Submitted:
24 April 2025
Posted:
24 April 2025
You are already at the latest version
Abstract
Anti-neutrophil cytoplasmic antibodies (ANCA)-associated vasculitis (AAV) is a heterogeneous group of small-vessel vasculitides characterized by the presence of antibodies binding to myeloperoxidase (MPO) and proteinase-3 (PR3), found in neutrophil granules. Apart from being the target of ANCA, neutrophils actively contribute to the vicious cycle of inflammation and vascular damage in AAV. On the other hand, platelets have recently been recognized as essential for thrombosis and as inflammatory effectors that collaborate with neutrophils, reinforcing the generation of reactive oxygen species (ROS) and the formation of neutrophil extracellular traps (NETs) in those diseases. Neutrophils exhibit morphological and functional heterogeneity in AAV, reflecting the complexity of their contribution to disease pathogenesis. Since long-term immunosuppression may be related to serious infections and malignancies, there is an urgent need for reliable biomarkers of disease activity to optimize the management of AAV. This review summarizes the current understanding of the role of neutrophils and platelets in the pathogenesis of AAV, focusing on their crosstalk, and highlights the potential for identifying novel biomarkers relevant for predicting the disease course and its relapses.
Keywords:
ANCA-associated vasculitis (AAV)
; neutrophils
; platelets
; myeloperoxidase (MPO)
; proteinase-3 (PR3)
; neutrophil extracellular traps (NETs)
; extravesicles (EVs)
; biomarkers
1. Introduction
The anti-neutrophil cytoplasmic antibodies (ANCA)-associated vasculitis (AAV) comprises a group of autoimmune diseases characterized by inflammation of small and medium-sized blood vessels, often leading to multi-organ damage. According to the 2012 Revised International Chapel Hill Consensus, AAV encompasses three main subtypes: granulomatosis with polyangiitis (GPA), microscopic polyangiitis (MPA), and eosinophilic granulomatosis with polyangiitis (EGPA)[1], with EGPA being the rarest and most distinct form due to its eosinophilic component. The presence of ANCA is a characteristic feature of AAV, with PR3-ANCA (proteinase-3 ANCA) associated with GPA and MPO-ANCA (myeloperoxidase ANCA) observed in MPA. Unlike GPA and MPA, where ANCA is detected in approximately 90% of cases, only 30–40% of EGPA cases are ANCA-positive, primarily with MPO-ANCA [2,3].
The heterogeneity of AAV presents significant challenges in diagnosis and management, as symptoms and disease severity vary. Although any tissue can be involved, the respiratory tract and kidneys are most frequently affected, with complications such as pulmonary hemorrhage and rapidly progressive glomerulonephritis (RPGN) posing serious risks, including death [3,4,5]. Additionally, some patients exhibit overlapping features of different AAV subtypes, further complicating classification and prognosis [2,6,7]. Finally, the clinical presentation and disease trajectory may also change, making the long-term prognosis and treatment plans complex and challenging [8].
Due to the typical relapse-remission pattern of AAV, remission induction and remission maintenance are fundamental paradigms in management. Remission induction in GPA and MPA depends on the severity of disease presentation. For patients with life- or organ-threatening manifestations, both the 2021 American College of Rheumatology/Vasculitis Foundation (ACR/VF)[9] and 2022 European Alliance of Associations for Rheumatology (EULAR) guidelines [10] recommend a combination of glucocorticoids (GC) and either rituximab (RTX) or cyclophosphamide (CYC). Maintenance of remission relies on intensive and long-term immunosuppression. In GPA and MPA, maintenance treatment with repeated doses of RTX is superior to azathioprine (AZA) and methotrexate (MTX) in reducing relapse rates, while AZA and MTX are recommended when RTX is contraindicated [11,12]. Mycophenolate mofetil (MMF) is considered a third-line treatment [9,10]. However, in some patients, discontinuing GC despite immunosuppressive therapy use may still be unfeasible [10].
Immunosuppressive treatment in AAV patients is associated with various complications, some of which can be severe and potentially fatal, particularly during long-term maintenance therapy. It has been shown that infections and malignancies rank among the top three long-term causes of death in AAV, being surpassed only by cardiovascular diseases, whose risk is dose-dependent on GC use [13,14,15]. Therefore, it is crucial to balance therapy to reduce the risk of disease relapse but also minimize the side effects and toxicity of immunosuppression. Since no clinical parameters are available, reliable laboratory biomarkers are urgently needed to predict clinical trajectory and disease relapse.
Vasculitides are related to the inflammatory process that damages the walls of small and medium-sized blood vessels, resulting in impaired blood flow and ischemia [16,17]. The exact mechanisms underlying immunologic and cell responses, including preliminary triggers, have not been fully understood; however, the central role of neutrophils in the pathogenesis of AAV is unquestionable [18,19,20,21]. Platelets, on the other hand, have recently been considered as potential contributors to the inflammatory process underlying AAV. Moreover, there is growing evidence of crosstalk between neutrophils and platelets in various conditions [22,23,24], including AAV [25,26,27,28].
Given the relatively novel evidence on platelet involvement in AAV pathogenesis, we aim to review that issue. We summarize the data on how platelets burden and modulate the inflammatory response and contribute to vascular damage in AAV. Furthermore, we recapitulate the known facts on the interplay between neutrophils and platelets in AAV and highlight the impact of this novel research on disease prognosis and optimal therapeutic approach in those patients.
2. Materials and Methods
A literature search was performed on PubMed and Embase for studies published in English by April 2025, using MeSH and Emtree terms related to ANCA-associated vasculitis, neutrophils, and platelets. Titles and abstracts were screened for relevance to the review topic. Articles that did not provide new insights were excluded. Additional relevant articles were identified through citation searching. Due to substantially different pathogenesis and rarity, this review did not include data on EGPA.
3. Neutrophils
3.1. Development
Neutrophils, or polymorphonuclear (PMN) leukocytes, are the most abundant type of leukocytes, constituting 50-70% of their total population. The average lifespan of neutrophils is relatively short, limited by programmed cell death, ranging from 7-9 hours [29] to over 5 days [30]. Therefore, to maintain their count, about 1-2 × 1011 neutrophils are produced daily in the bone marrow via neutropoiesis [31], a process dependent on granulocyte colony-stimulating factor (G-CSF) activity [32]. After maturation, neutrophils are retained in bone marrow for the next 4-6 days, forming a reserve pool, ready to be deployed in case of an infection or trauma [33]. Neutrophil retention and release are regulated by chemokine signaling, the expression of which is controlled also by G-CSF [34]. In MPA, high initial levels of circulating G-CSF were associated with the severity of central nervous system involvement and disease activity, according to the Birmingham Vasculitis Activity Scale (BVAS)[35]. After fulfilling a biological function, neutrophils undergo apoptosis in the bone marrow, spleen, or liver. Then, they are cleared by macrophages through efferocytosis [36], which modulates the neutropoiesis by a negative feedback loop. Macrophages release large amounts of interleukin (IL)-23, triggering IL-17 production by T, NK, and NKT cells and further increasing G-CSF production. Interestingly, phagocytosis of neutrophils reduces IL-23 production by macrophages, leading to diminished neutropoiesis and cell release to the peripheral blood. On the other hand, when inflammation is amplified, the same cytokine axis intensifies neutrophil proliferation and maturation [37,38]. Patients with AAV exhibit a markedly elevated circulating neutrophil count [39]. In AAV, according to the study by Nogueira et al. [40], serum IL-17A levels were significantly higher in the acute phase of GPA than in healthy controls. Moreover, in the same study, patients with elevated IL-23 had more active disease and higher ANCA titers. Additionally, a study by Huang et al. documented a positive correlation between the neutrophil-to-lymphocyte ratio (NLR) and C-reactive protein (CRP) and an inverse with C3 level in those with MPO-ANCA [41]. Interestingly, a threshold NLR of at least 5.9 was related to increased disease severity and risk of relapse in follow-up [42]. Furthermore, an elevated delta neutrophil index (DNI) of at least 0.65, reflecting the proportion of immature granulocytes within the total neutrophil count, was associated with more severe disease at diagnosis; thus, it could likely serve as a predictor of relapse in GPA or MPA [43].
3.2. Interactions with Endothelium and Migration
Since damage to the vascular wall in small vessels is a central issue in AAV, the interactions between neutrophils and the endothelium are vital to their pathogenesis. Under physiological conditions, neutrophils do not adhere to resting endothelium. Upon receiving inflammatory stimuli, neutrophils migrate towards the inflammation site [44]. Leukocyte adhesion cascade initiating extravasation involves capture, rolling, activation, firm adhesion, and intraluminal crawling. This process depends on interactions of neutrophil glycoproteins with endothelial E- and P-selectins, whose expression is upregulated in inflammatory loci. Firstly, E- and P-selectin capture circulating neutrophils to the surface of endothelium by interacting with E-selectin ligand-1 (ESL-1) and P-selectin glycoprotein ligand 1 (PSGL1), respectively. The interaction of β2 integrins, namely lymphocyte function-associated antigen-1 (LFA-1, αLβ2, CD11a/CD18) and macrophage-1 antigen (Mac-1, αMβ2, CD11b/CD18) with ligands such as vascular cell adhesion molecule (VCAM-1), intercellular adhesion molecule-1 (ICAM-1, CD54), and ICAM-2 on endothelial cells, ensures firm adhesion of neutrophils [45,46]. The expression of adhesion molecules, along with the deposition of plasma fibronectin on leukocytes, is enhanced by inflammation and endothelial damage. Both ANCA and tumor necrosis factor (TNF) increase β2 integrin expression on neutrophils, further promoting their attachment and neutrophil aggregation, enhancing trapping of cell aggregates in capillaries [47]. Adhesion of neutrophils is followed by crawling along the surface of the endothelium, following the gradient of chemoattractants, adhesion receptors, and endothelial stiffness. Finally, neutrophils travel through the vascular wall via paracellular or, to a lesser extent, transcellular routes in the process of diapedesis, facilitated by the interaction of β2 integrins, ICAM-1, PECAM-1, and junctional adhesion molecules [48]. In this context, the biological role of CD177, a novel IgG Fc receptor found on the neutrophil surface, must also be mentioned. It has been demonstrated that CD177 ligation causes the migratory arrest of neutrophils, likely triggered by β2 integrins as signaling partners, including their higher expression and affinity, reduced internalization, and extracellular signal-regulated kinases (ERK)-driven suppression of chemoattractants stimuli, potentially leading to endothelial injury [49]. Next, studies on murine models and in vitro have shown that some adhesive molecules can increase neutrophil extracellular trap (NETs) formation, a process essential to AAV pathogenesis and tremendously enhancing local inflammatory response. For instance, Mac-1 augments lipopolysaccharides (LPSs)-related NETs formation (NETosis), P-selectin after ligation to PSGL-1, while the β2 integrin subfamily when triggered by β-glucan [47].
Semaphorin A4D (SEMA4D) is a glycoprotein engaged in neutrophil-endothelium interactions, considered a potential biomarker of disease activity. In typical physiological conditions, the membrane complex of CD100/SEMA4D serves as an inhibitory receptor on neutrophils, where it interacts with plexin-B2 (PLXNB2) expressed by endothelial cells. This interaction shields the endothelium from damage, mainly by suppressing neutrophil-driven immune responses by reducing the production of reactive oxygen species (ROS) and NETosis [50]. A disintegrin and metalloproteinase domain-containing protein 17 (ADAM17) is a protease responsible for cleavage membrane-bound proteins, including SEMA4D, converting it to a soluble form [50,51]. A study by Bertram et al. [51] has reported a higher circulating ADAM17 concentration in patients with active PR3-AAV with renal involvement compared to those in remission or with nonvascular renal diseases. Similarly, markedly elevated serum SEMA4D levels have been demonstrated in patients with AAV, depicting a positive association with the BVAS score, and they were higher than in other autoimmune diseases, such as rheumatoid arthritis or systemic lupus erythematosus (SLE). Furthermore, Wang et al. [52] demonstrated that neutrophils from AAV patients exhibited a notable reduction in cell-surface SEMA4D compared to healthy controls.
3.3. Effector Mechanisms
Neutrophils possess an armamentarium of mechanisms aimed at pathogen detection and destruction. Neutralization of ingested particles and pathogens relies on two main cytotoxic mechanisms – ROS generation in the oxidative burst, catalyzed by nicotinamide adenine dinucleotide phosphate (NADPH)[53], and activity of antibacterial proteins, originating from neutrophil granules [54]. Depending on the stage of neutrophil differentiation, the granules can be divided into four main types–azurophil, specific, gelatinase, and secretory [55]. Degranulation through exocytosis also serves as a targeted, defensive mechanism and further enhances inflammatory response, with secretory granules being the most available for exocytosis [56]. Azurophil granules, however, are particularly relevant in the context of autoimmune vasculitis. Among various potent antimicrobial proteins - such as defensins, cathelicidins, cathepsin B, and lysozyme – they also include MPO and PR3 [57,58,59].
The presence of antibodies that bind to PR3 or MPO is a distinguishing feature of AAV compared to other types of vasculitis. Notably, neutrophils in AAV are characterized by an elevated content of PR3 and MPO in the cytoplasm, which, in normal circumstances, is downregulated before cells enter circulation. This phenomenon is mainly related to the epigenetic dysregulation [60]. Furthermore, expression of those proteins on the neutrophil surface is augmented upon stimulation by complement component 5a (C5a) and cytokines released after ANCA binding, followed by neutrophil degranulation, ROS generation, NETosis, and release of extracellular vesicles (EVs)[61]. EVs are an umbrella term for phospholipid bilayer-enclosed vesicles released by eukaryotic cells. Depending on their size, EVs can be divided into exosomes (50-150 nm) and microparticles (100-1,000 nm)[62]. EVs regulate cell-to-cell communication, with different stimuli influencing the composition of their cargo, including proteins, nucleic acids, and lipids [63]. EVs cargo has been profiled in numerous studies investigating potential biomarkers in various conditions, such as cancer [64,65], neurological disorders [66], autoimmune diseases [67,68], and thrombosis [69]. Neutrophil-derived EVs were shown to carry ANCA-antigens, pro-inflammatory miRNA, and oxylipins. Interestingly, studies report a positive correlation between the BVAS and the levels of C3a, C5a, pentraxin 3 (PTX3), and high mobility group box 1 (HMGB1) expressed on MPO-positive microparticles [70,71]. A recent study by Surmiak et al. [72] has reported that EVs from GPA patients, likely originating from neutrophils, encompass higher leukotriene B4 (LTB4) and 5-oxo-eicosatetraenoic acid (5-oxo-ETE) content compared to EVs from healthy controls. This study also shows that primed neutrophils stimulated with LTB4 or 5-oxo-ETE exhibit a concentration-dependent increase in ROS production and dsDNA release. Another study by the same team [73] shows that EVs released by IgG anti-PR3-activated neutrophils induce human umbilical vein endothelial cells (HUVECs) to produce pro-inflammatory cytokines via their miRNA content (most notably miR-223-3p and miR-142-3p). Furthermore, the expression of EVs-derived miR-223-3p and miR-664a-3p has been elevated in active GPA and correlated positively with BVAS, but also with circulating DNA-MPO complexes, suggesting a link to NETosis [74]. In a study by Glémain et al. [75], miR-142-3p and miR-451 from neutrophil-derived EVs have been implicated in promoting endothelial inflammation and damage. Furthermore, increased expression of miR-223, miR-142-3p, and miR-451 has been observed in peripheral blood of those with kidney involvment, suggesting their role in AAV progression.
ROS damage cellular components by causing double-stranded DNA breaks [76], oxidizing proteins, leading to their misfolding and loss of function [77], and inducing lipid peroxidation that compromises membrane integrity [78]. In addition to direct endothelium damage, oxidative burst might play a role in the stimulation of NETosis [79], as its inhibition decreases NETs formation [80]. A study by Hilhorst et al. [81] suggested that renal damage in AAV depends on an imbalance between ROS and antioxidant defenses. Oxidative burst and hypochlorous acid (HOCl) production in neutrophils due to MPO activation at diagnosis have been linked to the presence of active cellular crescents in MPA, whereas higher serum thiol concentrations correlated with fewer cellular crescents and reduced interstitial fibrosis. According to the same study, MPA patients display higher oxidative stress, characterized by increased serum HOCl and advanced oxidation protein products (AOPP) concentration compared to healthy individuals. Interestingly, Sun et al. [82] reported a decrease in AAV-associated organ damage and MPO deposition in the kidneys and lungs of rat models treated with astaxanthin, a potent antioxidant. Therefore, the level of oxidative stress response could reflect disease activity in AAV, making it a compelling direction for further research. Hydroperoxides of amino acids and their residues are generated during oxidative modification driven by ROS, propagating oxidative damage further within proteins [83]. Therefore, real-time monitoring of oxidation product formation could be a future biomarker of AAV activity.
3.4. Heterogeneity
In the past, neutrophils were thought to form a homogenous population of cells possessing uniform properties. However, this view has been challenged by findings enabled by the advent of high-resolution technologies [57]. Despite a short lifespan, neutrophils change in the process of aging. In addition to downregulating CXCR2 and L-selectin, it manifests by upregulating CXCR4, CD11b, and CD49 [84], accompanied by increased capacity to form NETs. These processes can be linked to the activation by pathogen-associated molecular patterns receptors (PAMPs)[85], e.g., by endotoxins, and depend on changes in cortisol level and expression of circadian-clock genes, remaining in relation to circadian rhythm [86]. The abovementioned alterations promote neutrophils’ homing and infiltrating tissues, where they act as sentinels. In the lungs, for instance, neutrophils adhere to the vascular lumen and reside in the interstitial spaces, retained by a mechanism that depends on CXCR4, making them a possible destination for aged neutrophils [87]. Evidence indicates that neutrophils might display differences in phenotype also across tissue [88,89]. Moreover, neutrophils display substantial diversity in morphology and function in pathological contexts of inflammation, infection, and cancer [90]. For instance, the proportion of circulating neutrophils expressing CD177 remains stable within an individual, regardless of age, gender, or activation state. However, their percentage can increase during pregnancy [91], GM-CSF therapy, or in patients with polycythemia vera [92]. Studies have revealed that variability in CD177 expression affects the surface display of PR3 [93]. Only CD177(+) neutrophils express PR3, and an increased proportion of these cells is linked to a higher risk of developing AAV [94] and a higher risk of relapse in GPA [95]. Stimulation of CD177 triggers degranulation and ROS production, especially in neutrophils expressing high levels of CD177 and PR3 [96]. Interestingly, that effect is attenuated by blocking the complement receptor integrin Mac-1 [97]. On the other hand, neutrophils of MPO-ANCA-positive individuals exhibit higher expression of LFA-1 compared to PR3-ANCA-positive ones, with this integrin expression associated with systemic and pulmonary components of the BVAS [98]. Furthermore, in AAV, neutrophils coexpressing CD177, LFA-1, Mac-1, CD80, CD86, CD100/SEMA4D, class II major histocompatibility complex (MHC II), TNF receptor (TNFR)1, and TNFR2 are detected in higher percentages, also in positive correlation with the BVAS [50,98,99,100]. Among neutrophil subsets described in a pathological context, particular attention is given to low-density neutrophils (LDNs), first identified in individuals with SLE [101]. LDNs can be separated from other granulocytes using a discontinuous density gradient and exhibit a density comparable to peripheral blood mononuclear cells (PBMCs). They have been observed in various conditions, including cancer [102], infections [103], and autoimmune disorders, such as rheumatoid arthritis [104] and psoriasis [105]. Studies based on autoimmune conditions suggest that LDGs are characterized by more pronounced proinflammatory properties [106,107], a higher tendency to undergo NETosis [108,109], and better promotion of Th17 cell differentiation compared to other subsets of neutrophils [110]. Interestingly, LDGs consisting of both mature CD10(+) and immature CD10(-) cells have been identified in AAV [111]. The CD10(-) subset displays downregulated CD16 and CD88/C5aR expression, an immature nuclear structure, and diminished responsiveness to stimulation by MPO-ANCA, which undermines their relevance in AAV pathogenesis [111]. On the other hand, LDGs co-expressing CD10, CD16, and CD88/C5aR can be induced by ANCA stimulation and likely play a role in activating the alternative complement pathway [112]. LDGs have been reported to correlate with disease activity in AAV [113]. However, it is important to note that LDNs encompass a broad spectrum of neutrophil phenotypes and have been observed in lower fraction also in healthy individuals [114]. The actual distinctiveness and relevance of neutrophil phenotypes have been questioned and attributed to the limitations of cell isolation methodology, making it one of the most controversial current aspects of neutrophil research [115,116].
3.5. Cell Death
After completing their function, senescent neutrophils are removed from the inflamed tissue to avoid unnecessary perpetuation of the inflammation [57]. Besides homing to undergo apoptosis and efferocytosis, they can migrate in reverse or perish via different death processes, including pyroptosis, ferroptosis, necroptosis, necrosis, and NETosis [117]. In AAV, neutrophils show a decreased rate of spontaneous apoptosis in vitro, resulting in a prolonged lifespan, which suggests a tendency to accumulate at the inflammation site in vivo [118]. Delayed apoptosis observed in neutrophils of GPA patients involves both the intrinsic and extrinsic pathways and is likely driven by elevated levels of circulating factors such as GM-CSF, TNF, and soluble fatty acid synthase (sFas) [119]. On the other hand, the opposite situation occurs in the case of NETosis in AAV. NETosis is a particularly distinct type of cell death – as opposed to apoptosis, it involves the disintegration of neutrophil cellular membrane paired with the release of NETs, which consist of granule contents combined with double-stranded DNA [120]. The formation of NETs inside neutrophils is facilitated by granule elastase and MPO, which help chromatin decondensation by cleaving the histones. Upon release, chromatin immobilizes pathogens, and granule-derived proteins exert their antimicrobial effect, making this process a type of cellular death and an effector neutrophil mechanism [121]. Moreover, it has been observed that, as in the case of unregulated necrosis, NETosis can drive a proinflammatory response instead of mitigating it, contributing to organ injury [122]. In AAV, neutrophils display enhanced NETosis compared to healthy individuals [123,124]. Although NETs formation in AAV is often driven by ANCA, Kraaij et al. [124] suggest it can occur independently of them. Even when IgG and IgA were depleted from AAV patient serum, NETosis still occurred in vitro, indicating the presence of additional humoral factors contributing to neutrophil activation and disease progression. MPO and PR3 are integral components of NETs, so their exposure may further drive ANCA production. In AAV, NETs enriched with PR3 and MPO have been detected in kidney tissues during active disease [125,126]. Interestingly, while both MPO and PR3 can form complexes with DNA, a study by Surmiak et al. [123] shows that MPO-DNA complexes are consistently present in all GPA patients, whereas PR3-DNA complexes can be detected only in some. This suggests that PR3 binding to DNA may be less specific than the complex formation between DNA and MPO or neutrophil elastase, making MPO-DNA complexes a more universal circulating marker of NETs formation. In the same study, patients with active GPA exhibited elevated circulating levels of serine proteases, DNA-histone complexes, and MPO-DNA complexes. However, DNA-histone complexes were similarly elevated in GPA patients in the remission and did not correlate with the BVAS score, making MPO-DNA complexes a more reliable marker of NETosis. Additionally, increased levels of free circulating mitochondrial and genomic DNA have been identified as serum markers of GPA. Elevated circulating nucleosome levels, previously observed in AAV, further support the role of NETosis in disease pathology [127]. Moreover, fibrous DNA deposits containing PR3 and MPO can activate plasmacytoid dendritic cells (pDCs) and autoreactive B cells through a Toll-like receptor 9 (TLR-9)–dependent mechanism [128], potentially amplifying the autoimmune response. Interestingly, NETs formation is not inextricably associated with cell death. In AAV, it can also occur via vital NETosis, contributing to the persistent, pathological immune response [129]. Another type of cell death that leads to NETs formation is necroptosis. Necroptosis is a regulated form of cell death that relies on receptor-interacting serine-threonine kinase 3 (RIPK3) and mixed lineage kinase domain-like (MLKL). It is highly dependent on TNFR1 system, and morphologically resembles necrosis [130]. Schreiber et al. [131] has shown that ANCA-induced NETs formation is also regulated by RIPK1/3 and MLKL pathway. Ferroptosis is a relatively newly recognized type of programmed cell death preceded by iron accumulation and lipid peroxidation [132]. There is a scarcity of studies investigating ferroptosis in vasculitis. However, a recent paper by Rousselle et al. [133] has demonstrated that ANCA-activated neutrophils trigger ferroptosis in endothelial cells in vitro, and endothelial cell ferroptosis plays a role in ANCA-associated glomerulonephritis in a murine AAV model.
4. Platelets
Platelets are small, anucleate cells circulating for about 7–10 days in humans before being cleared in the spleen and liver. Platelet production, or thrombopoiesis, takes place primarily in the bone marrow. This process begins with hematopoietic stem cells (HSCs) differentiating into large, polyploid megakaryocytes, which then extend long, cytoplasmic protrusions known as proplatelets, ultimately demarcating their tips and releasing platelets into circulation [134]. Approximately 1011 platelets are produced daily [135]. While the roles of platelets in hemostasis and thrombosis have been extensively studied, their contributions to the immune response, particularly in autoimmune diseases, have only recently been recognized.
Higher platelet counts are observed in GPA compared to healthy controls; thus, they are suggested as GPA biomarkers. Still, no difference between patients in remission and those with active disease has been observed [123,136]. Proposed mechanisms of count increase comprise interactions between platelet-derived sCD40L and matrix metalloproteinase-9 (MMP-9)[136] and between platelet-derived CCL5 and megakaryocytes [137]. Lower platelet count at diagnosis has been associated with more severe AAV manifestation and impaired renal function. A recent study by Lee et al. [138] has proposed a novel predictor based on platelet, neutrophil, monocyte, and lymphocyte counts - pan-immune-inflammation value (PIIV = neutrophil count (× 1000/m3) × monocyte count (× 1000/ m3) × platelet count (×1000/mm3) / lymphocyte count (× 1000/m3)). A high PIIV (cut-off point of at least 1011.3) at the onset of AAV has been postulated as an independent risk factor for all-cause mortality. Additionally, patients with GPA and MPA—especially those with renal and pulmonary involvement—exhibit significantly elevated peripheral blood platelet count and platelet-to-lymphocyte ratio (PLR). PLR of 272.0 or greater has been linked to more severe disease activity [139,140]. On the other hand, in a study by Jin et al., low platelet counts (cut-off point of 264.5 × 109/L) are suggested to be a predictor of worse renal function, including end-stage renal disease (ESRD) and death [141]. This observation is supported by a study by Álamo et al., where low platelet count was one of the death predictors in AAV, linked also to treatment resistance [142].
Platelets possess granules of three types: 1) alpha granules containing chemokines (CXCL7, CXCL4/PF4, CXCL1/GROα, CXCL5, CCL5/RANTES, CCL3/MIP1α) [143], coagulation factors, PDGF receptors, TGF-β, P-selectin, fibrinogen, von Willebrand factor (vWF), and fibronectin; 2) dense granules storing calcium, magnesium, nucleotides (ADP, ATP), serotonin, histamine; and 3) lysosomal granules which include glycohydrolases and proteases, such as cathepsin, acid phosphatase, collagenase, and elastase. Dense bodies play a role in vasoconstriction and cytokine production primarily due to their high serotonin content [144,145]. In addition, platelets participate in immune responses by producing and releasing proinflammatory cytokines, most notably IL-1β [146]. They interact with specific receptors on leukocytes, such as monocytes, leading to cell activation. Platelets also express a diverse range of immune receptors, including chemokine receptors [147], Fc receptors [148], TLRs [24,149], and complement receptors [150,151,152]. The complement system plays a vital role in the pathogenesis of AAV [153]. Activation of the complement pathway leads to deposition of membrane attack complex (MAC) on the platelet surface and generation of C3a and C5a, further stimulating platelets [26].
In response to vascular damage, platelets undergo immediate changes in shape and function to participate in hemostasis or contribute to thrombosis in an unfavorable scenario. However, as platelets form heterogeneous subpopulations, chronic diseases involve shifts in their composition, altering platelet phenotypes [154]. Changes in platelet phenotypes can be driven by interactions between fully formed platelets and circulating inflammatory molecules or cells and by modulating platelet production by alterations in the megakaryocyte microenvironment. Such observations have been made in several disorders characterized by prolonged systemic inflammation, such as SLE [155], systemic sclerosis [156], and cancer [157]. For instance, chronic inflammatory states are characterized by increased platelet receptor expression and activity and the loss of surface receptors through cleavage and shedding [154]. However, as this is a relatively novel topic, it remains understudied and highlights a new direction for research in AAV monitoring.
5. Neutrophil-Platelet Crosstalk
Recent studies have shown that the progression of AAV relies on the interactions between activated platelets and neutrophils, and the interplay of the coagulation and complement systems (Figure 1).
A study by Miao et al.[27] has investigated in AAV platelet-derived microparticles (PMP) containing proinflammatory cytokines, adhesion molecules, and growth factors, and has showed a positive correlation between PMP levels and erythrocyte sedimentation rate (ESR), CRP concentrations, and the BVAS score, particularly in relation to renal involvement. The PMP level was also positively associated with both platelet and neutrophil counts and the percentage of neutrophils among leukocytes during the active phase of AAV. Upon exposure to cytokines such as TNF, but also C5a, and ANCA, neutrophils undergo an oxidative burst and degranulate, releasing microparticles containing tissue factor, which enhance NETs formation. In turn, NETs not only enhance platelet adhesion [158] and aggregation [120], but also contribute to the activation of the coagulation cascade [159].
Both tissue factor and NETs components initiate the coagulation cascade, ultimately generating thrombin [22]. Thrombin then activates platelets through protease-activated receptors (PARs), leading to platelet degranulation and the release of P-selectin. Platelet receptors facilitate interactions with neutrophils, promoting ROS generation and further NETs formation (Figure 2) [25,28]. These interactions damage the endothelium and trigger vWF release, which mediates platelet adhesion and aggregation, exacerbating vascular injury. Meanwhile, the activated neutrophils and platelets, through their membranes, microparticles, and NETs, stimulate the alternative complement pathway, increasing C5a levels. This positive feedback loop amplifies thrombin production and platelet/endothelial activation, perpetuating the inflammatory processes [160,161]. Activated platelets with bound fibrinogen can stimulate neutrophils, triggering an oxidative burst and facilitating neutrophil extravasation. Elevated plasma levels of platelet-derived soluble CD40L and CD62P/P-selectin complex correlate with disease activity in GPA [162]. The interaction between soluble CD40L and CD40 facilitates communication between platelets and endothelial cells, triggering the production of proinflammatory cytokines, such as IL-1β, IL-2, and TNF. This signaling also upregulates the expression of endothelial adhesion molecules, including ICAM-1, VCAM-1, and P-selectin, promoting the recruitment of leukocytes to the site of vascular damage. The sCD40L increases endothelial tissue factor expression and contributes to endothelial dysfunction by stimulating neutrophil ROS production. sCD40L can directly activate neutrophils, trigger ROS release, and upregulate Mac-1 [163]. It has been suggested that soluble P-selectin can serve as a biomarker of platelet activation [164]. The interaction of neutrophils and platelets mediated by P-selectin and PSGL enables their aggregation [165]. Patients with active GPA have more platelet-neutrophil aggregates and higher CD11b expression on neutrophils compared to healthy individuals. The percentage of platelet-neutrophil aggregates among neutrophils increases with BVAS [123]. The release of P-selectin from platelets stimulates NETs formation; moreover, blocking its interaction with PSGL-1 substantially inhibits platelet-mediated NETosis [166]. In addition, according to a study by Matsumoto et al., platelets activated through TLR9 signaling release CXCL4, which in turn promotes the formation of NETs. Interestingly, the TLR9-CXCL4 axis is upregulated in AAV [28]. Platelets promote the formation of NETs also via activating neutrophils by TLR4 [149]. HMGB1 is a protein derived, i.a., from platelets, capable of activating TLR4-dependant NETosis [167]. Higher serum levels of HMGB1 have also been linked to AAV with renal involvement [168].
Finally, the abovementioned CD177 has been shown not only to recognize β2 integrins but also to bind to PR3 and platelet endothelial cell adhesion molecule 1 (PECAM-1), an immunoglobulin family protein found on endothelial cells, platelets, and neutrophils. However, evidence does not confirm its role in binding platelets to neutrophils or enhancing their interactions [49,169]. Further studies are needed to clarify that issue.
5. Conclusions
Growing evidence shows that all essential neutrophil functions can be altered in AAV and engaged in the disease pathogenesis. Furthermore, platelets play a crucial role in neutrophil activation and NETs formation, ultimately leading to vascular damage. Potential predictors of AAV prognosis and biomarkers of disease activity refer to neutrophils and platelets, ranging from indices calculated based on blood counts to serum cytokine levels and surface antigen expression. Further studies are needed to establish their relevance and implementation in clinical practice. Neutrophil and platelet phenotypes represent a relatively novel and understudied area; therefore, changes in their composition could provide valuable insights into AAV mechanisms and therapeutic strategies.
Author Contributions
Conceptualization, A.D. and K.W.; methodology, A.D. and K.W.; software, M.S..; investigation, A.D. and S.B-S.; writing—original draft preparation, A.D.; writing—review and editing, K.W., M.S., S.B-S, K.W-W., M.K. and J.M.; visualization, M.S.; supervision, K.W., M.S., K.W-W., M.K. and J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jennette, J.C.; Falk, R.J.; Bacon, P.A.; Basu, N.; Cid, M.C.; Ferrario, F.; Flores-Suarez, L.F.; Gross, W.L.; Guillevin, L.; Hagen, E.C.; et al. 2012 Revised International Chapel Hill Consensus Conference Nomenclature of Vasculitides. Arthritis Rheum. 2012, 65, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Drynda, A.; Padjas, A.; Wójcik, K.; Dziedzic, R.; Biedroń, G.; Wawrzycka-Adamczyk, K.; Włudarczyk, A.; Wilańska, J.; Musiał, J.; Zdrojewski, Z.; et al. Clinical Characteristics of EGPA Patients in Comparison to GPA Subgroup with Increased Blood Eosinophilia from POLVAS Registry. J. Immunol. Res. 2024, 2024, 4283928. [Google Scholar] [CrossRef] [PubMed]
- Wójcik, K.; Masiak, A.; Jeleniewicz, R.; Jakuszko, K.; Brzosko, I.; Storoniak, H.; Kur-Zalewska, J.; Wisłowska, M.; Madej, M.; Hawrot-Kawecka, A.; et al. Association of antineutrophil cytoplasmic antibody (ANCA) specificity with the demographic and clinical characteristics of patients with ANCA-associated vasculitides. Pol. Arch. Intern. Med. 2022, 132. [Google Scholar] [CrossRef] [PubMed]
- Biedroń, G.; Włudarczyk, A.; Wawrzycka-Adamczyk, K.; Wójcik, K.; Musiał, J.; Bazan-Socha, S.; Zdrojewski, Z.; Masiak, A.; Czuszyńska, Z.; Majdan, M.; et al. Respiratory involvement in antineutrophil cytoplasmic antibody-associated vasculitides: a retrospective study based on POLVAS registry. Clin. Exp. Rheumatol. 2021, 40, 720–726. [Google Scholar] [CrossRef]
- Koirala A, Sharma PD, Jhaveri KD, Jain K, Geetha D. Rapidly Progressive Glomerulonephritis. Adv Kidney Dis Health. 2024 Nov 1;31(6):485–95.
- Iudici, M.; Puéchal, X.; Pagnoux, C.; Courvoisier, D.S.; Hamidou, M.; Blanchard-Delaunay, C.; Maurier, F.; Ruivard, M.; Quéméneur, T.; Aumaître, O.; et al. Significance of eosinophilia in granulomatosis with polyangiitis: data from the French Vasculitis Study Group Registry. Rheumatology 2021, 61, 1211–1216. [Google Scholar] [CrossRef]
- Quan, M.V.; Frankel, S.K.; Maleki-Fischbach, M.; Tan, L.D. A rare case report of polyangiitis overlap syndrome: granulomatosis with polyangiitis and eosinophilic granulomatosis with polyangiitis. BMC Pulm. Med. 2018, 18, 181. [Google Scholar] [CrossRef]
- Tomasson, G.; Grayson, P.C.; Mahr, A.D.; LaValley, M.; Merkel, P.A. Value of ANCA measurements during remission to predict a relapse of ANCA-associated vasculitis--a meta-analysis. Rheumatology 2011, 51, 100–109. [Google Scholar] [CrossRef]
- Chung SA, Langford CA, Maz M, Abril A, Gorelik M, Guyatt G, et al. 2021 American College of Rheumatology/Vasculitis Foundation Guideline for the Management of Antineutrophil Cytoplasmic Antibody–Associated Vasculitis. Arthritis Rheumatol. 2021 Aug;73(8):1366–83.
- Hellmich, B.; Sanchez-Alamo, B.; Schirmer, J.H.; Berti, A.; Blockmans, D.; Cid, M.C.; Holle, J.U.; Hollinger, N.; Karadag, O.; Kronbichler, A.; et al. EULAR recommendations for the management of ANCA-associated vasculitis: 2022 update. Ann. Rheum. Dis. 2023, 83, 30–47. [Google Scholar] [CrossRef]
- Pagnoux, C.; Mahr, A.; Hamidou, M.A.; Boffa, J.-J.; Ruivard, M.; Ducroix, J.-P.; Kyndt, X.; Lifermann, F.; Papo, T.; Lambert, M.; et al. Azathioprine or Methotrexate Maintenance for ANCA-Associated Vasculitis. New Engl. J. Med. 2008, 359, 2790–2803. [Google Scholar] [CrossRef]
- Guillevin, L.; Pagnoux, C.; Karras, A.; Khouatra, C.; Aumaître, O.; Cohen, P.; Maurier, F.; Decaux, O.; Ninet, J.; Gobert, P.; et al. Rituximab versus Azathioprine for Maintenance in ANCA-Associated Vasculitis. New Engl. J. Med. 2014, 371, 1771–1780. [Google Scholar] [CrossRef]
- Flossmann, O.; Berden, A.; de Groot, K.; Hagen, C.; Harper, L.; Heijl, C.; Höglund, P.; Jayne, D.; Luqmani, R.; Mahr, A.; et al. Long-term patient survival in ANCA-associated vasculitis. Ann. Rheum. Dis. 2011, 70, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Smith, R. Complications of therapy for ANCA-associated vasculitis. Rheumatology 2020, 59, iii74–iii78. [Google Scholar] [CrossRef] [PubMed]
- Sayer, M.; Chapman, G.B.; Thomas, M.; Dhaun, N. Cardiovascular Disease in Anti-neutrophil Cytoplasm Antibody-Associated Vasculitis. Curr. Rheumatol. Rep. 2023, 26, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Woywodt, A.; Streiber, F.; de Groot, K.; Regelsberger, H.; Haller, H.; Haubitz, M. Circulating endothelial cells as markers for ANCA-associated small-vessel vasculitis. Lancet 2003, 361, 206–210. [Google Scholar] [CrossRef]
- Weidner, S.; Hafezi-Rachti, S.; Rupprecht, H.D. Thromboembolic events as a complication of antineutrophil cytoplasmic antibody–associated vasculitis. Arthritis Care Res. 2006, 55, 146–149. [Google Scholar] [CrossRef]
- Falk, R.J.; Jennette, J.C. Anti-Neutrophil Cytoplasmic Autoantibodies with Specificity for Myeloperoxidase in Patients with Systemic Vasculitis and Idiopathic Necrotizing and Crescentic Glomerulonephritis. New Engl. J. Med. 1988, 318, 1651–1657. [Google Scholar] [CrossRef]
- Niles, J.; Mccluskey, R.; Ahmad, M.; Arnaout, M. WEGENERS GRANULOMATOSIS AUTO-ANTIGEN IS A NOVEL NEUTROPHIL SERINE PROTEINASE. 1989, 74, 1888–1893.
- Falk, R.J.; Terrell, R.S.; A Charles, L.; Jennette, J.C. Anti-neutrophil cytoplasmic autoantibodies induce neutrophils to degranulate and produce oxygen radicals in vitro. Proc. Natl. Acad. Sci. 1990, 87, 4115–4119. [Google Scholar] [CrossRef]
- Ewert, B.H.; Jennette, J.C.; Falk, R.J. Anti-myeloperoxidase antibodies stimulate neutrophils to damage human endothelial cells. Kidney Int. 1992, 41, 375–383. [Google Scholar] [CrossRef]
- McDonald, B.; Davis, R.P.; Kim, S.-J.; Tse, M.; Esmon, C.T.; Kolaczkowska, E.; Jenne, C.N. Platelets and neutrophil extracellular traps collaborate to promote intravascular coagulation during sepsis in mice. Blood 2017, 129, 1357–1367. [Google Scholar] [CrossRef]
- Xu, X.R.; Yousef, G.M.; Ni, H. Cancer and platelet crosstalk: opportunities and challenges for aspirin and other antiplatelet agents. Blood 2018, 131, 1777–1789. [Google Scholar] [CrossRef]
- Tay, S.H.; Zharkova, O.; Lee, H.Y.; Toh, M.M.X.; Libau, E.A.; Celhar, T.; Narayanan, S.; Ahl, P.J.; Ong, W.Y.; Joseph, C.; et al. Platelet TLR7 is essential for the formation of platelet–neutrophil complexes and low-density neutrophils in lupus nephritis. Rheumatology 2023, 63, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, N.; Baldini, M.; Ramirez, G.A.; Rovere-Querini, P.; Manfredi, A.A. Platelet-leukocyte deregulated interactions foster sterile inflammation and tissue damage in immune-mediated vessel diseases. Thromb. Res. 2012, 129, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Miao, D.; Li, D.-Y.; Chen, M.; Zhao, M.-H. Platelets are activated in ANCA-associated vasculitis via thrombin-PARs pathway and can activate the alternative complement pathway. Arthritis Res. Ther. 2017, 19, 252–252. [Google Scholar] [CrossRef]
- Miao, D.; Ma, T.-T.; Chen, M.; Zhao, M.-H. Platelets release proinflammatory microparticles in anti-neutrophil cytoplasmic antibody-associated vasculitis. Rheumatology 2019, 58, 1432–1442. [Google Scholar] [CrossRef]
- Matsumoto, K.; Yasuoka, H.; Yoshimoto, K.; Suzuki, K.; Takeuchi, T. Platelet CXCL4 mediates neutrophil extracellular traps formation in ANCA-associated vasculitis. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Saverymuttu, S.H.; Peters, A.M.; Keshavarzian, A.; Reavy, H.J.; Lavender, J.P. The kinetics of 111Indium distribution following injection of 111Indium labelled autologous granulocytes in man. Br. J. Haematol. 1985, 61, 675–685. [Google Scholar] [CrossRef]
- Pillay, J.; Braber, I.D.; Vrisekoop, N.; Kwast, L.M.; de Boer, R.J.; Borghans, J.A.M.; Tesselaar, K.; Koenderman, L. In vivo labeling with 2H2O reveals a human neutrophil lifespan of 5.4 days. Blood 2010, 116, 625–627. [Google Scholar] [CrossRef]
- Dancey, J.T.; A Deubelbeiss, K.; A Harker, L.; A Finch, C. Neutrophil kinetics in man. J. Clin. Investig. 1976, 58, 705–715. [Google Scholar] [CrossRef]
- Richards, M.K.; Liu, F.; Iwasaki, H.; Akashi, K.; Link, D.C. Pivotal role of granulocyte colony-stimulating factor in the development of progenitors in the common myeloid pathway. Blood 2003, 102, 3562–3568. [Google Scholar] [CrossRef]
- Rankin, S.M. The bone marrow: a site of neutrophil clearance. J. Leukoc. Biol. 2010, 88, 241–251. [Google Scholar] [CrossRef]
- Summers, C.; Rankin, S.M.; Condliffe, A.M.; Singh, N.; Peters, A.M.; Chilvers, E.R. Neutrophil kinetics in health and disease. Trends Immunol. 2010, 31, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Ota, S.; Kotani, T.; Matsuda, S.; Nishioka, D.; Masuda, Y.; Unoda, K.; Hosokawa, T.; Ishida, S.; Takeuchi, T. Initial serum GM-CSF levels are associated with the severity of cerebral small vessel disease in microscopic polyangiitis patients. J. Neuroimmunol. 2021, 359, 577671. [Google Scholar] [CrossRef] [PubMed]
- Greenlee-Wacker, M.C. Clearance of apoptotic neutrophils and resolution of inflammation. Immunol. Rev. 2016, 273, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Stark, M.A.; Huo, Y.; Burcin, T.L.; Morris, M.A.; Olson, T.S.; Ley, K. Phagocytosis of Apoptotic Neutrophils Regulates Granulopoiesis via IL-23 and IL-17. Immunity 2005, 22, 285–294. [Google Scholar] [CrossRef]
- Ley K, Smith E, Stark MA. IL-17A-producing neutrophil-regulatory Tn lymphocytes. Immunol Res. 2006 Mar 1;34(3):229–42.
- Matsumoto, K.; Suzuki, K.; Yoshimoto, K.; Seki, N.; Tsujimoto, H.; Chiba, K.; Takeuchi, T. Significant association between clinical characteristics and immuno-phenotypes in patients with ANCA-associated vasculitis. Rheumatology 2019, 59, 545–553. [Google Scholar] [CrossRef]
- Nogueira, E.; Hamour, S.; Sawant, D.; Henderson, S.; Mansfield, N.; Chavele, K.-M.; Pusey, C.D.; Salama, A.D. Serum IL-17 and IL-23 levels and autoantigen-specific Th17 cells are elevated in patients with ANCA-associated vasculitis. Nephrol. Dial. Transplant. 2010, 25, 2209–2217. [Google Scholar] [CrossRef]
- Huang, L.; Shen, C.; Zhong, Y.; Ooi, J.D.; Zhou, Y.-O.; Chen, J.-B.; Wu, T.; Meng, T.; Xiao, Z.; Lin, W.; et al. The association of neutrophil-to-lymphocyte ratio with all-cause mortality in Chinese patients with MPO-ANCA associated vasculitis. Clin. Exp. Med. 2020, 20, 401–408. [Google Scholar] [CrossRef]
- Ahn, S.S.; Jung, S.M.; Song, J.J.; Park, Y.-B.; Lee, S.-W. Neutrophil to lymphocyte ratio at diagnosis can estimate vasculitis activity and poor prognosis in patients with ANCA-associated vasculitis: a retrospective study. BMC Nephrol. 2018, 19, 1–7. [Google Scholar] [CrossRef]
- Yoo, J.; Ahn, S.S.; Jung, S.M.; Song, J.J.; Park, Y.-B.; Lee, S.-W. Delta Neutrophil Index Is Associated with Vasculitis Activity and Risk of Relapse in ANCA-Associated Vasculitis. Yonsei Med J. 2018, 59, 397–405. [Google Scholar] [CrossRef]
- Halbwachs, L.; Lesavre, P. Endothelium-Neutrophil Interactions in ANCA-Associated Diseases. J. Am. Soc. Nephrol. 2012, 23, 1449–1461. [Google Scholar] [CrossRef]
- Bazan-Socha, S.; Bukiej, A.; Marcinkiewicz, C.; Musial, J. Integrins in Pulmonary Inflammatory Diseases. Curr. Pharm. Des. 2005, 11, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Lelliott, P.M.; Nishide, M.; Pavillon, N.; Okita, Y.; Shibahara, T.; Mizuno, Y.; Yoshimura, H.; Obata, S.; Kumanogoh, A.; I Smith, N. Cellular Adhesion Is a Controlling Factor in Neutrophil Extracellular Trap Formation Induced by Anti-Neutrophil Cytoplasmic Antibodies. ImmunoHorizons 2022, 6, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Halai K, Whiteford J, Ma B, Nourshargh S, Woodfin A. ICAM-2 facilitates luminal interactions between neutrophils and endothelial cells in vivo. J Cell Sci. 2014 Feb 1;127(3):620–9.
- Bai, M.; Grieshaber-Bouyer, R.; Wang, J.; Schmider, A.B.; Wilson, Z.S.; Zeng, L.; Halyabar, O.; Godin, M.D.; Nguyen, H.N.; Levescot, A.; et al. CD177 modulates human neutrophil migration through activation-mediated integrin and chemoreceptor regulation. Blood 2017, 130, 2092–2100. [Google Scholar] [CrossRef]
- Nishide, M.; Nojima, S.; Ito, D.; Takamatsu, H.; Koyama, S.; Kang, S.; Kimura, T.; Morimoto, K.; Hosokawa, T.; Hayama, Y.; et al. Semaphorin 4D inhibits neutrophil activation and is involved in the pathogenesis of neutrophil-mediated autoimmune vasculitis. Ann. Rheum. Dis. 2017, 76, 1440–1448. [Google Scholar] [CrossRef]
- Bertram, A.; Lovric, S.; Engel, A.; Beese, M.; Wyss, K.; Hertel, B.; Park, J.-K.; Becker, J.U.; Kegel, J.; Haller, H.; et al. Circulating ADAM17 Level Reflects Disease Activity in Proteinase-3 ANCA-Associated Vasculitis. J. Am. Soc. Nephrol. 2015, 26, 2860–2870. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Song, Y.; Song, D.; Huang, D. The emerging roles of semaphorin4D/CD100 in immunological diseases. Biochem. Soc. Trans. 2020, 48, 2875–2890. [Google Scholar] [CrossRef]
- Li H, Zhou X, Huang Y, Liao B, Cheng L, Ren B. Reactive Oxygen Species in Pathogen Clearance: The Killing Mechanisms, the Adaption Response, and the Side Effects. Front Microbiol [Internet]. 2021 Feb 4;11.
- Levy, O. Antimicrobial proteins and peptides of blood: templates for novel antimicrobial agents. Blood. 2000 Oct 15;96(8):2664–72.
- Pham, C.T.N. Neutrophil serine proteases: specific regulators of inflammation. Nat. Rev. Immunol. 2006, 6, 541–550. [Google Scholar] [CrossRef]
- Faurschou, M.; Borregaard, N. Neutrophil granules and secretory vesicles in inflammation. Microbes Infect. 2003, 5, 1317–1327. [Google Scholar] [CrossRef]
- Zhang, F.; Xia, Y.; Su, J.; Quan, F.; Zhou, H.; Li, Q.; Feng, Q.; Lin, C.; Wang, D.; Jiang, Z. Neutrophil diversity and function in health and disease. Signal Transduct. Target. Ther. 2024, 9, 1–49. [Google Scholar] [CrossRef]
- Cowland, J.B.; Borregaard, N. Granulopoiesis and granules of human neutrophils. Immunol. Rev. 2016, 273, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Nordenfelt, P.; Tapper, H. Phagosome dynamics during phagocytosis by neutrophils. J. Leukoc. Biol. 2011, 90, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Jennette JC, Falk RJ, Gasim AH. Pathogenesis of ANCA Vasculitis. Curr Opin Nephrol Hypertens. 2011 May;20(3):263–70.
- Hong, Y.; Eleftheriou, D.; Hussain, A.A.; Price-Kuehne, F.E.; Savage, C.O.; Jayne, D.; Little, M.A.; Salama, A.D.; Klein, N.J.; Brogan, P.A. Anti-Neutrophil Cytoplasmic Antibodies Stimulate Release of Neutrophil Microparticles. J. Am. Soc. Nephrol. 2012, 23, 49–62. [Google Scholar] [CrossRef] [PubMed]
- van der Pol, E.; Böing, A. N.; Harrison, P.; Sturk, A.; Nieuwland, R. , Classification, Functions, and Clinical Relevance of Extracellular Vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [CrossRef]
- Dalli, J.; Montero-Melendez, T.; Norling, L.V.; Yin, X.; Hinds, C.; Haskard, D.; Mayr, M.; Perretti, M. Heterogeneity in Neutrophil Microparticles Reveals Distinct Proteome and Functional Properties. Mol. Cell. Proteom. 2013, 12, 2205–2219. [Google Scholar] [CrossRef]
- Soung, Y.H.; Ford, S.; Zhang, V.; Chung, J. Exosomes in Cancer Diagnostics. Cancers 2017, 9, 8. [Google Scholar] [CrossRef]
- Sun, J.; Sun, Z.; Gareev, I.; Yan, T.; Chen, X.; Ahmad, A.; Zhang, D.; Zhao, B.; Beylerli, O.; Yang, G.; et al. Exosomal miR-2276-5p in Plasma Is a Potential Diagnostic and Prognostic Biomarker in Glioma. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef]
- Pulliam, L.; Sun, B.; Mustapic, M.; Chawla, S.; Kapogiannis, D. Plasma neuronal exosomes serve as biomarkers of cognitive impairment in HIV infection and Alzheimer’s disease. J. NeuroVirology 2019, 25, 702–709. [Google Scholar] [CrossRef]
- Zheng, C.; Xie, L.; Qin, H.; Liu, X.; Chen, X.; Lv, F.; Wang, L.; Zhu, X.; Xu, J. The Role of Extracellular Vesicles in Systemic Lupus Erythematosus. Front. Cell Dev. Biol. 2022, 10, 835566. [Google Scholar] [CrossRef]
- Ricco C, Eldaboush A, Liu ML, Werth VP. Extracellular Vesicles in the Pathogenesis, Clinical Characterization, and Management of Dermatomyositis: A Narrative Review. Int J Mol Sci. 2024 Feb 6;25(4):1967.
- Gasecka, A.; Böing, A.N.; Filipiak, K.J.; Nieuwland, R. Platelet extracellular vesicles as biomarkers for arterial thrombosis. Platelets 2016, 28, 228–234. [Google Scholar] [CrossRef]
- Antovic, A.; Mobarrez, F.; Manojlovic, M.; Soutari, N.; Baggemar, V.D.P.; Nordin, A.; Bruchfeld, A.; Vojinovic, J.; Gunnarsson, I. Microparticles Expressing Myeloperoxidase and Complement C3a and C5a as Markers of Renal Involvement in Antineutrophil Cytoplasmic Antibody–associated Vasculitis. J. Rheumatol. 2019, 47, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Manojlovic, M.; Juto, A.; Jonasdottir, A.; Colic, J.; Vojinovic, J.; Nordin, A.; Bruchfeld, A.; Gunnarsson, I.; Mobarrez, F.; Antovic, A. Microparticles expressing myeloperoxidase as potential biomarkers in anti-neutrophil cytoplasmic antibody (ANCA)-associated vasculitides (AAV). J. Mol. Med. 2020, 98, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Surmiak, M.; Gielicz, A.; Stojkov, D.; Szatanek, R.; Wawrzycka-Adamczyk, K.; Yousefi, S.; Simon, H.-U.; Sanak, M. LTB4 and 5-oxo-ETE from extracellular vesicles stimulate neutrophils in granulomatosis with polyangiitis. J. Lipid Res. 2020, 61, 1–9. [Google Scholar] [CrossRef]
- Surmiak, M.; Kosałka-Węgiel, J.; Polański, S.; Sanak, M. Endothelial cells response to neutrophil-derived extracellular vesicles miRNAs in anti-PR3 positive vasculitis. Clin. Exp. Immunol. 2021, 204, 267–282. [Google Scholar] [CrossRef]
- Surmiak, M.; Wawrzycka-Adamczyk, K.; Kosałka-Węgiel, J.; Polański, S.; Sanak, M. Profile of circulating extracellular vesicles microRNA correlates with the disease activity in granulomatosis with polyangiitis. Clin. Exp. Immunol. 2022, 208, 103–113. [Google Scholar] [CrossRef]
- Glémain, A.; Néel, M.; Néel, A.; André-Grégoire, G.; Gavard, J.; Martinet, B.; Le Bloas, R.; Riquin, K.; Hamidou, M.; Fakhouri, F.; et al. Neutrophil-derived extracellular vesicles induce endothelial inflammation and damage through the transfer of miRNAs. J. Autoimmun. 2022, 129, 102826. [Google Scholar] [CrossRef]
- Sharma, V.; Collins, L.B.; Chen, T.-H.; Herr, N.; Takeda, S.; Sun, W.; Swenberg, J.A.; Nakamura, J. Oxidative stress at low levels can induce clustered DNA lesions leading to NHEJ mediated mutations. Oncotarget 2016, 7, 25377–25390. [Google Scholar] [CrossRef]
- Sohal, R.S. Role of oxidative stress and protein oxidation in the aging process1,2 1Guest Editor: Earl Stadtman 2This article is part of a series of reviews on “Oxidatively Modified Proteins in Aging and Disease.” The full list of papers may be found on the homepage of the journal. Free. Radic. Biol. Med. 2002, 33, 37–44. [Google Scholar] [CrossRef]
- Mylonas, C.; Kouretas, D. Lipid peroxidation and tissue damage. Vivo Athens Greece 1999, 13, 295–309. [Google Scholar]
- Azzouz, D.; Khan, M.A.; Palaniyar, N. ROS induces NETosis by oxidizing DNA and initiating DNA repair. Cell Death Discov. 2021, 7, 1–10. [Google Scholar] [CrossRef]
- Leung, H.H.L.; Perdomo, J.; Ahmadi, Z.; Yan, F.; McKenzie, S.E.; Chong, B.H. Inhibition of NADPH oxidase blocks NETosis and reduces thrombosis in heparin-induced thrombocytopenia. Blood Adv. 2021, 5, 5439–5451. [Google Scholar] [CrossRef] [PubMed]
- Hilhorst, M.; Maria, A.T.; Kavian, N.; Batteux, F.; Borderie, D.; Le Quellec, A.; van Paassen, P.; Guilpain, P. Impact of MPO-ANCA-mediated oxidative imbalance on renal vasculitis. Am. J. Physiol. Physiol. 2018, 315, F1769–F1776. [Google Scholar] [CrossRef]
- Sun R lan, Shang J chun, Han R hong, Xing G qun. Protective effect of astaxanthin on ANCA-associated vasculitis. Int Immunopharmacol. 2024 May;132:111928.
- Michalski, R.; Zielonka, J.; Gapys, E.; Marcinek, A.; Joseph, J.; Kalyanaraman, B. Real-time Measurements of Amino Acid and Protein Hydroperoxides Using Coumarin Boronic Acid. J. Biol. Chem. 2014, 289, 22536–22553. [Google Scholar] [CrossRef]
- Casanova-Acebes, M.; Pitaval, C.; Weiss, L.A.; Nombela-Arrieta, C.; Chèvre, R.; A-González, N.; Kunisaki, Y.; Zhang, D.; van Rooijen, N.; Silberstein, L.E.; et al. Rhythmic Modulation of the Hematopoietic Niche through Neutrophil Clearance. Cell 2013, 153, 1025–1035. [Google Scholar] [CrossRef]
- Zhang, D.; Chen, G.; Manwani, D.; Mortha, A.; Xu, C.; Faith, J.J.; Burk, R.D.; Kunisaki, Y.; Jang, J.-E.; Scheiermann, C.; et al. Neutrophil ageing is regulated by the microbiome. Nature 2015, 525, 528–532. [Google Scholar] [CrossRef]
- Aroca-Crevillén, A.; Adrover, J.M.; Hidalgo, A. Circadian Features of Neutrophil Biology. Front. Immunol. 2020, 11, 576. [Google Scholar] [CrossRef]
- Devi, S.; Wang, Y.; Chew, W.K.; Lima, R.; A-González, N.; Mattar, C.N.; Chong, S.Z.; Schlitzer, A.; Bakocevic, N.; Chew, S.; et al. Neutrophil mobilization via plerixafor-mediated CXCR4 inhibition arises from lung demargination and blockade of neutrophil homing to the bone marrow. J. Exp. Med. 2013, 210, 2321–2336. [Google Scholar] [CrossRef]
- Watson, F.; Robinson, J.; Phelan, M.; Bucknall, R.; Edwards, S. Receptor expression in synovial fluid neutrophils from patients with rheumatoid arthritis. Ann. Rheum. Dis. 1993, 52, 354–359. [Google Scholar] [CrossRef]
- Fortunati, E.; Kazemier, K.M.; Grutters, J.C.; Koenderman, L.; Bosch, V.J.M.M.V.D. Human neutrophils switch to an activated phenotype after homing to the lung irrespective of inflammatory disease. Clin. Exp. Immunol. 2008, 155, 559–566. [Google Scholar] [CrossRef]
- Sumagin, R. Phenotypic and Functional Diversity of Neutrophils in Gut Inflammation and Cancer. Am. J. Pathol. 2023, 194, 2–12. [Google Scholar] [CrossRef]
- Rudin, A.D.; Torell, A.; Popovic, J.; Stockfelt, M.; Jacobsson, B.; Rudin, A.; Christenson, K.; Lundell, A.-C.; Bylund, J. Pregnancy is associated with a simultaneous but independent increase in circulating CD177pos and immature low-density granulocytes. J. Leukoc. Biol. 2024. [Google Scholar] [CrossRef]
- Göhring K, Wolff J, Doppl W, Schmidt KL, Fenchel K, Pralle H, et al. Neutrophil CD177 (NB1 gp, HNA-2a) expression is increased in severe bacterial infections and polycythaemia vera. Br J Haematol. 2004 Jul;126(2):252–4.
- Abdgawad, M.; Gunnarsson, L.; A Bengtsson, A.; Geborek, P.; Nilsson, L.; Segelmark, M.; Hellmark, T. Elevated neutrophil membrane expression of proteinase 3 is dependent upon CD177 expression. Clin. Exp. Immunol. 2010, 161, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Witko-Sarsat, V.; Lesavre, P.; Lopez, S.; Bessou, G.; Hieblot, C.; Prum, B.; Noël, L.H.; Guillevin, L.; Ravaud, P.; Sermet-Gaudelus, I.; et al. A Large Subset of Neutrophils Expressing Membrane Proteinase 3 Is a Risk Factor for Vasculitis and Rheumatoid Arthritis. J. Am. Soc. Nephrol. 1999, 10, 1224–1233. [Google Scholar] [CrossRef] [PubMed]
- Smargianaki, S.; Elmér, E.; Lilliebladh, S.; Ohlsson, S.; Pettersson, Å.; Hellmark, T.; Johansson, Å.C. Disease Activity and Tendency to Relapse in ANCA-Associated Vasculitis Are Reflected in Neutrophil and Intermediate Monocyte Frequencies. J. Immunol. Res. 2024, 2024, 6648265. [Google Scholar] [CrossRef] [PubMed]
- Marino, S.F.; Jerke, U.; Rolle, S.; Daumke, O.; Kettritz, R. Competitively disrupting the neutrophil-specific receptor–autoantigen CD177:proteinase 3 membrane complex reduces anti-PR3 antibody-induced neutrophil activation. J. Biol. Chem. 2022, 298, 101598. [Google Scholar] [CrossRef]
- Jerke, U.; Rolle, S.; Dittmar, G.; Bayat, B.; Santoso, S.; Sporbert, A.; Luft, F.; Kettritz, R. Complement Receptor Mac-1 Is an Adaptor for NB1 (CD177)-mediated PR3-ANCA Neutrophil Activation. J. Biol. Chem. 2011, 286, 7070–7081. [Google Scholar] [CrossRef]
- Matsumoto, K.; Kurasawa, T.; Yoshimoto, K.; Suzuki, K.; Takeuchi, T. Identification of neutrophil β2-integrin LFA-1 as a potential mechanistic biomarker in ANCA-associated vasculitis via microarray and validation analyses. Arthritis Res. Ther. 2021, 23, 1–12. [Google Scholar] [CrossRef]
- Haller, H.; Eichhorn, J.; Pieper, K.; Göbel, U.; Luft, F.C. Circulating leukocyte integrin expression in Wegener's granulomatosis. J. Am. Soc. Nephrol. 1996, 7, 40–48. [Google Scholar] [CrossRef]
- Hasegawa, M.; Nishii, C.; Ohashi, A.; Tomita, M.; Nakai, S.; Murakami, K.; Nabeshima, K.; Fujita, Y.; Ishii, J.; Hiki, Y.; et al. Expression of Tumor Necrosis Factor Receptors on Granulocytes in Patients with Myeloperoxidase Anti-Neutrophil Cytoplasmic Autoantibody-Associated Vasculitis. Nephron Clin. Pr. 2009, 113, c222–c233. [Google Scholar] [CrossRef]
- Hacbarth, E.; Kajdacsy-Balla, A. Low density neutrophils in patients with systemic lupus erythematosus, rheumatoid arthritis, and acute rheumatic fever. Arthritis Rheum. 1986, 29, 1334–1342. [Google Scholar] [CrossRef]
- Vanhaver, C.; Nana, F.A.; Delhez, N.; Luyckx, M.; Hirsch, T.; Bayard, A.; Houbion, C.; Dauguet, N.; Brochier, A.; van der Bruggen, P.; et al. Immunosuppressive low-density neutrophils in the blood of cancer patients display a mature phenotype. Life Sci. Alliance 2023, 7, e202302332. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Huang, J.; Yang, Y.; Liu, L.; Shao, Y.; Li, L.; Sun, B. Dysfunction of low-density neutrophils in peripheral circulation in patients with sepsis. Sci. Rep. 2022, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wright, H.L.; A Makki, F.; Moots, R.J.; Edwards, S.W. Low-density granulocytes: functionally distinct, immature neutrophils in rheumatoid arthritis with altered properties and defective TNF signalling. J. Leukoc. Biol. 2016, 101, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Qin, H.; Yu, Y.; Li, Y.; Lu, J.; Shi, Y. A Distinct Immature Low-Density Neutrophil Population Characterizes Acute Generalized Pustular Psoriasis. J. Investig. Dermatol. 2022, 142, 2831–2835.e5. [Google Scholar] [CrossRef]
- Liu, Y.; Xia, C.; Chen, J.; Fan, C.; He, J. Elevated circulating pro-inflammatory low-density granulocytes in adult-onset Still’s disease. Rheumatology 2020, 60, 297–303. [Google Scholar] [CrossRef]
- Denny MF, Yalavarthi S, Zhao W, Thacker SG, Anderson M, Sandy AR, et al. A Distinct Subset of Proinflammatory Neutrophils Isolated from Patients with Systemic Lupus Erythematosus Induces Vascular Damage and Synthesizes Type I IFNs. J Immunol. 2010 Mar 15;184(6):3284–97.
- Villanueva, E.; Yalavarthi, S.; Berthier, C.C.; Hodgin, J.B.; Khandpur, R.; Lin, A.M.; Rubin, C.J.; Zhao, W.; Olsen, S.H.; Klinker, M.; et al. Netting Neutrophils Induce Endothelial Damage, Infiltrate Tissues, and Expose Immunostimulatory Molecules in Systemic Lupus Erythematosus. J. Immunol. 2011, 187, 538–552. [Google Scholar] [CrossRef]
- Torres-Ruiz, J.M.; Carrillo-Vázquez, D.A.; Leal-Alanis, A.; Zentella-Dehesa, A.; Tapia-Rodríguez, M.; Maravillas-Montero, J.L.; Nuñez-Álvarez, C.A.; Carazo-Vargas, E.R.; Romero-Hernández, I.; Juárez-Vega, G.; et al. Low-Density Granulocytes and Neutrophil Extracellular Traps as Biomarkers of Disease Activity in Adult Inflammatory Myopathies. Am. J. Clin. Oncol. 2021, 28, e480–e487. [Google Scholar] [CrossRef]
- Wu, H.; Zhen, Y.; Ma, Z.; Li, H.; Yu, J.; Xu, Z.-G.; Wang, X.-Y.; Yi, H.; Yang, Y.-G. Arginase-1–dependent promotion of T H 17 differentiation and disease progression by MDSCs in systemic lupus erythematosus. Sci. Transl. Med. 2016, 8, 331ra40–331ra40. [Google Scholar] [CrossRef]
- Mhaonaigh, A.U.; Coughlan, A.M.; Dwivedi, A.; Hartnett, J.; Cabral, J.; Moran, B.; Brennan, K.; Doyle, S.L.; Hughes, K.; Lucey, R.; et al. Low Density Granulocytes in ANCA Vasculitis Are Heterogenous and Hypo-Responsive to Anti-Myeloperoxidase Antibodies. Front. Immunol. 2019, 10, 2603. [Google Scholar] [CrossRef]
- Yuan, J.; Gou, S.-J.; Huang, J.; Hao, J.; Chen, M.; Zhao, M.-H. C5a and its receptors in human anti-neutrophil cytoplasmic antibody (ANCA)-associated vasculitis. Arthritis Res. Ther. 2012, 14, R140–R140. [Google Scholar] [CrossRef]
- Grayson, P.C.; Carmona-Rivera, C.; Xu, L.; Lim, N.; Gao, Z.; Asare, A.L.; Specks, U.; Stone, J.H.; Seo, P.; Spiera, R.F.; et al. Neutrophil-Related Gene Expression and Low-Density Granulocytes Associated With Disease Activity and Response to Treatment in Antineutrophil Cytoplasmic Antibody–Associated Vasculitis. Arthritis Rheumatol. 2015, 67, 1922–1932. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Camarillo, C.; Alemán, O.R.; Rosales, C. Low-Density Neutrophils in Healthy Individuals Display a Mature Primed Phenotype. Front. Immunol. 2021, 12, 672520. [Google Scholar] [CrossRef] [PubMed]
- Silvestre-Roig, C.; Fridlender, Z.G.; Glogauer, M.; Scapini, P. Neutrophil Diversity in Health and Disease. Trends Immunol. 2019, 40, 565–583. [Google Scholar] [CrossRef] [PubMed]
- Hardisty, G.R.; Llanwarne, F.; Minns, D.; Gillan, J.L.; Davidson, D.J.; Findlay, E.G.; Gray, R.D. High Purity Isolation of Low Density Neutrophils Casts Doubt on Their Exceptionality in Health and Disease. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Pérez-Figueroa, E.; Álvarez-Carrasco, P.; Ortega, E.; Maldonado-Bernal, C. Neutrophils: Many Ways to Die. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Abdgawad, M.; Pettersson, Å.; Gunnarsson, L.; Bengtsson, A.A.; Geborek, P.; Nilsson, L.; Segelmark, M.; Hellmark, T. Decreased Neutrophil Apoptosis in Quiescent ANCA-Associated Systemic Vasculitis. PLOS ONE 2012, 7, e32439. [Google Scholar] [CrossRef]
- Surmiak, M.; Hubalewska-Mazgaj, M.; Wawrzycka-Adamczyk, K.; Musiał, J.; Sanak, M. Delayed neutrophil apoptosis in granulomatosis with polyangiitis: dysregulation of neutrophil gene signature and circulating apoptosis-related proteins. Scand. J. Rheumatol. 2019, 49, 57–67. [Google Scholar] [CrossRef]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2017, 18, 134–147. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Cahilog, Z.; Zhao, H.; Wu, L.; Alam, A.; Eguchi, S.; Weng, H.; Ma, D. The Role of Neutrophil NETosis in Organ Injury: Novel Inflammatory Cell Death Mechanisms. Inflammation 2020, 43, 2021–2032. [Google Scholar] [CrossRef]
- Surmiak, M.; Hubalewska-Mazgaj, M.; Wawrzycka-Adamczyk, K.; Szczeklik, W.; Musiał, J.; Brzozowski, T.; Sanak, M. Neutrophil-related and serum biomarkers in granulomatosis with polyangiitis support extracellular traps mechanism of the disease. . 2016, 34, S98–104. [Google Scholar] [PubMed]
- Kraaij, T.; Kamerling, S.W.; van Dam, L.S.; Bakker, J.A.; Bajema, I.M.; Page, T.; Brunini, F.; Pusey, C.D.; Toes, R.E.; Scherer, H.U.; et al. Excessive neutrophil extracellular trap formation in ANCA-associated vasculitis is independent of ANCA. Kidney Int. 2018, 94, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Kessenbrock, K.; Krumbholz, M.; Schönermarck, U.; Back, W.; Gross, W.L.; Werb, Z.; Gröne, H.-J.; Brinkmann, V.; Jenne, D.E. Netting neutrophils in autoimmune small-vessel vasculitis. Nat. Med. 2009, 15, 623–625. [Google Scholar] [CrossRef]
- Aendekerk, J.P.; Ysermans, R.; Busch, M.H.; Theunissen, R.O.; Bijnens, N.; Potjewijd, J.; Damoiseaux, J.G.; Reutelingsperger, C.P.; van Paassen, P. Assessment of longitudinal serum neutrophil extracellular trap–inducing activity in anti-neutrophil cytoplasmic antibody–associated vasculitis and glomerulonephritis in a prospective cohort using a novel bio-impedance technique. Kidney Int. 2023, 104, 151–162. [Google Scholar] [CrossRef]
- Surmiak, M.P.; Hubalewska-Mazgaj, M.; Wawrzycka-Adamczyk, K.; Szczeklik, W.; Musiał, J.; Sanak, M. Circulating mitochondrial DNA in serum of patients with granulomatosis with polyangiitis. Clin. Exp. Immunol. 2015, 181, 150–155. [Google Scholar] [CrossRef]
- Ford, S.L.; O’sullivan, K.M.; Kitching, A.R.; Holdsworth, S.R.; Gan, P.Y.; Summers, S.A. Toll-like Receptor 9 Induced Dendritic Cell Activation Promotes Anti-Myeloperoxidase Autoimmunity and Glomerulonephritis. Int. J. Mol. Sci. 2023, 24, 1339. [Google Scholar] [CrossRef]
- Yousefi, S.; Mihalache, C.; Kozlowski, E.; Schmid, I.; Simon, H.U. Viable neutrophils release mitochondrial DNA to form neutrophil extracellular traps. Cell Death Differ. 2009, 16, 1438–1444. [Google Scholar] [CrossRef]
- Khoury MK, Gupta K, Franco SR, Liu B. Necroptosis in the Pathophysiology of Disease. Am J Pathol. 2020 Feb;190(2):272–85.
- Schreiber, A.; Rousselle, A.; Becker, J.U.; von Mässenhausen, A.; Linkermann, A.; Kettritz, R. Necroptosis controls NET generation and mediates complement activation, endothelial damage, and autoimmune vasculitis. Proc. Natl. Acad. Sci. 2017, 114, 201708247–E9625. [Google Scholar] [CrossRef]
- Yan H fa, Zou T, Tuo Q zhang, Xu S, Li H, Belaidi AA, et al. Ferroptosis: mechanisms and links with diseases. Signal Transduct Target Ther. 2021 Feb 3;6(1):1–16.
- Rousselle, A.; Lodka, D.; Kling, L.; Kettritz, R.; Schreiber, A. Endothelial Cell Ferroptosis Promotes Renal Damage in ANCA-Induced Glomerulonephritis. J. Am. Soc. Nephrol. 2022, 33, 481–481. [Google Scholar] [CrossRef]
- Carminita, E.; Becker, I.C.; Italiano, J.E. What It Takes To Be a Platelet: Evolving Concepts in Platelet Production. Circ. Res. 2024, 135, 540–549. [Google Scholar] [CrossRef]
- Ghalloussi, D.; Dhenge, A.; Bergmeier, W. New insights into cytoskeletal remodeling during platelet production. J. Thromb. Haemost. 2019, 17, 1430–1439. [Google Scholar] [CrossRef] [PubMed]
- Willeke, P.; Kümpers, P.; Schlüter, B.; Limani, A.; Becker, H.; Schotte, H. Platelet counts as a biomarker in ANCA-associated vasculitis. Scand. J. Rheumatol. 2015, 44, 302–308. [Google Scholar] [CrossRef]
- CCL5 derived from platelets increases megakaryocyte proplatelet formation - PubMed [Internet].
- Lee, L.E.; Ahn, S.S.; Pyo, J.Y.; Song, J.J.; Park, Y.-B.; Lee, S.-W. Pan-immune-inflammation value at diagnosis independently predicts all-cause mortality in patients with antineutrophil cytoplasmic antibody-associated vasculitis. Clin. Exp. Rheumatol. 2021, 39, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Kucuk, H.; Tecer, D.; Goker, B.; Varan, O.; Babaoglu, H.; Guven, S.C.; Ozturk, M.A.; Haznedaroglu, S.; Tufan, A. Platelet/lymphocyte ratio and mean platelet volume in patients with granulomatosis with polyangiitis. Hortic. Bras. 2019, 60, 1–6. [Google Scholar] [CrossRef]
- Park, H.J.; Jung, S.M.; Song, J.J.; Park, Y.-B.; Lee, S.-W. Platelet to lymphocyte ratio is associated with the current activity of ANCA-associated vasculitis at diagnosis: a retrospective monocentric study. Rheumatol. Int. 2018, 38, 1865–1871. [Google Scholar] [CrossRef]
- Jin, Y.; Wang, F.; Tang, J.; Luo, L.; Huang, L.; Zhou, F.; Qi, E.; Hu, X.; Deng, S.; Ge, H.; et al. Low platelet count at diagnosis of anti-neutrophil cytoplasmic antibody-associated vasculitis is correlated with the severity of disease and renal prognosis. Clin. Exp. Med. 2024, 24, 1–10. [Google Scholar] [CrossRef]
- L H, C S, Y Z, Jd O, Yo Z, Jb C, et al. Risk factors for treatment resistance and relapse of Chinese patients with MPO-ANCA-associated vasculitis. Clin Exp Med [Internet]. 2020 May;20(2).
- Flad, H.-D.; Brandt, E. Platelet-derived chemokines: pathophysiology and therapeutic aspects. Cell. Mol. Life Sci. 2010, 67, 2363–2386. [Google Scholar] [CrossRef]
- Sonmez, O.; Sonmez, M. Role of platelets in immune system and inflammation. Porto Biomed. J. 2017, 2, 311–314. [Google Scholar] [CrossRef]
- Heijnen H, van der Sluijs P. Platelet secretory behaviour: as diverse as the granules … or not? J Thromb Haemost. 2015 Dec 1;13(12):2141–51.
- Berger, M.; Maqua, H.; Lysaja, K.; Mause, S.F.; Hindle, M.S.; Naseem, K.; Dahl, E.; Speer, T.; Marx, N.; Schütt, K. Platelets from patients with chronic inflammation have a phenotype of chronic IL-1β release. Res. Pr. Thromb. Haemost. 2023, 8, 102261. [Google Scholar] [CrossRef]
- Clemetson KJ, Clemetson JM, Proudfoot AEI, Power CA, Baggiolini M, Wells TNC. Functional expression of CCR1, CCR3, CCR4, and CXCR4 chemokine receptors on human platelets. Blood. 2000 Dec 15;96(13):4046–54.
- Worth, R.G.; Chien, C.D.; Chien, P.; Reilly, M.P.; McKenzie, S.E.; Schreiber, A.D. Platelet FcγRIIA binds and internalizes IgG-containing complexes. Exp. Hematol. 2006, 34, 1490–1495. [Google Scholar] [CrossRef]
- Clark SR, Ma AC, Tavener SA, McDonald B, Goodarzi Z, Kelly MM, et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat Med. 2007 Apr;13(4):463–9.
- Vik, D.P.; Fearon, D.T. Cellular distribution of complement receptor type 4 (CR4): expression on human platelets. J. Immunol. 1987, 138, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Peerschke, E.I.; Ghebrehiwet, B. Platelet Receptors for the Complement Component C1q: Implications for Hemostasis and Thrombosis. Immunobiology 1998, 199, 239–249. [Google Scholar] [CrossRef] [PubMed]
- del Conde, I.; Crúz, M.A.; Zhang, H.; López, J.A.; Afshar-Kharghan, V. Platelet activation leads to activation and propagation of the complement system. J. Exp. Med. 2005, 201, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Heras Benito, M. Complement in vasculitis associated with anti-neutrophil cytoplasm antibodies with renal involvement: pathogenic, prognostic and therapeutic implications. Med Clin (Barc). 2023 Aug 25;161(4):160–5.
- Aslan, J.E. Platelet Proteomes, Pathways, and Phenotypes as Informants of Vascular Wellness and Disease. Arter. Thromb. Vasc. Biol. 2021, 41, 999–1011. [Google Scholar] [CrossRef]
- Cornwell, M.G.; El Bannoudi, H.; Luttrell-Williams, E.; Engel, A.; Barrett, T.J.; Myndzar, K.; Izmirly, P.; Belmont, H.M.; Clancy, R.; Ruggles, K.V.; et al. Modeling of clinical phenotypes in systemic lupus erythematosus based on the platelet transcriptome and FCGR2a genotype. J. Transl. Med. 2023, 21, 1–17. [Google Scholar] [CrossRef]
- Yasuoka, H.; Sakata, K.; Yoshimoto, K.; Takeuchi, T. Phenotype of Platelets Are Altered and Activated in Circulation of Patients with Systemic Sclerosis. Blood 2018, 132, 3732–3732. [Google Scholar] [CrossRef]
- Holmes, C.E.; Levis, J.E.; Schneider, D.J.; Bambace, N.M.; Sharma, D.; Lal, I.; Wood, M.E.; Muss, H.B. Platelet phenotype changes associated with breast cancer and its treatment. Platelets 2016, 27, 703–711. [Google Scholar] [CrossRef]
- Massberg, S.; Grahl, L.; von Bruehl, M.-L.; Manukyan, D.; Pfeiler, S.; Goosmann, C.; Brinkmann, V.; Lorenz, M.; Bidzhekov, K.; Khandagale, A.B.; et al. Reciprocal coupling of coagulation and innate immunity via neutrophil serine proteases. Nat. Med. 2010, 16, 887–896. [Google Scholar] [CrossRef]
- Brill, A.; Fuchs, T.A.; Savchenko, A.S.; Thomas, G.M.; Martinod, K.; DE Meyer, S.F.; Bhandari, A.A.; Wagner, D.D. Neutrophil extracellular traps promote deep vein thrombosis in mice. J. Thromb. Haemost. 2011, 10, 136–144. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, H.; Wang, C.; Chen, M.; Zhao, M. Promotion of Hypercoagulability in Antineutrophil Cytoplasmic Antibody–Associated Vasculitis by C5a-Induced Tissue Factor–Expressing Microparticles and Neutrophil Extracellular Traps. Arthritis Rheumatol. 2015, 67, 2780–2790. [Google Scholar] [CrossRef]
- de Bont CM, Boelens WC, Pruijn GJM. NETosis, complement, and coagulation: a triangular relationship. Cell Mol Immunol. 2019 Jan;16(1):19–27.
- Tomasson, G.; Lavalley, M.; Tanriverdi, K.; Finkielman, J.D.; Davis, J.C.; Hoffman, G.S.; McCUNE, W.J.; Clair, E.W.S.; Specks, U.; Spiera, R.; et al. Relationship Between Markers of Platelet Activation and Inflammation with Disease Activity in Wegener’s Granulomatosis. J. Rheumatol. 2011, 38, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gorzelanny, C.; Schneider, S.W. Platelets in Skin Autoimmune Diseases. Front. Immunol. 2019, 10, 1453. [Google Scholar] [CrossRef] [PubMed]
- Ferroni, P.; Martini, F.; Riondino, S.; La Farina, F.; Magnapera, A.; Ciatti, F.; Guadagni, F. Soluble P-selectin as a marker of in vivo platelet activation. Clin. Chim. Acta 2009, 399, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Polanowska-Grabowska, R.; Wallace, K.; Field, J.J.; Chen, L.; Marshall, M.A.; Figler, R.; Gear, A.R.; Linden, J. P-Selectin–Mediated Platelet-Neutrophil Aggregate Formation Activates Neutrophils in Mouse and Human Sickle Cell Disease. Arter. Thromb. Vasc. Biol. 2010, 30, 2392–2399. [Google Scholar] [CrossRef]
- Etulain, J.; Martinod, K.; Wong, S.L.; Cifuni, S.M.; Schattner, M.; Wagner, D.D. P-selectin promotes neutrophil extracellular trap formation in mice. Blood 2015, 126, 242–246. [Google Scholar] [CrossRef]
- Tadie, J.-M.; Bae, H.-B.; Jiang, S.; Park, D.W.; Bell, C.P.; Yang, H.; Pittet, J.-F.; Tracey, K.; Thannickal, V.J.; Abraham, E.; et al. HMGB1 promotes neutrophil extracellular trap formation through interactions with Toll-like receptor 4. Am. J. Physiol. Cell. Mol. Physiol. 2013, 304, L342–L349. [Google Scholar] [CrossRef]
- Bruchfeld, A.; Wendt, M.; Bratt, J.; Qureshi, A.R.; Chavan, S.; Tracey, K.J.; Palmblad, K.; Gunnarsson, I. High-Mobility Group Box-1 Protein (HMGB1) Is Increased in Antineutrophilic Cytoplasmatic Antibody (ANCA)-Associated Vasculitis with Renal Manifestations. Mol. Med. 2010, 17, 29–35. [Google Scholar] [CrossRef]
- Pliyev, B.K.; Menshikov, M. Comparative evaluation of the role of the adhesion molecule CD177 in neutrophil interactions with platelets and endothelium. Eur. J. Haematol. 2012, 89, 236–244. [Google Scholar] [CrossRef]
Figure 1.
The vicious cycle of endothelial damage in anti-neutrophil cytoplasmic antibodies-associated vasculitis.
Figure 1.
The vicious cycle of endothelial damage in anti-neutrophil cytoplasmic antibodies-associated vasculitis.
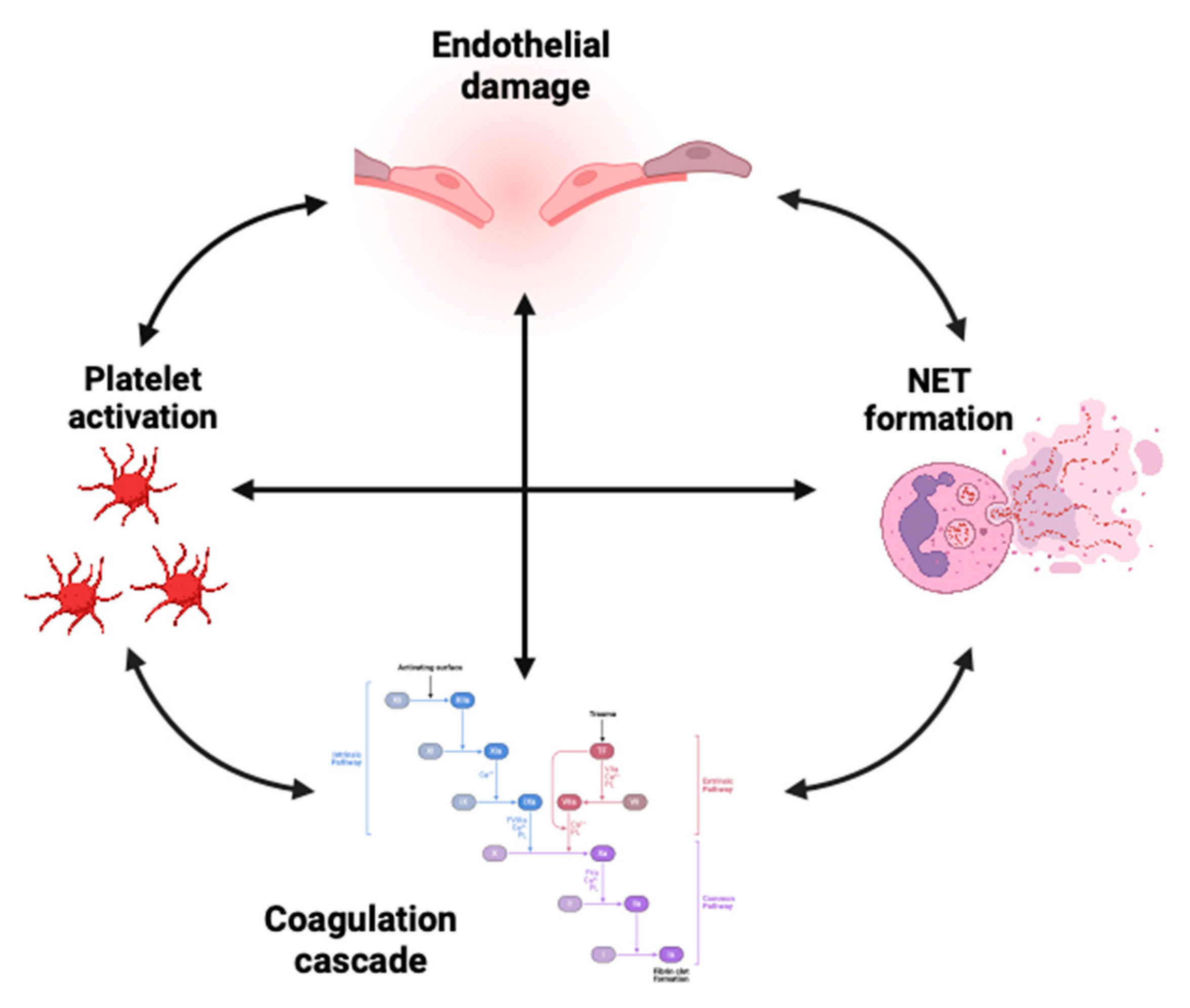
Figure 2.
Platelet-neutrophil interactions in AAV. Abbreviations: BLT1/2 – Leukotriene B4 receptors 1 and 2; GPIIb/IIIa – Glycoprotein IIb/IIIa; HMGB1 – High Mobility Group Box 1; LTB4 – Leukotriene B4; MAC-1 – Macrophage-1 Antigen (CD11b/CD18); PSGL-1 – P-Selectin Glycoprotein Ligand-1; RAGE – Receptor for Advanced Glycation End Products; TLR – Toll-like Receptor.
Figure 2.
Platelet-neutrophil interactions in AAV. Abbreviations: BLT1/2 – Leukotriene B4 receptors 1 and 2; GPIIb/IIIa – Glycoprotein IIb/IIIa; HMGB1 – High Mobility Group Box 1; LTB4 – Leukotriene B4; MAC-1 – Macrophage-1 Antigen (CD11b/CD18); PSGL-1 – P-Selectin Glycoprotein Ligand-1; RAGE – Receptor for Advanced Glycation End Products; TLR – Toll-like Receptor.
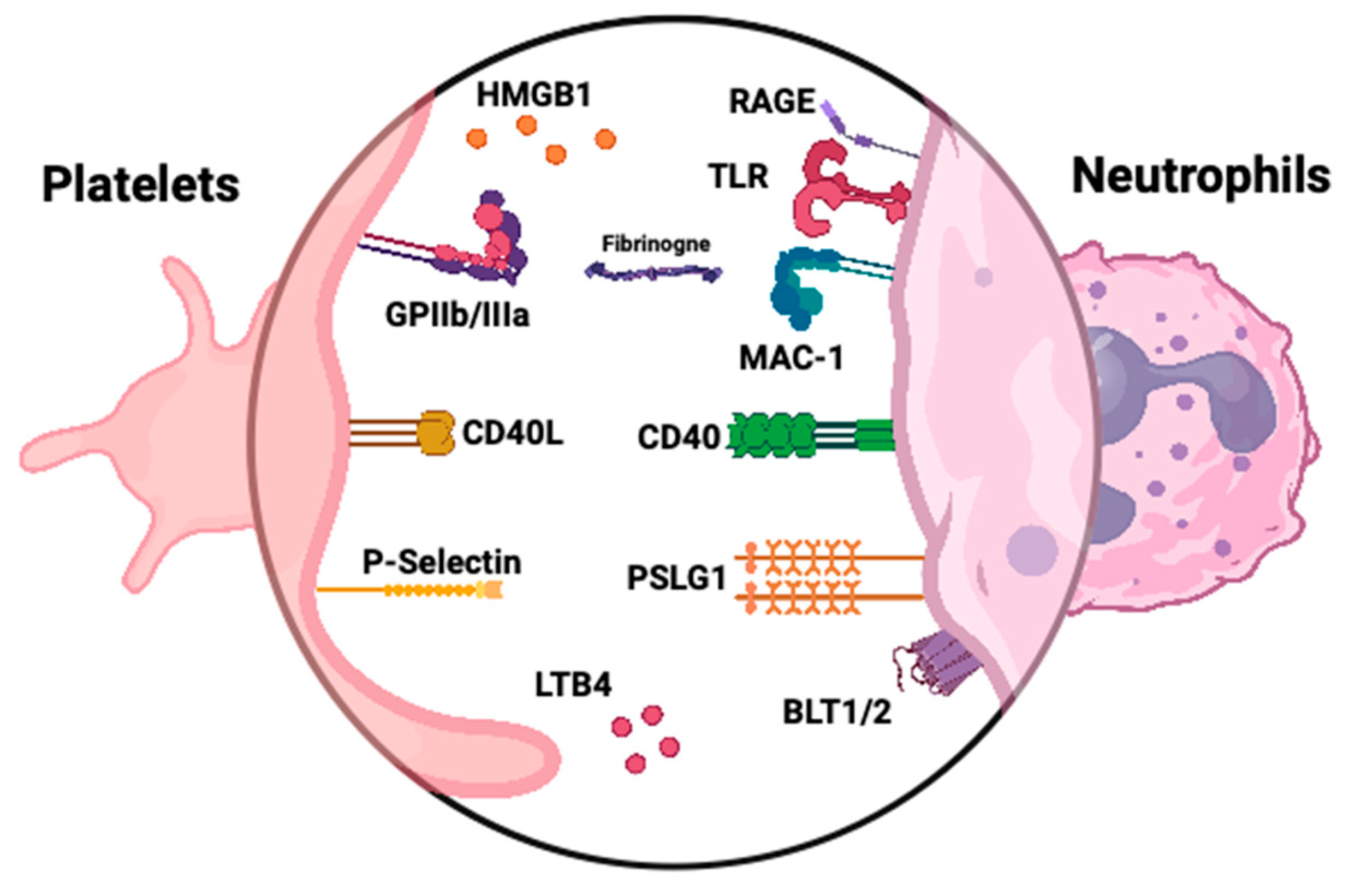
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



