Submitted:
13 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
Ovarian cancer remains a global health challenge characterized by frequently delayed diagnosis and alarming mortality rates. Recent progress in the crosstalk between gut and vaginal microbiomes in ovarian cancer provides new insights into the complicated pathogenesis of ovarian cancer. Intestinal and vaginal dysbiosis exerts a significant in-fluence on the occurrence and development of ovarian cancer mainly through three principal pathways: estrogen metabolism potentiation, chronic inflammatory priming, and DNA epigenetic reprogramming. Recent research has provided robust evidence of great associations between dysbiosis and ovarian cancer. In this review, we summarize the characteristics of gut and vaginal microbiomes in patients with ovarian cancer and the potential mechanisms of the microbiomes promoting the development of ovarian cancer. Moreover, we discuss evidence and mechanisms of the interaction between gut and vaginal microbiomes. We believe that effective prevention and early diagnostic models will be established in the future combining microbiomes with other associated factors of ovarian cancer including genetic, environmental, lifestyle factors, etc., which is essential for reducing the high morbidity and mortality of ovarian cancer. We hope to shed light on the gut and vaginal microbiomes in the development of ovarian cancer and the potential clinical applications in this review.
Keywords:
ovarian cancer
; gut microbiomes
; vaginal microbiomes
; diagnosis
1. Introduction
Ovarian carcer remains the deadliest gynecologic malignancy worldwide in 2022, with 324,398 incident cases and 206,839 mortality events globally. [1] It’s estimated that the new cases and related deaths in America by 2023 are 19,710 and 13,270, respectively [2]. Notably, more than 60% of women with ovarian cancer are diagnosed at the advanced stage [3], driving the 5-year survival rate of patients in stage IV below 20% [4]. Over the past 30 years, first-line treatment strategies—comprising debulking surgery combined with platinum-based chemotherapy and paclitaxel—have seen limited advancements since the mid-1990s [5]. Though progress in immunotherapies, including poly ADP-ribose polymerase (PARP) inhibitors (e.g. Olaparib, Rucaparib, and Veliparib) [6,7,8]and the anti-angiogenic agent (e.g. bevacizumab) have been deployed clinically, ovarian cancer continues to exhibit a high mortality rate [9]. Diagnostic delay and limited therapy are the leading factors accounting for the high mortality rate [10,11,12].
The risk factors of ovarian cancer remain not yet fully elucidated. Genetic factors [13,14], environmental factors [15], lifestyle factors [16], reproductive factors [17], and microbiome factors have all been implicated in the complex pathogenesis of ovarian cancer. (Figure 1) In recent years, microbiome factors in ovarian cancer have attracted more attention from researchers due to their potential roles in the pathogenesis.
The gut and vaginal microbiomes, as integral parts of the human microbiome, have attracted considerable attention for their relations with human’s health and the crosstalk between them [18,19,20,21,22,23]. Extensive studies have demonstrated a substantial correlation between gut and vaginal microbiomes and ovarian cancer [24,25]. Unique characteristics of gut and vaginal microbiomes of patients with ovarian cancer have been revealed by some clinical trials [25,26]. Further studies have revealed the potential mechanisms have been reported by some research. Gut and vaginal microbiomes affect the development of ovarian cancer mainly through increasing the levels of estrogen [27], regulating inflammatory responses [28], and modulating epigenetic modifications [29]. These results further indict the potential clinical applications of gut and vaginal microbiomes for prevention and early diagnosis.
In this review, we briefly summarize the characteristics of gut and vaginal microbiomes and discuss the correlation between intestinal and vaginal microbiomes and ovarian cancer. We hope to provide a possible solution to prevention and early diagnosis for patients with ovarian cancer from the microbiome point of view.
2. Characteristics of Gut and Vaginal Microbiota in Ovarian Cancer
2.1. Characteristics of Gut and Vaginal Microbiota in Health Women
Gut microbiota: The dominant phyla in the gut of healthy human are Bacteroidetes and Firmicutes, accounting for over 90% of gut microbiota [30]. Phyla Actinobacteria, Proteobacteria, and Verrucomicrobia are common but generally the minor components [31]. The composition of the gut microbiota can be influenced by a variety of factors such as genetic variation, age, diet, drug, etc. and is highly variable between different individuals [32]. Researchers have found obvious differences in the gut microbiota between males and females [33]. Compared with males, a higher α diversity of gut microbiota and a lower abundance of Bacteroidetes were observed in females [34,35]. In addition, one recent research revealed that the ratio of Firmicutes to Bacteroidetes in the gut was significantly higher in female in comparison with male [36]. A variety of metabolically protective bacteria including Akkermansia muciniphila and Alistipesshahii are notably abundant in the intestine of females over males [37]. The levels of estrogen may result in the difference of gut microbiota between male and female [38]. Study has proved that postmenopausal women present with a similarity of the gut microbiota with men [39].
Vaginal Microbiota: The vagina is colonized by a group of microbes. In healthy reproductive-age women, the vaginal microbiota is dominated by Lactobacillus spp. including Lactobacillus crispatus, Lactobacillus gasseri, Lactobacillus iners, and Lactobacillus jensenii [40]. The component of the vaginal microbiota changes dynamically with age, estrogen, living style, etc. [41] To describe its taxonomic composition the term "community state type"(CST) is raised [42]. Vaginal microbiota can be briefly divided into 5 main CSTs according to the subtypes of Lactobacillus, including CST I, II, III, V, and CST IV [43]. The CST I, II, III, and V are dominated by Lactobacillus while CST IV is characterized by a lower proportion of Lactobacillus spp. and a higher proportion of anaerobic bacteria [40,44,45]. CST IV is associated with the development of bacterial vaginosis, which is always accompanied with common gynecological diseases including sexually transmitted diseases, pelvic inflammatory disease, endometriosis, gynecological cancers, etc.[46,47]
2.2. Characteristics of Gut and Vaginal Microbiota in Patients with Ovarian Cancer
The change of components of the gut and vaginal microbiota in women with ovarian cancer has been observed in many studies. Understanding the unique characteristics of the microbiota in these patients can greatly help understand the development of ovarian cancer [48,49].
Patients with ovarian cancer show gut microbiota dysbiosis in comparison with healthy controls. Hu et al. [50] observed decreased Shannon index and α diversity in 20 patients with EOC. The β diversity was observed gradually decreasing with the progression of ovarian cancer in 30 patients [48]. D’ Amico et al. [51] analyzed the gut microbiota in 24 ovarian cancer patients and 24 health controls. They found a decrease in Lachnospiraceae, Bifidobacteriaceae, Clostridiaceae, Rikenellaceae, and Porphyromonadaceae while an increase in Coriobacteriace (Adlercreutzia and Collinsella) Lactococcus, and Lachnobacterium in ovarian cancer patients compared to the healthy control [51]. The critical role of intestinal dysbacteriosis in the development of ovarian cancer was further demonstrated by the studies of epithelium ovarian cancer (EOC) mice. The decreased relative abundance of Actinobacteria phyla, Bifidobacterium, Ruminococcaceae, and Ruminococcus and increased relative abundance of Proteobacteria, Bacteroides, and prevotella were also found in the EOC group. It should be noted that the differences were more significant in advanced stages. More importantly, the growth of tumors was significantly accelerated in the EOC mice with antibiotic deployment, compared with control mice. Chambers et al. [52] found in mice injected with EOC lines that the EOC tumor growth is significantly increased in mice treated with antibiotic-containing water compared to mice in the control group. These findings indicated that gut microenvironment homeostasis might exert inhibitory effects on ovarian cancer. (Table 1)
Obvious changes in the vaginal microbiota are also found in patients with ovarian cancer. In a multicentral case-control study including 360 ovarian cancer, 115 healthy controls, and 69 controls with benign gynecological conditions, Nené et al. [25] divided the vaginal microbiota into two community types, the community type O with less than 50% of Lactobacillus while the community type L on contrary, with at least 50% of Lactobacillus. They claimed that female patients younger than 50 years old had a significantly higher prevalence of community type O microbiota than age-matched controls. This indicates that community type L or Lactobacillus may be a protective factor in ovarian cancer. Jacobson et al. [26] also observed a less proportion of Lactobacillus-dominated vaginal community in ovarian cancer compared to age-matched healthy women. In addition, Asangba et al. [48] observed the difference in α diversity and β-diversity in the lower genital tract of patients with ovarian cancer compared with benign ones and a group of microbes in the lower genital tract that differ in different stages and grades of ovarian cancer. Moreover, vaginal infections including Human Papillomavirus (HPV) [53], Chlamydia trachomatis (CT) [54], and Neisseria gonorrhea [55] have been observed to be connected to the development of ovarian cancer. (Table 1)
These studies indicate the potential association between gut and vaginal microbiomes and ovarian cancer. The characteristics of the microbiomes in patients with ovarian cancer can possibly become the auxiliary diagnostic method of ovarian cancer. Possible mechanisms of how gut and vaginal microbiomes promote the development of ovarian cancer will be described as follow parts.
3. Interaction of Gut and Vaginal Microbiome
3.1. Evidences of the Interaction of Gut and Vaginal Microbiomes
The gut and vagina are anatomically neighbors so it’s possible to assume the existence of a relation between the microbiome in these two organs. In fact, some studies have proved that there is a degree of interaction between the human vaginal microbiota and gut microbiota. In the following, we summarize several evidences of the possible relationship of the gut and vaginal microbiota.
Similar compositions of gut and vaginal microbiota have been presented. A cross-sectional study revealed the correlation of rectal and vaginal microbiota in women who are not pregnant [56]. They found Lactobacilluscrispatus (16%), Lactobacillus jensenii (10%), and Lactobacillus gasseri (10%) were the prevalent lactobacilli colonizing the rectums in most females, which is surprisingly according with the proportion in their vagina. [56]. Another research assessed the rectal and vaginal microbiota in 132 pregnant women at 35-37 weeks of gestation. They found almost half of the bacterial species present in both the rectum and vagina. [57]. A recent study showed similar results. Shin et al. [58] found that from the last trimester of pregnancy to the second month after birth, the composition of the microbiota in these two places became closer. In another study, bacterial vaginosis associated bacteria were detected in both vagina and rectum in women with CST IV (non-Lactobacillus-dominant). This suggested that the rectum stored the bacterial vaginosis associated bacteria of CST IV [59]. The high homogeneity of the microbiota colonized between the vagina and rectum provides strong evidence that the rectal microbiota serves as a reservoir for the vaginal microbiota. Moreover, some evidences have vaguely presented interaction between gut and vaginal microbiomes. the probiotics given orally can change the microbiota in the vagina [60]and cure the Gardnerella vaginalis induced bacterial vaginitis [61]. Oral medication can regulate the vaginal microenvironment, indicating the innate relations between the gut and vagina. Current research also suggested that the vaginal microbiological environment of the mother has a close influence on the gut microbiota of the infant, which is called vaginal seeding [62]. The gut microbial composition of infants born by cesarean section differs from that of neonates born vaginally over a certain period. It may affect neonatal health in terms of obesity [63], and immune-related diseases [64].
Though no definite evidence confirms the interaction between gut and vagina microbiota, the similar composition and the Vaginal Seeding for infants strongly suggest potential association between them.
3.2. Possible Mechanisms of the Interaction of Gut and Vaginal Microbiomes
More researches on the interaction between microbiome located in different parts of the body are still warranted. With the improvement of research techniques and the accumulation of experimental content, we’ll complete the missing parts of the interaction between gut and vaginal microbiomes and get a better understanding of the link between gut and vaginal microbial interaction and ovarian cancer in the future.
Some evidences indicate that gut and vaginal microbiomes may interact with each other via the gut-vaginal axis. The gut-vagina axis is described as two-side interactions between gut and vaginal microbes. In the gut-vagina axis, the gut influences the vaginal microbiome primarily through the estrobolome, but the influence of the vaginal microbiome on the gut microbiome has not yet been revealed. Estrobolome is defined as a part of the gut microbiome with the ability to metabolize estrogen and regulate the levels of estrogen through the estrogen-gut microbiome axis. In other word, estrobolome is an aggregate of genes in gut microbes with the capability of secreting β-glucuronides for estrogen metabolism [65]. Conjugated estrogen can be deconjugated by those enzymes and then return to blood circulation through enterohepatic circulation, resulting in a high level of estrogen in the body [65,66,67]. High levels of circulating estrogen stimulate the production of vaginal glycogen which then promotes the colonization of Lactobacillus in the vagina, thus influencing the vaginal microbiome [68]. Although the interaction mechanisms between vaginal microbes and gut microbiota are largely unknown recently, the link between obesity, gut microbiome dysbiosis, and vaginal microbiota composition suggests the two-side effects of the gut-vagina axis.
More researches on the interaction between microbiome located in different parts of the body are still warranted. With the improvement of research techniques and the accumulation of experimental content, we’ll complete the missing parts of the interaction between gut and vaginal microbiomes and get a better understanding of the link between gut and vaginal microbial interaction and ovarian cancer in the future.
4. Potential Mechanisms of Gut and Vaginal Microbiomes in Ovarian Cancer
Emerging evidence substantiates a significant association between dysbiosis of gut and vaginal microbiota with ovarian cancer. The gut and vaginal microbiota affect the development of ovarian cancer mainly through two routines, the microbe-associated molecular patterns (MAMPs) and metabolites [28,69]. MAMPs represent conserved structural components of microorganisms, including bacterial flagellin and lipopolysaccharide (LPS) [70] which mediate host-microbiome interaction by combining with Pattern Recognition Receptors (PRRs) such as toll-like receptors (TLRs) and nucleotide-binding oligomerization domain-containing protein-like receptors (NLRs) and then activate downstream pathways to regulate the physiological and pathological processes of the host [69]. Metabolites produced by microbiota are also responsible for interacting with the host cells [28]. The gut microbiota can produce metabolites including estrogen, SCFAs, secondary bile acid, indolepropionic acid (IPA), conjugated linoleic acid (CLA), and even genotoxic metabolites. While the vaginal microbiota produces distinct metabolic products including lactic acid, hydrogen peroxide(H2O2), SCFAs, peptides, phospholipid, nucleotides, etc. [71] that contribute to regulate host physiological and pathological processes, even cancer genesis [72]. Herein, we conclude the most characterized mechanisms through which gut and vaginal microbiota influence ovarian cancer progression.
4.1. Gut and Vaginal Microbiomes and Estrogen in Ovarian Cancer
In general, estrogen is produced by the ovarian, adrenal gland, and adipose tissue with additional contributions from dietary sources [33]. Hepatic metabolism converts estrogens into conjugated forms via glucuronidation (UDP-glucuronosyltransferase) and sulfation (sulfotransferases), facilitating biliary excretion or enterohepatic recirculation [73]. Circulating estrogen exist in three main forms including estrone, estradiol, and estriol, in either free or protein-bound stats [74]. Estrogen combines with classical nuclear receptors (estrogen receptors α and β, ER-α and ER-β) or non-classical membrane receptors (G-protein-coupled estrogen receptor, GPER) [75,76]. Estrogen and their metabolites can also be processed by the gut microbiome [77], and enrolled in the enterohepatic circulation, reabsorbed into the circulation system to regulate physiological activities. It's widely known that high estrogen levels and ER-positive [78] are risk factors for ovarian cancer. Patients with early menarche, late menopause, and fertility drugs are at a higher risk of developing ovarian cancer [79].
The gut microbiome possibly affects the development of ovarian cancer via the regulation of estrogen levels in two main routines, estrobolome and gut-brain axis. (Figure 2) The gut microbiome can regulate the levels of estrogen through the estrobolome [80]. The estrobolome is mainly composed of Firmicutes and Bacteroidetes [81]. A total of 279 β-glucuronidase proteins encoded by gut microbes have been identified in the Human Microbiome Project database [67,81]. The β-glucuronides produced by estrobolome can uncouple the conjugated estrogen, promote the reabsorption of free estrogen through enterohepatic circulation and increase the total estrogen burden [65]. On the other hand, gut microbiome can also affect the generation of estrogen through gut-brain axis. Gut microbiome can produce metabolites like gamma-aminobutyric acid and norepinephrine acting as neurotransmitters, stimulating local nerve cells or vagus nerves and then affecting brain cells [82]. These effects may lead to stress reactions such as depression and anxiety [83], closely associated with over activation of the hypothalamic-pituitary-adrenal (HPA) axis [84]. The activation of HPA axis in stress circumstances is linked with the generation of gonadal hormones [85], leading to the change of estrogen levels. However, little is known about the impact of vaginal microbes on estrogen.
Available evidence suggested that high unconjugated estradiol levels are positively correlated with a high risk of non-plasmacytoid ovarian cancer [86,87]. Excessive estrogen may promote the proliferation of ovarian epithelial cells [88] and then accelerate the progression of ovarian cancer through receptor-dependent pathways (ER-α and GREP) and receptor-independent pathways (cytochrome P, CYP450 enzymes). ER-α combined with estrogen can activate downstream oncogenesis genes including c-fos, c-myc, and HER2/neu. Moreover, the combined ER-α can active mitogen-activated protein kinase (MAPK) [89] and Wnt/β-catenin signaling pathways [90], which are closely associated with the development of ovarian cancer [91,92]. The GREP induces the second messenger system such as extracellular regulated protein kinases (ERK) and the Phosphatidylinositol-3-kinase (PI3K) [93], promoting the proliferation of cancer cells [27]. In addition, the metabolites of estrogen like quinone intermediates via CYPs can elevate the level of free radicals and induce the mutation of genes [27]. (Figure 2)
In summary, gut and vaginal microbiomes interact with each other via gut-vaginal axis. The disturbance of the gut microbiome changes the estrogen levels via estrobolome and gut-brain axis. The estrogen then combines with the receptors and promotes the proliferation of cancer cells. Meanwhile, the metabolites of estrogen can lead to DNA damage, which results in the development of ovarian cancer [27].
4.2. Gut and Vaginal Microbiomes and Inflammation in Ovarian Cancer
The critical role of inflammation has been well proved in carcinogenesis. Acute inflammation mediates antitumor immunity through dendritic cell activation and antigen presentation while chronic inflammation leads to a cancer-promoting niche [94]. Long-term preserving acute inflammation turns into chronic inflammation, which leads to continuous inflammatory signaling activation and formation of hypoxic and acidic microenvironment. And then the aggregation of immunosuppressive cells and activation of protooncogene lead to damage of DNA, and at last tumor genesis [95]. Notably, the risk factors of ovarian cancer such as psychological factors [15,96,97]and obesity [98] have been proven to promote ovarian cancer through inflammation associated pathways with the help of cytokines including interleukins and interferons [99]. The inflammatory response is mainly regulated by microbe associated molecular patterns (MAMPs) and the metabolites [69] [100,101]. (Figure 3)
The MAMPs of the gut microbiota interact with PRRs and then cause the host inflammatory reaction. TLR-4 is one of the most reported PRRs in ovarian cancer. Gut microbiota-derived LPS activates TLR4/NF-κB signaling in ovarian carcinoma cells via myeloid differentiation factor 88 (MYD88) adaptor recruitment. [102] Then increase the levels of proinflammatory cytokines including tumor necrosis factor α (TNF-α), interleukin 6 (IL-6), interleukin 8 (IL-8) and monocyte chemotactic protein 1 (MCP-1) combining with anti-apoptotic family activation. This may promote the development of ovarian cancer by stimulating pro-angiogenic cytokines and anti-apoptotic responses [103,104].
Emerging evidence further implicates TLR4 crosstalk with Hedgehog (Hh) pathway. Hu et al. [50] demonstrated the pathway in murine EOC model. The gut microbiota from mice with EOC could promote the proliferation of tumors, while Hh inhibition (GANT61) significantly restrained the tumor progression [50]. Gut microbiota promotes EOC via Sonic Hedgehog (Shh) /Gli-1 axis. NF-κB regulates the Hh signaling pathway by increasing the production of TNF-α and IL-1β, enhancing the activity of Sonic Hedgehog (Shh) promoter, and promoting the expression of Gli-1[50,105]. As a result, it increases migration and invasion of cancer cells. [106] A retrospectively including 102 patients revealed association between TLR4 and tumor associated macrophage (TAM). The over expression of TLR4 on macrophage and the increasing levels of TAM biomarkers are linked to reduced progression-free survival [107]. This correlation was further confirmed by Xu et al. with murine model [108]. Gut microbiota dysbiosis induced by high-dose antibiotics was associated with ovarian cancer progression. Gut microbiota dysbiosis enhanced the secretion of TAM-derived TNF-α and IL-6, and then promoted the epithelial-mesenchymal transition (EMT) procedure. As a result, the development of ovarian cancer was accelerated. (Figure 3)
TLR5 is reported to be associated with ovarian cancer via activating MyD88/ TNF receptor associated factor 6 (TRAF6) signaling. Rutkowski et al. [24] suggested that TLR5 in microbiota increased the levels of IL-6 and activating the adaptor molecules MyD88 and TRAF6. Then the recruitment of Myeloid-derived suppressor cells and γδ lymphoid cells promoting the secretion of Galectin-1, which accelerated the growth of tumor cells. This result was further proved in EOC patients. Patients with TLR5 deficient were claimed to have a higher long-term survival rate when compared with those without TLR5 deficient [24]. Moreover, IL-6 was also demonstrated to be able to activate the Janus Kinase-signal transducer and activator of transcription 3 pathway (JAK-STAT3) and MAPK pathways, which are closely associated with the aggressive disease course and poor prognosis of high-grade ovarian cancer [109].(Figure 3)
Beyond MAMPs, gut microbiota-derived metabolites, including secondary bile acids, SCFAs, IPA, genotoxic metabolites, etc. pose critical influence on ovarian cancer development. [28] These metabolites act locally or at distal sites through circulation system [110] and affect ovarian carcinogenesis through promoting cell apoptosis, scavenging free-radicals and suppressing inflammation [100,101].
Gut microbiota can uncouple conjugated bile acids via Bacterial bile salt hydrolase (BSH) enzymes and further process and modify them to generate secondary bile acids, including deoxycholic acid (DCA) and lithocholic acid [111]. Secondary bile acids can inhibit the proliferation of ovarian cancer. DCA and ursodeoxycholic acid (UDCA) can induce cell apoptosis through a PKC-independent pathway in the ovarian cell line (A2780) and cisplatin-resistant cell line(A2780-CP) [112]. This result further proves the anti-tumor effect of secondary bile acid on ovarian cancer. SCFAs, generally including acetate, propionate, and butyrate, are a group of compounds fermented from undigested carbohydrates by intestinal flora such as Bacteroides, Roseburia, Bifidobacterium, Fecalibacterium, and Enterobacteria [113]. SCFAs were claimed to be implicated in the promotion of macrophage differentiation and inhibition of the NF-κB pathway in cancer patients, which can inhibit carcinogenesis [114]. Tryptophan can be metabolized by the gut microbiota, and then produce indole compounds including IPA [115]. IPA has been proven with anti-tumor effects in ovarian cancer. A study found a decreased level of IPA in patients with ovarian cancer. And it further suggested that IPA can scavenge free radicals and was responsible for anti-tumor function. [116] In addition, intestinal dysbacteriosis can also promote carcinogenesis by producing genotoxic metabolites (e.g. Colibactin) and promoting a pro-inflammatory state that contributes to cancer progression [117]. The research is still limited in the field of gut microbiota metabolisms and ovarian cancer. (Figure 3)
Moreover, the metabolism including secondary bile acids, CLAs, and SCFAs can suppress inflammation via anti-obesity mechanisms. Evidence shows those microbiota metabolites can suppress obesity and the adipocytokines such as TNF-α and IL-6. It has been shown that Bile acids may affect obesity via altering energy, lipid, and glucose metabolism [118]. Available evidence indicated that levels of secondary bile acids are abnormal in obese patients and may be associated with pathological changes. Secondary bile acids can reduce inflammation and lipoprotein uptake, and decrease the development of atherosclerosis [119]. CLAs are a family of fatty acids with weight-loss properties, widely found in beef and dairy products and produced by some bacteria in the Firmicutes and Actinobacteria phyla. Recent studies reported that CLAs can promote weight loss by reducing the synthesis of anti-inflammatory eicosanoids, activating PPARs involved in lipid metabolism and immune function, promoting the browning of white adipose tissue [120], and reducing adiponectin secreted by adipose cells [121]. There is a contradiction between the anti-obesity and pro-obesity effects of SCFAs. On the one hand, SCFAs can increase intestinal energy accumulation and promote the development of obesity. On the other hand, SCFAs increase energy expenditure and anorexia hormone production, reduce appetite, and maintain the intestinal mucosal barrier integrity to inhibit obesity. In conclusion, SCFAs contribute to the improvement of insulin sensitivity and play a beneficial role in maintaining normal metabolism and function of multiple organ systems [122,123].
The vaginal microbiota is also essential in the prevention of infection by external pathogens and the maintenance of an immune tolerant environment. Several studies have linked reproductive tract infections with an increased risk of ovarian cancer [124,125]. Commensal vaginal microbiota is dominated by lactobacillus, which can protect the host from infection by producing lactic acid and H2O2[56]. Rahbar et al. [126] found that Lactobacillus, isolated from the vagina, enabled to induce the apoptosis of CAOV-4 cells and suppress the development of ovarian cancer by down-regulation of miR-21, miR-200b, and TLR-4. Available evidence indicated that vaginal dysbacteriosis is associated with an increased risk of infection and ovarian cancer. Patients with ovarian cancer are observed with less Lactobacillus and more pathogens including Chlamydia trachomatis and HPV in the vagina [25]. Chlamydia trachomatis can block caspase 3 and cytochrome C release and inhibit apoptosis, which can promote the development of ovarian cancer [125]. In addition, Chlamydia trachomatis can also active the TLRs in the epithelial cells and promote the ovarian cancer carcinogenesis [127]. Other cancerogenic mechanisms associated with pathogen infection include regulation of DNA damage repairing, degeneration of p53, and stimulation of the MAPK signaling pathway [125].
4.3. Gut and Vaginal Microbiomes and Inflammation in Ovarian Cancer
The genetic factor is an essential risk factor for ovarian cancer. Although identified susceptibility genes have offered insights into the complex genetic basis of ovarian cancer, the identified risk loci only explain a small part of overall disease variance. The epigenetic modifications, mainly including methylation and histone modifications, mediating the interaction between genetics and environment, have been widely observed in the development of ovarian cancer. Some studies reported that the methylation of 59 cytosine phosphate guanine (CpG) rich regions was increased in many kinds of tumors, including ovarian cancer. methylated CpG islands are implicated in the regulation of cell cycle, apoptosis, and drug-sensitive gene silencing. Methylation of some risk genes such as BRCA1, RASSF1A, and PTEN has been demonstrated in ovarian cancer [29,128,129]. Histone modifications, such as acetylation and methylation are implicated in the pathogenesis of ovarian cancer [130]. The loss of acetylation of histone H4 lysine 16 (H4K16) in ovarian cancer tissue has been reported associated with the development of ovarian cancer [131]. Meanwhile, the histone methyltransferases, responsible for histone methylation, such as enhancers of zeste homologue 2 (EZH2), disruptors of telomeric silencing-1-like (DOT1L), and protein arginine methyltransferase (PRMT) are reported in correlation with the development of ovarian cancer [130]. Microbiota are closely related to host cell epigenetics. Helicobacter pylori can cause host gene methylation and suppress damaged DNA repairing in gastric cancer and colorectal malignant tissues [132,133,134]. Fusobacterium is also reported to accelerate the methylation of colorectal cancer in patients with UC, which is closely associated with the development of colorectal cancer [135]. In addition, some metabolites such as SCFAs can also influence the epigenetics in host cells [136,137]. Among SCFAs, butyrate has been reported with the activity of histone deacetylase (HDAC) inhibitor in the nucleus to prevent cell multiplication and trigger apoptosis [137]. Moreover, the folate and some B vitamins produced by gut microbiota can participate in the methylation of DNA and histone by providing methyl groups [138].
Although recent research has revealed epigenetic modifications in ovarian cancer, the role of microbiota remains unknown in this procedure [29,128]. Some studies indirectly suggested a link between microbe-induced alterations in host epigenetics and ovarian cancer development. In patients with vaginal Chlamydia trachomatis, Chlamydia trachomatis infection induces methylation of the calreticulin promoter, which in turn leads to a down-regulation of calreticulin expression and Epithelial–mesenchymal transition (EMT) [139]. Chlamydia trachomatis infection is the leading cause of pelvic inflammatory disease (PID). Chlamydia trachomatis infected women have been shown to have a higher risk of ovarian cancer than non-infected women [124]. The difference may be related to its ability to induce EMT in ovarian epithelial cells. In humans with Firmicutes dominated gut microbiota, a total of 803 genes with differentially methylated promoters are claimed to be associated with lipometabolism, obesity, and the inflammatory response, while obesity and inflammation have been verified as the risk factors of ovarian cancer [140]. More efforts are needed to clarify how specific microbiota or metabolites can regulate epigenetic modifications and then affect the development of ovarian cancer. Key metabolites and critical signaling pathways among them may become novel targets for epigenetic-based therapies for ovarian cancer.
5. Discussion
Ovarian cancer is a complex disease with an unclear pathogenesis and complex aetiologic factors. The disease prediction and early diagnosis of ovarian cancer are far from satisfactory. The unclear pathogenesis and delayed diagnosis in ovarian cancer are urgent to be solved. Recent advances in microbiomes suggest the potential of microbiota in the diagnosis and management of human disease [141,142,143] and offer us an entirely new way to better understand the mechanism of ovarian cancer and the opportunity to optimize the prevention and early diagnosis of ovarian cancer.
As we discussed above, ovarian cancer has been reported mainly associated with factors including environmental factors [144,145], genetic factors [13,146,147,148], demographic and lifestyle factors [96,149,150], reproductive factors [13,151,152], microbiome factors, and other factors [16,87]. (Figure 1) Though recent research revealed the separate effects of these risk factors, their cooperation in individuals is almost ignored, which may be a blockage of exploring the development of ovarian cancer. The risk factors exert different effects in specific patients with ovarian cancer. The Wheel of Causation and Web of Causation [153], models that focus on the etiology of chronic diseases, may help us understand the interaction of risk factors in ovarian cancer. The multi-causal models will help to further elucidate the effect of risk factors on a specific individual and the interactions of each factor, which are essential for individual prevention [154]. These will greatly help us make the strategy of ovarian cancer prevention.
Moreover, women with ovarian cancer are generally diagnosed at advanced stages (stage III/IV) [3], which is closely related to the poor prognosis. The main reason for the difficulties in early diagnosis should be the lack of effective screening methods. The present screening methods mainly include transvaginal ultrasonography (TVS) and serum tests for cancer antigen 125 (CA125) [155]. However, none of them shows satisfactory effects in decreasing the morbidity of ovarian cancer in asymptomatic, average-risk women [156]. Updating of ovarian cancer screening methods is in urgency. Microbiomes in the gut and vagina provide us with a potential way to screen for ovarian cancer. Combining gut and vaginal microbiomes and other associated factors, we may improve the diagnostic model of ovarian cancer, shortening the diagnostic delay and reducing mortality. To ensure the accuracy of the early diagnosis model, we need to make full use of modern molecular biology technology to screen and identify ovarian cancer biomarkers from the aspects of microbiology, metabolomics, genomics, transcriptomics, key signaling pathways, and tissue target cells. At the same time, combining with the traditional factors related to ovarian cancer (including symptoms, serum index, genetic index, etc.) is also essential to enhance the sensitivity and specificity of the early diagnosis prediction model.
6. Conclusions
The gut and vaginal microbiomes show extremely complex interactions in different kinds of diseases including ovarian cancer. Some important roles of microbiomes in the development and treatment of ovarian cancer have been demonstrated in several studies. The gut and vaginal microbiomes may affect the progression of ovarian cancer via the regulation of estrogen, modulation of inflammatory, and immune responses, and modulation of epigenetic modifications. There are great differences in gut and vaginal microbiomes between patients with ovarian cancer and those with benign ones, providing a possibility of using microbiomes for differential diagnosis. Moreover, incorporating gut and vaginal microbiomes into multi-aetiologic models will help to fully elucidate the mechanisms of ovarian cancer development. Combining multi-omics research for novel ovarian cancer-related markers with traditional ovarian cancer-associated factors is crucial for the prevention and early diagnosis of ovarian cancer, which reduces the morbidity and mortality rate of ovarian cancer.
Author Contributions
writing—original draft preparation, H.L. and Z.Z.; writing—review and editing, J.C and J.P.; visualization, H.Z. and Y.B.; supervision, H.Z.; All authors have read and agreed to the published version of the manuscript.”
Funding
This study was supported in part by the Sichuan International Science and Technology Innovation Cooperation/HongKong/Macao/Taiwan Science and Technology Innovation Cooperation Project (Grant No. 2021YFH0189), the Sichuan International Science Foundation Project (Grant No. 2022NSFSC1363), and the project for disciplines of excellence–Clinical Research Incubation Project, West China Hospital, Sichuan University (Grant No.2021HXFH065). Figures were created by BioRender.com.
Data Availability Statement
No new data were generated or analyzed in this review.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors report there are no competing interests to declare.
References
- BRAY F, LAVERSANNE M, SUNG H, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries [J]. CA Cancer J Clin 2024, 74, 229–63. [CrossRef]
- SIEGEL R L, MILLER K D, WAGLE N S, et al. Cancer statistics, 2023 [J]. CA Cancer J Clin 2023, 73, 17–48. [CrossRef]
- TORRE L A, TRABERT B, DESANTIS C E, et al. Ovarian cancer statistics, 2018 [J]. CA Cancer J Clin 2018, 68, 284–96. [CrossRef]
- HUANG J, CHAN W C, NGAI C H, et al. Worldwide Burden, Risk Factors, and Temporal Trends of Ovarian Cancer: A Global Study [J]. Cancers (Basel) 2022, 14. [CrossRef]
- RICHARDSON D L, ESKANDER R N, O'MALLEY D M. Advances in Ovarian Cancer Care and Unmet Treatment Needs for Patients With Platinum Resistance: A Narrative Review [J]. JAMA Oncol 2023, 9, 851–9. [CrossRef]
- CHIONH F, MITCHELL G, LINDEMAN G J, et al. The role of poly adenosine diphosphate ribose polymerase inhibitors in breast and ovarian cancer: current status and future directions [J]. Asia Pac J Clin Oncol 2011, 7, 197–211. [CrossRef]
- ITAMOCHI H, KIGAWA J. Clinical trials and future potential of targeted therapy for ovarian cancer [J]. Int J Clin Oncol 2012, 17, 430–40. [CrossRef]
- ZHOU P, WANG J, MISHAIL D, et al. Recent advancements in PARP inhibitors-based targeted cancer therapy [J]. Precis Clin Med 2020, 3, 187–201. [CrossRef]
- GONZáLEZ-MARTíN A, POTHURI B, VERGOTE I, et al. Niraparib in Patients with Newly Diagnosed Advanced Ovarian Cancer [J]. N Engl J Med 2019, 381, 2391–402. [CrossRef]
- KöBEL M, KANG E Y. The Evolution of Ovarian Carcinoma Subclassification [J]. Cancers (Basel) 2022, 14. [CrossRef]
- SCHOUTROP E, MOYANO-GALCERAN L, LHEUREUX S, et al. Molecular, cellular and systemic aspects of epithelial ovarian cancer and its tumor microenvironment [J]. Seminars In Cancer Biology 2022,86(Pt 3): 207-23. [CrossRef]
- MA R, TANG Z, WANG J. NLRP12 is a prognostic biomarker and correlated with immune infiltrates in epithelial ovarian cancer [J]. J Gene Med 2023, e3585. [CrossRef]
- WEBB P M, JORDAN S J. Epidemiology of epithelial ovarian cancer [J]. Best Pract Res Clin Obstet Gynaecol 2017, 41. [CrossRef]
- BIEGING K T, MELLO S S, ATTARDI L D. Unravelling mechanisms of p53-mediated tumour suppression [J]. Nat Rev Cancer 2014, 14, 359–70. [CrossRef]
- GENG S, ZHANG X, ZHU X, et al. Psychological factors increase the risk of ovarian cancer [J]. J Obstet Gynaecol 2023, 43, 2187573. [CrossRef]
- ALI A T, AL-ANI O, AL-ANI F. Epidemiology and risk factors for ovarian cancer [J]. Prz Menopauzalny 2023,22(2). [CrossRef]
- LA VECCHIA, C. Ovarian cancer: epidemiology and risk factors [J]. Eur J Cancer Prev 2017, 26, 55–62. [Google Scholar]
- WISHART D S, OLER E, PETERS H, et al. MiMeDB: the Human Microbial Metabolome Database [J]. Nucleic Acids Res 2023, 51, D611–D20. [CrossRef]
- BELKAID Y, HAND T W. Role of the microbiota in immunity and inflammation [J]. Cell 2014, 157, 121–41. [CrossRef]
- POSTLER T S, GHOSH S. Understanding the Holobiont: How Microbial Metabolites Affect Human Health and Shape the Immune System [J]. Cell Metab 2017, 26, 110–30. [CrossRef]
- CHEN Y, XIAO L, ZHOU M, et al. The microbiota: a crucial mediator in gut homeostasis and colonization resistance [J]. Front Microbiol 2024, 15, 1417864. [CrossRef]
- MA C, JIANG M, LI J, et al. Plasma Epstein-Barr Virus DNA load for diagnostic and prognostic assessment in intestinal Epstein-Barr Virus infection [J]. Front Cell Infect Microbiol 2024, 14, 1526633. [CrossRef]
- YOU L, ZHOU J, XIN Z, et al. Novel directions of precision oncology: circulating microbial DNA emerging in cancer-microbiome areas [J]. Precis Clin Med 2022, 5, pbac005. [CrossRef]
- RUTKOWSKI M R, STEPHEN T L, SVORONOS N, et al. Microbially driven TLR5-dependent signaling governs distal malignant progression through tumor-promoting inflammation [J]. Cancer Cell 2015, 27, 27–40. [CrossRef]
- NENé N R, REISEL D, LEIMBACH A, et al. Association between the cervicovaginal microbiome, BRCA1 mutation status, and risk of ovarian cancer: a case-control study [J]. Lancet Oncol 2019, 20, 1171–82. [CrossRef]
- JACOBSON D, MOORE K, GUNDERSON C, et al. Shifts in gut and vaginal microbiomes are associated with cancer recurrence time in women with ovarian cancer [J]. PeerJ 2021, 9, e11574. [CrossRef]
- MUNGENAST F, THALHAMMER T. Estrogen biosynthesis and action in ovarian cancer [J]. Front Endocrinol (Lausanne) 2014, 5, 192. [CrossRef]
- BORELLA F, CAROSSO A R, COSMA S, et al. Gut Microbiota and Gynecological Cancers: A Summary of Pathogenetic Mechanisms and Future Directions [J]. ACS Infect Dis 2021, 7, 987–1009. [CrossRef]
- MATEI D, NEPHEW K P. Epigenetic Attire in Ovarian Cancer: The Emperor's New Clothes [J]. Cancer Res 2020, 80, 3775–85. [CrossRef]
- BäCKHED F, LEY R E, SONNENBURG J L, et al. Host-bacterial mutualism in the human intestine [J]. Science 2005, 307, 1915–20.
- LOZUPONE C A, STOMBAUGH J I, GORDON J I, et al. Diversity, stability and resilience of the human gut microbiota [J]. Nature 2012, 489, 220–30. [CrossRef]
- RINNINELLA E, RAOUL P, CINTONI M, et al. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases [J]. Microorganisms 2019, 7. [CrossRef]
- YOON K, KIM N. Roles of Sex Hormones and Gender in the Gut Microbiota [J]. J Neurogastroenterol Motil 2021, 27, 314–25. [CrossRef] [PubMed]
- PUGH J N, LYDON K M, O'DONOVAN C M, et al. More than a gut feeling: What is the role of the gastrointestinal tract in female athlete health? [J]. Eur J Sport Sci 2022, 22, 755–64. [CrossRef] [PubMed]
- SIDDIQUI R, MAKHLOUF Z, ALHARBI A M, et al. The Gut Microbiome and Female Health [J]. Biology (Basel) 2022, 11. [CrossRef]
- KOLIADA A, MOSEIKO V, ROMANENKO M, et al. Sex differences in the phylum-level human gut microbiota composition [J]. BMC Microbiol 2021, 21, 131. [CrossRef]
- ZHANG D, XI Y, FENG Y. Ovarian cancer risk in relation to blood lipid levels and hyperlipidemia: a systematic review and meta-analysis of observational epidemiologic studies [J]. Eur J Cancer Prev 2021, 30, 161–70. [CrossRef]
- NEUMAN H, DEBELIUS J W, KNIGHT R, et al. Microbial endocrinology: the interplay between the microbiota and the endocrine system [J]. FEMS Microbiol Rev 2015, 39, 509–21. [CrossRef]
- SANTOS-MARCOS J A, RANGEL-ZUñIGA O A, JIMENEZ-LUCENA R, et al. Influence of gender and menopausal status on gut microbiota [J]. Maturitas 2018, 116, 43–53. [CrossRef]
- ANAHTAR M N, GOOTENBERG D B, MITCHELL C M, et al. Cervicovaginal Microbiota and Reproductive Health: The Virtue of Simplicity [J]. Cell Host Microbe 2018, 23, 159–68. [CrossRef]
- SMITH S B, RAVEL J. The vaginal microbiota, host defence and reproductive physiology [J]. J Physiol 2017, 595, 451–63. [CrossRef]
- GAJER P, BROTMAN R M, BAI G, et al. Temporal dynamics of the human vaginal microbiota [J]. Sci Transl Med 2012, 4, 132ra52. [CrossRef]
- RAVEL J, GAJER P, ABDO Z, et al. Vaginal microbiome of reproductive-age women [J]. Proc Natl Acad Sci U S A 2011, 108 (Suppl 1), 4680–7. [CrossRef] [PubMed]
- FRANCE M, ALIZADEH M, BROWN S, et al. Towards a deeper understanding of the vaginal microbiota [J]. Nat Microbiol 2022, 7, 367–78. [CrossRef] [PubMed]
- MANCABELLI L, TARRACCHINI C, MILANI C, et al. Vaginotypes of the human vaginal microbiome [J]. Environ Microbiol 2021, 23, 1780–92. [CrossRef]
- GREENBAUM S, GREENBAUM G, MORAN-GILAD J, et al. Ecological dynamics of the vaginal microbiome in relation to health and disease [J]. Am J Obstet Gynecol 2019, 220, 324–35. [CrossRef]
- CHAMPER M, WONG A M, CHAMPER J, et al. The role of the vaginal microbiome in gynaecological cancer [J]. BJOG 2018, 125, 309–15. [CrossRef]
- ASANGBA A E, CHEN J, GOERGEN K M, et al. Diagnostic and prognostic potential of the microbiome in ovarian cancer treatment response [J]. Sci Rep 2023, 13, 730. [CrossRef]
- EL BAIRI K, KANDHRO A H, GOURI A, et al. Emerging diagnostic, prognostic and therapeutic biomarkers for ovarian cancer [J]. Cell Oncol (Dordr) 2017, 40, 105–18. [CrossRef]
- HU X, XU X, ZENG X, et al. Gut microbiota dysbiosis promotes the development of epithelial ovarian cancer via regulating Hedgehog signaling pathway [J]. Gut Microbes 2023, 15, 2221093. [CrossRef]
- D'AMICO F, PERRONE A M, RAMPELLI S, et al. Gut Microbiota Dynamics during Chemotherapy in Epithelial Ovarian Cancer Patients Are Related to Therapeutic Outcome [J]. Cancers (Basel) 2021, 13. [CrossRef]
- CHAMBERS L M, ESAKOV RHOADES E L, BHARTI R, et al. Disruption of the Gut Microbiota Confers Cisplatin Resistance in Epithelial Ovarian Cancer [J]. Cancer Research 2022, 82, 4654–69. [CrossRef]
- ZHANG Y, ZHANG X-R, PARK J-L, et al. Genome-wide DNA methylation profiles altered by Helicobacter pylori in gastric mucosa and blood leukocyte DNA [J]. Oncotarget 2016, 7, 37132–44. [CrossRef]
- TRABERT B, WATERBOER T, IDAHL A, et al. Antibodies Against Chlamydia trachomatis and Ovarian Cancer Risk in Two Independent Populations [J]. J Natl Cancer Inst 2019, 111, 129–36. [CrossRef]
- SHARMA H, TAL R, CLARK N A, et al. Microbiota and pelvic inflammatory disease [J]. Semin Reprod Med 2014, 32, 43–9. [CrossRef]
- ANTONIO M A D, RABE L K, HILLIER S L. Colonization of the rectum by Lactobacillus species and decreased risk of bacterial vaginosis [J]. J Infect Dis 2005, 192, 394–8.
- EL AILA N A, TENCY I, CLAEYS G, et al. Identification and genotyping of bacteria from paired vaginal and rectal samples from pregnant women indicates similarity between vaginal and rectal microflora [J]. BMC Infect Dis 2009, 9, 167. [CrossRef]
- SHIN H, MARTINEZ K A, HENDERSON N, et al. Partial convergence of the human vaginal and rectal maternal microbiota in late gestation and early post-partum [J]. NPJ Biofilms Microbiomes 2023, 9, 37. [CrossRef]
- FUDABA M, KAMIYA T, TACHIBANA D, et al. Bioinformatics Analysis of Oral, Vaginal, and Rectal Microbial Profiles during Pregnancy: A Pilot Study on the Bacterial Co-Residence in Pregnant Women [J]. Microorganisms 2021, 9. [CrossRef]
- DI PIERRO F, CRISCUOLO A A, DEI GIUDICI A, et al. Oral administration of Lactobacillus crispatus M247 to papillomavirus-infected women: results of a preliminary, uncontrolled, open trial [J]. Minerva Obstet Gynecol 2021, 73, 621–31. [CrossRef]
- JANG S-E, JEONG J-J, CHOI S-Y, et al. Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus La-14 Attenuate Gardnerella vaginalis-Infected Bacterial Vaginosis in Mice [J]. Nutrients 2017, 9. [CrossRef]
- LIU Y, LI H-T, ZHOU S-J, et al. Effects of vaginal seeding on gut microbiota, body mass index, and allergy risks in infants born through cesarean delivery: a randomized clinical trial [J]. Am J Obstet Gynecol MFM 2023, 5, 100793. [CrossRef]
- YUAN C, GASKINS A J, BLAINE A I, et al. Association Between Cesarean Birth and Risk of Obesity in Offspring in Childhood, Adolescence, and Early Adulthood [J]. JAMA Pediatr 2016, 170, e162385. [CrossRef]
- KRISTENSEN K, HENRIKSEN L. Cesarean section and disease associated with immune function [J]. J Allergy Clin Immunol 2016, 137, 587–90. [CrossRef] [PubMed]
- PLOTTEL C S, BLASER M J. Microbiome and malignancy [J]. Cell Host Microbe 2011, 10, 324–35. [CrossRef]
- KWA M, PLOTTEL C S, BLASER M J, et al. The Intestinal Microbiome and Estrogen Receptor-Positive Female Breast Cancer [J]. J Natl Cancer Inst 2016, 108. [CrossRef]
- ERVIN S M, LI H, LIM L, et al. Gut microbial β-glucuronidases reactivate estrogens as components of the estrobolome that reactivate estrogens [J]. J Biol Chem 2019, 294, 18586–99. [CrossRef]
- BUCHTA, V. Vaginal microbiome [J]. Ceska Gynekol 2018, 83, 371–9. [Google Scholar]
- CLAES I J, VARGAS GARCíA C E, LEBEER S. Novel opportunities for the exploitation of host-microbiome interactions in the intestine [J]. Curr Opin Biotechnol 2015, 32, 28–34. [CrossRef]
- LEBEER S, VANDERLEYDEN J, DE KEERSMAECKER S C J. Host interactions of probiotic bacterial surface molecules: comparison with commensals and pathogens [J]. Nat Rev Microbiol 2010, 8, 171–84. [CrossRef]
- ILHAN Z E, ŁANIEWSKI P, THOMAS N, et al. Deciphering the complex interplay between microbiota, HPV, inflammation and cancer through cervicovaginal metabolic profiling [J]. EBioMedicine 2019, 44, 675–90. [CrossRef]
- SCOTT A J, ALEXANDER J L, MERRIFIELD C A, et al. International Cancer Microbiome Consortium consensus statement on the role of the human microbiome in carcinogenesis [J]. Gut 2019, 68, 1624–32. [CrossRef]
- RAFTOGIANIS R, CREVELING C, WEINSHILBOUM R, et al. Estrogen metabolism by conjugation [J]. J Natl Cancer Inst Monogr 2000, 113–24.
- RUGGIERO R J, LIKIS F E. Estrogen: physiology, pharmacology, and formulations for replacement therapy [J]. J Midwifery Womens Health 2002, 47, 130–8. [CrossRef]
- CUI J, SHEN Y, LI R. Estrogen synthesis and signaling pathways during aging: from periphery to brain [J]. Trends Mol Med 2013, 19, 197–209. [CrossRef]
- PROSSNITZ E R, BARTON M. The G-protein-coupled estrogen receptor GPER in health and disease [J]. Nat Rev Endocrinol 2011, 7, 715–26. [CrossRef]
- SEYED HAMEED A S, RAWAT P S, MENG X, et al. Biotransformation of dietary phytoestrogens by gut microbes: A review on bidirectional interaction between phytoestrogen metabolism and gut microbiota [J]. Biotechnol Adv 2020, 43, 107576. [CrossRef]
- TKALIA I G, VOROBYOVA L I, SVINTSITSKY V S, et al. Clinical significance of hormonal receptor status of malignant ovarian tumors [J]. Exp Oncol 2014, 36, 125–33.
- CHOI J-H, WONG A S T, HUANG H-F, et al. Gonadotropins and ovarian cancer [J]. Endocr Rev 2007, 28, 440–61.
- BAKER J M, AL-NAKKASH L, HERBST-KRALOVETZ M M. Estrogen-gut microbiome axis: Physiological and clinical implications [J]. Maturitas 2017, 103, 45–53. [CrossRef]
- POLLET R M, D'AGOSTINO E H, WALTON W G, et al. An Atlas of β-Glucuronidases in the Human Intestinal Microbiome [J]. Structure 2017, 25. [CrossRef]
- TETEL M J, DE VRIES G J, MELCANGI R C, et al. Steroids, stress and the gut microbiome-brain axis [J]. J Neuroendocrinol 2018, 30. [CrossRef]
- MAENG L Y, BEUMER A. Never fear, the gut bacteria are here: Estrogen and gut microbiome-brain axis interactions in fear extinction [J]. Int J Psychophysiol 2023, 189, 66–75. [CrossRef]
- DINAN T G, SCOTT L V. Anatomy of melancholia: focus on hypothalamic-pituitary-adrenal axis overactivity and the role of vasopressin [J]. J Anat 2005, 207, 259–64.
- OYOLA M G, HANDA R J. Hypothalamic-pituitary-adrenal and hypothalamic-pituitary-gonadal axes: sex differences in regulation of stress responsivity [J]. Stress 2017, 20, 476–94. [CrossRef]
- TRABERT B, COBURN S B, FALK R T, et al. Circulating estrogens and postmenopausal ovarian and endometrial cancer risk among current hormone users in the Women's Health Initiative Observational Study [J]. Cancer Causes Control 2019, 30, 1201–11. [CrossRef]
- SHI L-F, WU Y, LI C-Y. Hormone therapy and risk of ovarian cancer in postmenopausal women: a systematic review and meta-analysis [J]. Menopause 2016, 23, 417–24. [CrossRef]
- CUNAT S, HOFFMANN P, PUJOL P. Estrogens and epithelial ovarian cancer [J]. Gynecologic Oncology 2004, 94, 25–32.
- LIU A, ZHANG D, YANG X, et al. Estrogen receptor alpha activates MAPK signaling pathway to promote the development of endometrial cancer [J]. J Cell Biochem 2019, 120, 17593–601. [CrossRef]
- GOAD J, KO Y-A, KUMAR M, et al. Oestrogen fuels the growth of endometrial hyperplastic lesions initiated by overactive Wnt/β-catenin signalling [J]. Carcinogenesis 2018, 39, 1105–16. [CrossRef]
- LIU X, ZHAN T, GAO Y, et al. Benzophenone-1 induced aberrant proliferation and metastasis of ovarian cancer cells via activated ERα and Wnt/β-catenin signaling pathways [J]. Environ Pollut 2022, 292, 118370. [CrossRef]
- SIMPKINS F, JANG K, YOON H, et al. Dual Src and MEK Inhibition Decreases Ovarian Cancer Growth and Targets Tumor Initiating Stem-Like Cells [J]. Clin Cancer Res 2018, 24, 4874–86. [CrossRef]
- PETRIE W K, DENNIS M K, HU C, et al. G protein-coupled estrogen receptor-selective ligands modulate endometrial tumor growth [J]. Obstet Gynecol Int 2013, 2013, 472720. [CrossRef]
- DE VISSER K E, JOYCE J A. The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth [J]. Cancer Cell 2023, 41, 374–403. [CrossRef] [PubMed]
- ZHAO H, WU L, YAN G, et al. Inflammation and tumor progression: signaling pathways and targeted intervention [J]. Signal Transduct Target Ther 2021, 6, 263. [CrossRef] [PubMed]
- VAN TUIJL L A, VOOGD A C, DE GRAEFF A, et al. Psychosocial factors and cancer incidence (PSY-CA): Protocol for individual participant data meta-analyses [J]. Brain Behav 2021, 11, e2340. [CrossRef]
- ARDEN-CLOSE E, GIDRON Y, MOSS-MORRIS R. Psychological distress and its correlates in ovarian cancer: a systematic review [J]. Psychooncology 2008, 17, 1061–72. [CrossRef]
- HOTAMISLIGIL G, S. Inflammation and metabolic disorders [J]. Nature 2006, 444, 860–7. [Google Scholar] [CrossRef]
- HULDANI H, ABDUL-JABBAR ALI S, AL-DOLAIMY F, et al. The potential role of interleukins and interferons in ovarian cancer [J]. Cytokine 2023, 171, 156379. [CrossRef]
- GAO J, XU K, LIU H, et al. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism [J]. Front Cell Infect Microbiol 2018, 8, 13. [CrossRef]
- FACHI J L, SéCCA C, RODRIGUES P B, et al. Acetate coordinates neutrophil and ILC3 responses against C. difficile through FFAR2 [J]. J Exp Med 2020, 217. [CrossRef]
- WOODS D C, WHITE Y A, DAU C, et al. TLR4 activates NF-κB in human ovarian granulosa tumor cells [J]. Biochem Biophys Res Commun 2011, 409, 675–80. [CrossRef]
- KELLY M G, ALVERO A B, CHEN R, et al. TLR-4 signaling promotes tumor growth and paclitaxel chemoresistance in ovarian cancer [J]. Cancer Research 2006, 66, 3859–68.
- LUPI L A, CUCIELO M S, SILVEIRA H S, et al. The role of Toll-like receptor 4 signaling pathway in ovarian, cervical, and endometrial cancers [J]. Life Sci 2020, 247, 117435. [CrossRef]
- KASPERCZYK H, BAUMANN B, DEBATIN K-M, et al. Characterization of sonic hedgehog as a novel NF-kappaB target gene that promotes NF-kappaB-mediated apoptosis resistance and tumor growth in vivo [J]. FASEB J 2009, 23, 21–33. [CrossRef]
- WANG Y, JIN G, LI Q, et al. Hedgehog Signaling Non-Canonical Activated by Pro-Inflammatory Cytokines in Pancreatic Ductal Adenocarcinoma [J]. Journal of Cancer 2016, 7, 2067–76. [CrossRef]
- VLAD C, DINA C, KUBELAC P, et al. Expression of toll-like receptors in ovarian cancer [J]. J BUON 2018, 23, 1725–31.
- XU S, LIU Z, LV M, et al. Intestinal dysbiosis promotes epithelial-mesenchymal transition by activating tumor-associated macrophages in ovarian cancer [J]. Pathogens and Disease 2019, 77. [CrossRef]
- BROWNING L, PATEL M R, HORVATH E B, et al. IL-6 and ovarian cancer: inflammatory cytokines in promotion of metastasis [J]. Cancer Manag Res 2018, 10, 6685–93. [CrossRef]
- SIPOS A, UJLAKI G, MIKó E, et al. The role of the microbiome in ovarian cancer: mechanistic insights into oncobiosis and to bacterial metabolite signaling [J]. Mol Med 2021, 27, 33. [CrossRef]
- BEGLEY M, GAHAN C G M, HILL C. The interaction between bacteria and bile [J]. FEMS Microbiol Rev 2005, 29, 625–51.
- HOROWITZ N S, HUA J, POWELL M A, et al. Novel cytotoxic agents from an unexpected source: bile acids and ovarian tumor apoptosis [J]. Gynecologic Oncology 2007, 107, 344–9.
- SUN M, WU W, LIU Z, et al. Microbiota metabolite short chain fatty acids, GPCR, and inflammatory bowel diseases [J]. J Gastroenterol 2017, 52, 1–8. [CrossRef] [PubMed]
- ZHAO L-Y, MEI J-X, YU G, et al. Role of the gut microbiota in anticancer therapy: from molecular mechanisms to clinical applications [J]. Signal Transduct Target Ther 2023, 8, 201. [CrossRef]
- WIKOFF W R, ANFORA A T, LIU J, et al. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites [J]. Proc Natl Acad Sci U S A 2009, 106, 3698–703. [CrossRef] [PubMed]
- KE C, HOU Y, ZHANG H, et al. Large-scale profiling of metabolic dysregulation in ovarian cancer [J]. Int J Cancer 2015, 136, 516–26. [CrossRef] [PubMed]
- BOSSUET-GREIF N, VIGNARD J, TAIEB F, et al. The Colibactin Genotoxin Generates DNA Interstrand Cross-Links in Infected Cells [J]. mBio 2018, 9. [CrossRef]
- CHáVEZ-TALAVERA O, TAILLEUX A, LEFEBVRE P, et al. Bile Acid Control of Metabolism and Inflammation in Obesity, Type 2 Diabetes, Dyslipidemia, and Nonalcoholic Fatty Liver Disease [J]. Gastroenterology 2017, 152. [CrossRef]
- POLS T W H, NOMURA M, HARACH T, et al. TGR5 activation inhibits atherosclerosis by reducing macrophage inflammation and lipid loading [J]. Cell Metab 2011, 14, 747–57. [CrossRef]
- GOMES A C, HOFFMANN C, MOTA J F. The human gut microbiota: Metabolism and perspective in obesity [J]. Gut Microbes 2018, 9, 308–25. [CrossRef]
- OUCHI N, WALSH K. Adiponectin as an anti-inflammatory factor [J]. Clin Chim Acta 2007, 380, 24–30. [CrossRef]
- CANFORA E E, JOCKEN J W, BLAAK E E. Short-chain fatty acids in control of body weight and insulin sensitivity [J]. Nat Rev Endocrinol 2015, 11, 577–91. [CrossRef]
- YOU H, TAN Y, YU D, et al. The Therapeutic Effect of SCFA-Mediated Regulation of the Intestinal Environment on Obesity [J]. Front Nutr 2022, 9, 886902. [CrossRef]
- SHANMUGHAPRIYA S, SENTHILKUMAR G, VINODHINI K, et al. Viral and bacterial aetiologies of epithelial ovarian cancer [J]. Eur J Clin Microbiol Infect Dis 2012, 31, 2311–7. [CrossRef]
- PATHAK S, WILCZYŃSKI J R, PARADOWSKA E. Factors in Oncogenesis: Viral Infections in Ovarian Cancer [J]. Cancers (Basel) 2020, 12. [CrossRef]
- RAHBAR SAADAT Y, POURSEIF M M, ZUNUNI VAHED S, et al. Modulatory Role of Vaginal-Isolated Lactococcus lactis on the Expression of miR-21, miR-200b, and TLR-4 in CAOV-4 Cells and In Silico Revalidation [J]. Probiotics Antimicrob Proteins 2020, 12, 1083–96. [CrossRef]
- XIE X, YANG M, DING Y, et al. Microbial infection, inflammation and epithelial ovarian cancer [J]. Oncology Letters 2017, 14, 1911–9. [CrossRef]
- MOUFARRIJ S, DANDAPANI M, ARTHOFER E, et al. Epigenetic therapy for ovarian cancer: promise and progress [J]. Clin Epigenetics 2019, 11, 7. [CrossRef]
- SCHöNDORF T, EBERT M P, HOFFMANN J, et al. Hypermethylation of the PTEN gene in ovarian cancer cell lines [J]. Cancer Lett 2004, 207, 215–20.
- YANG Q, YANG Y, ZHOU N, et al. Epigenetics in ovarian cancer: premise, properties, and perspectives [J]. Mol Cancer 2018, 17, 109. [CrossRef]
- CAI M, HU Z, LIU J, et al. Expression of hMOF in different ovarian tissues and its effects on ovarian cancer prognosis [J]. Oncol Rep 2015, 33, 685–92. [CrossRef]
- SANTOS J C, RIBEIRO M L. Epigenetic regulation of DNA repair machinery in Helicobacter pylori-induced gastric carcinogenesis [J]. World J Gastroenterol 2015, 21, 9021–37. [CrossRef]
- ZHANG P-P, ZHOU L, CAO J-S, et al. Possible Epithelial Ovarian Cancer Association with HPV18 or HPV33 Infection [J]. Asian Pac J Cancer Prev 2016, 17, 2959–64.
- PAPASTERGIOU V, KARATAPANIS S, GEORGOPOULOS S D. Helicobacter pylori and colorectal neoplasia: Is there a causal link? [J]. World J Gastroenterol 2016, 22, 649–58. [CrossRef]
- TAHARA T, HIRATA I, NAKANO N, et al. Potential link between Fusobacterium enrichment and DNA methylation accumulation in the inflammatory colonic mucosa in ulcerative colitis [J]. Oncotarget 2017, 8, 61917–26. [CrossRef]
- JEFFERY I B, O'TOOLE P W. Diet-microbiota interactions and their implications for healthy living [J]. Nutrients 2013, 5, 234–52. [CrossRef]
- HAQUE S, RAINA R, AFROZE N, et al. Microbial dysbiosis and epigenetics modulation in cancer development - A chemopreventive approach [J]. Seminars In Cancer Biology 2022, 86 (Pt 3), 666–81. [CrossRef]
- WOO V, ALENGHAT T. Epigenetic regulation by gut microbiota [J]. Gut Microbes 2022, 14, 2022407. [CrossRef]
- RAJIĆ J, INIC-KANADA A, STEIN E, et al. Chlamydia trachomatis Infection Is Associated with E-Cadherin Promoter Methylation, Downregulation of E-Cadherin Expression, and Increased Expression of Fibronectin and α-SMA-Implications for Epithelial-Mesenchymal Transition [J]. Front Cell Infect Microbiol 2017, 7, 253. [CrossRef]
- KUMAR H, LUND R, LAIHO A, et al. Gut microbiota as an epigenetic regulator: pilot study based on whole-genome methylation analysis [J]. mBio 2014, 5. [CrossRef]
- LI L, CHENG R, WU Y, et al. Diagnosis and management of inflammatory bowel disease [J]. J Evid Based Med 2024, 17, 409–33. [CrossRef]
- ZENG Z, JIANG M, LI X, et al. Precision medicine in inflammatory bowel disease [J]. Precis Clin Med 2023, 6, pbad033. [CrossRef]
- MA C, CHEN K, LI L, et al. Epstein-Barr virus Infection Exacerbates Ulcerative Colitis by Driving Macrophage Pyroptosis via the Upregulation of Glycolysis [J]. Precis Clin Med 2025. [CrossRef]
- MADDAMS J, PARKIN D M, DARBY S C. The cancer burden in the United Kingdom in 2007 due to radiotherapy [J]. Int J Cancer 2011, 129, 2885–93. [CrossRef]
- ZAMWAR U M, ANJANKAR A P. Aetiology, Epidemiology, Histopathology, Classification, Detailed Evaluation, and Treatment of Ovarian Cancer [J]. Cureus 2022, 14, e30561. [CrossRef]
- STRATTON J F, PHAROAH P, SMITH S K, et al. A systematic review and meta-analysis of family history and risk of ovarian cancer [J]. Br J Obstet Gynaecol 1998, 105, 493–9.
- YANG-HARTWICH Y, SOTERAS M G, LIN Z P, et al. p53 protein aggregation promotes platinum resistance in ovarian cancer [J]. Oncogene 2015, 34, 3605–16. [CrossRef]
- BEWTRA C, WATSON P, CONWAY T, et al. Hereditary ovarian cancer: a clinicopathological study [J]. Int J Gynecol Pathol 1992, 11, 180–7.
- BANDERA E V, LEE V S, RODRIGUEZ-RODRIGUEZ L, et al. Racial/Ethnic Disparities in Ovarian Cancer Treatment and Survival [J]. Clin Cancer Res 2016, 22, 5909–14.
- FABER M T, KJæR S K, DEHLENDORFF C, et al. Cigarette smoking and risk of ovarian cancer: a pooled analysis of 21 case-control studies [J]. Cancer Causes Control 2013,24(5). [CrossRef]
- GAITSKELL K, GREEN J, PIRIE K, et al. Histological subtypes of ovarian cancer associated with parity and breastfeeding in the prospective Million Women Study [J]. Int J Cancer 2018, 142, 281–9. [CrossRef]
- GREISER C M, GREISER E M, DöREN M. Menopausal hormone therapy and risk of ovarian cancer: systematic review and meta-analysis [J]. Hum Reprod Update 2007, 13, 453–63.
- ROTHMAN K, GREENLAND S, LASH T. Modern Epidemiology, 3rd Edition [J]. Lippincott Williams & Wilkins 2008,P2272.
- JU L, SUO Z, LIN J, et al. Fecal microbiota and metabolites in the pathogenesis and precision medicine for inflammatory bowel disease [J]. Precis Clin Med 2024, 7, pbae023. [CrossRef]
- ANDRYKOWSKI M, A. Psychological and Behavioral Impact of Participation in Ovarian Cancer Screening [J]. Diagnostics (Basel, Switzerland) 2017, 7. [Google Scholar] [CrossRef]
- MENON U, GRIFFIN M, GENTRY-MAHARAJ A. Ovarian cancer screening--current status, future directions [J]. Gynecologic Oncology 2014, 132, 490–5. [CrossRef]
Figure 1.
Associated factors of ovarian cancer Associated factors of ovarian cancer have been reported mainly including environmental factors, genetic factors, demographic and lifestyle factors, reproductive factors, microbiome factors, and other factors. These factors together change the risk of developing ovarian cancer.
Figure 1.
Associated factors of ovarian cancer Associated factors of ovarian cancer have been reported mainly including environmental factors, genetic factors, demographic and lifestyle factors, reproductive factors, microbiome factors, and other factors. These factors together change the risk of developing ovarian cancer.
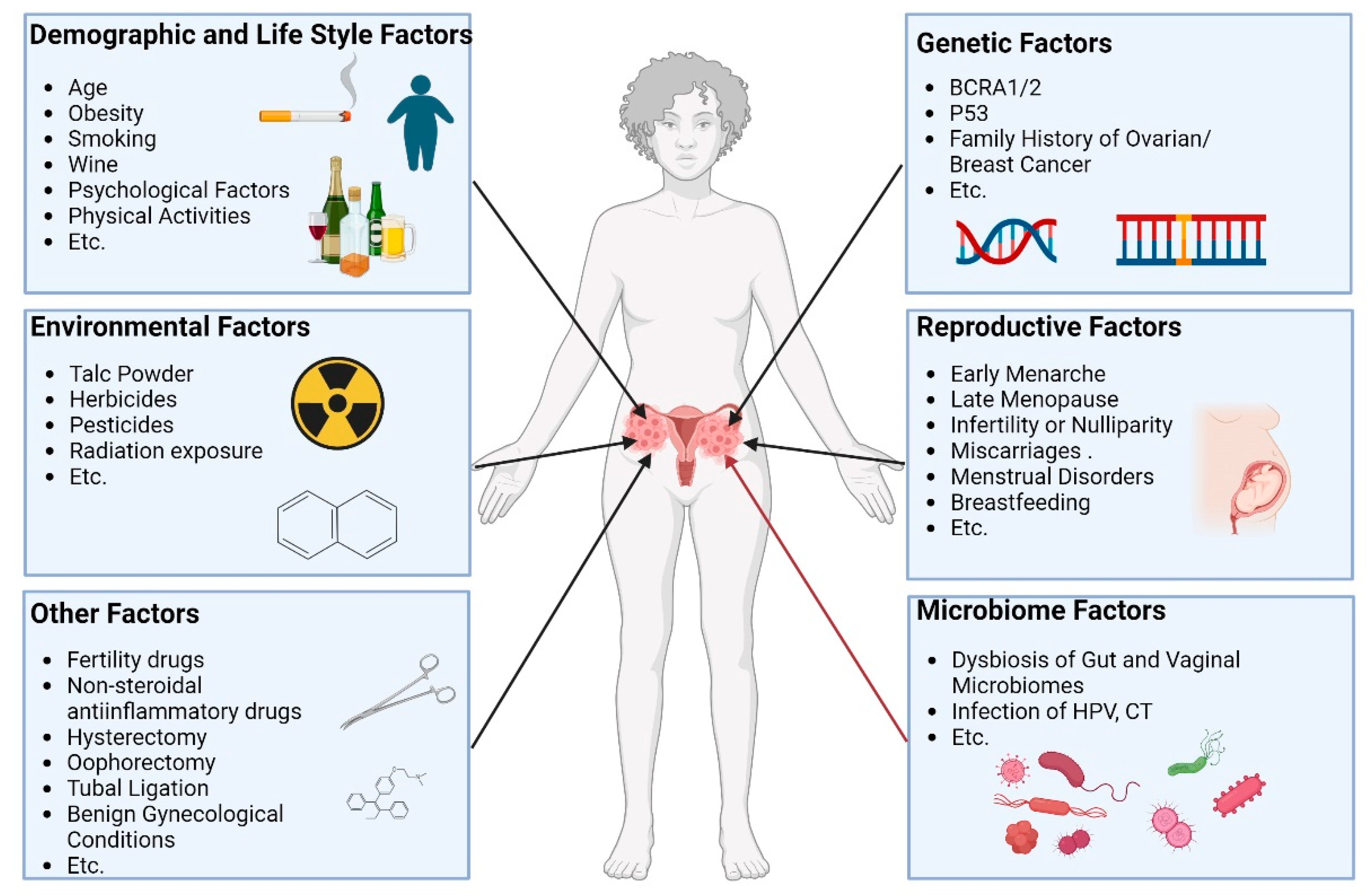
Figure 2.
Gut and vaginal microbiomes and estrogen in ovarian cancer Gut and vaginal microbiomes interact with each other via gut-vaginal axis. Gut and vaginal microbiomes change in patients with ovarian cancer. Gut microbiome changes circulatory estrogen levels by estrobolome and gut-brain axis. Estrobolome increases the levels of circulation estrogen by β-glucuronides. Gut-brain axis leads to stress and then activates hypothalamic-pituitary-adrenal (HPA) axis, regulating the generating of estrogen. Excessive estrogen combines with the ER-α and G-protein-coupled estrogen receptor (GPER) then activates signaling pathways including ERK, PI3K, etc. and oncogenesis genes. Meanwhile, the metabolites of estrogen via CYPs elevate the level of free radicals causing DNA damage, promoting tumor genesis.
Figure 2.
Gut and vaginal microbiomes and estrogen in ovarian cancer Gut and vaginal microbiomes interact with each other via gut-vaginal axis. Gut and vaginal microbiomes change in patients with ovarian cancer. Gut microbiome changes circulatory estrogen levels by estrobolome and gut-brain axis. Estrobolome increases the levels of circulation estrogen by β-glucuronides. Gut-brain axis leads to stress and then activates hypothalamic-pituitary-adrenal (HPA) axis, regulating the generating of estrogen. Excessive estrogen combines with the ER-α and G-protein-coupled estrogen receptor (GPER) then activates signaling pathways including ERK, PI3K, etc. and oncogenesis genes. Meanwhile, the metabolites of estrogen via CYPs elevate the level of free radicals causing DNA damage, promoting tumor genesis.
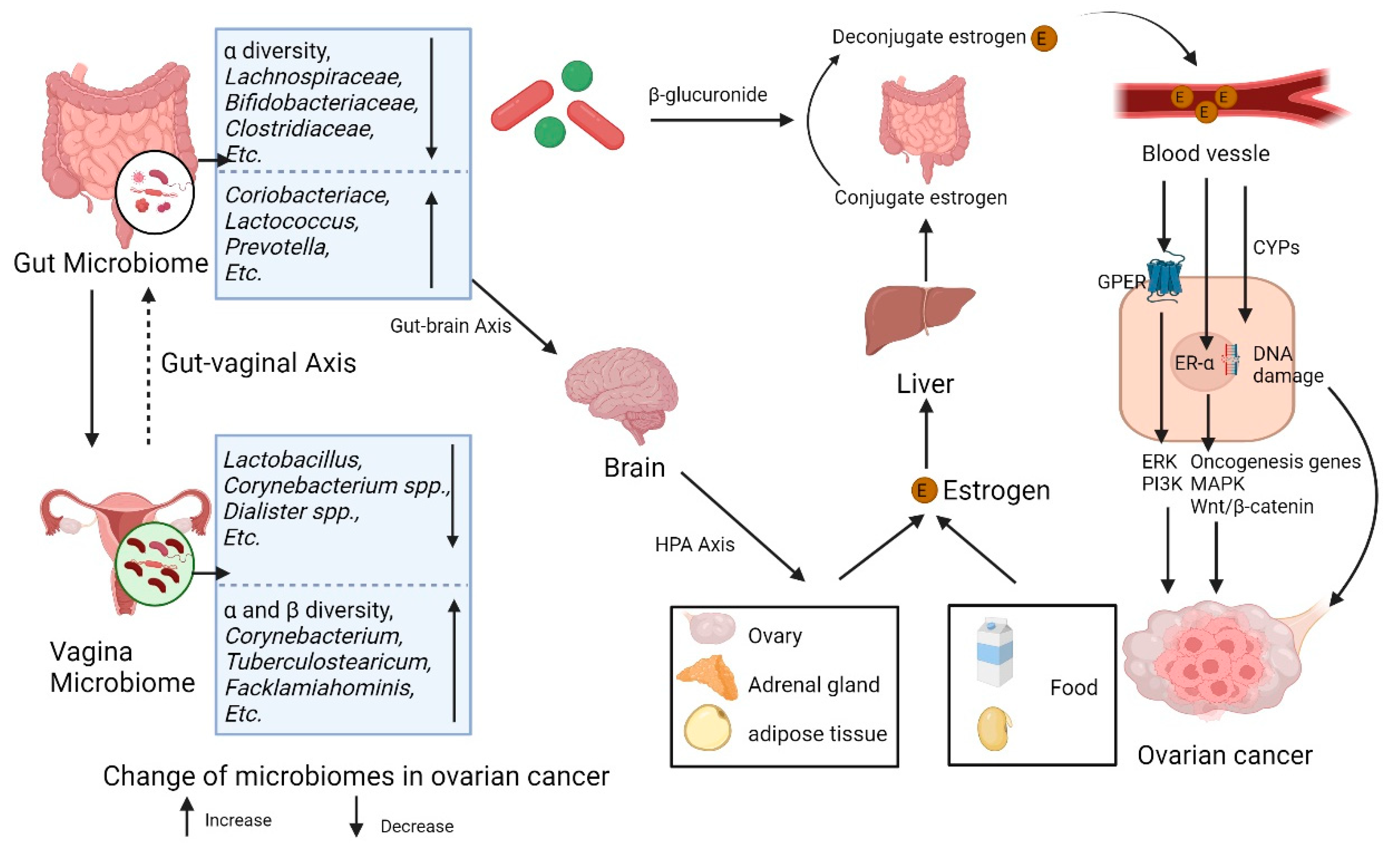
Figure 3.
Microbiomes and inflammation in ovarian cancer Gut and vaginal microbiomes interact with each other via gut-vaginal axis. Gut and vaginal microbiomes change in patients with ovarian cancer. The gut and vaginal microbiomes are associated with inflammation mainly through MAMPs and metabolites. The MAMPs interact with TLR-4 and TLR-5, activating the downstream signaling and leading to tumor development by promoting inflammation. The metabolites of microbes can inhibit the inflammatory reaction, while some of metabolisms (Colibactin) cause DNA damage and promoting tumor genesis.
Figure 3.
Microbiomes and inflammation in ovarian cancer Gut and vaginal microbiomes interact with each other via gut-vaginal axis. Gut and vaginal microbiomes change in patients with ovarian cancer. The gut and vaginal microbiomes are associated with inflammation mainly through MAMPs and metabolites. The MAMPs interact with TLR-4 and TLR-5, activating the downstream signaling and leading to tumor development by promoting inflammation. The metabolites of microbes can inhibit the inflammatory reaction, while some of metabolisms (Colibactin) cause DNA damage and promoting tumor genesis.
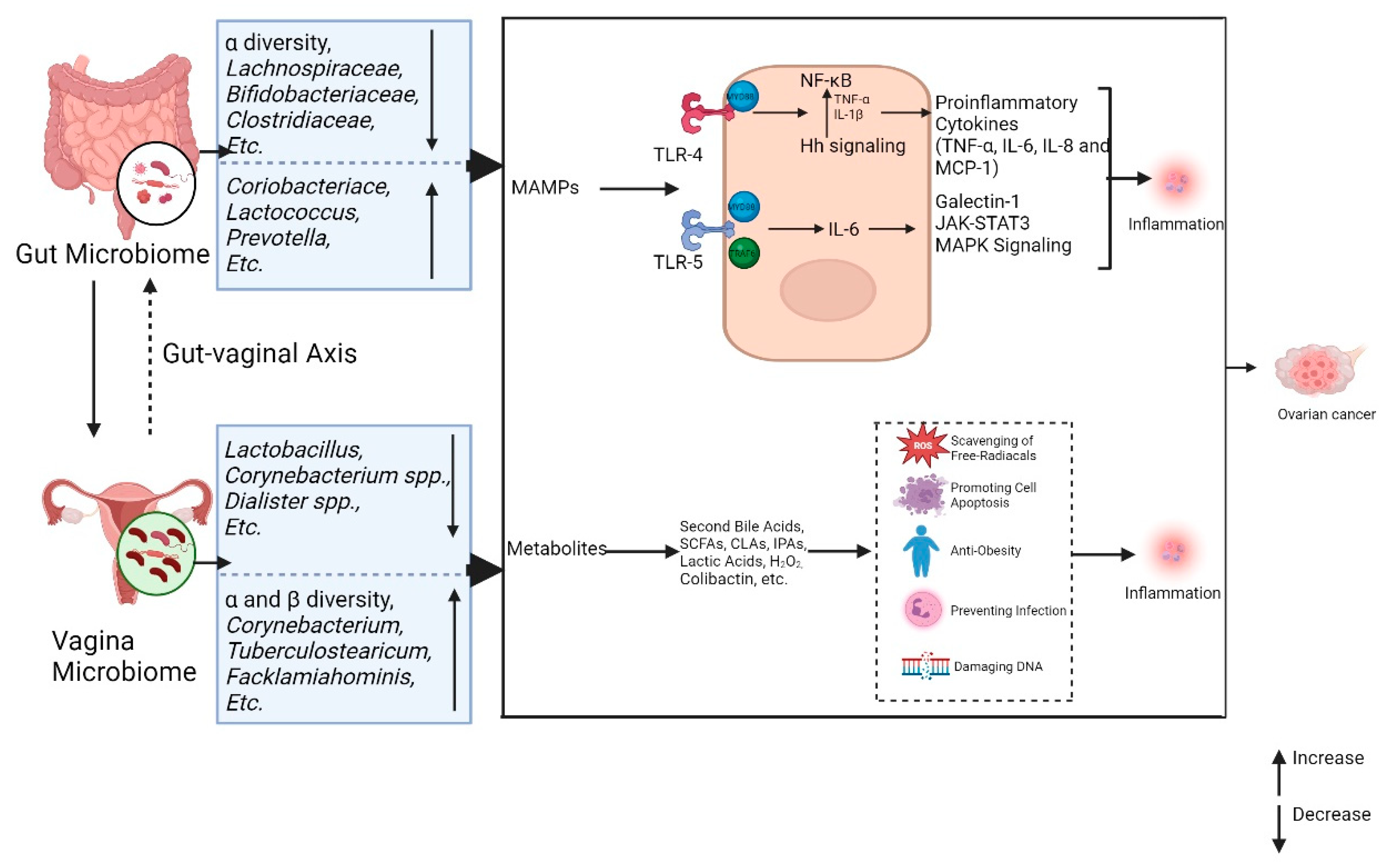

Table 1.
Characteristics of the gut and vaginal microbiomes in patients with ovarian cancer.
| Study | Study design | Participants | Associated outcomes |
| D'Amico 2021[47] | Cohort study |
|
Gut microbiota:
|
| Hu 2023[46] | Case-control study |
|
Gut microbiota:
|
| Asangba 2023[44] | Cohort study |
|
Vaginal microbiota:
|
| Nené 2019[21] | Case-control study |
|
Vaginal microbiota:
|
| Jacobson 2021[22] | Cohort Study |
|
Gut microbiota:
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



