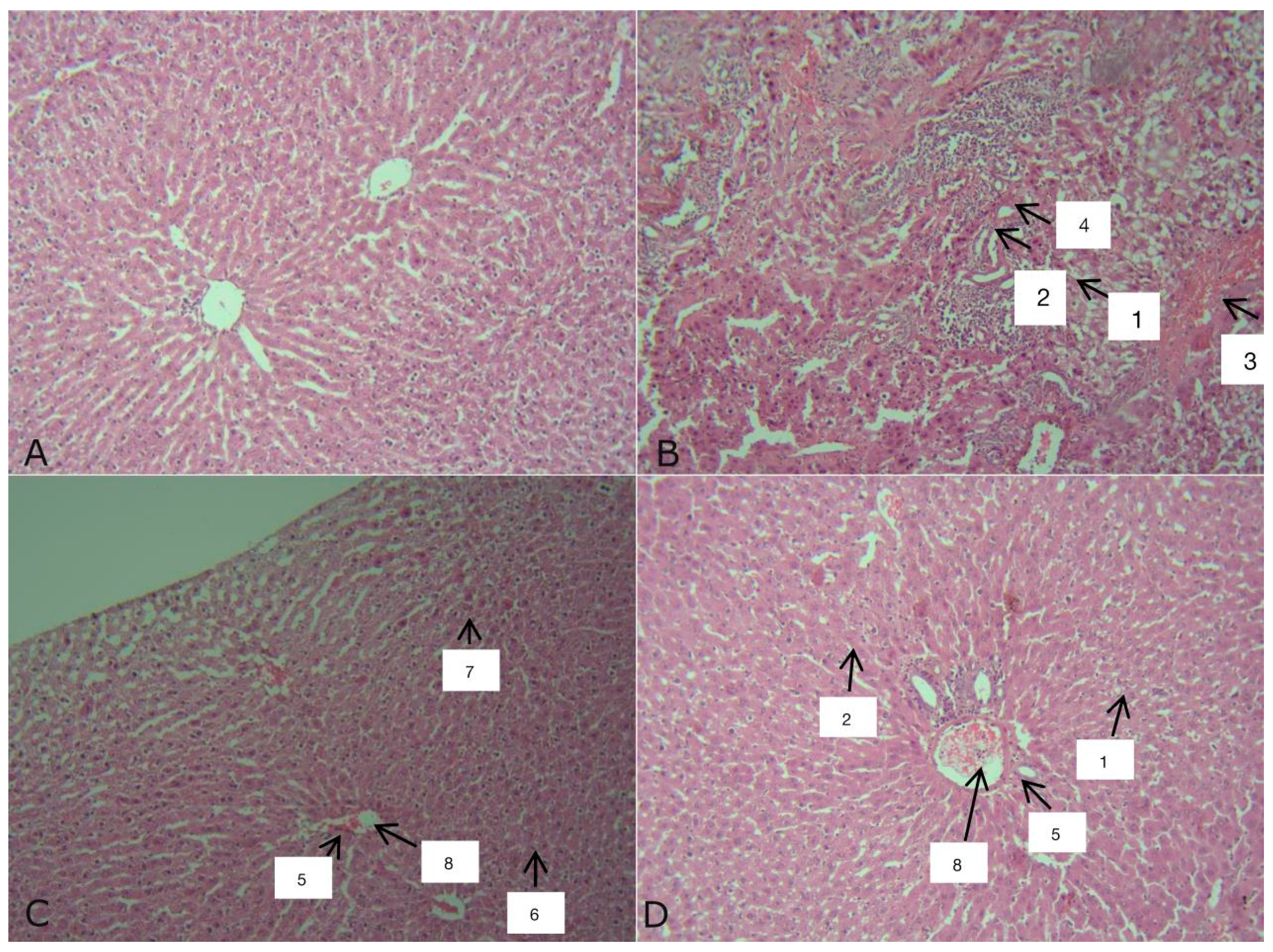
Figure 9.
Change in rat weight during the experiment.
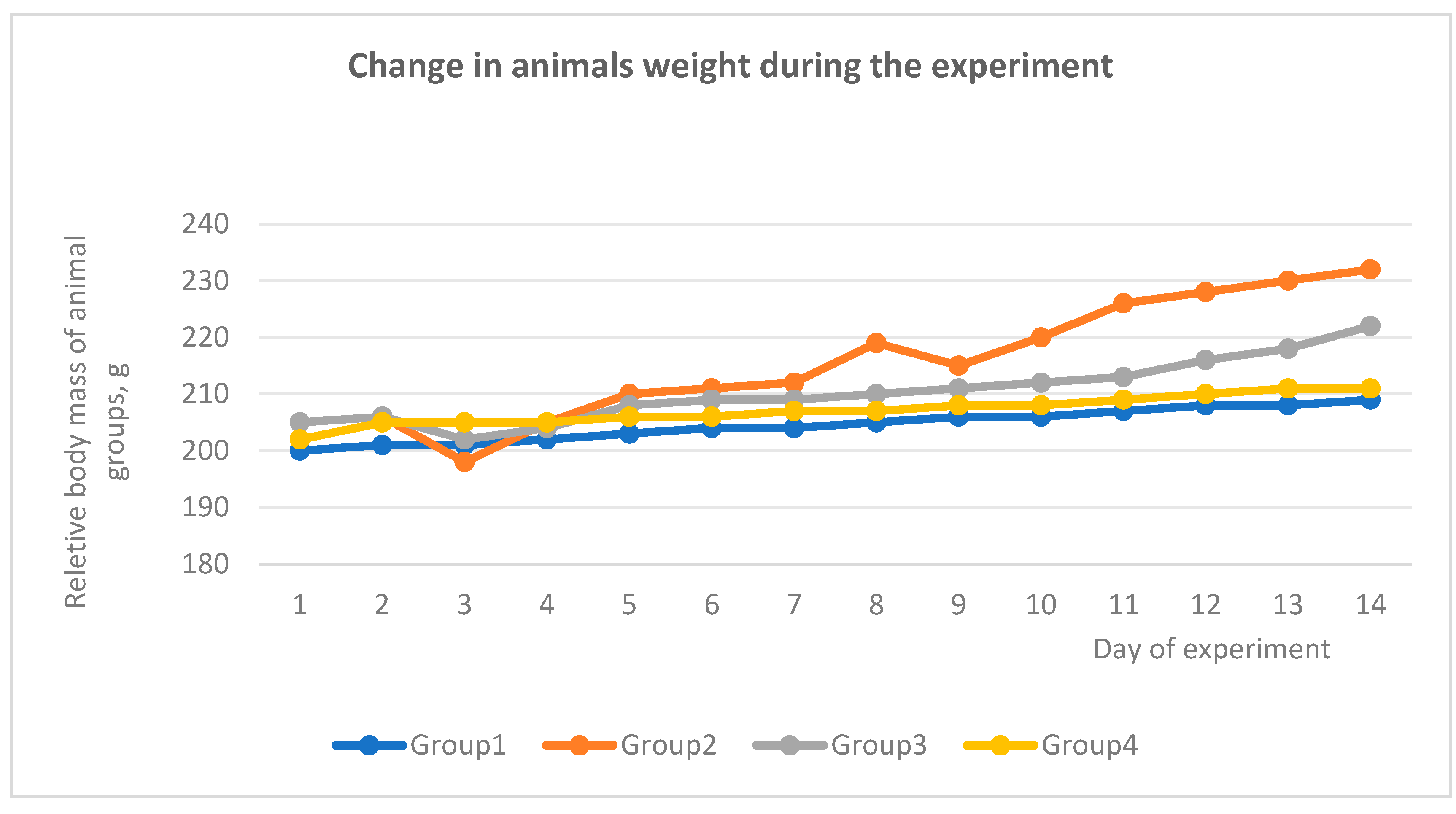
Figure 10.
photo of the liver in normal conditions and with pathology A - intact, B - control group.
Figure 10.
photo of the liver in normal conditions and with pathology A - intact, B - control group.
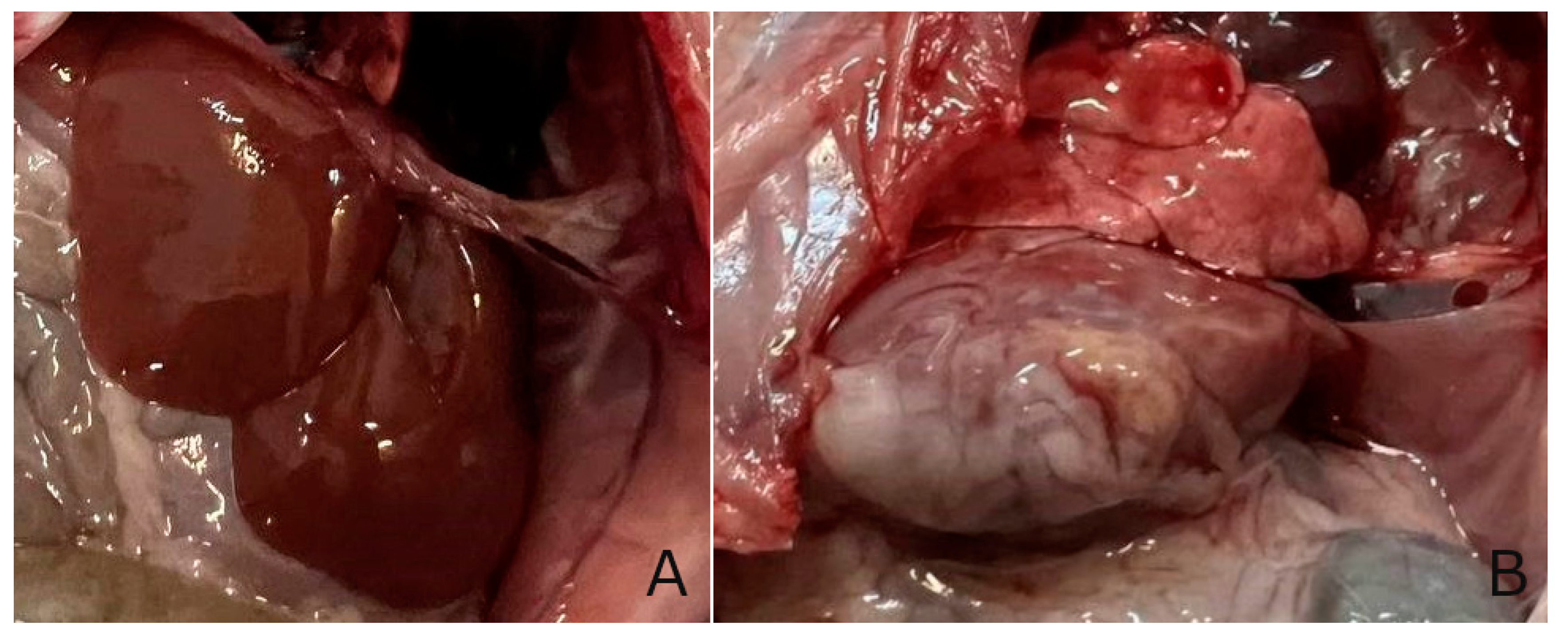

Table 1.
Biochemical parameters of blood serum of animals in the studied groups.
| Indicators | Experimental groups | |||
| Group I (intact) | Group II (control) | Group III (TTZ) | Group IV (DCTP) | |
| АLТ (Units/L) | 42.60±2.80 | 147.90±9.49* | 63.03±10.50*/** | 60.50±6.80*/** |
| АSТ (Units/L) | 78.10±5.30 | 162.70±12.80* | 129.50±11.49*/** | 92.70±9.80*/**/*** |
| De Ritis coefficient | 1.80±0.10* | 1.10±0.03* | 2.12±0.49** | 1.56±0.29*/**/*** |
| Alkaline phosphatase Units/L | 90.50±15.80 | 228.30±44.70* | 101.50±5.45** | 89.40±21.00** |
| Total bilirubin (mmol/L) | 9.60±1.80 | 19.18±1.80* | 10.35±1.05** | 12.90±2.10** |
| Direct bilirubin (mmol/L) | 3.30±0.10 | 5.58±0.78* | 3.56±0.19** | 3.30±0.40** |
| Thymol test (Sh) | 0.90±0.05 | 4.62±0.56* | 1.93±0.46** | 1.90±0.20** |
| Mean values and standard errors are presented, there were 10 surviving rats in the intact group, 7 surviving rats in the control group (tetrachloromethane intoxication), 8 surviving rats in the group receiving TTZ (Thiotriazoline) at a dose of 10 mg/100 g, and 10 surviving rats in the group receiving DCTP at a dose of 6 mg/100 g. *Significant difference from intact (P<0.05). **Significant difference from control (P<0.05). ***Significant difference from TTZ (P<0.05) | ||||
Table 2.
Content of diene conjugates and Schiff bases in the liver in the studied groups of animals: intact animals (intact); animals with hepatitis (control); animals with hepatitis that received Thiotriazoline (TTZ) and animals with hepatitis that received DCTP (DCTP).
Table 2.
Content of diene conjugates and Schiff bases in the liver in the studied groups of animals: intact animals (intact); animals with hepatitis (control); animals with hepatitis that received Thiotriazoline (TTZ) and animals with hepatitis that received DCTP (DCTP).
| Animal group | Diene conjugates (nmol/g) | Schiff bases (nmol/g) |
| Group I (intact) | 25.19±2.38 | 38.80±17.69 |
| Group II (control) | 48.34±1.06* | 116.00±28.33* |
| Group III (TTZ) | 34.22±2.63** | 83.11±14.94** |
| Group IV (DCTP) | 27.48±1.34**/*** | 47.56±10.67**/*** |
| The mean values and standard errors are presented. There were 10 surviving rats in the intact group, 7 surviving rats in the control group (tetrachloromethane intoxication), 8 surviving rats in the group receiving TTZ (Thiotriazoline) at a dose of 10 mg/100 g, and 10 surviving rats in the group receiving DCTP at a dose of 6 mg/100 g. *Significant difference from intact (P<0.05). **Significant difference from control (P<0.05). ***Significant difference between DCTP and TTZ (P<0.05). | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



