
Submitted:
31 March 2025
Posted:
01 April 2025
You are already at the latest version
Abstract
Modern agriculture faces critical challenges due to increasing soil degradation resulting from overcultivation and excessive chemical use. These factors have led to declining agricultural productivity, reduced soil fertility, and a significant loss of microbial biodiversity. This review examines the role of plant growth-promoting bacteria (PGPB) as symbiotic microorganisms in restoring degraded soils. Key molecular and physiological mechanisms that enhance soil structure, improve water retention, and increase the bioavailability of essential nutrients for plants are explored. Additionally, the capacity of halotolerant bacteria to mitigate salt stress—an urgent issue in soils affected by salinization due to organic matter decomposition and irrigation practices—is analyzed. The role of plant growth regulators (PGRs) in both plant adaptation to adverse conditions and the stimulation of plant growth and development is also highlighted. Finally, current limitations in understanding biotic and abiotic stress resistance mechanisms in plants treated with growth-promoting strains are discussed, emphasizing the need for further research to optimize soil restoration strategies. This work provides a comprehensive overview of microorganism-based approaches for fostering more sustainable and resilient agriculture.
Keywords:
abiotic and biotic stress
; agricultural sustainability
; Plant Growth Promoting Bacteria (PGPB)
; restoration of degraded soils
; soil microbiota
; Plant Growth Regulators (PGRs)
1. Soil Degradation Challenges Stemming from Excessive Agricultural Cultivation.
Food security is a fundamental pillar of global health, human well-being, and economic stability [1]. Ensuring the availability, accessibility, and quality of food is essential for sustaining global populations [2]. However, while agriculture is vital for humanity, unsustainable and excessive practices can have severe environmental consequences. Soil degradation resulting from intensive agricultural cultivation is a global concern, affecting millions of hectares worldwide [3]. One of the primary consequences of overcultivation is nutrient depletion, which compromises soil fertility and long-term productivity [4,5,6]. This decline reduces the soil’s capacity to support healthy crops and maintain balanced ecosystems [7]. The intensive use of fertilizers, often required to sustain high yields, exacerbates this issue by promoting nutrient leaching, particularly of nitrogen and phosphorus, into deeper soil layers or groundwater, leading to water contamination [8]. Additionally, this process contributes to salt accumulation and soil acidification [9], further diminishing soil fertility and increasing dependence on external inputs, thereby perpetuating the cycle of degradation [10,11].
The widespread practice of intensive monoculture aggravates this situation by selectively depleting specific nutrients required by a single crop species, resulting in nutritional imbalances and progressively declining yields [12,13,14,15,16]. This problem is further exacerbated by excessive tillage [17] and the use of heavy machinery [18], which compact the soil and form a dense, impermeable layer known as a plow pan [19]. Soil compaction restricts root penetration and impedes water infiltration, reducing soil porosity and its capacity to retain nutrients and oxygen, both of which are essential for plant growth [20]. Furthermore, compacted soils are more susceptible to erosion caused by wind and water, which depletes the fertile topsoil, degrades soil structure, and negatively impacts microbial biodiversity and ecosystem functionality [21,22].
Soil degradation reduces crop resilience to diseases and environmental stress [23] and facilitates the runoff of nutrients and organic matter into water bodies, leading to pollution and eutrophication [24]. Excessive tillage and the use of disc plows disrupt soil aggregates, which are essential for maintaining soil porosity, thereby promoting the formation of compacted subsurface layers that can persist for years [25]. To reduce these negative impacts, is essential implementing sustainable soil management strategies such as diversifying crops, utilizing cover crops, and incorporating organic matter. These practices can be effective, as they help restore soil structure and reduce compaction. Additionally, limiting machinery traffic through direct seeding and the establishment of designated access roads can contribute to the long-term preservation of soil health [26,27,28].
Soil harbors an extraordinary diversity of microorganisms, including bacteria, fungi, actinomycetes, and protozoa, which play fundamental roles in maintaining soil health and fertility [13,15]. However, the loss of microbial biodiversity due to excessive agricultural intensification poses a significant challenge with severe repercussions for soil functionality and the sustainability of agroecosystems [29]. The extensive use of pesticides and herbicides drastically reduces populations of beneficial microorganisms, such as nitrogen-fixing bacteria, mycorrhizal fungi, and actinomycetes, impairing key ecological functions, including organic matter decomposition, nutrient recycling, and plant health maintenance [30]. This decline in microbial biodiversity negatively impacts crop yields [31] and increases dependence on external inputs, such as fertilizers and pesticides, with long-term economic and environmental consequences [32].
The indiscriminate use of agrochemicals, combined with soil physical degradation, pollution, wildfires, and droughts, exerts direct pressures on soil microbial communities, significantly reducing their population density and diversity (Figure 1"A"). Additionally, pesticides and herbicides can disrupt microbial ecosystems by selectively promoting the proliferation of certain species at the expense of native ones, ultimately compromising soil functionality [33]. The widespread adoption of intensive monocultures further exacerbates this issue by creating a selective environment that favors a limited subset of microbial species, reducing overall diversity and weakening soil resilience to pathogens and environmental stressors [34]. These factors, by destroying natural habitats and disrupting critical biogeochemical cycles, restrict microbial activity, diminish soil porosity, and hinder the movement of air and water within the soil matrix [35].
To address these challenges, adopting sustainable agricultural practices is essential to restore soil structure, reduce compaction, and enhance microbial activity. Additionally, the application of plant growth-promoting bacteria (PGPB) plays a pivotal role in conserving and restoring soil microbial diversity. These bacteria contribute to soil health and resilience by facilitating phosphorus solubilization, nitrogen fixation, and the production of plant growth regulators (PGRs) that stimulate plant growth (Figure 1B). Moreover, PGPB help mitigate environmental stressors, such as salinity and drought, by producing osmoprotective compounds and enzymes that alleviate ethylene-induced stress in plants [36].
This integrated approach fosters the sustainability of agricultural systems while ensuring the long-term health of both soils and the environment. By combining responsible agricultural practices with PGPB applications, soil fertility can be restored, crop productivity enhanced, and reliance on chemical inputs reduced, ultimately contributing to a more sustainable and ecologically balanced agricultural system [37,38].
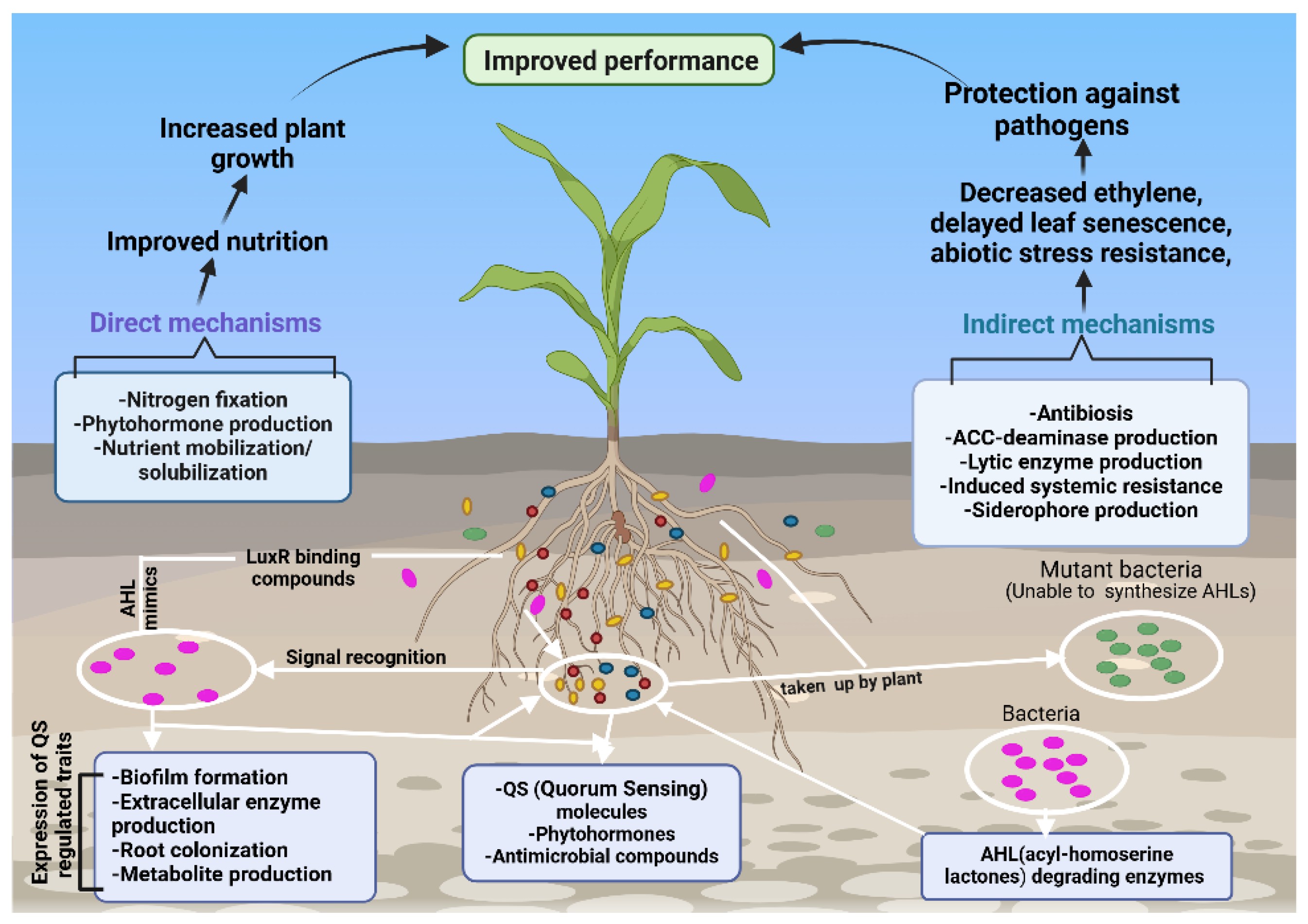
2. Direct and Indirect Mechanisms of Plant Growth-Promoting Bacteria
Plants have coevolved with a diverse microbiota that plays a crucial role in their growth and health [39]. These microbes form symbiotic relationships with plant tissues, offering multiple advantages in return for reduced carbon and other metabolites provided by the host plant [40]. This symbiosis has existed since the earliest stages of terrestrial plant evolution, enabling plants to overcome challenges such as nutrient acquisition, abiotic stress, and pathogen defense [41,42,43] (Figure 2).
Considerable advancements have been achieved in understanding the composition and dynamics of plant microbiota, along with the functional capabilities of strains isolated from these communities. For instance, PGPB have been identified in various plant species, including those of both agricultural [44] and ecological significance [45,46]. The bioactive compounds secreted by these bacteria function as biostimulants and play a critical role in modulating plant stress responses. Moreover, PGPB facilitates nutrient uptake and assimilation, enhance soil structure, and generate extracellular molecules like plant growth regulators (PGRs), secondary metabolites, antibiotics, and signaling compounds, all of which support plant development and resilience. [47].
Considering the potential of these bacterial strains and the actual challenges in crop production, it is crucial to incorporate microbial innovations into agricultural practices. The following sections explore the primary mechanisms by which PGPB enhance plant growth, such as biological nitrogen fixation, phosphate solubilization, siderophore production, and the synthesis of plant growth regulators (PGRs).
Figure 2.
Direct and indirect mechanisms of action of PGPB in plants. Symbiotic bacteria within the plant-associated microbiota play a crucial role in enhancing plant growth and resilience through various direct and indirect mechanisms.
Figure 2.
Direct and indirect mechanisms of action of PGPB in plants. Symbiotic bacteria within the plant-associated microbiota play a crucial role in enhancing plant growth and resilience through various direct and indirect mechanisms.
2.1. Indirect Mechanisms
2.1.1. Siderophores Production
Iron plays a fundamental role in the photosynthetic system of plants, as it is an essential component of chlorophyll, which absorbs light, and is involved in various biosynthetic pathways. However, the amount of soluble iron available in soil is often insufficient to optimize crop yields [37]. To address this limitation, bacteria, fungi, and plants release specific low-molecular-weight substances called siderophores, which bind to iron in the soil [48].
Siderophores are primarily produced by bacteria to alleviate stress caused by iron deficiency, thereby promoting plant growth. These bacteria possess surface proteins that regulate the uptake of ferric iron (Fe³⁺), ensuring its availability for metabolic processes [49]. Notably, siderophores produced by PGPB exhibit exceptionally high affinity for Fe³⁺. Once the iron-siderophore complex is formed, it is recognized by specific receptors on bacterial or plant cell surfaces, internalized, and subsequently reduced to ferrous iron (Fe²⁺) or released upon siderophore degradation, making iron available for metabolic use [48].
Importantly, siderophores produced by PGPB have a significantly higher affinity for iron than those synthesized by plants or fungi, enabling them to sequester large amounts of this nutrient [50]. This trait not only facilitates iron acquisition under conditions of scarcity but also plays a crucial role in plant protection. By releasing siderophores with exceptionally high iron-binding affinity, PGPB effectively limit iron availability for pathogenic fungi and bacteria, thereby restricting their growth [51]. The effectiveness of this biocontrol mechanism relies on the fact that PGPB siderophores outcompete fungal siderophores in iron chelation [52].
Structurally, microbial siderophores commonly contain functional groups such as hydroxamates and catecholates, as well as carboxylates, citrates, or ethylenediamine moieties, which may coexist within the same molecule [53]. Hydroxamate-type siderophores are predominant in fungi, whereas catecholate siderophores, which exhibit stronger iron-binding capabilities, are more prevalent in bacteria [54]. In contrast, plant-derived siderophores, such as mugineic and avenic acids, belong to the family of iminocarboxylic acids substituted with linear hydroxyl and amino groups. These compounds exhibit higher iron-chelating efficiency compared to bacterial siderophores [55]. Additionally, bacterial siderophores can chelate other trivalent and divalent metal ions, albeit with considerably lower affinity [56].
2.1.2. Enzymatic Mechanisms in Indirect Plant Defense.
Several compounds play key roles in indirect mechanisms, including lytic enzymes produced by PGPB. These enzymes exhibit antimicrobial activity, acting as a defense against bacterial pathogens [57]. Similarly, proteases contribute significantly to plant protection by degrading pathogen-derived proteins. These enzymes, produced by species such as Bacillus clausii and Bacillus lentus, are classified as alkaline, acidic, or neutral based on their optimal pH [58].
Another indirect mechanism that promotes root growth involves catalase-positive bacteria, which protect plant roots from oxidative damage caused by hydrogen peroxide. This process enhances plant resilience under oxidative stress conditions [59]. Notable bacterial species associated with this function include Bacillus insolitus, Bacillus pasteurii, Bacillus laterosporus, and Staphylococcus aureus [60].
Hydrogen cyanide (HCN) is another relevant compound due to its volatility and high toxicity, which interfere with cellular respiration [61]. Synthesized by rhizospheric bacteria, HCN inhibits the growth of pathogenic fungi, nematodes, insects, and termites. Additionally, it functions as a natural herbicide by colonizing the roots of undesired plants, suppressing their growth without affecting the host plant [62].
Enzymes such as amylases also play a crucial role in plant protection. Amylases, primarily classified as α-amylases, β-amylases, and γ-amylases, are produced by bacteria, actinobacteria and fungi and are involved in the degradation of phytopathogen cell walls. α-Amylases, in particular, are commonly found in endophytic bacteria associated with medicinal plants and agricultural crops [63]. Bacterial species of the genus Bacillus, such as Bacillus licheniformis, Bacillus stearothermophilus, and Geobacillus bacterium, are notable for their high production of these enzymes [64].
Finally, ureases are essential enzymes in soil ecosystems, catalyzing the hydrolysis of urea into ammonium and nitrate—two nitrogen forms readily assimilated by plants as a vital nutrient source [65]. Additionally, ureolytic bacteria contribute to calcite precipitation by increasing pH and generating carbonate ions, a process with applications in soil nutrient enhancement, concrete crack repair, and biomineralization [66].
2.2. Direct Mechanisms
2.2.1. Plant Growth Regulators and Their Role in Plant Growth and Signaling
PGRs are essential chemical compounds that enable plants to adapt and respond to environmental conditions [67]. Both plants and microorganisms produce PGRs such as cytokinins and auxins. However, research on cytokinin biosynthesis remains limited due to the structural diversity of these compounds and their typically low concentrations, which make detection and analysis challenging [68]. Unlike animal hormones, plant hormones are not synthesized in specialized organs but can be produced in virtually any plant tissue. Currently, plant development is known to be regulated by five major PGRs groups: auxins (AUXs), gibberellins (GBRs), cytokinins (CTKs), ethylene (ETH), and abscisic acid (ABA) [69].
In addition, other signaling molecules with hormone-like functions have been identified, playing essential roles in herbivore defense and disease resistance. These include salicylic acid (SA), nitric oxide (NO), jasmonic acid (JA), brassinosteroids (BRs), strigolactones (SLs) and systemin peptides [70]. Although NO is recognized as a key signaling molecule and an intermediary in metabolic processes, it is not yet classified as a PGR due to its inorganic nature [71]. Research on gibberellic acid also remains scarce, with little progress in the past two decades [72]. Among all PGRs, indole-3-acetic acid (IAA) is the most extensively studied and is considered a key factor in plant growth promotion by plant growth-promoting bacteria (PGPB) [73]. Both plants and microbes synthesize IAA through different pathways, one of which is tryptophan (Trp)-dependent. Microbial IAA production varies according to physiological factors such as temperature, pH and the availability of carbon and nitrogen sources [74].
Most PGRs in plants are thought to undergo long-distance translocation, which is crucial for inter-organ chemical communication. This transport mainly takes place through the phloem and xylem. For instance, buds produce growth hormones like auxins, which are quickly moved to the roots via the phloem, while roots produce cytokinins that are transported to the buds through the xylem. Additionally, some hormones, such as weak acids, can diffuse across cell membranes in their protonated form [75]. As research advances and new evidence emerges, the list of plant growth regulators and signaling molecules is expected to expand, further refining our understanding of plant biochemical pathways.
2.2.2. Contribution of Microbial Activity to Nutrient Solubilization and Plant Growth.
Phosphorus (P) is a crucial nutrient for plant growth, and its deficiency can significantly hinder development [76]. Some bacteria possess the ability to transform inorganic phosphorus compounds like tricalcium phosphate, hydroxyapatite, and rock phosphate into soluble forms by releasing organic acids such as citric acid and gluconic acid. These acids bind to phosphate cations through their hydroxyl and carboxyl groups, improving the bioavailability of phosphorus for plants [77].
Zinc acts as a crucial cofactor for many enzymes required for plant growth. Some bacteria facilitate zinc solubilization by producing siderophores, which enhance the release of soluble zinc ions [78]. The optimal zinc concentration in plants ranges from 30 to 100 mg/kg, and its deficiency can lead to stunted growth and the formation of necrotic spots [79].
Similarly, while potassium is naturally present in the soil, it is not always available in a form that plants can readily absorb. PGPBs enhance potassium solubilization by producing organic acids such as citric, oxalic, and tartaric acids [80]. This macronutrient is essential for the activation of over 60 enzymes involved in plant metabolism, including those regulating photosynthesis, protein synthesis, and starch metabolism. Additionally, potassium is essential for stomatal regulation, which affects water use efficiency and enhances drought resistance. Higher potassium concentrations also improve plant disease resistance, enhance the quality of cotton fibers, and extend the shelf life of fruits and vegetables [81].
3. Physiological Mechanisms of Plant-Microorganism Interaction
The beneficial interaction between plants and bacteria occurs in a dynamic environment, where molecular and biochemical signals play a fundamental role [82]. This symbiotic relationship relies on a continuous exchange of signals that mediate communication between plants and free-living bacteria, leading to mutual benefits for both organisms [83]. Among these bacteria, PGPB enhance plant development through both direct and indirect mechanisms, as previously discussed. This chapter examines the physiological mechanisms underlying plant-microorganism interactions, with the aim of elucidating the modes of action through which bacteria exert their beneficial effects.
3.1. Bacterial Contribution to Plant Nutrient Acquisition
3.1.1. Nitrogen
According to Liebig's law of the minimum, nitrogen (N) is usually the most limiting nutrient for plant growth, followed by phosphorus (P) [84]. Nitrogen is an essential element for all living organisms, as it is a key component of nucleic acids and proteins [85]. While atmospheric nitrogen (N₂) is abundant, most organisms cannot directly assimilate it. Only a specific group of bacteria and archaea, known as diazotrophs, have developed the ability to fix N₂, converting it into ammonia (NH₃) through a process called biological nitrogen fixation (BNF). This process is facilitated by nitrogenase, an enzyme complex that is sensitive to oxygen [86].
Diazotrophic bacteria do not release ammonia freely, as nitrogen assimilation and fixation are tightly coupled and regulated by intricate control mechanisms [87]. These regulatory pathways include, transcriptional regulators, post-translational protein-modifying enzymes, and PII signal transduction proteins that control the expression of nitrogen metabolism genes [88,89,90,91]. In diazotrophic proteobacteria, these mechanisms interact with the nitrogen-fixing gene regulator (NifA), ensuring that nitrogenase expression is adjusted according to N demand [92]. The ammonia produced during nitrogen fixation diffuses across the bacterial cell membrane and is actively recovered by the ammonium transporter (AmtB), despite the high energetic cost of this process [92]. BNF is an energy-intensive process, requiring approximately 16 moles of ATP per mole of fixed N₂ [94].
Nitrogenase plays a critical role in nitrogen fixation, a highly conserved evolutionary process [94]. All nitrogen-fixing organisms possess the nif genes, which encode the nitrogenase complex. Additionally, key genes involved in nutrient cycling, such as cbbL, nifH, amoA, and apsA, highlight the significance of nitrogen fixation in microbial metabolism and soil health [96]. Nitrogenase consists of two metalloproteins: molybdenum-iron nitrogenase (MoFe) and iron nitrogenase reductase (Fe) [97]. MoFe nitrogenase (NifDK) functions as an oxygen-sensitive dinitrogenase, while the Fe protein (NifH) facilitates electron transfer to the MoFe protein using two molecules of MgATP per electron [98]. The MoFe₇S₉-homocitrate cofactor in the MoFe protein serves as the active site for N₂ reduction, as well as for other substrates such as acetylene and protons [99].
The ATP-hydrolyzing nitrogenase complex consists of two primary proteins: the γ₂ homodimer of dinitrogenase reductase (NifH) and the α₂β₂ heterotetramer of dinitrogenase (where NifD represents the α subunit and NifK the β subunit). The MoFe₇S₉ metal cluster, located in the α subunit, is essential for dinitrogen reduction [100]. In some microorganisms, such as Azotobacter vinelandii and Methanosarcina acetivorans, molybdenum (Mo) can be replaced by iron (Fe) or vanadium (V), giving rise to the alternative nitrogenase variants Anf and Vnf, respectively. Among these, FeMo nitrogenase is the most efficient at reducing dinitrogen to ammonia, followed by Anf and Vnf [101]. Since nitrogenase is highly oxygen-sensitive, nitrogen-fixing organisms develop specialized mechanisms, such as oxygen barriers around infected cells, to maintain a microaerobic environment conducive to nitrogen fixation [102].
Bacterial nitrogen assimilation occurs through the production of glutamate from ammonium and 2-oxoglutarate via two alternative pathways: the glutamine synthetase-glutamate synthase (GS-GOGAT) pathway and the glutamate dehydrogenase (GDH) pathway. The GDH enzyme catalyzes the reductive amination of 2-oxoglutarate to glutamate using NADPH. This pathway is mainly active under conditions of high ammonium and low carbon/energy availability due to its high Km (~1 mM) for ammonium and its lack of ATP dependence [105].
The GS-GOGAT pathway, on the other hand is more prevalent in bacteria, and functions under low-ammonium and high-carbon/energy conditions. This pathway involves the activity of glutamine synthetase (GS), an enzyme with a much lower Km for ammonium that requires ATP to convert glutamate into glutamine. Subsequently, glutamate synthase (GOGAT) transfers the amide group from glutamine to 2-oxoglutarate in a reaction that requires NADPH, resulting in the production of two molecules of glutamate [106] (Figure 3). This pathway represents the primary route for ammonia assimilation in bacteria [103].
Glutamate plays a critical role as a nitrogen transporter in plants, facilitating the translocation of this element from the roots through the xylem to various plant tissues [107]. Once glutamate-derived amino acids reach their target sites, they are efficiently utilized in seed germination, protein synthesis, nucleic acid biosynthesis, and the production of other essential metabolites required for growth and development [108]. Additionally, glutamate-derived metabolites regulate key metabolic pathways, including photosynthesis, respiration, and responses to environmental stress, ensuring proper physiological function and adaptation to changing environmental conditions [104]. The integration of nitrogen into diverse biological molecules through glutamate highlights the complexity and efficiency of plant nitrogen metabolism, enabling plants to optimize nitrogen utilization for sustained growth and productivity.
3.1.2. Phosphorus
After N, P is the most limiting essential mineral element for plant growth, as it is only absorbed in its soluble monobasic (H₂PO₄⁻) or dibasic (HPO₄²⁻) forms [109]. In soils, (P) is predominantly found in inorganic forms, either adsorbed onto soil mineral surfaces or in poorly available precipitates. It is also present in organic forms, where it is incorporated into biomass or associated with soil organic matter [86].
P is one of the nineteen essential elements for plant life, playing a vital role in energy uptake, storage, and transfer. It is also a key structural component of important biomolecules like DNA, RNA, and phospholipids in both plant and animal cells [76]. P is involved in fundamental physiological processes, including photosynthesis, root development, stem strengthening, flower and seed formation, crop maturation, energy metabolism, cell division and expansion, nitrogen fixation in legumes, disease resistance, starch synthesis, and genetic information transfer [110]. Additionally, adequate P availability is crucial for the formation of reproductive primordia during the early stages of plant development [111]. Therefore, P is indispensable for virtually all physiological processes in plants.
Within the soil-plant system, approximately 90% of total phosphorus is found in the soil, while less than 10% remains available outside of it. However, only a small portion of soil-bound (P) is directly available to plants, as over 90% is strongly attached to soil particles and inorganic minerals like apatite, hydroxyapatite, and oxyapatite. Furthermore, phosphorus is found in organic forms, such as inositol phosphates (soil phytate), phosphomonoesters, and phosphotriesters, which need to undergo mineralization before they become bioavailable [112].
Phosphate-solubilizing bacteria (PSBs) are key members of the plant microbiome that facilitate the conversion of insoluble organic and inorganic phosphate compounds into plant-available forms, particularly under phosphorus-deficient conditions [113]. This microbial activity is crucial for enhancing soil P availability, ultimately promoting plant growth and development [86].
The primary mechanism by which PSBs solubilize phosphate is through the acidification of the surrounding environment via organic acid production or proton release. This decrease in pH increases P solubility [114]. In alkaline soils, phosphate tends to precipitate as calcium phosphates, such as fluorapatite and francolite, rendering it insoluble [115]. As soil pH increases, the divalent (HPO₄²⁻) and trivalent (H₂PO₄⁻) forms of inorganic phosphorus become more prevalent. PSBs release various organic acids as metabolic byproducts, primarily through oxidative respiration or fermentation, particularly when utilizing glucose as a carbon source [114]. The composition and concentration of these acids can differ, and the efficiency of phosphate solubilization depends on the strength and type of acids produced. Di- and tricarboxylic acids are generally more effective than monobasic and aromatic acids. Organic acids that play a role in phosphate solubilization include gluconic, lactic, citric, isovaleric, succinic, glycolic, oxalic, formic, 2-ketogluconic, and acetic acids [112,116].
Among these, gluconic acid and 2-ketogluconic acid are the most commonly reported mineral phosphate solubilizers [117]. Gluconic acid is primarily produced by Pseudomonas spp. [118,119] and Burkholderia spp. [120,121], whereas 2-ketogluconic acid is produced by Nguyenibacter sp. [122] and Rhizobium tropici [123]. Additionally, Priestia megaterium and Bacillus velezensis produce lactic, acetic, and citric acids, while Enterobacter sp. synthesizes acetic acid [119,124]. Gram-negative bacteria have been found to be more efficient dissolving mineral phosphates because their ability to release a wide range of organic acids into the surrounding soil environment [125].
Mineralization plays a crucial role in soil ecosystems, as it is driven by the decomposition of plant and animal residues, which contain significant amounts of organic phosphorus compounds, such as sugar phosphates, nucleic acids, phospholipids, phytic acid, phosphonates and polyphosphates. [126]. The mineralization and immobilization of organic P in soil play a fundamental role in its biogeochemical cycle, particularly in agricultural lands and degraded soils. Through mineralization, PSBs release inorganic phosphate from these complex organic compounds, thereby increasing its availability to plants [127].
PSBs facilitate the mineralization of soil organic P by producing phosphatases—enzymes that hydrolyze organic phosphate compounds, liberating inorganic P in a bioavailable form for plant uptake. The production of these enzymes has been reported in both Gram-positive bacteria, such as those of the genus Bacillus [128], and Gram-negative bacteria, including Burkholderia sp., Gluconacetobacter sp. [120], and Pseudomonas sp. [128]. Additionally, fungi of the genera Aspergillus and Penicillium have also been shown to produce phosphatases [129].
Phytases, a specific class of phosphatases, differ in their cellular localization between Gram-negative and Gram-positive bacteria. In Gram-negative bacteria, phytases are intracellular or periplasmic proteins [130], whereas in Gram-positive bacteria, they are primarily extracellular enzymes [131]. Two main types of phytases have been identified: acidic and alkaline [132]. Acidic phytases, with an optimal pH range of 2.5–5.5, belong to the histidine acid phosphatase family and are characterized by a highly conserved motif near the N-terminal region [133]. These enzymes do not require cofactors for optimal activity and are typically produced by Gram-negative bacteria and fungi. In contrast, alkaline phytases, which exhibit optimal activity at pH 6–8, are considered a distinct class due to their high specificity for phytic acid and their dependence on calcium ions for enzymatic function [132].
Another mechanism by which PSBs facilitate phosphorus solubilization is chelation. This process involves the production of organic and inorganic acids that dissolve insoluble phosphate compounds in the soil by chelating metal cations and competing with phosphate for adsorption sites [126]. These acids bind to insoluble aluminum and iron oxides, stabilizing them and forming chelates [134]. Additionally, their hydroxyl and carboxyl functional groups interact with phosphate-associated cations, converting phosphorus into soluble forms that plants can absorb [135]. Reported inorganic acids produced by PSBs include carbonic, hydrogen sulfide, nitric, and hydrochloric acid [136].
3.2. Bacterial Modulation of Plant Hormonal Pathways and Signaling Mechanisms
3.2.1. Plant Growth Regulators
PGRs are essential molecules that govern plant growth and development by stimulating both the root and aerial systems [137]. Beyond their role in growth, these regulators function as molecular signals in response to biotic factors that may limit plant development or even become lethal if not properly regulated [138]. Rhizospheric bacteria have been documented to secrete hormones that enhance nutrient absorption in roots or modulate the plant’s hormonal balance, thereby promoting growth and improving stress responses [139,140].
Indole-3-acetic acid (IAA) is the most widely studied and prevalent natural auxin produced by plant growth-promoting bacteria (PGPB). It plays a central role in regulating various plant developmental processes, including vascular tissue differentiation, cell elongation, apical dominance, lateral root initiation, and fruit ripening [141]. In addition to its structural roles, IAA acts as an important signaling molecule that regulates several aspects of plant development [142]. Among its primary functions are the regulation of cell division, expansion, and differentiation, as well as the modulation of gene expression necessary for plant growth and adaptation to environmental conditions [143]. For instance, IAA promotes organ formation and modulates responses to abiotic stresses such as salinity, drought, nutrient deficiencies and extreme temperatures [144,145]. These regulatory effects are mediated by the activation of specific genes that enable plants to adjust their development under adverse conditions.
IAA is also fundamental to plant cell signaling, controlling multiple growth and developmental processes through complex signal transduction mechanisms. One of its key roles is promoting cell expansion by activating protein synthesis [146]. In roots, IAA stimulates cell elongation in the elongation zone, facilitating root system development, whereas in stems, it regulates cell elongation, contributing to vertical growth and branch formation [147]. Another crucial function of IAA is its involvement in apical dominance, a phenomenon in which the growth of the terminal shoot suppresses the development of lateral shoots. This regulation is mediated by a negative feedback mechanism: IAA synthesized in the apical meristem is transported downward along the stem, inhibiting the outgrowth of lateral shoots from axillary buds [148]. Furthermore, IAA accumulates in the rhizosphere in response to internal and external signals, inducing the formation of lateral and adventitious roots through the activation of specific genes that promote cell division and differentiation in root initiation zones [149]. Additionally, IAA plays a critical role in ovary development, fruit ripening [150], and the differentiation of floral tissues, including the determination of floral structure and reproductive organ formation [151].
Another key function of IAA is the regulation of gene expression, influencing plant development, growth, and responses to environmental stress [152]. Its mechanism of action begins with the binding of IAA to specific receptors, such as Transport Inhibitor Response 1 (TIR1) and Auxin Signaling F-Box (AFB) proteins, which are components of the Skp1-Cullin-F-box (SCF) ubiquitin ligase complex. This interaction triggers a series of molecular events in which the SCF complex targets auxin repressor proteins (Aux/IAA) for ubiquitination and subsequent degradation by the 26S proteasome. The degradation of Aux/IAA repressors releases auxin response factors (ARFs), which then control the transcription of auxin-responsive genes [153]. ARFs are transcription factors that bind to specific DNA sequences, called auxin response elements (AuxREs), found in the promoters of target genes. Depending on their function, ARFs can either activate or repress gene transcription [154,155]. Activating ARFs promote gene expression, whereas repressive ARFs inhibit it [156]. Moreover, ARFs do not function in isolation; they interact with other transcription factors and co-regulatory proteins to fine-tune gene expression in response to plant physiological needs and environmental cues [152]. This tightly regulated system ensures a precise response to fluctuations in IAA levels, optimizing plant development and adaptation.
IAA plays a crucial role in plants' response to stress, inducing the expression of genes that increase their resistance to pathogens and improve their adaptation to adverse conditions such as drought, salinity, and extreme temperatures [157,158]. Among the genes regulated by IAA, GH3 genes stand out, which encode enzymes responsible for conjugating IAA with amino acids, which allows the levels of free IAA in the plant to be regulated. Another important group is the SAUR (Small Auxin-Up RNA) genes, which are rapidly activated in response to IAA and are involved in processes such as cell elongation. In addition, the TIR1/AFB genes, which encode auxin receptors, generate a feedback loop that reinforces IAA signaling [159]. These molecular mechanisms allow plants to adjust their growth and development based on internal and external stimuli, ensuring a dynamic and adaptive response to their environment. Research in this field continues to uncover new components and processes that expand our understanding of how IAA regulates gene expression and, ultimately, plant development and adaptation.
IAA plays a crucial role in plant stress responses by inducing the expression of genes that enhance resistance to pathogens and improve adaptation to adverse conditions such as drought, salinity, and extreme temperatures [157,158]. Among the genes regulated by IAA, the GH3 gene family is particularly notable, encoding enzymes responsible for conjugating IAA with amino acids, thereby regulating free IAA levels in the plant. Another important group includes the SAUR (Small Auxin-Up RNA) genes, which are rapidly activated in response to IAA and participate in processes such as cell elongation. Additionally, the TIR1/AFB genes, which encode auxin receptors, establish a feedback loop that reinforces IAA signaling [159]. These molecular mechanisms enable plants to dynamically adjust their growth and development in response to internal and external stimuli, ensuring an adaptive response to their environment. Ongoing research continues to reveal novel components and processes that expand our understanding of how IAA regulates gene expression and, ultimately, plant development and adaptation.
In addition to IAA, other key PGR and development include cytokinins—purine-derived hormones that play a critical role in promoting and maintaining cell division [160]. These hormones play a key role in several differentiation processes, such as callus induction, bud formation, and primary root growth [161]. One of their crucial functions is to maintain totipotent stem cell populations in the shoot and root meristems, which regulate essential developmental processes and influence plant architecture [68]. Due to these functions, cytokinins are essential for plant growth plasticity, ensuring adaptability to different environmental conditions.
Gibberellins constitute the most extensive group of PGRs, with over 100 identified molecules exhibiting varying biological activities [162]. These diterpene-derived hormones share a common chemical structure based on a 19–20 carbon backbone, which is directly related to their function. A key feature of gibberellins is their ability to translocate from the roots to the aerial parts of the plant, where they exert significant effects on growth, flowering, and germination [163]. While gibberellins are essential for plant development, little is known about microorganisms capable of producing them. However, it has been established that symbiotic bacteria, such as rhizobia in legume nodules, synthesize gibberellins, auxins, and cytokinins at low concentrations, primarily during nodule formation and phases of rapid cell division [164].
Abscisic acid (ABA) acts as a stress-responsive signal that is transported through the xylem from the roots to the plant's aerial parts [165]. Upon arrival, it regulates stomatal movement and the activity of the shoot meristem [166]. The intensity of ABA signaling in plants is regulated by several mechanisms, including the activity of PGPB in the rhizosphere. Furthermore, the lateral transport of ABA in the root cortex across apoplastic barriers, pH variations in the leaf apoplast, the activity of β-glucosidases in both the apoplast and the mesophyll cytosol, and their redistribution in the stem contribute significantly to ABA regulation. [167].
Ethylene is another essential PGR that acts as a key regulator of plant responses to biotic interactions [168]. It plays a critical role in plant defense against pathogens and in mediating beneficial interactions with symbiotic microorganisms [169,170,171]. These interactions enable plants to balance disease resistance with the promotion of symbiotic relationships that enhance growth and development [172]. PGPB positively influence ethylene signaling via the acdS gene, which encodes the enzyme ACC deaminase [163]. During plant-microbe interactions, this enzyme metabolizes 1-aminocyclopropane-1-carboxylic acid (ACC), the immediate precursor of ethylene, thereby reducing its availability [173]. A significant portion of ACC synthesized by ACC synthase (ACS) in the roots is exuded into the rhizosphere, where it is taken up by bacteria and degraded by ACC deaminase (Figure 4). This process lowers ACC levels in roots, consequently reducing ethylene biosynthesis [174]. By attenuating ethylene’s inhibitory effects on root development, this mechanism ultimately promotes plant growth [175]. These interactions highlight the critical role of PGPB not only in enhancing nutrient availability but also in modulating hormonal pathways that govern plant development. This underscores the potential of PGPB as biotechnological tools for optimizing plant growth and stress adaptation.
3.2.2. Volatile Organic Compounds
In addition to producing PGRs, PGPB generate volatile organic compounds (VOCs), which are essential for long-distance communication within microbial communities and between microorganisms and plants in the rhizosphere [142]. These small, lipophilic molecules are produced through several metabolic pathways and act as chemical mediators for signal transduction [163]. Both bacteria and fungi produce a broad range of VOCs, such as ketones, terpenoids, alcohols, alkanes, alkenes, and sulfur-containing compounds [176]. Among these, 2-heptanol, 2-undecanone, and pentadecane have been identified as key signaling molecules in plant-microbe interactions [177]. Upon detection by plant roots, these VOCs modulate root system architecture, enhancing the mutual association between roots and PGPBs [178]. This process facilitates bacterial colonization of the rhizosphere, creating a more favorable environment for plant growth.
VOCs perform two primary functions: acting as chemical signals and exhibiting antimicrobial activity. Compounds such as pyrrolnitrin (PRN) and 2,4-diacetylphloroglucinol (DAPG), produced by bacterial genera such as Pseudomonas [179], Butiauxella and Serratia [180], play a crucial role in plant defense against bacterial and fungal pathogens. These molecules not only possess direct antimicrobial properties but also stimulate systemic defense responses in plants, thereby enhancing their resistance mechanisms [181,182]. PRN and DAPG modulate plant hormonal pathways, activating internal defense mechanisms that lead to the production of protective molecules and the reinforcement of structural barriers [183,184]. This process, known as induced systemic resistance (ISR), primes plants for a more efficient and rapid response to future pathogen attacks [185]. Thus, VOCs provide both immediate protection and long-term preparedness against biotic stressors.
Additionally, VOCs influence quorum sensing (QS) systems by interfering with intra- and interspecies microbial communication [186]. They also regulate the expression of genes involved in hormone signaling, thereby affecting root development and enhancing plant defenses against herbivorous insects, primarily through the modulation of auxin pathways [187]. Furthermore, VOCs contribute to plant tolerance to both biotic and abiotic stressors [188,189], as well as processes such as virulence [190] and biofilm formation [191]. The volatile profile of these metabolites is unique to each genotype or species, reflecting their ecological and functional significance in plant-microbe interactions [192].
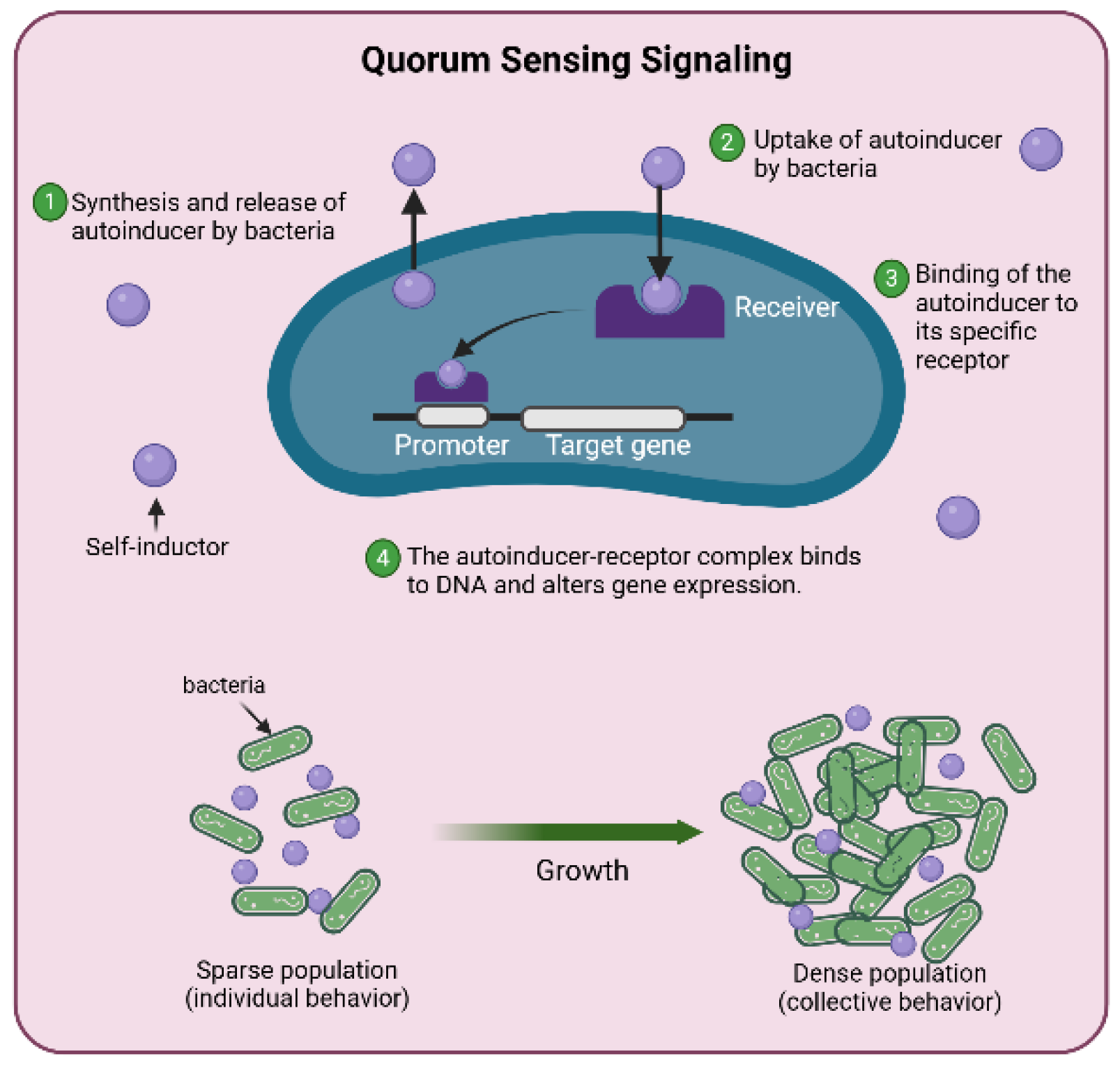
3.2.3. Quorum Sensing Detection
Inter- and intraspecies signaling in the rhizosphere primarily occurs through quorum-sensing (QS) molecules, which enable microbial communities to synchronize and coordinate their behavior [193]. This cell-to-cell communication mechanism is essential for the colonization of PGPBs in plant roots [194]. QS signals, also known as autoinducers, facilitate communication both within bacterial species (intraspecific) and between different species (interspecific), regulating gene expression in response to population density [85] (Figure 5). These signals control various microbial phenotypes, including conjugation, virulence, competition in the rhizosphere, and the production of hydrolytic enzymes and secondary metabolites [142].
In Gram-negative bacteria, N-acyl homoserine lactones (AHLs) serve as the primary QS signaling molecules [195]. These compounds regulate gene expression and influence key processes such as symbiotic interactions with plants, root development, stress responses, activation of plant defense mechanisms, metabolic regulation, and hormonal balance [196] (Figure 6). Studies have shown that various rhizosphere-associated bacteria, including Serratia, Aeromonas, Acinetobacter, and other proteobacteria, produce and respond to QS signaling molecules, particularly AHLs [197,198,199]. This ability underscores the significance of QS communication in rhizosphere dynamics, facilitating complex and beneficial interactions between microbes and plants while promoting a more favorable environment for plant growth and health.
Figure 5.
In low-density populations, bacteria exhibit individual behaviors, whereas in high-density populations, they transition to collective behavior through the production, release, and detection of signaling molecules known as autoinducers. This QS mechanism regulates gene expression associated with group behaviors, including biofilm formation, virulence, and extracellular enzyme production.
Figure 5.
In low-density populations, bacteria exhibit individual behaviors, whereas in high-density populations, they transition to collective behavior through the production, release, and detection of signaling molecules known as autoinducers. This QS mechanism regulates gene expression associated with group behaviors, including biofilm formation, virulence, and extracellular enzyme production.
Some Gram-negative bacteria, such as Burkholderia, produce diffusible signaling factor (DSF), a QS molecule with a chemical structure corresponding to cis-11-methyl-2-dodecenoic acid [200]. DSF facilitates bacterial communication and enhances innate immunity in various crops, strengthening their natural defenses against pathogens [186].
In addition to DSF, antibiotics also function as signaling molecules in QS systems [201]. At low, non-inhibitory concentrations, antibiotics act as both intraspecific and interspecific communication signals, regulating collective bacterial behaviors and coordinating community responses [202]. This dual role—as antimicrobial agents and signaling molecules—underscores their significance in microbial ecosystems, where they contribute to the organization and adaptation of bacterial populations [203].
Fungal species also produce a diverse array of QS molecules, including γ-heptalactone, γ-butyrolactone, tyrosol, farnesol, and dodecanol, which facilitate communication with bacterial species [204]. Certain fungi, such as Saccharomyces cerevisiae, secrete alcohols as signaling molecules, primarily associated with specific developmental processes [205]. Fungal-bacterial interactions mediated by QS molecules represent an intricate evolutionary strategy that enables microorganisms to compete for host infection and space colonization [206]. This cross-communication allows fungi and bacteria to coordinate their activities, shaping microbial community dynamics. By engaging in QS-mediated interactions, these organisms optimize their survival and competitive advantage in the rhizosphere, highlighting the intricate balance between cooperation and competition that drives adaptation and ecological dominance in specific niches [207].
3.3. Plant Molecular Signaling and Microbial Communication in the Rhizosphere
In addition to the signaling molecules produced by PGPB, plants secrete a wide range of compounds through their root exudates. These compounds, which include both low- and high-molecular-weight molecules, as well as volatile and non-volatile substances, play a crucial role in mediating interactions with various microorganisms in the rhizosphere [209]. By facilitating plant-microbe communication, these exudates influence the function and composition of microbial communities, promoting positive beneficial interactions that improve plant growth and its overall health. [208].
Plants activate multiple signaling pathways in response to environmental stimuli, including mitogen-activated protein kinase (MAPK) cascades, which are central to both abiotic and biotic stress responses [210,211]. These pathways are tightly regulated by PGRs, which coordinate defense mechanisms and key developmental processes [212]. Through the integration of external and internal signals, plants adapt to their environment to ensure survival and optimal growth. MAPK pathways exhibit remarkable evolutionary conservation, underscoring their fundamental role in cellular signaling networks [213]. The MAPK cascade consists of three tiers: MAPKKK, MAPKK, and MAPK, which relay signals through phosphorylation events. Although MAPK pathways are generally activated in response to specific stimuli, they can also exhibit cross-talk, leading to finely tuned and coordinated cellular responses. Additionally, MAPKs interact with other signaling molecules to process stimuli and elicit specific physiological outcomes [214] (Figure 7). These proteins are involved in diverse signaling processes, including stress responses and developmental regulation [215]. This intricate and highly adaptable signaling system enables plants to efficiently respond to environmental cues while optimizing their interactions with rhizosphere microorganisms, ultimately enhancing survival and growth [216].
The chemical compounds secreted by both PGPB and plants function as signaling molecules that significantly influence rhizosphere associations and plant development. By producing specialized metabolites, plants can shape the composition of their surrounding microbiota, which in turn modulates plant metabolism and physiology [217]. This extensive, dynamic, and cross-kingdom signaling network plays a pivotal role in structuring and adapting the rhizomicrobiome. The diverse chemical signals produced by plants are essential for shaping microbial communities associated with the roots, facilitating microbial recruitment and interaction [218]. Both microbial and plant-derived signaling molecules can be leveraged to manipulate plant responses, enabling plants to communicate with and adapt to their microbiome. These signaling interactions regulate multiple biological processes, including plant growth, gene expression, hormonal signaling, and defense responses, ultimately ensuring a balanced and beneficial relationship between plants and their associated microbial communities [142].
4. PGPB-Mediated Soil Restoration of Soils Degraded by Excessive Cultivation
Soil is a complex system composed of diverse biotic and abiotic components that collectively create a favorable environment for plant growth and support a wide range of microorganisms [219]. These elements play essential roles in maintaining soil health and providing critical ecosystem services, including nutrient cycling, climate regulation, and organic matter decomposition (Figure 8) [220]. However, intensive and prolonged agricultural use has led to soil degradation, characterized by fertility loss, compaction, erosion, and a decline in microbial biodiversity [30]. Consequently, the soil's capacity to sustain efficient and sustainable agricultural production has been significantly compromised, affecting both crop yield and quality [221]. To mitigate these issues, it is essential to implement strategies that incorporate bio-inputs, which contribute to soil restoration and preservation, promote sustainable agricultural practices, and ensure long-term food security.
Plant growth and productivity are adversely affected by a variety of stressors, broadly classified as abiotic and biotic [222]. These stress factors can arise naturally or result from anthropogenic activities, which have been exacerbated by continuous population growth and the subsequent expansion of agricultural practices to ensure food security [223].
Abiotic stressors include drought, heavy metal contamination, salinity, suboptimal temperatures, and nutrient deficiencies, all of which induce physiological and morphological alterations in plants [224,225]. Additionally, intensive agricultural practices have led to the excessive application of chemical fertilizers, pesticides, and herbicides, significantly reducing the diversity of beneficial soil microorganisms [222].
The decline in microbial diversity, reduced crop productivity, loss of soil fertility, and increased competition for nutrients are direct consequences of these stressors, which often act synergistically to exacerbate their negative impacts [226]. In response to these adverse conditions, plants activate rapid defense mechanisms, including the synthesis of PGRs and the accumulation of protective compounds such as phenolic acids and flavonoids [41]. While these adaptive strategies enhance plant resilience to unfavorable environments, they often come at the expense of reduced growth and productivity, highlighting the trade-off between survival and agricultural yield (Figure 9).
4.1. Nutrient Dynamics in the Restoration of Degraded Soils
PSBs not only enhance soil fertility but also play a crucial role in restoring degraded soils [117]. These bacteria facilitate P solubilization by converting insoluble forms into bioavailable phosphates, thereby improving nutrient accessibility for plants. This process improves soil structure, boosts aggregate stability and promotes root development, leading to reduced erosion and enhanced water retention capacity [227]. Additionally, PSBs support the establishment of cover crops, which mitigate soil exposure and further prevent erosion in degraded areas [114]. Moreover, PSBs actively contribute to the P cycle, ensuring its sustained availability in ecosystems where nutrient balance is often disrupted, a key factor in soil restoration [228]. Beyond their direct benefits to plants, PSBs also stimulate the growth of other beneficial microbial communities, enhancing organic matter decomposition and nutrient recycling, thereby improving overall soil health [229].
Diazotrophic bacteria are essential for the restoration of degraded soils due to their ability to fix atmospheric N and convert it into plant-assimilable forms. This process is critical for revitalizing soils that have lost fertility due to intensive cultivation and nutrient depletion. Fixed N not only directly benefits host plants by enhancing their growth and development but also enriches the surrounding soil, making N available to non-N-fixing plants [230].
Beyond N fixation, these bacteria contribute to soil structure improvement through multiple mechanisms. They promote root growth by producing PGRs, such as auxins, cytokinins, and gibberellins [231], and facilitate soil aggregate formation via the exudation of polysaccharides, exopolysaccharides (EPS), proteins, glycoproteins, and lipids. EPS, which include sugars such as glucose, galactose, mannose, rhamnose, and uronic acids, form a biofilm that enhances bacterial adhesion to surfaces and promotes microbial aggregation. This biofilm increases soil aggregate stability, improving porosity and water retention capacity [232].
Finally, the presence of diazotrophic bacteria enhances microbial biodiversity, thereby contributing to ecosystem stability and resilience [233]. Collectively, these bacteria play a fundamental role in soil restoration strategies by facilitating fertility recovery, improving soil structure, and sustaining the growth of robust, healthy vegetation.
4.2. Biological Strategies for Mitigating Salt-Induced Stress in Degraded Soils
Soil salinization is a complex phenomenon that can result from processes such as nitrification, denitrification, organic matter decomposition, and excessive irrigation. These processes lead to the accumulation of salts on the soil surface, including sodium sulfate, sodium chloride, magnesium sulfate and magnesium chloride [234,235,236]. Excessive salt concentrations disrupt plant biochemical and physiological processes, significantly reducing photosynthetic efficiency [237].
The removal of salts through physical and chemical methods is often slow, costly, and unsustainable. Moreover, these approaches can negatively impact soil structure, particularly in soils with high salinity levels [238]. As an alternative, halophilic and halotolerant bacteria, capable of surviving in extreme saline conditions—including concentrations of up to 150 g/L NaCl—have been identified. Among these, PGPBs play a crucial role in enhancing plant tolerance to salinity through multiple mechanisms.
These mechanisms include the accumulation of osmolytes, increased proline production, selective uptake of K+ ions, reduced electrolyte leakage, enhanced nutrient absorption, N fixation, and the solubilization of P and other essential elements. Additionally, these bacteria produce plant growth regulators that strengthen plant cellular defense systems, siderophores that enhance Fe availability, and EPS, which function as biocontrol agents and osmoprotectants, improving stress tolerance. PGPBs also exhibit ACC deaminase activity, which mitigates the negative effects of ethylene accumulation, reducing stress and promoting plant growth under saline conditions [36].
Another key mechanism involves the regulation of aquaporins, proteins whose synthesis is induced under water stress. Aquaporins facilitate water uptake and optimize photosynthetic efficiency, thereby mitigating the adverse effects of salinity [239]. Collectively, these bacteria represent a promising biological strategy for enhancing plant resilience to salinity and restoring degraded soils.
Phosphate uptake and transport in plants are highly sensitive to salinity. Under saline conditions, P tends to precipitate, rendering it unavailable for plant growth [240]. To counteract this issue, PGPBs play a critical role by solubilizing inorganic P, a key mechanism for mitigating salt stress [241]. This process enhances P availability and supports plant development under saline conditions, helping plants overcome the challenges associated with salt stress [242].
4.3. Bioremediation of Heavy Metal-Contaminated Soils Using PGPB.
Industrialization, anthropogenic activities, and the excessive use of chemical fertilizers have contributed to soil contamination with heavy metals (HMs), profoundly impacting the plant microbiome and posing risks to human and animal health [243]. This contamination is not confined to soil; HMs can leach into groundwater and accumulate in agriculturally important plant species, increasing the risk of exposure through consumption [244]. These toxic metals induce DNA damage and exhibit carcinogenic effects in animals and humans due to their mutagenic capacity [245]. Moreover, prolonged exposure to high HMs concentrations has been linked to severe health effects in humans and wildlife [246]. As a result, HMs contamination is a globally significant environmental issue.
Heavy metals are defined as metals with densities greater than 5 g/cm³ [247]. The most common HMs in soils include manganese (Mn), copper (Cu), cadmium (Cd), lead (Pb), chromium (Cr), aluminum (Al) and zinc (Zn). Of the 53 identified HMs, 17 are biologically active and essential for the proper functioning of organisms and ecosystems [245]. Some, such as copper (Cu), iron (Fe), molybdenum (Mo), manganese (Mn) and boron (B), function as micronutrients [248], while others, including selenium (Se), arsenic (As), chromium (Cr), nickel (Ni), vanadium (V), and zinc (Zn), are classified as trace elements [249]. However, metals such as silver (Ag), mercury (Hg), lead (Pb), cadmium (Cd), and uranium (U), have no known biological function and are highly toxic to plants and microorganisms [250] (Figure 10).
Plant-associated microorganisms play a crucial role in alleviating stress in HM-contaminated soils by producing PGRs such as gibberellins, cytokinins, and IAA, as well as ACC deaminase and siderophores [243].
PGPB exhibit resistance to HM toxicity through various mechanisms, including metal exclusion via permeability barriers, active transport, extracellular and intracellular sequestration, modification of cellular metal targets, detoxification, and passive transport systems. These mechanisms are primarily regulated by genes located on chromosomal or plasmid DNA. The permeability barrier preventing metal entry consists of the bacterial cell wall, extracellular layers, exopolysaccharides EPS, and the capsule, which collectively block the uptake of toxic metals. Additionally, the genetic modulation of protein channels in the bacterial membrane enables selective permeability, allowing PGPB to regulate metal influx. These metals can non-specifically bind to extracellular polymeric substances, lipopolysaccharides, membrane proteins, and the outer membrane envelope, thereby preventing their interference with metal-sensitive biochemical processes [251].
Despite the detrimental effects of HMs on bacterial viability, activity, and abundance in soils, certain bacterial populations have adapted to their presence. This adaptation leads to shifts in the composition of soil and rhizosphere microbial communities, as HM resistance is frequently encoded in plasmids [244].
Under HMs stress conditions, PGPB contribute to metal removal and promote plant development through mechanisms such as sequestration via EPS, efflux pumps, volatilization, metal complex formation, reduced cell permeability, and enzymatic detoxification [245]. These bacteria also facilitate key ecological functions, including N fixation, siderophore production, nutrient mobilization, and phosphate solubilization, which support the establishment of plant species used in phytoremediation and microbially mediated transformations [252].
Moreover, PGPB activity reduces ethylene production and modulates IAA and ACC deaminase levels in plants, thereby mitigating stress responses and decreasing disease incidence in HM-contaminated soils [253]. The application of inoculants containing growth-promoting bacterial strains, in combination with phytoremediation strategies, has emerged as an effective approach for HM removal [83].
The interaction between microorganisms and plants represents a promising biotechnological strategy for the remediation of HM-contaminated soils. Through direct and indirect plant growth-promoting mechanisms, PGPB mitigate HM toxicity, restore soil health, and enhance agricultural productivity in polluted environments.
Figure 11.
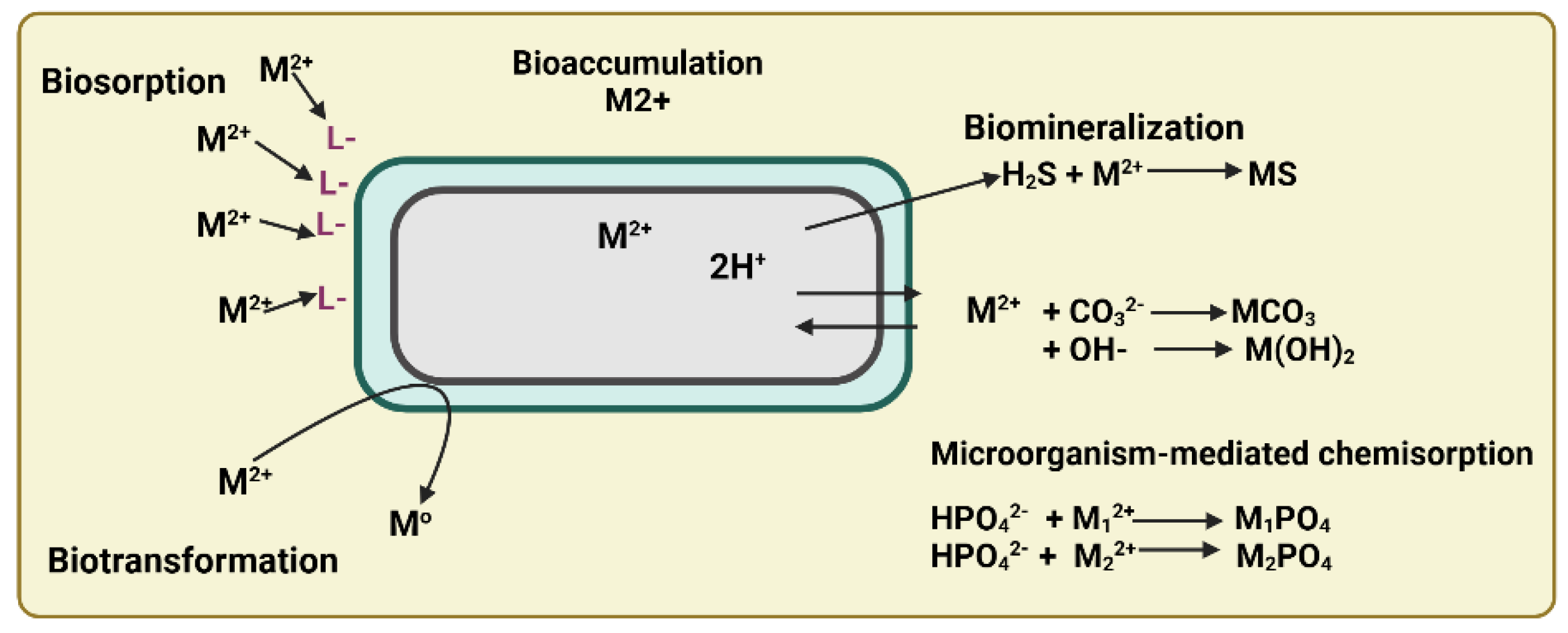
Mechanisms of interaction between HMs and microorganisms. Microorganisms engage with HMs through five primary pathways that facilitate their transformation, immobilization, or accumulation: biosorption (binding of metals to cell surface components), bioaccumulation (active transport and intracellular storage), biomineralization (conversion into mineral compounds such as sulfides or carbonates), biotransformation (modification of oxidation states, such as reduction to less toxic forms), and microbially mediated chemoadsorption (formation of complexes with phosphates, leading to immobilization).
Figure 11.
Mechanisms of interaction between HMs and microorganisms. Microorganisms engage with HMs through five primary pathways that facilitate their transformation, immobilization, or accumulation: biosorption (binding of metals to cell surface components), bioaccumulation (active transport and intracellular storage), biomineralization (conversion into mineral compounds such as sulfides or carbonates), biotransformation (modification of oxidation states, such as reduction to less toxic forms), and microbially mediated chemoadsorption (formation of complexes with phosphates, leading to immobilization).
Initially, PGPB were utilized and studied to improve crop yields and to control plant diseases. However, their role in bioremediation has recently become more significant because of their ability to degrade and even mineralize organic compounds in association with plants [245]. This expanded functionality broadens the potential applications of PGPB, which could significantly influence future bioremediation strategies [251]. To date, various native bacterial strains capable of degrading specific contaminants have been isolated from diverse environments [254,255,256]. Additionally, the genetic pathways underlying these degradation processes have been extensively characterized [257].
5. Perspectives on Innovative Strategies for Soil Restoration Using PGPB in Overcultivated Lands
According to the report “The State of the World's Land and Water Resources for Food and Agriculture”, the biophysical condition of 5,670 million hectares of land is declining worldwide. Of this, 1,660 million hectares (29%) have been degraded due to human activities, while the remaining 4,026 million hectares are experiencing degradation due to natural processes or human-induced factors. Approximately half of these deteriorated lands are in poor condition, making them highly susceptible to further degradation. Additionally, 656 million hectares (12% of globally degraded land) are under moderate pressure, which could accelerate degradation processes. Overall, it is estimated that 41% of global land degradation is directly attributed to anthropogenic pressures [258].
This scenario underscores the urgent need for viable and effective solutions to soil degradation, given its strong link to human activity and its implications for food security. As agricultural land expansion is increasingly constrained while global food demand continues to rise, implementing alternative strategies to restore both vulnerable and degraded agricultural soils is essential. Such efforts are critical to sustaining agricultural productivity and meeting future food demands [259].
At the 2023 United Nations Sustainable Development Goals (SDG) Summit, the 17 SDGs were evaluated, with particular emphasis on those directly related to soil and natural resource management: Zero Hunger (SDG 2), Climate Action (SDG 13), and Life on Land (SDG 15). These goals are fundamental to achieving food security and fostering sustainable economic growth [260]. In this context, preserving PGPB biodiversity in soils has become a key component of sustainable agricultural strategies, particularly in degraded agricultural ecosystems [261,262].
The implementation of these strategies would significantly enhance global food production, addressing one of the major barriers to sustainable development—hunger [263]. In this critical context, integrating microbial technologies into agricultural systems is essential, with PGPB playing a central role as an environmentally friendly solution. Their application has proven effective in sustainable land management, restructuring rhizospheres in degraded agricultural soils, and improving both crop health and yield [264].
Microbial inoculants have the potential to enhance soil health and sustain agricultural productivity across various agroecosystems. However, while numerous studies have demonstrated the effectiveness of PGPB under controlled or optimal conditions, their application in degraded soils at the field level has not consistently produced satisfactory results. To overcome these challenges, researchers have focused on developing PGPB consortia tailored to address soil degradation in real-world environments [265].
A promising strategy involves combining these consortia with compostable organic residues, which significantly enhances their effectiveness. For instance, Menguala et al. [266] conducted a field trial to assess the effects of inoculating Lavandula dentata L. seedlings with a consortium of rhizobacteria in combination with sugar beet residues. The results demonstrated a significant increase in dry shoot and root biomass, as well as an improvement in foliar nutrient content (NPK). Additionally, P and N availability in the rhizosphere increased by 29% and 46%, respectively, compared to the control. These treatments also improved the microbiological and biochemical properties of the soil. The selection of efficient rhizobacterial strains and their integration with organic residues has been identified as a key factor in maximizing the effectiveness of these biotechnological tools for revegetating and rehabilitating degraded soils, particularly in semi-arid conditions.
The combined application of PGPB and biochar (BC) offers multiple benefits. BC enhances soil physicochemical properties, serves as a carrier for PGPB inoculation, provides a carbon source for bacterial proliferation, and offers a protective microenvironment against competitors due to its porous structure [209,267]. Recent studies have shown that the combination of PGPB and BC is an effective strategy for the remediation and restoration of degraded soils, as it increases organic carbon content, improves soil water retention, and enhances microbial activity, particularly in mitigating ethylene-induced stress [268].
Notable examples include the remediation of soils contaminated with heavy metals [206,269], hydrocarbons [270,271,272,273], and those affected by salinity stress [274,275,276]. Despite these promising findings, further research is required to confirm the long-term efficacy of this approach under diverse field and climatic conditions.
Natural revegetation in soils subjected to prolonged agricultural activity, particularly in rainfed cropping systems, is often a slow process due to recurrent water scarcity, which limits plant establishment and growth [277]. Other factors, such as soil type, suitability, and nutrient availability, also play a critical role in the regeneration of vegetation cover [278]. To address these challenges, revegetation programs using native plant species have been implemented, as they are considered the most suitable strategy for restoring degraded lands [180]. For example, Calle et al. [279] propose the use of shrubs and grasses to rehabilitate arid areas, a strategy that has proven effective in both arid and semi-arid regions [280,281].
Moreover, the application of PGPB, either through seed inoculation [44] or direct soil application [282], has emerged as an effective tool for restoring degraded soils. Successful revegetation strategies based on microbial inoculation require the development of inoculants that perform optimally under specific environmental conditions, thereby enhancing plant growth, nutrient uptake, and water status [283].
Plants and soil microorganisms often share adaptations to the same environmental conditions, suggesting that native strains may be more effective in promoting plant growth. This approach not only supports the preservation of local biodiversity but also minimizes the risks associated with introducing exogenous microbial species. As previously discussed, free-living bacteria can colonize the rhizosphere, facilitating root system establishment and, consequently, enhancing plant health and nutrition. Additionally, PGPB have been shown to synthesize essential vitamins that play a crucial role in key physiological processes involved in plant-microorganism interactions [280].
6. Conclusions
Ensuring sufficient agricultural production to sustain human life amid accelerating soil degradation, pollution, and water scarcity represents one of the greatest challenges of our time. Additionally, the growing demand for organic products reflects concerns over the excessive use of agrochemicals, which have been shown to compromise soil integrity, reduce crop yields, and exacerbate environmental issues.
Extensive research has been dedicated to developing sustainable alternatives to synthetic agricultural inputs. Among these, the application of PGPB stands out as a promising strategy, demonstrating immediate and positive effects on soil health and plant development. However, field-level application remains challenging, particularly in degraded soils, highlighting the need for optimized microbial formulations.
Recent studies indicate that combining PGPB with organic amendments such as biochar or compostable residues significantly enhances their effectiveness. These combinations improve soil structure, increase nutrient availability, and provide a more favorable microenvironment for microbial survival. Additionally, integrating microbial inoculants into revegetation programs using native plant species has proven to be an effective strategy for restoring degraded soils, particularly in arid and semi-arid ecosystems.
Leveraging PGPB-based strategies in conjunction with complementary soil improvement approaches presents a viable and environmentally friendly solution for soil restoration. Prioritizing research and large-scale implementation of these integrated solutions is essential to mitigating soil degradation and ensuring the sustainability of global food production in the face of climate change.
The latest findings suggest that adopting multiple soil restoration strategies, particularly those that enhance soil biodiversity, yields the most promising results. The coming years will be critical in addressing these challenges, but the scientific community is well-prepared to advance innovative solutions that contribute to sustainable agriculture and ecosystem resilience.
Author Contributions
Conceptualization, M-RM, M-VFD; writing—original draft preparation, M-RM; writing—review and editing, M-VFD, S-RG and P-EM; visualization, M-RM; supervision, M-VFD and P-EM. methodology, M-VFD and P-EM; formal analysis, M-VFD and P-EM; investigation S-RG, M-VFD, P-EM and M-RM. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting this review will be made available by the authors on request.
Acknowledgments
Mario Maciel-Rodríguez acknowledges SECIHTI, which provided a doctoral scholarship (CVU: 668802), and to the Secretariat of Research and Postgraduate Studies (SIP) of the National Polytechnic Institute (IPN), which provided a BEIFI program scholarship. This work was supported by the National Polytechnic Institute, Mexico. Francisco David Moreno-Valencia acknowledges SECIHTI, which provided a postdoctoral scholarship (CVU: 309635), and the Meritorious Autonomous University of Puebla and Microbiological Sciences Research Centre, for providing the laboratories and resources to carry out the postdoctoral work.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| PGPB | Plant Growth Promoting Bacteria |
| PGRs | Plant Growth regulators |
| PSB | Phosphate Solubilizing Bacteria |
| QS | Quorum Sensing |
| HM | Heavy Metals |
References
- Subramaniam, Y.; Masron, T.A. Food security and environmental degradation: evidence from developing countries. GeoJournal 2021, 86, 1141–1153. [Google Scholar] [CrossRef]
- Sartori, M.; Ferrari, E.; M'Barek, R.; Philippidis, G.; et al. Remaining loyal to our soil: A prospective integrated assessment of soil erosion on global food security. Ecological Economics 2024, 219, 108103. [Google Scholar] [CrossRef]
- Pozza, L.E.; Field, D.J. The science of Soil security and food security. Soil Security 2020, 1, 100002. [Google Scholar] [CrossRef]
- Lal, R.; Moldenhauer, W.C. Effects of soil erosion on crop productivity. Crit. Rev. Plant Sci. 1987, 5, 303–367. [Google Scholar] [CrossRef]
- Bravo-Espinosa, M.; Mendoza, M.E.; Medina-Orozco, L.; Prat, C.; García-Oliva, F.; López-Granados, E. Runoff, soil loss, and nutrient depletion under traditional and alternative cropping systems in the Transmexican Volcanic Belt, Central Mexico. Land Degrad. Develop. 2009, 20, 640–653. [Google Scholar] [CrossRef]
- Yang, T.; Siddique, K.H.M.; Liu, K. Cropping systems in agriculture and their impact on soil health: A review. Glob. Ecol. Conserv. 2020, 23, e01118. [Google Scholar] [CrossRef]
- Visha Kumari, V.; Balloli, S.S.; Kumar, M.; Ramana, D.B.V.; et al. Diversified cropping systems for reducing soil erosion and nutrient loss and for increasing crop productivity and profitability in rainfed environments. Agricultural Systems 2024, 217, 103919. [Google Scholar] [CrossRef]
- Hunt, N.D.; Hill, J.D.; Liebman, M. Cropping system diversity effects on nutrient discharge, soil erosion, and agronomic performance. Environ. Sci. Technol. 2019, 53, 1344–1352. [Google Scholar] [CrossRef]
- Goel, R.; Chaturvedi, P.; Kumar, S.; Soni, R.; Suyal, D.C. Editorial: Hazardous pollutants in agricultural soil and environment. Front. Microbiol. 2024, 15, 1411735. [Google Scholar] [CrossRef]
- Ju, X.T.; Kou, C.L.; Christie, P.; Dou, Z.X.; Zhang, F.S. Changes in the soil environment from excessive application of fertilizers and manures to two contrasting intensive cropping systems on the North China Plain. Environ. Pollut. 2007, 145, 497–506. [Google Scholar] [CrossRef]
- Dai, Q.; Zhang, L.; Jiang, S.; Su, B.; Li, Z.; Shuai, Y.; Wang, J. Improved Salt Tolerance in Brassica napus L. Overexpressing a Synthetic Deinocuccus Stress-Resistant Module DICW. Int. J. Mol. Sci. 2025, 26, 2500. [Google Scholar] [CrossRef]
- Thomas, E.; Thomas Kurien, V.; Prabha, V.S.; Thomas, A.P. Monoculture vs mixed-species plantation impact on the soil quality of an ecologically sensitive area. J. Agric. Environ. Int. Dev. 2020, 114, 41–62. [Google Scholar] [CrossRef]
- Qian, L.; Zhang, J.; Lisha, C. Impact of monoculture of poplar on rhizosphere microbial communities over time. Pedosphere 2020, 30, 487–495. [Google Scholar] [CrossRef]
- Mera, J.L.A.; Vinces, L.M.G.; Murillo, D.M.S.; Chilán, G.R.M. The monoculture of corn (Zea mayz) and its impact on fertility soil. Int. J. Chem. Mater. Sci. 2021, 4, 7–12. [Google Scholar]
- Gui, H.; Fan, L.; Wang, D.; Yan, P.; et al. Variations in soil nutrient dynamics and bacterial communities after the conversion of forests to long-term tea monoculture systems. Front. Microbiol. 2022, 13, 896530. [Google Scholar] [CrossRef]
- Ilampooranan, I.; Van Meter, K.J.; Basu, N.B. Intensive agriculture, nitrogen legacies, and water quality: intersections and implications. Environ. Res. Lett. 2022, 17, 035006. [Google Scholar] [CrossRef]
- Purwanto, B. H.; Alam, S. Impact of intensive agricultural management on carbon and nitrogen dynamics in the humid tropics. J. Soil Sci. Plant Nutr. 2019, 66, 50–59. [Google Scholar] [CrossRef]
- Cárceles Rodríguez, B.; Durán-Zuazo, V.H.; Soriano Rodríguez, M.; García-Tejero, I.F.; Gálvez Ruiz, B.; Cuadros Tavira, S. Conservation agriculture as a sustainable system for soil health: A review. Soil Systems 2022, 6, 87. [Google Scholar] [CrossRef]
- Hamada, K.; Inoue, H.; Mochizuki, H.; Miyamoto, T.; Asakura, M.; Shimizu, Y. Effect of hardpan on the vertical distribution of water stress in a converted paddy field. Soil Tillage Res. 2021, 214, 105161. [Google Scholar] [CrossRef]
- Zhang, M.; Dai, S.; Gul, S.; He, L.; Chen, H.; Liu, D. Effect of plow pan on the redistribution dynamics of water and nutrient transport in soils. Sustainability 2024, 16, 8859. [Google Scholar] [CrossRef]
- Issaka, S.; Ashraf, M.A. Impact of soil erosion and degradation on water quality: a review. Geol. Ecol. Landsc. 2017, 1, 1–11. [Google Scholar] [CrossRef]
- Shaheb, M.R.; Venkatesh, R.; Shearer, S.A. A review on the effect of soil compaction and its management for sustainable crop production. J. Biosyst. Eng. 2021, 46, 417–439. [Google Scholar] [CrossRef]
- Fernández-Ojeda, P.R.; Acevedo, D.C.; Villanueva-Morales, A.; Uribe-Gómez, M. State of the essential chemical elements in the soils of natural, agroforestry and monoculture systems. Rev. Mex. Cienc. For. 2016, 7, 65–77. [Google Scholar]
- Shah, A.N.; Tanveer, M.; Shahzad, B.; Yang, G.; et al. Soil compaction effects on soil health and crop productivity: an overview. Environ. Sci. Pollut. Res. 2017, 24, 10056-10067. [CrossRef]
- Haruna, S.I.; Anderson, S.H.; Udawatta, R.P.; Gantzer, C.J.; et al. Improving soil physical properties through the use of cover crops: A review. Agrosyst. Geosci. Environ. 2020, 3, e20105. [CrossRef]
- Hamza, M.A.; Anderson, W.K. Soil compaction in cropping systems: A review of the nature, causes and possible solutions. Soil Till Res. 2005, 82, 121–145. [Google Scholar] [CrossRef]
- Yu An, Le Wang, Mingye Zhang, Shouzheng Tong, Yifan Li, Haitao Wu, Ming Jiang, Xuan Wang, Yue Guo, Li Jiang, Synergies and trade-offs between aboveground and belowground traits explain the dynamics of soil organic carbon and nitrogen in wetlands undergoing agricultural management changes in semi-arid regions, Agric. Ecosyst. Environ. 2025, Volume 381. [CrossRef]
- Piccoli, I.; Seehusen, T.; Bussell, J.; Vizitu, O.; et al. Opportunities for mitigating soil compaction in Europe—case studies from the soilcare project using soil-improving cropping systems. Land 2022, 11, 223. [Google Scholar] [CrossRef]
- Swain, C.K. Environmental pollution indices: a review on concentration of heavy metals in air, water, and soil near industrialization and urbanisation. Discov. Environ. 2024, 2, 5. [Google Scholar] [CrossRef]
- Bedolla-Rivera, H.I.; Negrete-Rodríguez, M.L.X.; Gámez-Vázquez, F.P.; Álvarez-Bernal, D.; Conde-Barajas, E. Analyzing the impact of intensive agriculture on soil quality: a systematic review and global meta-analysis of quality indexes. Agronomy 2023, 13, 2166. [Google Scholar] [CrossRef]
- Tsiafouli, M. A.; Thébault, E.; Sgardelis, S. P.; De Ruiter, P. C.; et al. Intensive agriculture reduces soil biodiversity across Europe. Glob. Change Biol. 2015, 21, 973–985. [Google Scholar] [CrossRef]
- Tripathi, S.; Srivastava, P.; Devi, R. S.; Bhadouria, R. Chapter 2: Influence of synthetic fertilizers and pesticides on soil health and soil microbiology. In: Narasimha, M., & Prasad, V. (Eds). Agrochemicals detection, treatment and remediation. Butterworth-Heinemann 2020, Pp:25-54. [CrossRef]
- Baweja, P.; Kumar, S.; Kumar, G. Fertilizers and pesticides: Their impact on soil health and environment. In: Giri, B., Varma, A. (eds) Soil health. Soil Biol. 2020, vol 59. Springer, Cham. [CrossRef]
- Bourke, P.M.; Evers, J.B.; Bijma, P.; et al. Breeding beyond monoculture: Putting the “intercrop” into crops. Front. Plant Sci. 2021, 12, 734167. [Google Scholar] [CrossRef]
- Wei, Z.; Wang, H.; Ma, C.; Li, S.; et al. Unraveling the impact of long-term rice monoculture practice on soil fertility in a rice-planting meadow soil: A perspective from microbial biomass and carbon metabolic rate. Microorganisms 2022, 10, 2153. [Google Scholar] [CrossRef]
- Reang, L., Bhatt, S., Tomar, R.S. et al. (2022). Plant growth promoting characteristics of halophilic and halotolerant bacteria isolated from coastal regions of Saurashtra Gujarat. Sci. Rep. 2022, 12, 4699. [CrossRef]
- Tanya Morocho and Leiva-Mora, M. Efficient microorganisms, functional properties and agricultural applications. Ctro. Agr. 2019, vol.46, n.2 [citado 2025-03-04], pp. 93-103.
- Chen, Q.; Song, Y.; An, Y.; Lu, Y.; Zhong, G. Mechanisms and Impact of Rhizosphere Microbial Metabolites on Crop Health, Traits, Functional Components: A Comprehensive Review. Molecules 2024, 29, 5922. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; et al. Plant–microbiome interactions: from community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Figueiredo-Santos, L.; Lopes-Olivares, F. Plant microbiome structure and benefits for sustainable agriculture. Curr. Plant Biol. 2021, 26, 100198. [Google Scholar] [CrossRef]
- Fitzpatrick, C.R.; Salas-González, I.; Conway, J.M.; Finkel, O.M.; et al. The plant microbiome: From ecology to reductionism and beyond. Annu. Rev. Microbiol. 2020, 74, 81–100. [Google Scholar] [CrossRef]
- Trivedi, P.; Batista, B.D.; Bazany, K.E. y Singh, B.K. Plant–microbiome interactions under a changing world: responses, consequences and perspectives. New Phytol. 2022, 234, 1951–1959. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Berg, G.; Tack, A.J.M.; Carolina Lobato, C.; Wassermann, B. From seed to seed: the role of microbial inheritance in the assembly of the plant microbiome. Trends Microbiol. 2023, 31, 346-355. [CrossRef]
- Molina-Romero, D.; Juárez-Sánchez, S.; Venegas, B.; Ortíz-González, C.S.; Baez, A.; Morales-García, Y.E.; Muñoz-Rojas, J. A bacterial consortium interacts with different varieties of maize, promotes the plant growth, and reduces the application of chemical fertilizer under field conditions. Front. Sustain. Food Syst. 2021, 4, 616757. [CrossRef]
- Heredia-Acuña, C.; Almaraz-Suarez, J.J.; Arteaga-Garibay, R.; Ferrera-Cerrato, R.; Pineda-Mendoza, D.Y. Isolation, characterization and effect of plant-growth-promoting rhizobacteria on pine seedlings (Pinus pseudostrobus Lindl.). J. For. Res. 2019, 30, 1727-1734. [CrossRef]
- Domínguez-Castillo, C.; Alatorre-Cruz, J.M.; Castañeda-Antonio, D.; et al. Potential seed germination-enhancing plant growth-promoting rhizobacteria for restoration of Pinus chiapensis ecosystems. J. For. Res. 2021, 32, 2143–2153. [Google Scholar] [CrossRef]
- Patel, J. S., Yadav, S. K., Bajpai, R., Teli, B., & Rashid, M. PGPR secondary metabolites: an active syrup for improvement of plant health. In: Molecular aspects of plant beneficial microbes in agriculture, Sharma, V., Salwan, R., Khalil, L. & Al-Ani, T (eds), Academic Press, 2020, pp. 195-208. [CrossRef]
- Saha, R.; Saha, N.; Donofrio, R.S.; Bestervelt, L.L. Microbial siderophores: a mini review. J. Basic Microbiol. 2013, 53, 303–317. [Google Scholar] [CrossRef]
- Sayyed, R.Z.; Chincholkar, S.B.; Reddy, M.S.; Gangurde, N.S.; Patel, P.R. Siderophore Producing PGPR for Crop Nutrition and Phytopathogen Suppression. In: Bacteria in Agrobiology: Disease Management, Maheshwari, D. (eds), Springer, Berlin, Heidelberg, 2013. [CrossRef]
- Saha, M.; Sarkar, S.; Sarkar, B.; et al. Microbial siderophores and their potential applications: a review. Environ. Sci. Pollut. Res. 2016, 23, 3984–3999. [Google Scholar] [CrossRef]
- Perez, A.; Rossano, S.; Trcera, N.; Verney-Carron, A.; Rommevaux, C.; Fourdrin, C.; Agnello, A.C. Direct and indirect impact of the bacterial strain Pseudomonas aeruginosa on the dissolution of synthetic Fe (III)-and Fe (II)-bearing basaltic glasses. Chem. Geol. 2019. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Leong, J.; Teintze, M.; Schroth, M.N. Enhanced plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature 1980, 286, 885–886. [Google Scholar] [CrossRef]
- Laschat, S.; Bilitewski, U.; Blodgett, J.; Duhme-Klair, A.-K.; Dallavalle, S.; Routledge, A.; Schobert, R. Chemical and biological aspects of nutritional immunity-perspectives for new anti-infectives targeting iron uptake systems. Angew. Chem. Int. Ed. 2017. [Google Scholar] [CrossRef]
- Grobelak, A.; Napora, A.; Kacprzak, M. Using plant growth-promoting rhizobacteria (PGPR) to improve plant growth. Ecol. Eng. 2015, 84, 22–28. [Google Scholar] [CrossRef]
- Glick, Bernard. Beneficial Plant-Bacterial Interactions. Springer Nature 2015, 10.1007/978-3-319-13921-0. [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- AlKahtani, M.D.F.; Fouda, A.; Attia, K.A.; et al. Isolation and characterization of plant growth promoting endophytic bacteria from desert plants and their application as bioinoculants for sustainable agriculture. Agronomy 2020, 10, 1325. [Google Scholar] [CrossRef]
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial proteases applications. Front. Bioeng. Biotechnol. 2019, 7, 451237. [Google Scholar] [CrossRef] [PubMed]
- Bumunang, E.; Babalola, O.O. Characterization of rhizobacteria from field grown genetically modified (GM) and non-GM maizes. Braz. Arch. Biol. Technol. 2014, 57, 1–8. [Google Scholar] [CrossRef]
- Talebi Atouei, M.; Pourbabaee, A.A.; Shorafa, M. Alleviation of salinity stress on some growth parameters of wheat by exopolysaccharide-producing bacteria. Iran. J. Sci. Technol. Trans. A: Sci. 2019, 43, 2725–2733. [Google Scholar] [CrossRef]
- Alemu, F. Isolation of Pseudomonas flurescens from rhizosphere of faba bean and screen their hydrogen cyanide production under in vitro study, Ethiopia. Am. J. Life Sci. 2016, 4, 13. [Google Scholar] [CrossRef]
- Sehrawat, A.; Sindhu, S.; Glick, B.R. Hydrogen cyanide production by soil bacteria: biological control of pests and promotion of plant growth in sustainable agriculture. Pedosphere 2022, 32, 15–38. [Google Scholar] [CrossRef]
- Ismail, M.A.; Amin, M.A.; Eid, A.M.; et al. Comparative study between exogenously applied plant growth hormones versus metabolites of microbial endophytes as plant growth-promoting for Phaseolus vulgaris L. Cells 2021, 10, 1059. [Google Scholar] [CrossRef]
- Far, B.E.; Ahmadi, Y.; Khosroshahi, A.Y.; Dilmaghani, A. Microbial alpha-amylase production: progress, challenges and perspectives. Adv. Pharm. Bull. 2020, 10, 350–358. [Google Scholar] [CrossRef]
- Witte, C.P. Urea metabolism in plants. Plant. Sci. 2011, 180, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Teng, A.; Chu, J.; Cao, B. A quantitative, high-throughput urease activity assay for comparison and rapid screening of ureolytic bacteria. Environ. Res. 2022, 208, 112738. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—a review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Mekureyaw, M.F.; Pandey, C.; Hennessy, R.C.; Nicolaisen, M.H.; Liu, F.; Nybroe, O.; Roitsch, T. The cytokinin-producing plant beneficial bacterium Pseudomonas fluorescens G20-18 primes tomato (Solanum lycopersicum) for enhanced drought stress responses. J. Plant Physiol. 2022, 270, 153629. [Google Scholar] [CrossRef]
- Davies, P.J. The Plant Hormones: Their Nature, Occurrence, and Functions. In: Plant Hormones, Davies, P.J. (eds). Springer, Dordrecht, 1995, pp 1-12. [CrossRef]
- Smith, S.; Li, C.; Li, J. Hormone Function in Plants. In: Hormone metabolism and signaling in plants, Li J., Li C. Smith S. (eds), Academic Press, 2017, pp 1-38. [CrossRef]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, L. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Front. Plant Sci. 2013, 4, 1–19. [Google Scholar] [CrossRef]
- Gutiérrez-Mañero, F.J.; Ramos-Solano, B.; Probanza, A.; Mehouachi, J.; Tadeo, F.R.; Talon, M. The plant-growth-promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiol. Plant. 2011, 111, 206–211. [Google Scholar] [CrossRef]
- Rehman, F.; Kalsoom, M.; Adnan, M.; Toor, M.D.; Zulfiqar, A. Plant growth promoting rhizobacteria and their mechanisms involved in agricultural crop production: a review. SunText Rev. Biotechnol. 2022, 1, 110. [Google Scholar] [CrossRef]
- Chandra, S.; Askari, K.; Kumari, M. Optimization of indole acetic acid production by isolated bacteria from Stevia rebaudiana rhizosphere and its effects on plant growth. J. Genet. Eng. & Biotechnol. 2018, 16, 581-586. [CrossRef]
- Vanneste, S. and Friml J. Auxin: a trigger for change in plant development. Cell 2009, 136, 1005-1016. [CrossRef]
- Satyaprakash, M.; Satyaprakash, M.; Nikitha, T.; Reddi, E.U.B.; Sadhana, B.; Satya Vani, S. Phosphorous and phosphate solubilising bacteria and their role in plant nutrition. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2133–2144. [Google Scholar] [CrossRef]
- Verma, J.P.; Jaiswal, D.K.; Singh, S.; Kumar, A.; Prakash, S.; Curá, J.A. Consequence of phosphate solubilising microbes in sustainable agriculture as efficient microbial consortium: a review. Climate Change and Environmental Sustainability 2017. 5, 1. [CrossRef]
- Eshaghi, E.; Nosrati, R.; Owlia, P.; Malboobi, M.A.; Ghaseminejad, P.; GanjaliIranian, M.R. Zinc solubilization characteristics of efficient siderophore-producing soil bacteria. J. Microbiol. 2019, 11, 419–430. [Google Scholar] [CrossRef]
- Fasim, F.; Ahmed, N.; Parsons, R.; Gadd, G.M. Solubilization of zinc salts by a bacterium isolated from the air environment of a tannery. FEMS Microbiol. Lett. 2002; 213, 1–6. [CrossRef]
- Olaniyan, F.T.; Alori, E.T.; Adekiya, A.O.; Ayorinde, B.B.; Daramola, F.Y.; Osemwegie, O.O.; Babalola, O.O. The use of soil microbial potassium solubilizers in potassium nutrient availability in soil and its dynamics. Ann. Microbiol. 2022; 72, 1–12. [CrossRef]
- Rawat, J.; Pandey, N.; Saxena, J. Role of Potassium in Plant Photosynthesis, Transport, Growth and Yield. In: Role of Potassium in Abiotic Stress, Iqbal, N., Umar, S. (eds), Springer, Singapore, 2022. [CrossRef]
- Bukhat, S.; Imran, A.; Javaid, S.; Shahid, M.; Majeed, A.; Naqqash, T. Communication of plants with microbial world: Exploring the regulatory networks for PGPR mediated defense signaling. Microbiol. Res. 2020, 238, 126486. [Google Scholar] [CrossRef]
- Song, S.; Liu, Y.; Wang, N.R.; Haney, C.H. Mechanisms in plant-microbiome interactions: lessons from model systems. Curr. Opin. Plant Biol. 2021, 62, 102003. [Google Scholar] [CrossRef] [PubMed]
- Yahaya, S.M.; Mahmud, A.A.; Abdullahi, M.; Haruna, A. Recent advances in the chemistry of nitrogen, phosphorus and potassium as fertilizers in soil: A review. Pedosphere 2023, 33, 385–406. [Google Scholar] [CrossRef]
- Santoyo, G.; Urtis-Flores, C.A.; Loeza-Lara, P.D.; Orozco-Mosqueda, M.d.C.; Glick, B.R. Rhizosphere colonization determinants by Plant Growth-Promoting Rhizobacteria (PGPR). Biology 2021, 10, 475. [CrossRef]
- Sharma, P.; Kumawat, K.C.; Kaur, S. Plant growth promoting rhizobacteria in nutrient enrichment: Current perspectives. In: Biofortification of Food Crops, Singh, U., Praharaj, C., Singh, S., Singh, N. (eds). Springer, New Delhi, 2016, pp263-289. [CrossRef]
- Geddes, B.A.; Oresnik, I.J. The Mechanism of Symbiotic Nitrogen Fixation. In: The Mechanistic Benefits of Microbial Symbionts. Advances in Environmental Microbiology vol 2. Hurst, C. (eds), Springer, Cham., 2016, pp 69-97. [CrossRef]
- Huergo, L.F. Huergo, L.F. Chandra, G.; Merrick, M. PII signal transduction proteins: nitrogen regulation and beyond. FEMS Microbiol. Rev. 2013, 37, 251-283. [CrossRef]
- Merrick, M. Post-translational modification of PII signal transduction proteins. Front. Microbiol. 2015, 5, 763. [Google Scholar] [CrossRef]
- Bueno Batista, M.; Dixon, R. Manipulating nitrogen regulation in diazotrophic bacteria for agronomic benefit. Biochem. Soc. Trans. 2019, 47, 603–614. [Google Scholar] [CrossRef]
- Gerhardt, E.C.M.; Parize, E.; Gravina, F.; Pontes, F.L.D.; et al. The protein-protein interaction network reveals a novel role of the signal transduction protein PII in the control of c-di-GMP homeostasis in Azospirillum brasilense. mSystems 2020, 5, e00817-20. [Google Scholar] [CrossRef]
- Zeng, Y.; Guo, L.; Gao, Y.; Cui, L.; et al. Formation of NifA-PII complex represses ammonium-sensitive nitrogen fixation in diazotrophic proteobacteria lacking NifL. Cell Reports 2024, 43, 114476. [Google Scholar] [CrossRef] [PubMed]
- Plunkett, M.H.; Knutson, C.M.; Barney, B.M. Key factors affecting ammonium production by an Azotobacter vinelandii strain deregulated for biological nitrogen fixation. Microb. Cell Fact. 2020, 19, 107. [Google Scholar] [CrossRef]
- Cherkasov, N.; Ibhadon, A.O.; Fitzpatrick, P. A review of the existing and alternative methods for greener nitrogen fixation. Chemical Engineering and Processing: Process Intensif. 2015, 90, 24-33. [CrossRef]
- Rutledge, H.L.; Tezcan, F.A. Electron transfer in nitrogenase. Chem. Rev. 2020, 120, 5158–5193. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, Y.; Huang, K.; Wang, F.; Mei, Z. Molecular mechanism and agricultural application of the NifA–NifL system for nitrogen fixation. Int. J. Mol. Sci. 2023, 24, 907. [Google Scholar] [CrossRef] [PubMed]
- Inomura, K.; Bragg, J.; Follows, M.J. A quantitative analysis of the direct and indirect costs of nitrogen fixation: a model based on Azotobacter vinelandii. ISME J. 2017, 11, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Addo, M. A.; Dos Santos, P. C. Distribution of nitrogen-fixation genes in prokaryotes containing alternative nitrogenases. Chem. Bio. Chem. 2020, 21, 1749–1759. [Google Scholar] [CrossRef]
- Swain, H.; Abhijita, S. Nitrogen fixation and its improvement through genetic engineering. Journal of Global Biosciences 2013, 2, 98–112. [Google Scholar]
- Einsle, O.; Rees, D.C. Structural enzymology of nitrogenase enzymes. Chem. Rev. 2020, 120, 4969–5004. [Google Scholar] [CrossRef] [PubMed]
- Chanderban, M.; Hill, C.A.; Dhamad, A.E.; Lessner, D.J. Expression of V-nitrogenase and Fe-nitrogenase in Methanosarcina acetivorans is controlled by molybdenum, fixed nitrogen, and the expression of Mo-nitrogenase. Appl. Environ. Microbiol. 2023, 89, e01033-23. [Google Scholar] [CrossRef]
- Lery, L.M.; Bitar, M.; Costa, M.G.; Rössle, S.C.S.; Bisch, P.M. Unraveling the molecular mechanisms of nitrogenase conformational protection against oxygen in diazotrophic bacteria. BMC Genom. 2010, 11 (Suppl 5), S7. [Google Scholar] [CrossRef]
- Beato, V.M.; Rexach, J.; Navarro-Gochicoa, M.T.; Camacho-Cristóbal, J.J.; Herrera-Rodríguez, M.B.; González-Fontes, A. Boron deficiency increases expressions of asparagine synthetase, glutamate dehydrogenase and glutamine synthetase genes in tobacco roots irrespective of the nitrogen source. Soil. Sci. Plant Nutr. 2014, 60, 314–324. [Google Scholar] [CrossRef]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: current status and future prospects. J. Genet. Genom. 2022, 49, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Grzechowiak, M.; Sliwiak, J.; Link, A.; Ruszkowski, M. Legume-type glutamate dehydrogenase: Structure, activity, and inhibition studies. Int. J. Biol. Macromol. 2024, 278, 134648. [Google Scholar] [CrossRef]
- Luque-Almagro, V.M.; Gates, A.J.; Moreno-Vivián, C.; Ferguson, S.J.; Richardson, D.J.; Roldán, M.D. Bacterial nitrate assimilation: gene distribution and regulation. Biochem. Soc. Trans. 2011, 39, 1838–1843. [Google Scholar] [CrossRef]
- Mokhele, B.; Zhan, X.; Yang, G.; Zhang, X. Nitrogen assimilation in crop plants and its affecting factors. Can. J. Plant Sci. 2012, 92, 399–405. [Google Scholar] [CrossRef]
- Qiu, X.M.; Sun, Y.Y.; Ye, X.Y.; Li, Z.G. Signaling role of glutamate in plants. Front. Plant Sci. 2020, 10, 1743. [CrossRef]
- Malhotra, H.; Vandana Sharma, S.; Pandey, R. Phosphorus Nutrition: Plant Growth in Response to Deficiency and Excess. In: Plant Nutrients and Abiotic Stress Tolerance, Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B. (eds). Springer, Singapore, 2018, pp 171-190. [CrossRef]
- Khan, M.S.; Ahmad, E.; Zaidi, A.; Oves, M. Functional aspect of phosphate-solubilizing bacteria: importance in crop production. In: Bacteria in agrobiology: Crop productivity, Maheshwari, D., Saraf, M., Aeron, A. (eds), Springer, Berlin, Heidelberg, 2013, pp237-263. [CrossRef]
- Billah, M., Khan, M., Bano, A., Hassan, T. U., Munir, A., & Gurmani, A. R. Phosphorus and phosphate solubilizing bacteria: Keys for sustainable agriculture. Geomicrobiol. J. 2019, 36, 904-916. [CrossRef]
- Rodríguez, H.; Fraga, H. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17(4-5):319-339. [CrossRef]
- Bargaz, A.; Elhaissoufi, W.; Khourchi, S.; Benmrid, B.; et al. Benefits of phosphate solubilizing bacteria on belowground crop performance for improved crop acquisition of phosphorus. Microbiol. Res. 2021, 252, 126842. [Google Scholar] [CrossRef] [PubMed]
- Rawat, P.; Das, S.; Shankhdhar, D.; Shankhdhar, S. C. Phosphate-solubilizing microorganisms: Mechanism and their role in phosphate solubilization and uptake. J. Soil Sci. Plant Nutr. 2021, 21, 4968. [Google Scholar] [CrossRef]
- Adnan, M.; Shah, Z.; Fahad, S.; et al. Phosphate-solubilizing bacteria nullify the antagonistic effect of soil calcification on bioavailability of phosphorus in alkaline soils. Sci. Rep. 2017, 7, 16131. [Google Scholar] [CrossRef]
- Pande, A.; Pandey, P.; Mehra, S.; Singh, M.; Kaushik, S. Phenotypic and genotypic characterization of phosphate solubilizing bacteria and their efficiency on the growth of maize. J. Genet. Eng. Biotechnol. 2017, 15, 379-391. [CrossRef]
- Prabhu, N.; Borkar, S.; Garg, S. Chapter 11 - Phosphate solubilization by microorganisms: Overview, mechanisms, applications and advances. In: Advances in Biological Science Research, A Practical Approach, Meena, S.N & Naik, M.M (eds). Academic Press, 2019, pp161-176. [CrossRef]
- Vyas, P.; Gulati, A. Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbiol. 2009, 9, 174. [Google Scholar] [CrossRef]
- Suleman, M.; Yasmin, S.; Rasul, M.; et al. Phosphate solubilizing bacteria with glucose dehydrogenase gene for phosphorus uptake and beneficial effects on wheat. PLoS ONE 2018, 13, e0204408. [Google Scholar] [CrossRef]
- Stephen, J.; Shabanamol, S.; Rishad, K.S.; Jisha, M.S. Growth enhancement of rice (Oryza sativa) by phosphate solubilizing Gluconacetobacter sp. (MTCC 8368) and Burkholderia sp. (MTCC 8369) under greenhouse conditions. 3 Biotech 2015, 5, 831–837. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, Y.; Huang, S.; et al. Genome-based identification of phosphate-solubilizing capacities of soil bacterial isolates. AMB Expr. 2024, 14, 85. [Google Scholar] [CrossRef]
- Li, X.L.; Zhao, X.Q.; Dong, X.Y.; Ma, J.F.; Shen, R.F. Secretion of gluconic acid from Nguyenibacter sp. L1 is responsible for solubilization of aluminum phosphate. Front. Microbiol. 2021, 12, 784025. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.S.; Park, R.D.; Kim, Y.W.; et al. PQQ-dependent organic acid production and effect on common bean growth by Rhizobium tropici CIAT 899. J. Microbiol. Biotechnol. 2003, 13, 955–959. [Google Scholar]
- Mani, G.; Senthilkumar, R.; Venkatesan, K.; et al. Halophilic phosphate-solubilizing microbes (Priestia megaterium and Bacillus velezensis) isolated from Arabian Sea seamount sediments for plant growth promotion. Curr. Microbiol. 2024, 81, 405. [Google Scholar] [CrossRef]
- Wan, W.; Qin, Y.; Wu, H.; et al. Isolation and characterization of phosphorus solubilizing bacteria with multiple phosphorus sources utilizing capability and their potential for lead immobilization in soil. Front. Microbiol. 2020, 11, 752. [Google Scholar] [CrossRef]
- Pan, L.; Cai, B. Phosphate-solubilizing bacteria: Advances in their physiology, molecular mechanisms and microbial community effects. Microorganisms 2023, 11, 2904. [Google Scholar] [CrossRef]
- Cheng, Y.; Narayanan, M.; Shi, X.; et al. Phosphate-solubilizing bacteria: Their agroecological function and optimistic application for enhancing agro-productivity. Sci. Total Environ. 2023, 901, 166468. [Google Scholar] [CrossRef]
- Rawat, P.; Das, S.; Shankhdhar, D.; Shankhdhar, S.C. Phosphate-Solubilizing Microorganisms: Mechanism and Their Role in Phosphate Solubilization and Uptake. J. Soil Sci. Plant Nutr. 2020, 21, 49 - 68. [CrossRef]
- Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi - current perspective. Arch. Agron. Soil Sci. 2009, 56, 73–98. [Google Scholar] [CrossRef]
- Lim, B.L.; Yeung, P.; Cheng, C.; Hill, J.E. Distribution and diversity of phytate-mineralizing bacteria, ISME J. 2007, 1, 321-330. [CrossRef]
- Choi, Y.M.; Suh, H.J.; Kim, J.M. Purification and properties of extracellular phytase from Bacillus sp. KHU-10. J. Protein Chem. 2001, 20, 287-292. [CrossRef]
- Kramer, S.; Green, D.M. Acid and alkaline phosphatase dynamics and their relationship to soil microclimate in a semiarid woodland. Soil Biol. Biochem. 2000, 32, 179–188. [Google Scholar] [CrossRef]
- Behera, B.C., Singdevsachan, S.K. Mishra, R.R., Dutta, S.K., Thatoi, H.N. Diversity, mechanism and biotechnology of phosphate solubilising microorganism in mangrove - A review. Biocatal. Agric. Biotechnol. 2014, 3, 97-110. [CrossRef]
- Panhwar, Q.A.; Naher, U.A.; Jusop, S.; Othman, R.; Latif, M.A.; Ismail, M.R. Biochemical and molecular characterization of potential phosphate-solubilizing bacteria in acid sulfate soils and their beneficial effects on rice growth. PLoS ONE 2014, 9, e97241. [Google Scholar] [CrossRef]
- Qureshi, M. A., Ahmad, Z. A., Akhtar, N., Iqbal, A., Mujeeb, F., & Shakir, M. A. Role of phosphate solubilizing bacteria (PSB) in enhancing P availability and promoting cotton growth. J. Anim. Plant Sci. 2012, 22, 204-210.
- Salsabila, N.; Fitriatin, B. N.; Hindersah, R. The role of phosphate-solubilizing microorganisms in soil health and phosphorus cycle: A review. IJLSAR 2023, 2, 281-287. [CrossRef]
- Alberton, D.; Valdameri, G.; Moure, V.R.; Monteiro, R.A.; et al. What did we learn from plant growth-promoting rhizobacteria (PGPR)-grass associations studies through proteomic and metabolomic approaches. Front. Sustain. Food Syst. 2020, 4, 607343. [Google Scholar] [CrossRef]
- Shan, X.; Yan, J.; Xie, D. Comparison of phytohormone signaling mechanisms. Curr. Opin. Plant Biol. 2012, 15, 84–91. [Google Scholar] [CrossRef]
- Drogue, B.; Combes-Meynet, E.; Moënne-Loccoz, Y.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Control of the cooperation between plant growth-promoting rhizobacteria and crops by rhizosphere signals. In: Molecular microbial ecology of the rhizosphere, 1st ed., de Bruijn, F.J. (ed.), Hoboken, New Jersey, USA, 2013, John Wiley & Sons Ltd., pp 279-293. [CrossRef]
- Venturi, V.; Keel, C. Signaling in the rhizosphere. Trends Plant Sci. 2026, 21, 187–198. [Google Scholar] [CrossRef]
- Agudelo-Morales, C.E.; Lerma, T.A.; Martínez, J.M.; Palencia, M.; Combatt, E.M. Phytohormones and plant growth regulators: A review. J. Sci. Technol. Appl. 2021, 10, 27–65. [Google Scholar] [CrossRef]
- Ortíz-Castro, R., Contreras-Cornejo, H. A., Macías-Rodríguez, L., & López-Bucio, J. The role of microbial signals in plant growth and development. Plant Signal. Behav. 2009, 4, 701-712. [CrossRef]
- Emenecker, R.J.; Strader, L.C. Auxin-abscisic acid interactions in plant growth and development. Biomolecules 2020, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V., Bhatt, I.D. y Nandi, S.K. Chapter 20 - Role and regulation of auxin signaling in abiotic stress tolerance. In: Plant signaling molecules, Khan, M.I.R., Reddy, P.S., Ferrante, A. & Khan, N.A. (Eds.). Woodhead Publishing, 2019, pp: 319-331. [CrossRef]
- Jing, H.; Wilkinson, E.G.; Sageman-Furnas, K.; Strader, L.C. Auxin and abiotic stress responses. J. Exp. Bot. 2023, 74, 7000–7014. [Google Scholar] [CrossRef] [PubMed]
- Duca, D.; Lorv, J.; Patten, C.L.; et al. Indole-3-acetic acid in plant-microbe interactions. Antonie van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Bano, A.; et al. Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: consequences for changing environment. Environ. Sci. Pollut. Res. 2015, 22, 4907–4921. [Google Scholar] [CrossRef]
- Morel, M.A. y Castro-Sowinski, S. The complex molecular signaling network in microbe-plant interaction. In: Plant microbe symbiosis: Fundamentals and advances, Arora, N. (eds), Springer, New Delhi, 2013, pp 169-199. [CrossRef]
- Di, D.W.; Zhang, C.; Luo, P.; et al. The biosynthesis of auxin: how many paths truly lead to IAA. J. Plant Growth Regul. 2016, 78, 275–285. [Google Scholar] [CrossRef]
- Fenn, M.A.; Giovannoni, J.J. Phytohormones in fruit development and maturation. Plant J. 2021, 105, 446–458. [Google Scholar] [CrossRef]
- Chandler, J.W. The hormonal regulation of flower development. J. Plant Growth Regul. 2011, 30, 242–254. [Google Scholar] [CrossRef]
- Roosjen, M.; Paque, S.; Weijers, D. Auxin response factors: output control in auxin biology. J. Exp. Bot. 2018, 69, 179–188. [Google Scholar] [CrossRef]
- Gomes, G.L.B.; Scortecci, K.C. Auxin and its role in plant development: structure, signalling, regulation and response mechanisms. Plant Biol. 2021, 23, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.W. Auxin response factors. Plant, Cell & Environment 2015, 39, 1014-1028. [CrossRef]
- Bouzroud, S.; Gouiaa, S.; Nan Hu, N.; et al. Auxin Response Factors (ARFs) are potential mediators of auxin action in tomato response to biotic and abiotic stress (Solanum lycopersicum). PloS one 2018, 13, e0193517. [Google Scholar] [CrossRef] [PubMed]
- Li, S.B.; Xie, Z.Z.; Hu, C.G.; Zhang, J.Z. A review of auxin response factors (ARF’s) in plants. Front. Plant Sci. 2016, 7, 47. [Google Scholar] [CrossRef]
- Hartmann, A.; Klink, S.; Rothballer, M. Plant growth promotion and induction of systemic tolerance to drought and salt stress of plants by quorum sensing auto-inducers of the N-acyl-homoserine lactone type: recent developments. Front. Plant Sci. 2021, 12, 683546. [Google Scholar] [CrossRef]
- Verma, S.; Negi, N.P. Pareek, S., Mudgal, G.; Kumar, D. Auxin response factors in plant adaptation to drought and salinity stress. Physiol. Plant. 2022, 174, e13714. [Google Scholar] [CrossRef]
- Bao, D.; Chang, S.; Li, X.; Qi, Y. Advances in the study of auxin early response genes: Aux/IAA, GH3, and SAUR. Crop J. 2024, 12, 964–978. [Google Scholar] [CrossRef]
- Fanai, A.; Bohia, B.; Lalremruati, F.; et al. Plant growth promoting bacteria (PGPB)-induced plant adaptations to stresses: an updated review. Peer J. 2024, 12, e17882. [Google Scholar] [CrossRef]
- Sevim, G.; Ozdemir-Kocak, F.; Unal, D. Chapter 15 - Hormonal signaling molecules triggered by plant growth-promoting bacteria. In: Phytohormones and Stress Responsive Secondary Metabolites, Ozturk, M., Bath R.A., Asharf M., Policarpo-Tonelli F.M., Unal B. T., Dar G. H. (eds). Academic Press, 2023, pp 187-196. [CrossRef]
- Kang, S.M.; Khan, A. L., You, Y.-H., et al. Gibberellin production by newly isolated strain Leifsonia soli se134 and its potential to promote plant growth. J. Microbiol. Biotechnol. 2014, 24, 106-112. [CrossRef]
- Orozco-Mosqueda, M.C.; Santoyo, G.; Glick, B.R. Recent advances in the bacterial phytohormone modulation of plant growth. Plants 2023, 12, 606. [Google Scholar] [CrossRef] [PubMed]
- Castro-Camba, R.; Sánchez, C.; Vidal, N.; Vielba, J.M. Plant development and crop yield: The role of gibberellins. Plants 2022, 11, 2650. [Google Scholar] [CrossRef]
- Belimov, A.A.; Dodd, I.C.; Safronova, V.I.; et al. Abscisic acid metabolizing rhizobacteria decrease ABA concentrations in planta and alter plant growth. Plant Physiol. Biochem. 2014, 74, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.C.; Bottini, R.; Piccoli, P. Role of Abscisic Acid Producing PGPR in Sustainable Agriculture. In: Bacterial Metabolites in Sustainable Agroecosystem. Sustainable Development and Biodiversity vol 12, Maheshwari, D. (eds), Springer Cham., 2015, pp259-282. [CrossRef]
- Khan, N.; Bano, A.; Ali, S.; et al. Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regul. 2020, 90, 189–203. [Google Scholar] [CrossRef]
- Binder, B.M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, A.; Shemansky, J.M.; Chang, C.; et al. History of research on the plant hormone ethylene. J Plant Growth Regul. 2015, 34, 809–827. [Google Scholar] [CrossRef]
- Ju, C.; Van de Poel, B.; Cooper, E.; et al. Conservation of ethylene as a plant hormone over 450 million years of evolution. Nature Plants 2015, 1, 14004. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Rossi, M.J.; Glick, B.R. Ethylene and 1-aminocyclopropane-1-carboxylate (ACC) in plant–bacterial interactions. Front. Plant Sci. 2018, 9, 114. [Google Scholar] [CrossRef]
- Shekhawat, K., Fröhlich, K., García-Ramírez, G. X., Trapp, M. A., & Hirt, H. Ethylene: A master regulator of plant–microbe interactions under abiotic stresses. Cells 2023, 12, 31. [CrossRef]
- Ravanbakhsh, M.; Sasidharan, R.; Voesenek, L.A.C.J.; et al. Microbial modulation of plant ethylene signaling: ecological and evolutionary consequences. Microbiome 2018, 6, 52. [Google Scholar] [CrossRef]
- Esquivel-Cote, R.; Gavilanes-Ruiz, M.; Cruz-Ortega, R.; Huante, P. Importancia agrobiotecnológica de la enzima ACC desaminasa en rizobacterias, una revisión. Rev. Fitotec. Mex. 2013, 36, 251–258. [Google Scholar] [CrossRef]
- Singh, R.P.; Ma, Y.; Shadan, A. Perspective of ACC-deaminase producing bacteria in stress agriculture. J. Biotechnol. 2022, 352, 36–46. [Google Scholar] [CrossRef]
- Effmert, U.; Kalderás, J.; Warnke, R.; et al. Volatile mediated interactions between bacteria and fungi in the soil. J. Chem. Ecol. 2012, 38, 665–703. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Hung, R.; Lee, S.; Padhi, S. 18 Fungal and Bacterial Volatile Organic Compounds: An Overview and Their Role as Ecological Signaling Agents. In: Hock, B. (eds) Fungal Associations. The Mycota 2012 vol 9. Springer, Berlin, Heidelberg. [CrossRef]
- Minerdi, D.; Maggini, V.; Fani, R. Volatile organic compounds: from figurants to leading actors in fungal symbiosis. FEMS Microbiol. Ecol. 2021, 97, fiab067. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Carrillo, R.; Guerra Ramírez, P. Pseudomonas spp. benéficas en la agricultura. Rev. Mexicana Cienc. Agric. 2022, 13, 715–725. [Google Scholar] [CrossRef]
- Moreno-Valencia, F.D.; Plascencia-Espinosa, M.Á.; Morales-García, Y.E.; Muñoz-Rojas, J. Selection and effect of Plant Growth-Promoting Bacteria on pine seedlings (Pinus montezumae and Pinus patula). Life 2024, 14, 1320. [Google Scholar] [CrossRef]
- Sabu, R.; Aswani, R.; Jishma, P.; et al. Plant growth promoting endophytic Serratia sp. zob14 protecting ginger from fungal pathogens. Proc. Natl. Acad. Sci., India, Sect. B Biol. Sci. 2019, 89, 213-220. [CrossRef]
- Kankariya, R.A.; Chaudhari, A.B.; Gavit, P.M.; Dandi, N.D. 2,4-Diacetylphloroglucinol: A novel biotech bioactive compound for agriculture. In: Microbial interventions in agriculture and environment, Singh, D., Gupta, V., Prabha, R. (eds). Springer, Singapore, 2019, pp491-452. [CrossRef]
- Jani, J.; Parvez, N.; Mehta, D. Metabolites of Pseudomonads: A new avenue of plant health management. In: New horizons in insect science: Towards sustainable pest management, Chakravarthy, A. (eds) Springer, New Delhi, 2015, pp61-69. [CrossRef]
- Lahiri, D.; Nag, M.; Sayyed, R.Z.; Gafur, A.; Ansari, M.J.; Ray, R.R. PGPR in biofilm formation and antibiotic production. In: Antifungal metabolites of rhizobacteria for sustainable agriculture, Fungal Biology series, Sayyed, R., Singh, A., Ilyas, N. (eds) Springer, Cham., 2022, pp 65-82. [CrossRef]
- Kour, D.; Negi, R.; Khan, S.S.; et al. Microbes mediated induced systemic response in plants: A review. Plant Stress 2024, 11, 100334. [Google Scholar] [CrossRef]
- Singh, A.A, Singh, & A.K. Role of bacterial Quorum Sensing in plant growth promotion. World J. Microbiol. Biotechnol. 2025, 41, 18. [CrossRef]
- Tyagi, S.; Mulla, S.I.; Lee, K.J.; Chae, J.C.; Shukla, P. VOCs-mediated hormonal signaling and crosstalk with plant growth promoting microbes. Crit. Rev. Biotechnol. 2018, 38, 1277–1296. [Google Scholar] [CrossRef]
- Liu, X.M.; Zhang, H. The effects of bacterial volatile emissions on plant abiotic stress tolerance. Front. Plant Sci. 2015, 6, 774. [Google Scholar] [CrossRef]
- Laller, R.; Khosla, P.K.; Negi, N.; et al. Bacterial volatiles as PGPRs: Inducing plant defense mechanisms during stress periods. S. Afr. J. Bot. 2023, 159, 131–139. [Google Scholar] [CrossRef]
- Plyuta, V.; Lipasova, V.; Popova, A.; et al. Influence of volatile organic compounds emitted by Pseudomonas and Serratia strains on Agrobacterium tumefaciens biofilms. APMIS 2016, 124, 586–594. [Google Scholar] [CrossRef]
- Wang, J.; Raza, W.; Jiang, G.; et al. Bacterial volatile organic compounds attenuate pathogen virulence via evolutionary trade-offs. ISME J. 2023, 17, 443–452. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Dubery, I.A. Profiling of volatile organic compounds from four Plant Growth-Promoting Rhizobacteria by SPME–GC–MS: A metabolomics study. Metabolites 2022, 12, 763. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Schikora, A. AHL-priming for enhanced resistance as a tool in sustainable agriculture. FEMS Microbiol. Ecol. 2020, 96, fiaa226. [Google Scholar] [CrossRef]
- Chamkhi, I.; El Omari, N.; Benali, T.; Bouyahya, A. Quorum sensing and plant-bacteria interaction: role of quorum sensing in the rhizobacterial community colonization in the rhizosphere. In: Quorum Sensing: Microbial Rules of Life, vol 1374, Dhiman S. S. (eds), ACS Publishers, Washington D.C. USA, 2020, pp. 139-153. [CrossRef]
- Cortez, M.; Handy, D.; Headlee, A.; et al. Quorum Sensing in the rhizosphere. In: Microbial cross-talk in the rhizosphere. Rhizosphere Biology, Horwitz, B.A., Mukherjee, P.K. (eds), Springer, Singapore, 2022, pp 99-134. [CrossRef]
- Hartmann, A. Quorum sensing N-acyl-homoserine lactone signal molecules of plant beneficial Gram-negative rhizobacteria support plant growth and resistance to pathogens. Rhizosphere 2020, 16, 100258. [Google Scholar] [CrossRef]
- Hanif, M.K.; Malik, K.A.; Hameed, S.; Saddique, M. J.; et al. Growth stimulatory effect of AHL producing Serratia spp. from potato on homologous and non-homologous host plants. Microbiol. Res. 2020, 238, 126506. [CrossRef]
- Viswanath, G.; Sekar, J.; Ramalingam, P.V. Detection of diverse N-Acyl Homoserine Lactone signalling molecules among bacteria associated with rice rhizosphere. Curr. Microbiol. 2020, 77, 3480–3491. [Google Scholar] [CrossRef]
- Nawaz, M.S.; Arshad, A.; Rajput, L.; Fatima, K.; Ullah, S.; Ahmad, M.; Imran, A. Growth-stimulatory effect of quorum sensing signal molecule N-acyl-homoserine lactone-producing multi-trait Aeromonas spp. on wheat genotypes under salt stress. Front. Microbiol. 2020, 11, 553621. [Google Scholar] [CrossRef]
- Ramírez-Mata, A.; Fernández-Domínguez, I.J.; Nuñez-Reza, K.J.; Xiqui-Vázquez, M.L.; Baca, B.E. Redes de señalización en la producción de biopelículas en bacterias: quorum sensing, di-GMPc y óxido nítrico. Rev. Argent. Microbiol. 2014, 46, 242-255. [CrossRef]
- Khan, M.; Bhargava, P.; Goel, R. Quorum sensing molecules of rhizobacteria: A Trigger for developing systemic resistance in plants. In: Plant Growth Promoting Rhizobacteria for sustainable stress management. Microorganisms for Sustainability vol 12, Sayyed, R., Arora, N., Reddy, M. (eds), Springer, Singapore, 2019, pp117-138. [CrossRef]
- Lahiri, D.; Nag, M.; Sayyed, R.Z.; Gafur, A.; Ansari, M.J.; Ray, R.R. PGPR in biofilm formation and antibiotic production. In: Antifungal metabolites of rhizobacteria for sustainable agriculture. Fungal Biology Series, Sayyed, R., Singh, A., Ilyas, N. (eds) Springer, Cham., 2022, pp 65-82. [CrossRef]
- Kenawy, A.; Dailin, D.J.; Abo-Zaid, G.A.; et al. Biosynthesis of antibiotics by PGPR and their roles in biocontrol of plant diseases. In: Plant Growth Promoting Rhizobacteria for sustainable stress management. Microorganisms for Sustainability vol. 13, Sayyed, R. (eds), Springer, Singapore, 2019, pp1-35. [CrossRef]
- Mehmood, A.; Liu, G.; Wang, X.; Meng, G.; Wang, C.; Liu, Y. Fungal Quorum-Sensing molecules and inhibitors with potential antifungal activity: A review. Molecules 2019, 24, 1950. [Google Scholar] [CrossRef]
- Albuquerque, P.; Casadevall, A. Quorum sensing in fungi – a review. Med. Mycol. 2012, 50, 337–345. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, H.; Xu, S.; et al. Bacterial-fungal interactions under agricultural settings: from physical to chemical interactions. Stress Biol. 2022, 2, 22. [Google Scholar] [CrossRef]
- Khalid, S., Keller, N.P. Chemical signals driving bacterial–fungal interactions. "Environ. Microbiol. 2021, 23, 1334-1347. [CrossRef]
- Khashi u Rahman, M.; Zhou, X.; Wu, F. The role of root exudates, CMNs, and VOCs in plant–plant interaction. J. Plant Interact. 2019, 14, 630–636. [Google Scholar] [CrossRef]
- Wang, Y.; Wenqing LI, W.; Binghai Du, B.; LI, H. Effect of biochar applied with plant growth-promoting rhizobacteria (PGPR) on soil microbial community composition and nitrogen utilization in tomato. Pedosphere 2021, 31, 872-881. [CrossRef]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, A.; Marczak, M.; Ludwikow, A. Mitogen-activated protein kinase cascades in plant hormone signaling. Front. Plant Sci. 2018, 9, 1387. [Google Scholar] [CrossRef]
- Jalmi, S.K.; Sinha, A.K. ROS mediated MAPK signaling in abiotic and biotic stress-striking similarities and differences. Front. Plant Sci. 2015, 6, 769. [Google Scholar] [CrossRef] [PubMed]
- Krysan, P.J.; Colcombet, J. Cellular complexity in MAPK signaling in plants: Questions and emerging tools to answer them. Front. Plant Sci. 2018, 9, 1674. [CrossRef]
- Hamel, L.P.; Nicole, M.C.; Sritubtim, S.; et al. Ancient signals: comparative genomics of plant MAPK and MAPKK gene families. Trends Plant Sci. 2006, 11, 192–198. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, S. Mitogen-activated protein kinase cascades in plant signaling. J. Integr. Plant Biol. 2022, 64, 301–341. [Google Scholar] [CrossRef] [PubMed]
- Danquah, A.; De Zélicourt, A.; Colcombet, J.; Hirt, H. The role of ABA and MAPK signaling pathways in plant abiotic stress responses. Biotechnol. Adv. 2014, 32, 40–52. [Google Scholar] [CrossRef]
- Bigeard, J.; Hirt, H. Nuclear signaling of plant MAPKs. Front. Plant Sci. 2018, 9, 469. [Google Scholar] [CrossRef]
- Wogene, S. Tibor J. Zoltán M. Unveiling the significance of rhizosphere: Implications for plant growth, stress response, and sustainable agriculture, Plant Physiol. Biochem. 2024, Volume 206 108290. [CrossRef]
- Jamil, F.; Mukhtar, H.; Fouillaud, M.; Dufossé, L. Rhizosphere Signaling: Insights into Plant-Rhizomicrobiome Interactions for Sustainable Agronomy. Microorganisms 2022, Apr 25;10, 899. [CrossRef]
- Pardo Díaz, S.; Mazo Molina, D.C.; Rojas Tapias, D.F. Bacterias promotoras del crecimiento vegetal: filogenia, microbioma, y perspectivas. In: Rol de las bacterias promotoras de crecimiento vegetal en sistemas de agricultura sostenible. Bonilla Buitrago, R. González de Bashan, L. E., & Pedraza, R. O. (eds). Corporación Colombiana de Investigación Agropecuaria – AGROSAVIA, Cundinamarca, Colombia, 2021, pp 46-77. [CrossRef]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; et al. Plant health: feedback effect of root exudates-rhizobiome interactions. Appl. Microbiol. Biotechnol. 2019, 103, 1155–1166. [Google Scholar] [CrossRef]
- Shah, W.U.H.; Lu, Y.; Liu, J.; Rehman, A.; Yasmeen, R. The impact of climate change and production technology heterogeneity on China's agricultural total factor productivity and production efficiency. Sci. Tot. Environ. 2024, 907, 168027. [Google Scholar] [CrossRef]
- Pervaiz, Z. H., Iqbal, J., Zhang, Q., Chen, D., Wei, H., & Saleem, M. Continuous cropping alters multiple biotic and abiotic indicators of soil health. Soil Systems 2020, 4, 59. [CrossRef]
- Zandalinas, S.I.; Mittler, R. Plant responses to multifactorial stress combination. New Phytol. 2022, 234, 1161–1167. [Google Scholar] [CrossRef]
- Jeyasri, R.; Muthuramalingam, P.; Satish, L.; et al. An overview of abiotic stress in cereal crops: Negative Impacts, regulation, biotechnology and integrated omics. Plants 2021, 10, 1472. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez Guzman, M.; Cellini, F.; Fotopoulos, V.; Balestrini, R.; Arbona, V. New approaches to improve crop tolerance to biotic and abiotic stresses. Physiol. Plant. 2022, 174, e13547. [Google Scholar] [CrossRef] [PubMed]
- Teshome, D. T.; Zharare, G. E.; Naidoo, S. The threat of the combined effect of biotic and abiotic stress factors in forestry under a changing climate. Front. Plant Sci. 2020, 11, 601009. [Google Scholar] [CrossRef]
- Li, M.; Li, X.; Xue, D.; et al. Enhanced plant growth through composite inoculation of phosphate-solubilizing bacteria: Insights from plate and soil experiments. Agronomy 2024, 14, 2461. [Google Scholar] [CrossRef]
- Liang, J.L.; Liu, J.; Jia, P.; et al. Novel phosphate-solubilizing bacteria enhance soil phosphorus cycling following ecological restoration of land degraded by mining. ISME J. 2020, 14, 1600–1613. [Google Scholar] [CrossRef]
- Bashir, Z.; Hamid, B.; Yatoo, A.M.; et al. Phosphorus solubilizing microorganisms: An eco-friendly approach for sustainable plant health and bioremediation. J. Soil Sci. Plant Nutr. 2024, 24, 6838–6854. [Google Scholar] [CrossRef]
- Liu-Xu, L. González-Hernández, A.I. Camañes, G. Vicedo, B. Scalschi, L. Llorens, E. Harnessing Green Helpers: Nitrogen-Fixing Bacteria and Other Beneficial Microorganisms in Plant–Microbe Interactions for Sustainable Agriculture. Horticulturae 2024, 10, 621. [CrossRef]
- Dobbelaere, S.; Vanderleyden, J.; Okon, Y. Plant growth-promoting effects of diazotrophs in the rhizosphere. Crit. Rev. Plant Sci. 2003, 22, 107–149. [Google Scholar] [CrossRef]
- Ghosh, A.; Sah, D.; Chakraborty, M.; Rai, J.P.N. Mechanism and application of bacterial exopolysaccharides: An advanced approach for sustainable heavy metal abolition from soil. Carbohydr. Res. 2024, 544, 109247. [Google Scholar] [CrossRef]
- Thiebaut, F., Urquiaga, M.C. d. O., Rosman, A. C., da Silva, M. L., & Hemerly, A. S. The impact of non-nodulating diazotrophic bacteria in agriculture: understanding the molecular mechanisms that benefit crops. Int. J. Mol. Sci. 2022, 23, 11301. [CrossRef]
- Cuevas, J., Daliakopoulos, I. N., del Moral, F., Hueso, J. J., & Tsanis, I. K. A review of soil-improving cropping systems for soil salinization. Agronomy 2019, 9, 295. [CrossRef]
- Mirza, B.S.; McGlinn, D.J.; Bohannan, B.J.M.; et al. Diazotrophs show signs of restoration in amazon rain forest soils with ecosystem rehabilitation. Appl. Environ. Microbiol. 2020, 86, e00195-20. [Google Scholar] [CrossRef]
- Munir, N.; Hanif, M.; Abideen, Z.; Sohail, M.; et al. Mechanisms and strategies of plant microbiome interactions to mitigate abiotic stresses. Agronomy 2022, 12, 2069. [Google Scholar] [CrossRef]
- Acosta-Motos, J. R., Ortuño, M. F., Bernal-Vicente, A., Diaz-Vivancos, P., Sanchez-Blanco, M. J., & Hernandez, J. A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [CrossRef]
- Stavi, I.; Thevs, N.; Priori, S. Soil salinity and sodicity in drylands: A review of causes, effects, monitoring, and restoration measures. Front. Environ. Sci. 2021, 9, 712831. [Google Scholar] [CrossRef]
- Okur, B.; Örçen, N. Soil salinization and climate change. In: Climate change and soil interactions. Prasad, M.N.V., Pietrzykowski, M. (Eds.), Elsevier, Amsterdam, 2020, pp 331-350. [CrossRef]
- Madline, A.; Benidire, L.; Boularbah, A. Alleviation of salinity and metal stress using plant growth-promoting rhizobacteria isolated from semiarid Moroccan copper-mine soils. Environ. Sci. Pollut. Res. 2021, 28, 67185–67202. [Google Scholar] [CrossRef]
- Etesami, H.; Noori, F. Soil salinity as a challenge for sustainable agriculture and bacterial-mediated alleviation of salinity stress in crop plants. In: Saline soil-based agriculture by halotolerant microorganisms, Kumar, M., Etesami, H., Kumar, V. (eds), Springer, Singapore, 2019, pp 1-22. [CrossRef]
- Shultana, R., Zuan, A. T. K., Naher, U. A., et al. The PGPR mechanisms of salt stress adaptation and plant growth promotion. Agronomy 2022, 12, 2266. [CrossRef]
- Hnini, M., Rabeh, K., & Oubohssaine, M. (). Interactions between beneficial soil microorganisms (PGPR and AMF) and host plants for environmental restoration: A systematic review. Plant Stress 2024, 11, 100391. [CrossRef]
- Sharma, I., Sharma, S., Sharma, V., et al. (). PGPR-Enabled bioremediation of pesticide and heavy metal-contaminated soil: A review of recent advances and emerging challenges. Chemosphere 2024, 362, 142678. [CrossRef]
- Gupta, R.; Khan, F.; Alqahtani, F.M.; et al. Plant Growth–Promoting Rhizobacteria (PGPR) assisted bioremediation of heavy metal toxicity. Appl. Biochem. Biotechnol. 2024, 196, 2928–2956. [Google Scholar] [CrossRef]
- Sobariu, D.L.; Fertu, D.I.T.; Diaconu, M.; et al. Rhizobacteria and plant symbiosis in heavy metal uptake and its implications for soil bioremediation. New Biotechnol. 2017, 39, 125–134. [Google Scholar] [CrossRef]
- Hawkes, S.J. What is a" heavy metal" Journal of chemical education 1997, 74, 1374. [CrossRef]
- Fageria, N.K.; Baligar, V.C.; Clark, R.B. Micronutrients in crop production. Adv. Agron. 2002, 77, 185–268. [Google Scholar] [CrossRef]
- Banuelos, G. S., & Ajwa, H. A. Trace elements in soils and plants: an overview. J. Environ. Sci. Health 1999, Part A 34, 951-974. [CrossRef]
- Zhao, H.; Wu, Y.; Lan, X.; et al. Comprehensive assessment of harmful heavy metals in contaminated soil in order to score pollution level. Sci. Rep. 2022, 12, 3552. [Google Scholar] [CrossRef]
- Mondal, S.; Mukherjee, S.K.; Hossain, S.T. Exploration of plant growth promoting rhizobacteria (PGPRs) for heavy metal bioremediation and environmental sustainability: Recent advances and future prospects. In: Modern approaches in waste bioremediation, Shah, M.P. (eds), Springer Cham., 2023, pp 29-55. [CrossRef]
- Ortiz, J.; Dias, N.; Alvarado, R.; Soto, J.; et al. N-acyl homoserine lactones (AHLs) type signal molecules produced by rhizobacteria associated with plants that growing in a metal (oids) contaminated soil: A catalyst for plant growth. Microbiol. Res. 2024, 281, 127606. [Google Scholar] [CrossRef] [PubMed]
- Alves, A. R., Yin, Q., Oliveira, R. S., Silva, E. F., & Novo, L. A. Plant growth-promoting bacteria in phytoremediation of metal-polluted soils: Current knowledge and future directions. Sci. Total Environ. 2022, 838, 156435. [CrossRef]
- Pandey, S.; Ghosh, P.K.; Ghosh, S.; et al. Role of heavy metal resistant Ochrobactrum sp. and Bacillus spp. strains in bioremediation of a rice cultivar and their PGPR like activities. J. Microbiol. 2013, 51, 11–17. [Google Scholar] [CrossRef]
- Hou, J.; Liu, W.; Wang, B.; Wang, Q.; Luo, Y.; Franks, A.E. PGPR enhanced phytoremediation of petroleum contaminated soil and rhizosphere microbial community response. Chemosphere 2015, 138, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Riseh, R.S.; Vazvani, M.G.; Hajabdollahi, N.; et al. Bioremediation of heavy metals by rhizobacteria. Appl. Biochem. Biotechnol. 2023, 195, 4689–4711. [Google Scholar] [CrossRef] [PubMed]
- Chitara, M.K.; Chauhan, S.; Singh, R.P. Bioremediation of polluted soil by using plant growth–promoting rhizobacteria. In: Microbial rejuvenation of polluted environment. Microorganisms for sustainability vol. 25, Panpatte, D.G., Jhala, Y.K. (eds), Springer, Singapore, 2021, pp203-226. [CrossRef]
- Food and Agriculture Organization (FAO). El estado de los recursos de tierras y aguas del mundo para la alimentación y la agricultura - Sistemas al límite, Informe de síntesis. FAO, Rome, Italy, 2021, 1-86. [CrossRef]
- Gomiero, T. Soil degradation, land scarcity and food security: Reviewing a complex challenge. Sustainability 2016, 8, 281. [Google Scholar] [CrossRef]
- United Nations Organization. 2023 United Nations Sustainable Development Goals (SDGs) Summit. Available online: https://www.un.org/sustainabledevelopment/es/2023/05/un-2023-sdg-summit/ (accessed on 22-05- 2024).
- Sharma, I.P.; Chandra, S.; Kumar, N.; Chandra, D. PGPR: Heart of soil and their role in soil fertility. In: Agriculturally important microbes for sustainable agriculture, Meena, V., Mishra, P., Bisht, J., Pattanayak, A. (eds). Springer, Singapore, 2017, pp 51-67. [CrossRef]
- Hasan, A.; Tabassum, B.; Hashim, M.; Khan, N. Role of Plant Growth Promoting Rhizobacteria (PGPR) as a plant growth enhancer for sustainable agriculture: A review. Bacteria 2024, 3, 59–75. [Google Scholar] [CrossRef]
- Mitra, D.; Pellegrini, M.; Guerra-Sierra, B.E. Interaction between plants and Growth-Promoting Rhizobacteria (PGPR) for sustainable development. Bacteria 2024, 3, 136–140. [Google Scholar] [CrossRef]
- Patel, P.; Sayyed, R.Z.; Patel, H. PGPR: A sustainable agricultural mitigator for stressed agro-environments. In: Plant growth promoting microorganisms of arid region, Mawar, R., Sayyed, R.Z., Sharma, S.K., Sattiraju, K.S. (eds). Springer, Singapore, 2023, pp303-318. [CrossRef]
- Morales-García, Y.E.; Juárez-Hernández, D.; Hernández-Tenorio, A.L.; et al. Inoculante de segunda generación para incrementar el crecimiento y salud de plantas de jardín. AyTBUAP 2020, 5, 136-154. [CrossRef]
- Menguala, C.; Schoebitz, M.; Azcón, R. y Roldán, A. Microbial inoculants and organic amendment improves plant establishment and soil rehabilitation under semiarid conditions. J. Environ. Manag. 2014, 134, 1–7. [Google Scholar] [CrossRef]
- Saeed, M.; Ilyas, N.; Jayachandran, K.; Shabir, S.; et al. Advances in biochar and PGPR engineering system for hydrocarbon degradation: A promising strategy for environmental remediation. Environ. Pollut. 2022, 305, 119282. [Google Scholar] [CrossRef]
- Anbuganesan, V.; Vishnupradeep, R.; Bruno, L.B.; Sharmila, K.; Freitas, H.; Rajkumar, M. Combined Application of Biochar and Plant Growth-Promoting Rhizobacteria Improves Heavy Metal and Drought Stress Tolerance in Zea mays. Plants 2024, 13, 1143. [Google Scholar] [CrossRef]
- Abdelkrim, S.; Jebara, S.H.; Saadani, O.; Abid, G.; et al. In situ effects of Lathyrus sativus-PGPR to remediate and restore quality and fertility of Pb and Cd polluted soils. Ecotoxicol. Environ. Saf. 2020, 192, 110260. [Google Scholar] [CrossRef]
- Hussain, F.; Hussain, I.; Khan, A.H.A.; Muhammad, Y.S.; et al. Combined application of biochar, compost, and bacterial consortia with Italian ryegrass enhanced phytoremediation of petroleum hydrocarbon contaminated soil. Environ. Exp. Bot. 2018, 153, 80–88. [Google Scholar] [CrossRef]
- Ali, M.H.; Khan, M.I.; Bashir, S.; Azam, M.; et al. Biochar and Bacillus sp. MN54 assisted phytoremediation of diesel and plant growth promotion of maize in hydrocarbons contaminated soil. Agronomy 2021, 11, 1795. [Google Scholar] [CrossRef]
- Minnikova, T.; Kolesnikov, S.; Minin, N.; Gorovtsov, A.; Vasilchenko, N.; Chistyakov, V. The influence of remediation with Bacillus and Paenibacillus strains and Biochar on the biological activity of petroleum-hydrocarbon-contaminated haplic chernozem. Agriculture 2023, 13(3):719. [CrossRef]
- Fang, X.; Zhang, M.; Zheng, P.; et al. Biochar bacteria-plant combined potential for remediation of oil-contaminated soil. Front. Microbiol. 2024, 15, 1343366. [Google Scholar] [CrossRef]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.D.; Rashwan, E.; Kamara, M.M. Synergistic effect of biochar and plant growth promoting rhizobacteria on alleviation of water deficit in rice plants under salt-affected soil. Agronomy 2019, 9, 847. [CrossRef]
- Nehela, Y.; Mazrou, Y.S.A.; Alshaal, T.; Rady, A.M.S.; et al. The integrated amendment of sodic-saline soils using biochar and Plant Growth-Promoting Rhizobacteria enhances maize (Zea mays L.) resilience to water salinity. Plants 2021, 10, 1960. [CrossRef]
- Malik, L.; Sanaullah, M.; Mahmood, F.; Hussain, S.; Shahzad, T. Co-application of biochar and salt tolerant PGPR to improve soil quality and wheat production in a naturally saline soil. Rhizosphere 2024, 29, 100849. [Google Scholar] [CrossRef]
- Wade, M.R.; Gurr, G.M.; Wratten, S.D. Ecological restoration of farmland: progress and prospects. Philos. Trans. R. Soc. B: Biol. Sci. 2008, 363, 831-847. [CrossRef]
- Gairola, S.U.; Bahuguna, R.; Bhatt, S.S. Native plant species: A tool for restoration of mined lands. J. Soil Sci. Plant. Nutr. 2023, 23, 1438–1448. [Google Scholar] [CrossRef] [PubMed]
- Calle, C.; Bonifacio, A.; Villca, M.; et al. Arbustos y pastos para restablecer la cobertura vegetal en zonas áridas del Sur de Bolivia. CIMMYT, Mexico, 2022, pp 1-12. https://hdl.handle.net/10883/22242.
- de-Bashan, L. E., Hernandez, J. P., & Bashan, Y. The potential contribution of plant growth-promoting bacteria to reduce environmental degradation – A comprehensive evaluation. Appl. Soil Ecol. 2012, 61, 171-189. Lopez-Lozano et al., 2016. [CrossRef]
- Li, H.; Qiu, Y.; Yao, T.; et al. Effects of PGPR microbial inoculants on the growth and soil properties of Avena sativa, Medicago sativa, and Cucumis sativus seedlings. Soil Till. Res. 2020, 199, 104577. [Google Scholar] [CrossRef]
- Khoshru, B., Mitra, D.; Khoshmanzar, E., Myo, E. M., Uniyal, N., Mahakur, B., et al. Current scenario and future prospects of plant growth-promoting rhizobacteria: an economic valuable resource for the agriculture revival under stressful conditions. J. Plant Nut. 2020, 43, 3062–3092. [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: Recent developments, constraints, and prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
Figure 1.
Factors contributing to soil degradation and microbiota-based remediation strategies. The primary causes of soil degradation (A) are predominantly anthropogenic, leading to adverse effects on soil structure and function and ultimately compromising its long-term sustainability. Microbiota-based remediation strategies (B) aim to restore soil integrity and functionality, promoting sustainable recovery. These strategies include the application of biofertilizers, biopesticides, and phytoremediation techniques.
Figure 1.
Factors contributing to soil degradation and microbiota-based remediation strategies. The primary causes of soil degradation (A) are predominantly anthropogenic, leading to adverse effects on soil structure and function and ultimately compromising its long-term sustainability. Microbiota-based remediation strategies (B) aim to restore soil integrity and functionality, promoting sustainable recovery. These strategies include the application of biofertilizers, biopesticides, and phytoremediation techniques.
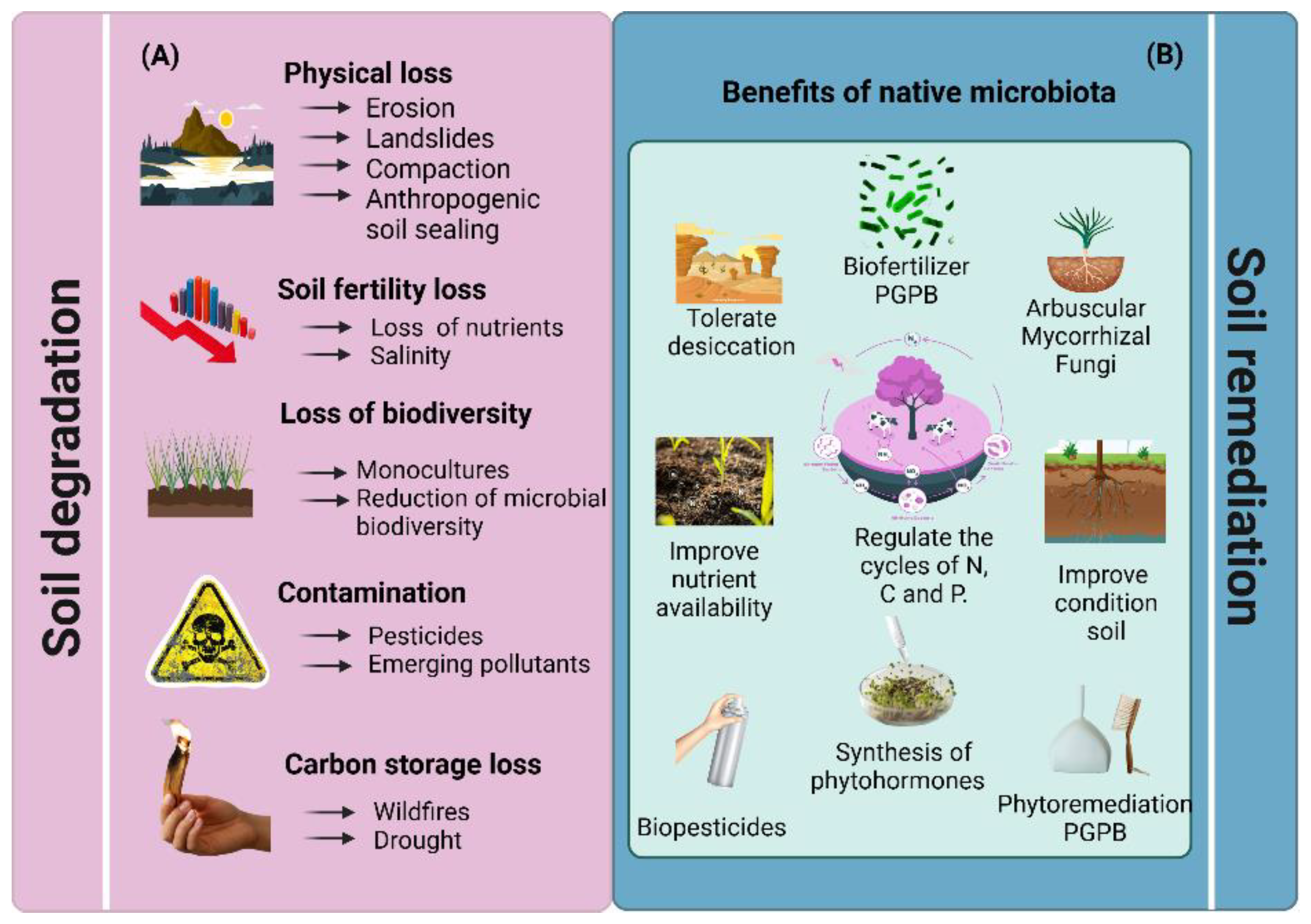
Figure 3.
The enzyme GS catalyzes the conversion of ammonium (15NH4+) and glutamate into glutamine in an ATP-dependent reaction that generates ADP and inorganic phosphate (Pi). Subsequently, GOGAT transfers the amide group from glutamine to α-ketoglutarate, utilizing NADPH as a reducing cofactor, to produce two glutamate molecules. This pathway, which predominates under low-ammonium and high-carbon/energy conditions, is highly efficient in glutamate synthesis, a key intermediate in bacterial nitrogen and carbon metabolism.
Figure 3.
The enzyme GS catalyzes the conversion of ammonium (15NH4+) and glutamate into glutamine in an ATP-dependent reaction that generates ADP and inorganic phosphate (Pi). Subsequently, GOGAT transfers the amide group from glutamine to α-ketoglutarate, utilizing NADPH as a reducing cofactor, to produce two glutamate molecules. This pathway, which predominates under low-ammonium and high-carbon/energy conditions, is highly efficient in glutamate synthesis, a key intermediate in bacterial nitrogen and carbon metabolism.
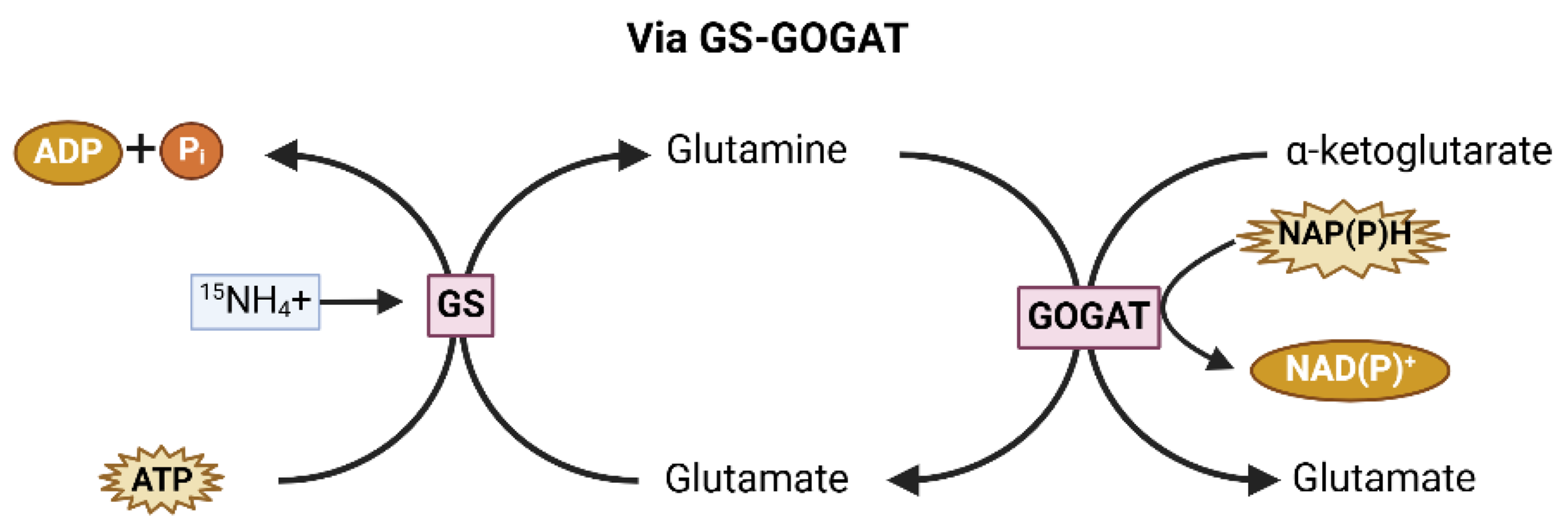
Figure 4.
The enzyme ACC deaminase lowers ethylene levels by degrading ACC, an ethylene precursor, thereby alleviating its inhibitory effects on plant growth. In addition, bacteria produce IAA, a PGR derived from Trp, which promotes root development. They also solubilize Pi and chelate Fe³⁺ through siderophore production, enhancing the bioavailability of these essential nutrients to plants. The synthesis of exopolysaccharides contributes to soil aggregation and water retention, promoting root health. On the plant side, the activation of genes associated with defense (CaPR-10), stress tolerance (VA, sHSP), and ion transport (K⁺, Na⁺, and Ca²⁺) strengthens resilience to abiotic and biotic stress.
Figure 4.
The enzyme ACC deaminase lowers ethylene levels by degrading ACC, an ethylene precursor, thereby alleviating its inhibitory effects on plant growth. In addition, bacteria produce IAA, a PGR derived from Trp, which promotes root development. They also solubilize Pi and chelate Fe³⁺ through siderophore production, enhancing the bioavailability of these essential nutrients to plants. The synthesis of exopolysaccharides contributes to soil aggregation and water retention, promoting root health. On the plant side, the activation of genes associated with defense (CaPR-10), stress tolerance (VA, sHSP), and ion transport (K⁺, Na⁺, and Ca²⁺) strengthens resilience to abiotic and biotic stress.
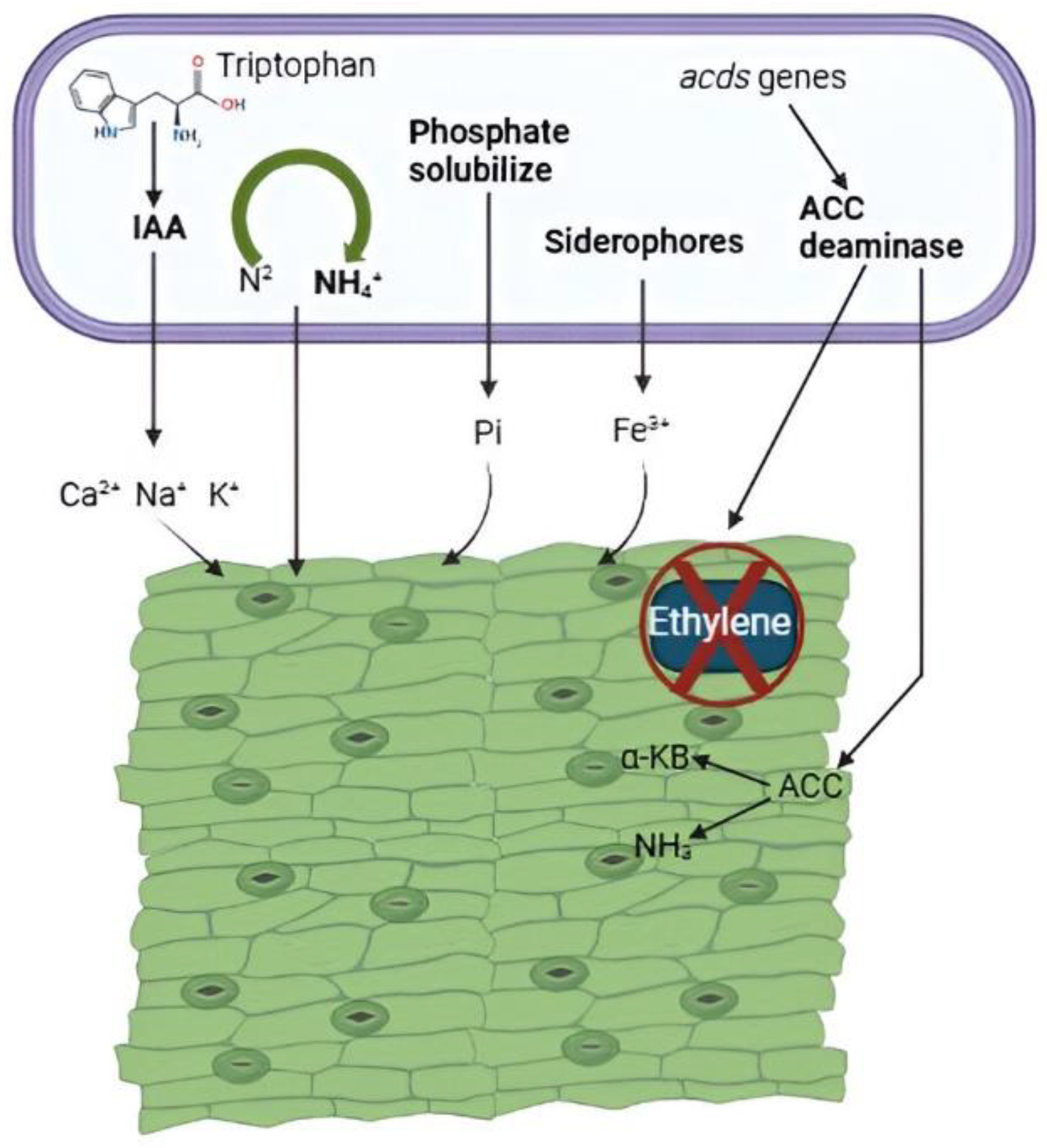
Figure 6.
Plant-microorganism interactions mediated by signaling molecules and communication mechanisms. Plants release chemical compounds that influence microbial behavior in the rhizosphere through processes such as QS and antibiosis. In response, microorganisms produce specific signaling molecules, including AHLs and other chemical signals, which can induce plant resistance, modulate root development, and shape the surrounding microbiota. Additionally, quorum quenching (QQ) serves as a regulatory mechanism that degrades or interferes with QS signals, modulating inter-organism interactions and controlling processes such as virulence and microbial competition.
Figure 6.
Plant-microorganism interactions mediated by signaling molecules and communication mechanisms. Plants release chemical compounds that influence microbial behavior in the rhizosphere through processes such as QS and antibiosis. In response, microorganisms produce specific signaling molecules, including AHLs and other chemical signals, which can induce plant resistance, modulate root development, and shape the surrounding microbiota. Additionally, quorum quenching (QQ) serves as a regulatory mechanism that degrades or interferes with QS signals, modulating inter-organism interactions and controlling processes such as virulence and microbial competition.
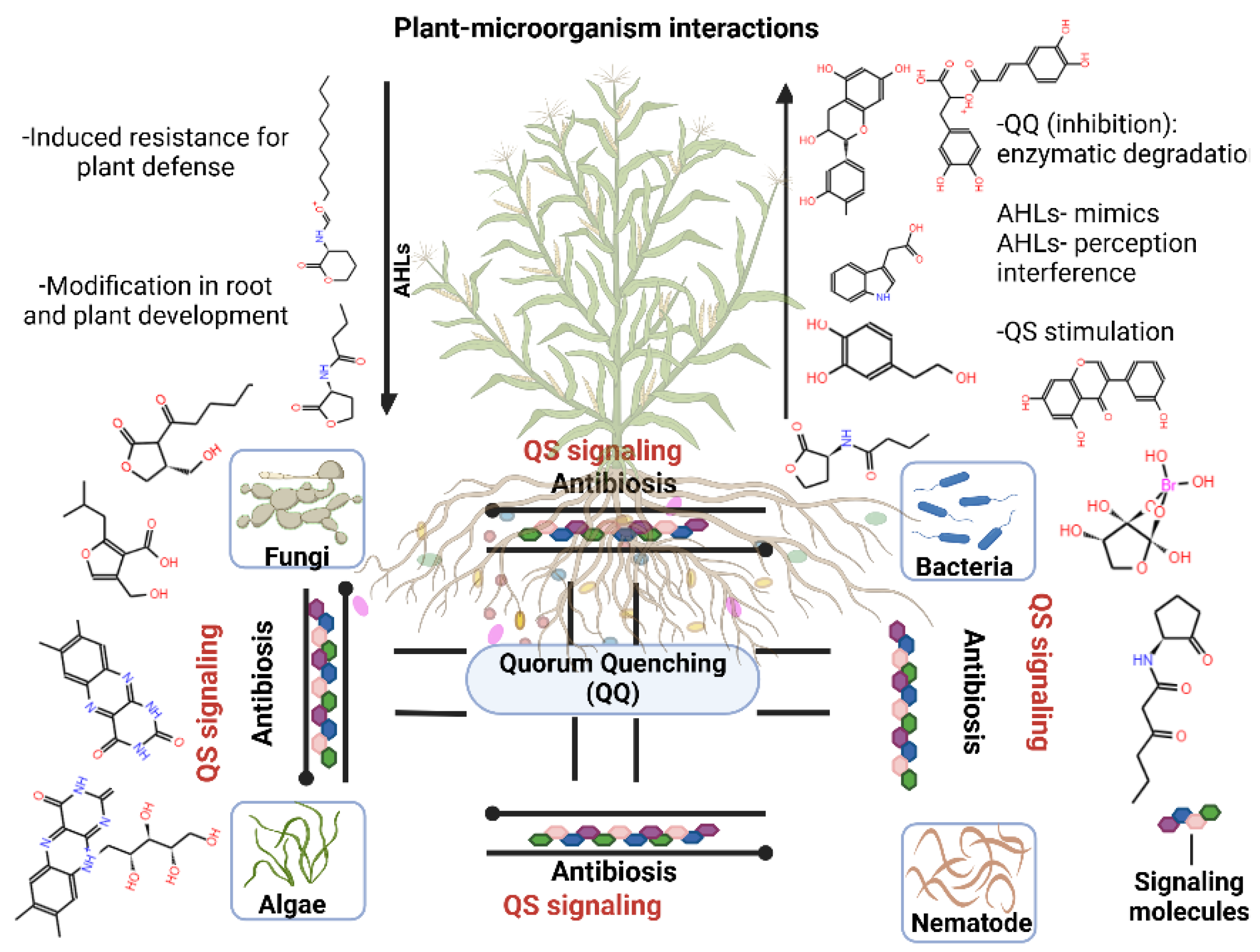
Figure 7.
Signaling pathways involved in plant stress responses induced by pathogens and rhizobacteria. External signals, including reactive oxygen species (ROS), nitric oxide (NO), and calcium (Ca²⁺) fluctuations, activate hormonal signaling pathways and MAPKs, which regulate gene expression to enhance defense mechanisms and stress adaptation.
Figure 7.
Signaling pathways involved in plant stress responses induced by pathogens and rhizobacteria. External signals, including reactive oxygen species (ROS), nitric oxide (NO), and calcium (Ca²⁺) fluctuations, activate hormonal signaling pathways and MAPKs, which regulate gene expression to enhance defense mechanisms and stress adaptation.
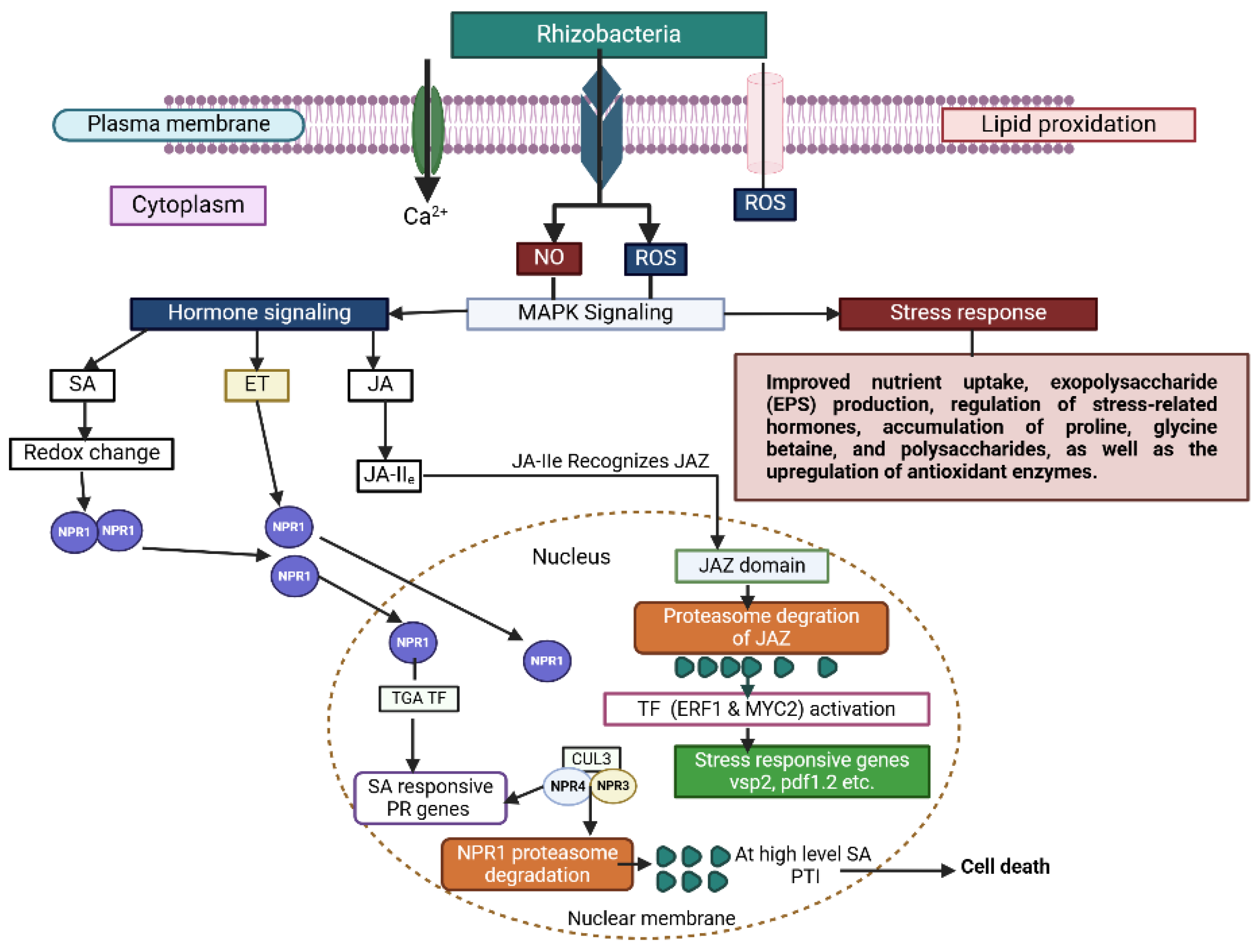
Figure 8.
Key factors influencing soil quality and stability, which determine erosion dynamics and their impact on environmental recovery. These factors are categorized into five interrelated groups: soil properties, microbial communities, contaminants, economic considerations, and non-technical factors.
Figure 8.
Key factors influencing soil quality and stability, which determine erosion dynamics and their impact on environmental recovery. These factors are categorized into five interrelated groups: soil properties, microbial communities, contaminants, economic considerations, and non-technical factors.
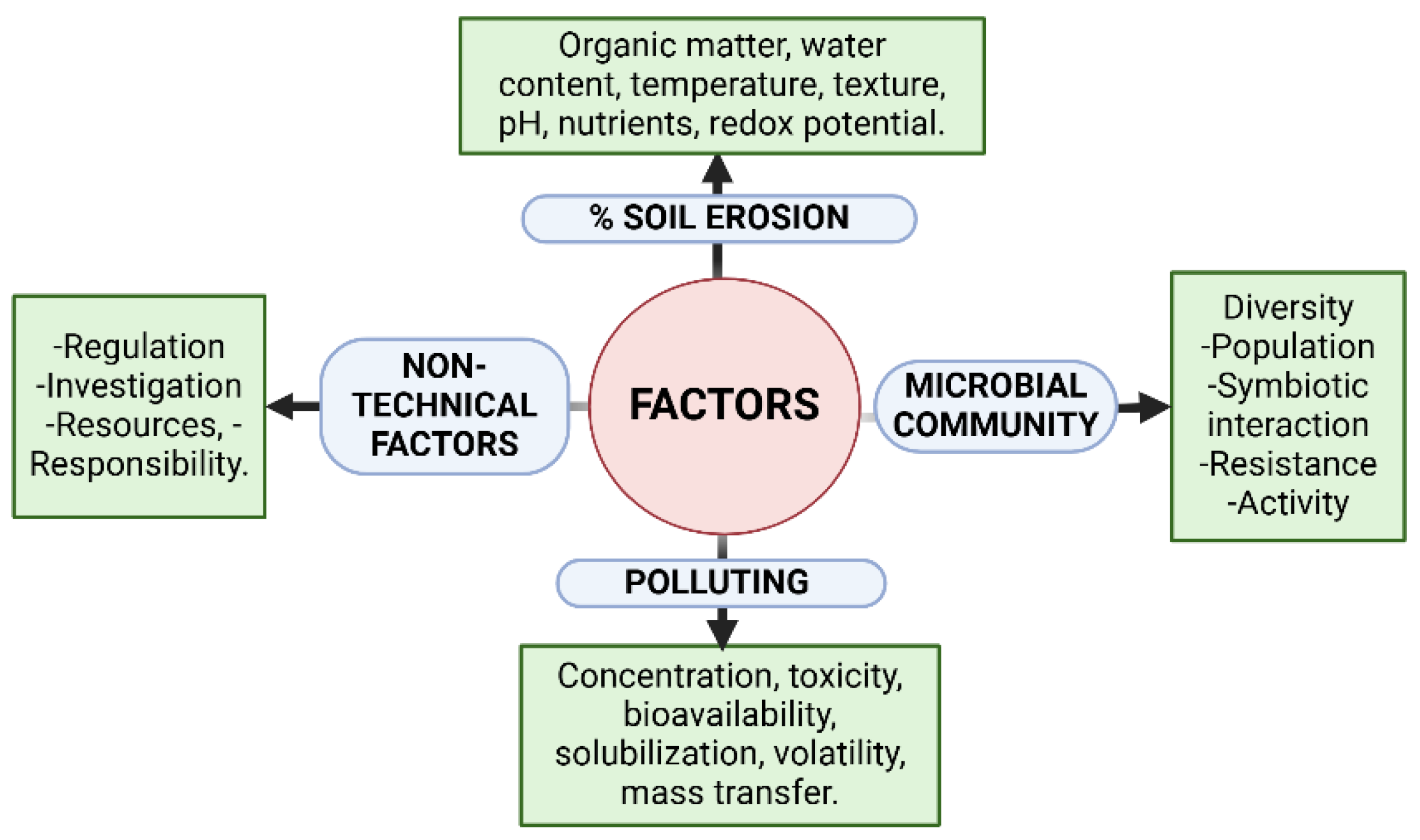
Figure 9.
Primary adaptive responses promoted by PGPB to mitigate stress factors such as heavy metals, hydrocarbons, salinization, and drought, acting as microbial strategies to alleviate biotic and abiotic stress in plants. These microbial interactions contribute to the regulation of plant growth, enhancement of stress tolerance, and restoration of soil functionality, ultimately supporting plant resilience and sustaining agricultural productivity under environmental stress conditions.
Figure 9.
Primary adaptive responses promoted by PGPB to mitigate stress factors such as heavy metals, hydrocarbons, salinization, and drought, acting as microbial strategies to alleviate biotic and abiotic stress in plants. These microbial interactions contribute to the regulation of plant growth, enhancement of stress tolerance, and restoration of soil functionality, ultimately supporting plant resilience and sustaining agricultural productivity under environmental stress conditions.
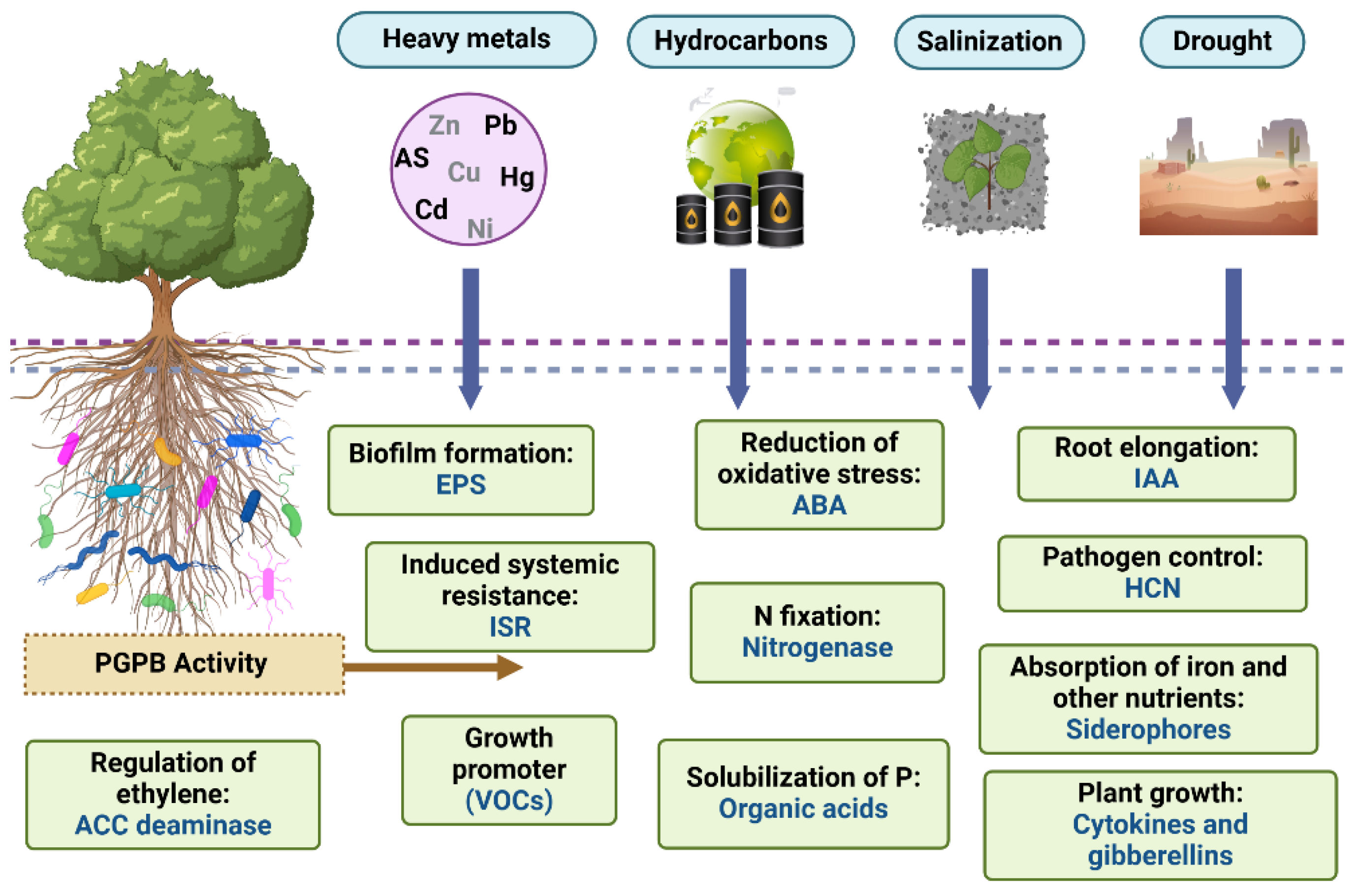
Figure 10.
Soil degradation is driven by human activities such as industrialization, urbanization, chemical usage, extensive agriculture, waste accumulation, and mining, leading to the buildup of heavy metals (Hg, Cd, Cr, Pb, As) and the depletion of essential nutrients (N, P, K). In healthy soil, phytoremediation is enhanced by stimulating biological processes such as phytomineralization, which facilitate contaminant detoxification and nutrient recovery. Additionally, the use of nanoparticles (NPs) and sulfidation processes aids in stabilizing toxic elements.
Figure 10.
Soil degradation is driven by human activities such as industrialization, urbanization, chemical usage, extensive agriculture, waste accumulation, and mining, leading to the buildup of heavy metals (Hg, Cd, Cr, Pb, As) and the depletion of essential nutrients (N, P, K). In healthy soil, phytoremediation is enhanced by stimulating biological processes such as phytomineralization, which facilitate contaminant detoxification and nutrient recovery. Additionally, the use of nanoparticles (NPs) and sulfidation processes aids in stabilizing toxic elements.
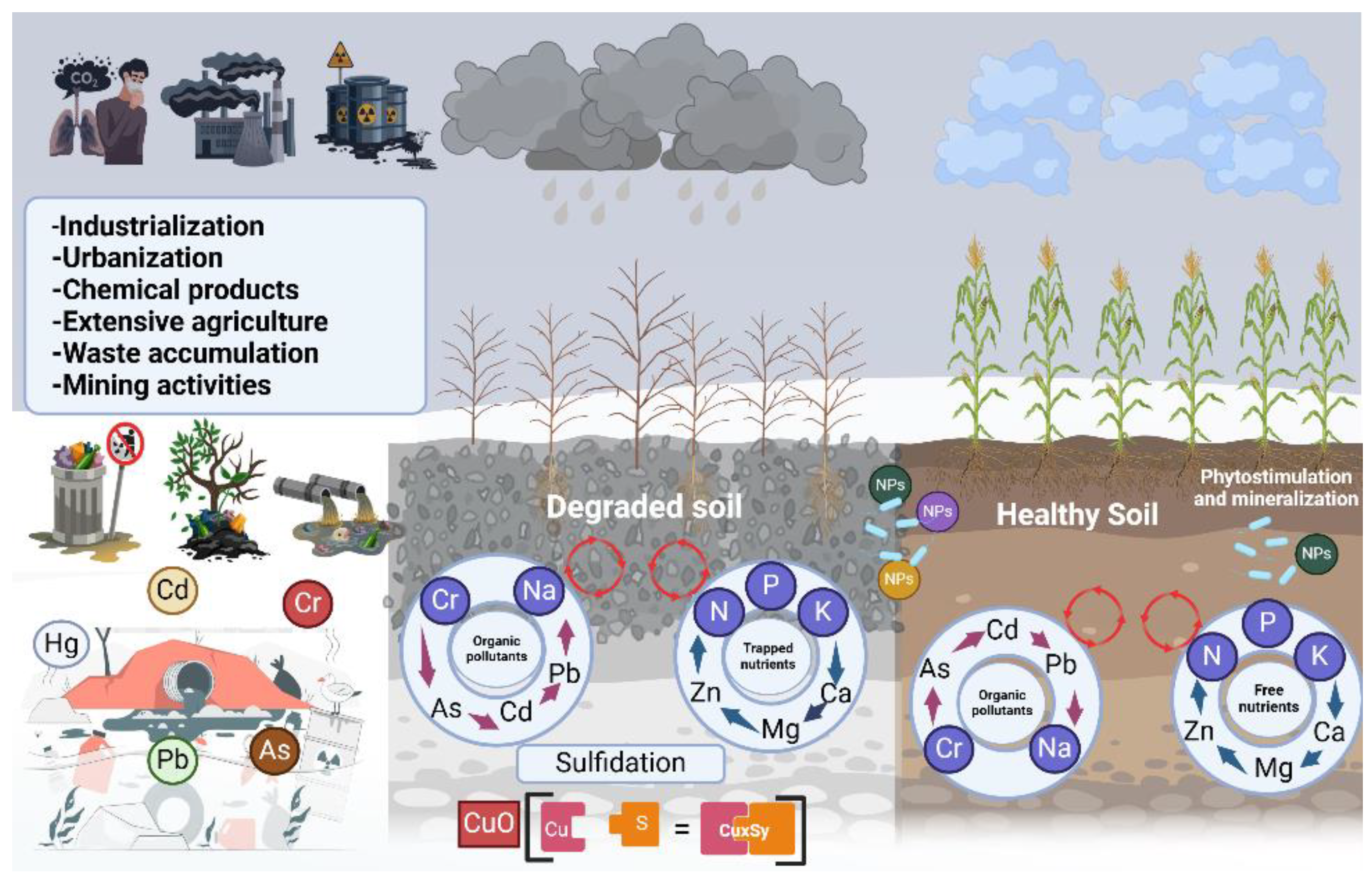
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



