Submitted:
17 February 2025
Posted:
18 February 2025
You are already at the latest version
Abstract
In recent years, bipolar disorder (BD), a multifaceted mood disorder marked by severe episodic mood fluctuations, has been shown to have an impact on disability-adjusted life-years (DALYs). The increasing prevalence of BD highlights the need for better di-agnostic tools , particularly those involving genetic insights. Genetic association studies can play a crucial role in identifying variations linked to BD, shedding light on its ge-netic underpinnings and potential therapeutic targets. Our research utilized data from the Taiwan Precision Medicine Array (TPM Array), aiming to identify significant single nucleotide polymorphisms (SNPs) associated with BD. Notably, we identified the var-iant rs11156606 in the ABCD1 gene, which plays a role in fatty acid metabolism—a process potentially connected to BD pathophysiology. Subsequent linkage disequilib-rium (LD) analysis of rs11156606 revealed a strongly associated variant, rs73640819, located within the ABCD1 3’ untranslated region (3’UTR), suggesting a regulatory role on ABCD1 RNA functionality. Additionally, our findings indicate that the variant rs3829533 is in strong LD with rs3751800 and rs3751801, which significantly affect the coding sequence of the MTHFSD gene. These findings enhance the genetic under-standing of BD in the Taiwanese Han population and highlight the value of genetic research in advancing our knowledge of bipolar disorder.
Keywords:
Bipolar disorder
; Genetic association study
; Genetic variants
; Taiwanese Han popula-tion
; Fatty acid metabolism
1. Introduction
Bipolar disorder (BD) is a clinically severe mood disorder with a lifetime prevalence of 4%[1]. From 1990 to 2017, the incidence of BD increased by 47.74% and the disability-adjusted life years (DALYs) associated with the condition increased by 54.4% during this period [2], taking away years of healthy functioning from individuals with the illness.
Currently, BDs are primarily diagnosed by careful assessment of behavior combined with subjective reports of abnormal experiences to group patients into disease categories standardized in The Diagnostic and Statistical Manual of Mental Disorders (DSM) and The International Statistical Classification of Diseases and Related Health Problems (ICD). However, BD types I and II are difficult to diagnose accurately in clinical practice, particularly in early stages. Besides, nearly a quarter of adults (22.5%) and adolescents with major depressive disorder (MDD), followed up for a mean length of 12-18 years developed BD [3]. Underestimated prevalence and difficulties in diagnosis have resulted in an overarching need for more precise and early diagnostic methods.
Functional studies of candidate genes and their locations within genomic regions linked to schizophrenia (SCZ) or bipolar disorder (BD) through linkage studies were prevalent until around 2006, when genome-disease association studies began to explore potential risk gene variants. This is particularly important in psychiatry, given the unconvincing and inconsistent evidence from candidate gene studies and the genetic architecture for most diseases that seem to be polygenic [4]. Since the first genome-disease association study of BD in 2007, a handful of risk loci have been identified, notably ANK3 [5], NCAN [6], CACNA1C [7] and ODZ4 [8].In recent years, an increasing number of studies have reported hundreds of genes and proteins related to BD, many of which have been suggested as potential biomarkers for this disease, including BDNF [9], RELN [10] and ANK [5]. More importantly, many genes have been reported to influence the pathogenesis of BD through multiple possible mechanisms [4].
In this study, we aimed to demonstrate the capabilities of genome-disease association analysis using the TPM array to detect genetic variations of bipolar disorder (BD) in the Taiwanese population. This analysis aids in predicting specific Taiwanese risk-relevant single nucleotide polymorphisms (SNPs), which could be applied in the diagnosis of BD, by investigating individuals who participated in the Tri-Service General Hospital (TSGH) genetic study project. Additionally, we focused on identifying the potential mechanisms of the most influential characterized variants, as well as related variants in ABCD1 and MTHFSD, which may further inform treatment strategies for BD.
2. Materials and Methods
2.1. Study Participants and Ethical Approval
The data was collected from the TPM array, which aims to create a database containing detailed clinical information and genetic profiles of one million individuals from the Taiwanese Han population. Participants in this study were recruited from medical centers and genotyped by Academia Sinica. DNA extraction and SNPs identification were performed as follows: First, each participant’s genomic DNA was extracted and purified from 3 mL of peripheral blood collected in EDTA vacutainers using the QIAsymphony SP system (QIAGEN, Hilden, Germany). Second, the purified DNA from each participant was loaded onto the TPM array chip, and genome-type signals were detected using the Axiom GeneTitan system (Thermo Fisher Scientific, Sunnyvale, CA, USA). Conversion and quality control of SNP calling and sample annotation was performed using the Axiom Analysis Suite (Thermo Fisher Scientific, Sunnyvale, CA, USA).
2.2. Disease Association Analysis
Figure 1 illustrates the steps involved in the disease association analysis. Initially, 128 individuals diagnosed with BD (ICD-10 code F31), determined through diagnostic interviews conducted by clinical psychiatrists according to the DSM, Fifth Edition (DSM-5) criteria, were recruited from the TSGH. The control group comprised 26,122 individuals without BD. Comprehensive participant information is presented in Table 1.
The case group (128 bipolar disorder patients) and the control group (26,122 participants). Genotyping was performed using the Taiwan Precision Medicine (TPM) Array Chip. Data from 493,852 SNPs were filtered, and 280,177 SNPs were passed through the chi-squared test to detect risk factors.
To analyze the genotyping data from the TPM array, SNPs with a low typing call rate (< 80%) were filtered out, and the remaining data was subjected to chi-squared test association analysis using PLINK 1.9 software (https://zzz.bwh.harvard.edu/plink/) [11]. Variants with low quality (minor allele frequency less than 0.05 and Hardy-Weinberg equilibrium less than 1 × 10⁻⁶) were removed, and highly significant p-values (less than 1 × l0-5) were selected for further analysis.
2.3. Variant Annotations and Functional Analysis
Genes were identified for variant annotations utilizing the RefSeq Database (https://www.ncbi.nlm.nih.gov/refseq/), as described in wANNOVAR (https://wannovar.wglab.org/) [12]. To compare the allele frequency in different racial populations, the public domain databases 1000 Genomes[13], Genome Aggregation Database (gnomAD) [14], and Taiwan BioBank (https://taiwanview.twbiobank.org.tw/index) [15] were used. Further characterization of the genes associated with the identified variants involved examining their biological functions and molecular pathways. The biological processes of the genes, as defined by Gene Ontology [16], were elucidated using Enrichr (https://maayanlab.cloud/Enrichr/) [17].
2.4. Linkage Disequilibrium (LD) Analysis
LD analysis helps researchers understand which genes are likely to be functionally relevant to a trait or disease, as well as the number and location of the contributing genes[18]. In our study, we uploaded the variants rs11156606 and rs3829533 obtained from association study using the LDproxy Tool, which is part of LDlink (https://ldlink.nih.gov/?tab=home) [19,20] and analysis parameters were as follows: 1. Genome Build Version: GRCH37 ( rs11156606) and GRCH38 ( rs3829533). 2. Population: CDX (Chinese Dai in Xishuangbanna, China), CHB (Han Chinese in Beijing, China), and CHS (Southern Han Chinese). 3. LD measurements: D’ (D prime) and R2 (R-squared) were calculated based on allele frequencies. The measured values of D' and R² range from 0 to 1, where a value of 1 indicates complete disequilibrium, and a value of 0 indicates complete equilibrium. The base-pair window is 500,000 bp. Regulatory potential prediction was based on FORGEdb. [21]
3. Results
3.1. Demographics of the Selected Patients
Table 1 lists the demographic composition of the participants enrolled in the study stratified by sex and age group. Within the BD cohort, female participants slightly outnumbered male participants, with 69 females and 59 males. The control group was more extensive, comprising 11,793 males and 14,329 females, indicating a demographic balance that mirrors the expected population distributions. The age distribution within the BD cohort revealed the highest prevalence in the 50-59 age demographic, with a progressive decrease observed in both the younger and older cohorts. By contrast, the control group demonstrated substantial inclusivity across all adult age ranges, ensuring adequate age-matched controls for robust comparative analyses.
3.2. Study Workflow
Figure 1 shows the genetic-disease association study pipeline implemented in our investigation to identify genetic susceptibilities associated with BD. This study differentiated between a case cohort of 128 individuals diagnosed with BD and a sizable control cohort consisting of 26,122 participants. Genotyping was performed using the Taiwan Precision Medicine (TPM) array chip, designed to accurately detect a comprehensive spectrum of SNPs. The initial genotypic yield from this high-throughput platform was an extensive array of 493,852 SNPs.
The subsequent stage involved a filtering process to ensure data integrity and relevance, in which a substantial number of SNPs were excluded from the initial pool owing to factors such as low minor allele frequency, deviations from Hardy-Weinberg equilibrium, or missing data. After filtering out the data that could potentially confound the analysis, a curated set of 280,177 high-quality SNPs was obtained.
The selected SNPs were analyzed using the chi-square test to detect significant associations between SNP frequencies and BD incidence. Variants exhibiting a p-value of less than 10-5 were deemed significant and flagged for in-depth investigation to ascertain their potential as risk factor indicators.
3.3. Genetic Variants Identified, Statistical Analysis and Significance
According to our association study results, there were 11 genetic variants that demonstrated significant (p<10-5) associations with the condition (refer to the data points above the red line in the Manhattan plot in Figure 2). The Q-Q plot (Figure 2) shows significant deviations above the diagonal line, suggesting a potential link with BD. The genomic inflation factor, lambda, derived from the median of the observed and expected chi-square values was close to 1. This indicates minimal inflation and suggests that our test statistics are robust. Therefore, we were able to confidently interpret the results, affirming the minimal bias in the findings of our study.
The variants identified with high significance (specifically rs6427761, rs6658422, rs7530898, rs74419649, rs79221549, rs6496485, rs3829533, rs755981, rs6081464, rs7289613, and rs11156606) were distributed across various chromosomes. These SNPs were associated with several coding and non-coding genes, such as ABCD1, C20orf78, DPP10, LOC284930, LPP, MTHFSD, NR5A2, and non-coding RNA genes, including LINC01221 and NTRK3-AS1 (Table 2). Each variant demonstrated a high odds ratio (>1.5), indicating robust association with BD.
We conducted a search of the GWAS Catalog (https://www.ebi.ac.uk/gwas/) to further understand the novelty and relevance of these findings. Our search revealed that all variants except rs11156606 have not been previously reported to be associated with BD or any physical disease. Notably, rs11156606, located at the 153,741,041 bp position on the X chromosome, was recorded in the GWAS Catalog (accession ID GCST000821) as being associated with both BD and SCZ [22]. In our study, this variant showed a particularly high odds ratio of 2.36, suggesting a more than two-fold increase in the risk of BD in carriers of the allele.
The allele frequencies of the identified variants in our study population varied significantly, ranging from 15.7% to 33.2% in cases and 7.3% to 21.6% in controls. These variations highlight the distinct prevalence of genetic markers. Moreover, the allele frequencies in our study were closely aligned with those recorded in the Taiwan Biobank, which profiles the genetics of healthy Taiwanese individuals. This similarity suggests that the genetic makeup in our control group was representative of the general healthy population in Taiwan (Table 3).
Furthermore, a comparison with other global populations revealed that the allele frequencies in our control group were similar to those found in Asian populations. Notably, the frequencies of rs6496485, rs3829533, rs755987, and rs6081464 were higher in our study than in African, American, and European populations (Table 3). This suggests that these four variants may serve as specific biomarkers for BD in the Taiwanese population, offering the potential for more targeted diagnostic approaches.
3.4. Functional Annotations of Risk Genes
As shown in Table 4, we identified 64 significant Gene Ontology (GO) biological process terms with p-values less than 0.05. These terms are predominantly related to biological functions such as fatty acid metabolism, protein localization, potassium ion transport, vascular transport, and viral replication, which may be linked to BD. Notably, fatty acid metabolism emerged as a major significant GO term, particularly associated with the variant rs11156606 located in the ABCD1 gene.
Analysis of similar GO terms revealed three key cluster networks: (1) Fatty Acid Metabolism/Oxidative Stress Regulation, (2) Lipid Homeostasis and Membrane Potential Regulation, and (3) Localization of Lipid Transport and Membrane Proteins, as shown in Figure 3. Additionally, four ABCD1-related GO terms were identified with direct relevance to neuronal functions: GO:0106027 (neuron projection organization), GO:1990535 (neuron projection maintenance), GO:0043217 (myelin maintenance), and GO:0042552 (myelination). These findings suggest that mutations in ABCD1 play a crucial role in the functioning of the nervous system, potentially contributing to the pathology of BD and other neurological conditions.
The BP functions of the genes were identified using the GO database, with terms selected based on p-values < 0.05. The GO terms are listed in Table 4. Similarity networks were constructed using Navier–GO.
3.5. LD Analysis
The significant variant rs11156606, identified in the association study and located in intron 7 of ABCD1, was linked through LDproxy analysis (Figure 4)to a rare missense mutation, rs782720024 (c.436T>A, p.Phe146Ile), in exon 1. This mutation exhibits low allele frequencies, approximately 0.002, specifically within the EAS and Taiwanese populations, indicating its rarity in the Chinese population. This suggests that individuals in populations with this variant may have a higher risk of BD. Additionally, another SNP rs73640819 (c.131G>A), found in the 3' UTR with high LD values, plays a crucial role in gene expression regulation, including transcription, translation, RNA stability, and localization. Its allele frequency was notably the lowest in the Taiwanese population at 0.07 (Figure S1), highlighting population-specific genetic differences and their potential impact on disease susceptibility and gene regulation mechanisms.
Furthermore, LDproxy analysis revealed that the variant rs3829533, located in exon 6 of MTHFSD, is closely associated with two other SNPs, rs3751800 (D’=1, R2=0.987) and rs3751801 (D’=1, R2=1), based on data from the Chinese population. Notably, the associated SNPs located in MTHFSD exon 8 resulted in missense coding changes (rs3751800: c.727C>T/p. Arg243Cys and rs3751801: c.730G>C/p.Ala244Pro) (Supplemental Table S2 and Figure 5). (Figure 5) This finding is significant because it suggests that while the Taiwanese population may possess the synonymous variant rs3829533 at a relatively high frequency (0.238 in cases and 0.138 in controls, as shown in Table 2), which does not alter coding, the associated missense variants rs3751800 and rs3751801 could potentially modify the molecular function of MTHFSD owing to changes in amino acids, thereby increasing the risk of BD. Figure S2 illustrates that the allele frequencies of the two variants in the East Asian and Taiwanese populations (0.16 and 0.15, respectively) were higher than those in the African, American, and European populations.
Additionally, FORGEdb scores, as depicted in Figure 4 and Figure 5, predicted the likelihood of genetic variants functioning as regulatory elements. These scores range from 0 to 10, and are based on various regulatory DNA datasets, including transcription factor (TF) binding and chromatin accessibility. The FORGEdb scores for rs11156606, rs73640819, rs3829533, rs3751800, and rs3751801 are 7, 10, 7, 8, and 8, respectively. These high scores indicated a significant regulatory effect on ABCD1 and MTHFSD, further suggesting that these variants could be specific risk factors for BD in the Taiwanese population.
We investigated the nucleotide polymorphisms in the three variants within the MTHFSD gene (Figure 6). The variants rs3829533 and rs3751801 were synonymous; the former retained the amino acid threonine at position 175 of the MTHFSD protein translated from exon 6. Conversely, rs3751800 represented a missense variant, whereas its LD-associated variant rs3751801 induced amino acid substitutions at positions 243 and 244 of the MTHFSD protein (arginine to cysteine and alanine to proline, respectively). These changes occurred within exon 8 of MTHFSD.
The synonymous variant rs3829533 preserves the amino acid threonine at position 175 of the MTHFSD protein (translated from exon 6). In contrast, the LD-associated variants rs3751800 and rs3751801 result in amino acid substitutions at positions 243 and 244, respectively: arginine to cysteine and alanine to proline. These alterations occur within exon 8 of MTHFSD.
4. Discussion
This study represents an advancement in our understanding of the genetic underpinnings of BD, particularly in the Taiwanese population. By identifying 11 novel genetic variants associated with BD, our research not only contributes to the growing body of genomic knowledge, but also highlights the importance of conducting population-specific genetic studies. The discovery of these variants, most of which have not been previously reported in relation to BD, opens new avenues for understanding the complex genetic architecture of this condition. In particular, the high odds ratios observed for these variants underscore their potential role as robust biomarkers, offering insights for the development of more precise diagnostic tools and personalized treatment strategies. Furthermore, the alignment of allele frequencies with those in the Taiwan Biobank reinforces the relevance of our findings to Taiwanese demographics, providing a foundation for future studies to explore genetic risk factors and their implications in a targeted manner. BD is a complex disease with unknown causes, involving various factors such as demographics, genetics, and environment, some of which have strong evidence supporting their link to BD. Therefore, this study not only enriches our genetic understanding of BD but also underscores the critical need for tailored genetic research in diverse populations to uncover the nuanced nature of complex psychiatric disorders.
There is growing attention on the interaction between specific genes and the environment as well as the suggested involvement of fatty acid metabolism and neuronal function in the disorder [23]. The variant rs11156606, located intronically in ABCD1 gene, presents intriguing possibilities regarding its mechanistic role in BD. The protein encoded by this gene is a member of the ATP-binding cassette (ABC) transporter superfamily. ABC genes are divided into seven distinct subfamilies (ABC1, MDR/TAP, MRP, ALD, OABP, GCN20, White). This protein is a member of the ALD subfamily, which is involved in the peroxisomal import of fatty acids and/or fatty acyl-CoAs in organelles [24]. It also plays a crucial role in cellular processes such as membrane transport [25,26,27], circulatory system stability [28], lipid transport and homeostasis [28]. Dysregulation of ABC transporters has been implicated in various diseases, including Mendelian disorders [25], cancer, and drug resistance [27]. Several studies have indicated that fatty acid metabolism plays a significant role in BD development and manifestation. In this study, Leclercq S. et al. tested PUFA as potent inducers of the ABCD genes. The expression levels of ABCD2 and ABCD3 were significantly higher in n-3-deficient rats than in rats fed ALA- or DHA-supplemented diets, indicating sensitivity towards dietary PUFA [29]. In a study comparing patients with BD and healthy controls, subjects with BD had distinctly lower levels of omega-3 eicosapentaenoic acid (EPA) and higher omega-6 arachidonic acid levels, coupled with increased plasma IL-6 and TNF-α levels [30]. We established a connection between the genotype of the fatty acid desaturase gene cluster and altered polyunsaturated fatty acid levels in BD. In one study, fatty acids in the erythrocyte membranes of patients with bipolar manic disorder and healthy controls were analyzed using thin-layer chromatography and gas chromatography. The results showed lower levels of arachidonic acid (20:4n-6) and docosahexaenoic acid (22:6n-3) in BD patients with BD compared to normal controls [31]. The association of the ABCD1 variant rs11156606 with altered fatty acid metabolism in BD underscores its potential role in the pathophysiology of the condition. This link, highlighted by the distinct PUFA profiles in patients with BD, suggests that dysregulation of ABC transporters, which are key in lipid transport and homeostasis, could be a critical factor in BD development.
The discovery of rs11156606 within intron region 7 of ABCD1 via disease-gene association interpretation exemplifies the effectiveness of such research in identifying genetic variants that are potentially crucial for disease predisposition. This study combined data from people with SCZ and BD to find shared genetic factors, and revealed that SNP rs11156606 is associated with BD and SCZ [22] Notably, the unique presence of the rare allele rs782720024 in East Asian and Taiwanese populations, along with the discovery of rs73640819, which shows high LD and is situated at the 3’ UTR of ABCD1, underlines the significance of population-specific genetic studies. These findings reveal the intricate nature of gene regulation and its influence on disease, highlighting how allele frequency variations across populations, especially their scarcity in Taiwanese individuals, may shed light on the unique regulatory mechanisms that affect gene expression and disease outcomes.
The discovery of the high LD-associated variant rs73640819 in the 3' untranslated region (3’ UTR) of ABCD1 highlights its importance beyond a mere sequence tailing the coding region. The 3’-UTR plays a pivotal role in the regulation of gene expression by influencing mRNA stability, translation efficiency, and subcellular localization, thereby fine-tuning protein synthesis. This region is integral to the complex post-transcriptional gene regulation machinery and acts as a critical regulator of the pathway from DNA transcription to mRNA translation into proteins [32]. Such insights open avenues for further exploration of how variations in the 3’-UTR affect gene function and disease mechanisms, indicating a rich field for ongoing research.
The variant with c.131G>A changes introduces polyadenylation in the 3 ‘-UTR, highlighting the nuanced role of 3’-UTRs in genetic regulation. Unlike coding regions, 3’-UTRs are marked by AT-rich sequences that play a critical role in directing polyadenylation to these regions, thus influencing mRNA stability and cellular localization. This highlights the importance of 3’-UTRs in controlling gene expression through management of the mRNA lifecycle. Additionally, the diversity of mRNA isoforms attributed to alternative cleavage and polyadenylation (APA) allows for the production of multiple 3’ UTR variants from a single gene, each with unique regulatory capabilities [33]. This complexity underscores the significance of APA in the broader context of post-transcriptional regulation, opening paths for further investigation of its role in gene expression diversity and implications for cellular function and disease.
The Methenyltetrahydrofolate Synthetase Domain Containing (MTHFSD) gene plays a pivotal role in diverse physiological processes, including vascular health and cancer susceptibility, through its cytoplasmic activity in RNA binding, which facilitates crucial cellular functions [34]. It encodes a protein that is essential for the conversion of 5,10-methylenetetrahydrofolate to 5-methyltetrahydrofolate, a key step in the re-methylation of homocysteine to methionine, which is vital for cell health. Notably, the potential of S-adenosyl methionine (SAM) to alter lipid raft arrangements and influence receptor and transporter functions has been linked to therapeutic prospects in depression, BD, and SCZ [35]. Recent studies, including those by Lyu, highlight nutrient deficiencies, particularly in the conversion process from homocysteine to methionine, as is common in BD and SCZ [36], echoing Ozdogan et al.'s findings from measuring homocysteine levels in patients [37]. Animal models such as those of Akahoshi et al. suggest that a high-methionine diet could induce BD-like behaviors [38], pointing to a potential connection between high methionine intake and emotional states. Further research on MTHFSD expression identified it as a top differentially expressed gene in SCZ at various illness stages [39]. A 2016 study on methamphetamine-associated psychosis (MAP) identified MTHFSD as a candidate biomarker for RNA degradation [40], highlighting its role in folate catabolism and methionine metabolism. This connection is further supported by a meta-analysis by Hsieh et al., which found that individuals with BD generally have lower serum folate levels than healthy controls [41], underscoring the enzyme's role in maintaining active folate levels for vital biosynthetic pathways.
LD analysis revealed two correlated variants (rs3751800: c.727C>T/p. Arg243Cys and rs3751801: c.730G>C/p.Ala244Pro) within the MTHFSD gene, indicating its potential as a disease marker for BD in the Taiwanese population owing to its effect on MTHFSD function. These findings emphasize MTHFSD's critical involvement of MTHFSD in essential biological processes and their impact on human health, warranting further exploration of its mechanisms and therapeutic potential.
5. Conclusions
This study successfully identified novel genetic variants associated with BD in a Taiwanese Han population. These findings illuminate the intricate genetic basis of BD and highlight the significance of research on diverse populations. The discovered variants implicated pathways related to fatty acid metabolism, lipid homeostasis, and neuronal function, suggesting potential targets for future research. This study advances our understanding of BD's complex genetic architecture of BD, potentially paving the way for improved diagnostic and targeted therapeutic approaches.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1. LDproxy Analysis Results of rs11156606 in ABCD1 Two single nucleotide polymorphisms (SNPs) were identified: rs782720024 (missense variant; D' = 1, R² = 0.191) and rs73640819 (3'-UTR; D' = 1, R² = 0.929). Both SNPs exhibited strong linkage disequilibrium (LD). Table S2. LDproxy Analysis Results of rs3829533 in MTHFSD Sixteen SNPs were identified in high LD with both D' and R² values greater than 0.95. Among these, rs3751800 and rs3751801 are missense variants, resulting in amino acid substitutions in the encoded protein. Figure S1. Allele Frequency of rs73640819 in ABCD1 Allele frequencies were obtained from the 1000 Genomes Project and the Taiwan Biobank (Taiwanese population). Populations: AFR (African), AMR (American), EAS (East Asian), EUR (European). Figure S2. Allele Frequency of rs3751800 and rs3751801 in MTHFSD Allele frequencies were obtained from the 1000 Genomes Project and the Taiwan Biobank (Taiwanese population). Populations: AFR (African), AMR (American), EAS (East Asian), EUR (European).
Author Contributions
Conceptualization, Chin-Bin Yeh and Yi-Jen Hung; methodology, Wei-Chou Chang and Ajeet B Singh; data curation, Ta-Chuan Yeh; formal analysis, Chih-Chung Huang; supervision, Hsin-An Chang; writing—original draft preparation, Yi-Guang Wang and Wan-Ting Chen; writing—review and editing, Kuo-Sheng Hung. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The writing of this original article was approved by the Institutional Review Board of Tri-Service General Hospital, and all patients provided informed consent (IRB# 2-105-05-038).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author due to legal regulations of the database.
Acknowledgments
We extend our gratitude to all the participants and investigators involved in the Taiwan Precision Medicine Initiative. This study was supported by the Academia Sinica 40-05-GMM and AS-GC-110-MD02. We are also grateful to the Center for Precision Medicine and Genomics (CPMG) at Tri-Service General Hospital, National Defense Medical Center, for their invaluable assistance in gene information support (founding numbers: TSGH-B-112019, TSGH-B-112020, TSGH-B-113022). Additionally, we acknowledge PET-MRI support from the Department of Radiology at the Tri-Service General Hospital. Preliminary results are presented as posters at the 2023 50th Annual Military Medicine Symposium held in Taipei, Taiwan.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| APA | Polyadenylation |
| BD | Bipolar Disorder |
| DALYs | Disability-Adjusted Life Years |
| DSM | The Diagnostic And Statistical Manual of Mental Disorders |
| DSM-5 | DSM, Fifth Edition |
| EPA | Omega-3 Eicosapentaenoic Acid |
| GWAS | Genome-Wide Association Studies |
| ICD | The International Statistical Classification of Diseases and Related Health Problems |
| LD | Linkage Disequilibrium |
| MAP | Methamphetamine-Associated Psychosis |
| MDD | Major Depressive Disorder |
| MTHFSD | Methenyltetrahydrofolate Synthetase Domain Containing |
| SAM | S-Adenosyl Methionine |
| SCZ | Schizophrenia |
| SNP/SNPs | Single Nucleotide Polymorphism/Single Nucleotide Polymorphisms |
| TPM | Taiwan Precision Medicine |
| TPMI | Taiwan Precision Medicine Initiative |
| TSGH | Tri-Service General Hospital |
| UTR | Untranslated Region |
References
- Ketter, T.A. Diagnostic Features, Prevalence, and Impact of Bipolar Disorder. J. Clin. Psychiatry 2010, 71, e14. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Hu, C.; Ren, Z.; Bai, L.; Gao, F.; Lyu, J. Trends in the Incidence and DALYs of Bipolar Disorder at Global, Regional, and National Levels: Results from the Global Burden of Disease Study 2017. Journal of Psychiatric Research 2020, 125, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Ratheesh, A.; Davey, C.; Hetrick, S.; Alvarez-Jimenez, M.; Voutier, C.; Bechdolf, A.; McGorry, P.D.; Scott, J.; Berk, M.; Cotton, S.M. A Systematic Review and Meta-analysis of Prospective Transition from Major Depression to Bipolar Disorder. Acta Psychiatr Scand 2017, 135, 273–284. [Google Scholar] [CrossRef]
- Visscher, P.M.; Yengo, L.; Cox, N.J.; Wray, N.R. Discovery and Implications of Polygenicity of Common Diseases. Science 2021, 373, 1468–1473. [Google Scholar] [CrossRef]
- Leussis, M.P.; Berry-Scott, E.M.; Saito, M.; Jhuang, H.; de Haan, G.; Alkan, O.; Luce, C.J.; Madison, J.M.; Sklar, P.; Serre, T.; et al. The ANK3 Bipolar Disorder Gene Regulates Psychiatric-Related Behaviors That Are Modulated by Lithium and Stress. Biol Psychiatry 2013, 73, 683–690. [Google Scholar] [CrossRef]
- Schultz, C.C.; Mühleisen, T.W.; Nenadic, I.; Koch, K.; Wagner, G.; Schachtzabel, C.; Siedek, F.; Nöthen, M.M.; Rietschel, M.; Deufel, T.; et al. Common Variation in NCAN, a Risk Factor for Bipolar Disorder and Schizophrenia, Influences Local Cortical Folding in Schizophrenia. Psychol Med 2014, 44, 811–820. [Google Scholar] [CrossRef]
- Starnawska, A.; Demontis, D.; Pen, A.; Hedemand, A.; Nielsen, A.L.; Staunstrup, N.H.; Grove, J.; Als, T.D.; Jarram, A.; O’Brien, N.L.; et al. CACNA1C Hypermethylation Is Associated with Bipolar Disorder. Transl Psychiatry 2016, 6, e831. [Google Scholar] [CrossRef]
- A, H.; A, L.; J, T.; S, V.-K.; M, B.; S, S.; C, B.; L, P.; T, B.; G, B.; et al. The Risk Variant in ODZ4 for Bipolar Disorder Impacts on Amygdala Activation during Reward Processing. Bipolar disorders 2013, 15. [Google Scholar] [CrossRef]
- Chiou, Y.-J.; Huang, T.-L. Brain-Derived Neurotrophic Factor (BDNF) and Bipolar Disorder. Psychiatry Research 2019, 274, 395–399. [Google Scholar] [CrossRef]
- Ovadia, G.; Shifman, S. The Genetic Variation of RELN Expression in Schizophrenia and Bipolar Disorder. PLoS One 2011, 6, e19955. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am J Hum Genet 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, K. Genomic Variant Annotation and Prioritization with ANNOVAR and wANNOVAR. Nat Protoc 2015, 10, 1556–1566. [Google Scholar] [CrossRef] [PubMed]
- The 1000 Genomes Project Consortium; Corresponding authors; Auton, A. Abecasis, G.R.; Steering committee; Altshuler, D.M.; Durbin, R.M.; Abecasis, G.R.; Bentley, D.R.; Chakravarti, A.; et al. A Global Reference for Human Genetic Variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef]
- Gudmundsson, S.; Singer-Berk, M.; Watts, N.A.; Phu, W.; Goodrich, J.K.; Solomonson, M.; Genome Aggregation Database Consortium; Rehm, H. L.; MacArthur, D.G.; O’Donnell-Luria, A. Variant Interpretation Using Population Databases: Lessons from gnomAD. Human Mutation 2022, 43, 1012–1030. [Google Scholar] [CrossRef]
- Feng, Y.-C.A.; Chen, C.-Y.; Chen, T.-T.; Kuo, P.-H.; Hsu, Y.-H.; Yang, H.-I.; Chen, W.J.; Su, M.-W.; Chu, H.-W.; Shen, C.-Y.; et al. Taiwan Biobank: A Rich Biomedical Research Database of the Taiwanese Population. Cell Genomics 2022, 2, 100197. [Google Scholar] [CrossRef]
- Carbon, S.; Ireland, A.; Mungall, C.J.; Shu, S.; Marshall, B.; Lewis, S. the AmiGO Hub; the Web Presence Working Group AmiGO: Online Access to Ontology and Annotation Data. Bioinformatics 2009, 25, 288–289. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Current Protocols 2021, 1, e90. [Google Scholar] [CrossRef]
- Neale, B.M. Introduction to Linkage Disequilibrium, the HapMap, and Imputation. Cold Spring Harb Protoc 2010, 2010, pdb.top74. [Google Scholar] [CrossRef]
- Machiela, M.J.; Chanock, S.J. LDlink: A Web-Based Application for Exploring Population-Specific Haplotype Structure and Linking Correlated Alleles of Possible Functional Variants. Bioinformatics 2015, 31, 3555–3557. [Google Scholar] [CrossRef]
- Machiela, M.J.; Chanock, S.J. LDassoc: An Online Tool for Interactively Exploring Genome-Wide Association Study Results and Prioritizing Variants for Functional Investigation. Bioinformatics 2018, 34, 887–889. [Google Scholar] [CrossRef]
- Breeze, C.E.; Haugen, E.; Gutierrez-Arcelus, M.; Yao, X.; Teschendorff, A.; Beck, S.; Dunham, I.; Stamatoyannopoulos, J.; Franceschini, N.; Machiela, M.J.; et al. FORGEdb: A Tool for Identifying Candidate Functional Variants and Uncovering Target Genes and Mechanisms for Complex Diseases. Genome Biol 2024, 25, 3. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-S.; Liu, X.-F.; Aragam, N. A Genome-Wide Meta-Analysis Identifies Novel Loci Associated with Schizophrenia and Bipolar Disorder. Schizophrenia Research 2010, 124, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Rowland, T.A.; Marwaha, S. Epidemiology and Risk Factors for Bipolar Disorder. Therapeutic Advances in Psychopharmacology 2018, 8, 251–269. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-P.; Xu, D.; Wang, L.; Mao, Y.-X.; Li, Y.; Cheng, M.-T.; Zhou, C.-Z.; Hou, W.-T.; Chen, Y. Structural Basis of Substrate Recognition and Translocation by Human Very Long-Chain Fatty Acid Transporter ABCD1. Nat Commun 2022, 13, 3299. [Google Scholar] [CrossRef]
- Dean, M. The Genetics of ATP-Binding Cassette Transporters. In Methods in Enzymology; Elsevier, 2005; Vol. 400, pp. 409–429 ISBN 978-0-12-182805-9.
- Dean, M. ATP-binding Cassette (ABC) Transporter Supergene Family: Genetics and Evolution. In Encyclopedia of Life Sciences; Wiley, 2008 ISBN 978-0-470-01617-6.
- Wada, M. Role of ABC Transporters in Cancer Development and Malignant Alteration. YAKUGAKU ZASSHI 2022, 142, 1201–1225. [Google Scholar] [CrossRef]
- Salajegheh, A. Adenosine Triphosphate-Binding Cassette (ABC) Lipid Transporters. In Angiogenesis in Health, Disease and Malignancy; Springer International Publishing: Cham, 2016; ISBN 978-3-319-28138-4. [Google Scholar]
- Leclercq, S.; Skrzypski, J.; Courvoisier, A.; Gondcaille, C.; Bonnetain, F.; André, A.; Chardigny, J.-M.; Bellenger, S.; Bellenger, J.; Narce, M.; et al. Effect of Dietary Polyunsaturated Fatty Acids on the Expression of Peroxisomal ABC Transporters. Biochimie 2008, 90, 1602–1607. [Google Scholar] [CrossRef]
- Koga, N.; Ogura, J.; Yoshida, F.; Hattori, K.; Hori, H.; Aizawa, E.; Ishida, I.; Kunugi, H. Altered Polyunsaturated Fatty Acid Levels in Relation to Proinflammatory Cytokines, Fatty Acid Desaturase Genotype, and Diet in Bipolar Disorder. Transl Psychiatry 2019, 9, 208. [Google Scholar] [CrossRef]
- Chiu, C.-C.; Huang, S.-Y.; Su, K.-P.; Lu, M.-L.; Huang, M.-C.; Chen, C.-C.; Shen, W.W. Polyunsaturated Fatty Acid Deficit in Patients with Bipolar Mania. European Neuropsychopharmacology 2003, 13, 99–103. [Google Scholar] [CrossRef]
- Hong, D.; Jeong, S. 3’UTR Diversity: Expanding Repertoire of RNA Alterations in Human mRNAs. Molecules and Cells 2023, 46, 48–56. [Google Scholar] [CrossRef]
- Zhang, Q.; Tian, B. The Emerging Theme of 3′UTR mRNA Isoform Regulation in Reprogramming of Cell Metabolism. Biochemical Society Transactions 2023, 51, 1111–1119. [Google Scholar] [CrossRef]
- Castello, A.; Fischer, B.; Eichelbaum, K.; Horos, R.; Beckmann, B.M.; Strein, C.; Davey, N.E.; Humphreys, D.T.; Preiss, T.; Steinmetz, L.M.; et al. Insights into RNA Biology from an Atlas of Mammalian mRNA-Binding Proteins. Cell 2012, 149, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Karas Kuželički, N. S-Adenosyl Methionine in the Therapy of Depression and Other Psychiatric Disorders. Drug Development Research 2016, 77, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Lyu, N.; Xing, G.; Yang, J.; Zhu, X.; Zhao, X.; Zhang, L.; Wang, G. Comparison of Inflammatory, Nutrient, and Neurohormonal Indicators in Patients with Schizophrenia, Bipolar Disorder and Major Depressive Disorder. Journal of Psychiatric Research 2021, 137, 401–408. [Google Scholar] [CrossRef]
- Ozdogan, M.G.; Aydin, E.F.; Ustundag, M.F.; Ceyhun, H.A.; Oral, E.; Bakan, E. Homocysteine, Chronotype and Clinical Course in Bipolar Disorder Patients. Nordic Journal of Psychiatry 2020, 74, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Ishii, I.; Kamata, S.; Ito, S.; Shimonaga, A.; Koizumi, M.; Tsushima, M.; Miura, A.; Nagata, T.; Tosaka, Y.; Ohtani, H.; et al. A High-Methionine Diet for One-Week Induces a High Accumulation of Methionine in the Cerebrospinal Fluid and Confers Bipolar Disorder-like Behavior in Mice. IJMS 2022, 23, 928. [Google Scholar] [CrossRef]
- Narayan, S.; Tang, B.; Head, S.R.; Gilmartin, T.J.; Sutcliffe, J.G.; Dean, B.; Thomas, E.A. Molecular Profiles of Schizophrenia in the CNS at Different Stages of Illness. Brain Research 2008, 1239, 235–248. [Google Scholar] [CrossRef]
- Breen, M.S.; Uhlmann, A.; Nday, C.M.; Glatt, S.J.; Mitt, M.; Metsalpu, A.; Stein, D.J.; Illing, N. Candidate Gene Networks and Blood Biomarkers of Methamphetamine-Associated Psychosis: An Integrative RNA-Sequencing Report. Transl Psychiatry 2016, 6, e802–e802. [Google Scholar] [CrossRef]
- Hsieh, Y.-C.; Chou, L.-S.; Lin, C.-H.; Wu, H.-C.; Li, D.-J.; Tseng, P.-T. Serum Folate Levels in Bipolar Disorder: A Systematic Review and Meta-Analysis. BMC Psychiatry 2019, 19, 305. [Google Scholar] [CrossRef]
Figure 1.
Disease-Gene Association Analysis Pipeline Used in This Study.

Figure 2.
Manhattan and Q-Q Plots of Association Study Results in BD Patients. (A) A total of 280,177 variants were detected in TSGH participants through a chi-squared test. Eleven highly significant variants were selected based on a p-value < 10⁻⁵ (red spots above the blue dashed line): rs7538098, rs6427761, rs7538098, rs74419649, rs79221549, rs6496485, rs3829533, rs75581, rs6081464, rs7289613, and rs11156606. (B) The Q-Q (quantile-quantile) plot shows the quality of the association test. The observed distribution of low p-values (with high observed -log₁₀(P) values) indicates true associations with bipolar disorder. The genomic inflation factor (λ) was calculated using the median of the observed and expected chi-squared values.
Figure 2.
Manhattan and Q-Q Plots of Association Study Results in BD Patients. (A) A total of 280,177 variants were detected in TSGH participants through a chi-squared test. Eleven highly significant variants were selected based on a p-value < 10⁻⁵ (red spots above the blue dashed line): rs7538098, rs6427761, rs7538098, rs74419649, rs79221549, rs6496485, rs3829533, rs75581, rs6081464, rs7289613, and rs11156606. (B) The Q-Q (quantile-quantile) plot shows the quality of the association test. The observed distribution of low p-values (with high observed -log₁₀(P) values) indicates true associations with bipolar disorder. The genomic inflation factor (λ) was calculated using the median of the observed and expected chi-squared values.
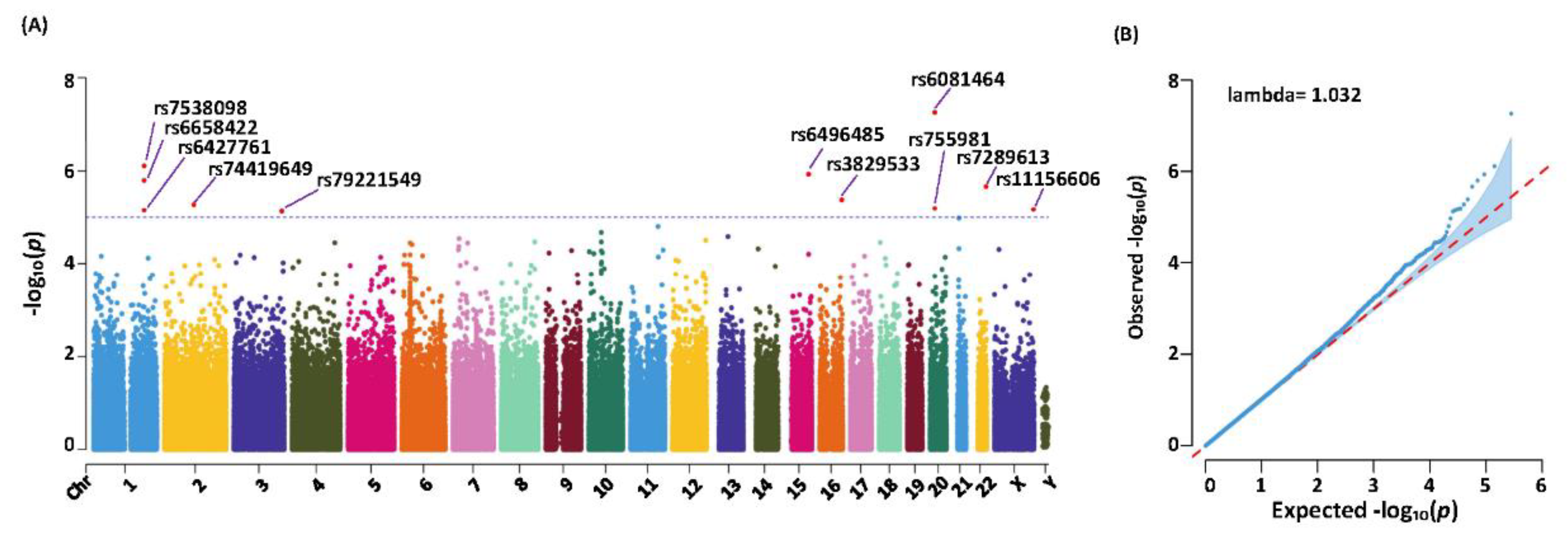
Figure 3.
Significant GO BP Term Similarity Networks of ABCD1 Analysis.
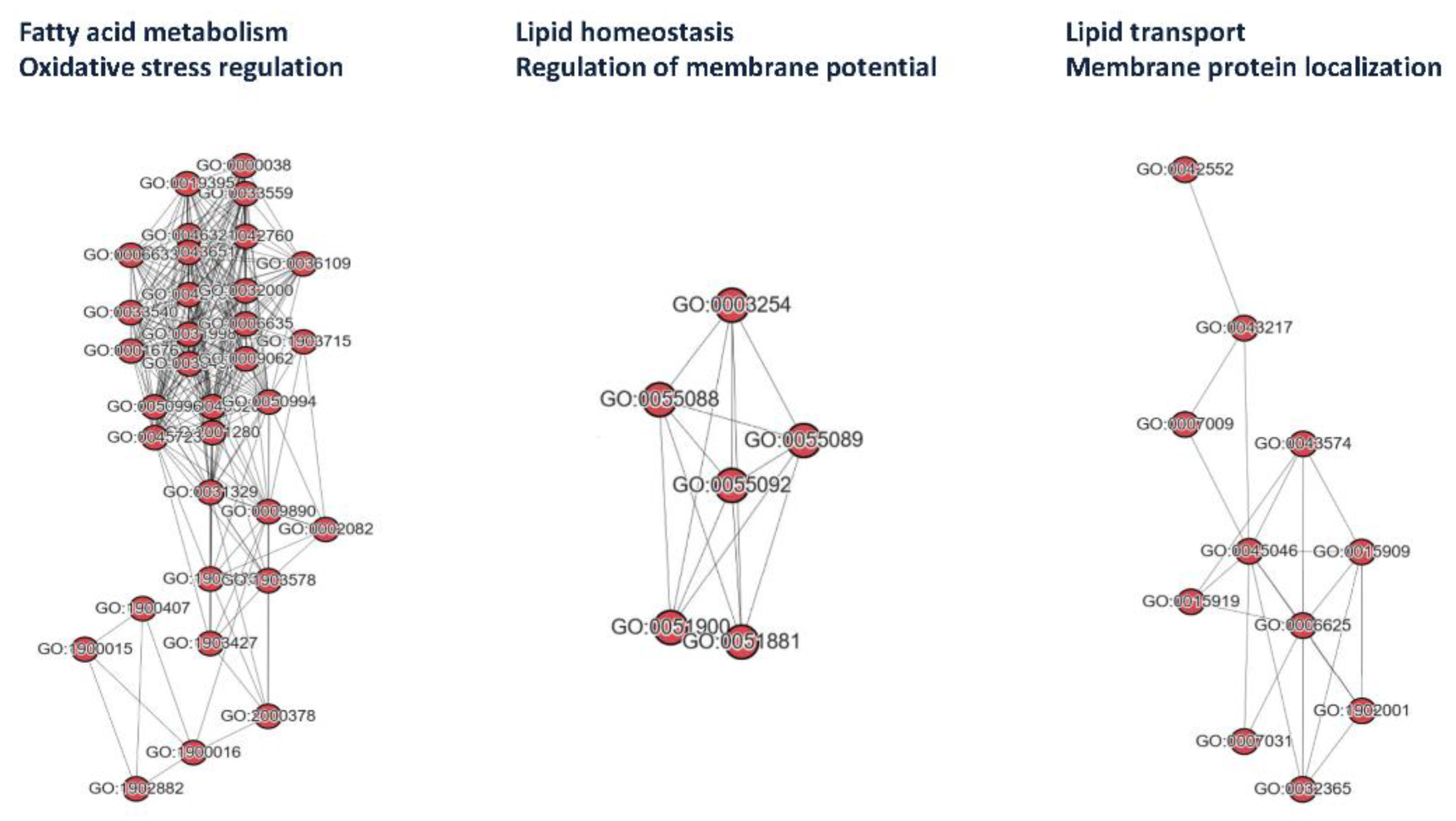
Figure 4.
LD Analysis of Variant rs11156606 on Chromosome X (152.96–153.06 Mb). (A) Overview of LD-associated variants within the ABCD1 gene region, highlighted between the orange dashed lines. The transcription direction is indicated by the arrow from 3' to 5' on Chromosome X. (B) Close-up of the genomic locations of rs11156606, rs782720024, and rs73640819. The missense variant rs782720024 is found in exon 1 of ABCD1, while rs73640819 is located in the non-coding 3' UTR region.
Figure 4.
LD Analysis of Variant rs11156606 on Chromosome X (152.96–153.06 Mb). (A) Overview of LD-associated variants within the ABCD1 gene region, highlighted between the orange dashed lines. The transcription direction is indicated by the arrow from 3' to 5' on Chromosome X. (B) Close-up of the genomic locations of rs11156606, rs782720024, and rs73640819. The missense variant rs782720024 is found in exon 1 of ABCD1, while rs73640819 is located in the non-coding 3' UTR region.
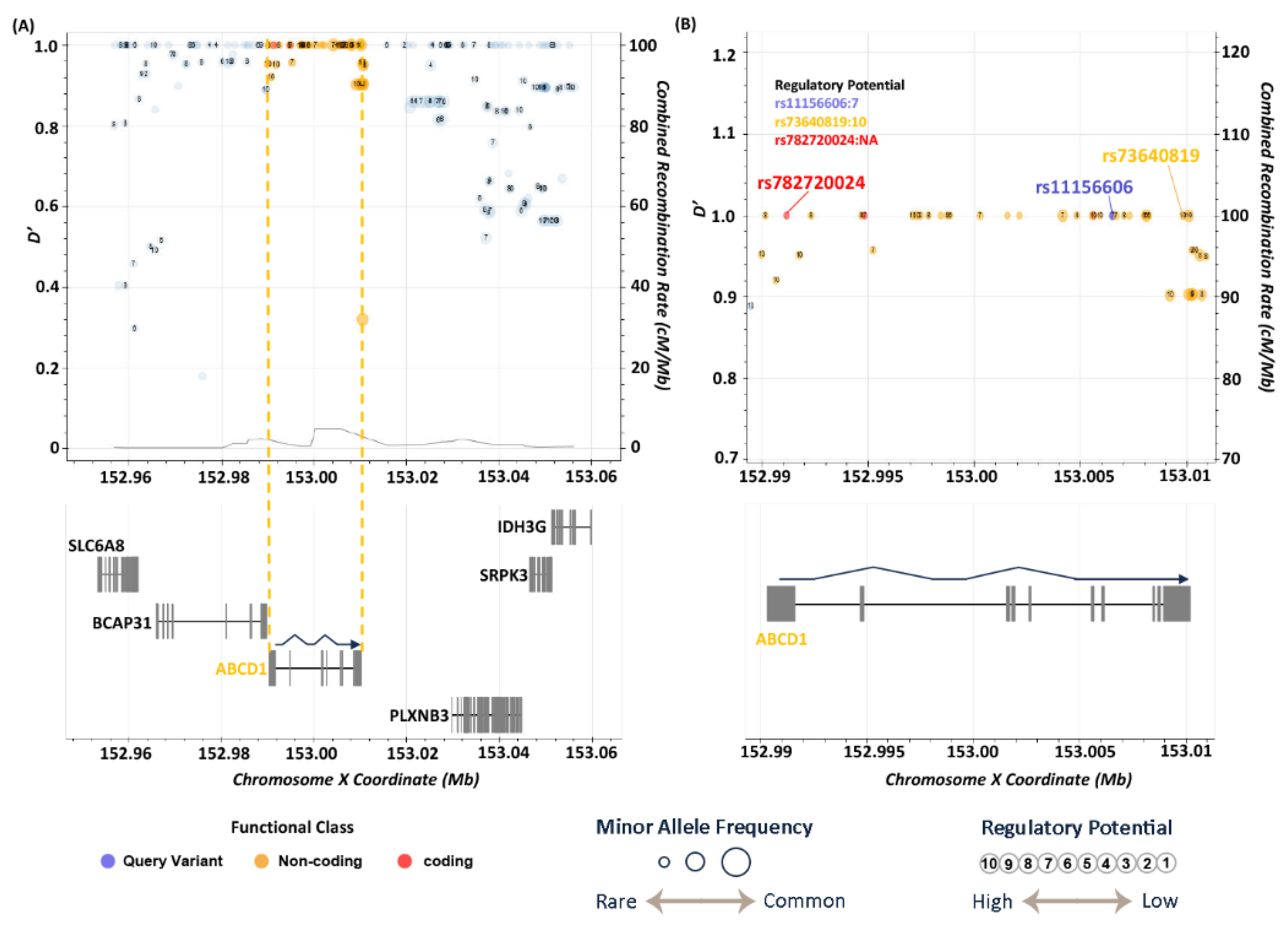
Figure 5.
LD Analysis of Variant rs3829533 on Chromosome 16 (86.45–86.65 Mb). (A) Overview of LD-associated variants within the MTHFSD gene region, including its promoter sequence (light gray box). The gene region is marked between the orange dashed lines, and the arrow indicates the transcription direction from 3' to 5' on Chromosome 16. (B) Close-up of the locations of rs3829533, rs3751800, and rs3751801. The missense variants rs3751800 and rs3751801 are located in exon 8 of MTHFSD.
Figure 5.
LD Analysis of Variant rs3829533 on Chromosome 16 (86.45–86.65 Mb). (A) Overview of LD-associated variants within the MTHFSD gene region, including its promoter sequence (light gray box). The gene region is marked between the orange dashed lines, and the arrow indicates the transcription direction from 3' to 5' on Chromosome 16. (B) Close-up of the locations of rs3829533, rs3751800, and rs3751801. The missense variants rs3751800 and rs3751801 are located in exon 8 of MTHFSD.
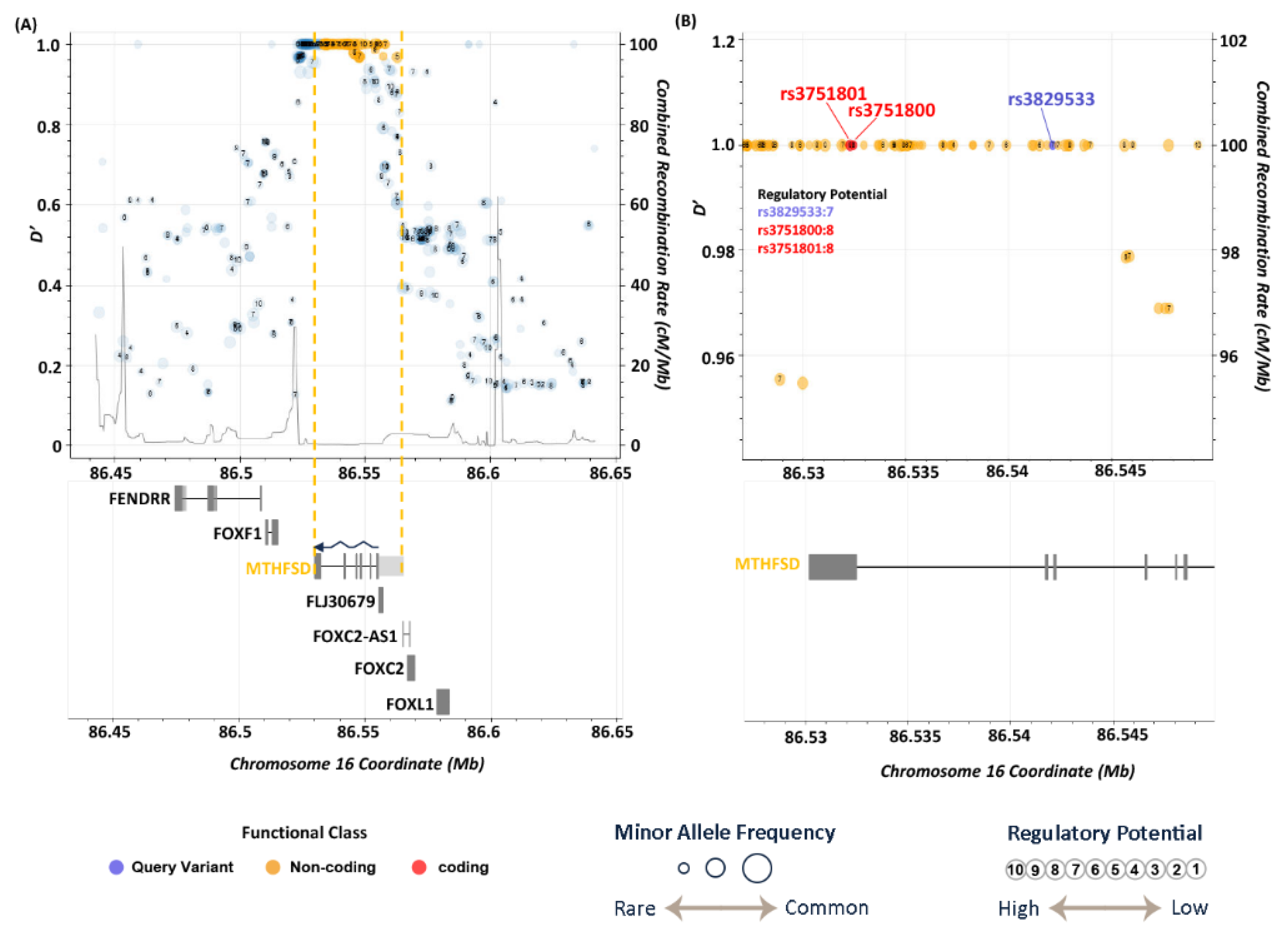
Figure 6.
Coding Changes of rs3829533, rs3751800, and rs3751801 in the MTHFSD Gene.
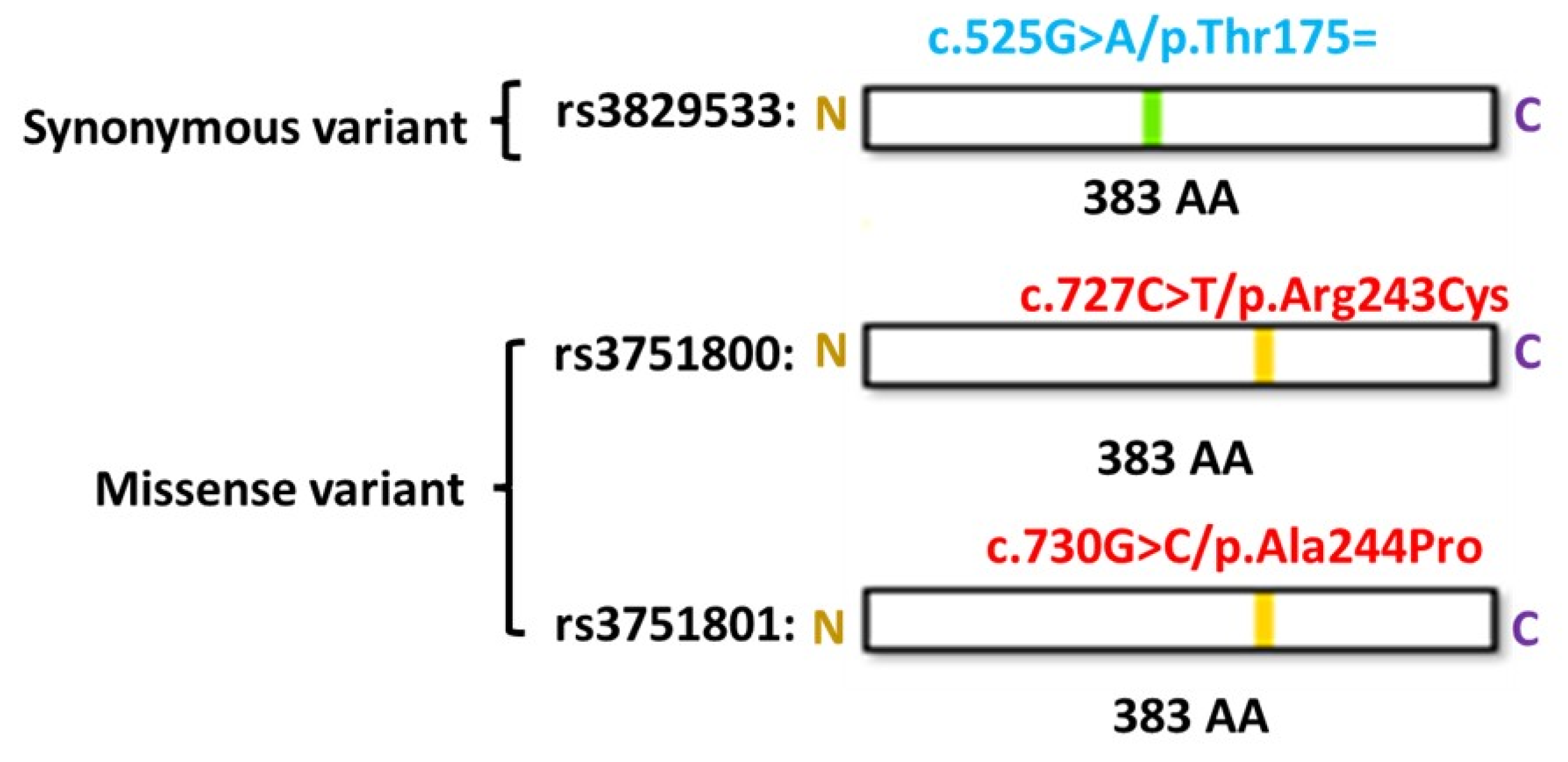

Table 1.
Participant Information in This Study.
| Table 1. Participant Information in This Study | ||||
|---|---|---|---|---|
| Group | BD | Control | p-value | |
| Sex | Male | 59 | 11793 | 0.63 |
| Female | 69 | 14329 | ||
| Age (Average ± SD) |
>90 | 0 | 197 (93 ± 2) | - |
| 80-89 | 1 (81 ± 0) | 895 (83 ± 3) | - | |
| 70-79 | 7 (73 ± 3) | 2970 (73 ± 3) | 0.86 | |
| 60-69 | 25 (64 ± 3) | 5681 (64 ± 3) | 0.26 | |
| 50-59 | 28 (54 ± 3) | 5124 (55 ± 3) | 0.54 | |
| 40-49 | 25 (44 ± 3) | 4096 (44 ± 3) | 0.16 | |
| 30-39 | 24 (35 ± 3) | 3424 (35 ± 3) | 0.21 | |
| 20-29 | 15 (25 ± 3) | 3002 (25 ± 3) | 0.56 | |
| 10-19 | 3 (15 ± 2) | 586 (15 ± 3) | 0.07 | |
| <9 | 0 | 147 (8 ± 1) | - | |
Table 2.
Variants Obtained from the Association Study Results with High Significance.
| Table 2. Variants Obtained from the Association Study Results with High Significance | ||||||||
|---|---|---|---|---|---|---|---|---|
| CHR | Position | SNP ID | Refa | Altb | p-valuec | Odds Ratio | Region | Relative Gene |
| 1 | 199219779 | rs6427761 | A | G | 1.56E-06 | 2.02 | intergenic | LINC01221;NR5A2 |
| 1 | 199223240 | rs6658422 | C | T | 7.01E-06 | 1.96 | intergenic | LINC01221;NR5A2 |
| 1 | 199262608 | rs7538098 | T | G | 7.63E-07 | 2.08 | intergenic | LINC01221;NR5A2 |
| 2 | 114648288 | rs74419649 | T | C | 5.17E-06 | 1.88 | intronic | DPP10 |
| 3 | 188587806 | rs79221549 | C | T | 7.47E-06 | 2.09 | intronic | LPP |
| 15 | 88454801 | rs6496485 | G | A | 1.18E-06 | 2.01 | intergenic | NTRK3-AS1;MRPL46 |
| 16 | 86542131 | rs3829533 | C | T | 4.06E-06 | 1.95 | exonic | MTHFSD |
| 20 | 19027381 | rs755981 | G | T | 6.48E-06 | 1.81 | intergenic | C20orf78;SLC24A3 |
| 20 | 19031000 | rs6081464 | T | C | 5.41E-08 | 2.09 | intergenic | C20orf78;SLC24A3 |
| 22 | 480s87568 | rs7289613 | C | T | 2.14E-06 | 2.06 | intergenic | LOC284930;MIR3201 |
| X | 153741041 | rs11156606 | A | C | 6.69E-06 | 2.36 | intronic | ABCD1 |
|
a Allele in control. b Allele in case. c Filter by p < 10-5 | ||||||||
Table 3.
Allele Frequencies of Variants from Other Genetic Projects.
| Table 3. Allele Frequencies of Variants from Other Genetic Projects | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| This study | TPMIa | TWBankb | 1000gc | gnomADd | ||||||||||
| SNP ID | Alt | Case | Control | - | - | AFRe | AMR | EAS | EUR | AFR | AMR | EAS | FIN | NFE |
| rs6427761 | G | 0.231 | 0.129 | 0.134 | 0.134 | 0.200 | 0.098 | 0.160 | 0.022 | 0.195 | 0.087 | 0.123 | 0.044 | 0.025 |
| rs6658422 | T | 0.215 | 0.122 | 0.127 | 0.127 | 0.290 | 0.100 | 0.150 | 0.047 | 0.249 | 0.101 | 0.115 | 0.089 | 0.043 |
| rs7538098 | G | 0.224 | 0.122 | 0.127 | 0.126 | 0.290 | 0.099 | 0.150 | 0.041 | 0.268 | 0.086 | 0.118 | 0.076 | 0.044 |
| rs74419649 | C | 0.273 | 0.167 | 0.168 | 0.157 | 0.011 | 0.260 | 0.180 | 0.110 | 0.021 | 0.318 | 0.160 | 0.213 | 0.133 |
| rs79221549 | T | 0.169 | 0.089 | 0.089 | 0.084 | 0.002 | 0.037 | 0.091 | 0.033 | 0.006 | 0.039 | 0.084 | 0.038 | 0.028 |
| rs6496485 | A | 0.242 | 0.137 | 0.137 | 0.131 | 0.076 | 0.085 | 0.140 | 0.076 | 0.091 | 0.076 | 0.138 | 0.074 | 0.084 |
| rs3829533 | T | 0.238 | 0.138 | 0.138 | 0.150 | 0.008 | 0.110 | 0.160 | 0.110 | 0.023 | 0.106 | 0.144 | 0.102 | 0.092 |
| rs755981 | T | 0.332 | 0.216 | 0.220 | 0.205 | 0.550 | 0.420 | 0.210 | 0.430 | 0.543 | 0.370 | 0.213 | 0.561 | 0.470 |
| rs6081464 | C | 0.285 | 0.160 | 0.163 | 0.152 | 0.022 | 0.110 | 0.160 | 0.130 | 0.039 | 0.075 | 0.158 | 0.140 | 0.127 |
| rs7289613 | T | 0.203 | 0.110 | 0.113 | 0.120 | 0.160 | 0.069 | 0.095 | 0.098 | 0.168 | 0.041 | 0.121 | 0.133 | 0.096 |
| rs11156606 | C | 0.157 | 0.073 | 0.075 | 0.081 | 0.698 | 0.135 | 0.099 | 0.115 | 0.640 | 0.131 | 0.087 | 0.110 | 0.119 |
|
a Taiwan Precision Medicine Initiative. b Taiwan Biobank. c 1000 Genomes global minor allele frequency. d gnomAD (genomes) allele frequencies. e AFR: African. AMR: American. EAS: East Asian. EUR: European. FIN: Finnish. NFE: Non-Finnish European. | ||||||||||||||
Table 4.
GO annotation of variant genes.
| Table 4. GO annotation of variant genes a | |||
|---|---|---|---|
| GO_ID | Term | p-value | Gene |
| GO:0042758 | long-chain fatty acid catabolic process | 0.003 | ABCD1 |
| GO:0042760 | very long-chain fatty acid catabolic process | 0.004 | ABCD1 |
| GO:2001280 | positive regulation of unsaturated fatty acid biosynthetic process | 0.004 | ABCD1 |
| GO:0032000 | positive regulation of fatty acid beta-oxidation | 0.005 | ABCD1 |
| GO:0043574 | peroxisomal transport | 0.005 | ABCD1 |
| GO:1900407 | regulation of cellular response to oxidative stress | 0.005 | ABCD1 |
| GO:0043217 | myelin maintenance | 0.006 | ABCD1 |
| GO:1990535 | neuron projection maintenance | 0.006 | ABCD1 |
| GO:0046321 | positive regulation of fatty acid oxidation | 0.007 | ABCD1 |
| GO:0051900 | regulation of mitochondrial depolarization | 0.007 | ABCD1 |
| GO:0055089 | fatty acid homeostasis | 0.007 | ABCD1 |
| GO:1902882 | regulation of response to oxidative stress | 0.007 | ABCD1 |
| GO:0030497 | fatty acid elongation | 0.008 | ABCD1 |
| GO:0031998 | regulation of fatty acid beta-oxidation | 0.008 | ABCD1 |
| GO:0036109 | alpha-linolenic acid metabolic process | 0.008 | ABCD1 |
| GO:0045723 | positive regulation of fatty acid biosynthetic process | 0.008 | ABCD1 |
| GO:1903427 | negative regulation of reactive oxygen species biosynthetic process | 0.008 | ABCD1 |
| GO:0033540 | fatty acid beta-oxidation using acyl-CoA oxidase | 0.009 | ABCD1 |
| GO:1903715 | regulation of aerobic respiration | 0.009 | ABCD1 |
| GO:0002082 | regulation of oxidative phosphorylation | 0.010 | ABCD1 |
| GO:0009792 | embryo development ending in birth or egg hatching | 0.010 | NR5A2 |
| GO:0046320 | regulation of fatty acid oxidation | 0.010 | ABCD1 |
| GO:0106027 | neuron projection organization | 0.010 | ABCD1 |
| GO:0003254 | regulation of membrane depolarization | 0.011 | ABCD1 |
| GO:0009890 | negative regulation of biosynthetic process | 0.011 | ABCD1 |
| GO:1900016 | negative regulation of cytokine production involved in inflammatory response | 0.011 | ABCD1 |
| GO:1902001 | fatty acid transmembrane transport | 0.011 | ABCD1 |
| GO:0032365 | intracellular lipid transport | 0.013 | ABCD1 |
| GO:0043651 | linoleic acid metabolic process | 0.013 | ABCD1 |
| GO:0045046 | protein import into peroxisome membrane | 0.013 | ABCD1 |
| GO:0050994 | regulation of lipid catabolic process | 0.013 | ABCD1 |
| GO:0050996 | positive regulation of lipid catabolic process | 0.013 | ABCD1 |
| GO:2000378 | negative regulation of reactive oxygen species metabolic process | 0.014 | ABCD1 |
| GO:0007031 | peroxisome organization | 0.015 | ABCD1 |
| GO:0015919 | peroxisomal membrane transport | 0.016 | ABCD1 |
| GO:0006625 | protein targeting to peroxisome | 0.017 | ABCD1 |
| GO:0045070 | positive regulation of viral genome replication | 0.017 | NR5A2 |
| GO:1903426 | regulation of reactive oxygen species biosynthetic process | 0.017 | ABCD1 |
| GO:0043266 | regulation of potassium ion transport | 0.018 | DPP10 |
| GO:1904062 | regulation of cation transmembrane transport | 0.018 | DPP10 |
| GO:0015909 | long-chain fatty acid transport | 0.019 | ABCD1 |
| GO:0000038 | very long-chain fatty acid metabolic process | 0.020 | ABCD1 |
| GO:0051881 | regulation of mitochondrial membrane potential | 0.024 | ABCD1 |
| GO:1900015 | regulation of cytokine production involved in inflammatory response | 0.026 | ABCD1 |
| GO:0042552 | myelination | 0.028 | ABCD1 |
| GO:0006635 | fatty acid beta-oxidation | 0.031 | ABCD1 |
| GO:1903078 | positive regulation of protein localization to plasma membrane | 0.031 | DPP10 |
| GO:1904377 | positive regulation of protein localization to cell periphery | 0.031 | DPP10 |
| GO:0033559 | unsaturated fatty acid metabolic process | 0.032 | ABCD1 |
| GO:1901379 | regulation of potassium ion transmembrane transport | 0.032 | DPP10 |
| GO:1903578 | regulation of ATP metabolic process | 0.033 | ABCD1 |
| GO:0019395 | fatty acid oxidation | 0.035 | ABCD1 |
| GO:0048524 | positive regulation of viral process | 0.037 | NR5A2 |
| GO:0007009 | plasma membrane organization | 0.038 | ABCD1 |
| GO:0031329 | regulation of cellular catabolic process | 0.038 | ABCD1 |
| GO:0055088 | lipid homeostasis | 0.038 | ABCD1 |
| GO:0045069 | regulation of viral genome replication | 0.039 | NR5A2 |
| GO:0009062 | fatty acid catabolic process | 0.041 | ABCD1 |
| GO:0006633 | fatty acid biosynthetic process | 0.042 | ABCD1 |
| GO:0055092 | sterol homeostasis | 0.042 | ABCD1 |
| GO:1903076 | regulation of protein localization to plasma membrane | 0.047 | DPP10 |
| GO:0001676 | long-chain fatty acid metabolic process | 0.049 | ABCD1 |
| GO:0010232 | vascular transport | 0.049 | SLC24A3 |
| GO:0150104 | transport across blood-brain barrier | 0.050 | SLC24A3 |
| aSelected based on p-value < 0.05. | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



