Submitted:
20 February 2025
Posted:
21 February 2025
You are already at the latest version
Abstract
The idea that race is a purely social construct appears to have achieved almost universal acceptance within academia. However, this consensus is artificial and maintained by a taboo on the subject of race, particularly in the biological sciences. Here, I argue that the common-sense view that race has a biological basis but is unimportant for traits such as intelligence or character best fits the scientific data. I start by showing that most past biological discussions of race have focused on the wrong concepts. Specifically, population structure at the genomic level is irrelevant to most notions of race, which stress functional differences in a handful of traits. I therefore argue that the concept of ecotype, which is associated with local adaptation of a handful of traits in a highly connected network of populations, is the best descriptor of human races. I then explain the relationship between the social construction of race and its biological basis. The essential point here is that race is socially constructed from real biological variation. I conclude with a brief discussion of scientific racism, which I argue is better labeled pseudoscientific racism. I posit that stifling discussion of race by biologists has been counterproductive and misguided. When scientists refuse to refute the racist spin doctoring of biological data that is central to scientific racism, it gives the public the notion that the data are not on their side. Moreover, ideologically driven positions, such as races do not exist, erode public confidence in the scientific community.
Keywords:
race
; genetics
; scientific racism
; evolution
; local adaptation
; ecotype
1. Introduction
Science alleging that some races are superior to others has a long history. In some sense, it goes back to Darwin and his cousin Galton, who along with Spencer started the field of Social Darwinism1 (Darwin 1859, 1872; Spencer 1866; Galton 1891; Rogers 1972; Claeys 2000). Indeed, until the horrors of the Nazis in World War 2, eugenics and Social Darwinism were mainstream evolutionary biology (Galton 1904; Huxley 1936). Ideas we find repugnant today, such as sterilizing those who are supposedly “less fit,” were taught in textbooks (reviewed in Tontonoz 2008). This, in fact, more than the theological questions raised by evolution, was why William Jennings Bryan, for example, was so incensed at the teaching of evolution in public schools. As a Christian, he did not want children taught that some people are fundamentally better than others2. This fact is not widely known, as contemporary discussion of the Scopes Monkey Trial tends to leave this out, whitewashing this period in the history of evolutionary biology.
After World War II, racist pseudoscience died down, for obvious reasons, until the 1960s, when Jensen resurrected it in a paper titled “How Much Can We Boost IQ and Scholastic Achievement?” (Jensen 1969). Here he basically argued that black Americans perform poorly in school because they are endowed with genes that ensure low IQ. His work was widely condemned but also celebrated by some such as the Nobel prize winning physicist Shockley. It is not so well known that Watson and Crick, the most famous biologists of the day, also supported these ideas. We found out Watson’s views years later, for which he was disgraced in the popular press (Shapiro 2019). Crick made clear his views at the time, however, albeit under the radar3. The prominent evolutionary biologist and author E. O. Wilson was also a supporter of this work.
Since the time of Jensen, the notion that black people are genetically inferior for traits like intelligence has never gone away, although it has waxed and waned in popularity (Lynn et al 2002; Rushton and Jensen 2005; Haier 2017). The publication of The Bell Curve was a high point for what is sometimes called “scientific racism”4. This work continued the tradition of arguing that programs like “Head Start” are a waste of time since minorities, chiefly black Americans, have low IQ for genetic reasons. The authors made this argument with a host of correlational analyses exploring various measures of IQ and economic outcome. This work was refuted, at the time, by numerous scholars (Heckman 1995; Sternberg 1995; Hauser 1995), and was not taken seriously in the academy, but it remained a darling of the far right, and some mainstream journalists. The chief rebuttal to this, and other pseudoscientific racist scholarship, incidentally, was by Stephen Jay Gould (1996) in his book, The Mismeasure of Man. Here he said, correctly I think, that scientific racism undergoes a revival, with new terminology but the same old ideas, every time there is a major shift to the political right. He said we must be vigilant and stamp it out each time with facts. This was an important statement that unfortunately has not been taken seriously by scholars, who have subsequently chosen to ignore, rather than combat far right ideology in science.
The last decade or so has seen a resurgence in the popularity of The Bell Curve, and race science in general. This started with some farfetched arguments for why Jewish persons are supposedly smarter than everybody else (Cochran et al 2006). These ideas were briefly championed by Steven Pinker (2006), who, with his large public platform, popularized them. Sam Harris, another person with a large audience, then championed The Bell Curve in several of his podcasts. His position was that Murray, the surviving author, was maligned not because he got the science wrong, but rather because his views are politically incorrect. One could go on, but this is the general position of the far right at present. They tell their listeners, who seem to be growing in number, that the science is, and always has been, on their side, but the debate does not happen because the left fears the truth. This is in keeping with what they like to call their field now, “race realism.” I brought up the prominent scientists who supported scientific racism earlier, incidentally, not to simply malign them but because the support of such objectively important scientists obviously has an effect. You cannot simply ignore people of that stature or condemn them socially but not go into the scientific details. You must respond categorically and, preferentially, with a face-to-face debate.
The primary response of the academic community to this resurgence of racist pseudoscience has been to ignore it. Moreover, the question of race has been ceded almost entirely to the humanities and social sciences (Omi and Winant 1994; Obach 1999; Selden 2000; Nieto 2009; Omi and Winant 2012; Pierce 2014; Bryant et al 2022; Amemiya et al 2024). According to scholars in those fields, races are social constructs with no biological basis. Authors in those fields have gone even further, in fact, and argued that discussion of race (from any perspective other than their own) is bigoted and unscientific (reviewed in Carl 2018). Within this climate, most biological papers on the subject simply rubber stamp their perspective (Kaplan 2011; Graves 2001, 2005; Duello et al 2021; Lala and Feldman 2024; but see Pigliucci and Kaplan 2003). The most recent example I could find of the now conventional biological view, and one we will use to frame some of our discussion in the present paper, is a recent paper by Lala and Feldman (2024). Here, the authors refute many specious arguments about genetics and race. However, they do so after making a tangled argument that race is a social construct.
In the present paper, I will explore the biological nature of race. This will involve providing the counter argument to authors such as Lala and Feldman (2014), while also starting over with the correct biological concepts, which relate to local adaptation, not genome-wide population structure. Essentially, no one, not even the racists, believe that humans have genome wide genetic differentiation between populations that is strong or discrete. Pretending that they do is a strawman argument. The question has always been one of local adaptation for a few traits, something for which genome-level statistics are inappropriate. After correctly describing race in biological terms, we will briefly go over the nature of racist pseudoscience, which is a testament to spin doctoring and lying by omission. We will then tackle the intersection between the social construction of race and its biological basis. Finally, I conclude with a brief discussion of The Bell Curve.
1.1. Are There Rules for Defining Race in Biology?
Lala and Feldman (2024) give a classic perspective on race from a biological viewpoint that focuses on the use of population genetics. The basic idea is to show that humans do not fall into discrete groups. This approach was pioneered by Lewontin5, and has subsequently been used many times by those trying to argue that races do not exist (Lewontin 1970, 1972; Bamshad et al 2004; Graves 2005; Dupre 2008; Shen and Feldman 2022). They primarily focus on measurements of Fst, or closely related statistics, which measure genetic differentiation between populations at either a single locus or averaged across the whole genome (Wright 1949; Falconer 1989). Fst is a number between 0 and 1, with 0 being no differentiation and 1 complete differentiation. When there is genetic differentiation across populations, we say that there is population structure. Population structure can be caused by a lack of gene flow, which causes alleles to drift apart, by selection, founder effects, or other forces (Wright 1949; Weir and Cockerham 1984; Meirmans and Hedrick 2011). A key point to understand is that lack of gene flow causes genome-wide population structure, while selection has a local effect on population structure (that is, it changes allele frequencies at the target of selection and near it on the chromosome). Within this framework, Lala and Feldman (2024) tell us that race has a precise meaning in biology, which is that a genome wide Fst of greater than 0.25 warrants a racial distinction.
Contrary to what some may assert there is no formal definition of race, or any closely related idea, in biology. First, the utility of the notion of subspecies, race, and so forth has always been debated, independent of its value for classifying humans (Wilson and Brown 1953; Mayr 1982; Phillimore and Owens 2006; Burbrink et al 2022). It is off-topic, so we won’t go deep into it, but I think the issue is that most species do not have subspecies, races, or ecotypes (to be introduced later), so most biologists do not have to deal with strong and geographically structured phenotypic variation. It is thus easy for many scientists to dismiss race, particularly since it is a loaded topic6. The reason most species do not have strong population level variation, incidentally, is because it is usually associated with occupying a broad geographic range, and most species seem to have narrow ranges and habitat requirements7. Extreme cases, similar to our own, in which there is a single species that has a vast distribution covering multiple continents are quite uncommon.
If we limit discussion to just those biologists who study species with significant geographical variation, then the study of race, or subspecies, is common and important but has no agreed upon rules (Mallet 2007; Stevens 2023; Hapeman and Smith 2024; Krohn et al 2024). I study social behavior and evolution, for example, and use honey bees as a model system. The western honey bee has a distribution that covers all of Africa, Europe, and the Middle East. It is locally adapted to the northern temperate zone, to savannas, jungles, mountaintops, etc. The bees in the tropics have many differences in physiology and behavior relative to those in Northern Europe, although they are clearly the same species. The honey bee is much like us in that it is flexible enough to live almost anywhere. Also, as for humans, there is gene flow across the whole range of the western honey bee, and hybrid zones exist between one race and the next (all variation is therefore continuous from one region to the next). So, how is this variation dealt with in the honey bee? There are currently 33 subspecies of honey bees (Ilyasov et al 2020). Subspecies, also commonly referred to as races in this literature, are based either on morphology (this population differs from that in size or shape) or genetic differences (Sheppard et al 1997; Oleksa and Tofliski 2015; Chen et al 2016; Momeni et al 2021). The distinctions based on genetics are not necessarily based on any specific cutoffs, or on analyses like the calculation of Fst values. Sometimes people look for population structure, other times they simply make an evolutionary tree, based on whatever genetic data are available, and call each branch a subspecies.
The situation I describe for the honey bee is not unique; it is general. This is to say that the biologists studying a particular species simply decide what sort differences are important enough to warrant some designation. Hence, for some species, like honey bees, there are many subspecies, but for other species, with similar variation, you find few subspecies or none. This situation is typically thought to be the result of researchers falling into the classes of “lumpers” and “splitters.” Lumpers are people who tend to view variation as relatively unimportant. They focus on similarities. Splitters are drawn to differences and focus on those. Obviously, similarities and differences are both important but how much value to attach to one and the other does not lend itself to a simple resolution (Corliss 1976; Endersby 2009). In other words, the personal preferences of the most prominent scientists in a field determine whether there are many taxonomic classes or few. Moreover, this is true for nearly all levels of biological classification. For example, there are seven bee families, but only one ant family (Brothers 2021). Ants are probably an older clade than bees and are arguably more variable as well in terms of morphology, physiology and behavior (Brady et al 2006; Cardinal and Danforth 2014). If the systematists who work on ants chose to, they could create more ant families. They are just lumpers, and the bee systematics splitters.
Let’s expand this discussion and try to draw some general conclusions. The purpose of this section is to show the reader that there are few rigorous rules for classifying biological variation. Hence, the argument that humans do not have races because we fail some well-known test is specious. But that is not even the worst of it, because even in the few cases where there are rules that most people think are reasonable, people break them whenever they feel like it. For example, the lowest taxonomic distinctions, like subspecies and race, and the higher ones, like family and order, are universally agreed to be less rigorous and fundamental than species. The most mechanistic of the species concepts, and the strongest in my opinion, is the biological species concept (Wright 1940; Mayr 1942). It says that two animals are members of the same species if they can interbreed and produce viable offspring. Of course, the idea cannot be applied to fossils and is difficult to apply to extant populations that never meet one another (reviewed in De Queiroz 2007). Nevertheless, when these, and other, problems are absent, this is often considered the main rule of species classification.
People ignore the biological species concept all the time. For example, polar bears and brown bears diverged about 500-600,000 years ago, and can and do hybridize and produce fertile offspring (Pongracz et al 2017; Miller et al 2012). Wolves (of several species) and coyotes also interbreed and produce fertile offspring (Hailer and Leonard 2008). To give an example where there is debate, the African savanna and forest elephants are sometimes considered different species, although some disagree and consider them subspecies (Shetty et al 2011; Mondol et al 2015). In this last case, the subspecies of elephants are arguably as different from one another as are the several species of wolf that exist in North America. In these cases (I could give many more), there is little reason why the biological species concept is sometimes applied and sometimes not, other than the subjective opinions of the leading scientists in each field.
There is another way to think about this problem that is relevant for the present discussion of human differences. In general, some scientists prefer to use genetic data to address questions of classification (and every other question for that matter). They like to come up with relatively arbitrary numerical thresholds or argue that every branch on a phylogenetic tree with statistical significance is deserving of a name. Such people would classify the forest and savannah elephants of Africa as separate species. Other biologists prefer to focus on behavior and physiology (and functional phenotypes in general). This is partly because genetics is under the hood and selection operates on phenotypes (Levins and Lewontin 1985; Lewontin 2000). With respect to species, animals either form one breeding pool or not. People in this second group are more comfortable calling the savannah and forest elephant types of the same species8. I fall into the second camp, but the take home message here is that there is no agreement on these issues and all of what I just said is bitterly contested.
To conclude this section, biological classification systems are arbitrary. Although there are rules, the rules vary and are not agreed upon. We are, therefore, not going to get answers to the question of human race by arbitrarily defining race to equal this or that number measured in this or that way. That does not even work for animals, a case in which there is not a huge amount of baggage and controversy. The best we can do is explore the nature of human variation and decide what biological concepts seem most relevant. We must further try to distance ourselves from the problem and objectively treat the data the way we would for any other species.
1.2. The Relevant Biological Concepts
In this section, we will start with what has now become the traditional biological perspective on race, which focuses on genome-wide population structure. We will argue that this perspective is useful in some ways, but mostly irrelevant for biologically defining race and for countering the arguments made by scientific racists. This is because the modern notion of race is associated with functional differences between populations, which could only result from local adaptation. Natural selection on traits has local not genome-wide effects. Hence, genome-wide statistics, which have more to do with drift or founder effects, are of little use. Moreover, the genetic basis of human intelligence is a mystery, meaning population genetics approaches at the individual locus level also cannot be used. We are thus left with phenotypic data, which should be understood to be paramount anyway since looking for genetic effects without strong proof at the phenotypic level that differences exist is pointless, and in the present case, could only be driven by stubborn bigoted beliefs in differences. Our discussion will thus focus on the nature of local adaptation and how we uncover whether it exists and, if so, what causes it? This will lead us to the concept of ecotype, which we will argue is the best biological description of human races.
1.3. Population Structure
Lewontin was the first to note that the level of genetic variation in the human species is small and does not support the idea that humans can be neatly broken up into meaningful categories (Lewontin 1970, 1972). Many have followed his lead and stressed that about 93% of our genetic variation is within populations, only about 7% of alleles are specific to continents, and overall, Fst is about 0.052-0.083 (Rosenberg et al 2002; Li et al 2008; Rosenberg 2011; Pemberton 2013; Auton et al 2015; Rosenberg et al 2018; Novembre 2022). Humans are thus remarkably homogeneous across our range. All of this is true and important because it sets an obvious ceiling to how different races can be. It also shows that the overall differences between populations are small, and probably trivial9. It does not, however, suggest that human races do not exist, nor does it show that human groups cannot vary for important traits like intelligence.
It is easy to make clear the second point of the previous chapter, because it is conceded by Lala and Feldman (2014) in their review. They state:
“In principle, a low global FST would not preclude a small number of intelligence- related genes of major effect that differed in frequency between socially defined races, but in practice, no such genes have been found.”
In other words, they admit that their long-winded discussion about genome-wide genetic variation, which follows that of many others, was irrelevant because those statistics would not pick up the signal from even the most obvious genetic mechanism that would cause significant variation in a trait like intelligence. They then say, correctly, that no such genes have been found, but this is also mostly irrelevant, because we do not know the genetic basis of human intelligence. Moreover, even if we found a locus that contributes to IQ, with allelic variation across races, it could still not prove the racial superiority of any group. This is because there are undoubtedly many loci that contribute to IQ, and other loci could contribute to IQ in an opposing manner. In other words, if an allele that increases IQ is found to be higher in Europeans (the holy grail of scientific racist scholarship), there could easily be other alleles that also increase IQ and are higher in Africans10. They would not have been found because nearly all the effort goes into finding such effects in European populations. Hence, until we know the total genetic architecture underlying intelligence, and all the extant variation within it, genetic explorations of intelligence will not determine whether differences exist across races.
2. Conflating Ancestry with Race
The answer of the last section is complete in that genome-wide population structure is irrelevant to the question of racial differences, and whether races exist, because races relate to variation in a handful of traits. Nevertheless, there is an additional aspect to the problem, which we should briefly discuss, as it seems to confuse both some scholars and many members of the public. This is that various companies can tell where your ancestors came from, and the categories they use are geographic in nature. Many people believe they are learning their racial history from such tests (Bolnick et al 2007).
A complete explanation of what these tests measure, and how it relates to evolutionary history, would require a long digression, but the fact that what they measure is independent of what is meant by race, especially in the context of scientific racism, is easy to explain. In short, these tests measure ancestry, or in other words, relatedness. The reason people of Swedish ancestry can be identified by their DNA is that they share a more recent common ancestor than they do with people from Asia or Africa, or even Germany. The key point here is that ancestry is independent, in principle, of functional differences due to variable selective history11. It’s now easy, for example, to distinguish between Irish and English ancestry. Does anyone (anymore) really think there are functional biological differences between the English and Irish? Obviously, the racists of the past thought so, but to bring that into the present discussion, which focuses on groups with obvious physical differences, is anachronous in the extreme12. Given that we can identify groups, like Irish and English, with this approach, or even more fine-grained groups such descent from regions within England, it should be apparent that what is being measured is not the sort of functional differences that are the subject of the racial debate. What is being measured is ancestry (relatedness), which is most usefully associated with families, but which can be extended to far larger groups.
Local Adaptation
Local adaptation refers to natural selection causing functional variation within a species due to variable selection pressures across their range (Kawecki et al 2004; Blanquart et al 2013; Savolainen et al 2013). For example, there are countless species that inhabit valleys and mountaintops. High-altitude populations often have adaptations to their harsh climate, which are sometimes genetic in origin. Let’s imagine, for argument’s sake, that a low altitude population is larger in size, due to greater resources, than the high-altitude population of the same species, a common pattern. Furthermore, let’s imagine that as one goes up the mountain the animals get continuously smaller. In such cases, there is typically not discrete variation at the species level (big and small forms with few intermediates) for at least two reasons. First, the selection pressure might vary continuously as one moves to higher altitude, and this could select for a gradient in phenotype. Second, and probably more relevant to the present discussion, migrants from each zone of adaptation move into the other and prevent selection from operating in a vacuum. Hence, large-bodied animals move up the mountain when they disperse and look for mates, bringing their alleles causing large size with them. They might even have an advantage in mating given their large size and have high reproductive output outside their place of origin. The result is continuous change (large to small size) as one moves up the mountain. In short, the likelihood of migrants arriving with optimal phenotypes from another area decreases with distance from that area, creating a gradient. In cases like the one I describe, the valley and mountaintop populations would be called, “ecotypes.” This is meant to illustrate the idea that a variable environment is selecting for adaptive variation in just those traits related to it (Turesson 1922; Gregor 1944; Engelhard et al 2011). There is thus a mountain ecotype and a valley one. No assumption is made regarding change in traits not related to the variable selection pressure, other than that differences in those traits are unlikely since the effects of selection are local at the genomic level. Hence, this notion of ecotypic variation is, in principle, independent of genome wide population structure, although in some cases the two can go hand in glove13.
The study of local adaptation in humans, mainly at the genomic level, is a rapidly growing field (reviewed in Rees et al 2020). Early successes related to the evolution of lighter skin and the ability to digest lactose, both of which turned out to be more complex than initially thought (Jablonski 2004; Trivedi and Gandhi 2017; Sabeti et al 2007; Field et al 2016; Segurel and Bon 2017; Rocha 2020). Overall, we now have relatively clear examples of local adaptation related to variable selective pressures associated with climate, diet, pathogen exposure, and a handful of physical characteristics (Yi et al 2010; Macholdt et al 2014; Heurta-Sanchez et al 2014; Schlebusch et al 2015; Fumagalli et al 2015; Nedelec et al 2016; Illardo and Nielsen 2018). Most of these examples were discovered at the genomic level (reviewed in Rees et al 2020). However, most of these cases have plausible stories about why selection favored the beneficial alleles at the phenotypic level. Examples include respiratory adaptations to high altitude and the ability to accommodate diets high or low in key vitamins or minerals (Yi et al 2010; Bigham and Lee 2014; Huerta-Sánchez et al 2014; Fumagalli et al 2015). Given how recent these discoveries are, and the currently poor sampling of most tropical human populations, undoubtedly many more cases of human local adaptation are awaiting discovery. In short, much of the obvious physical variation we see across the globe in humans, and probably quite a lot of variation that is not obvious, is due to variable selection pressures related to climate, habitat, or resource availability (Jeong and Rienzo 2014; Fan et al 2016; Rees et al 2020).
I agree with Pigliucci and Kaplan (2003) that human races are ecotypes. Broadly speaking, we seem to have some number of tropical and temperate zone ecotypes with continuous variation between them due to gene flow14. Let’s now consider some common anti scientific-racism arguments considering our introduction to local adaptation. Lala and Feldman (2014), for example, make much of the fact that human variation, while pronounced for some traits, is clinal and varies continuously. The argument, commonly expressed, seems to be that either human variation is discrete, in which case races exist, or human variation is continuous, in which case races cannot exist (Templeton 2013). This is a poor argument, for obvious reasons. To put ourselves in the shoes of the scientific racists, their argument has never been that people vary in discrete ways, either phenotypically or genetically. That cannot possibly be true. They say that some regions are home to superior people, and everyone else is worse. Now whether everyone else gets continuously worse as one gets further away from the zone of optimal phenotype, or whether there is a sharp boundary where people suddenly get worse is irrelevant. Admixture is also irrelevant for the same reason. Hence, bringing up the clinal nature of human variation as though it undermines their position is absurd. Moreover, continuous variation never stops biologists from using ecotypic, or racial, terms for other species. When referencing individuals intermediate between good ecotypes, scientists simply refer to the intermediates as intermediates or hybrids.
All of this begs the question of how biologists handle local adaptation? As for our discussion of biological classification, the answer is somewhat dependent on the whims of the people involved. But to be formal, there is a two-pronged attack. First the phenotypic nature of local adaptation is explored (Falconer and Mackay 1983; Kawecki and Ebert 2004). This involves describing the differences in terms of their qualitative and quantitative characteristics. An attempt is also made to explain why selection has favored the variable forms. For example, we discussed African elephants earlier. For those who view the two types as members of the same species, there is a forest ecotype, which is smaller, allowing it to move in dense forest, and a savannah ecotype, free from this selective pressure, and hence larger. There are also hybrid zones, where elephants intermediate in form between the savannah and forest forms predominate. Scientists also study the strength of selection against intermediate forms, since it characterizes the transition in form from one region to another.
Once the phenotypic nature of local adaptation is sufficiently explored, work can begin at the genetic level. This can take many forms. Classically, what is supposed to happen first is that studies are done showing that the phenotypic variation is genetic in nature. It could be the case, to return to our mountain top example, that a lack of food at high altitude stunts the growth of individuals who would otherwise grow much larger (a purely environmental basis for the population difference). Such a basis for population level variation is commonplace, although complex genotype environment interactions are perhaps more likely (Levins and Lewontin 1985, Lewontin 2000). A common garden, or cross-fostering experiment, in which animals adapted to the different places are raised in the same place, or in the opposite place, can show that differences are genetic in origin since genetically induced differences in phenotype should persist no matter the state of the environment. Unfortunately, common garden experiments are sometimes skipped, and work begins at looking for the genetic basis for the phenotypic differences found by behaviorists or physiologists. Such work comes in many forms, with genomics approaches based on QTL mapping, or GWAS, both of which are correlational in nature, being currently the most popular preliminary techniques (Borevitz and Chory 2004; Korte and Farlow 2013). I could continue with how we further explore the biology of local adaptation, and show causation, but this is sufficient for our purpose, since scientific racists never get beyond the approaches so far outlined.
3. Making Biological Sense of Race
The reader might think that an ecotype is just another word for subspecies, or folk race, and we are playing a game of semantics. This is not the case. Focusing on ecotypes is a distinct way of organizing biological variation. For example, sometimes ecotypes can map onto what are called folk races. They can also map onto traditional notions of population structure. However, ecotypes can lead to the formation of human groups that neither make sense from a population structure or folk race perspective. In other words, the idea that human races are ecotypes is a radical departure from past notions of race, and getting this idea across as succinctly as possible is the goal of this section.
3.1. A Basic Introduction to Evolutionary Trees
This paper will hopefully be read by nonscientists, as well as academics, so we need several sections where we step back from the question at hand to introduce briefly the necessary basic concepts and terminology of evolutionary biology. In this section, we will introduce evolutionary trees. An evolutionary tree is basically a complex genealogy. It is a series of splitting events that leads to a tree like the one shown in Figure 1. This shows the major animal groups. Each group (big or small) is called a clade. To qualify as a clade, all the living individuals that descend from a common ancestor have to be within it. The Ecdysozoa, animals who molt, are thus a clade, even though the nematodes (round worms) and arthropods (insects, crustacen, etc.), look quite different. When you give a name to a clade that does not include all living beings descended from a common ancestor you are creating a non-monophyletic group, a crime in the eyes of many evolutionary biologists. If one tried to make a worm group, for example, based on the superficial similarity of the annelids, which includes the earthworms, and the nematodes, the roundworms, you would be making a paraphyletic group (another expression for non-monophyletic).
The logic of the previous paragraph extends to population structure within species. Population geneticists, the biologists who have been addressing the biological basis of race for the most part, will not call a group a clade unless it is monophyletic. This need constrains the sort of groups that one can form with genomic data. Hence, you could not get a monophyletic tree that matches folk races, although you could get one that breaks the folk races up into many groups. This sort of thing has been the subject of much discussion in the literature on race. In this paper, we have argued that this work is irrelevant since race refers to functional biology, not genealogy.
3.2. Stasis and Convergence
Ecotypes are defined by function, not evolutionary history, so it is important that we also introduce some basic concepts relating to functional similarities and differences across groups. In general, we are interested in forces that might lead to clades that are distantly related being more similar to each other than they are to their nearer evolutionary relatives. When I say “similar” I mean either phenotypically overall more similar or genetically more similar (in terms of functional sequence) for traits of interest. One way this can happen is if there is stasis, which refers to a lack of change over time (Eldredge and Gould 1972; Wake et al 1983; Eldredge et al 2005).
Suppose, for example, we have a species that splits into two populations due to the emergence of some barrier to gene flow within their range (Figure 2A). Suppose further that after a long period of time, the two populations become distinct species, and that one of the species splits again, this time caused by migration. That is, a fraction of one species leaves the habitat in which they evolved and moves to a radically different habitat, such as an island off the mainland. Islands are places where radical change in phenotype can rapidly occur. There are predatory caterpillars in Hawaii, for example. After some period of time, this third population will also become a distinct species.
The hypothetical example of the previous chapter will lead to a simple tree with three branches as in Figure 2B. Species 2 and 3 are closer related to one other than they are to species 1, from a genealogical perspective, which is to say they share a more recent common ancestor with one another than with a member of species 1. Hence, when thinking from a classic evolutionary biology perspective, species 2 and 3 are closer together. However, in our example, there was no change in the environment at the first split of the population, which was caused by the emergence of a barrier to gene flow. Hence, species 1 and 2 may not have undergone any meaningful functional change. In fact, we have what are called cryptic species in which enormous effort has to go into showing that two populations are actually different species given how exactly similar they are in practically every way (Bickford et al 2007; Jorger and Schrodl 2013). Moreover, in our example, species 3 migrated to an island and underwent radical change, perhaps changing its entire way of life (from herbivore to predator, or predator to parasite). Species 3 might be hugely different in form and function across many traits from its ancestors. Hence, from a functional perspective, species 1 and 2 are similar (maybe even identical), and species 3 is really different. Hopefully, this little thought experiment shows why tree building, based mainly on neutral genetic variation, does not necessarily get at questions of similarity and difference from a functional perspective.
A second way we can get important functional variation that does not map onto the genealogical history of groups is when there is convergent evolution (Endler 1986; Stern 2013). This refers to cases in which the same phenotype evolves independently in two groups. In other words, two clades share a phenotype that they did not inherit from a common ancestor. This is caused by the same selective pressure leading to the same phenotype in both groups. When the molecular basis for the shared phenotype is identical in the two groups this is called parallel evolution, which is thought to be less common than when the molecular changes underlying the shared phenotypes are different (either completely or partially). We do not need a long example to make clear how this can lead to a similar situation to what we saw in Figure 2 for stasis, as the reader can think this through. A key difference perhaps between the two situations, however, is that stasis between clades that have never occupied different environments, but have had no gene flow for a long time, might lead to more pronounced and widespread similarity than what we would expect for convergence, which might be more trait specific, or lead to incomplete similarity depending on how long selection has had to operate on the relevant phenotypes and the nature of constraints resisting change.
4. Human Races from an Ecotypic Perspective
Figure 3 shows the phylogeny of human groups (broken up geographically) based on genomic data sampled from people around the world (adapted from Rees et al 2020). This is a tree that reflects genealogy, or in other words the order in which groups split from their common ancestor. When population geneticists say that folk races do not map onto biological variation what they mean is that there is no way to create a black race, for example, because it would have to include black people in Africa and black people in Melanesia, which would be a non-monophyletic group (reviewed in Lala and Feldman 2024). In other words, black people from the Andaman Islands, for example, who are as dark as anyone in Africa, are closer related to Europeans and Asians than they are to Africans. As I said before, one could just call every node on this tree a race and then argue that the white one is better than all rest, which is what scientific racists do. This is why approaches that stress that folk races are not good evolutionary groups are useless for combating scientific racism.
How would we think about whether there are black people from an ecotypic perspective? To make a long story short, this was the hypothetical example we explored in Figure 2. Humans arose in Africa. Of course, there are strongly distinct populations in Africa (omitted from our figure for convenience), but if we focus on one selective context, adaptation to a tropical environment, then the simple story of Figure 2 is relevant. Dark-skinned humans left Africa, and some stayed within the tropics (or close to it), while others migrated to the temperate zone (in many waves of course and over many time periods). The result is that there was likely stasis in many phenotypic traits for those populations that never left the tropics, while those that moved into the temperate zone changed more strongly in phenotype. Hence, for many traits Africans and Melanesians are probably more similar to each other than they are to East Asians or Europeans15, and it is this similarity which they recognize when they meet and recognize each other as “black” people.
Few scientists are interested in proving that folk races have a biological basis. It is a trivial question. I point out that folk races can be made sense of from a biological basis that makes use of more evolutionary biology than just genealogy to point out again that classification is quite flexible from a biological point of view and many notions of race are consistent with science. Let’s turn now to how a mainstream biologist not interested in race matters would use the ecotype idea (whether explicitly or implicitly) in a practical way to explore objectively important scientific questions. This discussion will show why classifying variation can be both interesting and important for understanding our biology.
As we previously mentioned, humans evolved in Africa, most of which is tropical. We then moved out of Africa and populated the rest of the world, adapting to most habitats on earth. This involved local adaptation making us better suited to variations in climate, resources, pathogens, and so forth. Within this context, a major question is what genes underlie adaptation to the climatic conditions of the temperate zone, and conversely what genes are important for tropical conditions. In both cases, the genes would likely underlie what are sometimes called an adaptive suite (Lande and Arnold 1983). The idea is that several traits together make an animal suited to a particular context. For example, light skin might be better suited to a lack of sunshine, but other physiological adaptations to cold tolerance might also co-evolve. To get at the many genes involved in cold tolerance, for example, one might break up humans into three ecotypes: tropical, temperate, and tropical reversion, using skin color and evolutionary history. Tropical reversion would indicate populations that were once temperate zone adapted but then moved back into the tropics (such as Native Americans in the Amazon). Candidate genes for cold tolerance would then be genes that experienced positive selection when the move into the temperate zone occurred but then experienced a reverse in selective pressure (or just a relaxation) in the populations that returned to the tropics16. I say candidate here because there are many other reasons why genes might show this pattern of change other than being associated with cold tolerance (reviewed in Korte and Farlow 2013). Experiments like this narrow down the list of genes that could be involved (the candidates) to a smaller number that is more amenable to functional studies that can show whether they actually are associated with the trait of interest (most turn out not to be).
In the previous example, the three groups we identified might also have been identified by a folk race perspective, or even by population structure. Let’s consider a case where this would not be true. Some interesting work has focused on human adaptation to high altitude (reviewed in Azad et al 2017). Three populations—Tibetan, East African, and Peruvian—have been the focus of study. To follow our last example, if we wish to know what genes are associated with adaptation to low oxygen levels, and other characteristics of living at high altitude, we might decide to break up humans into two groups, a high-altitude ecotype and low altitude ecotype17. In this case, folk races would be in both ecotypes. Asians, for example, can be high altitude or low altitude ecotypes. In such a study, we would be looking for genes that are shared by all three high altitude ecotypes and which show signatures of selection consistent with when the groups moved to high altitude. Of course, this experimental design would not get at genes that changed in one population alone, but other studies would look for those.
In the first example black people were treated as a single ecotype because we were interested in those genes associated with adaptation to a topical or temperate climate. Later we broke up black people into two groups, those with or without a long history of living at altitude. We could further break up the black people of just Africa into many groups that experienced different pathogen pressures to look for genes associated with resistance. The point here is that ecotypes are groups of convenience for testing particular hypotheses. Two populations might be linked together in one study because they have something in common but broken up in the next study because they differ in another way. I used black people as an example but such making and breaking of groups by function would occur across all groups depending on the question at hand. Incidentally, what I mention here about how we would use the ecotypic idea to get at gene function is exactly what most population geneticists are currently doing, albeit sometimes without being explicitly clear about it (reviewed in Rees et al 2020).
In summary, the notion of an ecotype differs from either a subspecies based on genome-wide differentiation, or a folk race based on traits that are striking to us socially. It is a practical way of organizing the large amount of biological variation in our species (at the phenotypic level) in such a way as to enable scientific discovery. With respect to scientific racism, this perspective argues that human beings have meaningful variation that allows for classification into groups. This should not be considered racist and, in fact, is not considered racist by most people. What is racist is claiming that some groups are superior to others for particular traits in the absence of good evidence.
5. Scientific Racism
The basic argument of scientific racism is that because humans vary functionally for some traits, such as skin color or size, we probably also vary for others like intelligence and character. To explain such supposed differences, they hypothesize, for example, that adaptation to the temperate zone requires more intelligence (Rushton and Jenson 2005). The initial inspiration for this hypothesis, of course, was the European conquest of the world during the colonial era. The modern form of scientific racism emphasizes IQ. They claim IQ measures intelligence, independent of the environment, and that they have shown clear differences between races due to genetic effects (Jensen 1969; Gottfredson 1997; Lynn et al 2002; Rushton and Jensen 2005; Rindermann et al 2016). There are many refutations of these arguments made regarding race, genes and IQ (Gould 1996; Fagan and Holland 2002, 2007; Dickens and Flynn 2006; Nisbett 2011; Nisbett et al 2012; Thomas 2016; Johnson 2025). Here I want to drill down on why ideologically driven perspectives, like race does not exist, are so damaging to combating such work.
The crux of the matter is that what is called scientific racism is not scientific at all. It is rather pseudoscientific, much like creationism (Hauser 1995). Like creationists, so-called hereditarians begin their exploration with a firm belief that has nothing to do with scientific data. Their belief stems from their bigotry, which they acquired prior to their training as scholars. Scientific racists support their beliefs, which are elaborated into long-winded ‘sciency’ narratives, by first ignoring, or downplaying, all the best data, as such data invariably does not support their position. Second, they emphasize whatever cherry-picked data they can argue support their views. These are often ambiguous results that are open to interpretation. Their favored topics of discussion are also often off topic, but spin doctored to seem relevant, or on topic but shoddy in nature. To give a few examples, there was a transracial adoption study in Minnesota which attracted much attention because the data was construed to support the notion of black inferiority in IQ (Scarr and Weinberg 1976; Weinberg et al 1992: Herrnstein and Murray 1994; Gottfredson 1997). The study compared children of various racial backgrounds adopted into middle class white households. The study had terribly confounding variables, and for this reason, no serious scientist, including the original authors, would draw strong conclusions from it18. A short time later, another transracial adoption study was conducted in Arizona with better control (it largely eliminated the confounding variables) and found no evidence for genetic differences (Moore 1986). This study looked at biracial and black children adopted into middle class black or white homes (a neater natural experiment).19 A 50% difference in the level of European ancestry had no effect on IQ (Figure 4). This second study is either ignored by scientific racists, or downplayed in an almost farcical manner, as they pontificate based on the Minnesota study (reviewed in Nisbett 2011).
A second example relates to the idea that the races differ in scholastic outcome in the United States in a manner supporting the notion of black inferiority. Scientific racists often state that the ranking of IQ, and academic performance, in the US, is Asian, white, Hispanic, black20. They cite nationwide statistics in support of this, or cherry-picked studies (Gottfredson 1997). However, there is strong regional variation in Hispanic and black socioeconomic outcome. Nationwide statistics comparing black and Hispanic achievement, meant to suggest a genetic effect, are therefore bound to suffer from confounding variables. Nevertheless, comparing Hispanics and blacks who live in similar environments might be a reasonable way to start testing the notion that genetics are at play in the poor performance of black Americans. In Johnson (2025), I did this using the national report card, a long running standardized testing program conducted across the US. In New England, where Hispanics and blacks live in similar urban neighborhoods, there is no meaningful difference in academic achievement (Table 1). Given that black Americans have far more African ancestry than the Hispanics of this area, the data do not support the notion that such ancestry is associated with poor performance. In short, a real scientist would devise a comparison that at least tries to control for obvious confounding variables when testing hypotheses, such as this group does better than that one independent of the environment. Asians in the US, for example, have either high or low IQs depending on their country of origin and economic class (reviewed in Johnson 2025). One never finds discussion of such facts in the scientific racism literature. It is all lies by omission, shoddy tests that ignore confounding variables, or taking weak data out of context.
We can now explain the main problem with the “races do not exist” approach to combating scientific racism. First, scientific racism is not serious science, or even science at all. It is not difficult to debunk this work. Whenever even remotely non racist people are exposed to the facts, the pseudoscientific nature of scientific racism is apparent. This does not mean that it will go away, of course, as the same is true of creationism, which is not supported by the facts either. However, scientific racism, unlike creationism, is a pernicious set of beliefs that most people perceive as morally reprehensible.21 The only way to lose the audience, in fact, is to behave in a pseudoscientific manner that shakes the audience’s confidence in your objectivity, and this is exactly what one does when they push the idea that race has no biological basis. This belief is inconsistent with common sense, and with how we treat every other animal from a biological perspective. Thus, we snatch defeat from the jaws of victory, when we push ideology over scientific objectivity.
Race Is Both a Social and Biological Construct
We have focused on the biological basis of race. But race is also a social construct, so we should explore how this interacts with the biology (Omi and Winant 1994; Obach 1999; Amemiya et al 2024). Like the other topics discussed in this paper, this seemingly complex question is straightforward, although a little complicated. Generally speaking, of course, race is socially constructed. We are deeply social animals and many of our concepts are socially constructed. This fact, however, is independent of whether there is a biological basis to what we socially construct. In short, we can socially construct categories in cases in which there is no meaningful biological variation22, and we can construct categories where meaningful biological variation exists. Moreover, we can choose to ignore meaningful biological variation23. Essentially, the idea that the social construction of concepts is paramount is correct, but its application to the context of race at present is flawed. We will develop these ideas in this section.
Race has been defined in myriad ways through the ages. It used to be common to see each nationality as a race (Darwin 1859, 1872), for example, while more recent ideas posited a small number of races associated with broad geographic regions (reviewed in Gould 1996). Many discussions of race from a biological point of view attempt to show that human variation cannot be partitioned (using some manner of cluster analysis with genomic data) in a manner that supports these racial concepts, often called “folk races” (reviewed in Feldman and Lala 2024). One problem with this approach is that it is a textbook example of the continuum fallacy (Graves 2001; Pigliucci and Kaplan 2003b; Edwards 2003). This fallacy is best illustrated with an example. Consider the concepts of tall and short. What if someone argued that tall and short do not exist because height is continuously variable and there is no way to decide how to define them? They would be wrong, of course, because human variation in height is real, and this variation is the objective thing that we are referencing with the concepts of tall and short. Within this context, we socially decide, that is, subjectively determine, only where to draw the lines for tall or short. Hence, you can disagree with where we should subjectively draw the line demarcating tall or short, but not the objective fact that tall and short exist. Moreover, even if you decide that being tall or short does not matter, and is undeserving of a name, it does not change the fact that such variation exists. Essentially, in such cases, we socially construct from real biological variation. Further, if you decide, and then act, as though height does not matter, when it clearly does for millions of other people, then you are being delusional about the present state of the world.
Let’s consider race in light of our discussion of the continuum fallacy. Race fundamentally refers to the biological variation within the human species. As for height, or weight, or countless other variables, the variation people consider with respect to race is continuous. What one can disagree with in this case is where we draw the line demarcating this or that racial group. We can also decide that the relevant variation is not socially meaningful and does not warrant categories. With respect to the first point, whether there are 3, 5, or 500 races does not change the fact that humans vary, and that the variation is structured in various ways. Moreover, from an evolutionary perspective, the fact that variation is continuous between one region of local adaptation, and another is to be expected and does not preclude the use of the term race. With respect to the second point, I personally think racial differences are socially meaningless and I think we will eventually reach a point at which this is the conventional view. The problem is that we are not there yet. Most people, on the left and right, do not think this way. Hence, you cannot simply declare racial variation meaningless and move on. When you do this, you do not end the discussion. You run away from it, and it carries on without your input.
6. The Bell Curve
The Bell Curve is the most high-profile modern example of scientific racism (Herrnstein and Murray 1994). The core idea of this book is that society is stratified based on intellectual ability. The authors argue that IQ is an accurate measure of raw intelligence, which they suggest determines a person’s place in society. They argue that this applies both within and across races. The most contentious points they raise concern their reasons for the low economic success of black Americans. They argue that the one standard deviation difference in IQ, which they estimated, is genetically determined and cannot be changed. Hence, black people will always be at the bottom and programs designed to help the economically disadvantaged are pointless.
Every component of The Bell Curve’s argument is scientifically invalid. IQ does not measure raw intelligence, as the environment has been shown to massively affect it (reviewed in Nisbett 2011). Genetic causes for IQ differences have not been shown (reviewed in Johnson 2005), but even if they existed, they could be changed (Lewontin 1972; Levins and Lewontin 1985). I could elaborate further, but all of this was said at the time of The Bell Curve’s publication, which is why it was not taken seriously within academia (Heckman 1989; Sternberg 1995; Hauser 1995). A key development in this story was the recent study by Chetty et al (2020). Here they analyzed the largest dataset yet on economic outcome in the United States and found that the achievement gap between white and black Americans is entirely due to differences in the outcome of black and white males. Black females had slightly higher upward economic mobility compared to white females when initial economic class was controlled for. The key point to take into consideration here is that there is no mean difference in male and female IQ, in either race. Therefore, IQ is likely not a strong determinant of economic success in the United States because the achievement gap is strongly sex specific while IQ scores are not. This simple fact greatly undermines the argument of The Bell Curve.
Despite this decisive blow to the core idea of The Bell Curve, it made a resurgence in popularity after the publication of Chetty’s work. Charles Murray made the rounds to many well watched right-wing podcasts, all of whom warmly welcomed him and let him make his case in classic scientific racism fashion, which is to say he ignored the Chetty study and presented his cherry-picked data with no push back. I think this chain of events was mainly possible because the significance of the Chetty study, which did receive a lot of press attention, was not much discussed in light of The Bell Curve.
7. Conclusions
This paper is organized into a narrative that moves from topic to topic in a somewhat offhand manner. To conclude, I will therefore summarize the main points as distinctly as possible in bullet points.
- If an alien arrived and treated human beings the way we scientifically treat every other animal species, they would almost certainly identify many ecotypes. “Ecotype” seems the correct expression for human races, given that our differences relate to local adaptation and are few in number. Subspecies, the other possible term, seems better suited to cases of genome-wide differentiation, which is weak across human populations.
- Attempts to show that races do not exist because population structure does not map onto folk races, i.e. black, white, Asian are without merit for two reasons. First, they commit the continuum fallacy in that race is fundamentally about biological variation within our species. How we carve the variation up, or if we choose to do so at all, does not affect the existence of the variation. Proving that variation does not matter requires phenotypic experiments, not exploration of population structure. Second, with respect to structure, scientific racists could not care less about how race is defined. They do not care about the number of races, what they are called, whether they are based on large scale genomic differentiation, or on a few mutations. All they care about is whether functional differences exist for traits like intelligence. Hence, studies of genome-wide human population structure are strawman arguments with respect to both race and scientific racism.
- Given that we do not know the genetic basis of intelligence (even in gross approximation), the only data that can address beliefs in racial superiority for this trait are phenotypic in nature. The relevant data suggests no difference whatsoever in the phenotypes, meaning genetics studies looking for differences are without merit. In short, genetic studies that claim to inform us on this topic are of little value no matter their political agenda.
- Ideologically driven attempts to show that races do not exist, although well intentioned, erode the public’s confidence in the objectivity of scientists when it comes to this contentious issue. These attempts have thus ironically strengthened the position of scientific racists.
| 1 | It seems to be a convention in evolutionary biology, which I will not follow because I think it is historically inaccurate, to forgive Darwin all his discussion of the inferiority of so called “savage races,” by which he meant all nonwhites. Darwin also expressed belief in the idea that the so-called lesser races will eventually face extinction in the face of competition from what he called the higher races of Western Europe. His belief in such ideas, and especially his unwavering support for the even worse bigotry of his cousin Galton, gave immense scientific credibility to scientific racism. |
| 2 | Bryan also cared about the theological issues. I focus here on the neglected part of this history. |
| 3 | Crick’s views can be found in his correspondence which is available online. E. O. Wilson’s support was also made clear in his personal correspondence, which was published after his death. |
| 4 | I prefer to call it pseudoscientific racism, which I think better captures what it is. I will cover this later in this paper. |
| 5 | Lewontin was a great scientist who radically expanded our understanding of evolution. Unfortunately, this is not as appreciated, even in science, as it should be because he was more prone to mixing ideology and science than any other foundational figure in biology and this has hurt his legacy. |
| 6 | This is conjecture, of course; the authors I discuss here do not admit this. |
| 7 | I do not know of a study that quantifies this claim, but whether it is true or not, the main point would stand, which is that many, and probably most, species do not have races. |
| 8 | “Type” can refer to race, subspecies population, or ecotype depending on the whim of the author. |
| 9 | This is to say that the idea that the races are fundamentally different, that is, strongly different across the entire genome cannot be true. Rather, if there are differences, they must relate to particular, traits. In other words, race must be skin deep. The problem with this perspective is that racists do in fact think race is skin deep, they just want to substitute the trait skin with intelligence or character. |
| 10 | It would probably have to be variation in the expression levels, or timing or place of expression, of one or more genes, or some combination of alleles, but the argument is the same whether the genetic basis is simple or complex. |
| 11 | I say in principle because shared ancestry can correlate with functional differences, but they are separate concepts. In other words, if all humans were limited to one region, Europe, for example, and no concept of race even existed we would still be able to do all the 23andme type analyses to identify people’s ancestry by their country of origin. |
| 12 | Clearly the racist notion of Irish inferiority, popular in the past, was socially constructed and we will return to this topic later when we discuss the social and biological construction of race. |
| 13 | When there is a barrier to gene flow (of variable force) between two populations and their habitats differ, then both genome wide population structure and stronger variation in the traits related to the selection are both predicted to occur. |
| 14 | The number of races, and their places of origin, is a classic lumper splitter problem that will never be resolved to everyone’s satisfaction. We clearly have many more tropical ecotypes, but a dispassionate biologist looking at human variation would likely hypothesize that we have many ecotypes in both the tropics and in the temperate zone. Founder effects, sexual selection and other forces in addition to natural selection probably add further complexity to our patterns of variation. |
| 15 | This is certainly true at the phenotypic level but at the genetic level it maybe true for some loci but not others. Even in cases where the genetic basis for a shared trait is different, however, it does not change the fact that from a functional perspective there is strong similarity, or even identical function. |
| 16 | Needless to say, I am giving a simple example to illustrate a general point, while ignoring much that goes into the design and interpretation of such studies. |
| 17 | Depending on the goals of the study we may or may not break up the three ecotypes into more groups. |
| 18 | The black children were adopted at an older age, and into poorer homes than the white children, making the inference that their IQs were lower due to genetic reasons unsound. |
| 19 | This study found a strong environmental effect suggesting white household are superior to black ones when it comes to providing a place where students can excel in school. This result likely led to this work also being ignored by the left as it neither supports the genetic argument for low black achievement, nor the popular idea that poor black achievement is caused by systemic racism. |
| 20 | They invariably ignore Native Americans, as their low scores, and close ancestry with Asians, cannot be made to fit their narrative. |
| 21 | I only compare scientific racism and creationism in the sense that both are pseudoscientific. Other than this, they could not be more different. I am not one of the many evolutionary biologists with a dim view of religion. I am not a believer, but I think religion does a lot of people good. |
| 22 | Medieval superstitious beliefs about handedness or hair color fall into this category. |
| 23 | Something like this is happening now as the age at which one becomes an adult seems to drift higher and higher. College students are called ‘kids” and the category “teenager” seems to be dissolving away as people in this clear developmental class are referred to as children, something that would have shocked people a century ago. I do not argue, or course, that this is a bad thing; the rise in standard of living has allowed us to make these social changes, which are generally a good thing. These examples simply illustrate that good biological reasons to have clear categories can be be ignored if society sees fit to do so. |
References
- Amemiya, J.; Sodré, D.; Heyman, G.D. Early developmental insights into the social construction of race. Journal of Experimental Psychology: General 2024, 153, 3062–3073. [Google Scholar] [CrossRef] [PubMed]
- Azad, P.; Stobdan, T.; Zhou, D.; Hartley, I.; Akbari, A.; Bafna, V.; Haddad, G.G. High-altitude adaptation in humans: From genomics to integrative physiology. Journal of Molecular Medicine 2017, 95, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- Bamshad, M.; Wooding, S.; Salisbury, B.A.; Stephens, J.C. Deconstructing the relationship between genetics and race. Nature reviews genetics 2004, 5, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends in ecology & evolution 2007, 22, 148–155. [Google Scholar]
- Bigham, A.W.; Lee, F.S. Human high-altitude adaptation: Forward genetics meets the HIF pathway. Genes & development 2014, 28, 2189–2204. [Google Scholar]
- Blanquart, F.; Kaltz, O.; Nuismer, S.L.; Gandon, S. A practical guide to measuring local adaptation. Ecology letters 2013, 16, 1195–1205. [Google Scholar] [CrossRef]
- Bolnick, D.A.; Fullwiley, D.; Duster, T.; Cooper, R.S.; Fujimura, J.H.; Kahn, J.; Kaufman, J.S.; Marks, J.; Morning, A.; Nelson, A.; Ossorio, P. The science and business of genetic ancestry testing. Science 2007, 318, 399–400. [Google Scholar] [CrossRef]
- Borevitz, J.O.; Chory, J. Genomics tools for QTL analysis and gene discovery. Current Opinion in Plant Biology 2004, 7, 132–136. [Google Scholar] [CrossRef]
- Brady, S. G.; Schultz, T. R.; Fisher, B. L.; Ward, P. S. Evaluating alternative hypotheses for the early evolution and diversification of ants. Proceedings of the National Academy of Sciences 2006, 103, 18172–18177. [Google Scholar] [CrossRef]
- Brothers, D.J. Aculeate Hymenoptera: Phylogeny and classification. In Encyclopedia of Social Insects; Springer International Publishing: Cham, 2021; pp. 3–11. [Google Scholar]
- Bryant, B.E.; Jordan, A.; Clark, U.S. Race as a social construct in psychiatry research and practice. JAMA psychiatry 2022, 79, 93–94. [Google Scholar] [CrossRef]
- Burbrink, F.T.; Crother, B.I.; Murray, C.M.; Smith, B.T.; Ruane, S.; Myers, E.A.; Pyron, R.A. Empirical and philosophical problems with the subspecies rank. Ecology and Evolution 2022, 12, e9069. [Google Scholar] [CrossRef] [PubMed]
- Cardinal, S.; Danforth, B. N. The Antiquity of Bees Is Derived from Extant Diversity and Resurrected Fossils. Proceedings of the National Academy of Sciences 2013, 110, 13118–13121. [Google Scholar] [CrossRef]
- Carl, N. How stifling debate around race, genes and IQ can do harm. Evolutionary Psychological Science 2018, 4, 399–407. [Google Scholar] [CrossRef]
- Chen, C.; Liu, Z.; Pan, Q.; Chen, X.; Wang, H.; Guo, H.; Liu, S.; Lu, H.; Tian, S.; Li, R.; Shi, W. Genomic analyses reveal demographic history and temperate adaptation of the newly discovered honey bee subspecies Apis mellifera sinisxinyuan n. ssp. Molecular biology and evolution 2016, 33, 1337–1348. [Google Scholar] [CrossRef]
- Chetty, R.; Hendren, N.; Jones, M.R.; Porter, S.R. Race and economic opportunity in the United States: An intergenerational perspective. The Quarterly Journal of Economics 2020, 135, 711–783. [Google Scholar] [CrossRef]
- Claeys, G. The" survival of the fittest" and the origins of social darwinism. Journal of the History of Ideas 2000, 61, 223–240. [Google Scholar] [CrossRef]
- Cochran, G.; Hardy, J.; Harpending, H. Natural history of Ashkenazi intelligence. Journal of Biosocial Science 2006, 38, 659–693. [Google Scholar] [CrossRef]
- Corliss, J.O. On lumpers and splitters of higher taxa in ciliate systematics. Transactions of the American Microscopical Society 1976, 95, 430–442. [Google Scholar] [CrossRef]
- Darwin, C.; Darwin C. On the Origin of Species by Means of Natural Selection; Murray: London, 1859. [Google Scholar]
- Darwin, C. The descent of man, and selection in relation to sex (Vol. 2). D. Appleton. 1872. [Google Scholar]
- De Queiroz, K. Species concepts and species delimitation. Systematic biology 2007, 56, 879–886. [Google Scholar] [CrossRef]
- Dickens, W.T.; Flynn, J.R. Black Americans reduce the racial IQ gap - Evidence from standardization samples. Psychological Science 2006, 17, 913–920. [Google Scholar] [CrossRef]
- Duello, T.M.; Rivedal, S.; Wickland, A.; Weller, A. Race and genetics versus “race” in genetics: A systematic review of the use of African ancestry in genetic studies. Evol. Med. Public Health 2021, 9, 232–245. [Google Scholar] [CrossRef]
- Edwards, A. W. F. Human genetic diversity: Lewontin’s fallacy. BioEssays 2003, 25, 798–801. [Google Scholar] [CrossRef]
- Eldredge, N.; Gould, S.J. in Models in Paleobiology, ed Schopf TJM (Freeman, San Francisco), pp 82–115. 1972. [Google Scholar]
- Eldredge, N.; Thompson, J.N.; Brakefield, P.M.; Gavrilets, S.; Jablonski, D.; Jackson, J.B.; Lenski, R.E.; Lieberman, B.S.; McPeek, M.A.; Miller, W. The dynamics of evolutionary stasis. Paleobiology 2005, 31, 133–145. [Google Scholar] [CrossRef]
- Endersby, J. Lumpers and splitters: Darwin, Hooker, and the search for order. science 2009, 326, 1496–1499. [Google Scholar] [CrossRef] [PubMed]
- R. M., May (Ed.) Natural Selection in the Wild; Princeton Univ: Princeton.
- Engelhard, G.H.; Ellis, J.R.; Payne, M.R.; Hofstede, R. Ter; Pinnegar, J.K. Ecotypes as a concept for exploring responses to climate change in fish assemblages. ICES Journal of Marine Science 2011, 68, 580–591. [Google Scholar] [CrossRef]
- Fagan J.F., Holland; R. Equal opportunity and racial differences in IQ. Intelligence 2002, 30, 361–387. [Google Scholar] [CrossRef]
- Fagan, J.F.; Holland, C.R. Racial equality in intelligence: Predictions from a theory of intelligence as processing. Intelligence 2007, 35, 319–334. [Google Scholar] [CrossRef]
- Falconer, D.S.; Mackay, T.F. Introduction to quantitative genetics; Longman Press: London, 1996. [Google Scholar]
- Fan, S.; Hansen, M.E.; Lo, Y.; Tishkoff, S.A. Going global by adapting local: A review of recent human adaptation. Science 2016, 354, 54–59. [Google Scholar] [CrossRef]
- Field, Y.; Boyle, E.A.; Telis, N.; Gao, Z.; Gaulton, K.J.; Golan, D.; Yengo, L.; Rocheleau, G.; Froguel, P.; McCarthy, M.I.; Pritchard, J.K. Detection of human adaptation during the past 2000 years. Science 2016, 354, 760–764. [Google Scholar] [CrossRef]
- Fumagalli, M.; Moltke, I.; Grarup, N.; Racimo, F.; Bjerregaard, P.; Jørgensen, M.E.; Korneliussen, T.S.; Gerbault, P.; Skotte, L.; Linneberg, A.; Christensen, C. Greenlandic Inuit show genetic signatures of diet and climate adaptation. Science 2015, 349, 1343–1347. [Google Scholar] [CrossRef]
- Galton, F. Hereditary genius. D. Appleton. 1891. [Google Scholar]
- Galton, F. Eugenics: Its definition, scope, and aims. American Journal of Sociology 1904, 10, 1–25. [Google Scholar] [CrossRef]
- Gottfredson, L.S. Mainstream science on intelligence: An editorial with 52 signatories, history, and bibliography (Reprinted from The Wall Street Journal, 1994). Intelligence 1997, 24, 13–23. [Google Scholar] [CrossRef]
- Gould, S.J. The Mismeasure of Man. WW Norton & Company. 1996. [Google Scholar]
- Graves, J.L. The emperor's new clothes: Biological theories of race at the millennium. Rutgers University Press. 2001. [Google Scholar]
- Graves, J. The race myth: Why we pretend race exists in America. Penguin. 2005. [Google Scholar]
- Gregor, J.W. The ecotype. Biological Reviews 1944, 19, 20–30. [Google Scholar] [CrossRef]
- Haier, H. No Voice at Vox: Sense and nonsense about discussing IQ and Race. Quilette, https://quillette.com/2017/06/11/no-voice-vox-sense-nonsense-discussing-iq-race/. 2017. [Google Scholar]
- Hailer, F.; Leonard, J.A. Hybridization among three native North American Canis species in a region of natural sympatry. PLoS ONE 2008, 3, e3333. [Google Scholar] [CrossRef]
- Hauser, R.M. The bell curve. CONTEMPORARY SOCIOLOGY-A JOURNAL OF REVIEWS 1995, 24, 149–153. [Google Scholar] [CrossRef]
- Hapeman, P.; Smith, L.M. Genetics, geography, and subspecies status of American mink in Florida, with an emphasis on Neogale vison evergladensis. Systematics and Biodiversity 2024, 22, 2330371. [Google Scholar] [CrossRef]
- Heckman, J.J. Lessons from the bell curve. Journal of Political Economy 1995, 103, 1091–1120. [Google Scholar] [CrossRef]
- Herrnstein, R.; Murray, C. The bell curve. Intelligence and Class Structure in American Life, New York. 1994. [Google Scholar]
- Huerta-Sánchez, E.; Jin, X.; Asan, Bianba; Z, Peter; B.M, Vinckenbosch; N, Liang; Y.U, Yi; X, He; M., Somel; P., Ni. Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA. Nature 2014, 512, 194–197. [Google Scholar] [CrossRef] [PubMed]
- Huxley, J.S. Eugenics and society. The Eugenics Review 1936, 28, 11. [Google Scholar]
- Ilyasov, R.A.; Lee, M.L.; Takahashi, J.I.; Kwon, H.W.; Nikolenko, A.G. A revision of subspecies structure of western honey bee Apis mellifera. Saudi Journal of Biological Sciences 2020, 27, 3615–3621. [Google Scholar] [CrossRef]
- Jablonski, N.G. The evolution of human skin and skin color. Annu. Rev. Anthropol. 2004, 33, 585–623. [Google Scholar] [CrossRef]
- Jensen, A. How much can we boost IQ and scholastic achieve ment? Harvard Educational Review 1969, 39, 1–123. [Google Scholar] [CrossRef]
- Jeong, C.; Rienzo, A. Di. Adaptations to local environments in modern human populations. Current opinion in genetics & development 2014, 29, 1–8. [Google Scholar]
- Johnson, B. Race, Genes, Evolution, and IQ: The Key Datasets and Arguments. 2025. [Google Scholar] [CrossRef]
- Jörger, K.M.; Schrödl, M. How to describe a cryptic species? Practical challenges of molecular taxonomy. Frontiers in zoology 2013, 10, 1–27. [Google Scholar] [CrossRef]
- Kaplan, J.M. ‘Race’: What biology can tell us about a social construct. eLS, 2011. [Google Scholar]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecology letters 2004, 7, 1225–1241. [Google Scholar] [CrossRef]
- Korte, A.; Farlow, A. The advantages and limitations of trait analysis with GWAS: A review. Plant methods 2013, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Krohn, A.R.; Apodaca, J.J.; Collins, L.; Hudson, B.D.; Barrett, K. Phylogenetics, subspecies, and conservation of the eastern pinesnake. The Journal of Wildlife Management 2024, 88, e22599. [Google Scholar] [CrossRef]
- Lala, K.N.; Feldman, M.W. Genes, culture, and scientific racism. Proceedings of the National Academy of Sciences 2024, 121, e2322874121. [Google Scholar] [CrossRef]
- Lande, R.; Arnold, S. J. The measurement of selection on correlated characters. Evolution 1983, 37, 1210–1226. [Google Scholar] [CrossRef]
- Levins, R.; Lewontin, R. The dialectical biologist. Harvard University Press. 1985. [Google Scholar]
- Lewontin, R.C. Race and intelligence. Bulletin of the Atomic Scientists 1970, 26, 2–8. [Google Scholar] [CrossRef]
- Lewontin, R. The Triple Helix: Gene, Organism, and Environment. Harvard University Press. 2000. [Google Scholar]
- Lewontin, R.C. Dobzhansky, T., Hecht, MK, Steere, WC, Eds.; The apportionment of human diversity. In Evolutionary biology; Appleton-Century-Crofts: New York, 1972; Volume 6, pp. 381–398. [Google Scholar]
- Li, J.Z.; Absher, D.M.; Tang, H.; Southwick, A.M.; Casto, A.M.; Ramachandran, S.; Cann, H.M.; Barsh, G.S.; Feldman, M.; Cavalli-Sforza, L.L.; Myers, R.M. Worldwide human relationships inferred from genome-wide patterns of variation. science 2008, 319, 1100–1104. [Google Scholar] [CrossRef]
- Lynn, R.; Vanhanen, T.; Stuart, M. IQ and the wealth of nations. Greenwood Publishing Group. 2002. [Google Scholar]
- Macholdt, E.; Lede, V.; Barbieri, C.; Mpoloka, S.W.; Chen, H.; Slatkin, M.; Pakendorf, B.; Stoneking, M. Tracing pastoralist migrations to southern Africa with lactase persistence alleles. Current Biology 2014, 24, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Mallet, J. Subspecies, semispecies, superspecies. Encyclopedia of biodiversity 2007, 5, 523–526. [Google Scholar]
- Mayr, E. Systematics and the origin of species; Columbia Univer sity Press: New York, 1942. [Google Scholar]
- Mayr, E. Of what use are subspecies? The Auk 1982, 99, 593–595. [Google Scholar]
- Meirmans, P.G.; Hedrick, P.W. Assessing population structure: FST and related measures. Molecular ecology resources 2011, 11, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Meixner, M.D.; Leta, M.A.; Koeniger, N.; Fuchs, S. The honey bees of Ethiopia represent a new subspecies of Apis mellifera—Apis mellifera simensis n. ssp. Apidologie 2011, 42, 425–437. [Google Scholar] [CrossRef]
- Miller, W.; Schuster, S.C.; Welch, A.J.; Ratan, A.; Bedoya-Reina, O.C.; Zhao, F.; Kim, H.L.; Burhans, R.C.; Drautz, D.I.; Wittekindt, N.E.; Tomsho, L.P. Polar and brown bear genomes reveal ancient admixture and demographic footprints of past climate change. Proceedings of the National Academy of Sciences 2012, 109, E2382–E2390. [Google Scholar] [CrossRef]
- Momeni, J.; Parejo, M.; Nielsen, R.O.; Langa, J.; Montes, I.; Papoutsis, L.; Farajzadeh, L.; Bendixen, C.; Căuia, E.; Charrière, J.D.; Coffey, M.F. Authoritative subspecies diagnosis tool for European honey bees based on ancestry informative SNPs. BMC genomics 2021, 22, 1–12. [Google Scholar] [CrossRef]
- Mondol, S.; Moltke, I.; Hart, J.; Keigwin, M.; Brown, L.; Stephens, M.; Wasser, S.K. New evidence for hybrid zones of forest and savanna elephants in Central and West Africa. Molecular Ecology 2015, 24, 6134–6147. [Google Scholar] [CrossRef]
- Moore, E.G.J. Family socialization and the IQ test performance of traditionally and transracially adopted black children. Developmental Psychology 1986, 22, 317–326. [Google Scholar] [CrossRef]
- Nisbett, R.E. The Achievement Gap: Past, Present & Future. Daedalus 2011, 140, 90–100. [Google Scholar]
- Nisbett, R.E,. J. Aronson; Blair, C.; Dickens, W.; Flynn, J.; Halpern, D.F.; Turkheimer, E. Group Differences in IQ Are Best Understood as Environmental in Origin. The American Psychologist 2012, 67, 503–504. [Google Scholar] [CrossRef]
- Novembre, J. The background and legacy of Lewontin's apportionment of human genetic diversity. Philosophical Transactions of the Royal Society B 2022, 377, 20200406. [Google Scholar] [CrossRef]
- Obach, B.K. Demonstrating the social construction of race. Teaching Sociology 1999, 27, 252–257. [Google Scholar] [CrossRef]
- Oleksa, A.; Tofilski, A. Wing geometric morphometrics and microsatellite analysis provide similar discrimination of honey bee subspecies. Apidologie 2015, 46, 49–60. [Google Scholar] [CrossRef]
- Omi, M; Winant, H. Racial Formation in the United States: From the 1960s to the 1990s, 2nd ed.; Routledge: New York, 1994. [Google Scholar]
- Omi, M.; Winant, H. Racial formation rules. Racial formation in the twenty-first century, 2012. [Google Scholar] [CrossRef]
- Pemberton, T.J.; DeGiorgio, M.; Rosenberg, N.A. Population structure in a comprehensive genomic data set on human microsatellite variation. G, 8: Genomes, Genetics 3, 2013. [Google Scholar]
- Phillimore, A.B.; Owens, I.P. Are subspecies useful in evolutionary and conservation biology? Proceedings of the Royal Society B: Biological Sciences 2006, 273, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Pierce, J.L. Pierce, J.L., 2014, March. Why teaching about race as a social construct still matters. In Sociological Forum (Vol. 29, No. 1, pp. 259–264). Wiley, Springer.
- Pigliucci, M.; Kaplan, J. On the concept of biological race and its applicability to humans. Philosophy of Science 2003a, 70, 1161–1172. [Google Scholar] [CrossRef]
- Pigliucci, M.; Kaplan, J. Making Sense of Evolution: The Conceptual Foundations of Evolutionary Biology; University of Chicago Press: Chicago, 2003b. [Google Scholar]
- Pinker, S. Groups and Genes. New Republic. https://newrepublic.com/article/77727/groups-and-genes. 2006. [Google Scholar]
- Pongracz, J.D.; Paetkau, D.; Branigan, M.; Richardson, E. Recent hybridization between a polar bear and grizzly bears in the Canadian Arctic. Arctic 2017, 70, 151–160. [Google Scholar] [CrossRef]
- Rees, J.S.; Castellano, S.; Andrés, A.M. The genomics of human local adaptation. Trends in Genetics 2020, 36, 415–428. [Google Scholar] [CrossRef]
- Rindermann, H.; Becker, D.; Coyle, T.R. Survey of Expert Opinion on Intelligence: Causes of International Differences in Cognitive Ability Tests. Frontiers in Psychology 2016, 7, 399. [Google Scholar] [CrossRef]
- Rocha, J. The evolutionary history of human skin pigmentation. Journal of molecular evolution 2020, 88, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.A. Darwinism and social Darwinism. Journal of the History of Ideas 1972, 33, 265–280. [Google Scholar] [CrossRef]
- Rosenberg, N.A. A population-genetic perspective on the similarities and differences among worldwide human populations. Human biology 2011, 83, 659. [Google Scholar] [CrossRef]
- Rosenberg, N.A.; Pritchard, J.K.; Weber, J.L.; Cann, H.M.; Kidd, K.K.; Zhivotovsky, L.A.; Feldman, M.W. Genetic structure of human populations. science 2002, 298, 2381–2385. [Google Scholar] [CrossRef]
- Rosenberg, N.A.; Mahajan, S.; Ramachandran, S.; Zhao, C.; Pritchard, J.K.; Feldman, M.W. Clines, clusters, and the effect of study design on the inference of human population structure. PLoS genetics 2005, 1, e70. [Google Scholar] [CrossRef]
- Rosenberg, N.A.; Edge, M.D.; Pritchard, J.K.; Feldman, M.W. Interpreting polygenic scores, polygenic adaptation, and human phenotypic differences. Evolution, medicine, and public health 2019, 2019, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Rushton, J.P.; Jensen, A.R. Thirty years of research on race differences in cognitive ability. Psychology, Public Policy, and Law 2005, 11, 235–294. [Google Scholar] [CrossRef]
- Sabeti, P.C.; Varilly, P.; Fry, B.; Lohmueller, J.; Hostetter, E.; Cotsapas, C.; Xie, X.; Byrne, E.H.; McCarroll, S.A.; Gaudet, R.; Schaffner, S.F. Genome-wide detection and characterization of positive selection in human populations. Nature 2007, 449, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, O.; Lascoux, M.; Merilä, J. Ecological genomics of local adaptation. Nature Reviews Genetics 2013, 14, 807–820. [Google Scholar] [CrossRef]
- Scarr, S.; Weinberg, R.A. IQ test performance of black children adopted by white families. American Psychologist 1976, 31, 726–739. [Google Scholar] [CrossRef]
- Schlebusch, C.M.; Gattepaille, L.M.; Engström, K.; Vahter, M.; Jakobsson, M.; Broberg, K. Human adaptation to arsenic-rich environments. Molecular biology and evolution 2015, 32, 1544–1555. [Google Scholar] [CrossRef] [PubMed]
- Ségurel, L.; Bon, C. On the evolution of lactase persistence in humans. Annual review of genomics and human genetics 2017, 18, 297–319. [Google Scholar] [CrossRef] [PubMed]
- Selden, S. Eugenics and the social construction of merit, race and disability. Journal of Curriculum Studies 2000, 32, 235–252. [Google Scholar] [CrossRef]
- Shapiro, J.I. On Watson, Racism, and Standardized Tests. Marshall Journal of Medicine 2019, 5, 3. [Google Scholar] [CrossRef]
- Sheppard, W.S.; Meixner, M.D. Apis mellifera pomonella, a new honey bee subspecies from Central Asia. Apidologie 2003, 34, 367–375. [Google Scholar] [CrossRef]
- Shen, H.; Feldman, M.W. Diversity and its causes: Lewontin on racism, biological determinism and the adaptationist programme. Philosophical Transactions of the Royal Society B 2022, 377, 20200417. [Google Scholar] [CrossRef]
- Shetty, N.R.; Vidya, T.N.C. To split or not to split: The case of the African elephant. Current Science (00113891) 2011, 100, 810. [Google Scholar]
- Spencer, H. Social Statics: Or, The Conditions Essential to Human Happiness Specified, and the First of Them Developed. D. Appleton. 1866. [Google Scholar]
- Stern, D.L. The genetic causes of convergent evolution. Nature Reviews Genetics 2013, 14, 751–764. [Google Scholar] [CrossRef]
- Sternberg, R.J. For whom the bell curve tolls: A review of The Bell Curve. Psychological Science 1995, 6, 257–261. [Google Scholar] [CrossRef]
- Stevens, R.D. Wither the subspecies? An ecological perspective on taxonomic, environmental and sexual determinants of phenotypic variation in big-eared woolly bats, Chrotopterus auritus. Biological Journal of the Linnean Society 2023, 139, 203–213. [Google Scholar] [CrossRef]
- Templeton, A. R. Biological races in humans. Stud. Hist. Philos. Biol. Biomed. Sci. 2013, 44, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D. Racial IQ Differences among Transracial Adoptees: Fact or Artifact? Journal of Intelligence 2016, 5, 1. [Google Scholar] [CrossRef]
- Tontonoz, M.J. The Scopes Trial Revisited: Social Darwinism versus Social Gospel. Science as Culture 2008, 17, 121–143. [Google Scholar] [CrossRef]
- Trivedi, A.; Gandhi, J. The evolution of human skin color. JAMA dermatology 2017, 153, 1165. [Google Scholar] [CrossRef] [PubMed]
- Turesson, G. ‘The Genotypical Response of the Plant Species to the Habitat. Hereditas 1922, 3, 211–350. [Google Scholar] [CrossRef]
- Weinberg, R.A.; Scarr, S.; Waldman, I.D. The Minnesota transracial adoption study – a follow up of test performance at adolescence. Intelligence 1992, 16, 117–135. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef]
- Wilson, E.O.; Brown, W.L. The subspecies concept and its taxonomic application. Systematic zoology 1953, 2, 97–111. [Google Scholar] [CrossRef]
- Wright, S. The statistical consequences of Mendelian heredity in relation to speciation. Pages 161–183 in The new systematics (J. Huxley, ed.). Oxford University Press, London. 1940. [Google Scholar]
- Wright, S. The genetical structure of populations. Annals of eugenics 1949, 15, 323–354. [Google Scholar] [CrossRef]
- Yi, X.; Liang, Y.; Huerta-Sanchez, E.; Jin, X.; Cuo, Z.X.P.; Pool, J.E.; Xu, X.; Jiang, H.; Vinckenbosch, N.; Korneliussen, T.S.; Zheng, H. Sequencing of 50 human exomes reveals adaptation to high altitude. science 2010, 329, 75–78. [Google Scholar] [CrossRef]
Figure 1.
Simplified phylogeny of some of the major groups of animals.
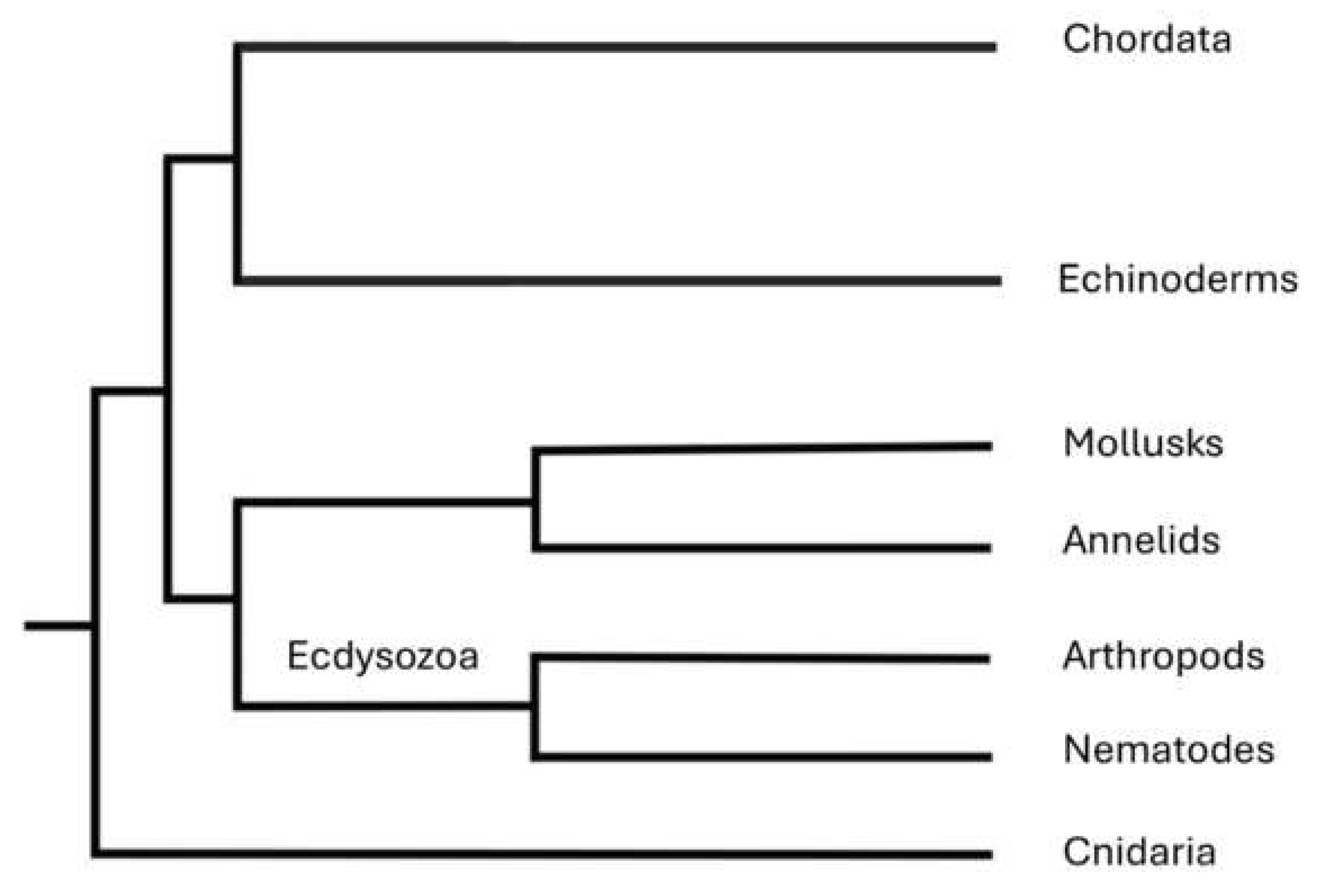
Figure 2.
Illustration of a common process that can lead to greater functional similarity in more distantly related clades. B. Speciation is first via the introduction of a barrier to gene flow within an otherwise unchanged region. A second speciation event then follows when an island off the coast is inhabited by one of the species. B. The resulting phylogeny of the process depicted in part A.
Figure 2.
Illustration of a common process that can lead to greater functional similarity in more distantly related clades. B. Speciation is first via the introduction of a barrier to gene flow within an otherwise unchanged region. A second speciation event then follows when an island off the coast is inhabited by one of the species. B. The resulting phylogeny of the process depicted in part A.
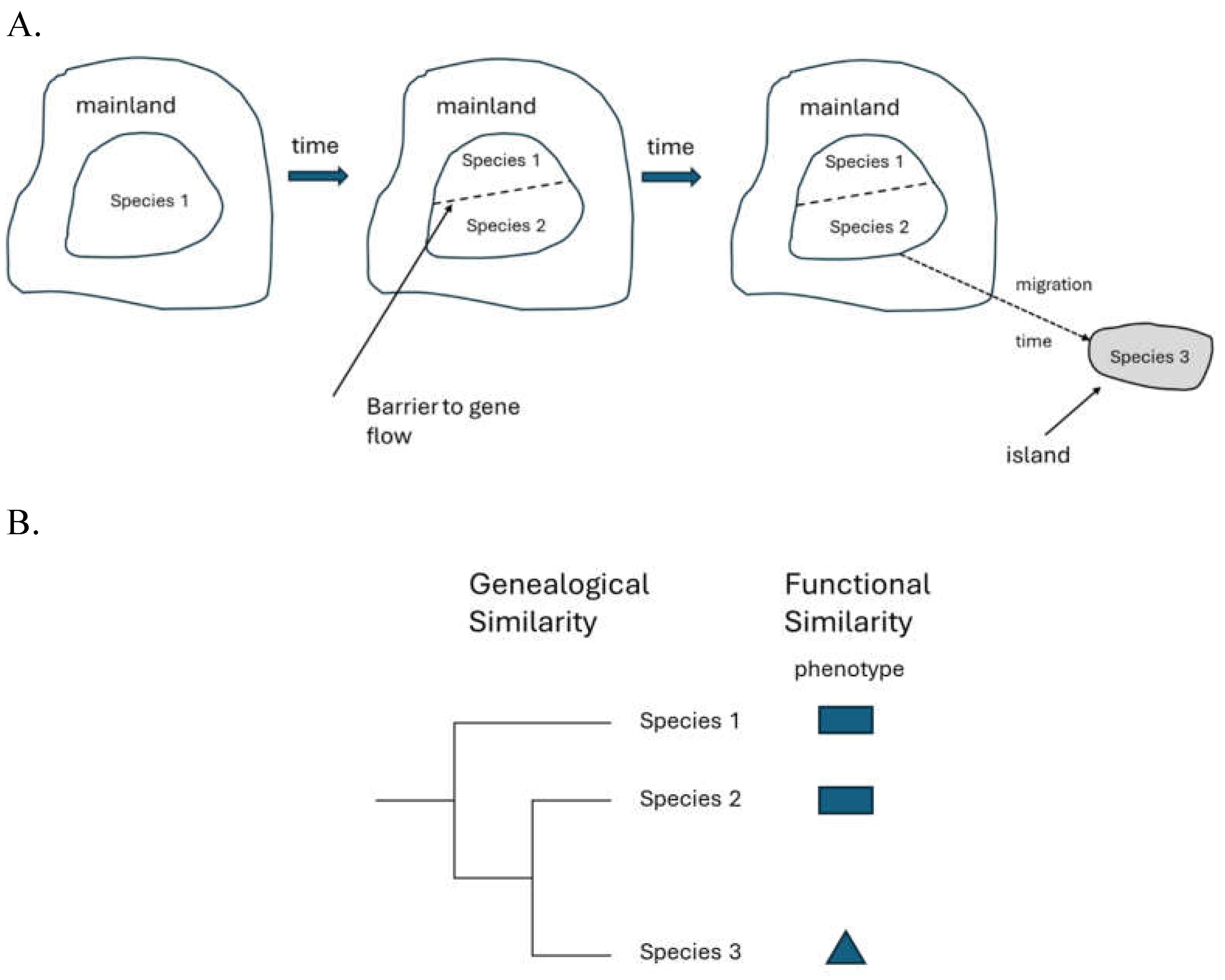
Figure 3.
Phylogeny of human populations by geographic region (adapted from Rees et al 2020).
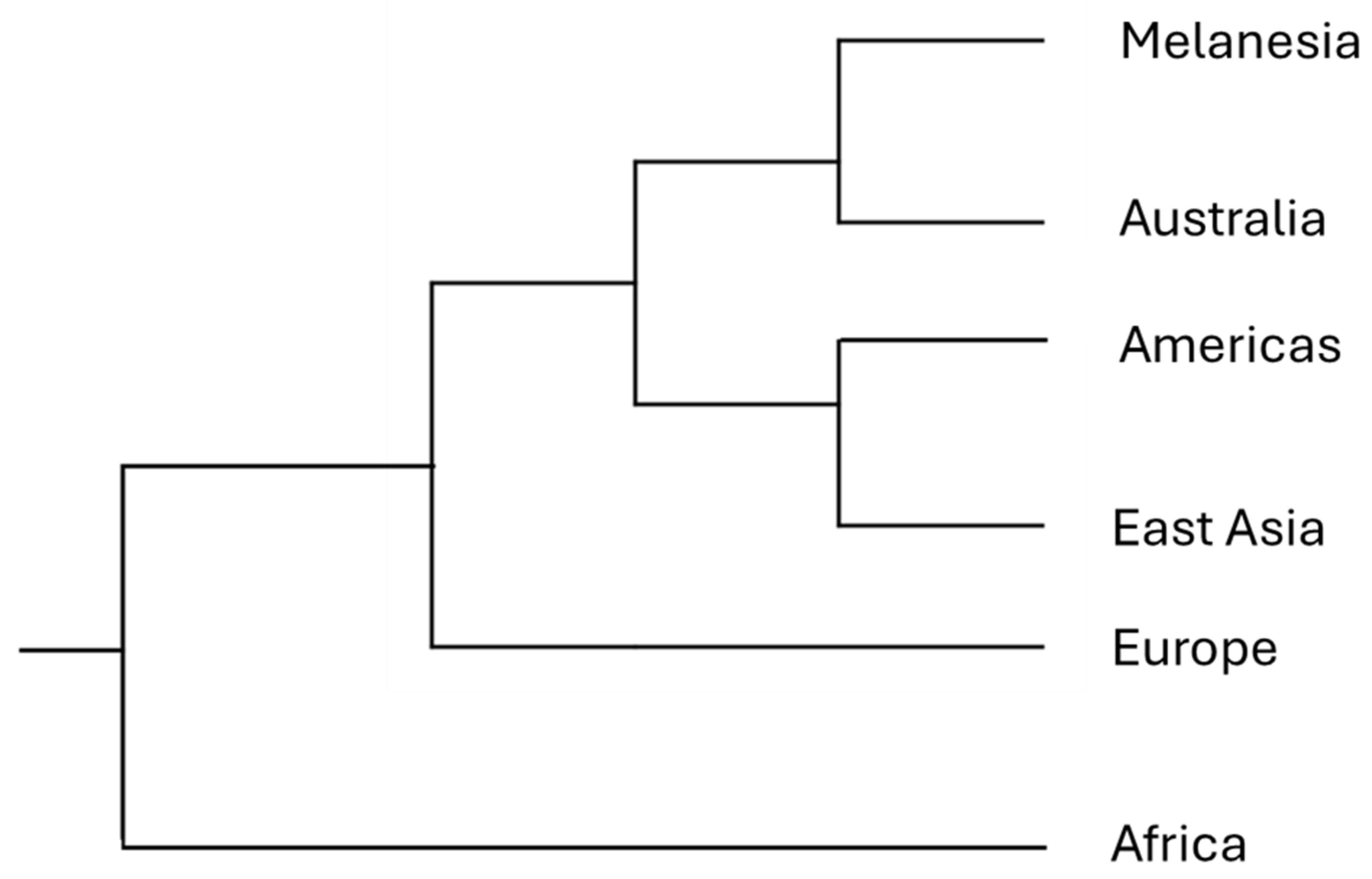
Figure 4.
Black and biracial children had higher mean IQs when adopted by middle class white families than when adopted by middle class black families, but percentage of African ancestry had no effect on IQ in either environment (Moore (1986).
Figure 4.
Black and biracial children had higher mean IQs when adopted by middle class white families than when adopted by middle class black families, but percentage of African ancestry had no effect on IQ in either environment (Moore (1986).
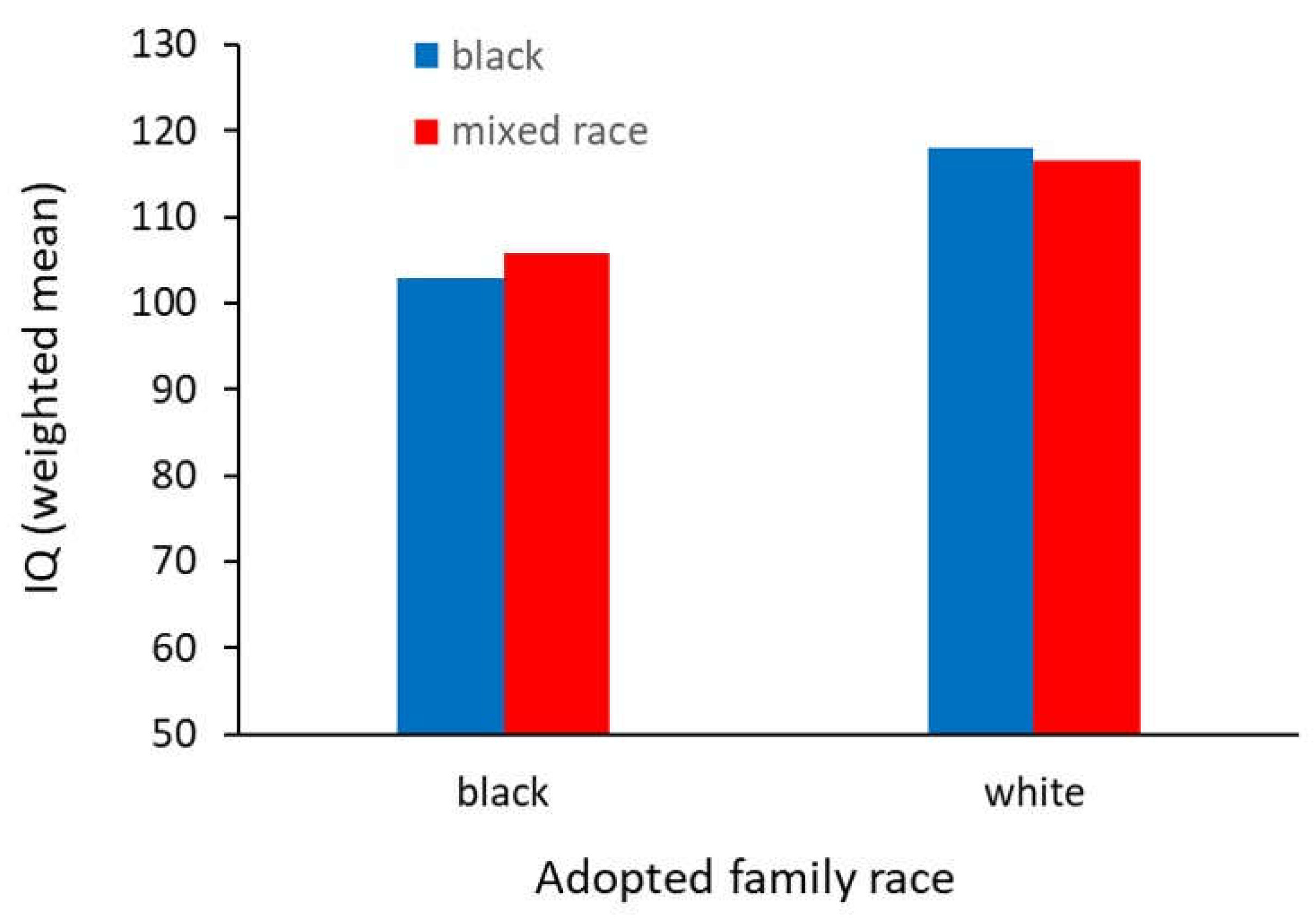

Table 1.
Fourth grade math NAEP scores by racial/ethnic group. See Johnson (2005) for a complete discussion of this data.
Table 1.
Fourth grade math NAEP scores by racial/ethnic group. See Johnson (2005) for a complete discussion of this data.
| Connecticut | 2011 | 2013 | 2015 | 2017 |
| white | 253 | 253 | 249 | 249 |
| black | 220 | 219 | 218 | 222 |
| Hispanic | 222 | 224 | 224 | 223 |
| Massachusetts | 2011 | 2013 | 2015 | 2017 |
| white | 258 | 260 | 256 | 255 |
| black | 235 | 230 | 230 | 229 |
| Hispanic | 236 | 234 | 232 | 234 |
| New York | 2011 | 2013 | 2015 | 2017 |
| white | 245 | 248 | 245 | 244 |
| black | 224 | 225 | 221 | 224 |
| Hispanic | 226 | 229 | 228 | 223 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



