
Submitted:
19 March 2025
Posted:
20 March 2025
You are already at the latest version
Abstract
Background/Objectives: Parietin (PTN), a blue-light absorbing pigment from Teloschistes spp. lichens, exhibits photosensitizing properties via Type I (superoxide anion, O2•-) and Type II (singlet oxygen, 1O2) mechanism, inactivating bacteria in vitro after photo-stimulation. We evaluate the in vitro antifungal activity of PTN against Candida tropicalis biofilms under actinic irradiation, its role in O2•- and 1O2 production, and the cellular stress response. Methods: Minimum Inhibitory Concentration (MIC) of PTN was determined in C. tropicalis NCPF 3111 under dark and actinic light conditions. Biofilm susceptibility was assessed at MIC/2, MIC, MICx2, MICx4 and MICx6 in the same conditions, and viability was measured by colony forming units. Photodynamic mechanisms were examined using Tiron (O2•- quencher) or sodium azide (1O2 scavenger). O2•- production was measured by the Nitro-Blue tetrazolium (NBT) reduction and nitric oxide (NO) by Griess assay. Total antioxidant capacity was studied by FRAP (Ferrous Reduction Antioxidant Potency) assay and superoxide dismutase (SOD) activity by NBT assay. Results: Photo-stimulation of PTN reduced C. tropicalis biofilm viability by 4-logs at MICx2. Sodium azide partially reversed the effect, whereas Tiron fully inhibited it, indicating the critical role of O₂•⁻. PTN also increased O₂•⁻ and NO levels, enhancing SOD activity and FRAP. However, this antioxidant response was insufficient to prevent biofilm photoinactivation. Conclusions: Photoinactivation of C. tropicalis biofilms by PTN is primarily mediated by O2•-, with a minor contribution from ¹O₂ and an imbalance in NO levels. These findings suggest PTN as a promising photosensitizer for antifungal photodynamic therapy.
Keywords:
natural anthraquinone
; superoxide anion
; singlet oxygen
; Candida biofilm
; photodynamic therapy
; photoinactivation
; reactive oxygen species
; oxidative stress
1. Introduction
Antimicrobial (ATM) resistance is currently a global problem that hinders the treatment of common infections, increasing the risk of serious and even fatal complications. Treatment times are prolonged, which has an impact on other aspects such as increased healthcare costs, both for patients and healthcare systems, as more complex and expensive treatments are used. It also reduces the safety of some medical procedures (surgeries, transplants, cancer treatments) by increasing the risk of resistant infections. This is compounded by the rapid spread of infection, which makes it difficult to control and increases the risk of epidemic outbreaks [1].
Invasive fungal infections represent a significant challenge in the clinical setting, especially those caused by species of the genus Candida, an opportunistic fungus responsible for most hospital-acquired infections with a high morbidity and mortality rate. The resistance of Candida spp. to ATM has been associated with their ability to form biofilms, which have the particularity of being formed by both morphotypes: the yeast and hyphal forms that intertwine to form a complex three-dimensional structure with greater resistance to traditional antifungals than their planktonic counterpart. Within this group, Candida tropicalis stands out for its ability to form biofilms that are highly resistant to conventional treatments, which makes eradication difficult and favors the persistence of infections in immunocompromised patients [2,3].
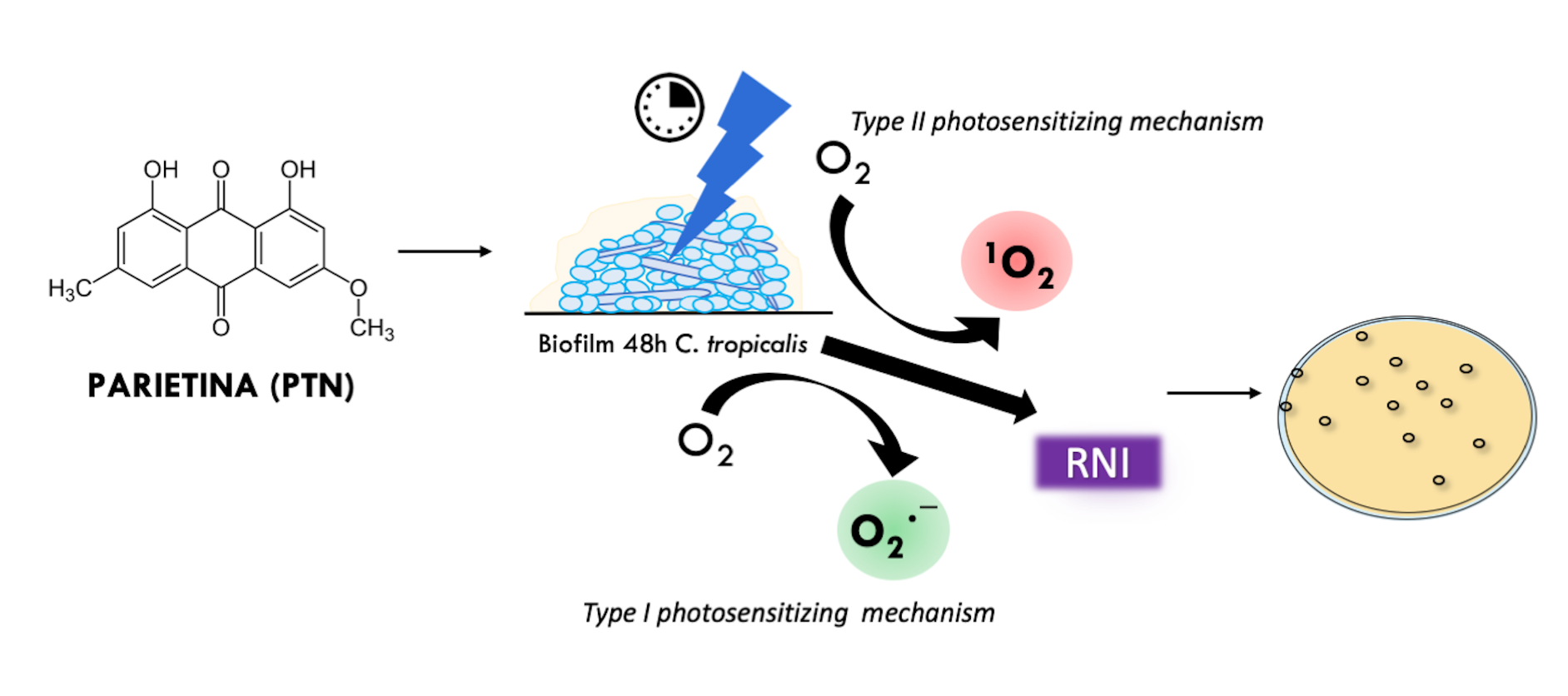
Among the many strategies to address ATM resistance, the antimicrobial photodynamic inactivation (aPDI) is an option, which is based on the generation of reactive oxygen species (ROS) following stimulation of a photosensitizer (PS) with light of a specific wavelength. This approach has proven to be effective against various pathogenic bacteria, parasites, viruses and fungi, while minimizing toxicity to human cells. The excited specie of PS reacts with molecular oxygen (3O2) by two photosensitizing mechanisms: Type I involving charge transfer (electrons or hydrogen atoms) to generate superoxide anion radical (O2∙¯), or Type II by energy transfer processes, generating singlet oxygen (1O2). These species thus generated and the species derived from them by secondary reactions (hydrogen peroxide: H2O2; hydroxyl radical: HO∙), are ultimately those that oxidize the biological molecules in their direct environment (proteins, lipids, DNA), triggering cellular oxidative stress that eventually leads to the death of the microorganisms, which translates into regression of the infection [4, 5].
In the search for new therapeutic agents, natural products have become of interest due to their structural diversity and bioactive potential [6]. Among them, lichens are an important source of secondary metabolites with antimicrobial, antioxidant and cytotoxic properties [7,8]. These symbiotic associations between fungi and algae or cyanobacteria produce a wide variety of terpenoid and phenolic derivatives, including among the latter anthraquinones (AQs), many of which have demonstrated activity against pathogenic bacteria and fungi [7].
AQs are a family of compounds found in higher plants and lichens with photoactive properties and potential antimicrobial activity. Some AQ derivatives have shown the ability to generate ROS under irradiation, suggesting their possible application in the inactivation of pathogens. However, the mechanism of action of these molecules and their impact on the viability and structure of fungal biofilms still require further investigation [9,10].
Parietin (PTN), 1,8-dihydroxy-3-methoxy-6-methyl-9,10-anthraquinone (Figure 1), is found as orange crystals in the upper crust of lichens of the Teloschistaceae family, acting as an Ultraviolet-B photoprotective pigment [11]. However, we have previously shown that, in dilute solution in chloroform, PTN is an efficient PS of 1O2 with a high quantum yield (ΦΔ = 0.69) as well as an O2∙¯ photo-generator in aqueous media [12].
In this work, the in vitro antifungal and antibiofilm activity of PTN, an AQ obtained from a lichen, was evaluated both in dark conditions and under irradiation. The aim is to determine its potential as a PS against C. tropicalis biofilms. In addition, the photo-sensitizing mechanisms (Type I and Type II) involved in the generation of ROS were studied; and because of this, the generation of reactive nitrogen intermediates (RNI) was assessed, since these were formed as a process of deactivation of O2∙¯. The activation of the superoxide dismutase enzyme (SOD) and the total antioxidant capacity of the biological system (by means of FRAP: ferric reducing antioxidant potency assay) are also evaluated, to establish whether oxidative or nitrosative stress would be responsible for the observed biological effect [13-15].
The results of this study indicate that the photoinactivation of Candida tropicalis biofilms by PTN is predominantly driven by the O₂•⁻ generation (Type I mechanism). Although the ¹O₂ (Type II mechanism) also plays a role in this process, its contribution is considerably smaller compared to O₂•⁻. Furthermore, the study revealed an imbalance in NO levels, which likely exacerbates the overall cellular stress response, contributing to the effectiveness of the photodynamic treatment. Given these findings, PTN shows strong potential as an effective PS in antifungal photodynamic therapy (aPDT), offering a promising therapeutic approach for treating fungal biofilm-related infections.
2. Materials and Methods
Reagents and solvents
Sabouraud Dextrose Broth (SDB) and Sabouraud Dextrose Agar (SDA) were obtained from Britania, CABA, Argentina. Sodium nitrite (NaNO2,) and ferrous sulfate (FeSO4) were acquired from Cicarelli, Sta. Fe, Argentina. Sodium hydroxide (NaOH, Biopack, Zárate, Argentina) and Amphotericin B (AmB, Richet, Tres Arroyos, Argentina), Phosphate-buffered saline (PBS). Roswell Park Memorial Institute 1640 (RPMI), morpholine propane sulfonic acid (MOPS), sodium azide (NaN3), Tiron, nitro blue tetrazolium (NBT), methionine and riboflavin were purchased from Sigma, St. Louis, MO, USA. Water and distilled solvents (Sintorgan, Buenos Aires, Argentina) were used.
Yeast strain and growth conditions
A standard strain was used: Candida tropicalis NCPF 3111 (National Collection of Pathogenic Fungi, Bristol, UK). It was conserved and reactivated according to Clinical & Laboratory Standards Institute guidelines (CLSI, 2002)[16]. Sabouraud Dextrose Broth (SDB) was used as a growth medium at 37 °C.
Natural photosensitizer tested
Parietin (PTN) (Figure 1) was obtained from the lichen Teloschistes nodulifer (Nyl.) Hillman (Teloschistáceas, identified as CORDC00005354 (Museo Botánico de Córdoba, Universidad Nacional Córdoba). In the purification and identification process, the methodology previously developed was followed to obtain a purity of 95%, according to HPLC [17,18]. A hydroalcoholic solution of PTN (3.5 mM) was used as a stock solution, containing 1% ethanol (EtOH). The tested concentrations were prepared by diluting the stock with culture medium, depending on whether the assay was carried out on planktonic yeasts or biofilm.
Irradiation system
Photo-inactivation assays were carried out using an actinic Phillips 20W lamp (380 ± 480 nm, 0.65 mWcm-2) with an emission maximum at 420 nm, which was placed inside a black box at 20 cm above the samples [18].
Photoactive Minimum Inhibitory Concentration
The lowest concentration of the compound that inhibits the growth of the planktonic form of C. tropicalis was determined. The protocol described by CLSI (2008) [19] was applied to determine the Minimum Inhibitory Concentration (MIC), adapted to be carried out under dark and irradiated conditions [20]. This allows establishing the photoactivated MIC (pMIC) in comparison to the MIC in darkness, giving an idea of the photodynamic potential of the PS. The assay was carried out on 96-well microplates (Greiner Bio-One, Germany) containing the following culture medium: RPMI 1640 with glutamine and 0.2% glucose, without sodium bicarbonate, buffered with 0.164 M MOPS, and adjusting the pH to 7 with 1N NaOH. PTN solutions (100 μL) were added to each well following the additive serial double dilution method, and twelve concentrations between 0.24 to 500 µg/mL were tested in triplicate. AmB was the positive control (+C), used at its MIC. The yeast suspension (100 μL) was then added to each well with a final concentration of 0.5 x 103 - 2.5 x 103 CFU/mL. One set of these concentrations was irradiated for 15 min at room temperature, whereas another set was shielded from light. Immediately after this time, both sets were incubated at 37ºC for 48 h. Finally, optical density (OD) of each well was measured at 490 or 530 nm on a microplate reader (Tecan Sunrise Model, TECAN, Australia). RPMI and yeast suspension with 1% EtOH, both without PTN were embraced as controls (in triplicate) in both experimental conditions.
In addition, a colony-forming unit (CFU) count assay was performed. A sample (100 µl) was taken from each well, which was diluted 1:10 in PBS until final dilution 1:1000000. Subsequently, each suspension (100 μL) was seeded on Sabouraud Dextrose Agar (SDA)-plates and incubated for 48 h at 37 ºC. Before incubation, the CFU were counted to establish the Minimum Fungicidal Concentration (MFC), defined as the lowest concentration that of a compound that kills 99.9% of a fungal inoculum.
Photo-inactivation biofilm procedures
Biofilm were formed in flat-bottomed 96-well microplates (Greiner Bio-One, Germany), following an adaptation from the method of O’Toole & Kolter [18]. Once a 48 h biofilms formation in SDB was achieved, the microplates were rinsed twice with PBS (200 µL x 2) at pH=7 to remove non-adherent cells. PTN was tested on C. tropicalis biofilms at five concentrations in triplicate: pMIC/2, pMIC, pMICx2, pMICx4, pMICx6, under darkness and irradiation conditions. SDB alone and SDB with 1% EtOH were included as negative controls. AmB (+C) was used as positive control at MIC. After treatment, microplates were incubated at 37ºC for 48 h. The supernatant was replaced by PBS (100 µL per well) and sonicated (40 kHz, 60 s in Codyson CD4831, Shenzhen, China). The biofilm of each well was removed and serially diluted with PBS. Each dilution was plated (10 μL) on SDA plate in triplicate, and the number of CFU formed after 48 h incubation at 37 °C was counted. The counting of CFU/mL was log-transformed.
Mechanism action studies
To determine the mechanism of photosensitizing action, the scavenging effect of sodium azide on singlet oxygen (1O2 quencher) or Tiron on superoxide anion (O2•¯quencher) was evaluated in PTN-treated and untreated biofilm both in the presence and absence of light [21]. Quenchers and PTN solutions were added on a dense biofilm (48 h) at the same time, so that the final quencher concentration was 200 μM and the PTN concentrations pMIC/2, pMIC, pMICx2, pMICx4, pMICx6. After darkness and irradiation treatment (15 min), the procedure continued as described above for CFU quantification.
Each supernatant of both microplates (dark and light) was extracted to assess the production of O2•¯, reactive nitrogen intermediates (RNI) and the activation of the antioxidant system: SOD and total non-enzymatic system by means of the ferric reducing antioxidant potency assay (FRAP).
Production of O2•¯was determined by the nitro blue tetrazolium (NBT) reduction [22]. Blue diformazan formation is proportional to the generated O2•¯in biofilms and its OD was measured at 540 nm on the same microplate reader. Results were expressed as OD540nm/CFU (Superoxide anion/CFUml-1) [18].
RNI generation was evaluated as nitrite formation, by using Griess reaction and a calibration curve of NaNO2 as standard [23]. OD was measured spectrophotometrically at 540 nm [18] and results were expressed as the ratio between nitrite concentration values/CFU (RNI/ CFUml-1).
SOD activity was evaluated by the ability of this enzyme to inhibit NBT reduction in the presence of O2•¯, generated by the photostimulation of riboflavin in the presence of oxygen and an electron donor (methionine). Results were expressed as SOD activation (%SOD/ CFUml-1) [18,24].
FRAP assay was used following the methodology described by Benzie & Strain (1996) [25]. The absorbance was measured at 593 nm and results were expressed as the Fe+2 concentration values/CFU (FRAP/ CFUml-1) by using a FeSO4 calibration curve [18].
Statistical analysis
All assays were made in triplicate and in 3 independent experiments. Data were expressed as means ± standard deviation. A p value < 0.05 was considered statistically significant, obtained by the t- Student-Newman-Keuls test for multiple comparisons. * denotes statistical significance at p < 0.05 when was compared to untreated biofilms. # indicates statistical significance at p < 0.05 when darkness and irradiation were compared.
3. Results
3.1. Photoactive Minimum Inhibitory Concentration (pMIC)
PTN produced growth inhibition on planktonic yeasts of C. tropicalis at 0.98 μg/mL (≈3.45 nM) under irradiation conditions, defined as the photoactive MIC (Figure 2). The photoactive MFC was also achieved at this concentration (data not shown). Furthermore, these values are below the cytotoxic concentration of PTN, since the cell viability of Vero cells was 80% and 60% at 100 µg/ml, under darkness and light respectively [12]. In contrast, in the dark, PTN only produces 20% growth inhibition; therefore, it is not possible to determine its MIC.
3.2. PTN Antimicrobial Photodynamic Therapy (APDT)
The antifungal activity of PTN was also tested on C. tropicalis biofilms at five concentrations taking as reference the pMIC value (0.98 μg/mL ≈3.45 μM), obtained in yeast suspensions. CFU were not affected by light conditions. Regarding the antibiofilm activity of PTN on C. tropicalis (Figure 3), we can state that it had no effect at the tested concentrations in the dark. On the other hand, when PTN was photo-stimulated it can be observed that the photodynamic effect was concentration-dependent, starting at the pMIC and the most active concentration being pMICx2 as it produced a 4 logs reduction. At the highest concentrations tested, no photoinactivation was observed, probably because in general AQs are aggregated in aqueous media [21].
3.2.1. Photodynamic Mechanismon Analysis after PTN-APDT
To clarify the photoinactivation mechanism, the ROS generated were determined by adding quenchers. Thus, the scavenging effect of 200 mM sodium azide (1O2 quencher) or Tiron (O2•- quencher) on biofilm photoinactivation by PTN is shown in Figure 4. Tiron completely inhibited the photodynamic effect of PTN at all bioactive concentrations (pMIC and pMICx2). The other ROS quencher, sodium azide, was less effective than Tiron, as it could not reverse the action of the O2•- although it was able to significantly increase the biofilm survival by almost 3 logs at these concentrations.
3.2.2. Biofilm Stress Response After PTN-APDT
The O2•¯generation and nitrosative metabolites production were studied for the photoinactivation process of PTN on C. tropicalis biofilms. An increment in O2•¯ levels was observed when PTN was photo-stimulated at pMIC/2, pMIC and pMICx2 respect to untreated biofilms (Figure 5, violet bars). In addition, at the higher concentrations tested of PTN no increase in this ROS was detected. In relation to RNI generation by this AQ when it was photo-stimulated on biofilms (Figure 6, yellow bars), the rise in RNI was greater for the most active concentration of PTN than other concentrations tested.
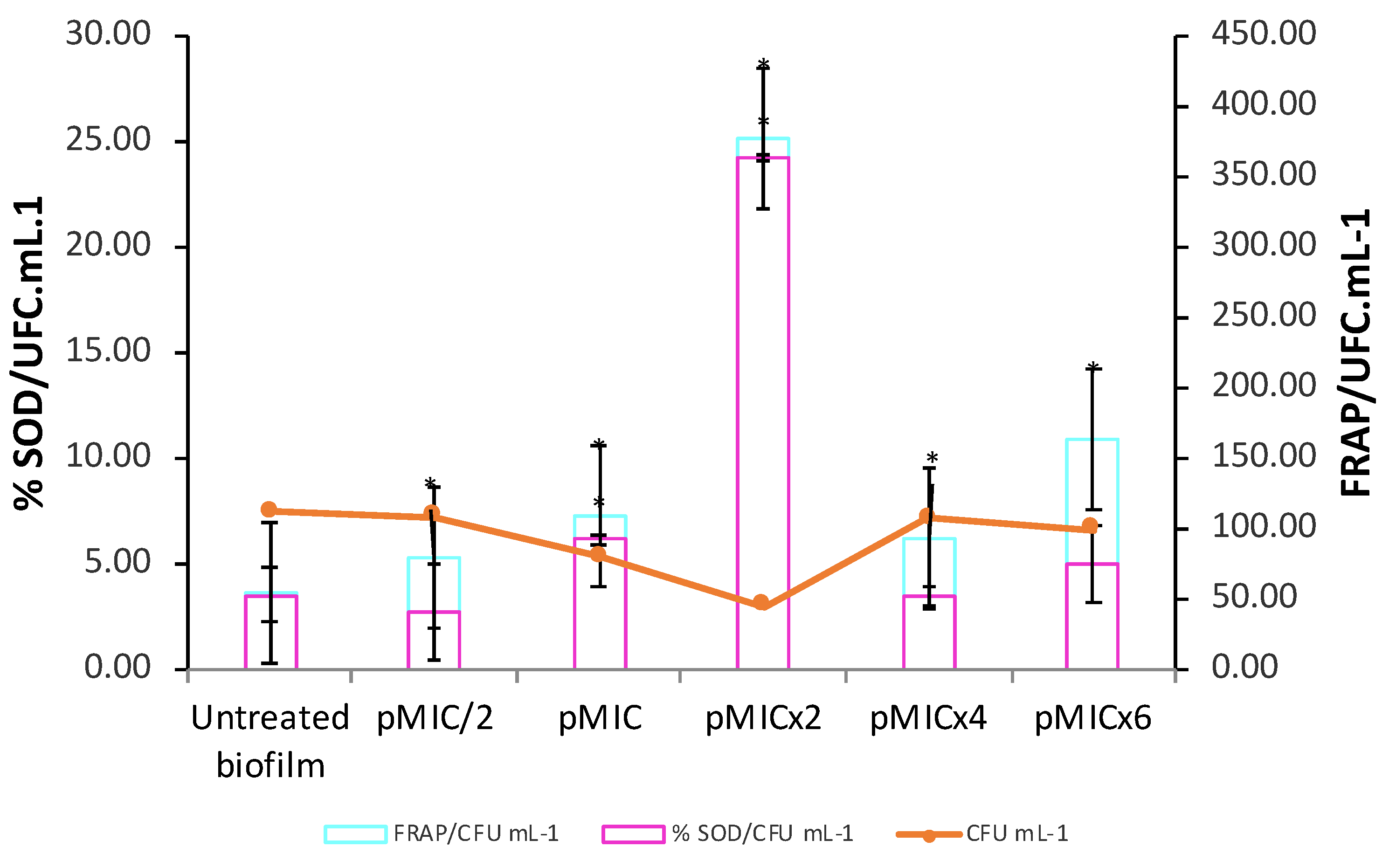
Furthermore, the stimulation of SOD activity (Figure 6, magenta bars) was observed at the photoactive concentrations of PTN (pMIC and pMICx2). The levels of total antioxidant capacity (FRAP) in the treated biofilm increased at all conditions evaluated compared to untreated biofilm (Figure 6, turquoise bars). Moreover, both defense systems (SOD and FRAP) were detected even in higher proportion for pMICx2.
4. Discussion
The treatment of candidiasis is complex due to the pathogenic mechanisms of Candida spp., particularly their ability to form biofilms and their acquired resistance or multi-resistance, which represents a significant public health concern [26]. Given this challenge, aPDI is being investigated as a possible alternative for treating fungal infections, which was the motivation for this study [27].
In this study, we started by investigating the effect of PTN-mediated PDI on planktonic yeast of C. tropicalis. The results showed negligible toxicity in samples treated with AQ and darkness (< 20 % growth inhibition). This is consistent with previous studies that reported PTN-induced cytotoxicity in Vero cells. Furthermore, in a previous work PTN had antibacterial effect on both gram-positive and -negative strains only after photostimulation [12]. The effect of light-activated PTN against C. tropicalis and other clinically relevant fungal pathogens, such as Candida albicans, Candida glabrata, Candida auris, has been described using a light-emitting diode (LED) irradiation system (λ= 428 nm). Although yeast growth inhibition was achieved at a lower concentration of PTN (0.156 μg/mL vs 0.98 μg/mL), a significantly higher irradiation dose was required (30 J/cm2 vs 0.59 J/cm2) [28]. Therefore, the lower irradiation dose employed was compensated by using a higher concentration of PS, without exhibiting cytotoxic effects on normal cells. Furthermore, the irradiation system employed in our study is widely used in phototherapy for the treatment of hyperbilirubinemia in humans (http://www.philips.com/phototherapy.
Similarly, Ma et al. investigated the in vitro PDI mediated by aloe-emodin on azole-sensitive and -resistant C. albicans planktonic yeasts [29]. After incubation with 10 μM aloe-emodin and irradiation with 96 J/cm2 of light, approximately 6.5 log10 reductions in the survival of azole-sensitive and -resistant C. albicans were achieved. Their findings further support the potential of anthraquinone derivatives as effective PSs against fungal pathogens, as they observed a significant reduction in yeast after light activation. In their study, a higher irradiation dose (96 J/cm²) was required to achieve substantial antifungal effects, in contrast to the lower doses employed in our work.
Our study also evaluated the PDI of an AQ on a Candida species, but this was PTN on C. tropicalis biofilms. In the dark, PTN showed no antifungal effect at any of the concentrations tested (up to 5.88μg/mL ≈20.7 μM). However, under actinic irradiation, the antibiofilm effect increased significantly, resulting in a 99.9% decrease in viability.
Other natural PS that align with the blue-light emission spectrum are curcumin and riboflavin. In this sense, Quishida et al. [30] investigated the PDI of curcumin and LED light against C. albicans, C. glabrata and Streptococcus mutans biofilm. The findings revealed that aPDT significantly decreased biofilm viability, metabolic activity, and biomass. However, the photo-antibiofilm effect required high concentrations of curcumin (80, 100, and 120 μM). Alshehri et al. [31] examined the effectiveness of PDI using 0.1% riboflavin on C. albicans biofilms. Blue LED light was utilized to activate riboflavin, and fungal cell viability was assessed using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Treatment with riboflavin and LED resulted in the lowest survival rate of C. albicans, with viability below 50%.
Furthermore, other AQs such as emodin, alizarin, alizarin red, chrysazin, quinalizarin, purpurin, and 1-hydroxyanthra-9, 10-quinone have been shown to inhibit biofilm formation of C. albicans. The efficacy of emodin against C. albicans biofilm of strains isolated from hospitalized patients was achieved in the range of 200-400 μg/mL [32]. Alizarin and chrysazin reduced the metabolic activity of C. albicans biofilms by >98% at 10 μg/ml and by 66% at 2 μg/ml, respectively [33]. Moreover, inhibition of C. albicans biofilm formation by anthraquinones-related compounds, namely pyrocatechol, alizarin red, quinalizarin, emodin, 1- hydroxyanthra-9,10-quinone, purpurin, chrysazin, and alizarin has shown to be dose-dependent [32,34]. In addition, the antifungal activity of alizarin has been largely attributed to the presence of aromatic hydroxyl groups (OH) in its AQ core [33]. This suggests that variations in the position of the OH groups significantly influence the antibiofilm properties. Among the AQs tested, those with a OH group at the position-para-exhibited the highest activity in all assays. Notably, purpurin, chrysazin, and alizarin, which share an OH group at the C1 position, demonstrated strong inhibitory effects against Candida albicans biofilms even at low concentrations (2 μg/ml) [33]. Therefore, the antibiofilm activity of PTN against C. tropicalis may be influenced by the presence of OH group at position-1 in the AQ structure.
Besides the AQs, hypericin (a dianthrone) and porphyrins have emerged as promising PSs with high selectivity for fungal pathogens. Hypericin-mediated aPDT demonstrated significant antifungal activity, achieving substantial reductions in Candida biofilm and planktonic cell viability. Furthermore, hypericin activated by yellow or orange light effectively affected fluconazole-resistant Candida strains with minimal cytotoxicity to host tissues [35,36].
Porphyrins are PS with high selectivity to pathogens. Thus, the photoinactivation of different Candida spp. by two cationic porphyrins derived from tetra-chloride salts, meso-tetra(4-N-methylpyridyl) porphyrin and meso-tetra(3-N-methylpyridyl)porphyrin derivatives (4- H2TMeP+ and 3-H2TMeP+), and anionic porphyrin tetra-sodium salt, meso-tetra(4-sulfonatophenyl)porphyrin (4-H2TPSP‒) was investigated by Amorin et al [37]. For the irradiation process, a white light LED and light dosage of 10.9 J/cm2 was used. Cationic porphyrins can photo-inactivate different Candida spp. in both planktonic and biofilm forms. C. tropicalis showed 35.8% interference when treated with 4-H2TMeP+, and 51.67% with 3-H2TMeP+ at 75uM for both porphyrins [37]. In addition, the tetracationic metalloporphyrin Zn(II) meso-tetrakis(N-n-hexylpyridinium-2-yl)porphyrin (ZnTnHex-2-PyP4+) was assayed against biofilms of C. albicans strains (ATCC 10231 and ATCC 90028) using a blue light-emitting diode. The PDI (4.3 J/cm2) at 0.8 μM showed about 89% decrease in the cell viability as assessed by the MTT assay[38].
Therefore, PTN stands out as an effective PS against Candida tropicalis, displaying strong antifungal and antibiofilm activity upon light activation, whereas remaining non-toxic in the dark. As an AQ-based compound, it benefits from a well-defined photoreactive core, structural stability, and required OH positioning, which enhance its photodynamic efficacy. Its potent effect, even at low irradiation doses, makes it a promising alternative to other natural PSs that require higher concentrations or light doses for comparable activity.
The photoinactivation mechanism of C. tropicalis biofilms by PTN was investigated to identify the photo-processes occurring in PTN-PDI.
Tiron inhibits the photoinactivation C. tropicalis biofilm by PTN at bioactive concentration. Although Tiron is a quencher of O2•−, it can react with hydroxyl radicals (OH•) [39]. Since production OH• occurs after the O2•− generation and Tiron was added before the irradiation period, it can be inferred that Tiron acted as a scavenger of O2•−. Thus, the Type I photosensitization mechanism would be involved in PTN-mediated PDI in the biofilm. Furthermore, our previous work showed that PTN can produce O2•−, being an efficient type I photosensitizer [12]. On the other hand, sodium azide only has a limited protective effect on C. tropicalis biofilm, despite PTN being an efficient ¹O₂. producer [12]. This type of behavior has also been reported for other AQs, such as rubiadin and its methylated derivative. For both AQ, it was shown that the main photosensitizing mechanism involved in the photoinduced antibiofilm activity is the O2•−formation through suppression of AQ excited states by electron transfer, whereas the role of ¹O₂ seems to be less significant [21].
Oxidative stress is a complex process that occurs because of two main factors: an overproduction of ROS, specifically by O2•− levels (Type I mechanism), given the short lifetime of singlet oxygen (3 μs) [40]; and a reduction in oxidative defenses. Thus, this excessive ROS cannot be compensated by the enzymatic and non-enzymatic antioxidant systems of the biofilm, which make them insufficient to remove free radicals, resulting in damage to macromolecules (DNA, protein, lipids) and cellular components (cell membrane) [41,42].
When studying the biofilm response to this oxidative burst, NO generation should also be assessed as a response to O2•− levels, since this occurs more rapidly than the enzymatic response of SOD, thus promoting biofilm photoinactivation [43]. Elevated levels of NO (µM concentration) can cause cytotoxicity by inducing oxidative and nitrosative stress, leading to DNA damage, enzyme modification, cellular dysfunction, inflammation, mitochondrial impairment, and apoptosis. NO can inhibit key mitochondrial enzymes, such as aconitase, and disrupt respiratory chain complexes. It also reacts with O2•−, at a faster rate than with oxygen, to form the toxic peroxynitrite (⁻OONO), which generates harmful radicals (OH and NO). It may react with H₂O₂ to produce ¹O₂, further intensifying damage [44,45].
Similar oxidative and nitrosative imbalances have been reported with other AQs, such as rubiadin and its 1-methyl ether derivative, isolated from Heterophyllaea pustulata. These compounds also induce significant photoreductions in biofilm formation of C. tropicalis, likely through similar mechanisms involving the overproduction of ROS and RNI [20].
In planktonic cells, oxidative stress induced by ROS overproduction has been extensively studied. Fiala et al. (2024), showed that PTN targets the cell membrane and induces cell death via ROS-mediated lipid peroxidation after light irradiation on Candida planktonic yeast [28]. Aloe-emodin has also been reported to induce photodynamic damage to the cell envelope of C. albicans planktonic form through ROS generation [29].
All this background suggests that the oxidative burst, resulting from ROS and RNI, is a common feature in the photo-antimicrobial action of AQs, as our results also show.
The study of natural PSs contributes to the development of aPDT, which has proven to be selective, fast acting and has not developed resistance to date. This selectivity is achieved through strict spatial control of irradiation on the affected area (infection), thus uniting the three agents responsible for the photodynamic effect (PS, light and 3O2). The PSs used in aPDT have not been shown to stimulate resistance mechanisms in microorganisms due to their multidimensional action, which generates a generalized oxidative imbalance that does not differentiate between resistant and non-resistant microorganisms. This photooxidation even includes the destruction of virulence factors, which are generally organic molecules susceptible to oxidation. Moreover, the PSs does not necessarily need to enter the cell to exert its action. Consequently, the pathogen cannot develop resistance through increased detoxification metabolism or export of the drug [46].
5. Conclusions
In conclusion, this study demonstrates that the photoinactivation of Candida tropicalis biofilms by PTN is primarily driven by O2•− (Type I mechanism), with ¹O₂ (Type II mechanism) playing a secondary role. The results also show an imbalance in NO levels, which likely contributes to the cellular stress response and enhances the effectiveness of the treatment. The dominant role of O₂•⁻ in biofilm inactivation highlights its significance in photodynamic mechanism of PTN. These findings suggest that PTN ability to disrupt fungal biofilms is linked to complex oxidative and nitrosative stress pathways, making PTN a promising candidate for antifungal PDT and potential clinical applications.
Author Contributions
Conceptualization, and methodology: S.C. Nuñez Montoya and J. Marioni. Software: J. Marioni, M L. Mugas, B. Romero, T.I. Gomez, and J.M. Morales. Formal analysis: J. Marioni, B. Romero, and S.C. Nuñez Montoya. Statical analysis: F. Martínez, and B.S. Konigheim. writing—original draft preparation: J. Marioni, and S.C. Nuñez Montoya. Supervision: J. Marioni, CD Bosarelli, B.S. Konigheim, and S.C. Nuñez Montoya. Resources, supervision, project administration and funding acquisition: J. Marioni and S.C. Nuñez Montoya. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by: SECyT (Consolidar, tipo 2, s/res. N◦ 411/18 y 155/22), FONCyT (PICT 2018 N◦ 4576, s/ res. ANPCyT n◦ 401/19), and CONICET (PIP 2021–2023, s/ res. 1639/2021). The APC was no funde.
Acknowledgments
GL, TIG, JMNM and BCR are doctoral fellow of CONICET. FM and MLM are pos-doctoral fellow of CONICET. JM, BSK, CB and SCNM are members of the Research Career of CONICET.
Conflicts of Interest
“The authors declare no conflicts of interest.”
Abbreviations
The following abbreviations are used in this manuscript:
| ΦΔ | Quantum yield of singlet oxygen |
| +C | Positive control |
| 1O2 | Singlet oxygen |
| 3O2 | Molecular oxygen |
| AmB | Amphotericin B |
| aPDI | Antimicrobial photodynamic inactivation |
| aPDT | Antimicrobial Photodynamic Therapy |
| AQ/s | Anthraquinone/s |
| ATM | Antimicrobial |
| CFU | Colony forming units |
| CLSI | Clinical and Laboratory Standards Institute |
| EtOH | Ethanol |
| FeSO4 | Ferrous sulfate |
| FRAP | Ferric Reducing Antioxidant Potency assay |
| H2O2 | Hydrogen peroxide |
| HO∙ | Hydroxyl radical |
| MIC | Minimal Inhibitory Concentration |
| MOPS | Morpholine propane sulfonic acid |
| NaN3 | Sodium azide |
| NaNO2 | Sodium nitrate |
| NaOH | Sodium hydroxide |
| NBT | Nitro BlueTetrazolium |
| NCPF | National Collection of Pathogenic Fungi |
| NO· | Nitric oxide |
| O2•- | Superoxide radical anion |
| OD | Optical density |
| PBS | Phosphate Buffered Saline |
| pMIC | Photoactive Minimal Inhibitory Concentration |
| PS | Photosensitizer |
| PTN | Parietin |
| RNI | Reactive nitrogen intermediates |
| ROS | Reactive oxygen species |
| RPMI | Roswell Park Memorial Institute 1640 |
| SDA | Sabouraud Dextrose Agar |
| SDB | Sabouraud Dextrose Broth |
| SOD | Superoxide dismutase |
| MFC | Minimum Fungicidal Concentration |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| OH | Hydroxyl groups |
| DNA | Deoxyribonucleic acid |
| ONOO⁻ | Peroxynitrite |
References
- Ahmed, S.K.; Hussein, S.; Qurbani, K.; Ibrahim, R.H.; Fareeq, A.; Mahmood, K.A.; Mohamed, M.G. Antimicrobial Resistance: Impacts, Challenges, and Future Prospects. J. Med. Surg. Public Health 2024, 2, 100081. [Google Scholar] [CrossRef]
- Riera, F.O.; Caeiro, J.P.; Angiolini, S.C.; Vigezzi, C.; Rodriguez, E.; Icely, P.A.; Sotomayor, C.E. Invasive Candidiasis: Update and Current Challenges in the Management of This Mycosis in South America. Antibiotics 2022, 11, 877. [Google Scholar] [CrossRef]
- Invasive Candidiasis | Nature Reviews Disease Primers. Nat. Rev. Dis. Primer.
- Cieplik, F.; Deng, D.; Crielaard, W.; Buchalla, W.; Hellwig, E.; Al-Ahmad, A.; Maisch, T. Antimicrobial Photodynamic Therapy – What We Know and What We Don’t. Crit. Rev. Microbiol. 2018, 44, 571–589. [Google Scholar] [CrossRef] [PubMed]
- Klausen, M.; Ucuncu, M.; Bradley, M. Design of Photosensitizing Agents for Targeted Antimicrobial Photodynamic Therapy. Molecules 2020, 25, 5239. [Google Scholar] [CrossRef] [PubMed]
- Aware, C.B.; Patil, D.N.; Suryawanshi, S.S.; Mali, P.R.; Rane, M.R.; Gurav, R.G.; Jadhav, J.P. Natural Bioactive Products as Promising Therapeutics: A Review of Natural Product-Based Drug Development. South Afr. J. Bot. 2022, 151, 512–528. [Google Scholar] [CrossRef]
- Lichens as a Potential Source of Bioactive Secondary Metabolites | SpringerLink.
- Thakur, M.; Chander, H. Potential of Lichens: A Review of Bioactive Compounds with Biological Activities.
- Christina Pires Gonçalves, L. Photophysical Properties and Therapeutic Use of Natural Photosensitizers. J. Photochem. Photobiol. 2021, 7, 100052. [Google Scholar] [CrossRef]
- Polat, E.; Kang, K. Natural Photosensitizers in Antimicrobial Photodynamic Therapy. Biomedicines 2021, 9, 584. [Google Scholar] [CrossRef]
- Fernández-Marín, B.; Artetxe, U.; Becerril, J.M.; Martínez-Abaigar, J.; Núñez-Olivera, E.; García-Plazaola, J.I. Can Parietin Transfer Energy Radiatively to Photosynthetic Pigments? Molecules 2018, 23, 1741. [Google Scholar] [CrossRef]
- Comini, L.R.; Morán Vieyra, F.E.; Mignone, R.A.; Páez, P.L.; Laura Mugas, M.; Konigheim, B.S.; Cabrera, J.L.; Núñez Montoya, S.C.; Borsarelli, C.D. Parietin: An Efficient Photo-Screening Pigment in Vivo with Good Photosensitizing and Photodynamic Antibacterial Effects in Vitro. Photochem. Photobiol. Sci. 2017, 16, 201–210. [Google Scholar] [CrossRef]
- Hurtado Bredda, F.J.; Nin Vaeza, N.; Rubbo Amonini, H. Estrés oxidativo y nitrosativo en la sepsis. Med. Intensiva 2005, 29, 159–165. [Google Scholar] [CrossRef]
- Baltazar, L.M.; Ray, A.; Santos, D.A.; Cisalpino, P.S.; Friedman, A.J.; Nosanchuk, J.D. Antimicrobial Photodynamic Therapy: An Effective Alternative Approach to Control Fungal Infections. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.S.; Mawang, C.I.; Daniel-Jambun, D.; Lim, Y.Y.; Lee, S.M. Current Anti-Biofilm Strategies and Potential of Antioxidants in Biofilm Control. Expert Rev. Anti Infect. Ther. 2018, 16, 855–864. [Google Scholar] [CrossRef] [PubMed]
- M27-A2 Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard—Second Edition.
- Mugas, M.L.; Calvo, G.; Marioni, J.; Céspedes, M.; Martinez, F.; Sáenz, D.; Di Venosa, G.; Cabrera, J.L.; Montoya, S.N.; Casas, A. Photodynamic Therapy of Tumour Cells Mediated by the Natural Anthraquinone Parietin and Blue Light. J. Photochem. Photobiol. B 2021, 214, 112089. [Google Scholar] [CrossRef] [PubMed]
- Marioni, J.; Arce, J.E.; Cabrera, J.L.; Paraje, M.G.; Núñez Montoya, S.C. Reduction of Candida Tropicalis Biofilm by Photoactivation of a Heterophyllaea Pustulata Extract. Pharm. Biol. 2016, 54, 2791–2801. [Google Scholar] [CrossRef]
- M27-A3: Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard—Third Edition.
- Marioni, J.; Da Silva, M.A.; Cabrera, J.L.; Montoya, S.C.N.; Paraje, M.G. The Anthraquinones Rubiadin and Its 1-Methyl Ether Isolated from Heterophyllaea Pustulata Reduces Candida Tropicalis Biofilms Formation. Phytomedicine 2016, 23, 1321–1328. [Google Scholar] [CrossRef]
- Marioni, J.; Bresolí-Obach, R.; Agut, M.; Comini, L.R.; Cabrera, J.L.; Paraje, M.G.; Nonell, S.; Montoya, S.C.N. On the Mechanism of Candida Tropicalis Biofilm Reduction by the Combined Action of Naturally-Occurring Anthraquinones and Blue Light. PLOS ONE 2017, 12, e0181517. [Google Scholar] [CrossRef]
- Mattila, H.; Khorobrykh, S.; Havurinne, V.; Tyystjärvi, E. Reactive Oxygen Species: Reactions and Detection from Photosynthetic Tissues. J. Photochem. Photobiol. B 2015, 152, 176–214. [Google Scholar] [CrossRef]
- Tsikas, D. Analysis of Nitrite and Nitrate in Biological Fluids by Assays Based on the Griess Reaction: Appraisal of the Griess Reaction in the l-Arginine/Nitric Oxide Area of Research. J. Chromatogr. B 2007, 851, (1–2). [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive Candidiasis. Nat. Rev. Dis. Primer 2018, 4, 18026. [Google Scholar] [CrossRef]
- Youf, R.; Müller, M.; Balasini, A.; Thétiot, F.; Müller, M.; Hascoët, A.; Jonas, U.; Schönherr, H.; Lemercier, G.; Montier, T.; Le Gall, T. Antimicrobial Photodynamic Therapy: Latest Developments with a Focus on Combinatory Strategies. Pharmaceutics 2021, 13, 1995. [Google Scholar] [CrossRef] [PubMed]
- Fiala, J.; Roach, T.; Holzinger, A.; Husiev, Y.; Delueg, L.; Hammerle, F.; Armengol, E.S.; Schöbel, H.; Bonnet, S.; Laffleur, F.; Kranner, I.; Lackner, M.; Siewert, B. The Light-Activated Effect of Natural Anthraquinone Parietin against Candida Auris and Other Fungal Priority Pathogens. Planta Med. 2024, 90, 588–594. [Google Scholar] [CrossRef]
- Ma, W.; Liu, C.; Li, J.; Hao, M.; Ji, Y.; Zeng, X. The Effects of Aloe Emodin-Mediated Antimicrobial Photodynamic Therapy on Drug-Sensitive and Resistant Candida Albicans. Photochem. Photobiol. Sci. 2020, 19, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Quishida, C.C.C.; De Oliveira Mima, E.G.; Jorge, J.H.; Vergani, C.E.; Bagnato, V.S.; Pavarina, A.C. Photodynamic Inactivation of a Multispecies Biofilm Using Curcumin and LED Light. Lasers Med. Sci. 2016, 31, 997–1009. [Google Scholar] [CrossRef]
- Alshehri, A.H. Mechanical and Antimicrobial Effects of Riboflavin-Mediated Photosensitization of in Vitro C. Albicans Formed on Polymethyl Methacrylate Resin. Photodiagnosis Photodyn. Ther. 2021, 36, 102488. [Google Scholar] [CrossRef]
- Janeczko, M.; Masłyk, M.; Kubiński, K.; Golczyk, H. Emodin, a Natural Inhibitor of Protein Kinase CK2, Suppresses Growth, Hyphal Development, and Biofilm Formation of Candida albicans . Yeast 2017, 34, 253–265. [Google Scholar] [CrossRef]
- Manoharan, R.K.; Lee, J.-H.; Kim, Y.-G.; Lee, J. Alizarin and Chrysazin Inhibit Biofilm and Hyphal Formation by Candida Albicans. Front. Cell. Infect. Microbiol. 2017, 7, 447. [Google Scholar] [CrossRef]
- Tsang, P.W.-K.; Bandara, H.M.H.N.; Fong, W.-P. Purpurin Suppresses Candida Albicans Biofilm Formation and Hyphal Development. PLoS ONE 2012, 7, e50866. [Google Scholar] [CrossRef]
- Galinari, C.B.; Biachi, T.D.P.; Gonçalves, R.S.; Cesar, G.B.; Bergmann, E.V.; Malacarne, L.C.; Kioshima Cotica, É.S.; Bonfim-Mendonça, P.D.S.; Svidzinski, T.I.E. Photoactivity of Hypericin: From Natural Product to Antifungal Application. Crit. Rev. Microbiol. 2023, 49, 38–56. [Google Scholar] [CrossRef]
- Łopaciński, M.; Fiegler-Rudol, J.; Niemczyk, W.; Skaba, D.; Wiench, R. Riboflavin- and Hypericin-Mediated Antimicrobial Photodynamic Therapy as Alternative Treatments for Oral Candidiasis: A Systematic Review. Pharmaceutics 2024, 17, 33. [Google Scholar] [CrossRef] [PubMed]
- Amorim, C.F.; Iglesias, B.A.; Pinheiro, T.R.; Lacerda, L.E.; Sokolonski, A.R.; Pedreira, B.O.; Moreira, K.S.; Burgo, T.A.L.; Meyer, R.; Azevedo, V.; Portela, R.W. Photodynamic Inactivation of Different Candida Species and Inhibition of Biofilm Formation Induced by Water-Soluble Porphyrins. Photodiagnosis Photodyn. Ther. 2023, 42, 103343. [Google Scholar] [CrossRef]
- Souza, S.O.; Raposo, B.L.; Sarmento-Neto, J.F.; Rebouças, J.S.; Macêdo, D.P.C.; Figueiredo, R.C.B.Q.; Santos, B.S.; Freitas, A.Z.; Cabral Filho, P.E.; Ribeiro, M.S.; Fontes, A. Photoinactivation of Yeast and Biofilm Communities of Candida Albicans Mediated by ZnTnHex-2-PyP4+ Porphyrin. J. Fungi 2022, 8, 556. [Google Scholar] [CrossRef] [PubMed]
- Bors, W. Pulse-Radiolytic Investigations of Catechols and Catecholamines II. Reactions of Tiron with Oxygen Radical Species. Biochim. Biophys. Acta BBA - Gen. Subj. 1979, 582, 537–542. [Google Scholar] [CrossRef]
- Snyder, J.W.; Skovsen, E.; Lambert, J.D.C.; Poulsen, L.; Ogilby, P.R. Optical Detection of Singlet Oxygen from Single Cells. Phys. Chem. Chem. Phys. 2006, 8, 4280. [Google Scholar] [CrossRef] [PubMed]
- Vatansever, F.; De Melo, W.C.M.A.; Avci, P.; Vecchio, D.; Sadasivam, M.; Gupta, A.; Chandran, R.; Karimi, M.; Parizotto, N.A.; Yin, R.; Tegos, G.P.; Hamblin, M.R. Antimicrobial Strategies Centered around Reactive Oxygen Species – Bactericidal Antibiotics, Photodynamic Therapy, and Beyond. FEMS Microbiol. Rev. 2013, 37, 955–989. [Google Scholar] [CrossRef]
- Elian, C.; Méallet, R.; Versace, D. Photoactive Dye-Loaded Polymer Materials: A New Cutting Edge for Antibacterial Photodynamic Therapy. Adv. Funct. Mater. 2024, 34, 2407228. [Google Scholar] [CrossRef]
- Wink, D.A.; Mitchell, J.B. Chemical Biology of Nitric Oxide: Insights into Regulatory, Cytotoxic, and Cytoprotective Mechanisms of Nitric Oxide. Free Radic. Biol. Med. 1998, 25, (4–5). [Google Scholar] [CrossRef]
- Fraix, A.; Sortino, S. Combination of PDT Photosensitizers with NO Photodononors. Photochem. Photobiol. Sci. 2018, 17, 1709–1727. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, H.; Tao, X.; Xie, Y.; Yang, L.; Mao, Z.; Xia, W. Light-Triggered Nitric Oxide Release by a Photosensitizer to Combat Bacterial Biofilm Infections. Chem. – Eur. J. 2021, 27, 5453–5460. [Google Scholar] [CrossRef]
- Tavares, A.; Carvalho, C.M.B.; Faustino, M.A.; Neves, M.G.P.M.S.; Tomé, J.P.C.; Tomé, A.C.; Cavaleiro, J.A.S.; Cunha, Â.; Gomes, N.C.M.; Alves, E.; Almeida, A. Antimicrobial Photodynamic Therapy: Study of Bacterial Recovery Viability and Potential Development of Resistance after Treatment. Mar. Drugs 2010, 8, 91–105. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of parietin (1,8-dihydroxy-3-methoxy-6-methyl-9,10-anthraquinone, PTN).
Figure 1.
Chemical structure of parietin (1,8-dihydroxy-3-methoxy-6-methyl-9,10-anthraquinone, PTN).
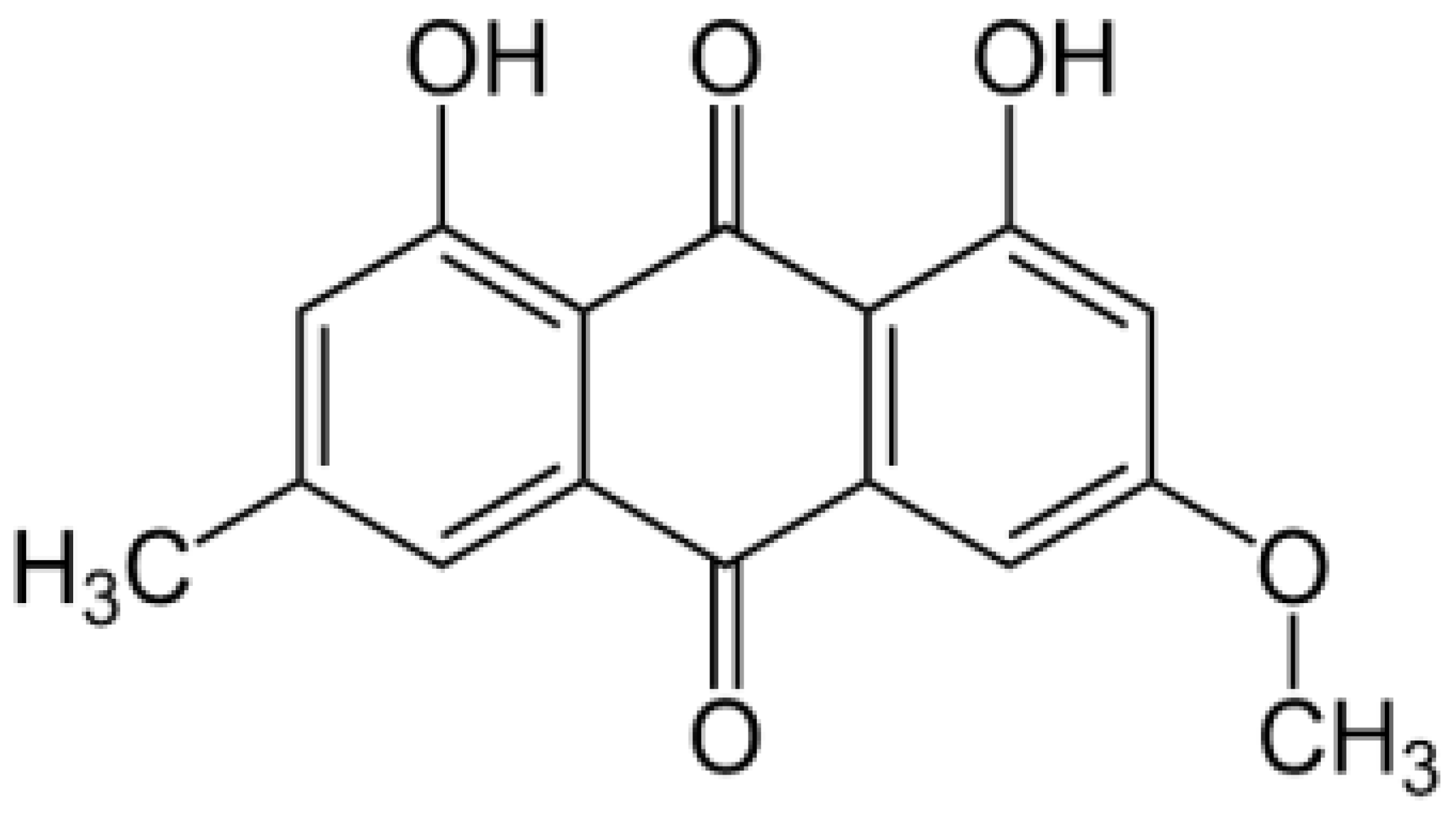
Figure 2.
MIC determination of parietin (PTN) under darkness and irradiation conditions. Optical density values obtained of planktonic cells at 530 nm, previously treated with different concentrations of PTN. * p < 0.05 respect to C. tropicalis. # p < 0.05 darkness vs. irradiation.
Figure 2.
MIC determination of parietin (PTN) under darkness and irradiation conditions. Optical density values obtained of planktonic cells at 530 nm, previously treated with different concentrations of PTN. * p < 0.05 respect to C. tropicalis. # p < 0.05 darkness vs. irradiation.
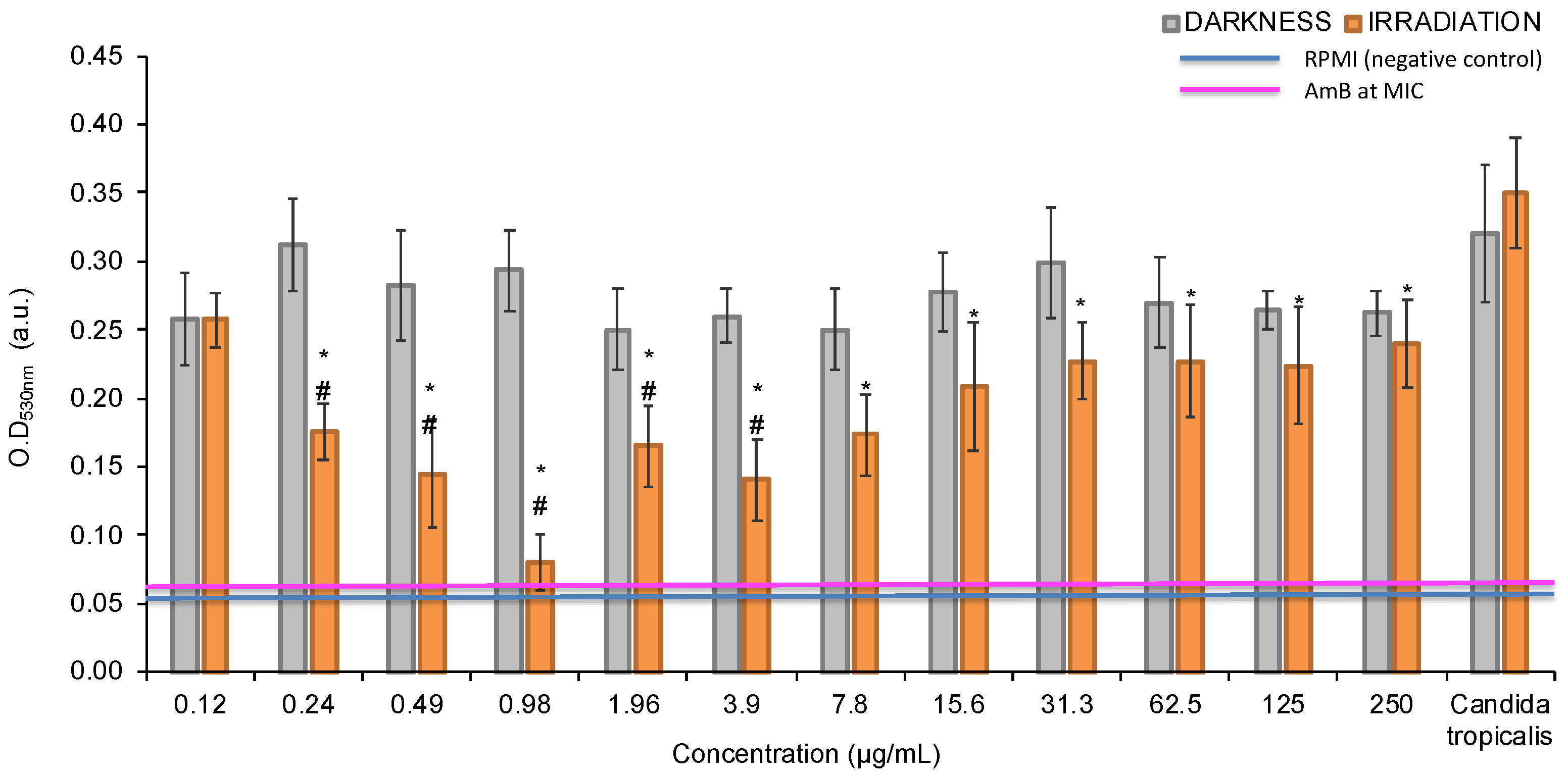
Figure 3.
Photoinactivation studies of C. tropicalis biofilms with parietin at five concentrations, using pMIC ((0.98 μg/mL ≈3.45 μM) as reference and actinic irradiation (L+) vs. darkness (L-). * p < 0.05 respect to C. tropicalis. # p < 0.05 darkness vs. irradiation.
Figure 3.
Photoinactivation studies of C. tropicalis biofilms with parietin at five concentrations, using pMIC ((0.98 μg/mL ≈3.45 μM) as reference and actinic irradiation (L+) vs. darkness (L-). * p < 0.05 respect to C. tropicalis. # p < 0.05 darkness vs. irradiation.
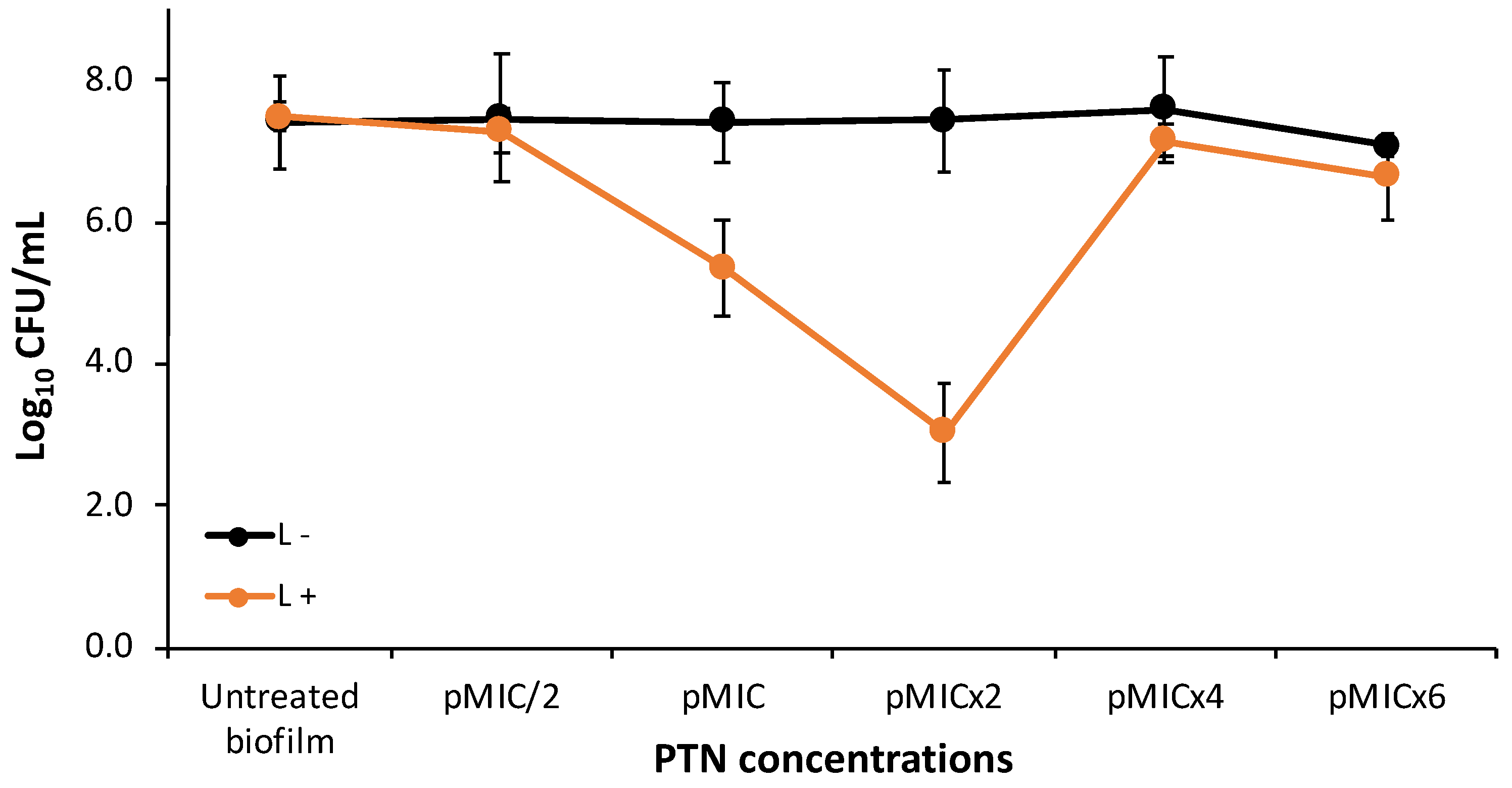
Figure 4.
Photoinactivation mechanism studies of C. tropicalis biofilms with parietin at five concentrations, using pMIC (0.98 μg/mL ≈3.45 μM) as reference and actinic irradiation (L+) vs. darkness (L-). * p < 0.05 respect to C. tropicalis. # p < 0.05 darkness vs. irradiation.
Figure 4.
Photoinactivation mechanism studies of C. tropicalis biofilms with parietin at five concentrations, using pMIC (0.98 μg/mL ≈3.45 μM) as reference and actinic irradiation (L+) vs. darkness (L-). * p < 0.05 respect to C. tropicalis. # p < 0.05 darkness vs. irradiation.
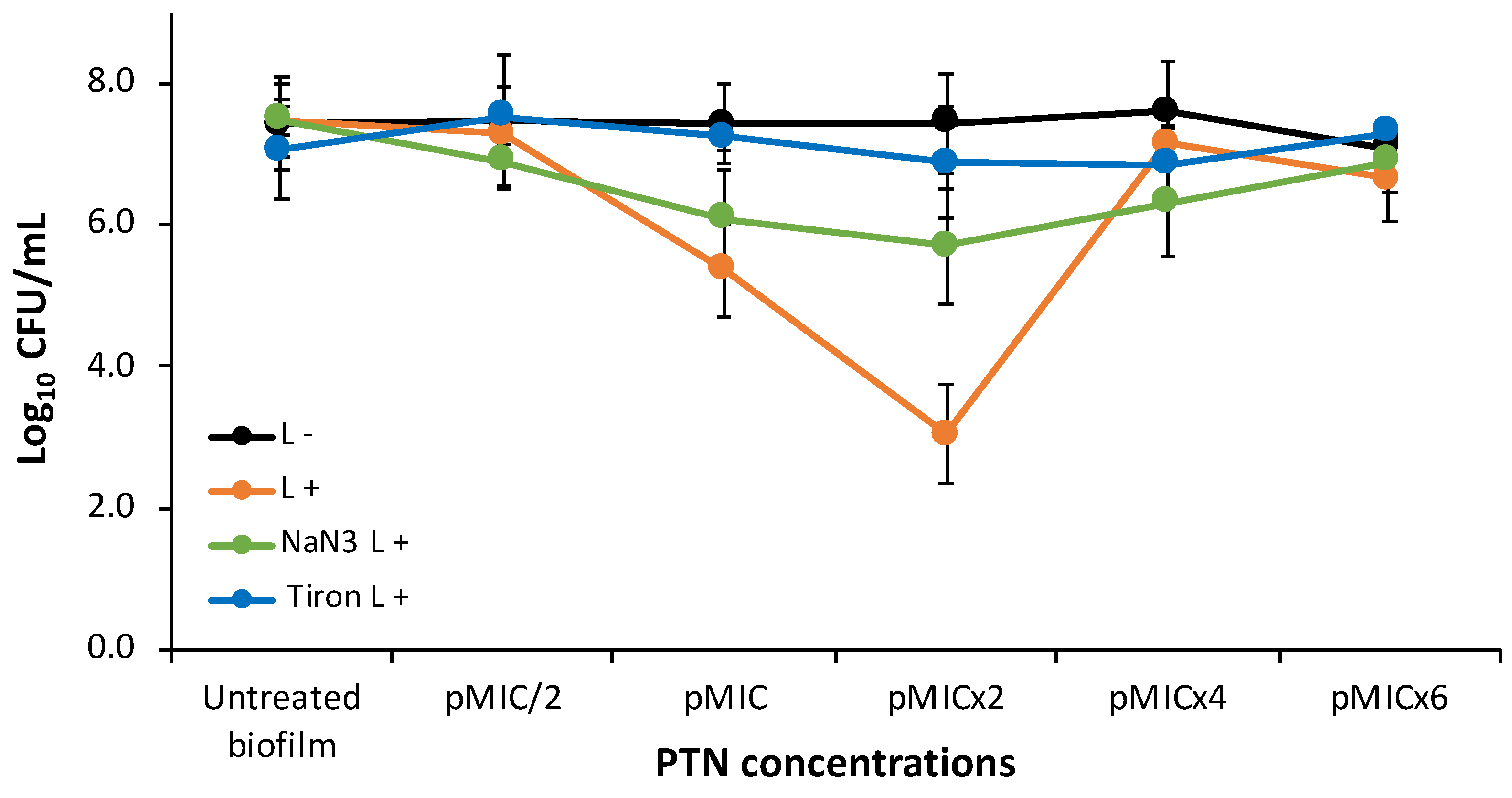
Figure 5.
Reactive oxygen species (ROS) and reactive nitrogen intermediates (RNI) production on C. tropicalis biofilm treated with parietin and irradiation, using pMIC (0.98 μg/mL ≈3.45 μM) as reference. Data obtained by the colony forming unit (CFU) count test, corresponding to viable biofilm. ON: nitric oxide. * p < 0.05 respect to C. tropicalis.
Figure 5.
Reactive oxygen species (ROS) and reactive nitrogen intermediates (RNI) production on C. tropicalis biofilm treated with parietin and irradiation, using pMIC (0.98 μg/mL ≈3.45 μM) as reference. Data obtained by the colony forming unit (CFU) count test, corresponding to viable biofilm. ON: nitric oxide. * p < 0.05 respect to C. tropicalis.
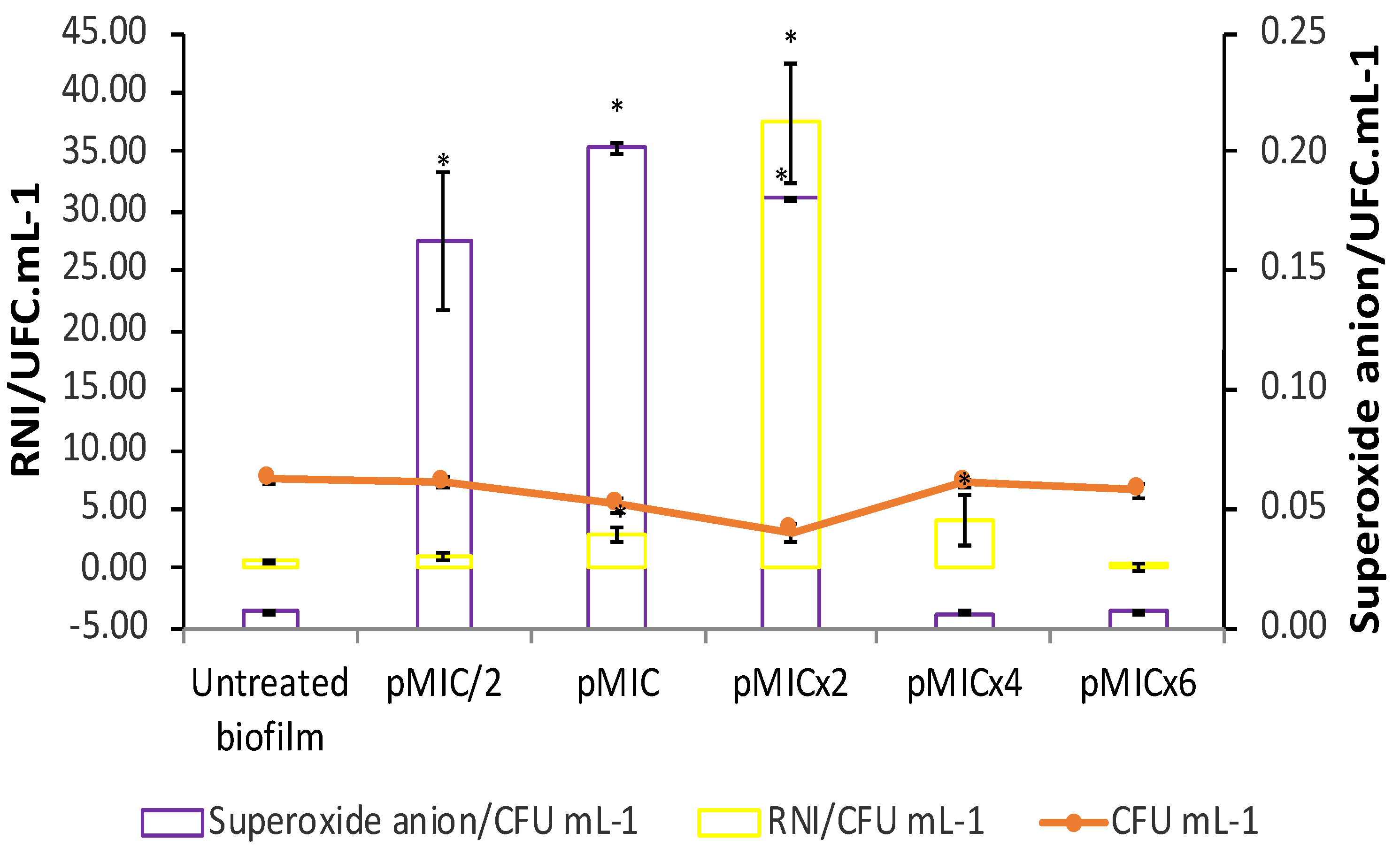
Figure 6.
Activation of superoxide dismutase enzyme (SOD) and total antioxidant capacity (FRAP) in C. tropicalis biofilm treated with parietin and irradiation, using pMIC (0.98 μg/mL ≈3.45 μM) as reference. Data obtained by the colony forming unit (CFU) count test, corresponding to viable biofilm. * p < 0.05 compared to C. tropicalis.
Figure 6.
Activation of superoxide dismutase enzyme (SOD) and total antioxidant capacity (FRAP) in C. tropicalis biofilm treated with parietin and irradiation, using pMIC (0.98 μg/mL ≈3.45 μM) as reference. Data obtained by the colony forming unit (CFU) count test, corresponding to viable biofilm. * p < 0.05 compared to C. tropicalis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



