
Submitted:
12 February 2025
Posted:
13 February 2025
You are already at the latest version
Abstract
The major cause for blindness worldwide is age-related macular degeneration (AMD), and its incidence and prevalence are rising. There is no treatment for the early or atrophic forms of AMD. In terms of pathogenesis, the complex interaction of metabolic, functional, genetic, and environmental factors creates a stage for chronically developing changes in ocular structures of the macular region which contribute to varying degrees to the onset and final picture of AMD. Oxidative stress is a constant condition of the retina. Brown seaweed contains bioactive compounds with promising activities that make them interesting for potential use in AMD, such as the polysaccharide fucoidan, the carotenoid fucoxanthin and fucoxanthinol. Considering fucoidans, their heterogeneity requires intensive research to find suitable fucoidans for further applications. Fucoxanthin and its metabolite, fucoxanthinol, have shown antioxidant activities attributed to free radical scavenging and singlet oxygen inactivation in vitro. Fucoxanthin has greater antioxidant activity and is considered a potential protective agent against oxidative stress. This review gives an overview of the recent studies regarding their potential and challenges.
Keywords:
fucoidan
; age-related macular degeneration
; oxidative stress
; fucoxanthin Laminaria hyperborea
; Saccharina latissima
1. Introduction
1.1. Age-Related Macular Degeneration (AMD)
A major cause for blindness worldwide is age-related macular degeneration (AMD), and its incidence and prevalence are rising [1]. Globally, AMD ranks third as a cause of blindness after cataract and glaucoma. Most of the affected individuals live in developed countries. Wang et al. (2022) carried out a study about the global incidence, progression and risks factors of AMD and made projection statistics for 30 years. As a result, their estimation was 32.44 million of AMD´s patients by 2030, rising to 39.05 million by 2050. As well, they consider that Asia, Europe, and Northern America have and will have the highest number of patients [2]. In general, advanced AMD is rare before the age of 55, and more common in people of 75 years and older. The prevalence of neovascular AMD and geographic atrophy appears to vary in different ethnic and racial groups throughout the world. The prevalence of advanced AMD increases with each decade after the age of 50 with the highest prevalence occurring after the age of 80 [3]. Due to longer life expectancy and general aging of populations, maculopathies and in particular AMD have markedly increased in their importance as causes of irreversible visual impairment and blindness within the past 2 decades [4,5].
AMD is a common, chronic, progressive degenerative disorder of the macula features loss of central vision because of abnormalities in the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choroid complex often resulting in geographic atrophy and/or neovascularization [6]. Advanced AMD can be classified broadly into two types: dry and wet. Although dry AMD accounts for most of all diagnosed cases, wet AMD is responsible for most of the severe vision loss, and it usually occurs from weeks to months [7]. Eye anatomy is depicted in Figure 1.
Although neovascularization has been the most common cause of severe vision loss, geographic atrophy, the most advanced form of dry AMD, can cause a significant loss of vision as well [8]. This central vision deficiency is illustrated through Figure 2.
AMD is a multifactorial disorder, with dysregulation in the complement, lipid, angiogenic, inflammatory, and extracellular matrix pathways implicated in its pathogenesis. More than 50 genetic susceptibility loci have been identified, of which the most important are in the CFH and ARMS2 genes [9]. The major non-genetic risk factors are smoking and low dietary intake of antioxidants (zinc and carotenoids). Progression from early-stage to late-stage disease can be slowed with high-dose zinc and antioxidant vitamin supplements [1].
In terms of pathogenesis, the complex interaction of metabolic, functional, genetic, and environmental factors creates a stage for chronically developing changes in ocular structures of the macular region which contribute to varying degrees to the onset and final picture of AMD [7].
A major factor in the development of AMD is oxidative stress [3,4]. Oxidative stress can be classified in basal (positive), low, intermediate, and high intensity; these can happen in acute form, chronic and repetitive states. Moreover, some forms of oxidative stress have been classified: physiological (eustress), nutritional, dietary, glycol-oxidative stress, proteotoxic stress, disulfide stress, photooxidative stress (UV-A, UV-B, infrared-A), radiation induced, nitrosative, nanoparticle-induced oxidative stress and environmental stress (related to sulfur and nitrogen dioxide and ozone) [10]. The oxidative cell damage can be displayed in Figure 3.
The retina is under constant exposure to light, while functioning in a surrounding with high oxygen tension because of the high blood supply in this tissue. In addition to the high amount of hydrogen peroxide produced in the RPE, this leads to a strong oxidative burden and during the cause of aging to RPE degeneration [11]. Another factor that has been implicated in the development of AMD is chronic inflammation [12,13]. Furthermore, in the exudative form of AMD, where vessels grow out of the choroid under and/or into the retina, the cytokine Vascular endothelial growth factor (VEGF) is a major pathogenic factor [14]. Several factors, such as hypoxia, inflammation or oxidative stress, can induce the increased secretion of VEGF, primarily by the RPE [15,16,17]. Enhanced VEGF secretion by the RPE may lead to the growth of pathological vessels from the choroid. Indeed, the only available therapy for AMD is the use of VEGF-antagonists for the exudative form of the disease [14].
There is no treatment for the early forms of AMD, and the treatment options for atrophic AMD are very limited, with complement inhibitors being approved in the US which show modest effect by slowing down the progression of the atrophy [18], so the demands of new treatment options are high. Progress is likely to be from improved efforts in prevention and risk-factor modification, personalized medicine targeting specific pathways, newer anti-vascular endothelial growth factor agents or other agents, and regenerative therapies [1,3].
1.2. Brown Algae
An interesting source for potential new therapies for AMD could be brown algae. The brown algae or Phaeophyta comprise around 3000 species, with about 250 genera and between 1500-2000 mainly marine species. Within the genus the most prominent is laminaria, and within the Fucales species, fucus vesiculosus [19]. The brown coloration, of a highly variable hue, is due to the presence of certain carotenoid pigments known as fucoxanthins which is a special xanthophyll derived from marine algae, has attracted more and more attention due to its various biological functions. However, reports on its ocular benefits are still limited, but in vitro trials suggest that fucoxanthin could be beneficial for the prevention of retinal lesions induced by visible light [20]. The interest in fucoxanthins and fucoidans as potential therapeutics for AMD is rather recent, with a hypothesis paper published in 2016. It was because it has various biological properties such as antioxidants, anticancer, antiangiogenic, anti-inflammatory, and has cytoprotective and neuroprotector effects [19]. Our working group is focused on those therapeutic alternatives with emphasis on the oxidative stress path and ROS impairment on the physiological functions of RPE cells like natural resources such as marine bioactive compounds.
2. Bioactive Compounds
Brown algae contain many bioactive compounds such as carotenoids and polyphenols that have diverse biological activities, including antioxidant, anticancer, anti-inflammatory, UV radiation protection, etc. There are many experimental works that have identified many bioactive compounds, among the most important are polysaccharides, proteins, unsaturated fatty acids, polyphenolic compounds, terpenes, and pigments [20,21]. One of the pigments that has been attracting the attention of researchers lately due to their biological activity is fucoxanthin and its metabolite fucoxanthinol [22,23] . Brown algae mainly synthesize the polysaccharides laminarin, cellulose, alginate, and fucoidan. A potentially versatile therapeutic component of fucose-containing sulfated polysaccharides are fucidans which play a structural component of the cell wall [19].
2.1. Fucoidan
Marine polysaccharides are classified as sulfated (SPs) and nonsulfated macromolecules that are mainly derived from micro- and macroalgae. In particular, macroalgal SPs show more diverse chemical characteristics than nonsulfated analogues, regarding their molecular weight, monosaccharide composition, and sulfate content and position, which interact with various biological targets at different levels leading to diverse and promising pharmacological activities. Depending on the algae species the purity of the extracted crude fucoidans was shown to differ and this characteristic turned out as the most important one for changes in the viability of the used cell lines. Also, other parameters such as the molecular weights are generally considered important for the biological activities of fucoidans [24].
They are found in different phyla, such as in Phaeophyta or brown algae. Despite brown algae, with 1,800 species, not being the most abundant class, their SP fucoidans show more potential applications in different areas than those isolated from ulvan and carrageenan [25,26]. Fucoidans are known as fucose-containing sulfated polysaccharides (FCSPs) where l-fucose always predominates other sugar monomers, such as galactose, mannose, glucose, and uronic acids. In general, it is known that the fucoidan skeleton is composed of α(1,3)-linked residues of L-fucopyranose or alternations of α(1,3) and α(1,4) L-fucopyranoses. Its structure also includes acetate and sulfate groups distributed in the C-2 and C-4 positions of the chain ( (Figure 4). L-fucose may exceed 90% of the total sugar composition of fucoidans. FCSP are a very heterogeneous group of polysaccharides with strong variations in sugar composition, degree of branching and sulfation, and molecular weight. Their structure depends on different aspects, like species, harvest time, place, and extraction method [27]. A simple extraction method comparison is shown in Figure 5.
Few years ago, several fucoidans’ structures have been solved, and many aspects of their biological activity have been described such as: anticoagulant, antithrombotic activity, antivirus activity, antitumor, immunomodulatory activity, reducing blood lipids, anticomplementary activity, therapeutic potential in surgery, anti-inflammatory, gastric protection, against hepatopathy, against uropathy and renalpathy and antioxidant activity [19].
Lots of studies show that fucoidan presents significant antioxidant activity in experiments in vitro. For example, Marudhupandi et al. 2014 [28] studied the evaluation of potential antioxidant activity in vitro of fucoidan, from the brown seaweed Sargassum tenerrimum, where they demonstrated that the antioxidant property of fucoidans depended on the sulfated fraction and they demonstrated that fucoidan does not have toxic effects, which ensures that fucoidan can be used as natural antioxidants in the treatment of many human diseases such as age-related macular degeneration. Another study by Huang et al. 2015 [29] also evaluated the in vitro antioxidant activity of fucoidans from Sargassum glaucescens and demonstrated the potential activity antioxidants and colon cancer inhibition.
Recently Zayet et al mentioned that the multiple biological activities of fucoidan are mediated by its interaction with various proteins and signaling pathways, including NF-κB, MAPKs, TLRs, transforming growth factor Beta (TGF-β), nuclear factor erythroid 2-related factor 2 (Nrf2), and others. In the case of Nrf2, it is a basic leucine zipper transcription factor made up of a cap ‘n’ collar and encoded by the gene NFE2L2. After activation, Nrf2, through the antioxidant-response element (ARE), upregulates the expression of antioxidant enzymes such as heme oxygenase 1 (HO-1), catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GPx). These antioxidant enzymes and molecules in turn mitigate the detrimental effects of oxidative stress on nucleic acids, proteins, and/or plasma lipids [30]. Fucoidan is an excellent natural antioxidant and has great potential for preventing free radical-mediated diseases. Fucoidan from L. japonica can prevent the increase of lipid peroxide (LPO) in serum, liver, and spleen of diabetic mice obviously, as well as being capable of interfering with VEGF and blood lipids [19].
Evaluations have been made to investigate the effects of fucoidans on the viability of cells. An important paper compares effect of fucoidans on tumor and non-tumor cell line, including ocular cells lines, using fucoidans extracted from different brown algal species such as Fucus vesiculosus, F. serratus, F. distichus subsp. evanescens, Dictyosiphon foeniculaceus, Laminaria digitata and Saccharina latissima. Analysis of the correlation between the cell viability and various characteristics of the six different fucoidans suggest that highly pure fucoidans do not impair the cell viability [24].
Considering fucoidans are very promising marine polysaccharides for possible new treatment development for AMD, the heterogeneity of fucoidans requires more research to find suitable species and extraction methods. Different fucoidan samples extracted enzymatically from Laminaria digitata (LD), Saccharina latissima (SL) and Fucus distichus subsp. evanescens (FE) were tested by Philipp Dörschmann et al for toxicity, oxidative stress protection and VEGF (vascular endothelial growth factor) inhibition.
The report showed that crude fucoidans were extracted from seaweed using different enzymes and SL fucoidans were further separated into three fractions (SL_F1-F3) by ion-exchange chromatography (IEX). As results, they were the most effective fucoidans regarding VEGF inhibition, which correlated with a high fucose and low alginate content. SL fucoidans, which were treated with SAT2 enzymes and further processed with ion-exchange chromatography, are the most promising extracts for a potential application in AMD [31]. The Philipp Dörschmann study group showed that the fucoidans from Saccharina latissima displayed the most promising results concerning oxidative stress protection and reduction of VEGF secretion. Effects were measured on a uveal melanoma cell line (OMM-1) (oxidative stress), retinal pigment epithelium (RPE) cell line ARPE19 (oxidative stress and VEGF), and primary RPE cells (VEGF). Oxidative stress was induced by H2O2 or tert-Butyl hydroperoxide (TBHP). Cell viability was investigated with methyl thiazolyl tetrazolium (MTT or MTS) assay, and VEGF secretion with ELISA. Affinity to VEGF was determined by a competitive binding assay. Of the many species tested, Saccharina latissima and Laminaria hyperborea seem to be reasonable choices for further investigation and development, as fucoidans from both species decrease VEGF in ARPE-19 cells and RPE cells, and both may confer some protection against oxidative stress. Additionally, the studies indicate that high molecular weight may be preferable considering anti-VEGF function [32]. Further data are included in Table 1.
collaborators investigated the effect of fucoidan (Fucus vesiculosus, Sigma Aldrich) on the retinal pigment epithelium (RPE), looking at physiology, VEGF secretion, and angiogenesis, thus investigating a potential use of fucoidan for the treatment of exudative age-related macular degeneration. As result fucoidan displays no toxicity and does not diminish proliferation or phagocytosis but reduces wound healing in RPE cells. Fucoidan decreases VEGF secretion in RPE/choroid explants and RPE cells. Furthermore, it diminishes VEGF expression in RPE cells even when co-applied with bevacizumab. Furthermore, fucoidan reduces RPE-supernatant- and VEGF-induced angiogenesis of peripheral endothelial cells [34]. Since then, a variety of fucoidans have exhibited VEGF-inhibiting properties in RPE cell models. Together with Fucus vesiculosus, fucoidans from Saccharina latissima and Laminaria hyperborea were identified as the most promising candidates for further development of therapeutic agents for age-related macular degeneration [24].
Shikov et al. 2020 [19] in one of their works on fucoidans raised their results and literature data suggest as well that molecular weight, sulfate content, fucose content, and polyphenols may contribute to antioxidant, anticancerin, and anti-inflammatory activities and that fucoidan with higher molecular weight has promising biological properties to be applied in therapy of different diseases such as age-related macular degeneration [35].
Macroalgal habitats (e.g., kelp forests and seaweed beds) are the most extensive and productive coastal ecosystems on a global scale. Laminaria hyperborea (LH), commonly known as tangle or cuvie, belongs to the large brown seaweed family of the Laminariaceae (alias kelp). It mainly grows in the northeast Atlantic Ocean, especially around Scandinavia. The large kelp Laminaria hyperborea functions as the dominant foundation species along much of the wave-exposed rocky coastline of the northeast. LH, like all brown algal species, contains FCSP, commonly known as fucoidan, as a cell wall component [25]. These fucoidans contained 97.0% fucose and a degree of sulfation of 1.7. The main distinction of these fucoidans was molecular weight, with a high-molecular weight fucoidan (1548 kDa), a medium-molecular weight fucoidan (499 kDa), and a low-molecular weight fucoidan (26.9 kDa). A studie compared three purified sulfated fucans from Laminaria hyperborea regarding cell viability, oxidative stress protection, and vascular endothelial growth factor (VEGF) secretion in ocular cells. From the results taken together, the sulfated fucan Fuc1 from LH with a molecular weight average of 1548 kDa seems to be the best candidate for further research concerning AMD-relevant mechanisms. High molecular weight appears to be desirable for VEGF inhibition. In general, the biological activity of sulfated fucans is dependent on the molecular weight, and the desired effect for the treatment of ocular diseases increases with the size of the sulfated fucan [36]. On the other hand, Kopplin investigated the action of high-molecular weight LH fucoidan Fuc1 on the retinal pigment epithelium (RPE). The results have shown that Fuc1 does not impair RPE cellular functions and shows antiangiogenic and anti-inflammatory activities, which indicates its safety and strengthens its suitability concerning ocular diseases [37].
Saccharina latissima, a species that is distributed in Europe and North America, is of high ecological and increasing economic interest in coastal areas. As one of the domain species in oceanic forest, it plays an important role in providing shelter and nursey for numerous animals and purifying the offshore seawater. The Fucus and Laminaria species are brown algae found on the coasts of the North European Sea, the Baltic Sea, the Atlantic and Pacific Oceans, meaning that these species are found in a large part of our planets. These species have a high content of polysaccharides made up of different monosaccharides, generally with sulfate groups and some of them with various biological properties. In recent years, the polysaccharides of these species have been shown to play an important role as free radical scavengers and antioxidants, for the prevention of oxidative damage in living organisms [38].
2.3. Fucoxanthin and Fucoxanthinol
Fucoxanthin belonging to the xanthophyll a type of carotenoid, orange pigment that is extracted from brown algae, such as Undaria pinnatifida (Wakame), Laminaria japonica (Ma-Kombu), Phaeodactylum tricornutum, Cylindrotheca closterium. Hijikia fusiformis, and Sargassum fulvellum. The diverse health-promoting effects of fucoxanthin are attributed to its unique chemical structure such as an acetyl group, an allenic bond, and a conjugated carbonyl, along with 5,6-monoepoxide (Figure 6). The excellent antioxidant properties of fucoxanthin against oxidative stress have been known for many years and have been confirmed by numerous studies. The allenic bond and the acetyl functional group present in its structure are considered responsible for its antioxidant properties [39].
Fucoxanthin has antioxidant, anti-inflammatory and anticancer activity in a variety of model systems. Antioxidants are likely to be scavenged free radicals, viz. singlet oxygen, hydrogen peroxide, superoxide anion, and DPPH to rescue cells from the devastating effects of oxidative stress. Oxidative stress is the consequence of an imbalanced situation between oxidant and antioxidant molecules. Fucoxanthin has greater antioxidant activity and is considered a potential protective agent against ROS and/or oxidative stress. The results of studies in animals have shown its potential value in the prevention and treatment of diseases related to lifestyle, such as obesity, diabetes, cancer, cardiovascular diseases, and other chronic diseases as well as AMD [39]. Marine carotenoid fucoxanthin has been reported as the most promising antioxidant metabolite with potent resistance against oxidative stress in vitro and, in vivo. It has been evidenced by many studies on animal and human subjects to establish the ocular protective functions of fucoxanthin [23].
Fucoxanthin has been shown to have the ability to trap different free radicals in in vitro study, such as DPPH, ABTS, hydrogen peroxide, hydroxyl radical, superoxide anion and singlet oxygen, and also 12-doxyl-stearic acid and nitrobenzene with linoneic acid [40,41]. Fucoxanthin and its metabolite, fucoxanthinol, have shown antioxidant activities attributed to free radical scavenging and singlet oxygen inactivation in vitro. The hydroxyl radical trapping capacity of fucoxanthin and fucoxanthinol was shown to be significantly greater than that of α-tocopherol, although the singlet oxygen inactivation capacity of fucoxanthin and fucoxanthinol was lower than that of β-carotene. Importantly, fucoxanthin acts as an antioxidant under anoxic conditions, that is, under conditions of little dissolved oxygen, while other carotenoids, such as β-carotene and lutein, show little or no activity under these extreme conditions [42].
In several studies it has been reported that the biological activities of fucoxanthin as antioxidants, anti-inflammatory, anticancer and antimicrobial in various experiments carried out in tissues and cells [43,44,45,46][. Chen et al demonstrated that if fucoxanthin has a cytoprotective effect on retinal cells and on AMD, which shows that phycoxanthin could be considered as a treatment natural for dry macular degeneration [47].
Fucoxanthinol (Fuol), a marine xanthophyll carotenoid, possesses unusual structures with an allelic bond, a conjugated carbonyl, and a 5, 6-monoepoxid. Fuol is a major gastrointestinal metabolite of dietary fucoxanthin. To date, there are few reports about natural sources for the preparation of this compound. Nitzschia laevis, a diatom, is rich in Fuol and can be cultivated on a large scale through heterotrophic methods with a high growth rate [48,49]. Therefore, Nitzschia laevis possesses the potential to be a candidate for industrial production of Fuol [48]
The anti-inflammatory potential shown by fucoxanthin isolated from a brown alga, Myagropsis myagroides, was analyzed through the inhibition of nitric oxide (NO) production, one of the determining factors in the inflammation process. The inflammatory response, a self-defense reaction against various pathogenic stimuli, is characterized by attracting large amounts of leukocytes to the inflamed area, in which these inflammatory cells are activated by inflammation mediators and generate radicals of anions, their peroxide and nitric oxide, becoming a harmful self-destructive process, exacerbating age-related macular degeneration disease [39].Heo et al. and Kim et al. authenticated the inhibitory effects of fucoxanthin on inflammatory cytokines and mediators in lipopolysaccharide-stimulated RAW 264.7 macrophages. These results showed that fucoxanthin inhibited the inducible nitric oxide synthase and cyclooxygenase 2 protein expressions, and reduced the levels of nitric oxide, prostaglandin E2, tumor necrosis factor-α, interleukin-1β, and interleukin-6 through the inhibition of nuclear factor-κB activation and the phosphorylation of mitogen-activated protein kinases [50,51]. Considering the above results, the biological potentialities of fucoxanthin and fucoxanthin´s anti-inflammatory effect could offer protection in AMD?
3. Additional Challenge for the Use of Fucoidan in AMD
In addition to the biological activities of fucoidan shown in vitro experiments, several beneficial effects of fucoidan have been reported in humans. Fucoidans are natural compounds that are present in brown-type marine algae in different species and in animals. Currently, many researchers are very interested in the topic of fucoidans due to their biological properties as anti-inflammatory, antioxidant and anticancer, however, its mechanism of action is not well understood so far. According to the review of the literature on fucoidans, their structures have been analyzed, as well as the different methods of obtaining it according to the species, where many authors have agreed that their bioactivity is very difficult to conclude due to the diversity of their structural, the extraction methods, as well as the experimental conditions of each working group.
It is known that fucoidans present a great absorption problem when administered orally, but many experiments are required in vitro and then in vivo to solve the problem of oral absorption of fucoidans, it is thought that, in structural analysis and extraction methods where structural relationship and pharmacokinetic parameters can be achieved that can solve this problem [52].
Therefore, detection of fucoidan in the serum or urine has been thought to be impossible. The beneficial effects of fucoidan in vivo have been speculated to reflect various physiological reactions induced in the gastrointestinal tract after fucoidan administration. Consequently, specific, and highly sensitive fucoidan assay methods were developed to elucidate the effects of fucoidan and its mechanism in vivo and its biological effects in blood, tissues, and organs [53,54,55]. In general, the findings suggest that acidic conditions in the stomach have a restricted effect on fucoidan and that fucoidan-digesting enzymes are not secreted by human intestinal flora. On the other hand, there is another challenge that is the molecular weight of the fucoidan excreted in urine was significantly less than that of orally administrated. As Tokita et al and Phuong et al have mentioned, the results suggest that fucoidan degradation occurs in the excretory system, possibly in the kidneys, but not in the intestine by bacterial flora. Although FCD has continuously demonstrated promising biological activities by oral administration, its suggested broad-spectrum use in therapy may lead to confusion for clinical applications [56,57].
Additionally, the poor bioavailability and instability of fucoxanthin in the gastrointestinal tract have major limitations. Several studies have been made to facilitate the application of fucoxanthin in the food industry and improve low bioavailability. For example, a combination of monocaprin with fucoxanthin-containing co-feeding algal oil (SO) is an effective treatment to improve fucoxanthin bioavailability [58]. Alternatively, encapsulation is a promising approach to overcome these challenges by enclosing fucoxanthin in a protective layer, such as liposomes or nanoparticles. Encapsulation can improve the stability of fucoxanthin by protecting it from exposure to heat, pH, illumination, gastric acids and enzymes that can accelerate its degradation [59].
4. Conclusions
A major cause for blindness worldwide is age-related macular degeneration, and its incidence and prevalence are rising. There is no treatment for the early or atrophic forms of AMD, so the demands of new treatment options are unmet. Brown seaweed contains bioactive compounds with promising activities that make them interesting for potential use in AMD, such as the poly-saccharide fucoidan, which exhibit many different biological activities: antioxidant activities, anti-inflammatory, antioxidant and immunomodulatory that do compromise against AMD.
In the past few years, several fucoidans’ structures from algal species have been solved, and many aspects of their biological activity were proved as possible alternative treatment for AMD. The fucoidans from Saccharina latissimi, Laminaria hyperborean and Fucoxanthins displayed the most promising results concerning oxidative stress protection and reduction of VEGF secretion, however more studies are needed to translate from lab to human clinical trials.
References
- Mitchell, P.; Liew, G.; Gopinath, B.; Wong, T.Y. Age-related macular degeneration. Lancet 2018, 1147–1159. [Google Scholar] [CrossRef]
- Wang, Y.; Zhong, Y.; Zhang, L.; Wu, Q.; Tham, Y.; Rim, T.H.; Kithinji, D.M.; Wu, J.; Cheng, C.; Liang, H.; et al. Global Incidence, Progression, and Risk Factors of Age-Related Macular Degeneration and Projection of Disease Statistics in 30 Years: A Modeling Study. Gerontology 2021, 68, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Shaw, P.X.; Stiles, T.; Douglas, C.; Ho, D.; Fan, W.; Du, H. ; Xiao, X Oxidative stress, innate immunity, and age-related macular degeneration. AIMS molecular science 2016, 196–221. [Google Scholar] [CrossRef] [PubMed]
- Jonas, J.B. Updates on the Epidemiology of Age-Related Macular Degeneration. Asia Pac J Ophthalmol (Phila) 2017, 493–497. [Google Scholar]
- Bhutto, I.; Lutty, G. Understanding age-related macular degeneration (AMD): Relationships between the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choriocapillaris complex. Mol. Asp. Med. 2012, 33, 295–317. [Google Scholar] [CrossRef]
- Fleckenstein, M.; Schmitz-Valckenberg, S.; Chakravarthy, U. Age-Related Macular Degeneration: A Review. JAMA 2024, 147–157. [Google Scholar] [CrossRef]
- Gheorghe, A.; Mahdi, L.; Musat, O. Age-related macular degeneration. Rom J Ophthalmol 2015, 74–77. [Google Scholar]
- Fritsche, L.; Igl, W.; Cooke-Bailey, J.N.; Grassmann, F.; Sengupta, S. Nat Genet 2016, 134–143. [CrossRef]
- Sies, H.; Carsten, B.; Jones, D.P. Oxidative Stress. Annu Rev Biochem 2017, 715–748. [Google Scholar] [CrossRef]
- Klettner, A. Oxidative stress induced cellular signaling in RPE cells. Frontiers in bioscience (Scholar edition) 2012, 392–411. [Google Scholar] [CrossRef]
- Upadhyay, M.; Milliner, C.; Bell, B.A.; Bonilha, V.L. Oxidative stress in the retina and retinal pigment epithelium (RPE): Role of aging, and DJ-1. Redox Biol 2020, 37, 101623. [Google Scholar] [CrossRef]
- Masuda, T.; Shimazawa, M.; Hara, H. Retinal Diseases Associated with Oxidative Stress and the Effects of a Free Radical Scavenger (Edaravone). Oxidative Med. Cell. Longev. 2017, 2017, 9208489. [Google Scholar] [CrossRef]
- Schmidt-Erfurth, U.; Chong, V.; Loewenstein, A.; Larsen, M.; Souied, E.; Schlingemann, R.; Eldem, B.; Monés, J.; Richard, G.; Bandello, F. Guidelines for the management of neovascular age-related macular degeneration by the European Society of Retina Specialists (EURETINA). Br. J. Ophthalmol. 2014, 98, 1144–1167. [Google Scholar] [CrossRef]
- Hytti, M.; Korhonen, E.; Hongisto, H.; Kaarniranta, K.; Skottman, H.; Kauppinen, A. Differential Expression of Inflammasome-Related Genes in Induced Pluripotent Stem-Cell-Derived Retinal Pigment Epithelial Cells with or without History of Age-Related Macular Degeneration. Int. J. Mol. Sci. 2021, 22, 6800. [Google Scholar] [CrossRef]
- Watkins, W.M.; McCollum, G.W.; Savage, S.R.; Capozzi, M.E.; Penn, J.S.; Morrison, D.G. Hypoxia-induced expression of VEGF splice variants and protein in four retinal cell types. Experimental eye research 2013, 116, 240–246. [Google Scholar] [CrossRef]
- Klettner, A.; Koinzer, S.; Meyer, T.; Roider, J. Toll-like receptor 3 activation in retinal pigment epithelium cells – Mitogen-activated protein kinase pathways of cell death and vascular endothelial growth factor secretion. Acta Ophthalmol. 2013, 91, e211–e218. [Google Scholar] [CrossRef]
- Girgis, S.; Lee, L.R. Treatment of dry age-related macular degeneration: A review. Clin. Exp. Ophthalmol. 2023, 51, 835–852. [Google Scholar] [CrossRef]
- Klettner, A. Fucoidan as a Potential Therapeutic for Major Blinding Diseases—A Hypothesis. Mar. Drugs 2016, 14, 31. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, Z.; Wang, S.; Liu, Y.; Wei, Y. Fucoxanthin Pretreatment Ameliorates Visible Light-Induced Phagocytosis Disruption of RPE Cells under a Lipid-Rich Environment via the Nrf2 Pathway. Mar. Drugs 2021, 20, 15. [Google Scholar] [CrossRef]
- Stengel, D.; Connan, S. ; Popper, Z Algal chemo diversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnology Advances 2011, 483–501. [Google Scholar] [CrossRef]
- Pangestuti, R. Biological activities and health beneft effects of natural pigments derived from marine algae. Journal of Functional Foods 2011, 3, 255–66. [Google Scholar] [CrossRef]
- Mohibbullah, M.; Haque, M.; Sohag, A.; Hossain, M.; Zahan, M.; Uddin, M.; Hannan, M.; Moon, I.; Choi, J. A Systematic Review on Marine Algae-Derived Fucoxanthin: An Update of Pharmacological Insights. Mar Drugs 2022, 20, 279. [Google Scholar] [CrossRef]
- Dörschmann, P.; Klettner, A. Fucoidans as Potential Therapeutics for Age-Related Macular Degeneration—Current Evidence from In Vitro Research. Int. J. Mol. Sci. 2020, 21, 9272. [Google Scholar] [CrossRef]
- Zayed, A.; El-Aasr, M.; Ibrahim, A.-R.S.; Ulber, R. Fucoidan Characterization: Determination of Purity and Physicochemical and Chemical Properties. Mar. Drugs 2020, 18, 571. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Nagahawatta, D.P.; Fernando, I.P.S.; Kim, Y.-T.; Kim, J.-S.; Kim, W.-S.; Lee, J.S.; Jeon, Y.-J. A Review on Fucoidan Structure, Extraction Techniques, and Its Role as an Immunomodulatory Agent. Mar. Drugs 2022, 20, 755. [Google Scholar] [CrossRef]
- Lee, H.G.; Jayawardena, T.U.; Liyanage, N.; Song, K.-M.; Choi, Y.-S.; Jeon, Y.-J.; Kang, M.-C. Antioxidant potential of low molecular weight fucoidans from Sargassum autumnale against H2O2-induced oxidative stress in vitro and in zebr. Food Chemistry 2022, 132591. [Google Scholar]
- Marudhupandi, T.; Kumar, T.A.; Lakshmana-Senthil, S.; Nanthini-Devi, K. Invitro antioxidant properties of fucoidans fractions from Sargassum tenerrimum. Pakistan Journal of Biological Sciences 2014, 402–407. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Wu, S.-J.; Yang, W.-N.; Kuan, A.-W.; Chen, C.-Y. Antioxidant activities of crude extracts of fucoidan extracted from Sargassum glaucescens by a compressional-puffing-hydrothermal extraction process. Food Chem. 2016, 197, 1121–1129. [Google Scholar] [CrossRef]
- Zayed, A.; Al-Saedi, D.A.; Mensah, E.O.; Kanwugu, O.N.; Adadi, P.; Ulber, R. Fucoidan’s Molecular Targets: A Comprehensive Review of Its Unique and Multiple Targets Accounting for Promising Bioactivities Supported by In Silico Studies. Mar. Drugs 2023, 22, 29. [Google Scholar] [CrossRef]
- Dörschmann, P.; Mikkelsen, M.D.; Thi, T.N.; Roider, J.; Meyer, A.S.; Klettner, A. Effects of a Newly Developed Enzyme-Assisted Extraction Method on the Biological Activities of Fucoidans in Ocular Cells. Mar. Drugs 2020, 18, 282. [Google Scholar] [CrossRef] [PubMed]
- Dörschmann, P.; Bittkau, K.S.; Neupane, S.; Roider, J.; Alban, S.; Klettner, A. Effects of Fucoidans from Five Different Brown Algae on Oxidative Stress and VEGF Interference in Ocular Cells. Mar. Drugs 2019, 17, 258. [Google Scholar] [CrossRef]
- Li, Q.; Wang, X.; Wan, Y.; Hu, X.; Liu, J.; Wang, J. In vivo immunomodulatory activity of fucoidan from brown alga Undaria pinnatifida in sarcoma 180-bearing mice. J. Funct. Foods 2023, 103. [Google Scholar] [CrossRef]
- Dithmer, M.; Fuchs, S.; Shi, Y.; Schmidt, H.; Richert, E.; Roider, J.; Klettner, A. Fucoidan Reduces Secretion and Expression of Vascular Endothelial Growth Factor in the Retinal Pigment Epithelium and Reduces Angiogenesis In Vitro. PLOS ONE 2014, 9, e89150. [Google Scholar] [CrossRef]
- Pozharitskaya, O.; Obluchinskaya, E.; Shikov, A.N. Mechanisms of Bioactivities of Fucoidan from the Brown Seaweed Fucus vesiculosus L. of the Barents. Sea. Mar Drugs 2020, 275. [Google Scholar] [CrossRef]
- Smale, D.A.; Pessarrodona, A.; King, N.; Burrows, M.T.; Yunnie, A.; Vance, T.; Moore, P. Environmental factors influencing primary productivity of the forest-forming kelp Laminaria hyperborea in the northeast Atlantic. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Dörschmann, P.; Kopplin, G.; Roider, J.; Klettner, A. Interaction of High-Molecular Weight Fucoidan from Laminaria hyperborea with Natural Functions of the Retinal Pigment Epithelium. Int. J. Mol. Sci. 2023, 24, 2232. [Google Scholar] [CrossRef] [PubMed]
- Stévant, P.; Rebours, C.; Chapman, A. Seaweed aquaculture in Norway: recent industrial developments and future perspectives. Aquac. Int. 2017, 25, 1373–1390. [Google Scholar] [CrossRef]
- Peng, J.; Yuan, J.-P.; Wu, C.-F.; Wang, J.-H. Fucoxanthin, a Marine Carotenoid Present in Brown Seaweeds and Diatoms: Metabolism and Bioactivities Relevant to Human Health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Liu, C.-L.; Liang, A.-L.; Hu, M.-L. Protective effects of fucoxanthin against ferric nitrilotriacetate-induced oxidative stress in murine hepatic BNL CL.2 cells. Toxicol. Vitr. 2011, 25, 1314–1319. [Google Scholar] [CrossRef]
- Wang, L.; Park, Y.-J.; Jeon, Y.-J.; Ryu, B. Bioactivities of the edible brown seaweed, Undaria pinnatifida: A review. Aquaculture 2018, 495, 873–880. [Google Scholar] [CrossRef]
- Neumann, U.; Derwenskus, F.; Flaiz Flister, V.; Schmid-Staiger, U.; Hirth, T.; Bischoff, S.C. Fucoxanthin, A Carotenoid Derived from Phaeodactylum tricornutum Exerts Antiproliferative and Antioxidant Activities In Vitro. Antioxidants 2019, 8, 183. [Google Scholar] [CrossRef]
- Morandi, A.C.; Molina, N.; Guerra, B.A.; Bolin, A.P.; Otton, R. Fucoxanthin in association with Vitamin c acts as modulators of human neutrophil function. Eur. J. Nutr. 2013, 53, 779–792. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, C.; Liu, C.; Sue, Y.; Chen, T.; Hsu, Y.; Hwang, P.; Chen, C. Alleviative effect of fucoxanth in containing extract from brown seaweed Laminaria japonica on renal tubular cell apoptosis through upregulating Na+/H+ exchanger NHE1 in chronic kidney disease mice J. Ethnopharmacol 2018, 391–399. [Google Scholar] [CrossRef]
- Karpi´nski, T.; Adamczak, A. Fucoxanthin—An Antibacterial Carotenoid. Antioxidants 2019, 239. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, H.; Liu, Y. Anti-Inflammatory and Apoptotic Signaling Effect of Fucoxanthin on Benzo(A)Pyrene-Induced Lung Cancer in Mice. J. Environ. Pathol. Toxicol. Oncol. 2019, 38, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-J.; Lin, T.-B.; Peng, H.-Y.; Liu, H.-J.; Lee, A.-S.; Lin, C.-H.; Tseng, K.-W. Cytoprotective Potential of Fucoxanthin in Oxidative Stress-Induced Age-Related Macular Degeneration and Retinal Pigment Epithelial Cell Senescence In Vivo and In Vitro. Mar. Drugs 2021, 19, 114. [Google Scholar] [CrossRef]
- Heo, S.J.; Yoon, W.J.; Kim, K.N.; Ahn, G.N.; Kang, S.M.; Kang, D.H.; Affan, A.; Oh, C.; Jung, W.K.; Jeon, Y.J. Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated raw 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Yuan, J.-P.; Wu, C.-F.; Wang, J.-H. Fucoxanthin, a Marine Carotenoid Present in Brown Seaweeds and Diatoms: Metabolism and Bioactivities Relevant to Human Health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Heo, S.-J.; Ko, S.-C.; Kang, S.-M.; Kang, H.-S.; Kim, J.-P.; Kim, S.-H.; Lee, K.-W.; Cho, M.-G.; Jeon, Y.-J. Cytoprotective effect of fucoxanthin isolated from brown algae Sargassum siliquastrum against H2O2-induced cell damage. Eur. Food Res. Technol. 2008, 228, 145–151. [Google Scholar] [CrossRef]
- Kim, K.; Heo, S.; Yoon, W.; Kang, S.; Ahn, G.; Yi, T.; Jeon, Y. Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MAPKs in lipopolysaccharide-induced RAW 264.7 macrophages. Eur. J. Pharmacol 2010, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Irhimeh, M.; Fitton, J.; Lowenthal, R. Pilot clinical study to evaluate the anticoagulant activity of fucoidan. Blood Coagulation & Fibrinolysis: A International Journal in Haemostasis and Thrombosis 2009, 607–610. [Google Scholar]
- Tokita, Y.; Hirayama, M.; Nakajima, K.; Tamaki, K.; Iha, M.; Nagamine, T. Detection of Fucoidan in Urine after Oral Intake of Traditional Japanese Seaweed, Okinawa mozuku (Cladosiphon okamuranus Tokida). J. Nutr. Sci. Vitaminol. 2017, 63, 419–421. [Google Scholar] [CrossRef] [PubMed]
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M.; Kongtawelert, P. A quantitative method to detect fucoidan in human plasma using a novel antibody. Methods Find. Exp. Clin. Pharmacol. 2005, 27, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Imbs, T.; Zvyagintseva, T.; Ermakova, S. Is the transformation of fucoidans in human body possible? Int. J. Biol. Macromol. 2019, 142, 778–781. [Google Scholar] [CrossRef]
- Tokita, Y.; Nakajima, K.; Mochida, H.; Iha, M.; Nagamine, T. Development of a Fucoidan-Specific Antibody and Measurement of Fucoidan in Serum and Urine by Sandwich ELISA. Biosci. Biotechnol. Biochem. 2010, 74, 350–357. [Google Scholar] [CrossRef]
- Tran, P.H.; Lee, B.-J.; Tran, T.T. Current developments in the oral drug delivery of fucoidan. Int. J. Pharm. 2021, 598, 120371. [Google Scholar] [CrossRef]
- Nagata, K.; Takatani, N.; Beppu, F.; Abe, A.; Tominaga, E.; Fukuhara, T.; Ozeki, M.; Hosokawa, M.M. Enhances Bioavailability of Fucoxanthin in Diabetic/Obese KK-Ay Mice. Mar. Drugs 2022, 446. [Google Scholar] [CrossRef]
- Fernandes, V.; Mamatha, B.S. Fucoxanthin, a Functional Food Ingredient: Challenges in Bioavailability. Curr. Nutr. Rep. 2023, 12, 567–580. [Google Scholar] [CrossRef]
Figure 1.
Eye anatomy. Iris is the colored part of the eye, and the dark hole in the middle is called the pupil. The sclera is the white and protective layer of tissue that covers most of the eyeball. By helping to focus light as it enters the eye, the lens plays important roles in giving clear vision. A tiny but very specialized area of the retina called the macula is responsible for giving detailed central vision. The retina sends light as electrical impulses through the optic nerve to the brain. Blood vessels are responsible for the aye blood supply.
Figure 1.
Eye anatomy. Iris is the colored part of the eye, and the dark hole in the middle is called the pupil. The sclera is the white and protective layer of tissue that covers most of the eyeball. By helping to focus light as it enters the eye, the lens plays important roles in giving clear vision. A tiny but very specialized area of the retina called the macula is responsible for giving detailed central vision. The retina sends light as electrical impulses through the optic nerve to the brain. Blood vessels are responsible for the aye blood supply.
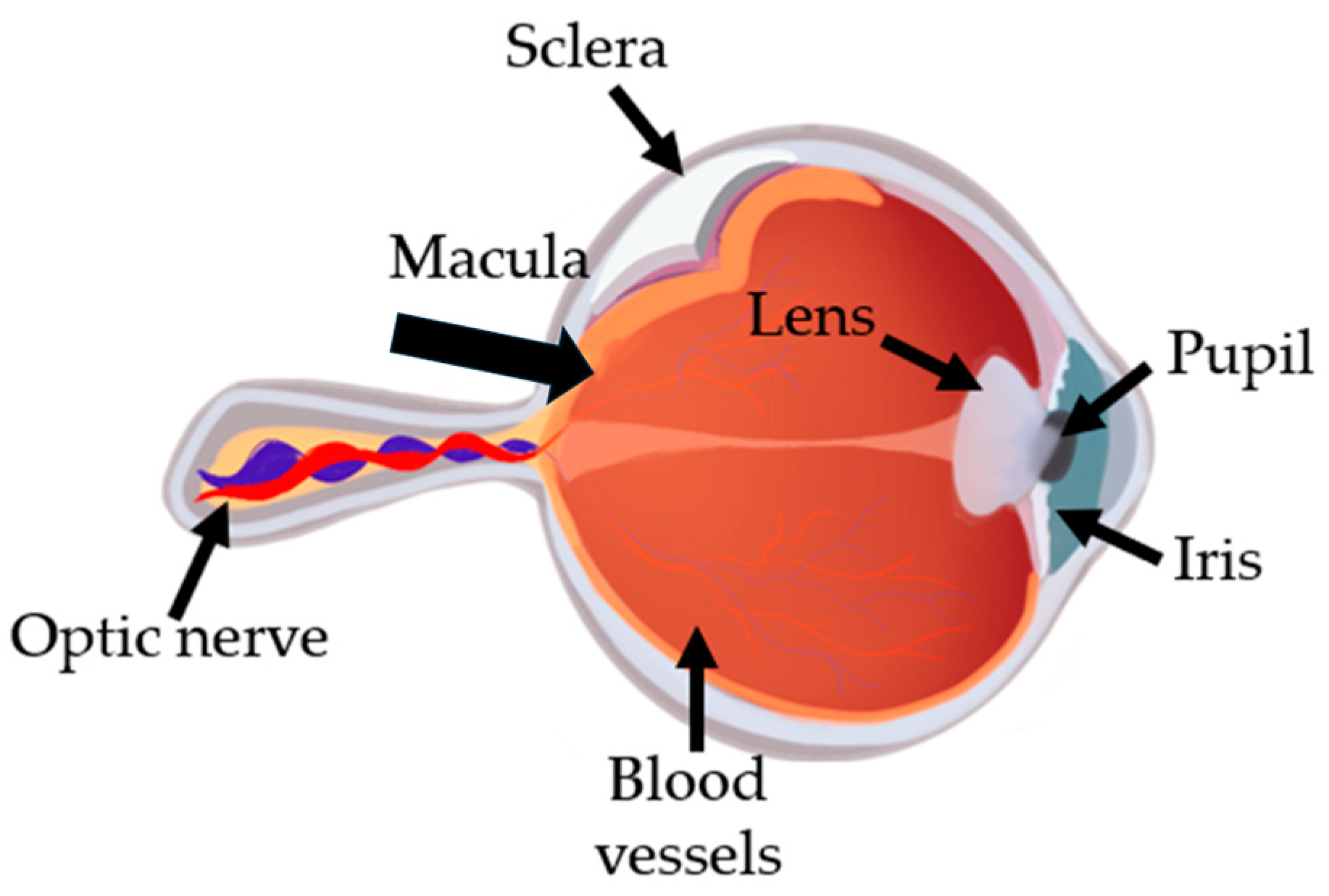
Figure 2.
Visualization of central vision loss due to AMD.
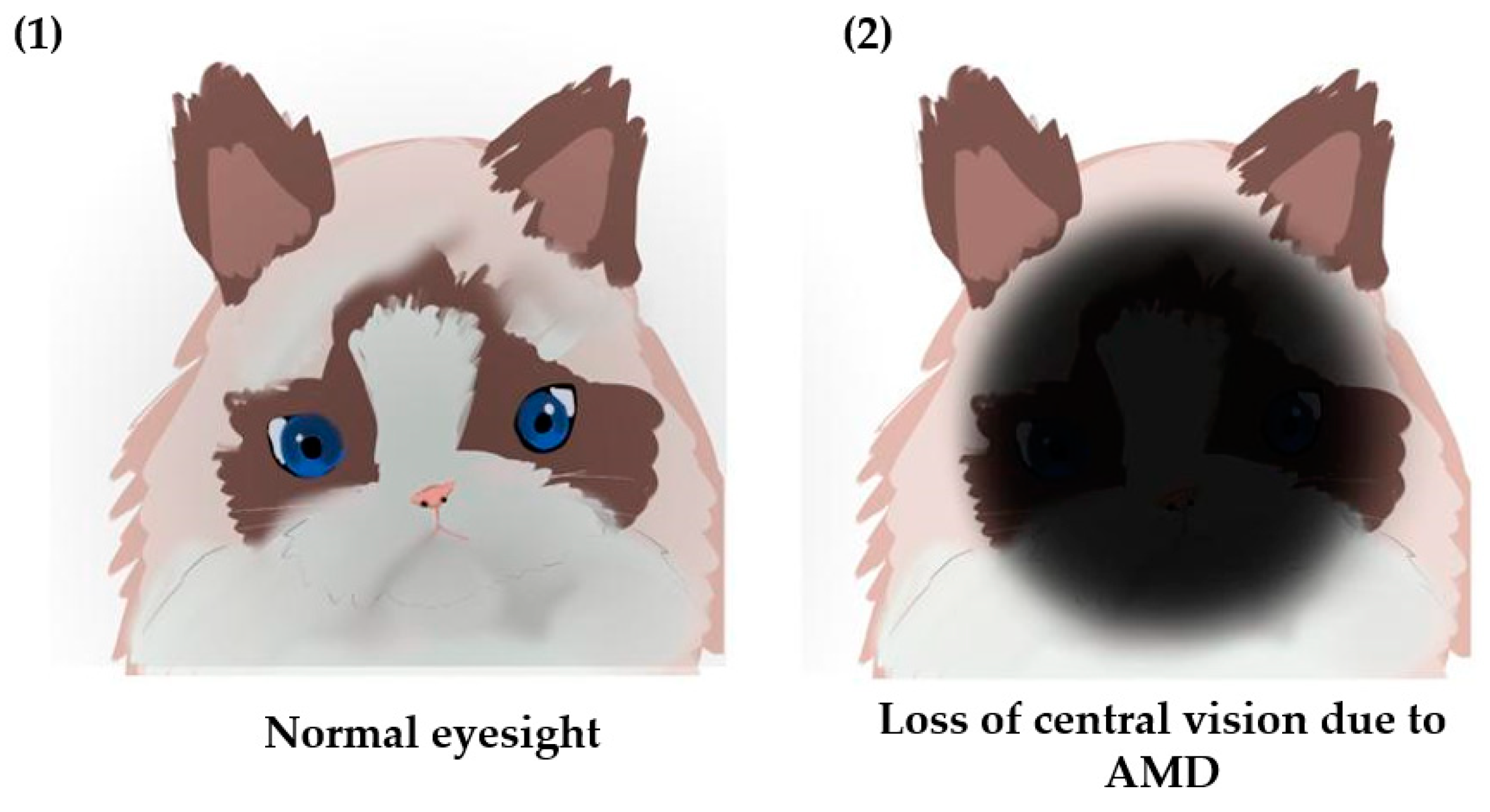
Figure 3.
Oxidative cell damage in the retinal cells.
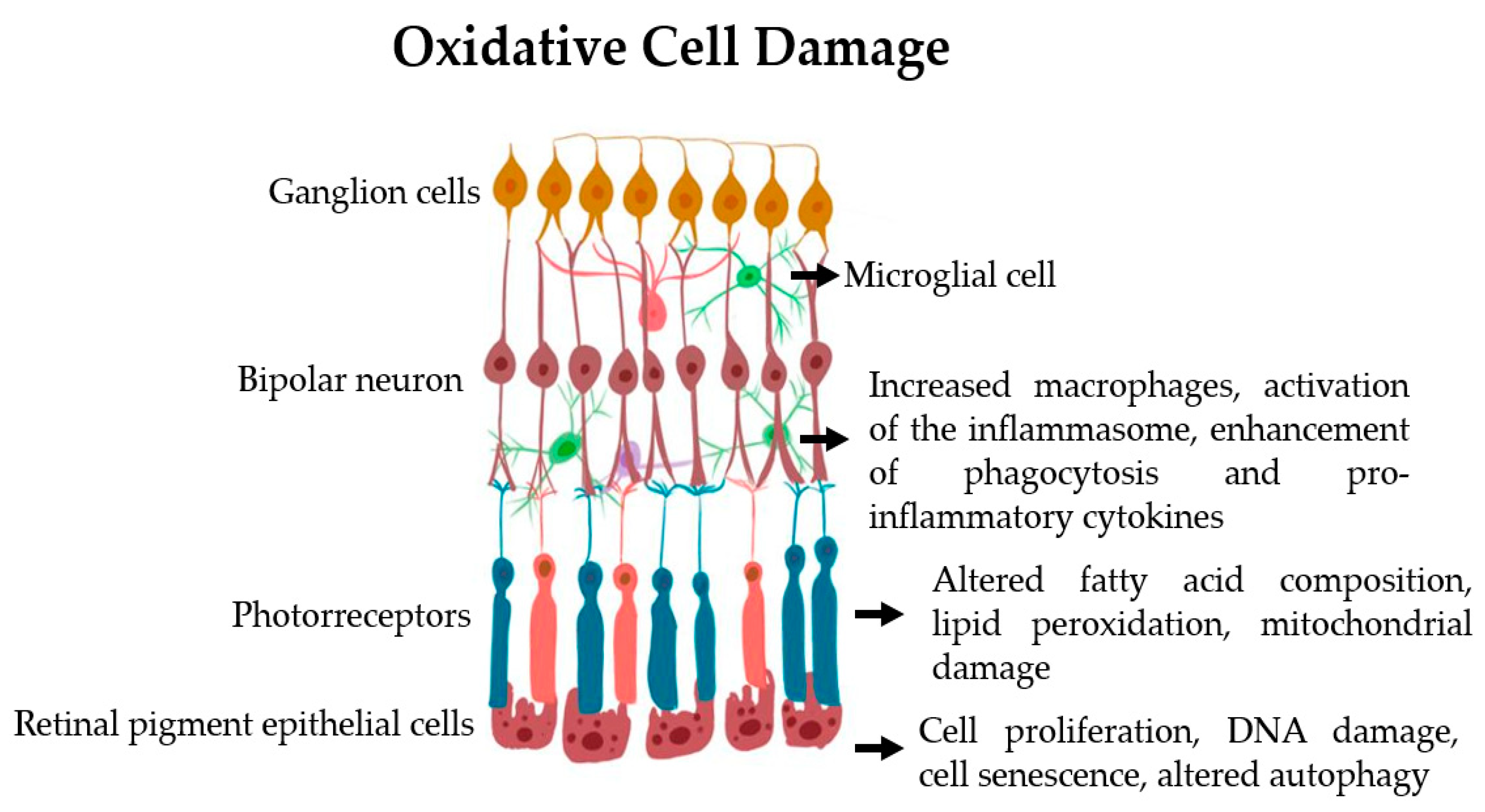
Figure 4.
Fucoidan structure from different brown seaweeds.
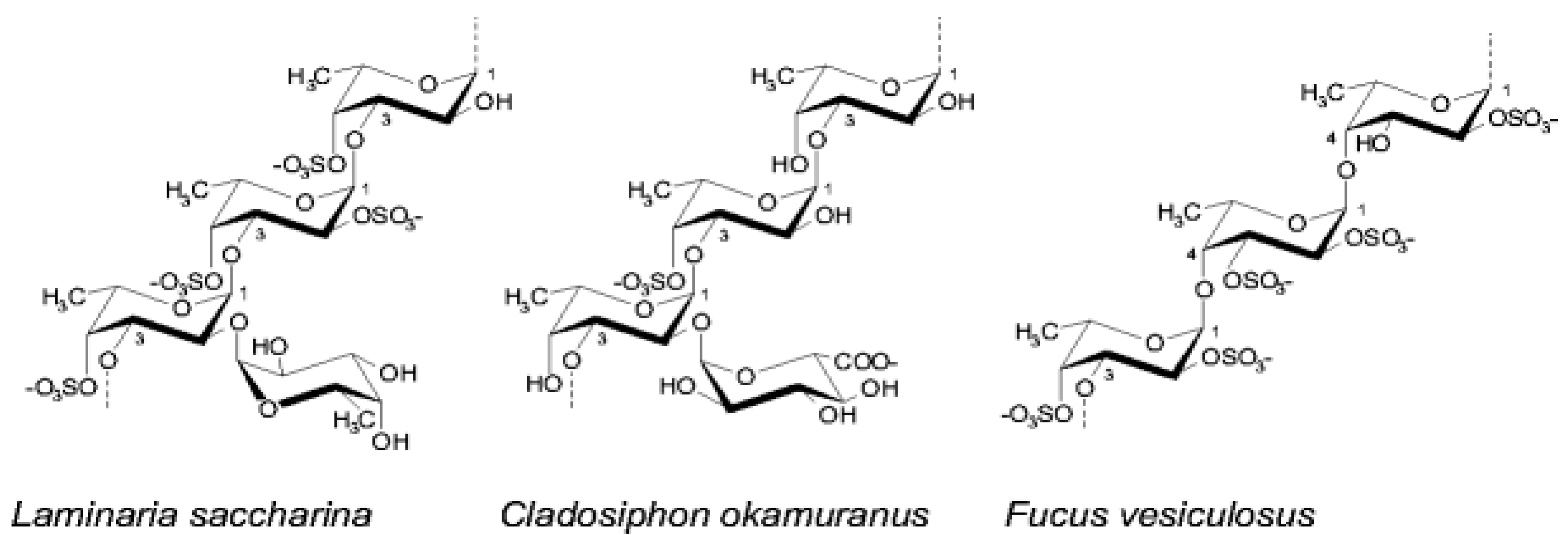
Figure 5.
Fucoidan Extraction Methods from Sargassum comparison, the disadvantages of every reagent. Adapted from Jayawardena et al [26].
Figure 5.
Fucoidan Extraction Methods from Sargassum comparison, the disadvantages of every reagent. Adapted from Jayawardena et al [26].
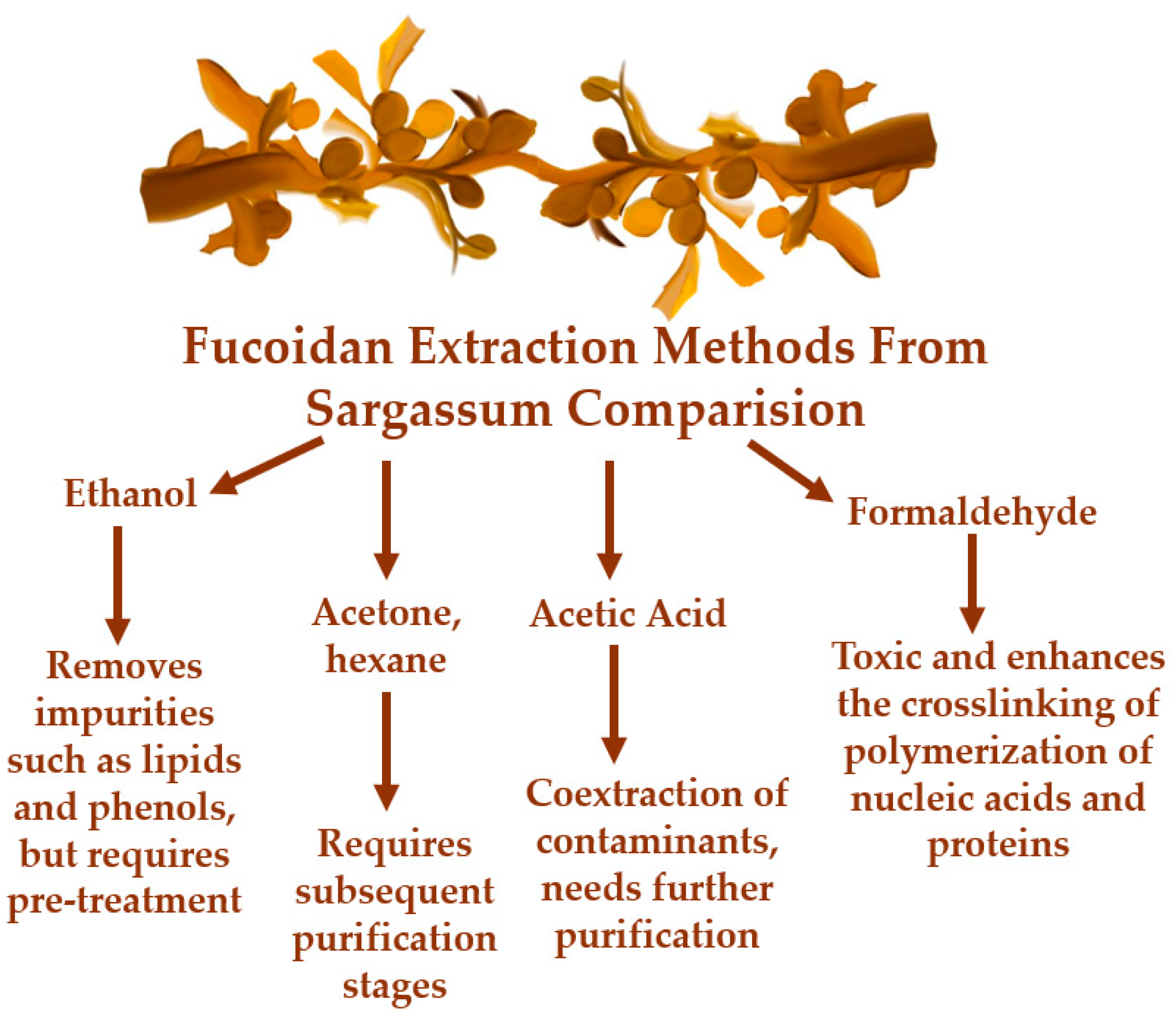
Figure 6.
The chemical structure of fucoxanthin.
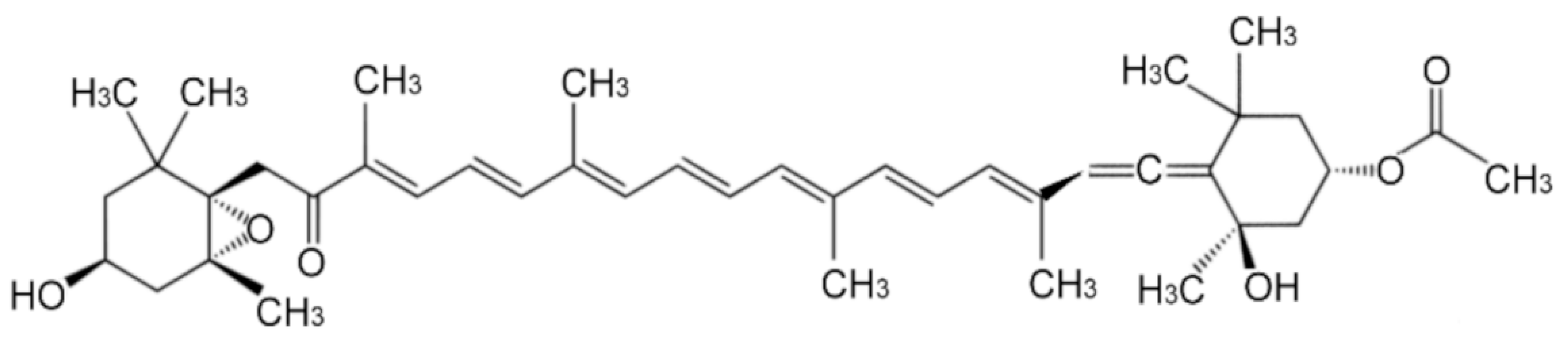

Table 1.
Effect of different fucoidans in different cell culture in vitro and in vivo models.
| Cell type | Disease Model | Concentration | Source | Effects | Reference |
|---|---|---|---|---|---|
| Microvascular endothelial cells |
High glucose | 12.5, 25, 50/mL | Fucoidan de L. japonica | Reduces VEGF (retina) | [19] Klettner, A et. 2016 |
| human colon carcinoma HT-29 | human colon | 0.272- 3.7mg/ml | Sargassum glaucescens | Antioxidant | [29] Huang, C.-Y et al. 2016 |
| primary porcine RPE cells (brighfield, 50×) | uveal melanoma cell line (OMM-1) | 1, 50 and 100 µgç |
Saccharina latissima And Laminaria hyperborea |
Reduces VEGF | [24] Klettner, A et al. 2020 |
| Murine S180 cell line | in vitro | 100, 200 and 300 mg/kg/day) | Undaria pinnatifida | Immunomodulatory | [33]. Qin Li et al. 2023 |
| RAW 264.7 macrophage cells | in vitro and in zebra fish | 25,50,100,200 µg/mL | Sargassum autumnale | anti-inflammatory | [27] Lee, H.U et al. |
| RAW 264.7 macrophage cells | in vitro | 10,50,250,5000 and 1000 µg/mL | Sargassum tenerrimum | Antioxidant | [28] Marudhupandi, T. et al. 2014 |
| primary porcine RPE cells (brighfield, 50×) | OMM-1 Cells | 100 µM, 200µM, 400 µM1000 µM | Fucus vesiculosus, Fucus distichus subsp. evanescens, Fucus serratus, Laminaria digitata, Saccharina latissima | Oxidative Stress and reduces VEGF | [32].P. Dörschmann et al. 2019 |
| ARPE-19 and porcine RPE-cells, | OMM-1 Cells | 100 µg/mL | Fucus vesiculosus | reduces VEGF |
[34] M. Dithmer et al. 2014 |
| ARPE-19 cells | In Vivo and In Vitro | 0.1,1 and 10 mg/kg | Hijikia fusiformis, Laminaria japonica and Sargassum fulvellum | Cytoprotective | [[47[M1] ]Chen, S.-J et al. 2021 |
| RAW 264.7 macrophage cells | In vitro | 12.5, 25 and 50 µM | Ishige okamurae | anti-inflammatory | [51]K.-N. Kim et al. 2010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



