Submitted:
03 April 2025
Posted:
04 April 2025
You are already at the latest version
Abstract
Microorganisms play an essential role in the biogeochemical processes of macroalgal cultivation ecosystems by participating in a complex network of interactions, significantly influencing the growth and development of macroalgae. This study used bibliometric analysis and VOSviewer based on Web of Science data to provide an overview by tracing the developmental footprint of the technology. Countries, institutions, authors, keywords, and key phrases were tracked and mapped accordingly. From 1 January 2003 to 31 December 2023, 619 documents by 2,516 authors from 716 institutions in 51 countries were analyzed. Keyword co-occurrence network analysis revealed five main areas of research on microbes in macroalgal cultivation ecosystems: (1) identification of microbial species and functional genes, (2) biogeochemical cycling of carbon in microbial communities, (3) microbial influences on macroalgae growth and development, (4) bioactivities, and (5) studies based on database. Thematic evolution and map research emphasized the centrality of microbial diversity research in this direction. Over time, the research hotspots and the core scientific questions of the microorganisms in the macroalgal cultivation ecosystems have evolved from single-organism interactions to the complex dynamics of microbial communities. The application of high-throughput techniques had become a hotspot, and the adoption of systems biology approaches had further facilitated the integrated analysis of microbial community composition and function. Our results provide valuable guidance and information for future research.
Keywords:
Microorganisms
; Bibliometric Analysis
; Macroalgal Cultivation Environment
; Functional Genes
; Biogeochemical Cycling of Carbon
1. Introduction
The escalating global interest in sustainable marine resource development has positioned macroalgae cultivation as a hot spot of research within marine aquaculture. This shift is attributed to the considerable economic advantages and environmental benefits associated with macroalgae farming. In 2019, accounting for 30% of the global aquaculture production by wet weight, seaweed aquaculture contributed 5.9% to the total value of global aquaculture, amounting to 34.6 million tons, the figure that surpassed 37.8 million tons by 2022 (Cai et al 2021, FAO 2024). Moreover, seaweed culture has demonstrated its ecological significance by increasing the levels of dissolved oxygen and absorbing inorganic nitrogen and phosphorus in the water column, effectively suppressing harmful algal blooms and showing significant potential for carbon sequestration (Gao et al 2021, Gao et al 2022, Zheng et al 2019, Racine et al 2021, Brown et al 2020).
The microbial community structure specific to the seaweed cultivation area is shaped by the seaweeds, which alter the chemical and physical conditions of their surroundings, including pH, dissolved oxygen, and nutrient salt concentration (Xie et al 2019, Wang et al 2020, Xu et al 2022). The microbial community within the algae cultivation zone establishes a sophisticated interactive network (Xie et al 2024), with its diverse microorganisms fulfilling multifaceted roles across the algal life cycle, thereby exerting a profound influence on the algae’s physiological status and ecological behavior. These communities play a crucial role in influencing overall algal health (Saravanan et al 2024), including promoting morphological development (Marshall et al 2006) and supplying growth factors (Croft et al 2005). They also contribute to nutrient provision (Pei et al 2024; Weigel et al 2022), supporting algae in adapting to environmental changes (Dittami et al 2016), and facilitate the release and settlement of algal spores (Joint et al 2007). Moreover, microbial communities also play an essential role in the ecological effects of seaweed cultivation areas. These communities are involved in the carbon cycle within seaweed farming environments through their metabolic activities, contributing to enhancing marine carbon sinks. Based on prior studies, microbes promoted the conversion of organic carbon from an active state to a more stable sedimentary form through the degradation of algal detritus, thereby storing carbon in marine sediments for long periods, a process termed the microbial carbon pump (MCP) (Jiao et al 2010). In the cultivation ecosystems, Bacteroidetes dominated the degradation of algal polysaccharides (Lu et al 2023, Zheng et al 2021). However, the metabolic capacity of seaweed varied significantly during different growth periods, so seaweed farming may sometimes become a source rather than a sink of CO2 due to the combined effects of macroalgae and microorganisms, even causing adverse environmental impact (Xiong et al 2023). In addition to contributing to carbon sinks, microbial communities play a vital role in the biogeochemical cycles of nitrogen and phosphorus in aquaculture areas (Pfister et al 2019, Pfister et al 2019). Through these cycles, microbial communities help maintain the health of water and sediments (Yang et al 2021, Xie et al 2019). While existing studies have partially characterized the functional roles of microorganisms in macroalgae cultivation systems (e.g., carbon sequestration and nutrient cycling), there is a lack of comprehensive synthesis tracing the transition from early descriptive community studies to contemporary functional genomics approaches. Moreover, existing findings exhibit pronounced fragmentation across disciplinary boundaries—while phycologists have documented microbial influences on algal growth rates, and biogeochemists have quantified nutrient cycling impacts, few studies bridge these perspectives to establish unified ecological frameworks.
Bibliometric analysis is a quantitative research methodology that systematically examines published scientific literature using mathematical and statistical techniques to assess thematic trends within a field of study. This method allows for a review of the field’s historical development and reveals the research trajectories of scholars who have made significant contributions to its advancement (Paul et al 2020). Furthermore, bibliometric analysis can statistically examine the relationship between keywords and literature, which can uncover latent patterns undetectable via manual literature synthesis, offering researchers new research opportunities (Marrone et al 2020). Current bibliometric studies on seaweed aquaculture have focused on various aspects, including the extraction techniques of bioactive compounds such as active peptides (Chukwudulue et al 2023), the antiviral activity of molecules like algal polysaccharides and algal lectins (Pagarete et al 2021, Baltz, 2021), the green synthesis of nanoparticles using algae (Lamilla-Tamayo, 2023), and the potential of seaweed aquaculture as carbon sinks (Zhong et al 2023, Wang et al 2025). Despite extensive summarize in related fields, systematic and in-depth studies on the bibliometrics of microorganisms in the cultivation ecosystems of macroalgae remain inadequate.
As microorganisms fulfill myriad functions within the complex ecosystems of macroalgae cultivation, a substantial body of research has explored their multifaceted roles. Despite this, the domain of microbiology within cultivated macroalgae settings remains relatively uncharted in bibliometric analysis. Consequently, there was a compelling need for an exhaustive bibliometric review that meticulously documented and synthesized the existing microbiological research in such macroalgae cultivation ecosystems. After a comprehensive review of the research status of microorganisms in the cultivation ecosystems of macroalgae, this study aims to identify the central scientific topics in current research through bibliometric analysis. It seeks to explore potentially crucial overlooked areas and outline future research directions based on development trends and research hotspots.
2. Methodology
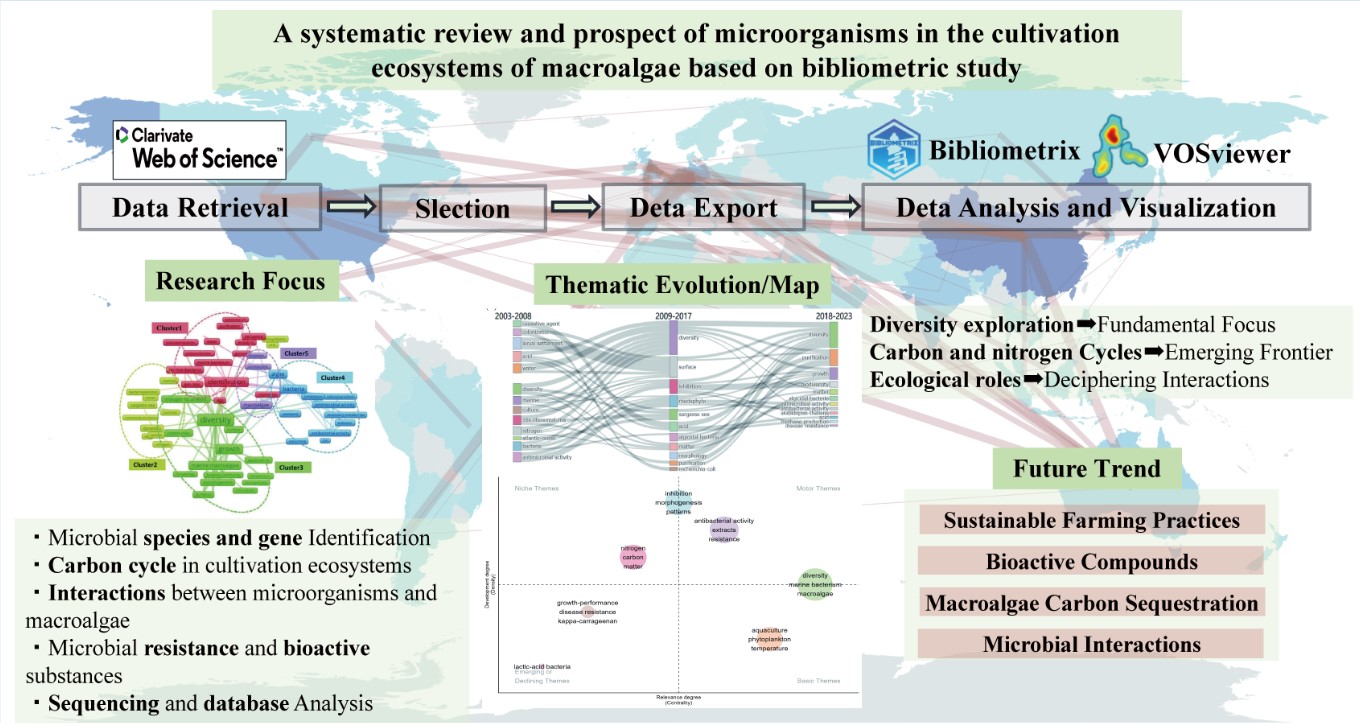
2.1. Study Framework and Data Sources
The research framework outlined all analysis processes and research content (Figure 1). The Web of Science Core Collection (WoSCC) was chosen for its data integrity and had been widely used in environmental bibliometric studies(Ding et al 2022, Li et al 2022). Similarly, WoSCC citation databases were utilized for the bibliometric analysis in this study.
2.2. Search Strategy
The literature collection focused on articles and review papers published from January 1, 2003, to December 31, 2023. We selected articles and reviews only, excluding editorials, conference papers, and book reviews, to avoid overlap in statistical data. The search process involved retrieval, screening, and storage. TS represented the "theme subject" search in the WoS database. By using keywords and Boolean logic, the TS retrieval technique allowed for the efficient and straightforward identification of a large volume of literature related to the topic (Mongeon et al 2016). The search strategy focused on three key terms: "macroalgae", "mariculture", and "microorganisms". Target macroalgae species included Kelp (Laminaria japonica), Gelidium spp., Laver (Pyropia/Porphyra spp.), Pelvetia spp., Sargassum spp., Undaria pinnatifida, Ulva spp., and Gracilaria spp., which collectively account for over 90% of global mariculture production (FAO 2024, Wang et al 2025). Literature was systematically collected for these species, with exclusion criteria applied to eliminate studies related to fermentation engineering or medical applications. The analysis specifically prioritized environmental aspects, including but not limited to bioremediation, nutrient cycling, and ecosystem services associated with macroalgal-microbial interactions.The specific retrieval methodology was described in the supplementary materials. However, the initial data often include duplicate articles or those with low relevance to the study topic. Processing this literature data was crucial for obtaining accurate analysis results. This step is necessary to ensure correct calculation of word frequencies, ultimately yielding reliable and potentially accurate results (Van Eck et al 2006). Therefore, duplicate articles and those with low relevance were removed. Ultimately, we downloaded all relevant records from WoSCC, including authors, titles, sources, abstracts, keywords, addresses, and cited references.
2.3. Data Analysis and Visualization
The dataset was imported into the Bibliometrix tool within the R Studio environment (version 4.3.1) to analyze the publication landscape comprehensively. This analysis included an overview of the publications, a detailed examination of associated keywords, a review of contributing authors, an assessment of the organizations affiliated with cited authors, an identification of the represented countries, and an evaluation of the journals where the work was published. The raw data were then processed using Microsoft Excel 2021 to facilitate further analysis. Finally, cluster analysis was conducted using VOSviewer software (version 1.6.18). A quantitative assessment of the environmental microbiology literature within the cultivation ecosystems of the macroalgae domain was performed. Publication volume data were exported from Bibliometrix and graphically represented using Microsoft Excel. Additional details, including information about the most prolific journals, authors, cited authors, their respective institutions, and countries of origin, were also extracted using Bibliometrix.
In bibliometrics, collaboration analysis typically involves a static examination of interrelated research articles authored by multiple contributors from various institutions. Following Kumar's (Kumar et al 2015) approach to comparative studies, network analysis was employed to explore distinct academic communities and highlight variations in relational models. A co-occurrence analysis of authors and institutions was performed using VOSviewer. For the authors' collaboration analysis, a threshold of 3 co-authors was established, and for institutions' cooperation analysis, a threshold of 5 was set. A geographical heatmap was created to illustrate publication volumes and collaboration between countries, with a minimum edge threshold of 2 for countries' collaborations.
VOSviewer was further utilized to determine the total citation count, average citation rate, volume of literature, and the aggregate number of collaborations for each nation. Keywords are crucial to scholarly articles, providing valuable insight into the core concepts discussed (SobirovIvanovna, 2020). Keyword Plus refers to additional keywords that the Web of Science (WoS) algorithmically clusters to enhance the relevance of the original article, thereby potentially increasing its visibility during searches (Li et al 2022). A bibliometric examination of these keywords was conducted using VOSviewer. Specific synonymous terms, such as "marine”, "ocean”, and "sea" were manually consolidated due to their overlapping meanings. For this study, a keyword frequency threshold of 5 was applied. Exploring keyword frontiers and their evolution requires a temporal dimension to identify research trends and shifts in focus over time (Catone et al 2020). Accordingly, Bibliometrix was used to analyze keyword trends in this study. The temporal analysis was divided into three periods based on key time markers and uniform intervals, guided by the keyword heatmap clustering results. The evolution of keyword topics also required consolidation, following the same methodology used for the keyword co-occurrence map. A minimum clustering frequency of 5 was established for the keyword topic evolution analysis.
3. Results and Discussions
3.1. Quantitative Analysis of the Publications
A systematic review of relevant articles from the past two decades identified 913 articles. After removing duplicates and articles with low relevance to the study topic, 610 documents published between 2003 and 2023 were included in the bibliometric analysis. These documents were authored by 2,516 researchers from 716 institutions across 51 countries (Table S1), comprising 543 original research articles and 67 review articles.
This study utilized a nonlinear fitting approach based on an exponential model to identify a growth trend in the annual research output concerning microorganisms within macroalgae cultivation ecosystems. The analysis yielded a coefficient of determination (R²) of 0.7755, as illustrated in Figure 2. The increase in international collaboration and global research initiatives likely drive this research growth. Notably, since 2015, there has been a more rapid increase in research output compared to the period between 2003 and 2015, following a period of fluctuation. This change may be linked to the 21st United Nations Climate Change Conference held in Paris at the end of 2015 and the subsequent signing of the Paris Agreement. The growing global focus on climate change and marine environmental issues has highlighted macroalgae aquaculture as a potential carbon sink and environmental remediation tool, contributing to reductions in greenhouse gas emissions, ocean acidification, and eutrophication. Consequently, research on the environmental microbiology of macroalgae aquaculture has also surged. Significantly, the number of publications in 2022 increased by 146% compared to the previous year, marking the highest annual growth rate since 2006, with 68 research papers published. In that year, the Food and Agriculture Organization of the United Nations (FAO) published a report (Cai et al 2021), which emphasized the benefits of macroalgae cultivation in global aquaculture development and its role in the food web, environmental benefits, and ecosystem services. As international recognition of the various functions of environmental microorganisms in macroalgae aquaculture grows, we can expect continued expansion in research and publication in this area (Wang et al 2025). We posit that this growth embodies the "Anthropocene microbiology" framework—where microbial processes are increasingly recognized as keystone mediators between anthropogenic pressures (e.g., ocean acidification) and nature-based solutions.
3.2. Collaboration Network Analysis
The number of publications in scientific research is often used as a quantitative indicator to measure a country’s research activity and influence within a specific academic field. A higher publication count generally signifies more robust research capabilities and significant contributions to the field. This study analyzed research articles from 51 countries, with details for the top 20 countries by publication volume provided in Table 2. Figure 3 offers the data and visualization of country publications and collaborations.
China was the leading contributor with 125 articles, followed by the United States with 73 articles, India with 45, Germany with 41, and Japan with 37. The MCP Ratio (Multinational Collaboration Proportion) was defined as the proportion of publications involving international co-authorship relative to the total number of articles within a specified research domain, reflecting cross-border academic collaboration intensity. Germany, for example, had an MCP Ratio of 0.537, indicating that over half of its publications involve international collaboration, reflecting its strong engagement in global research efforts. In contrast, Japan's MCP Ratio of 0.081 suggested lower participation in international cooperation.
Regarding total citations (TC), the United States led with 7,043 citations, underscoring its substantial research impact and academic contribution in this field. Regarding average citations per article, Australia ranked first with an average of 60.1 citations, suggesting high academic quality and influence of Australian research articles. Australia, the United States, and South Korea has shown exceptional performance in average citations per article, indicating that these countries produce high-quality research with considerable influence in the field.
The number of papers published by research institutions are critical indicators of their research scale and capabilities. In this field, 716 different institutions contributed to the publications. Of the top 25 institutions, 11 are from China, and three are from the United States, while Germany, France, and India each have two institutions represented. This distribution highlights China's significant research presence in the microorganisms in the cultivation ecosystems of macroalgae. The Chinese Academy of Sciences led with 62 articles, followed by the French National Centre for Scientific Research (53 articles), Sorbonne University (40 articles), and Ocean University of China (40 articles). Network analysis revealed that the CNRS, Chinese Academy of Sciences, and Sorbonne University had the highest PageRank, indicating their prominent roles in cooperation and influence (Figure 4a, Table S2).
A total of 2,516 authors contributed to this research area. Thomas Wichard was one of the most prolific authors, with 13 publications and the highest citation count. His research focused on the Ulva’s morphogenesis, cell differentiation, and bacterial interactions, utilizing multi-omics and genetic tools to enhance understanding of its ecological and molecular mechanisms. Jie Li from the Yellow Sea Fisheries Research Institute was second with ten articles, including studies on algal diseases and prevention techniques. Florian Weinberger had the highest mediating centrality (128.89) in the author collaboration network, reflecting extensive collaboration with many researchers, including influential scholars like Thomas Wichard and Gaoge Wang (Figure 4b, Table S3).
Regarding journal publications, the studies related to the microorganisms in the cultivation ecosystems of macroalgae were disseminated across 220 different journals. The distribution of periodical publications is illustrated in Figure 5 and Table S4. Among these, the Journal of Applied Phycology led with 41 articles, reflecting its broad focus on algae biology, ecology, and applied technology. The Applied and Environmental Microbiology journal stood out with the highest citation count (3,868) and the highest h-index, indicating its significant impact on the field. The top five journals by h-index were the Journal of Applied Phycology, Frontiers in Microbiology, Aquaculture, and Environmental Microbiology (Table S4). Frontiers in Microbiology, as an interdisciplinary open-access journal, provided a robust platform for the dissemination of cutting-edge research in microbiology. Aquaculture focused on advancements in seaweed farming technology and related microbiological studies. Environmental Microbiology examines the interactions between microorganisms and their environment, offering valuable insights into the role of microorganisms within ecosystems.
3.3. Term Co-Occurrence Network Analysis of Keywords
The Term Co-occurrence Network is a visualization tool used to reveal the knowledge structure and trends within a research field by analyzing and displaying frequently occurring vocabulary (terms, concepts, keywords, etc.) in the literature. Clusters within this network represented groups of often co-occurring terms, indicating their relevance to similar literature or contexts. Each cluster reflected a specific research topic, concept set, or knowledge domain. In the network visualization (Figure 6), node colors represent different clusters, and node sizes reflect the frequency of term occurrence. Terms appearing at least ten times are included and categorized into 5 clusters, each represented by a different color.
Cluster 1: Identification of Microbial Species and Functional Genes
Cluster 1 primarily focused on the identification of microbial species and functional genes within macroalgae culture environments. Key terms in this cluster included "identification", "gene", "nov.", "marine-bacteria", "polysaccharides", "evolution", "Escherichia coli", "microorganisms", "genome", and "colonization".
The methodological evolution in seaweed-associated microbiome research reveals a paradigm shift from cultivation-dependent phenotype characterization to multi-omics functional dissection. In traditional microbiological research, designing specific media for screening and isolating bacteria is crucial, facilitating subsequent physiological and biochemical studies. This approach remained in these days (Jonnadula et al 2009, Pei et al 2023, Wang et al 2023, Wang et al 2008). Sequence-based methods for identifying difficult-to-culture microorganisms, such as the 16S rRNA gene macrogenome analysis, are instrumental in studying microbial diversity. It has limitations in revealing functional roles within microbial communities. Expression analysis of specific functional genes can better predict the roles of these microbes in macroalgae culture environments. Research conducted by Thomas (Thomas et al 2011) utilized RT-qPCR technology to establish standardized methods for analyzing gene expression in Zobellia Galactanivorans. This work underscored the significance of functional gene analysis in elucidating the microbial physiology and metabolism associated with seaweed cultivation. Research conducted by Pei, combining high-throughput 16SrRNA sequencing with environmental factor analysis, found that the composition of Gracilaria lemaneiformis epiphytic bacterial communities correlates positively with nitrate nitrogen and dissolved inorganic nitrogen (DIN), and negatively with nitrite nitrogen. This study also involved screening with urea as the sole nitrogen source to identify urease-producing bacteria based on the ureC gene (Pei et al 2023, Pei et al 2021). The study of functional genes in biogeochemical cycles helps to deeply understand the response mechanisms of microorganisms to environmental changes and assess their role in ecosystem services. Based on the abundance of functional genes, Research conducted by Sun (Sun et al 2023) found through their research on kelp farming areas that macroalgae cultivation activities were able to significantly enhance the expression level of essential functional genes in bacterial communities, thereby strengthening the biogeochemical cycling capacity of coastal ecosystems. This combined approach provides a more comprehensive view of microbial dynamics in macroalgae farming. Integrating high-throughput sequencing technologies, such as metagenomics and transcriptomics, enhances our ability to explore microbial communities’ genetic diversity and functional complexity.
Knowledge of seaweed–bacterial symbioses has recently been applied to manipulate host microbiota with demonstrated benefits to seaweeds at the laboratory scale (Li et al 2023). The identification of keystone functional bacteria has emerged as a critical demand for elucidating bacterial metabolic mechanisms and optimizing aquaculture conditions. In recent years, single-cell transcriptomic techniques have been progressively implemented in microbial molecular ecology research. Their high-resolution gene expression profiling capabilities provide novel perspectives for deciphering microbial community dynamics, functional heterogeneity, and interaction mechanisms with hosts or environments. For microorganisms, the low RNA abundance and spatial resolution of bacteria limit the single-cell transcriptome technology of bacteria. Sarfaties et al. developed a high-throughput spatial transcriptomic methodology (bacterial-MERFISH) that not only delivers community-scale sequencing data but also preserves spatial information (Sarfaties et al 2025). This advancement extends single-cell transcriptomics to bacterial systems, demonstrating that gut bacteria can precisely regulate polysaccharide utilization gene expression at micrometer-scale spatial resolutions, with substantial behavioral heterogeneity observed among individual bacteria. This technology had the potential to migrate to the study of algae epiphytes and endophytes in the future. The mutualistic interactions between macroalgae and their symbiotic microbiota are pivotal for algal growth and disease resistance. However, current multi-omics studies of algal epiphytic bacteria predominantly rely on correlation-based analyses, lacking direct evidence of microenvironmental regulation (Pei et al 2023, Saravanan et al 2024). Furthermore, macroalgae cultivation frequently encounters environmental stressors such as pollution, temperature fluctuations, and salinity variations (Ulrich et al 2022). Single-cell transcriptomics enables the identification of microbial stress-responsive genes (e.g., encoding heat shock proteins and metabolic enzymes), thereby facilitating the evaluation of microbial tolerance mechanisms and their cascading impacts on farmed macroalgae.
Cluster 2: Biogeochemical Cycles of Carbon in Microbial Communities
Cluster 2 focused on understanding the biogeochemical cycles of carbon within microbial communities. Key terms in this cluster included "bacterioplankton", "community structure" and "carbon".
As our understanding of ecosystem functions and global impacts advances, the critical role of microorganisms in geochemical cycles, particularly the carbon cycle, has gained increasing attention. The absorption, transformation, and release of dissolved organic carbon (DOC) in cultivation ecosystems of macroalgae significantly impact the global carbon cycle and its potential as a carbon sink. Evidence suggests that macroalgae play a crucial role in sequestering carbon in the ocean (Duarte et al 2016, Kwan et al 2022). Carbon sequestration pathways include the burial and export of macroalgal organic carbon in sediments, its export to the deep sea, and the production of recalcitrant dissolved organic carbon (RDOC) in macroalgae ecosystems (Ortega et al 2019, Pedersen et al 2020, Chen et al 2020). Microorganisms in aquaculture areas facilitate the conversion of organic carbon from an active state (DOC) to a more stable state (RDOC) through the degradation of algal detritus, thereby storing carbon in marine sediments for extended periods. This process, known as the Microbial Carbon Pump (MCP), is crucial for long-term carbon storage (Jiao et al 2010). As documented in Hughes's publication, macroalgae aquaculture, as part of a Biological Carbon Capture and Storage (BECCS) strategy, could potentially become a carbon-negative fuel source in the future, offering a solution to global climate change (Hughes et al 2012).
Recent paradigm shifts challenge the conventional view of macroalgal systems as unequivocal carbon sinks. Metabolic coupling between algal exudation and microbial respiration may create carbon source dynamics, particularly during senescence phases when reduced photosynthetic activity coincides with enhanced extracellular carbon release (Zhang et al., 2024). Gallagher et al. (2022) further demonstrated that allochthonous carbon inputs and biogeochemical feedbacks can induce CO2 outgassing in certain cultivation scenarios. These findings underscore the context-dependent nature of carbon flux regulation, where environmental parameters (e.g., nutrient availability, light intensity, and microbial community composition) critically modulate DOC/POC release kinetics (Huang et al., 2024). The substantial input of organic matter in sediments from large-scale seaweed aquaculture areas enhances sediment bacterial diversity and promotes both carbon fixation and degradation processes (Huang et al 2023). This organic matter, primarily composed of seaweed detritus and microbial residues, exhibits high microbial availability. However, its significant contribution to carbon sequestration has not yet been incorporated into current carbon sink accounting systems. Simultaneously, microbial communities exhibit non-negligible primary productivity, with global chemolithoautotrophic carbon fixation in coastal sediments estimated at 175 Tg C yr-¹, accounting for approximately 22.7% of total marine carbon fixation (Middelburg 2011). While current evaluations of carbon sequestration in aquaculture ecosystems predominantly focus on macroalgae, the intrinsic carbon fixation capacity and carbon sink potential of environmental microbial communities remain unreported. Future investigations should address how microbial metabolism regulates organic carbon degradation, DOC generation transformation, and particulate organic carbon (POC) interactions, ultimately impacting the dissolved inorganic carbon (DIC) and marine carbonate system. The contribution of microbial communities in aquaculture ecosystems to primary productivity should be systematically incorporated into future carbon sink assessment frameworks.
Additionally, integrated multi-trophic aquaculture (IMTA) is defined as the integration of seaweeds into animal mariculture systems, which has been shown to have considerable potential to enhance the biological carbon sink (Wang et al 2025). For example, a comparative study of different mariculture systems showed that the annual carbon fixation rates were 4,387.5 kg ha−1 y−1 for kelp monoculture, 1,808.3 kg ha−1 y−1 for abalone monoculture, and 12,311.9 kg ha−1 y−1 for integrated multi-trophic aquaculture (IMTA) combining abalone and kelp (Tang et al 2014). This indicates that IMTA systems are significantly more efficient in carbon sequestration compared to monoculture practices. However, the research in the IMTA, specifically the establishment of aquaculture capacity, the dynamic monitoring of carbon in the aquaculture cycle, and the development of aquaculture facilities, was inadequate. Management strategies should be developed to optimize microbial communities and enhance the efficiency and sustainability of seaweed aquaculture systems as carbon sinks.
Cluster 3: Interactions between microorganisms and macroalgae in macroalgal cultivation environments
Cluster 3 focused on various aspects of growth, microbial interactions, and nutrient cycling in macroalgae culture environments. Key terms in this cluster included "growth", "gene nov.", "polysaccharides", "evolution", "bacterial community", "phylogenetic analysis", "surface", "photosynthesis" and "Chlorophyta".
The intricate interplay between macroalgae and their associated microbiomes represents a frontier in marine ecological research, particularly in the context of industrial-scale aquaculture systems. Macroalgae growth and development are not isolated phenomena; they are intricately linked to the complex dynamics of the surrounding microbial communities. In the context of extensive macroalgae cultivation areas, the role of environmental microorganisms emerges as a pivotal factor. Macroalgae exhibit rapid growth but are often limited by nitrogen availability. At the same time, there is substantial research on the effects of nitrogen on macroalgae, which is limited information on microbially mediated nitrogen cycling within these systems. Microorganisms play a crucial role in decomposing organic matter and cycling nutrients, transforming them into forms that macroalgae can absorb. Research conducted by Pei (Pei et al 2023) revealed that urease-producing bacteria (UPB) can hydrolyze urea to provide inorganic nitrogen to macroalgae, thereby promoting their growth. Under the condition of oligotrophication, heterotrophic bacteria (such as Rhodobacteraceae) transform atmospheric nitrogen into bioavailable forms through nitrogenase (nifH gene). The team of Chinese Academy of Sciences found that in the long-term symbiotic system of Synechococcus and heterotrophic bacteria, the nitrogen fixation ability of the flora is the key driving force to maintain the high carbon fixation activity of algae (Nair et al 2022). Metatranscriptomic analyses have revealed that environmental stressors, such as elevated temperature or nitrogen limitation, can activate the nitrate reduction (nar genes) and nitrite oxidation (nxr genes) pathways in the phycosphere microbial community (Lu et al 2023, Abdelrazek et al 2024). This activation forms a “nitrogen compensation” mechanism to alleviate the nitrogen-starved condition of seaweeds. Future research should explore the diversity of nitrogen-cycling microorganisms and their activities under varying environmental stresses, such as temperature changes and nutrient fluctuations, to provide a more holistic view of the nitrogen dynamics in macroalgae cultivation systems. Combined with single cell sequencing and stable isotope probe (SIP) technology, the nitrogen metabolic activity of specific strains can be analyzed in situ.
In addition to nutrient cycling, microorganisms in the cultivation ecosystems of macroalgae can produce bioactive substances, such as growth hormones, antibiotics, and antifungal compounds, to influence the growth of macroalgae. These substances can either directly enhance algae growth or inhibit harmful organisms. For instance, research conducted by Wichard (Wichard, 2015) revealed that large green algae like Ulvales lost their typical morphology under sterile conditions or without an appropriate microbiome. However, specific combinations of Rhodobacteriaceae and Flavobacteriaceae can restore growth in sterile cultures of Ulva. Additionally, Thallusin, a phytohormone-like bacterial compound found in algal culture systems, has been shown to induce morphogenesis in Ulva (Ulrich et al 2022). Yet, the studies did not fully elucidate the mechanisms by which these bacteria restore growth. Further research is needed to identify the specific bioactive compounds and their modes of action, which could lead to the development of probiotic strategies for enhancing macroalgae cultivation.
Microorganisms also form biofilms on the surface of algae, which can impact photosynthesis and gas exchange. Biofilms may impede efficient interactions between algae and their environment or provide protective benefits (Qian et al 2022). The dual role of biofilms in macroalgae health highlights the complexity of their interactions. While some studies have shown that biofilms can enhance algal resistance to environmental stressors, others have reported negative impacts on photosynthetic efficiency. This contradictory evidence suggests that the effects of biofilms are context-dependent and may vary with the composition of the microbial community and environmental conditions. Therefore, a more nuanced understanding of biofilm dynamics is required to optimize macroalgae cultivation practices. Future research should focus on the diversity and functional roles of microorganisms in nutrient cycling, the mechanisms of bioactive substance production, and the complex dynamics of biofilms. Addressing these gaps will not only enhance our fundamental knowledge but also provide practical insights for improving macroalgae cultivation efficiency and sustainability. In future, artificial intelligence (such as convolutional neural network) can be used to predict the relationship between the structure and function of biofilm, or gene editing (such as CRISPRi) can be used to silence the EPS synthesis gene of algal flora, so as to realize the precise regulation of biofilm.
Cluster 4: Microbial Resistance and Bioactive Substances in cultivation ecosystems of macroalgae
Cluster 4 focused on the microbial resistance mechanisms and bioactive substances produced by environmental microorganisms in the cultivation ecosystems of macroalgae. Key terms in this cluster included "bacteria", "antimicrobial activity", "secondary metabolites", "natural-products", "resistance", "extracts" and "inhibition".
As awareness of marine biodiversity and ecosystem services grows, the diverse metabolites produced by microorganisms in macroalgae farming environments are gaining attention. These metabolites can include antimicrobial compounds that protect macroalgae from pathogenic organisms, such as bacteria (Ashen et al 2000, Wang et al 2008), fungi (Lee et al 2015, West et al 2006), and viruses (Kim et al 2016), which may cause various aquatic diseases. In Integrated Multi-Trophic Aquaculture (IMTA) systems, macroalgae can mitigate the impact of harmful algal blooms and pathogenic microorganisms by producing bioactive substances. Research conducted by Goecke et al. (2013) revealed that microorganisms isolated from macroalgae species such as Fucus vesiculosus and Delesseria sanguinea display antibiotic properties effective against pathogens associated with seaweed and surface-associated strains. This discovery underscores the potential of macroalgae in IMTA systems to alleviate the detrimental effects of harmful algal blooms and pathogenic microorganisms through the production of bioactive compounds. The regulatory effects of seaweed aquaculture on the environment, such as nutrient competition, increased pH levels, and the epiphytic microorganisms on seaweeds, also played a part in this process. Research conducted by Sylvers (Sylvers et al 2021) revealed that the incorporation of macroalgae, notably Saccharina latissima and Ulva, into aquaculture systems markedly diminished both the population density of Alexandrium catenella and the toxin burden in mussels. The predominant mechanism of action was allelopathic, with nutritional competition, elevated pH, and macroalgae-associated bacteria contributing subsidiary roles in the mitigation process. Similarly, research conducted by Chai (Chai et al 2021) revealed that Gracilaria lemaneiformis inhibits the growth of Alexandrium sanguinea through chemotaxis and competition for resources. This research added to the growing body of evidence that underscores the multifaceted benefits of the cultivation ecosystems of macroalgae, where they serve as natural biofilters and disease-preventive agents.
Microbial colonization on seaweed surfaces can enhance the host's defense by secreting bioactive compounds with antimicrobial and antifouling properties. These compounds are also valuable in medicine, food, and other applications (Chukwudulue et al 2023). Bacillota, common in macroalgae culture environments, possess large genomes with approximately 9% dedicated to encoding new biosynthetic gene clusters (BGCs), which produce novel bioactive compounds (Baltz, 2016, Baltz, 2021). According to Nadarajan (Nadarajan et al 2021), BGCs within bacterial genomes regulate the synthesis of bioactive compounds like antimicrobial peptides, polyketides, and terpenoids. Identifying and characterizing these BGCs is crucial for developing new therapeutic agents and offers promising avenues for drug discovery from marine symbiotic bacteria. Understanding microbial resistance and the production of bioactive substances in aquaculture environments is essential for maintaining the health and productivity of these systems. However, Most of the predicted BGC products were unclassifed according to the research of Lu (Lu et al 2023), which refects our limited knowledge on secondary metabolites and substantiates that phycosphere bacteria represent a rich resource of as yet unexplored biosynthetic functions.
The study of microbial defense mechanisms in macroalgal aquaculture systems is transitioning from reductionist paradigms centered on single-molecule characterization to holistic frameworks integrating microbial consortia dynamics and cross-kingdom communication networks. This paradigm shift demands synergistic application of advanced methodologies such as in situ spatiotemporal metabolomics, artificial intelligence-driven ecological simulations, and multi-omics correlation analyses to unravel the tripartite interactions among macroalgae, their epiphytic microbiomes, and abiotic stressors. A seminal study by Carrell et al. (2022) exemplifies this approach, demonstrating that Sphagnum-Nostoc symbiosis suppresses host defense pathways (e.g., cysteine-rich peptides and glutathione S-transferases) while engaging in sulfur-mediated metabolic reciprocity. Through matrix-assisted laser desorption/ionization mass spectrometry imaging (MALDI-MSI), they identified Sphagnum-secreted choline-O-sulfate and taurine as bifunctional metabolites: serving both as sulfur donors for nitrogenase complex biosynthesis and osmoregulators enhancing cyanobacterial resilience under acidic conditions. This work establishes a conceptual framework for understanding how environmental gradients shape host-microbe metabolic bargaining and immunological accommodation. Nevertheless, translating these insights into mariculture applications faces persistent challenges. The dynamic nature of coastal ecosystems—marked by diurnal fluctuations in irradiance, tidal hydrodynamics, and thermal regimes—drives rapid compositional shifts in epiphytic microbial communities, complicating the identification of stable interaction signatures. Furthermore, technical limitations in in situ microbial sampling (e.g., biofilm adhesion strength variability) and RNA degradation risks under field conditions hinder high-resolution functional profiling.
Cluster 5: High-Throughput Sequencing and Database Utilization in microorganisms in the cultivation ecosystems of macroalgae
Cluster 5 focused on employing high-throughput sequencing technologies to construct and utilize databases to explore microorganisms’ genetic diversity and functional potential in macroalgae cultivation ecosystems. As depicted in Figure 6b, this area of research has become particularly prominent in recent years. Key terms in Cluster 5 included "macroalgae", "database" and "communities".
The advent of high-throughput sequencing (HTS) has revolutionized our capacity to decode microbial "dark matter" in macroalgae cultivation ecosystems, offering unprecedented resolution to investigate unculturable microorganisms and their functional interplay with algal hosts. As illustrated in Figure 6b, this field has experienced exponential growth since 2020, driven by synergistic advancements in sequencing technologies, computational biology, and multi-omics integration. However, the translation of sequencing-derived insights into actionable ecological or biotechnological outcomes remains hindered by methodological disparities and environmental complexity. This section critically evaluates recent breakthroughs, identifies persistent challenges, and proposes integrative frameworks to advance the field.
Traditional microbial culture techniques have significantly contributed to microbiology research. However, many microorganisms in natural environments remain unculturable, often called "dark matter" in the microbial world (Jiao et al 2021). Advances in high-throughput sequencing technologies now enable researchers to extract and analyze microbial DNA directly from environmental samples, including those from non-culturable microorganisms, thereby studying the genetic diversity and functional potential of microbial communities.
While traditional 16S rRNA amplicon sequencing remains foundational for profiling microbial communities (e.g., Lu et al 2023), its limited functional resolution has spurred innovation in linking taxonomy to metabolic potential. The development of RiboFR Seq (Zhang et al., 2016) and epicPCR (Spencer et al 2016) represents a paradigm shift, enabling physical linkage of ribosomal markers with adjacent protein-coding regions. These techniques have proven particularly valuable in macroalgae-associated systems, where host-specific microbes often encode specialized carbohydrate-active enzymes (CAZymes) for polysaccharide degradation. For instance, Reisky et al. (2024) recently identified a 12-gene ulvan utilization locus in Formosa agariphila through dbCAN2 and Pfam database mining, demonstrating how HTS-driven functional annotation can unravel microbial strategies for algal biomass conversion. Despite advances in platforms like dbCAN2, existing databases lack comprehensive annotations for algal-specific metabolic pathways, particularly for understudied red and brown macroalgae polysaccharide. Meanwhile, current methods still face limitations in resolving rare taxa or low-abundance functional genes, particularly in complex aquaculture environments with dynamic microbial turnover.
Systems biology approaches, including microbial metagenomics, metabolomics, and algal transcriptomics, have provided new insights into microorganism interactions with macroalgae. For instance, KleinJan (KleinJan et al 2022) explored how microbial communities might influence algal stress responses through compound production or quorum sensing effects. Additionally, Reisky (Reisky et al 2024) characterized the ulvan polysaccharide utilization locus (PUL) in Formosa agariphila using HMMs from dbCAN2 and Pfam, elucidating the metabolic pathway of ulvan degradation by marine microbes in algal farming environments. This groundbreaking work led to identifying and characterization of 12 carbohydrate-active enzymes within the ulvan PUL, shedding light on the biochemical strategies that marine microbes employ to harness algal-derived resources. However, the integration of multi-omics approaches, such as microbial metagenomics, metabolomics, and algal transcriptomics, while providing new insights, also introduces complexities in data interpretation. The mechanistic details of these interactions are often inferred from correlative data, and direct causal relationships are difficult to establish. The complexity of environmental microbiota and the dynamic nature of these ecosystems pose significant challenges for the consistent and reliable application of these techniques. For example, the variability in environmental conditions can lead to fluctuations in microbial community structure and function, which may not be captured by short-term or single-point sampling strategies. Therefore, a more integrated and holistic approach is needed, which includes the development of new methods combining species and functions to better capture the diversity of unculturable microorganisms, the use of advanced computational methods to integrate multi-omics data, and the implementation of long-term monitoring strategies to capture the dynamic changes in microbial communities. Emerging single-cell sequencing technologies, such as microfluidic-based platforms (Zheng et al 2022), are redefining our approach to microbial heterogeneity. By circumventing PCR amplification biases and enabling genome assembly from uncultured cells, these methods provide direct access to secondary metabolite biosynthesis gene clusters (BGCs) and host-microbe signaling pathways. For example, Hevroni et al. (2022) applied single-cell RNA sequencing, to profile virus and host transcriptomes of 12,000 single algal cells from a coccolithophore bloom. Their findings stressed the importance of studying host-virus dynamics in natural populations, linking single-cell infection state to host physiology revealed. However, technical challenges persist, including low DNA recovery from miniaturized reactions and difficulties in scaling these approaches for large-scale environmental studies.
3.4. Thematic Evolution
The heat maps in Figure 7 illustrate research trends based on Keyword Plus and author keywords. Analyzing author keywords helps identify research hotspots and core scientific issues within specific periods while examining extended keywords reveals interdisciplinary research trends and potential application areas. This analysis uncovered the evolution of research topics in this field.
From Keyword Plus data, terms such as "diversity", "marine bacteria", "macroalgae" and "growth" were frequently observed. Author keywords like "algae", "aquaculture" and "extracts" also appeared often. This trend indicated a strong focus on the impact of microorganisms on macroalgae growth and the study of bioactive substances. Terms such as "phylogenetic analysis" and "sequence" were prominent in both figures, highlighting the application of molecular biological methods to microbial taxonomy and genetic characteristics in aquaculture. The recurring term "diversity" featured prominently in earlier studies and held high centrality in the keywords co-occurrence network (Figure 6a), signifying that research on microbial diversity in macroalgae culture areas was foundational and intersected with various research directions.
Early studies prioritized elucidating microbial diversity under environmental stressors, driven by terms like "growth," "aquaculture," and "extracts.". Research during this period established baseline correlations between microbial communities and abiotic factors, such as seasonal nutrient fluctuations and anthropogenic impacts like oyster farming (Pinhassi et al 2006, Dubois et al 2007). For instance, Matsuo et al. (2003) identified the Cytophaga-Flavobacterium-Bacteroides (CFB) complex as critical for Monostroma oxyspermum morphogenesis, suggesting host-specific microbial recruitment mechanisms. Similarly, Lu et al. (2008) demonstrated Ulva clathrata's dual role in nutrient remediation and pathogen suppression, hinting at algal-microbe chemical signaling—a concept later expanded through metabolomics. While foundational, these studies were constrained by culture-dependent methods, overlooking unculturable microbial "dark matter".
The post-2008 era witnessed a paradigm shift toward molecular techniques, marked by keywords like "phylogenetic analysis," "sequence," and "biodegradation." The adoption of 16S rRNA sequencing and metagenomics enabled taxonomic profiling of epiphytic bacteria, revealing genera such as Formosa and Pseudoalteromonas as keystone degraders of algal polysaccharides (Albakosh et al 2016). Concurrently, terms like "antifouling" and "allelopathy" underscored microbial secondary metabolites' ecological roles, exemplified by Penesyan et al.'s work on bacterial antifouling agents (Penesyan et al 2009, Penesyan et al 2010, Penesyan et al 2011).
In the past five years (2018-2023), the field has advanced by adopting systems biology methods to analyze microbial communities comprehensively. The appearance of terms like "high-throughput sequencing", "microbial community" and "microbiome" in the author’s keywords reflected a growing interest in understanding the composition and function of microbial communities. This trend represented a methodological revolution and highlighted the need to grasp microbial community dynamics and ecological roles. Recent trends emphasize systems-level integration, with keywords like "high-throughput sequencing," "microbiome," and "holobiont" dominating. The rise of multi-omics (metagenomics, metabolomics, algal transcriptomics) has unraveled complex interaction networks. Additionally, keywords such as "holobiont", "bioactive compounds" and "biodegradation" in Keyword Plus indicated ongoing research into the overall functions of microbial communities, including their roles in biodegradation and bioactive compound production. Despite these advancements, traditional research methods, such as "culture" and "physiological analysis" remained crucial. Cultivation and functional development of species in large-scale seaweed culture environment are still in the initial stage (Li et al 2023. Despite significant advancements in understanding the interactions between macroalgae and their associated microbiota, the application of these findings in practical settings remains limited (Lu et al 2023, Pei et al 2024). For example, while research on the influence of microorganisms on macroalgae growth and development has gained prominence, the translation of these findings into improved cultivation practices and sustainable aquaculture systems is still in its infancy (Li et al 2023).
3.5. Thematic Map
A detailed thematic map (Figure 8) was created to visually represent the density and centrality of various research themes within large marine algae cultivation and environmental microbiology from 2003 to 2023. This map captured recent changes and emerging developments in the field, offering insights into the evolving research landscape.
At the forefront of the map, themes such as "diversity", "antibacterial activity", "extracts", and "resistance" were categorized as motor themes. These themes significantly impacted the field, representing fundamental concepts relevant to various studies. In the past, the themes of “diversity”, “aquaculture”, “phytoplankton”, and “temperature” were considered to fall within the basic themes category. These themes formed the foundation of the research field, having a broad scientific impact and being supported by substantial research and citations. The study on the diversity of symbiotic microorganisms in algae provides a resource pool for the development of new antibiotics, especially the screening of natural products for multi-drug resistant bacteria has become a hot spot (Nalini et al 2017). Marine algal endophytic actinomycetes are widely distributed across various algal species, including brown algae (e.g., Laminaria ochroleuca) and mangrove-associated algae. Studies have demonstrated that 45% of actinomycete strains isolated from Laminaria ochroleuca in northern Portugal exhibited significant inhibitory activity against Staphylococcus aureus and Candida albicans through their extracts, with some strains remaining effective even at low concentrations (Ulfah et al 2021). Moreover, marine-derived actinomycetes produce diverse secondary metabolites, including polyketides, alkaloids, and aminoglycosides. Notably, staurosporine, a representative compound, exhibits broad-spectrum antifungal activity against various phytopathogenic fungi (Xiong et al 2023). Research on extracts has undergone a paradigm shift, transitioning from conventional phytochemical profiling to function-driven precision development. For instance, brown algal polyphenols, renowned for their antioxidant properties, have been leveraged in aquaculture to mitigate reliance on antibiotics by modulating the microbial community structure of aquaculture water environments (Bogolitsyn et al 2024).
In contrast, "nitrogen," "carbon," and "matter" were identified as niche themes. Although these themes currently exhibited lower centrality, their developmental density suggested they held the potential to influence future research directions significantly. Their positioning close to the center of the map indicated a high likelihood of evolving into motor or basic themes. Research on the role of microorganisms in carbon and nitrogen cycles within macroalgal ecosystems was crucial for understanding system functionality and stability, enhancing aquaculture efficiency, promoting environmental sustainability, combating climate change, and developing new biological resources. Research elucidating the microbially-driven mechanisms of carbon (C) cycles has emerged as a critical nexus for marine ecological restoration and the advancement of the blue economy. Investigations into the marine carbon cycle have prioritized the degradation of algae-derived polysaccharides, with bacterial decomposition recognized as a key driver of carbon sequestration through the microbial carbon pump (MCP) mechanism (Hu et al 2023, Huang et al 2024). To address current scalability limitations, recent efforts synergistically integrate the C-N cycling functionalities of microbial communities with global carbon sink quantification frameworks, thereby providing a scientific foundation for climate policy formulation.
It was important to note that themes such as "inhibition," "morphogenesis" and "patterns" played dual roles in this context. They were central driving forces in the field and specialized areas of focus. "Inhibition" relates to microorganisms’ critical interactions, influencing ecological dynamics and disease resistance. "Morphogenesis" pertained to the developmental processes of algae, which were essential for understanding growth patterns and improving cultivation practices. "Patterns" reflected microbial communities’ complex ecological relationships and distribution, crucial for deciphering the mechanisms governing their structure and function. This dual categorization highlighted their significance as broad, field-defining themes and detailed research areas, offering nuanced insights and targeted innovations.
Additionally, themes such as "growth-performance", "disease resistance", and "lactic-acid bacteria" showed interesting trends when analyzed in terms of emerging or declining research foci. Analyzing these themes through co-occurrence patterns and temporal frequency shifts illuminate their current status and predict future trajectories. Notably, research on the influence of microorganisms on macroalgae growth and development has gained prominence since 2018, indicating it as a burgeoning research area. Phaeobacter sp. BS52, isolated from healthy D. pulchra, was antagonistic towards bleaching pathogens and significantly increased the proportion of healthy individuals when applied before the pathogen challenge (pathogen-only vs. BS52 + pathogen: 41–80%), and to a level similar to the control (Li et al 2021). Such a role of probiotics on the homeostasis of host microbiota, through directly or indirectly reducing the negative effect of biotic/ abiotic stress have also been reported in terrestrial plants and humans (Zhang et al 2020, Zhang et al 2020). Future research directions propose that targeted screening of functional probiotics and their application in microbial community engineering could enhance the ecological stability and disease resilience of aquaculture microbiomes, thereby optimizing host-microbe-environment interactions for sustainable aquaculture practices.
3.6. Prospects of the Environmental Microbiology of Macroalgae Aquaculture
The discipline of environmental microbiology in macroalgae aquaculture is undergoing a paradigm shift, driven by cutting-edge omics technologies and systems biology approaches. These advancements are unraveling the intricate metabolic networks and ecological interactions between macroalgae and their associated microbial communities, offering transformative potential for sustainable marine resource utilization. A critical frontier lies in leveraging microbial contributions to carbon cycling for climate mitigation. Recent studies highlight the pivotal role of microbial communities in converting labile dissolved organic carbon (DOC) into refractory DOC (RDOC) through the Microbial Carbon Pump (MCP) mechanism, thereby enhancing long-term carbon sequestration in marine sediments (Jiao et al 2010; Xiong et al 2024). However, emerging evidence challenges the simplistic view of macroalgae systems as universal carbon sinks, revealing context-dependent carbon flux dynamics influenced by microbial respiration during algal senescence (Gallagher et al 2022, Huang et al 2024). To address this, future research must integrate microbial carbon fixation into global carbon accounting frameworks, particularly in Integrated Multi-Trophic Aquaculture (IMTA) systems, which demonstrate 2.8-fold higher carbon sequestration rates compared to monocultures (Tang et al 2014; Wang et al 2025).
Innovations in microbial community engineering are equally promising. High-throughput sequencing and single-cell transcriptomics (e.g., bacterial-MERFISH) now enable spatially resolved functional profiling of unculturable taxa, uncovering novel biosynthetic gene clusters (BGCs) for antimicrobial and antifouling compounds (Sarfaties et al 2025; Lu et al 2023). For instance, Bacillota strains in macroalgae ecosystems harbor BGCs encoding polyketides and terpenoids, offering untapped potential for pharmaceutical and bioenergy applications (Baltz 2021,Chukwudulue et al 2023). Moreover, targeted probiotics such as Phaeobacter sp. BS52 have proven effective in suppressing pathogens and enhancing host resilience, reducing disease incidence by 39–80% in controlled trials (Li et al 2021). These findings underscore the need for precision microbiota manipulation to optimize algal growth and nutrient assimilation, particularly under nitrogen-limited conditions where urease-producing bacteria (UPB) and nitrogen-fixing Rhodobacteraceae play critical roles (Pei et al 2023, Nair et al 2022).
Interdisciplinary integration is paramount. Combining metabolomics, CRISPR-based gene editing, and AI-driven ecological modeling could decode host-microbe-environment interactions at unprecedented resolution. For example, convolutional neural networks may predict biofilm functionality, while CRISPRi silencing of exopolysaccharide genes could enable precise biofilm modulation (Qian et al 2022). Concurrently, stakeholder engagement and policy frameworks must evolve to align microbial-based solutions with Sustainable Development Goals (SDGs), emphasizing blue carbon strategies and circular bioeconomy principles. By bridging gaps between molecular insights and scalable aquaculture practices, this multidisciplinary synergy will not only enhance ecological resilience but also position macroalgae systems as keystones of global food security and climate action.
4. Conclusions
This study conducted an in-depth bibliometric analysis of 610 publications focused on microorganisms within macroalgae cultivation ecosystems. Utilizing the Bibliometrix package in R Studio and VOSviewer software for bibliometric analysis and network visualization, the research elucidated foundational information in this field, emphasizing the significance of sustainable development and ecological insights. The main conclusions are as follows:
- Research Focus Areas: Keyword co-occurrence network analysis identified five primary research areas within macroalgae aquaculture's environmental microbiology: i. identification of microbial species and functional genes; ii. biogeochemical cycles of carbon in microbial communities; iii. interactions between microorganisms and macroalgae in macroalgal cultivation environments; iv. microbial resistance and bioactive substances in macroalgae culture environments; v. high-throughput sequencing and database utilization in microorganisms in the cultivation ecosystems of macroalgae.
- Thematic Evolution: Exploring microbial diversity remains a fundamental focus within the field. The study of microbial carbon and nitrogen cycles emerges as a burgeoning frontier, ripe with potential for significant scientific breakthroughs. Meanwhile,Innovations in microbial community engineering are equally promising. Understanding the intricate web of microbial interactions and the ecological models governing microbial communities is essential for deciphering their ecological roles and contributions.
- Technological Advances: The integration of systems biology has significantly deepened our understanding of microbial microorganisms, laying a strong foundation for advanced aquaculture practices. Embracing interdisciplinary approaches and combining expertise from various fields is crucial to addressing the complex ecological challenges in aquatic ecosystems. With the advancement of single-cell transcriptomics and spatiotemporal metabolomics, the multi - omics technology is set to be increasingly applied to the study of microorganisms in macroalgal culture environments.
- Future Directions: Future investigations are anticipated to prioritize the development of sustainable farming practices and explore bioactive compounds to improve aquaculture systems' health and productivity. Moreover, the role of macroalgae in carbon sequestration and environmental remediation underscores its potential as a sustainable strategy for addressing climate change, thereby highlighting its ecological significance.
Author Contributions
P.P. and H.D. conceived and designed framework of this study. Y.C. and P. P. performed literature investigation and data collection. Y.C. analyzed data and wrote the draft. R.B. and P.L. contributed analytical method. P.P., M.A., M.S. and H.D. revised the manuscript. All authors reviewed and approved the final manuscript.
Funding
This research was supported by several funding sources, including National Key R & D Program of China (2023YFD2400100), research on breeding technology of candidate species for Guangdong modern marine ranching (2024-MRB-00-001), and the Program for University Innovation Team of Guangdong Province (2022KCXTD008).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The dataset(s) supporting the conclusions of this article is (are) included within the article and its additional file(s).
Acknowledgments
We thank Dr. Jia Yang of Shantou University for her help with collecting and collating document literature. We would like to express our sincere gratitude to editor-in-chief for handling and reviewing our manuscript in time, and several reviewers for their professional and careful guidance.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships.
References
- Abdelrazek H M, Shams El-Din N G, Ghozlan H A, Sabry S A and Abouelkheir S S 2024 Distribution and functional perspective analysis of epiphytic and endophytic bacterial communities associated with marine seaweeds, Alexandria shores, Egypt. BMC Microbiology, 24:293. [CrossRef]
- Albakosh M A, Naidoo R K, Kirby B and Bauer R 2016 Identification of epiphytic bacterial communities associated with the brown alga Splachnidium rugosum. Journal of Applied Phycology, 28(3), 1891–1901. [CrossRef]
- Ashen J B and Goff L J 2000 Molecular and Ecological Evidence for Species Specificity and Coevolution in a Group of Marine Algal-Bacterial Symbioses. Applied and Environmental Microbiology, 66(7), 3024–3030. https://journals.asm.org/journal/aem.
- Baltz R H 2017 Gifted microbes for genome mining and natural product discovery. Journal of Industrial Microbiology and Biotechnology, 44(4–5), 573–588. [CrossRef]
- Baltz R H 2021 Genome mining for drug discovery: Progress at the front end. In Journal of Industrial Microbiology and Biotechnology, Oxford University Press, 48, 9–10. [CrossRef]
- Bogolitsyn K, Dobrodeeva L, Samodova A, Parshina A 2024 In vitro Immunostimulant Activity of the Polyphenolic Extract from the Arctic Brown Algae Fucus vesiculosus. Plant Foods for Human Nutrition, 79(2), 511-517. [CrossRef]
- Brown A R, Lilley M, Shutler J, Lowe C, Artioli Y, Torres R, Berdalet E and Tyler C R 2020 Assessing risks and mitigating impacts of harmful algal blooms on mariculture and marine fisheries. Reviews in Aquaculture, 12(3), 1663-1688. [CrossRef]
- Cai J, Lovatelli A, Aguilar-Manjarrez J, Cornish L, Dabbadie L, Desrochers A, Diffey S, Garrido Gamarro E, Geehan J, Hurtado A, Lucente D, Mair G, Miao W, Potin P, Przybyla C, Reantaso M, Roubach R, Tauati M and Yuan X 2021 Seaweeds and microalgae: An overview for unlocking their potential in global aquaculture development. FAO Fisheries and Aquaculture Circular No. 1229. Rome: FAO. [CrossRef]
- Chai Z, Hu Z, Deng Y, Yang Y and Tang Y Z 2021 Interactions between the seaweed Gracilaria and dinoflagellate Akashiwo sanguinea in an indoor co-cultivation system and the interference of bacteria. Journal of Applied Phycology, 33(5), 3153–3163. [CrossRef]
- Chai Z Y, He Z L, Deng Y Y, Yang Y F and Tang Y Z 2018 Cultivation of seaweed Gracilaria lemaneiformis enhanced biodiversity in a eukaryotic plankton community as revealed via metagenomic analyses. Molecular Ecology, 27(4), 1081–1093. [CrossRef]
- Carrell A A, Veličković D, Lawrence T J, Bowen B P, Louie K B, Carper D L, Chu R K, Mitchell H D, Orr G, Markillie L M, Jawdy S S, Grimwood J, Shaw A J, Schmutz J, Northen T R, Anderton C R, Pelletier D A, Weston D J. Novel metabolic interactions and environmental conditions mediate the boreal peatmoss-cyanobacteria mutualism. ISME, 2022, 16(4): 1074–1085. [CrossRef]
- Chen J, Li H, Zhang Z, He C, Shi Q, Jiao N and Zhang Y 2020 DOC dynamics and bacterial community succession during long-term degradation of Ulva prolifera and their implications for the legacy effect of green tides on refractory DOC pool in seawater. Water Research, 185, 116268. [CrossRef]
- Chukwudulue U M, Barger N, Dubovis M and Luzzatto Knaan T 2023 Natural products and pharmacological properties of symbiotic Bacillota (Firmicutes) of marine macroalgae. Marine Drugs, 21(11), 569. [CrossRef]
- Croft M T, Lawrence A D, Raux-Deery E, Warren M J and Smith A G 2005 Algae acquire vitamin B12 through a symbiotic relationship with bacteria. Nature, 438(7064), 90–93. [CrossRef]
- Ding Q, Song X, Yuan M, Sun R, Zhang J, Yin L and Pu Y 2022 Removal of microcystins from water and primary treatment technologies-A comprehensive understanding based on bibliometric and content analysis, 1991–2020. Journal of Environmental Management, 305, 114349. [CrossRef]
- Dittami S M, Duboscq-Bidot L, Perennou M, Gobet A, Corre E, Boyen C and Tonon T 2016 Host-microbe interactions as a driver of acclimation to salinity gradients in brown algal cultures. ISME Journal, 10(1), 51–63. [CrossRef]
- Dubois S, Marin-Léal J C, Ropert M and Lefebvre S 2007 Effects of oyster farming on macrofaunal assemblages associated with Lanice conchilega tubeworm populations: A trophic analysis using natural stable isotopes. Aquaculture, 271(1–4), 336–349. [CrossRef]
- FAO 2024 The State of World Fisheries and Aquaculture 2024 – Blue Transformation in action. Rome. [CrossRef]
- Gallagher J B, Shelamoff V and Layton C 2022 Seaweed ecosystems may not mitigate CO2 emissions. ICES Journal of Marine Science, 79(3), 585–592. [CrossRef]
- Gao G, Beardall J, Jin P, Gao L, Xie S and Gao K 2022 A review of existing and potential blue carbon contributions to climate change mitigation in the Anthropocene. Journal of Applied Ecology, 59(7), 1686–1699. [CrossRef]
- Gao G, Gao L, Jiang M, Jian A and He L 2022 The potential of seaweed cultivation to achieve carbon neutrality and mitigate deoxygenation and eutrophication. Environmental Research Letters, 17(1). [CrossRef]
- Garland G, Edlinger A, Banerjee S, Degrune F, García-Palacios P, Pescador D S, Herzog C, Romdhane S, Saghai A, Spor A, Wagg C, Hallin S, Maestre F T, Philippot L, Rillig M C and van der Heijden M G A 2021 Crop cover is more important than rotational diversity for soil multifunctionality and cereal yields in European cropping systems. Nature Food, 2(1), 28–37. [CrossRef]
- Goecke F, Labes A, Wiese J and Imhoff J F 2013 Phylogenetic analysis and antibiotic activity of bacteria isolated from the surface of two co-occurring macroalgae from the Baltic Sea. European Journal of Phycology, 48(1), 47–60. [CrossRef]
- Huang H, Zhan S, Shao K, Chen H and Fan J 2024 Spatial distribution characteristics and interaction effects of DOM and microbial communities in kelp cultivation areas. Science of the Total Environment, 10(920), 170511. [CrossRef]
- Jarone P, Laura G, Laura A, Maria M S 2006 Seasonal changes in bacterioplankton nutrient limitation and their effects on bacterial community composition in the NW Mediterranean Sea. Aquatic Microbal Ecology, 44, 241–252. [CrossRef]
- Jiao J Y, Liu L, Hua Z S, Fang B Z, Zhou E M, Salam N, Hedlund B P and Li W J 2021 Microbial dark matter coming to light: Challenges and opportunities. National Science Review, 8(3), 280. [CrossRef]
- Jiao N, Herndl G J, Hansell D A, Benner R, Kattner G, Wilhelm S W, Kirchman D L, Weinbauer M G, Luo T, Chen F and Azam F 2010 Microbial production of recalcitrant dissolved organic matter: Long-term carbon storage in the global ocean. Nature Reviews Microbiology, 8(8), 593–599. [CrossRef]
- Joint I, Tait K and Wheeler G 2007 Cross-kingdom signalling: Exploitation of bacterial quorum sensing molecules by the green seaweed Ulva. In Philosophical Transactions of the Royal Society B: Biological Sciences, 362(1483), 1223–1233. [CrossRef]
- Jonnadula R, Verma P, Shouche Y S and Ghadi S C 2009 Characterization of microbulbifer strain CMC-5, a new biochemical variant of microbulbifer elongatus type strain DSM6810T isolated from decomposing seaweeds. Current Microbiology, 59(6), 600–607. [CrossRef]
- Kaur M, Saini K C, Mallick A and Bast F 2023 Seaweed-associated epiphytic bacteria: Diversity, ecological and economic implications. Aquatic Botany, 189, 103698. [CrossRef]
- Kim G H, Klochkova T A, Lee D J and Im S H 2016 Chloroplast virus causes green-spot disease in cultivated Pyropia of Korea. Algal Research, 17, 293–299. [CrossRef]
- KleinJan H, Frioux C, Califano G, Aite M, Fremy E, Karimi E, Corre E, Wichard T, Siegel A, Boyen C and Dittami S M 2023 Insights into the potential for mutualistic and harmful host–microbe interactions affecting brown alga freshwater acclimation. Molecular Ecology, 32(3), 703–723. [CrossRef]
- Krause-Jensen D and Duarte C M 2016 Substantial role of macroalgae in marine carbon sequestration. Nature Geoscience, 9(10), 737–742. [CrossRef]
- Kwan V, Fong J, Ng C S L, and Huang D 2022 Temporal and spatial dynamics of tropical macroalgal contributions to blue carbon. Science of the Total Environment, 828, 154369. [CrossRef]
- Lamilla-Tamayo L, Escobar-Calderón F and Skalický M 2023 Reviewing the potential of algae species as a green alternative to produce nanoparticles: Findings from a database analysis. Water, 15(12), 2208. [CrossRef]
- Lead J C, Gamarro E G, Geehan J A, Lucente D, Mair G C, Miao W, Reantaso M B, Roubach R, Yuan X and Potin P 2021 Seaweeds and microalgae: an overview for unlocking their potential in global aquaculture development. https://openknowledge.fao.org/handle/20.500.14283/cb5670en.
- Lee S J, Hwang M S, Park M A, Baek J M, Ha D S, Lee J E and Lee S R 2015 Molecular identification of the algal pathogen Pythium chondricola (Oomycetes) from Pyropia yezoensis (Rhodophyta) using ITS and cox1 markers. Algae, 30(3), 217–222. [CrossRef]
- Li J, Weinberger F, de Nys R, Thomas T, Egan S 2023 A pathway to improve seaweed aquaculture through microbiota manipulation. Trends in Biotechnology, 41(4), 545–556. [CrossRef]
- Li J, Majzoub ME, Marzinelli EM, Dai Z, Thomas T, Egan S 2022 Bacterial controlled mitigation of dysbiosis in a seaweed disease. ISME., 16(2), 378-387. [CrossRef]
- Li T, Huang J, Du H, Liu X, Zhong C and Lin S 2022 Coral bleaching from a nutrient perspective is understudied: A bibliometric survey. Frontiers in Marine Science, 9. [CrossRef]
- Lu D C, Wang F Q, Amann R I, Teeling H and Du Z J 2023 Epiphytic common core bacteria in the microbiomes of co-located green (Ulva), brown (Saccharina) and red (Grateloupia, Gelidium) macroalgae. Microbiome, 11(1), 126. [CrossRef]
- Lu K, Lin W and Liu J 2008 The characteristics of nutrient removal and inhibitory effect of Ulva clathrata on Vibrio anguillarum 65. Journal of Applied Phycology, 20(6), 1061–1068. [CrossRef]
- Marrone M and Linnenluecke M K 2020 Interdisciplinary Research Maps: A new technique for visualizing research topics. PLoS ONE, 15(11), e0242283. [CrossRef]
- Marshall K, Joint I, Callow M E and Callow J A 2006 Effect of marine bacterial isolates on the growth and morphology of axenic plantlets of the green alga Ulva linza. Microbial Ecology, 52(2), 302–310. [CrossRef]
- Matsuo Y, Suzuki M, Kasai H, Shizuri Y and Harayama S 2003 Isolation and phylogenetic characterization of bacteria capable of inducing differentiation in the green alga Monostroma oxyspermum. Environmental Microbiology, 5(1), 25-35. [CrossRef]
- Middelburg J 2011 Chemoautotrophy in the ocean. Geophysical Research Letters, 2011, 38(24): L24604.
- Mongeon P and Paul-Hus A 2016 The journal coverage of Web of Science and Scopus: a comparative analysis. Scientometrics, 106(1), 213–228. [CrossRef]
- Nair S, Zhang Z, Li H, Zhao H, Shen H, Kao S J, Jiao N and Zhang Y 2022 Inherent tendency of Synechococcus and heterotrophic bacteria for mutualism on long-term coexistence despite environmental interference. Science Advance, 8(39), eabf4792.
- Nalini MS, Prakash HS 2017 Diversity and bioprospecting of actinomycete endophytes from the medicinal plants. Letters in Applied Microbiology, 64(4), 261-270. [CrossRef]
- Ortega A, Geraldi N R, Alam I, Kamau A A, Acinas S G, Logares R, Gasol J M, Massana R, Krause-Jensen D and Duarte C M 2019 Important contribution of macroalgae to oceanic carbon sequestration. Nature Geoscience, 12(9), 748–754. [CrossRef]
- Pagarete A, Ramos A S, Puntervoll P, Allen M J and Verdelho V 2021 Antiviral potential of algal metabolites-a comprehensive review. Marine Drugs, 19(2), 94. [CrossRef]
- Paul J and Criado A R 2020 The art of writing literature review: What do we know and what do we need to know? International Business Review, 29(4), 101717. [CrossRef]
- Pedersen M F, Filbee-Dexter K, Norderhaug K M, Fredriksen S, Frisk N L, Fagerli C W and Wernberg T 2020 Detrital carbon production and export in high latitude kelp forests. Oecologia, 192(1), 227–239. [CrossRef]
- Pei P, Aslam M, Du H, Liang H, Wang H, Liu X and Chen W 2021 Environmental factors shape the epiphytic bacterial communities of Gracilariopsis lemaneiformis. Scientific Reports, 11(1), 13253. [CrossRef]
- Pei P, Aslam M, Wang H, Ye P, Li T, Liang H, Lin Q, Chen W and Du H 2024 Diversity and ecological function of urease-producing bacteria in the cultivation environment of Gracilariopsis lemaneiformis. Microbial Ecology, 87(1), 35. [CrossRef]
- Penesyan A, Kjelleberg S and Egan S 2010 Development of novel drugs from marine surface associated microorganisms. Marine Drugs. 8(3), 438–459. [CrossRef]
- Penesyan A, Marshall-Jones Z, Holmstrom C, Kjelleberg S and Egan S 2009 Antimicrobial activity observed among cultured marine epiphytic bacteria reflects their potential as a source of new drugs: Research article. FEMS Microbiology Ecology, 69(1), 113–124. [CrossRef]
- Penesyan A, Tebben J, Lee M, Thomas T, Kjelleberg S, Harder T and Egan S 2011 Identification of the antibacterial compound produced by the marine epiphytic bacterium Pseudovibrio sp. D323 and related sponge-associated bacteria. Marine Drugs, 9(8), 1391-1402. [CrossRef]
- Peng Z, Wang P, Luo X, Deng Q, Yang Z, Wu J, Xian W, Yan W, Mou X, Yuan Y, Li W and Li J 2024 Community structure and carbon metabolism functions of bacterioplankton in the Guangdong coastal zone. Marine Life Science and Technology, 6, 547-561. [CrossRef]
- Pfister C A, Altabet M A and Weigel B L 2019 Kelp beds and their local effects on seawater chemistry, productivity, and microbial communities. Ecology, 100(8), e02798. [CrossRef]
- Pfister C A and Altabet M A 2019 Enhanced microbial nitrogen transformations in association with macrobiota from the rocky intertidal. Biogeosciences, 16, 193-206. [CrossRef]
- Qian P Y, Cheng A, Wang R and Zhang R 2022 Marine biofilms: diversity, interactions and biofouling. Nature Reviews Microbiology. 20(11), 671–684. [CrossRef]
- Racine P, Marley A C, Froehlich H E, Gaines S D, Ladner I, MacAdam–Somer I, and Bradley D 2021 A case for seaweed aquaculture inclusion in U.S. nutrient pollution management. Marine Policy, 129, 104506. [CrossRef]
- Reisky L, Préchoux A, Zühlke M K, Bäumgen M, Robb C S, Gerlach N, Roret T, Stanetty C, Larocque R, Michel G, Song T, Markert S, Unfried F, Mihovilovic M D, Trautwein–Schult A, Becher D, Schweder T, Bornscheuer U T and Hehemann J H 2019 A marine bacterial enzymatic cascade degrades the algal polysaccharide ulvan. Nature Chemical Biology, 15(8), 803–812. [CrossRef]
- Saravanan P, Chatterjee A, Kiran K J, Bhowmick G D, Sappati P K and Nagarajan V 2024 Exploring Seaweed–Associated Marine Microbes: Growth Impacts and Enzymatic Potential for Sustainable Resource Utilization. Indian Journal of Microbiology 64, 593602. [CrossRef]
- Sarfatis A, Wang Y, Twumasi-Ankrah N, Moffitt J R 2025 Highly multiplexed spatial transcriptomics in bacteria. Science, 387(6732): eadr0932. [CrossRef]
- Singh R P and Reddy C R K 2014 Seaweed–microbial interactions: Key functions of seaweed–associated bacteria. FEMS Microbiology Ecology, 88(2), 213–230. [CrossRef]
- Spencer S J, Tamminen M V, Preheim S P, Guo M T, Briggs A W, Brito I L A, Weitz D, Pitkänen L K, Vigneault F, Virta M P and Alm E J 2016 Massively parallel sequencing of single cells by epicPCR links functional genes with phylogenetic markers. ISME Journal, 10(2), 427–436. [CrossRef]
- Sun Y, Li H, Wang X, Li H and Deng Y 2023 Kelp Culture Enhances Coastal Biogeochemical Cycles by Maintaining Bacterioplankton Richness and Regulating Its Interactions. mSystems, 8(2), 14. [CrossRef]
- Sylvers P H and Gobler C J 2021 Mitigation of harmful algal blooms caused by Alexandrium catenella and reduction in saxitoxin accumulation in bivalves using cultivable seaweeds. Harmful Algae, 105, 102056. [CrossRef]
- Tang Q 2014 Management strategies of marine food resources undermultiple stressors with particular reference of the Yellow Sea large marine ecosystem. Frontiers in Agricultural Science and Engineering, 1(1), 85-90. [CrossRef]
- Thomas F, Barbeyron T and Michel G 2011 Evaluation of reference genes for real–time quantitative PCR in the marine flavobacterium Zobellia galactanivorans. Journal of Microbiological Methods, 84(1), 61–66. [CrossRef]
- Ulrich J F, Gräfe M S, Dhiman S, Wienecke P, Arndt H D and Wichard T 2022 Thallusin Quantification in Marine Bacteria and Algae Cultures. Marine Drugs, 20(11), 609. [CrossRef]
- Ulfah M, Kasanah N, Wijayanti N. Antivibriosis and cytotoxicity of Actinobacteria associated with red seaweed Gelidiella acerosa. Aquaculture Research, 2021, 1, 1-9. [CrossRef]
- Van Eck N J and Waltman L 2007 VOS: A New Method for Visualizing Similarities Between Objects. In: Decker, R, Lenz, H.J eds) Advances in Data Analysis. Studies in Classification, Data Analysis, and Knowledge Organization. Springer, Berlin, Heidelberg. [CrossRef]
- Viju N, Punitha S M J and Satheesh S 2021 An Analysis of Biosynthesis Gene Clusters and Bioactivity of Marine Bacterial Symbionts. Current Microbiology, 78(7), 2522–2533. Springer. [CrossRef]
- Wang G, Shuai L, Li Y, Lin W, Zhao X and Duan D 2008 Phylogenetic analysis of epiphytic marine bacteria on Hole–Rotten diseased sporophytes of Laminaria japonica. Journal of Applied Phycology, 20(4), 403–409. [CrossRef]
- Wang W, Wu L, Xu K, Xu Y, Ji D, Chen C and Xie C 2020 The cultivation of Pyropia haitanensis has important impacts on the seawater microbial community. Journal of Applied Phycology, 32, 2561–2573. [CrossRef]
- Wang Q, Sun X, Lin S, Dong Y, Shen H, He Z, Luo H, Zou L, Chung I K, Yang Y 2025 Large-scale seaweed cultivation as a nature solution for carbon-negative economy and restorative environmental stewardship: Lessons from China. Renewable and Sustainable Energy Reviews, 207, 114954. [CrossRef]
- Wang Z, Xiao T, Pang S, Liu M and Yue H 2009 Isolation and identification of bacteria associated with the surfaces of several algal species. Chinese Journal of Oceanology and Limnology, 27(3), 487–492. [CrossRef]
- Wani H M, Chen C W, Huang C Y, Singhania R R, Sung Y J, Dong C and Patel A K 2023 Development of Bioactive Peptides Derived from Red Algae for Dermal Care Applications: Recent Advances. Sustainability, 15(11), 8506. [CrossRef]
- Weigel B L, Miranda K K, Fogarty E C, Watson A R and Pfister C A 2022 Functional Insights into the Kelp Microbiome from Metagenome–Assembled Genomes. MSystems, 7(3), e01422–21. [CrossRef]
- West J A, Klochkova T A, Kim G H and Loiseaux–De Goër S 2006 Olpidiopsis sp, an oomycete from Madagascar that infects Bostrychia and other red algae: Host species susceptibility. Phycological Research, 54(1), 72–85. http://www.botany.unimelb.
- Wichard T 2015 Exploring bacteria–induced growth and morphogenesis in the green macroalga order Ulvales (Chlorophyta Frontiers in Plant Science, 6, 1–19. [CrossRef]
- Xie X, He Z, Hu X, Yin H, Liu X and Yang Y 2017 Large–scale seaweed cultivation diverges water and sediment microbial communities in the coast of Nan’ao Island, South China Sea. Science of the Total Environment, 598, 97–108. [CrossRef]
- Xie X, He Z, Wang Q and Yang Y 2024 Diversity, composition and ecological networks of bacterial communities in response to a full cultivation cycle of the seaweed, Gracilariopsis lemaneiformis. Environmental Research, 240, 117453. [CrossRef]
- Xiong T, Li H, Hu Y, Zhai W, Zhang Z, Liu Y, Zhang J, Lu L, Chang L, Xue L, Wei Q, Jiao N and Zhang Y 2024 Seaweed farming environments do not always function as CO2 sink under synergistic influence of macroalgae and microorganisms. Agriculture Ecosystems and Environment, 361, 108824. [CrossRef]
- Xiong Z, Wang R, Xia T, Zhang S, Ma S, Guo Z. Natural Products and Biological Activity from Actinomycetes Associated with Marine Algae. Molecules, 2023, 28(13), 5138. [CrossRef]
- Xu N, Wang W, Xu K, Xu Y, Ji D, Chen C and Xie C 2022 Cultivation of different seaweed species and seasonal changes cause divergence of the microbial community in coastal seawaters. Frontiers in Microbiology, 13. [CrossRef]
- Yang Y, Luo H, Wang Q, He Z and Long A 2021 Large–scale Cultivation of Seaweed is Effective Approach to Increase Marine Carbon Sequestration and Solve Coastal Environmental Problems. Bulletin of Chinese Academy of Sciences, 36(3), 259–269. [CrossRef]
- Zhang J, Wei L, Yang J, Ahmed W, Wang Y, Fu L, Ji G 2020 Probiotic Consortia: Reshaping the Rhizospheric Microbiome and Its Role in Suppressing Root-Rot Disease of Panax notoginseng. Frontiers in Microbiology, 11:701. [CrossRef]
- Zhang J, Zhao J, Jin H, Lv R, Shi H, De G, Yang B, Sun Z, Zhang H 2020 Probiotics maintain the intestinal microbiome homeostasis of the sailors during a long sea voyage. Gut Microbes, 11(4):930-943. [CrossRef]
- Zhang Y, Ji P, Wang J and Zhao F 2016 RiboFR–Seq: A novel approach to linking 16S rRNA amplicon profiles to metagenomes. Nucleic Acids Research, 44(10), 99. [CrossRef]
- Zheng R, Cai R, Liu R, Liu G and Sun C 2021 Maribellus comscasis sp. nov, a novel deep–sea Bacteroidetes bacterium, possessing a prominent capability of degrading cellulose. Environmental Microbiology, 23(8), 4561–4575. [CrossRef]
- Zheng W, Zhao S, Yin Y, Zhang H, Needham D M, Evans E D, Dai C L, Lu P J, Alm E J and Weitz D A 2022 High–throughput, single–microbe genomics with strain resolution, applied to a human gut microbiome. Science, 376(6597), 1483. [CrossRef]
- Zheng Y, Jin R, Zhang X, Wang Q and Wu J 2019 The considerable environmental benefits of seaweed aquaculture in China. Stochastic Environmental Research and Risk Assessment, 33(4–6), 1203–1221. [CrossRef]
- Zhong C: Li T, Bi R, Sanganyado E, Huang J, Jiang S, Zhang Z and Du H 2023 A systematic overview, trends and global perspectives on blue carbon: A bibliometric study (2003–2021). Ecological Indicators, 148, 110063. [CrossRef]
Figure 1.
Research framework of the analysis process and research content on the microorganisms in the cultivation ecosystems of macroalgae.
Figure 1.
Research framework of the analysis process and research content on the microorganisms in the cultivation ecosystems of macroalgae.
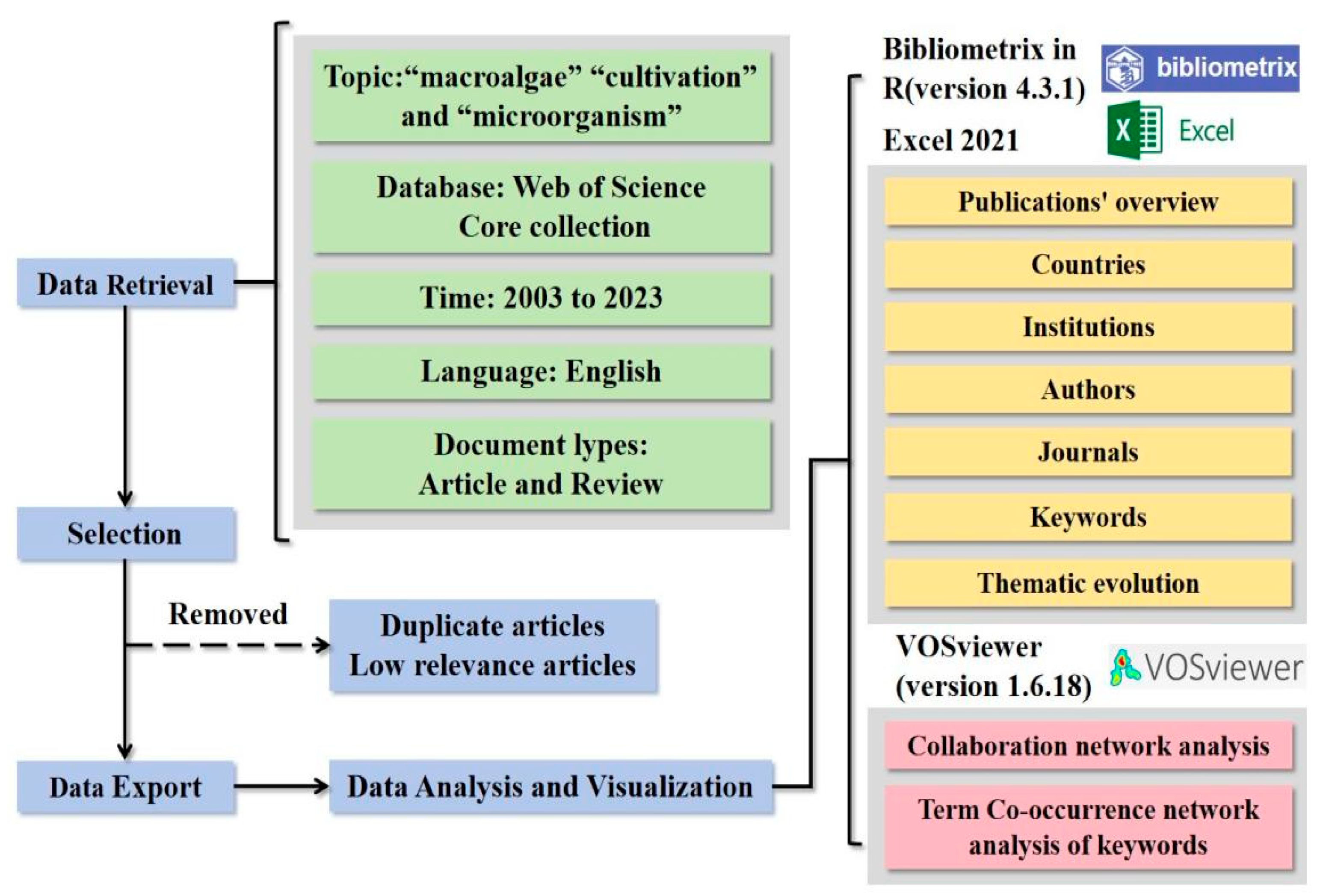
Figure 2.
Annual publications on the environmental microbiology of macroalgae cultivation in Web of Science. The bar represented the number of publications per year, and the curve represented the nonlinear fitting method (exponential model), revealing the temporal trend of annual publications.
Figure 2.
Annual publications on the environmental microbiology of macroalgae cultivation in Web of Science. The bar represented the number of publications per year, and the curve represented the nonlinear fitting method (exponential model), revealing the temporal trend of annual publications.
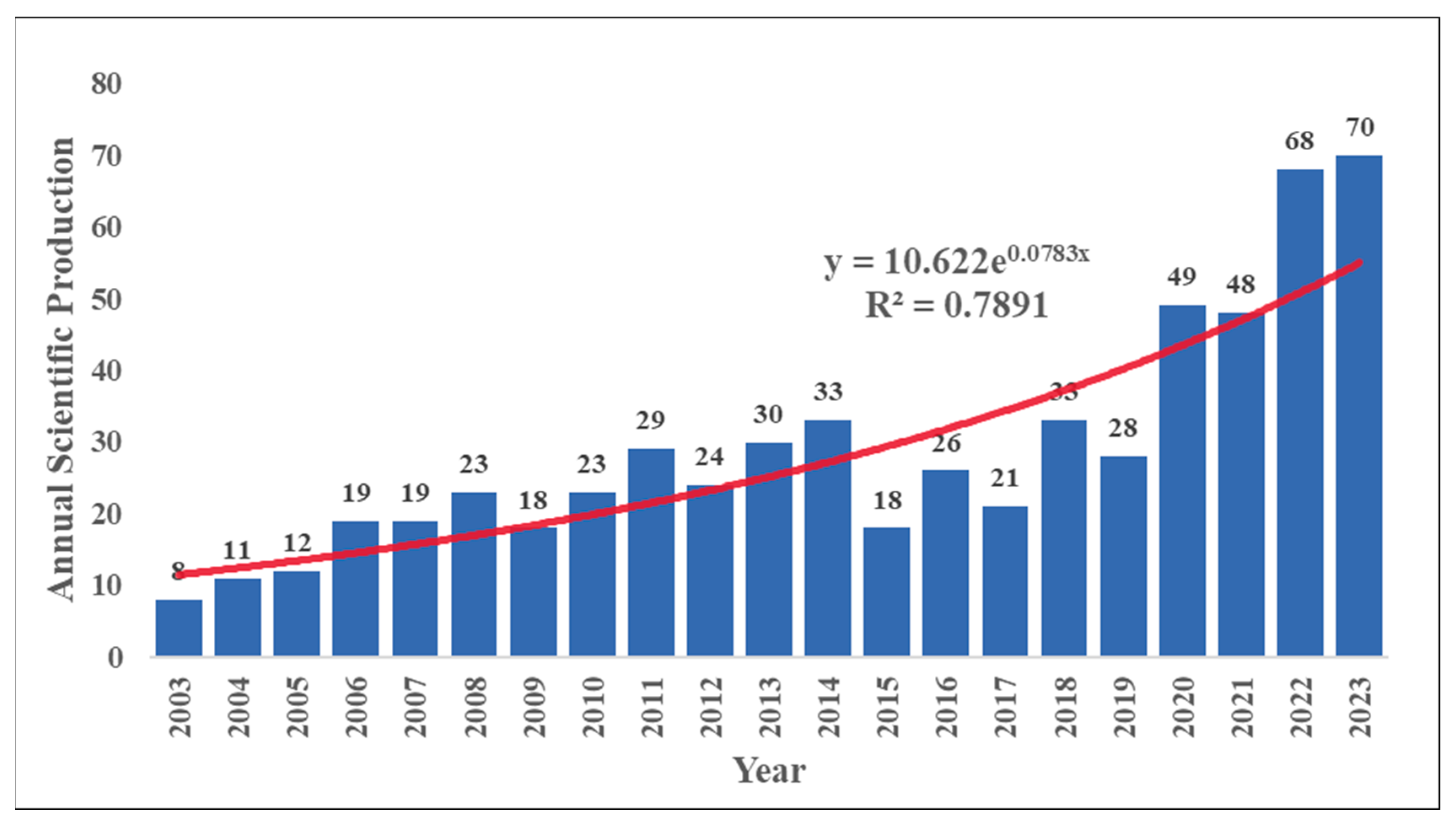
Figure 3.
Geographical heatmap based on the number of publications that countries are related to and countries' collaboration. The color depth of the countries in the map was related to the number of publications, and the thickness of the edges represents the cooperation between countries. Countries' collaboration min edges set at 2.
Figure 3.
Geographical heatmap based on the number of publications that countries are related to and countries' collaboration. The color depth of the countries in the map was related to the number of publications, and the thickness of the edges represents the cooperation between countries. Countries' collaboration min edges set at 2.
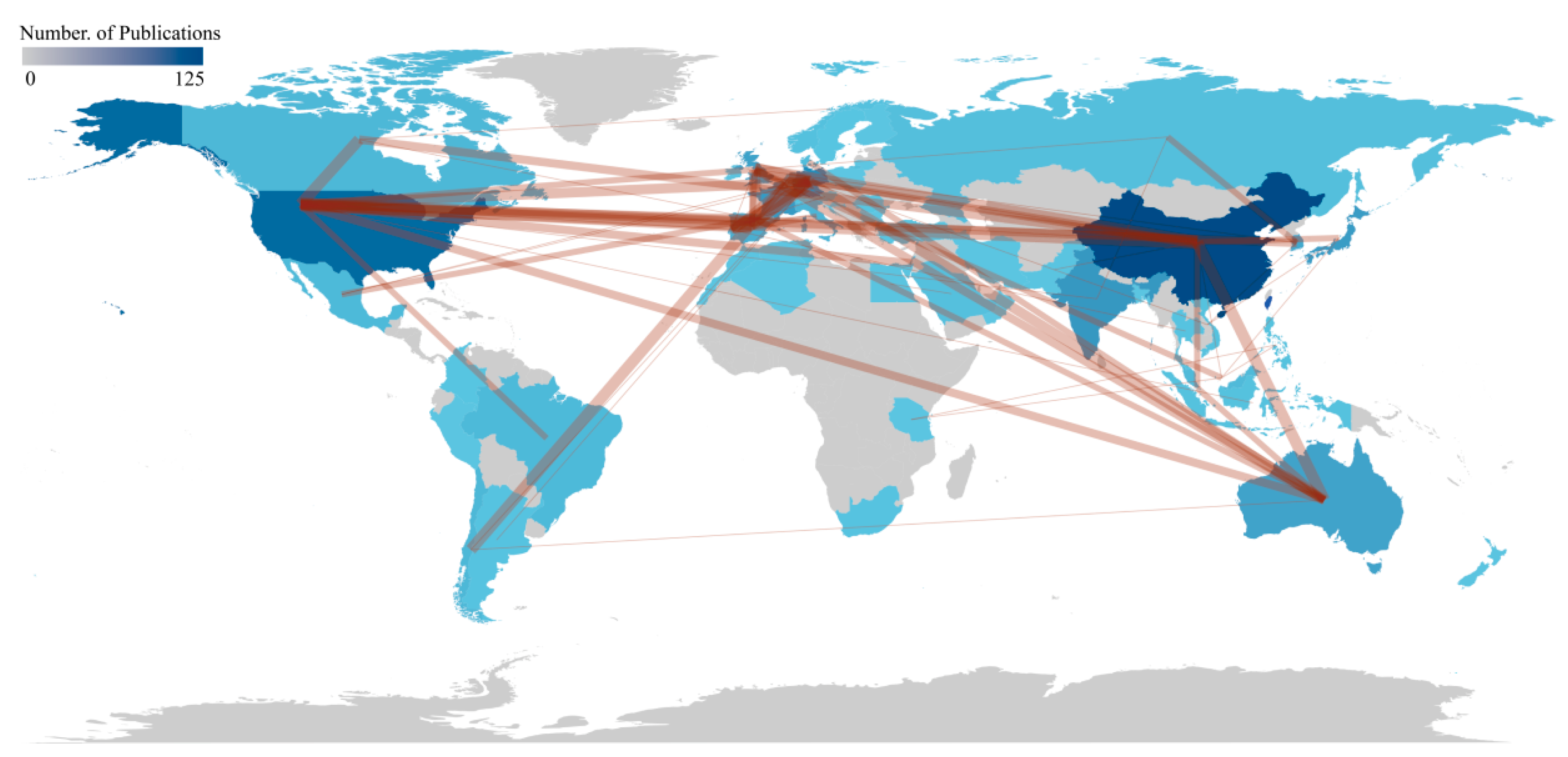
Figure 4.
(a) Network map of institutions and (b) co-authorship network of contributors involved in publishing articles on microorganisms within macroalgal cultivation ecosystems. The size of the circles represented the volume of publications, and the thickness of the lines was proportional to the frequency of collaboration. For the authors' collaboration analysis, a threshold of 3 co-authors was established, and for institutions' cooperation analysis, a threshold of 5 was set.
Figure 4.
(a) Network map of institutions and (b) co-authorship network of contributors involved in publishing articles on microorganisms within macroalgal cultivation ecosystems. The size of the circles represented the volume of publications, and the thickness of the lines was proportional to the frequency of collaboration. For the authors' collaboration analysis, a threshold of 3 co-authors was established, and for institutions' cooperation analysis, a threshold of 5 was set.
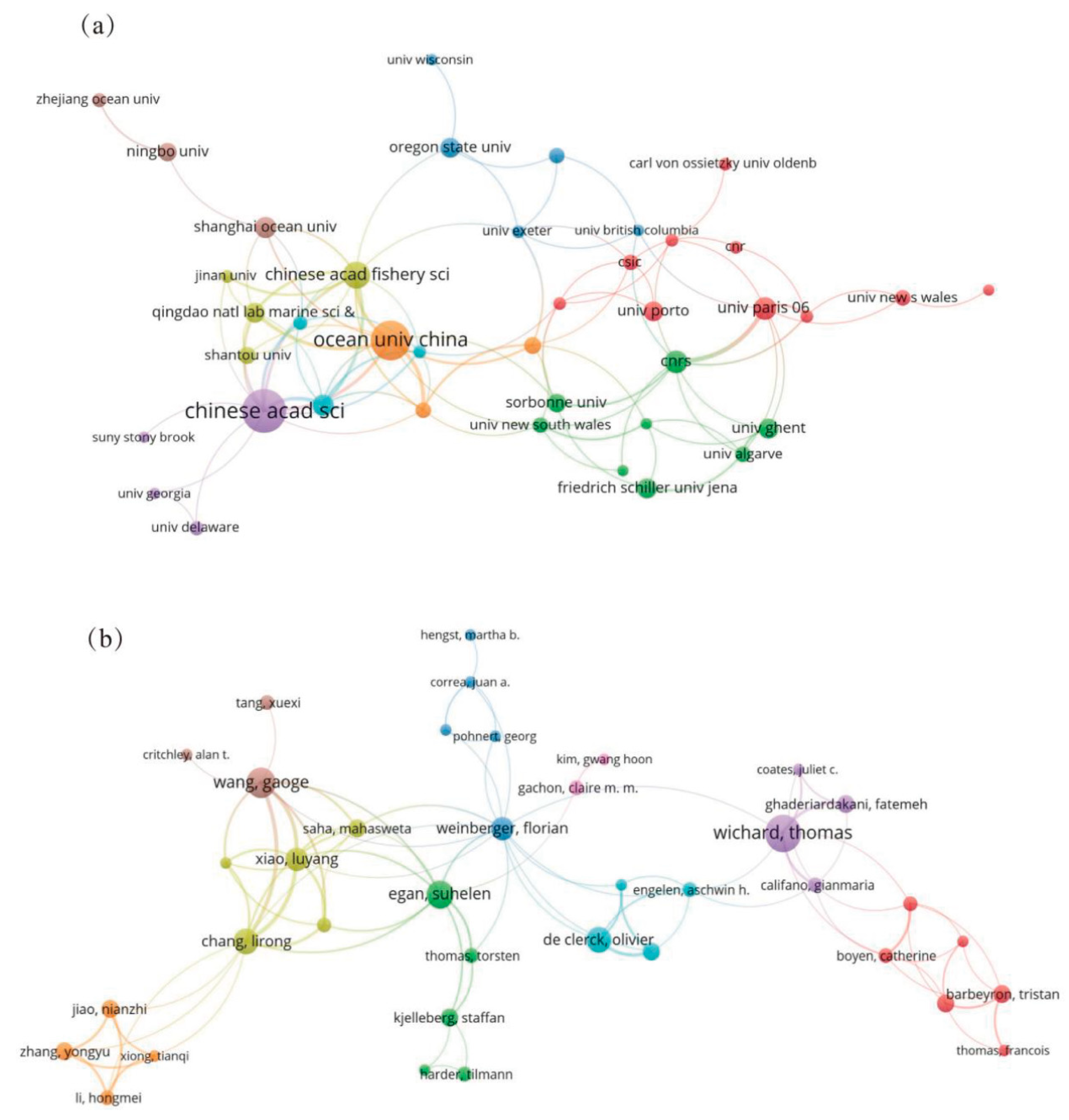
Figure 5.
The proportion of articles on the environmental microbiology of macroalgal cultivation published in various journals.
Figure 5.
The proportion of articles on the environmental microbiology of macroalgal cultivation published in various journals.
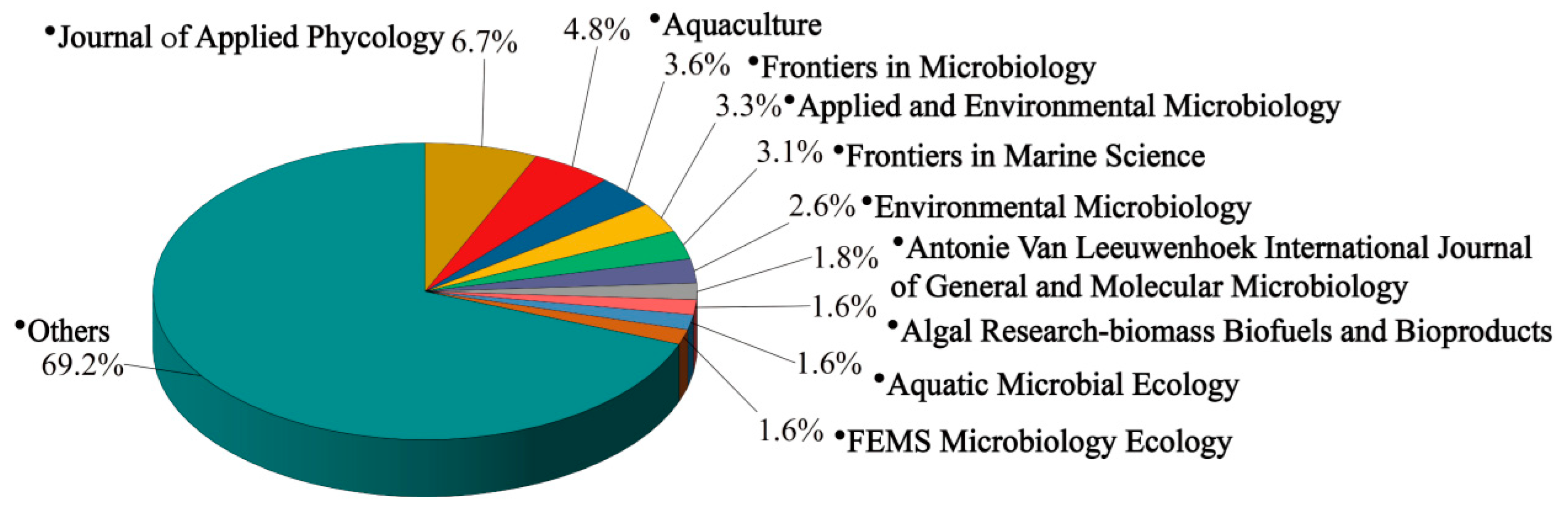
Figure 6.
Keyword cluster analysis, (a) network visualization and distinguish different thematic words with different colors. Red represents the identification of microbial species and functional genes; Yellow represents biogeochemical cycles of carbon in microbial communities; Green represents interactions between microorganisms and macroalgae in macroalgal cultivation environments; Blue represents microbial resistance and bioactive substances in macroalgae culture environments; Purple represents high-throughput sequencing and database utilization in microorganisms in the cultivation ecosystems of macroalgae. (b) overlay visualization. The color indicates the novelty of subject words.
Figure 6.
Keyword cluster analysis, (a) network visualization and distinguish different thematic words with different colors. Red represents the identification of microbial species and functional genes; Yellow represents biogeochemical cycles of carbon in microbial communities; Green represents interactions between microorganisms and macroalgae in macroalgal cultivation environments; Blue represents microbial resistance and bioactive substances in macroalgae culture environments; Purple represents high-throughput sequencing and database utilization in microorganisms in the cultivation ecosystems of macroalgae. (b) overlay visualization. The color indicates the novelty of subject words.
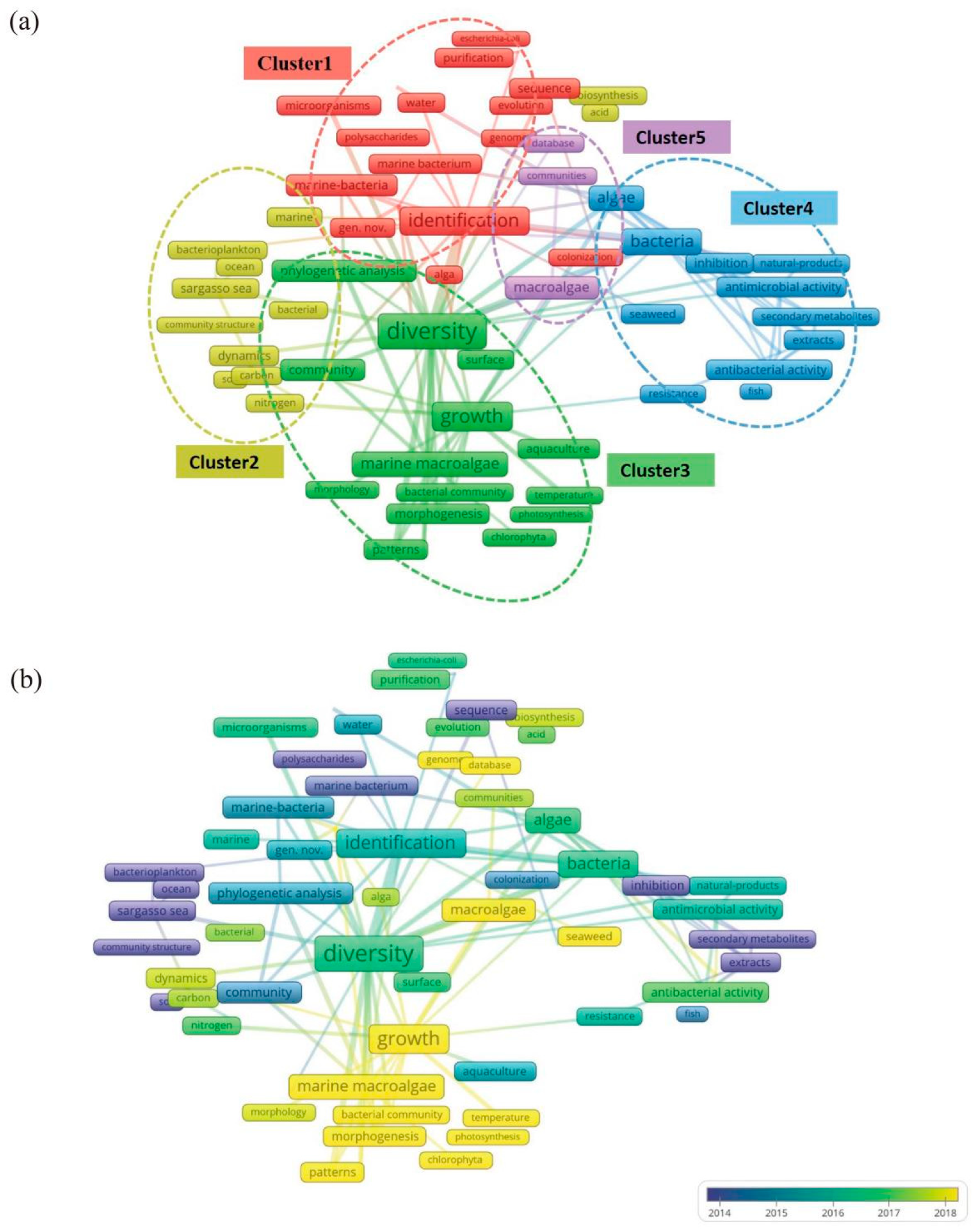
Figure 7.
Thematic evolution and map of research themes from 2003 to 2023. Transition of research themes from 2003-2008 to 2009-2017 and 2009-2017 to 2017-2023: The flow diagram illustrates the shifting focus of research themes over time. (a)(b) based on author keywords; (c)(d) based on keyword plus.
Figure 7.
Thematic evolution and map of research themes from 2003 to 2023. Transition of research themes from 2003-2008 to 2009-2017 and 2009-2017 to 2017-2023: The flow diagram illustrates the shifting focus of research themes over time. (a)(b) based on author keywords; (c)(d) based on keyword plus.
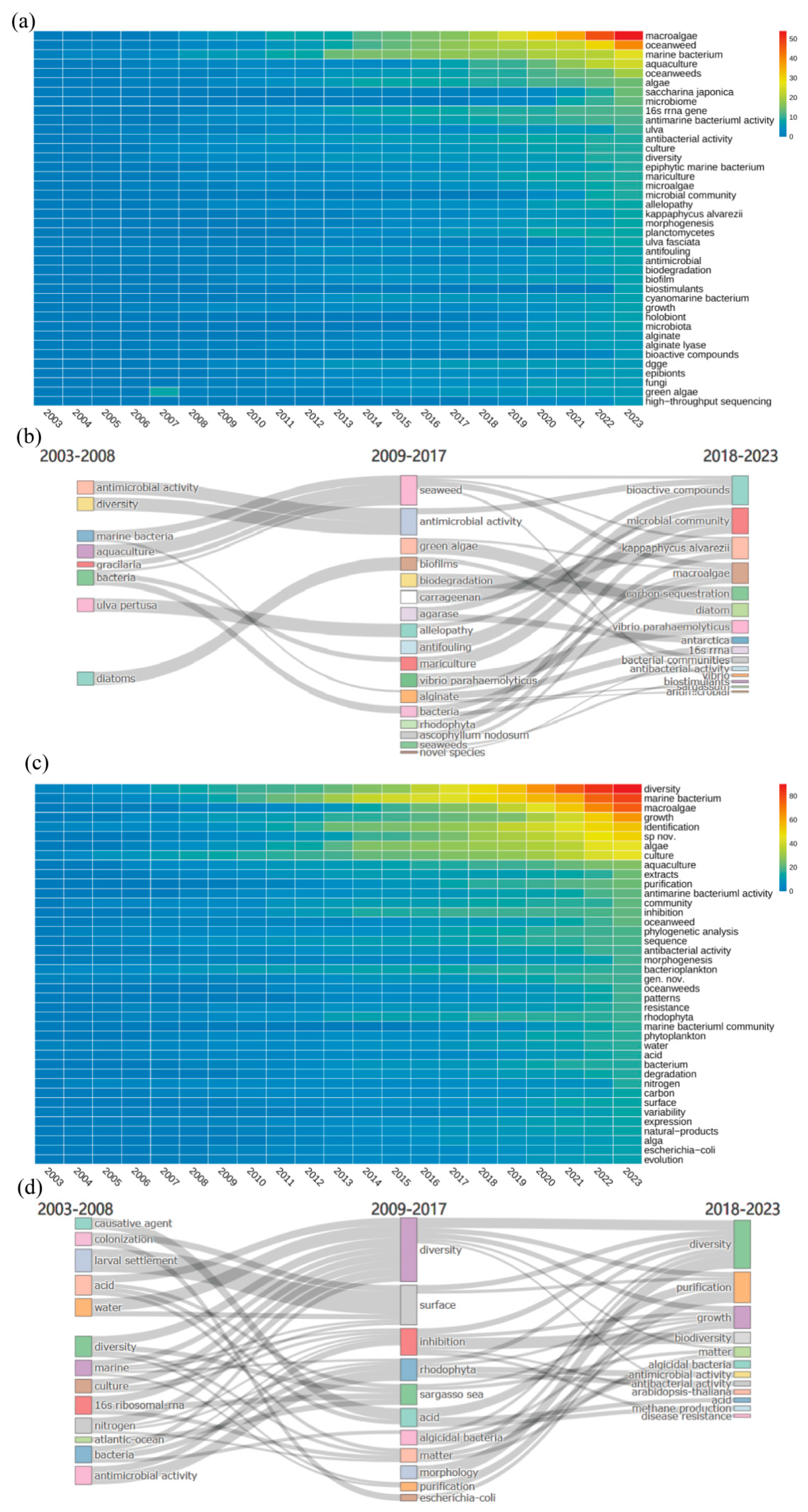
Figure 8.
Thematic map of 2003-2023. The thematic map categorizes research themes into four distinct areas based on their development density and centrality. Niche themes indicate specialized areas of research with growth potential. Motor themes represent the current and popular research direction of the research field. Basic themes show a sustained research interest. Emerging or Declining themes demonstrate newly emerging or declining topics and potential future significance.
Figure 8.
Thematic map of 2003-2023. The thematic map categorizes research themes into four distinct areas based on their development density and centrality. Niche themes indicate specialized areas of research with growth potential. Motor themes represent the current and popular research direction of the research field. Basic themes show a sustained research interest. Emerging or Declining themes demonstrate newly emerging or declining topics and potential future significance.
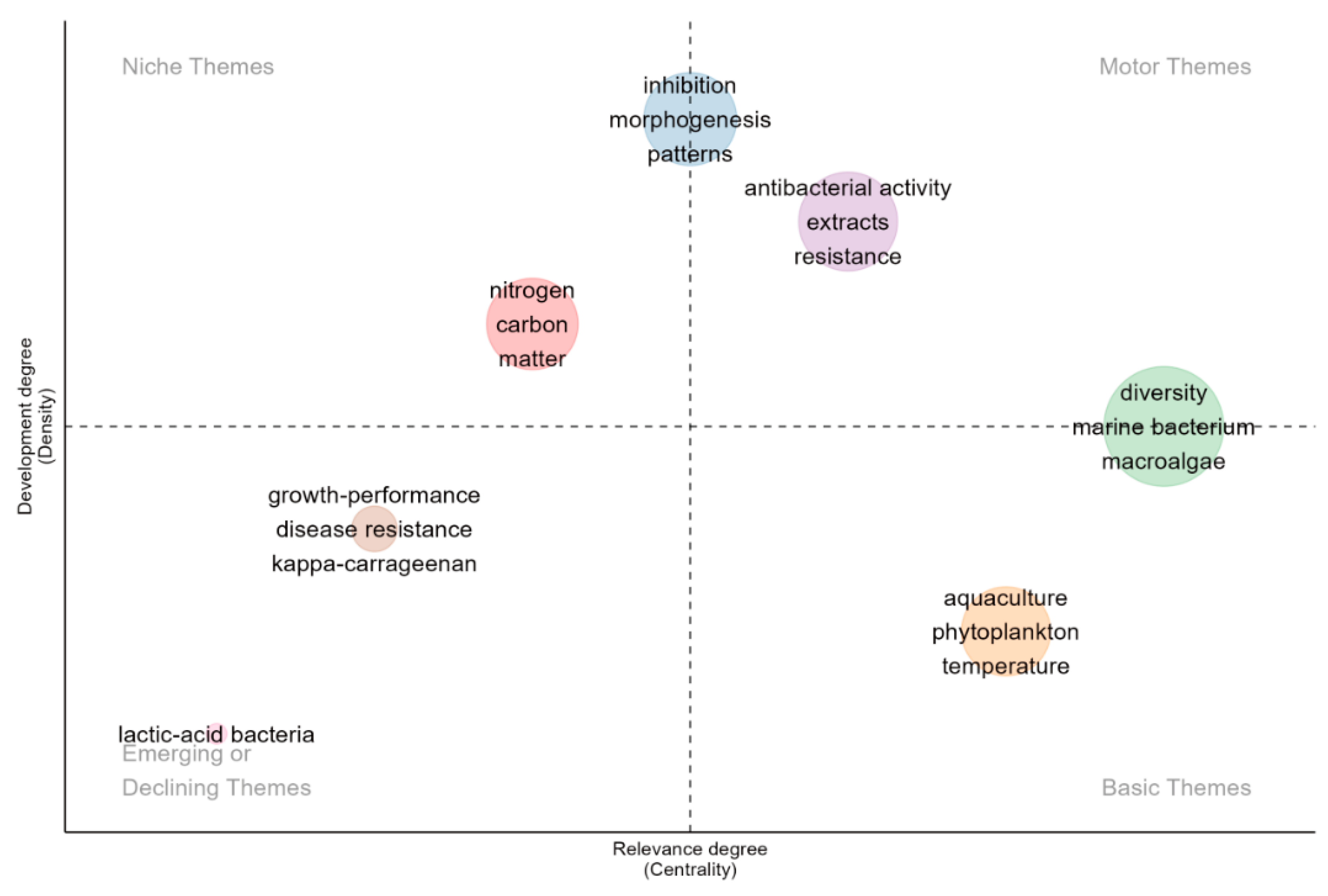

Table 2.
Literature data by country. Single Country Publications (SCP) denoted as the number of publications co-authored by authors of the same nationality; Multiple Country Publications (MCP) denoted as the number of co-authored publications with authors from other countries; Country's publications in the total number of publications (Freq.); Multiple Country Publications Ratio (MCP Ratio) denoted as the proportion of publications by countries in cooperation with other countries, serving as a key metric to assess the level of engagement in international scientific cooperation; Total Citations (TC); Average Article Citations (AAC). This table lists the top 20 countries by the number of publications.
Table 2.
Literature data by country. Single Country Publications (SCP) denoted as the number of publications co-authored by authors of the same nationality; Multiple Country Publications (MCP) denoted as the number of co-authored publications with authors from other countries; Country's publications in the total number of publications (Freq.); Multiple Country Publications Ratio (MCP Ratio) denoted as the proportion of publications by countries in cooperation with other countries, serving as a key metric to assess the level of engagement in international scientific cooperation; Total Citations (TC); Average Article Citations (AAC). This table lists the top 20 countries by the number of publications.
| Country | Articles | SCP | MCP | Freq. | MCP Ratio | TC | AAC |
|---|---|---|---|---|---|---|---|
| China | 125 | 101 | 24 | 0.205 | 0.192 | 2117 | 16.9 |
| USA | 73 | 61 | 12 | 0.12 | 0.164 | 7043 | 96.5 |
| India | 45 | 41 | 4 | 0.074 | 0.089 | 756 | 16.8 |
| Germany | 41 | 19 | 22 | 0.067 | 0.537 | 1660 | 40.5 |
| Japan | 37 | 34 | 3 | 0.061 | 0.081 | 815 | 22 |
| Korea | 25 | 19 | 6 | 0.041 | 0.24 | 581 | 23.2 |
| France | 24 | 16 | 8 | 0.039 | 0.333 | 505 | 21 |
| Australia | 21 | 12 | 9 | 0.034 | 0.429 | 1263 | 60.1 |
| United Kingdom | 20 | 10 | 10 | 0.033 | 0.5 | 774 | 38.7 |
| Italy | 14 | 10 | 4 | 0.023 | 0.286 | 311 | 22.2 |
| Portugal | 14 | 7 | 7 | 0.023 | 0.5 | 529 | 37.8 |
| Spain | 14 | 7 | 7 | 0.023 | 0.5 | 624 | 44.6 |
| Chile | 13 | 8 | 5 | 0.021 | 0.385 | 301 | 23.2 |
| Belgium | 12 | 3 | 9 | 0.02 | 0.75 | 300 | 25 |
| Brazil | 12 | 10 | 2 | 0.02 | 0.167 | 312 | 26 |
| Malaysia | 12 | 6 | 6 | 0.02 | 0.5 | 224 | 18.7 |
| Canada | 11 | 7 | 4 | 0.018 | 0.364 | 309 | 28.1 |
| Denmark | 10 | 5 | 5 | 0.016 | 0.5 | 549 | 54.9 |
| Mexico | 9 | 7 | 2 | 0.015 | 0.222 | 82 | 9.1 |
| Russia | 6 | 4 | 2 | 0.01 | 0.333 | 311 | 51.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



