Submitted:
06 February 2025
Posted:
06 February 2025
You are already at the latest version
Abstract
The AT-hook motif nuclear-localized (AHL) genes play crucial roles in chromatin remodeling and regulation of gene transcription, significantly influencing biological processes such as plant growth, development, and stress responses. Although AHL genes have been extensively studied in various plants, their biological roles in pepper (Capsicum annuum L.) remain unknown. In this study, 45 AHL genes were identified in the pepper genome through bioinformatics approaches. These genes were analyzed for their chromosomal distribution, phylogenetic relationships, and the structural and functional characteristics of their encoded proteins. Phylogenetic analysis categorized the CaAHL proteins into four subgroups. Transcriptome data revealed that CaAHL genes are expressed across various tissues, including roots, stems, flowers, seeds, and fruits, at different growth stages. Subcellular localization experiments confirmed that CaAHL23 predominantly localizes to the cell nucleus, consistent with the localization of its homolog TEK in Arabidopsis thaliana. A yeast two-hybrid assay revealed that CaAHL23 binds to the promoter of CaCYP703A2, which is the homolog of the genic male sterility gene SlCYP703A2 in tomato. These findings indicate a potential link between CaAHL23 and genic male fertility in pepper. In summary, this study provides valuable insights that establish a foundation for future functional studies of CaAHL genes in pepper.
Keywords:
pepper
; AT-hook motif nuclear-localized (AHL) gene family
; gene expression
; genic male sterility
1. Introduction
Capsicum annuum L., commonly known as chili pepper, is a globally significant crop renowned for its distinctive pungency and rich nutritional profile. Their economic importance is underscored by their extensive cultivation and the diverse products derived from them, including fresh produce, spices, and pharmaceuticals. Advancements in genomic research, notably the sequencing of the pepper genome, have paved the way for in-depth studies into gene families that influence key agronomic traits, thereby enhancing breeding programs aimed at improving yield, disease resistance, and stress tolerance [1,2,3].
The AT-hook motif nuclear-localized (AHL) gene family is defined by the presence of an AT-hook motif, a small DNA-binding domain that specifically interacts with AT-rich DNA regions, as well as a conserved plant and prokaryote conserved (PPC) domain. Over the past decade, the role of AHLs in regulating various plant growth and developmental processes has been increasingly recognized. These include the elongation of hypocotyls [4,5,6], the formation of pollen walls and flower development [7,8], root growth [9,10], petiole elongation [11], and the modulation of phytohormone signaling [11]. Furthermore, AHL genes have been implicated in plant responses to pathogen infections, as well as to environmental stresses such as salt and drought [12,13,14,15]. Recent studies have also broadened our understanding of the AHL gene family in important crops like cotton, soybean, maize, and rice [16,17,18,19]. These findings emphasize that diverse roles of AHLs in plant development, offering new strategies for crop improvement.
In recent years, significant progress has been made in understanding the roles of specific AHL genes in plant flower development and male sterility. In Arabidopsis thaliana, AHL22 was identified as a critical regulator of flowering time by modulating FLOWERING LOCUS T (FT) chromatin state [20]. Similarly, AHL16 suppressed transposon activation and ensured timely flowering [21]. TEK (TRANSPOSABLE ELEMENT SILENCING VIA AT-HOOK) was found essential for male fertility. TEK regulates AGPs in anthers critical for pollen wall formation. Mutations caused microspore development defects and sterility [22]. AHL15 promoted somatic embryogenesis, highlighting its biotechnological potential [23]. In rice, PERSISTENT TAPETAL CELL2 (PTC2) regulated tapetal programmed cell death and pollen wall patterning, with its loss leading to male sterility [7]. However, despite these advancements, comprehensive investigations into the AHL gene family in chili pepper remain scarce.
Given the economic importance of chili pepper and the potential roles of AHL genes in plant development and stress responses, a genome-wide analysis of the AHL gene family in chili pepper is warranted. This study aims to identify and characterize the CaAHL genes, analyze their expression profiles across different tissues and developmental stages, and explore their potential functions in relation to genic male sterility. The findings from this research will provide valuable insights into the molecular mechanisms governing chili pepper development and facilitate the development of improved cultivars through targeted breeding strategies.
2. Results
2.1. Identification and Basic Information About the CaAHL Gene Family
The Hidden Markov Model (HMM) profile of the PPC domain (PF03479) was used as a query sequence to identify AHL proteins in the Zhangshugang genome database. An initial search yielded 47 AHL candidates from the reference genome. After verification, partial AHL genes with incomplete PPC domains, considered pseudogenes, were excluded. Ultimately, 45 AHL proteins were confirmed for further analysis. To facilitate subsequent studies, these pepper AHL genes were designated as CaAHL1 to CaAHL45 based on their positions across the 12 chromosomes in the Zhangshugang reference genome. Comprehensive details, including gene names, chromosomal locations, protein lengths, molecular weights, theoretical isoelectric points (pI), instability indices, grand average of hydropathicity, and predicted subcellular localizations, are provided in Table 1.
The CaAHLs exhibited significant variation in protein length and physicochemical properties. The protein lengths ranged from 111 amino acids (CaAHL24) to 578 amino acids (CaAHL39). Correspondingly, the molecular weights (MW) varied from 11.71 kDa (CaAHL40) to 60.51 kDa (CaAHL39). The theoretical isoelectric points (pI) spanned from 4.44 (CaAHL34) to 11.25 (CaAHL40), while the instability indices ranged from 30.43 (CaAHL24) to 63.62 (CaAHL31). The grand average of hydropathicity (GRAVY) values ranged from -0.76 (CaAHL4) to 0.18 (CaAHL27). Subcellular localization predictions for the 45 CaAHL proteins revealed diverse distribution patterns: 20 localized to the nucleus, 17 to the chloroplast, five to the cytoplasm, and one each to the plasma membrane, vacuole membrane, and endoplasmic reticulum (ER) (Table 1).
2.2. Chromosome Distribution of the CaAHL Gene Family
The 45 CaAHLs were unevenly distributed across 9 of the 12 chromosomes in the Zhangshugang genome (Figure 1a). Most CaAHLs were concentrated near the distal ends of the chromosomes, with only a few located in central regions. Chromosome 1 had the highest number of CaAHLs, with a total of 21, followed by chromosomes 12 and 3, which contained 7 and 6 genes, respectively. In contrast, no CaAHLs were identified on chromosomes 10 and 11 (Figure 1b).
Interestingly, the CaAHLs were enriched in four specific regions of the genome: the proximal and distal ends of chromosome 1, the distal end of chromosome 3, and the proximal end of chromosome 12. These regions were designated as cluster 1, cluster 2, cluster 3, and cluster 4, respectively. Cluster 1 comprises five genes (CaAHL1 to CaAHL5), cluster 2 contains 12 genes (CaAHL10 to CaAHL21), cluster 3 includes three genes (CaAHL27 to CaAHL29), and cluster 4 consists of four genes (CaAHL39 to CaAHL42) (Figure. 1a). The uneven distribution and clustering of CaAHLs suggest a possible accumulation of genes through tandem duplication events in specific genomic regions.
2.3. Phylogenetic Analysis of the CaAHL Gene Family
We subsequently performed a phylogenetic analysis of the 45 CaAHLs, using their protein sequences to construct an unrooted tree for classification of the CaAHL gene family (Figure 2). The phylogenetic analysis revealed that the CaAHL gene family is divided into four main branches: branch A, containing 15 genes; branch B, containing 9 genes; branch C, containing 6 genes; and branch D, containing 15 genes. This classification provides insight into the evolutionary relationships within the CaAHL gene family. Combined with chromosome localization analysis, gene clusters were mostly distributed in the branch D, suggesting that the differentiation of gene function may gradually occur with gene duplication.
Next, we analyzed the tandem duplications within the CaAHL gene family and highlighted the genes in the four CaAHL gene clusters that may exhibit tandem repeats by using different colors in the phylogenetic tree (Figure 2). Overall, we observed that the five genes in cluster 1 grouped together with the five genes in cluster 2 in the phylogenetic tree, while the remaining three genes in cluster 1 clustered with the three genes in cluster 3 (Figure 2). We speculate that this pattern may result from the duplication of entire gene clusters on chromosomes during evolutionary processes. From the perspective of gene clusters, we found that five genes in cluster 1 (CaAHL13, CaAHL14, CaAHL16, CaAHL17, and CaAHL18) and four genes in cluster 4 (CaAHL39, CaAHL40, CaAHL41, and CaAHL42) were clustered together in the phylogenetic tree, respectively, suggesting the occurrence of tandem duplications within these clusters. In summary, the CaAHLs may undergo both inter-chromosomal replication and intra-cluster duplication.
2.4. Analysis of CaAHLs Conserved Motifs
We conducted an analysis of the phylogenetic relationships and conservative motifs of CaAHL proteins in pepper. The results indicated that most CaAHLs contained motifs 1, 2, 3, and 4. Nearly all CaAHLs included motif 1 and motif 3, except for CaAHL41, which lacked motif 1, and CaAHL17 and CaAHL18, which lacked motif 3. Additionally, 82.22% and 73.33% of CaAHLs contained motif 4 and motif 2, respectively (Figure 3 and Table S1). Some CaAHLs contained multiple identical motifs. For example, CaAHL11, CaAHL12, and CaAHL15 each contained two motif 4 elements, while CaAHL31 and CaAHL4 contained two and three motif 10 elements, respectively. Additionally, we found that CaAHL11 and CaAHL15, CaAHL13 and CaAHL14, as well as CaAHL23 and CaAHL24 exhibited highly similar motif structures, which is consistent with their clustering in the phylogenetic analysis (Figure 3 and Table S1). Overall, CaAHLs in the same subgroup in phylogenetic tree had similar structures and conserved motif distributions (Figure 3 and Table S1), indicating that CaAHLs contain highly conserved amino acid residues and that CaAHLs in the same cluster may have similar roles.
2.5. Cis-Regulatory Element Analysis of the CaAHLs Promoter
Since CaAHLs play important roles in response to plant growth, development, and stress responses, we utilized PlantCARE to analyze the cis-regulatory elements in the pre-2000 bp region of the CaAHLs promoter to explore their possible functions. For the promoter of the CaAHLs, we screened cis-regulatory elements related to growth, development, stress, and hormone responses. Using TBtools, these elements were categorized into 18 distinct classes. (Figure 4 and Figure 5). The analysis revealed that the promoters of CaAHL22 and CaAHL24 are enriched with abscisic acid-responsive cis-regulatory elements, containing 5 and 4 elements, respectively. Promoters of CaAHL7, CaAHL1, and CaAHL34 exhibit a higher abundance of anaerobic-responsive elements, with 5, 4, and 4 elements, respectively. The promoter of CaAHL28 shows a significant presence of gibberellin-responsive elements. Similarly, the promoters of CaAHL23 and CaAHL4 are enriched with jasmonic acid-responsive elements, containing 7 and 5 elements, respectively. CaAHL28 and CaAHL29 exhibit higher numbers of MYB binding site-related elements, with 6 and 5 elements, respectively, while the promoter of CaAHL21 contains 4 salicylic acid-responsive elements (Figure 4 and Figure 5). These distinct distributions of cis-regulatory elements suggest potential functional differentiation among the CaAHLs.
2.6. Tissue-Specific Expression Profiles of the CaAHLs in Peppers
To explore whether the CaAHLs plays a role in the tissue development of pepper, the expression profiles of CaAHLs in the roots, stems, leaves, pericarp, and placenta were observed using RNA-seq data from the pepper line 6421 (Figure 6). The results revealed that several genes, including CaAHL1, CaAHL26, CaAHL5, CaAHL8, CaAHL10, CaAHL44, CaAHL2, and CaAHL20, exhibited root- and stem-specific expression (Figure 6). Certain genes, such as CaAHL9, CaAHL20, CaAHL27, CaAHL28, and CaAHL11 to CaAHL18, showed higher expression levels in seeds at 20 and 25 days after flowering (DAF). Some genes were specifically expressed in certain tissues or developmental stages. For instance, CaAHL29 displayed high expression in seeds at 60 DAF, suggesting a potential role in seed maturation. CaAHL36, CaAHL23, CaAHL12, and CaAHL16 to CaAHL18 exhibited elevated expression during various stages of bud development, indicating their involvement in floral development. CaAHL21 showed specific expression in the placenta at 35 DAF, implying a role in placenta development (Figure 6). These findings suggest that most CaAHL genes are specifically expressed in roots, stems, and seeds at 20 and 25 DAF, highlighting their significant roles in root, stem, and seed development. The tissue-specific expression patterns of certain genes indicate functional differentiation among the CaAHL family members.
2.7. CaAHL23 as a Potential Regulator in Pepper Male Sterility
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn. To investigate the relationship between CaAHL genes and male sterility, we focused on genes specifically expressed during flower development. Through transcriptome data and real-time quantitative data (Figure S1), we found that CaAHL23 is specifically expressed in buds and exhibits a gene expression pattern highly similar to that of the well-characterized Arabidopsis TEK gene [22], suggesting a potential role in genic male sterility. To explore this hypothesis, we first conducted a subcellular localization analysis of CaAHL23, which confirmed its nuclear localization (Figure 7a), consistent with the previously reported localization of TEK. Subsequently, yeast one-hybrid assays were performed to identify downstream genes potentially regulated by CaAHL23. These assays demonstrated that CaAHL23 binds directly to the promoter region of CaCYP703A2 (Figure 7b). Given the high sequence homology between CaCYP703A2 and the tomato sterility-related gene SlCYP703A2 (Figure S2), we hypothesize that CaAHL23 may regulate pepper fertility by modulating the expression of CaCYP703A2. Subsequently, we detected the expression of CaCYP703A2 during flower development in the sterile line material WX2A and its fertile line WX2B, and expression profile data in the wild type (Figure 8). The results showed that compared with fertile lines, the expression of CaCYP703A2 was significantly reduced in sterile lines during flower development. These findings provide new insights into the genetic mechanisms underlying male sterility in pepper and highlights CaAHL23 as a candidate gene for further functional studies.
3. Discussion
The comprehensive analysis of the CaAHL gene family presented in this study offers valuable insights into the structure, evolutionary relationships, and functional roles of these genes in pepper. The identification and characterization of 45 CaAHL genes provide a foundational understanding of their molecular properties and potential regulatory functions in plant development and fertility. The identification of 45 CaAHL genes and detailed analysis of their protein characteristics, including length, molecular weight, theoretical isoelectric point, instability index, and predicted subcellular localization, reflect the molecular diversity of the CaAHL gene family (Table 1). These findings are consistent with previous studies that demonstrated functional diversification within gene families related to plant development [24,25,26]. Chromosomal distribution analysis revealed four gene clusters (Figure 1), suggesting that tandem duplications have contributed to the expansion of the CaAHL gene family. This observation aligns with studies in other species, where tandem duplications play a critical role in the evolution and functional diversification of gene families [27,28,29].
Phylogenetic analysis grouped the 45 CaAHL genes into four distinct subgroups and highlighted both inter-chromosomal replication and intra-cluster duplication as mechanisms driving gene family expansion (Figure 2). The conservation of motif distributions among members of the same subgroup indicates functional similarities and evolutionary conservation (Figure 3), consistent with observations in other plant species [30,31]. The presence of highly conserved amino acid residues underscores the evolutionary importance of these motifs in maintaining gene function.
Cis-regulatory element analysis revealed elements related to stress response, hormone signaling, and developmental processes, suggesting that CaAHL genes may be involved in diverse regulatory networks (Figure 4 and 5). The tissue-specific expression analysis showed that many CaAHL genes are highly expressed in roots, stems, and seeds at 20 and 25 days after flowering (DAF) (Figure 6), highlighting their potential roles in organ development. Notably, the functional differentiation indicated by distinct expression patterns among family members suggests specialization in different developmental or stress response pathways, as similarly reported in studies of AHL family genes in Arabidopsis [6,8,22,32,33].
A critical finding of this study is the potential regulatory role of CaAHL23 in pepper male sterility. Subcellular localization experiments confirmed its nuclear localization (Figure 7a), consistent with the Arabidopsis TEK gene, known for its role in pollen wall development and male sterility [22]. Yeast one-hybrid assays demonstrated that CaAHL23 binds directly to the promoter region of CaCYP703A2 (Figure 7b), a gene whose homologs in Arabidopsis, cotton, and rice are essential for pollen wall formation. Knockout studies of these homologs have resulted in male sterility [34,35]. At the same time, the expression of CaCYP703A2 gene in the sterile line was significantly reduced during flower development compared with fertile lines (Figure 8). Therefore, we hypothesize that CaAHL23 regulates pepper fertility by modulating CaCYP703A2 expression, making it a promising target for further studies on fertility regulation and hybrid breeding.
Our findings provide a comprehensive view of the CaAHL gene family's structure, evolution, and functional roles in pepper. The evidence for the involvement of CaAHL23 in male sterility opens new avenues for exploring its regulatory mechanisms and potential applications in hybrid breeding. Future research should focus on functional validation of CaAHL23 and its regulatory targets through genome-editing technologies and field trials to assess its role in fertility and crop improvement.
4. Materials and Methods
4.1. Retrieval and Identification of AHL Genes in Pepper
In this study, the candidate AHL proteins were retrieved as follows: first, the protein, nucleotide, and genome sequences of Zhangshugang genomes were downloaded from the Pepper Genomics Database (http://ted.bti.cornell.edu/cgi-bin/pepper/search) [36]. Second, the HMM of AHL protein (PF03479) was downloaded from the Pfam database (http://pfam-legacy.xfam.org/) [37]. Finally, the TBtools-II (v2.149) software [38] was used to search the predicted AHL proteins using the cut-off value of the default parameter.
4.2. Sequence Analysis and Structural Characteristics
Protein lengths, molecular weights, theoretical isoelectric points (pI), instability indices, and grand average of hydropathicity of CaAHL proteins were analyzed using the TBtools-II (v2.149) software [38]. The supposed subcellular localizations of CaAHL proteins were predicted using the online tool WoLF PSORT (https://wolfpsort.hgc.jp) [39]. The protein sequences were submitted to the MEME program (https://meme-suite.org/meme/tools/meme) [40] to assess conserved motifs.
4.3. Chromosome Localization, Tandem Duplication, and Synteny Analysis
Chromosome locations and gene position in pepper were obtained by searching the Sol Genomics Network. Chromosome mapping of the CaAHL gene family was visualized using MG2C (http://mg2c.iask.in/mg2c_v2.1) [41]. Tandem duplication events were further confirmed using the following criteria: (1) the alignment length had a coverage rate of more than 70% of the full length of the CaAHL genes; (2) the identity of the aligned region was over 70%, (3) and an array of two or more genes was less than 100 kb distance. TBtools-II (v2.149) software [38] was used to analyze the synteny of CaAHL genes among the three pepper genomes.
4.4. Phylogenetic Analysis
The phylogenetic tree was generated in the following three steps: first, the CaAHL protein sequences were imported into Clustal X to produce a multiple sequence alignment file. Second, the alignment result was used to build an unrooted tree using MEGA11 with a bootstrap of 1000 replicates and neighbor-joining (NJ) methods [42]. Third, the newly produced phylogenetic tree was visualized using the Interactive Tree of Life online website (https://itol.embl.de/) [43].
4.5. RNA-Seq Analysis of CaAHL Genes
Transcriptome sequencing (RNA-seq) data of development was used to explore the distribution of gene expression in pepper (the elite Capsicum line 6421) [44] to gain insight into the expression profiles of the CaAHL gene family in different tissues across periods. The treatment methods of all samples were based on those published by Liu et al.[44]. All data of AHL genes were normalized (log2(FPKM+1)), and a heatmap was drawn using TBtools-II (v2.149) software [38].
4.6. RNA Extraction and RT-qPCR Analysis
Capsicum line 6421 was cultivated under controlled conditions at 27°C during the day and 22°C at night, with a 16-hour light and 8-hour dark photoperiod. Total RNA was isolated using the RNAprep Pure Plant Kit (DP421, TIANGEN, China) following the manufacturer’s protocol and subsequently reverse-transcribed into cDNA using the RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific, China). Quantitative real-time PCR (RT-qPCR) was performed on a QuantStudio 3 Real-Time PCR System (ABI Life, USA) using 2× ChamQ Universal SYBR qPCR Master Mix (TransGen, China) in accordance with the provided instructions. The CaActin7 gene was used as the reference gene, with the forward primer sequence 5′-CTCGAGCAGTGTTTCCCAGT-3′ and the reverse primer sequence 5′-AGCTTCATCACCCACATAGGC-3′. Gene-specific primers were designed for CaAHL23 and CaCYP703A2 as follows: CaAHL23 forward primer, 5′-GAGTCTAGCGGTGGACCTAT-3′, and reverse primer, 5′-AGGTTCTCTCATCTCCAGGG-3′; CaCYP703A2 forward primer, 5′-TCAATCGACTCCCTCCTGGT-3′, and reverse primer, 5′-CGGAGATAGACCAATGGCCC-3′. RT-qPCR experiments were conducted with three biological replicates and three technical replicates per sample. Relative gene expression levels were determined using the 2^−ΔΔCt method.
4.7. Subcellular Localization
The full-length ORF sequences of CaAHL23 without the termination codon were cloned into the pCAMBIA1300-GFP vector and transformed into Agrobacterium tumefaciens GV3101. CaAHL23 fusion constructs was transformed into tobacco (Nicotiana benthamiana) leaves. After three days, the fluorescence signals were observed and captured using a confocal laser scanning microscope (Zeiss LSM510 META, Germany).
4.8. Y1H Assays
Pepper floral buds (F1-F9) were collected for cDNA library construction via a Matchmaker GAL4 One-Hybrid System (Clontech) following the manufacturer’s instructions. A 244-bp fragment (2769-3012 upstream of ATG) from the CaCYP703A2 promoter was synthesized by Tsingke Tech (Beijing, China) and subcloned and inserted into a pAbAi vector, and the pCaCYP703A2pro-AbAi bait was transformed into the yeast strain Y1HGold. Y1H library screening assays were performed against the cDNA expression library using the Matchmaker GAL4 One Hybrid System (Clontech). Competent Y1HGold yeast cells containing pCaCYP703A2pro-AbAi as bait were transformed with the contents of the cDNA library, plated onto selective SD/-Leu media supplemented with 100 ng/mL AbA, and incubated at 30℃. The p53-AbAi Control Vector and pGADT7-Rec53 Control Insert from Clontech were used as the positive controls. Yeast plasmids from positive colonies were extracted and transformed into Escherichia coli DH5α competent cells. The prey fragments from the positive colonies were identified by Sanger sequencing. To verify the interaction of CaAHL23 with the bait, the full-length cDNA sequences of CaAHL23 were amplified and fused in frame with the GAL4 activation domain of pGADT7rec (Clontech), forming pGADT7-CaAHL23.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Expression level analysis of CaAHL23 during flower development; Figure S2: CDS and protein sequence alignment of CaCYP703A2 and SlCYP703A2; Table S1: Number statistics of conserved motifs in the CaAHLs.
Author Contributions
Conceptualization, XY.S. and YL.L.; methodology, X.W.; software, XY.S.; validation, YL.L., Y.Z. and XY.S.; formal analysis, QZ.C. and Y.L.; investigation, BQ.T.; resources, F.L. and XX.Z.; data curation, XY.S. and YL.L.; writing—original draft preparation, XY.S.; writing—review and editing, YL.L. and F.L. and XX.Z.; supervision, YL.L.; project administration, F.L. and XX.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by China Agriculture Research System of MOF and MARA (CARS-24-A-15).
Data Availability Statement
Data is contained within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, S.; Park, M.; Yeom, S.-I.; Kim, Y.-M.; Lee, J.M.; Lee, H.-A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.-T.; et al. Genome Sequence of the Hot Pepper Provides Insights into the Evolution of Pungency in Capsicum Species. Nat Genet 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, U.; Li, X.; Fan, Y.; Chang, W.; Niu, Y.; Li, J.; Qu, C.; Lu, K. Multi-Omics Revolution to Promote Plant Breeding Efficiency. Front Plant Sci 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Islam, K.; Momo, J.; Rawoof, A.; Vijay, A.; Anusree, V.K.; Kumar, A.; Ramchiary, N. Integrated Use of Molecular and Omics Approaches for Breeding High Yield and Stress Resistance Chili Peppers BT - Smart Plant Breeding for Vegetable Crops in Post-Genomics Era. In; Singh, S., Sharma, D., Sharma, S.K., Singh, R., Eds.; Springer Nature Singapore: Singapore, 2023; pp. 279–335. ISBN 978-981-19-5367-5. [Google Scholar]
- Xu, L.; Zheng, S.; Witzel, K.; Van De Slijke, E.; Baekelandt, A.; Mylle, E.; Van Damme, D.; Cheng, J.; De Jaeger, G.; Inzé, D.; et al. Chromatin Attachment to the Nuclear Matrix Represses Hypocotyl Elongation in Arabidopsis Thaliana. Nat Commun 2024, 15, 1286. [Google Scholar] [CrossRef]
- Street, I.H.; Shah, P.K.; Smith, A.M.; Avery, N.; Neff, M.M. The AT-Hook-Containing Proteins SOB3/AHL29 and ESC/AHL27 Are Negative Modulators of Hypocotyl Growth in Arabidopsis. The Plant Journal 2008, 54, 1–14. [Google Scholar] [CrossRef]
- Xiao, C.; Chen, F.; Yu, X.; Lin, C.; Fu, Y.-F. Over-Expression of an AT-Hook Gene, AHL22, Delays Flowering and Inhibits the Elongation of the Hypocotyl in Arabidopsis Thaliana. Plant Mol Biol 2009, 71, 39–50. [Google Scholar] [CrossRef]
- Uzair, M.; Xu, D.; Schreiber, L.; Shi, J.; Liang, W.; Jung, K.-H.; Chen, M.; Luo, Z.; Zhang, Y.; Yu, J.; et al. PERSISTENT TAPETAL CELL2 Is Required for Normal Tapetal Programmed Cell Death and Pollen Wall Patterning1 [OPEN]. Plant Physiol 2020, 182, 962–976. [Google Scholar] [CrossRef]
- Tayengwa, R.; Sharma Koirala, P.; Pierce, C.F.; Werner, B.E.; Neff, M.M. Overexpression of AtAHL20 Causes Delayed Flowering in Arabidopsis via Repression of FT Expression. BMC Plant Biol 2020, 20, 559. [Google Scholar] [CrossRef]
- Machaj, G.; Grzebelus, D. Characteristics of the AT-Hook Motif Containing Nuclear Localized (AHL) Genes in Carrot Provides Insight into Their Role in Plant Growth and Storage Root Development. Genes (Basel) 2021, 12. [Google Scholar] [CrossRef]
- Feng, H.; Yang, H.; Sun, H.; Lu, P.; Yan, P.; Zhao, W.; Zeng, L.; Li, Z.; Zhang, L.; Hou, W. Characterization of AHL Transcription Factors and Functional Analysis of IbAHL10 in Storage Root Development in Sweetpotato. Sci Hortic 2024, 338, 113718. [Google Scholar] [CrossRef]
- Favero, D.S.; Kawamura, A.; Shibata, M.; Takebayashi, A.; Jung, J.-H.; Suzuki, T.; Jaeger, K.E.; Ishida, T.; Iwase, A.; Wigge, P.A.; et al. AT-Hook Transcription Factors Restrict Petiole Growth by Antagonizing PIFs. Current Biology 2020, 30, 1454–1466e6. [Google Scholar] [CrossRef]
- Jeong, H.-N.; Sun, H.-J.; Zuo, Z.-F.; Lee, D.H.; Song, P.-S.; Kang, H.-G.; Lee, H.-Y. Overexpression of ATHG1/AHL23 and ATPG3/AHL20, Arabidopsis AT-Hook Motif Nuclear-Localized Genes, Confers Salt Tolerance in Transgenic Zoysia Japonica. Plant Biotechnol Rep 2020, 14, 351–361. [Google Scholar] [CrossRef]
- Howden, A.J.M.; Stam, R.; Martinez Heredia, V.; Motion, G.B.; ten Have, S.; Hodge, K.; Marques Monteiro Amaro, T.M.; Huitema, E. Quantitative Analysis of the Tomato Nuclear Proteome during Phytophthora Capsici Infection Unveils Regulators of Immunity. New Phytologist 2017, 215, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Rayapuram, N.; Jarad, M.; Alhoraibi, H.M.; Bigeard, J.; Abulfaraj, A.A.; Völz, R.; Mariappan, K.G.; Almeida-Trapp, M.; Schlöffel, M.; Lastrucci, E.; et al. Chromatin Phosphoproteomics Unravels a Function for AT-Hook Motif Nuclear Localized Protein AHL13 in PAMP-Triggered Immunity. Proceedings of the National Academy of Sciences 2021, 118, e2004670118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Liu, Z.; Liu, Y.; Kong, D.; Li, T.; Yu, S.; Mei, H.; Xu, X.; Liu, H.; Chen, L.; et al. A Novel Gene OsAHL1 Improves Both Drought Avoidance and Drought Tolerance in Rice. Sci Rep 2016, 6, 30264. [Google Scholar] [CrossRef]
- Zhao, L.; Lü, Y.; Chen, W.; Yao, J.; Li, Y.; Li, Q.; Pan, J.; Fang, S.; Sun, J.; Zhang, Y. Genome-Wide Identification and Analyses of the AHL Gene Family in Cotton (Gossypium). BMC Genomics 2020, 21, 69. [Google Scholar] [CrossRef]
- Wang, M.; Chen, B.; Zhou, W.; Xie, L.; Wang, L.; Zhang, Y.; Zhang, Q. Genome-Wide Identification and Expression Analysis of the AT-Hook Motif Nuclear Localized Gene Family in Soybean. BMC Genomics 2021, 22, 361. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Mishra, A. Genome-Wide Identification and Analyses of the AHL Gene Family in Rice (Oryza Sativa). 3 Biotech 2023, 13, 248. [Google Scholar] [CrossRef]
- Bishop, E.H.; Kumar, R.; Luo, F.; Saski, C.; Sekhon, R.S. Genome-Wide Identification, Expression Profiling, and Network Analysis of AT-Hook Gene Family in Maize. Genomics 2020, 112, 1233–1244. [Google Scholar] [CrossRef]
- Yun, J.; Kim, Y.-S.; Jung, J.-H.; Seo, P.J.; Park, C.-M. The AT-Hook Motif-Containing Protein AHL22 Regulates Flowering Initiation by Modifying FLOWERING LOCUS T Chromatin in Arabidopsis*. Journal of Biological Chemistry 2012, 287, 15307–15316. [Google Scholar] [CrossRef]
- Xu, Y.; Gan, E.-S.; Ito, T. The AT-Hook/PPC Domain Protein TEK Negatively Regulates Floral Repressors Including MAF4 and MAF5. Plant Signal Behav 2013, 8, e25006. [Google Scholar] [CrossRef]
- Lou, Y.; Xu, X.-F.; Zhu, J.; Gu, J.-N.; Blackmore, S.; Yang, Z.-N. The Tapetal AHL Family Protein TEK Determines Nexine Formation in the Pollen Wall. Nat Commun 2014, 5, 3855. [Google Scholar] [CrossRef] [PubMed]
- Karami, O.; Rahimi, A.; Mak, P.; Horstman, A.; Boutilier, K.; Compier, M.; van der Zaal, B.; Offringa, R. An Arabidopsis AT-Hook Motif Nuclear Protein Mediates Somatic Embryogenesis and Coinciding Genome Duplication. Nat Commun 2021, 12, 2508. [Google Scholar] [CrossRef] [PubMed]
- Karlgren, A.; Gyllenstrand, N.; Källman, T.; Sundström, J.F.; Moore, D.; Lascoux, M.; Lagercrantz, U. Evolution of the PEBP Gene Family in Plants: Functional Diversification in Seed Plant Evolution. Plant Physiol 2011, 156, 1967–1977. [Google Scholar] [CrossRef]
- Ahmad, Z.; Tian, D.; Li, Y.; Aminu, I.M.; Tabusam, J.; Zhang, Y.; Zhu, S. Characterization, Evolution, Expression and Functional Divergence of the DMP Gene Family in Plants. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Hofer, J.; Ellis, N. Conservation and Diversification of Gene Function in Plant Development. Curr Opin Plant Biol 2002, 5, 56–61. [Google Scholar] [CrossRef]
- Feng, L.-Y.; Lin, P.-F.; Xu, R.-J.; Kang, H.-Q.; Gao, L.-Z. Comparative Genomic Analysis of Asian Cultivated Rice and Its Wild Progenitor (Oryza Rufipogon) Has Revealed Evolutionary Innovation of the Pentatricopeptide Repeat Gene Family through Gene Duplication. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Qanmber, G.; Liu, J.; Yu, D.; Liu, Z.; Lu, L.; Mo, H.; Ma, S.; Wang, Z.; Yang, Z. Genome-Wide Identification and Characterization of the PERK Gene Family in Gossypium Hirsutum Reveals Gene Duplication and Functional Divergence. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Qiao, X.; Li, Q.; Yin, H.; Qi, K.; Li, L.; Wang, R.; Zhang, S.; Paterson, A.H. Gene Duplication and Evolution in Recurring Polyploidization–Diploidization Cycles in Plants. Genome Biol 2019, 20, 38. [Google Scholar] [CrossRef]
- Chen, G.; He, W.; Guo, X.; Pan, J. Genome-Wide Identification, Classification and Expression Analysis of the MYB Transcription Factor Family in Petunia. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Cao, L.; Lu, X.; Zhang, P.; Wang, G.; Wei, L.; Wang, T. Systematic Analysis of Differentially Expressed Maize ZmbZIP Genes between Drought and Rewatering Transcriptome Reveals BZIP Family Members Involved in Abiotic Stress Responses. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, X.; Chen, J.; Guo, X.; Wang, H.; Zhen, W.; Zhang, J.; Hu, Z.; Zhang, X.; Botella, J.R.; et al. Overexpression of AHL9 Accelerates Leaf Senescence in Arabidopsis Thaliana. BMC Plant Biol 2022, 22, 248. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Favero, D.S.; Peng, H.; Neff, M.M. Arabidopsis Thaliana AHL Family Modulates Hypocotyl Growth Redundantly by Interacting with Each Other via the PPC/DUF296 Domain. Proceedings of the National Academy of Sciences 2013, 110, E4688–E4697. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wu, D.; Shi, J.; He, Y.; Pinot, F.; Grausem, B.; Yin, C.; Zhu, L.; Chen, M.; Luo, Z.; et al. Rice CYP703A3, a Cytochrome P450 Hydroxylase, Is Essential for Development of Anther Cuticle and Pollen Exine. J Integr Plant Biol 2014, 56, 979–994. [Google Scholar] [CrossRef]
- Ma, H.; Wu, Y.; Lv, R.; Chi, H.; Zhao, Y.; Li, Y.; Liu, H.; Ma, Y.; Zhu, L.; Guo, X.; et al. Cytochrome P450 Mono-Oxygenase CYP703A2 Plays a Central Role in Sporopollenin Formation and Ms5ms6 Fertility in Cotton. J Integr Plant Biol 2022, 64, 2009–2025. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhao, J.; Sun, H.; Xiong, C.; Sun, X.; Wang, X.; Wang, Z.; Jarret, R.; Wang, J.; Tang, B.; et al. Genomes of Cultivated and Wild Capsicum Species Provide Insights into Pepper Domestication and Population Differentiation. Nat Commun 2023, 14, 5487. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “One for All, All for One” Bioinformatics Platform for Biological Big-Data Mining. Mol Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein Localization Predictor. Nucleic Acids Res 2007, 35, W585–W587. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a Mixture Model by Expectation Maximization to Discover Motifs in Bipolymers. Proc Int Conf Intell Syst Mol Biol 1994, 2, 28–36. [Google Scholar]
- Chao, J.-T.; Kong, Y.-Z.; Wang, Q.; Sun, Y.-H.; Gong, D.-P.; Lv, J.; Liu, G.-S. MapGene2Chrom, a Tool to Draw Gene Physical Map Based on Perl and SVG Languages. Yi Chuan 2015, 37, 91–97. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yu, H.; Deng, Y.; Zheng, J.; Liu, M.; Ou, L.; Yang, B.; Dai, X.; Ma, Y.; Feng, S.; et al. PepperHub, an Informatics Hub for the Chili Pepper Research Community. Mol Plant 2017, 10, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chromosome mapping and gene number of CaAHLs in Zhangshugang genome. (a) Chromosome mapping of CaAHLs. Chromosome numbers are represented on the top, and the scale is shown on the left. (b) Statistical analysis for CaAHLs on 12 chromosomes.
Figure 1.
Chromosome mapping and gene number of CaAHLs in Zhangshugang genome. (a) Chromosome mapping of CaAHLs. Chromosome numbers are represented on the top, and the scale is shown on the left. (b) Statistical analysis for CaAHLs on 12 chromosomes.
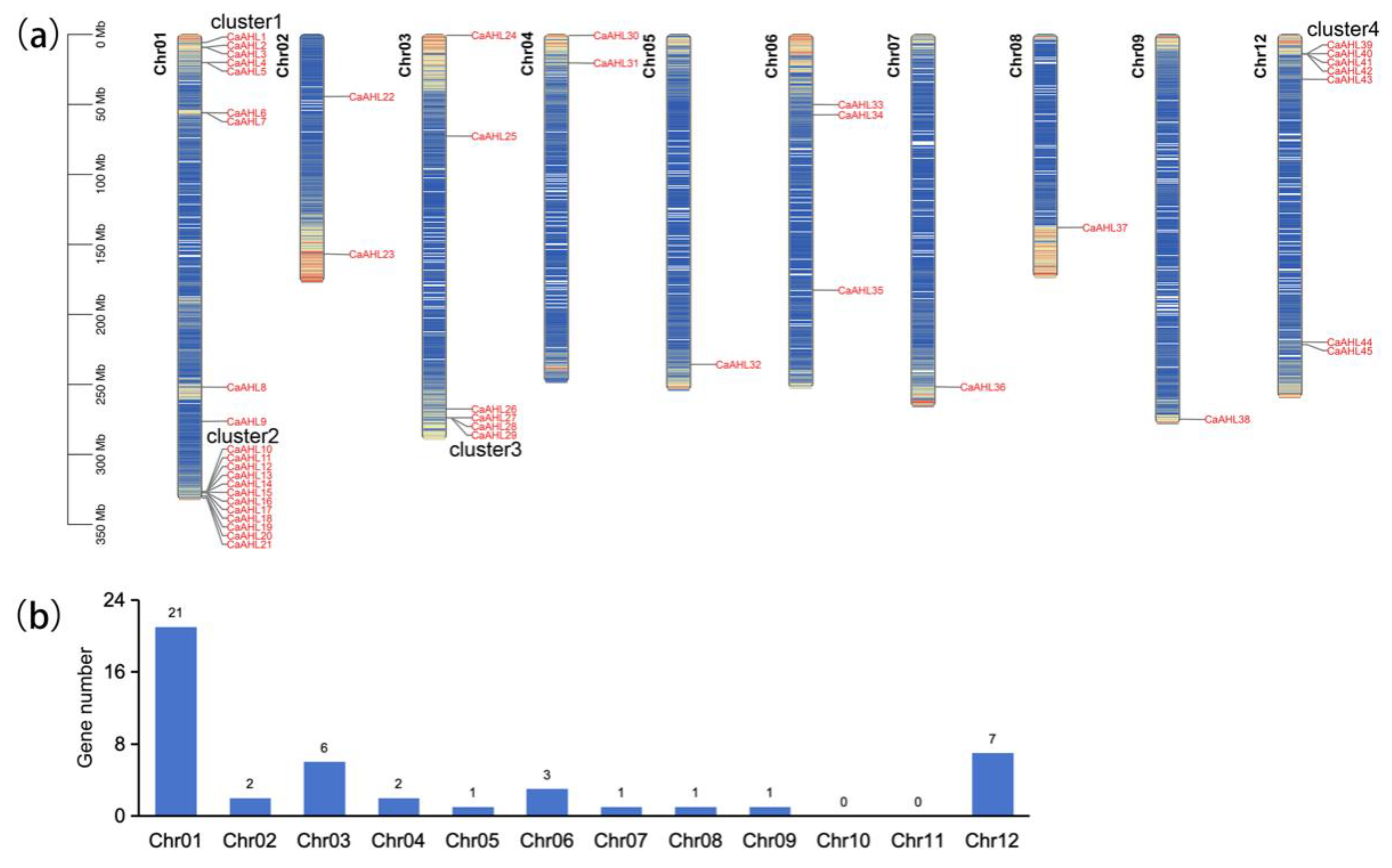
Figure 2.
Phylogenetic relationships among CaAHLs in the Zhangshugang genome. Branches are shaded in different colors to distinguish groups. Black dots indicate bootstrap support values, with only those greater than 0.5 displayed. Predicted tandem duplicate gene pairs are highlighted using various font colors: yellow for genes in cluster 1, red for cluster 2, blue for cluster 3, and purple for cluster 4.
Figure 2.
Phylogenetic relationships among CaAHLs in the Zhangshugang genome. Branches are shaded in different colors to distinguish groups. Black dots indicate bootstrap support values, with only those greater than 0.5 displayed. Predicted tandem duplicate gene pairs are highlighted using various font colors: yellow for genes in cluster 1, red for cluster 2, blue for cluster 3, and purple for cluster 4.
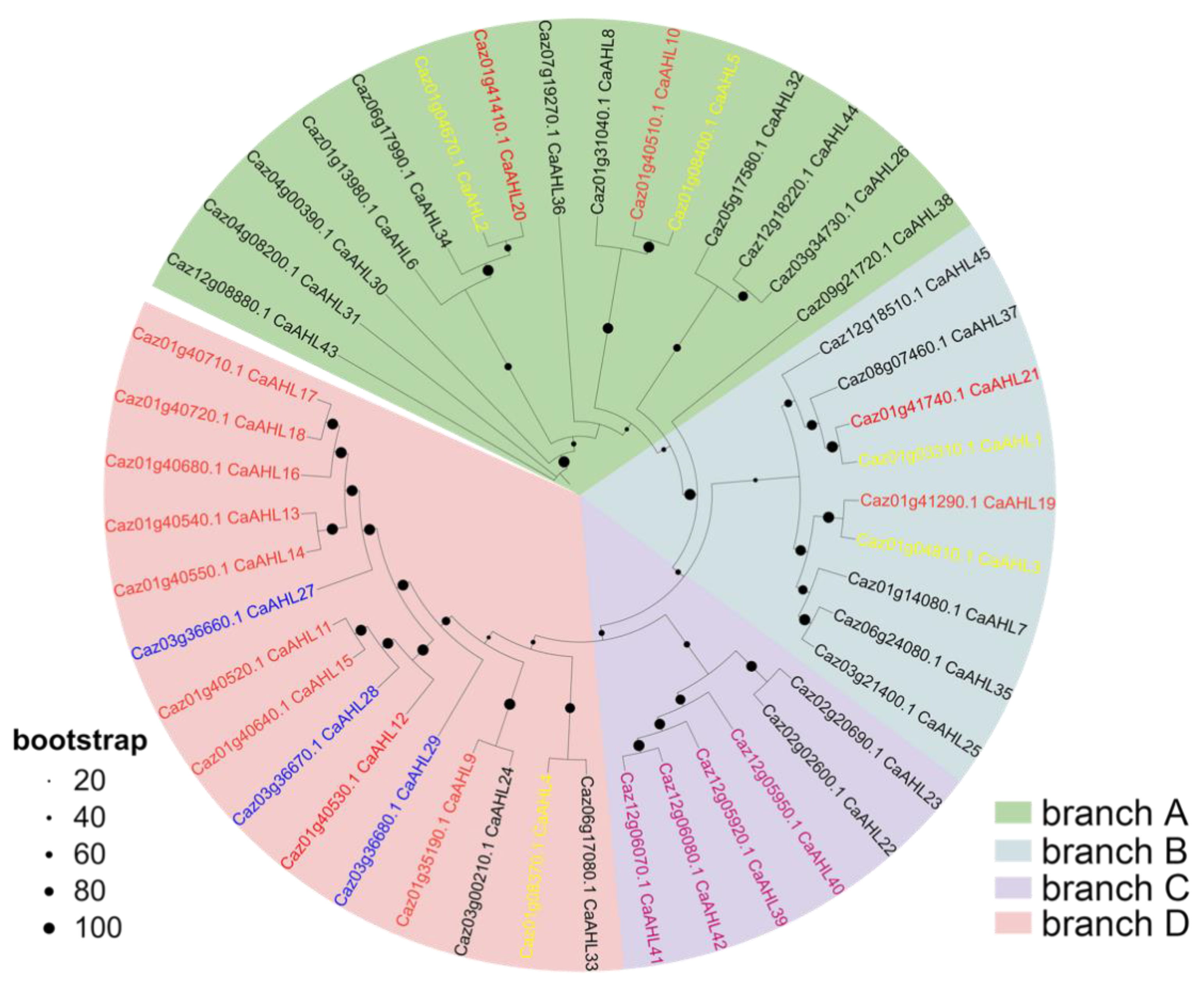
Figure 3.
Conserved motif analysis of CaAHLs. The evolutionary tree on the left is constructed based on the protein sequences of CaAHLs.
Figure 3.
Conserved motif analysis of CaAHLs. The evolutionary tree on the left is constructed based on the protein sequences of CaAHLs.
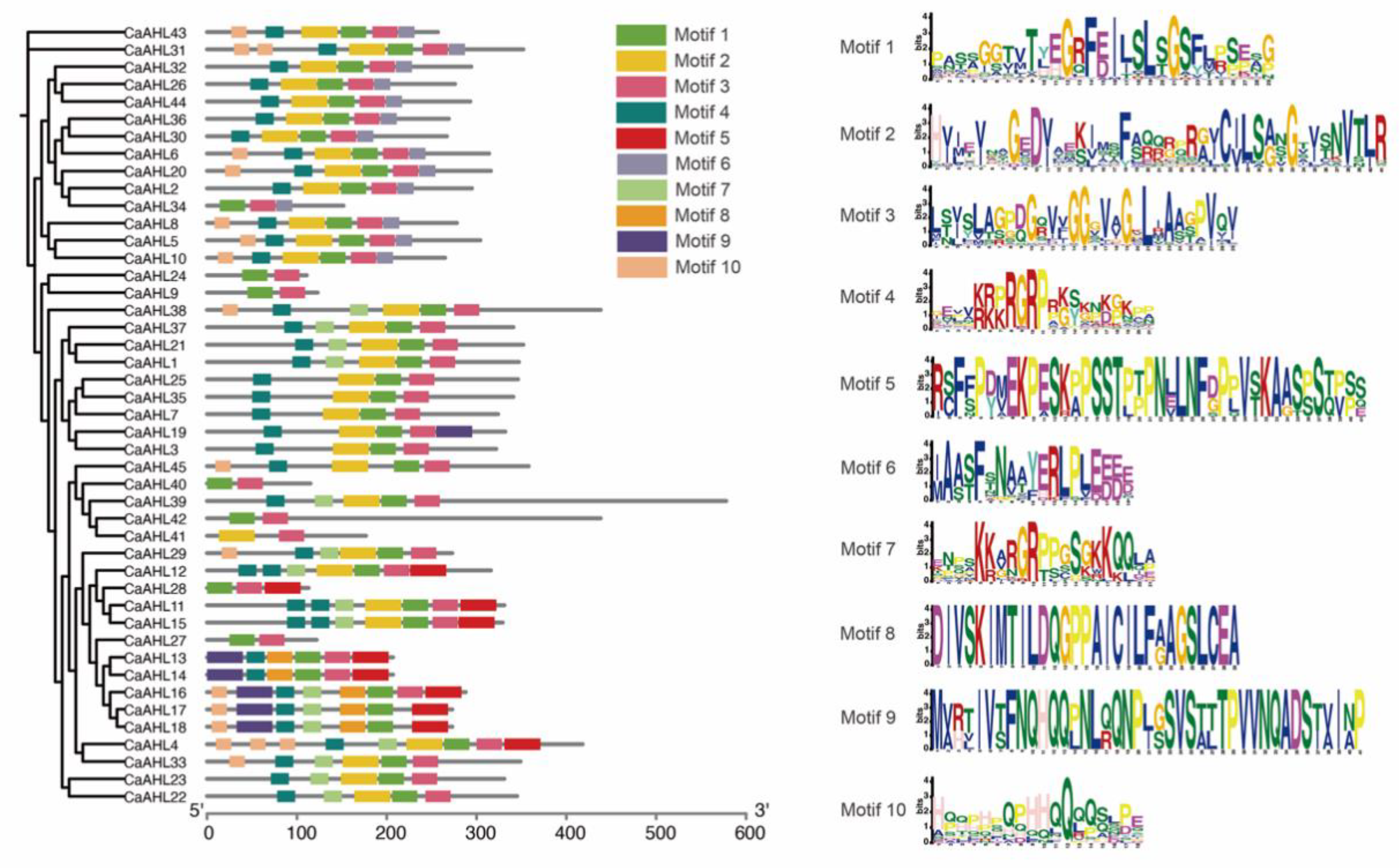
Figure 4.
The distribution of cis-regulatory elements predicted in the CaAHLs promoter. Different colored boxes represent different cis-regulatory elements. The evolutionary tree on the left is constructed based on the protein sequences of CaAHLs.
Figure 4.
The distribution of cis-regulatory elements predicted in the CaAHLs promoter. Different colored boxes represent different cis-regulatory elements. The evolutionary tree on the left is constructed based on the protein sequences of CaAHLs.
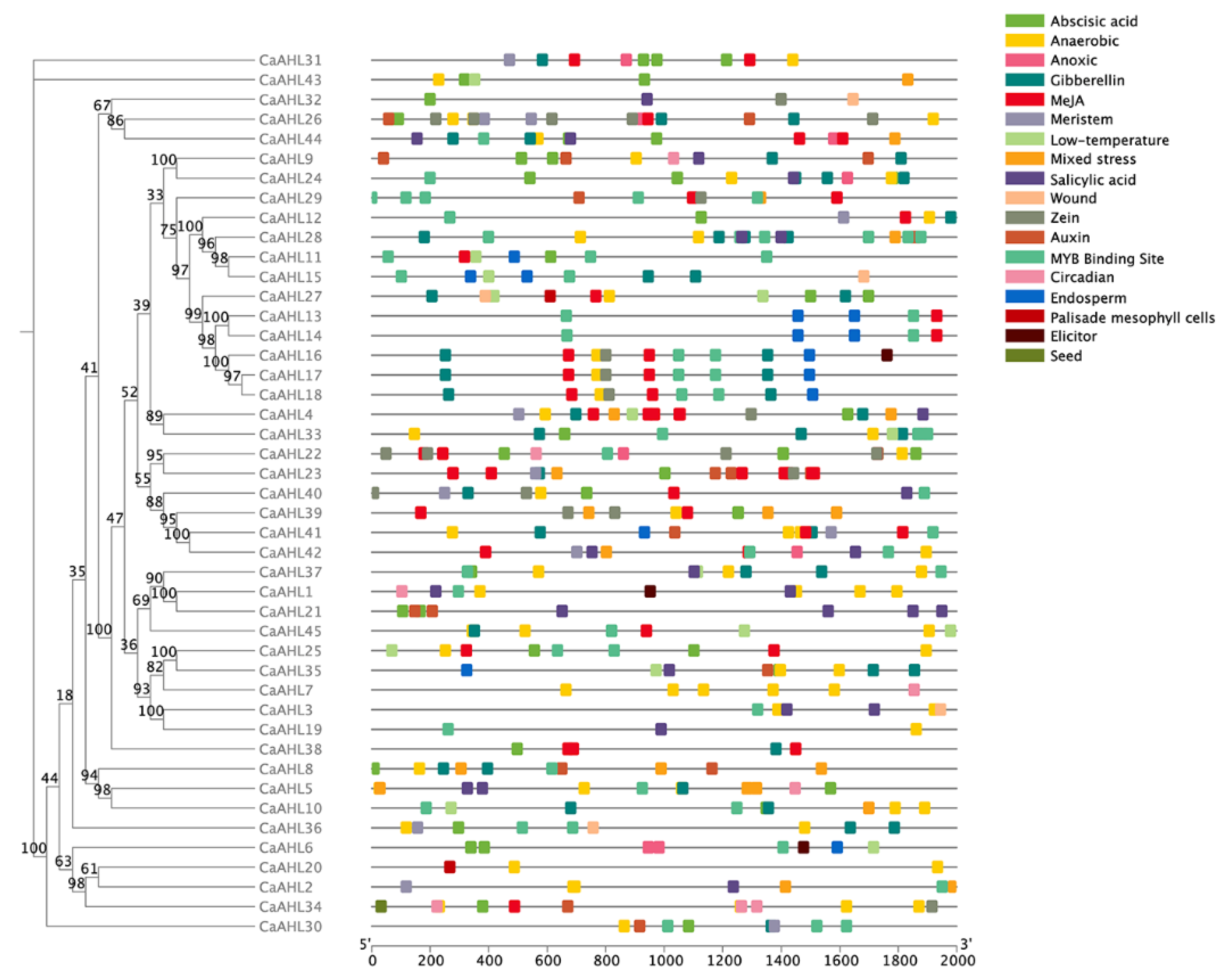
Figure 5.
Number statistics of cis-regulatory elements in the CaAHLs promoter regions. The evolutionary tree on the left is based on the analysis of the number and types of cis-regulatory elements.
Figure 5.
Number statistics of cis-regulatory elements in the CaAHLs promoter regions. The evolutionary tree on the left is based on the analysis of the number and types of cis-regulatory elements.
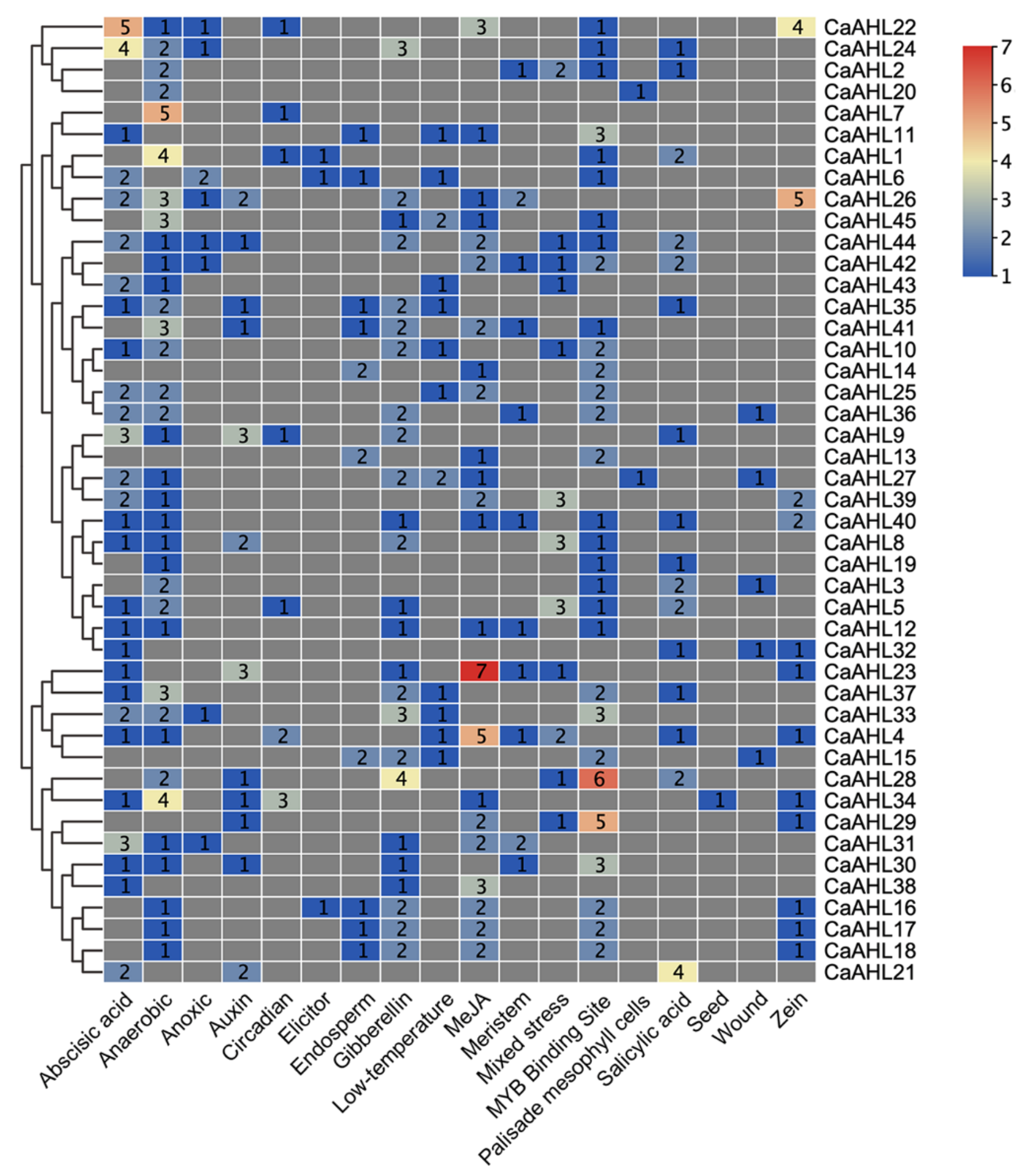
Figure 6.
Expression profile analysis of CaAHLs in various tissues and organs of pepper. Expression levels were determined in the following tissues and stages: leaf tissues were sampled at 60 days after emergence and marked correspondingly as AL; Stems and roots were marked AS and AR, respectively. Floral buds were sampled at 0.25, 0.35, 0.5, 0.8, 1.0, 1.2, and 1.7 cm, and marked correspondingly as F1, F2, F3, F4, F5, F6, F7, F8, and F9; fruits were collected at 3, 7, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, and 60 days after flowering (DAF), and marked correspondingly as FST0, FST1, G1, G2, G3, G4, G5, G6, G7, G8, G9, G10, G11; seed samples were collected at 10, 15, 20, 25, 30, 35, 40, 45, 50, 55 and 60 DAF, and marked correspondingly as ST1, ST2, S3, S4, S5, S6, S7, S8, S9, S10, S11; placenta samples were collected at 20, 25, 30, 35, 40, 45, 50, 55 and 60 DAF, and marked correspondingly as T3, T4, T5, T6, T7, T8, T9, T10, and T11. The FPKM values were log2-transformed, and a heatmap was generated using TBtools-II (v2.149) software. The evolutionary tree on the left is constructed based on gene expression levels. Expression values on the right are shown as a color gradient from low expression (blue) to high expression (red).
Figure 6.
Expression profile analysis of CaAHLs in various tissues and organs of pepper. Expression levels were determined in the following tissues and stages: leaf tissues were sampled at 60 days after emergence and marked correspondingly as AL; Stems and roots were marked AS and AR, respectively. Floral buds were sampled at 0.25, 0.35, 0.5, 0.8, 1.0, 1.2, and 1.7 cm, and marked correspondingly as F1, F2, F3, F4, F5, F6, F7, F8, and F9; fruits were collected at 3, 7, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, and 60 days after flowering (DAF), and marked correspondingly as FST0, FST1, G1, G2, G3, G4, G5, G6, G7, G8, G9, G10, G11; seed samples were collected at 10, 15, 20, 25, 30, 35, 40, 45, 50, 55 and 60 DAF, and marked correspondingly as ST1, ST2, S3, S4, S5, S6, S7, S8, S9, S10, S11; placenta samples were collected at 20, 25, 30, 35, 40, 45, 50, 55 and 60 DAF, and marked correspondingly as T3, T4, T5, T6, T7, T8, T9, T10, and T11. The FPKM values were log2-transformed, and a heatmap was generated using TBtools-II (v2.149) software. The evolutionary tree on the left is constructed based on gene expression levels. Expression values on the right are shown as a color gradient from low expression (blue) to high expression (red).
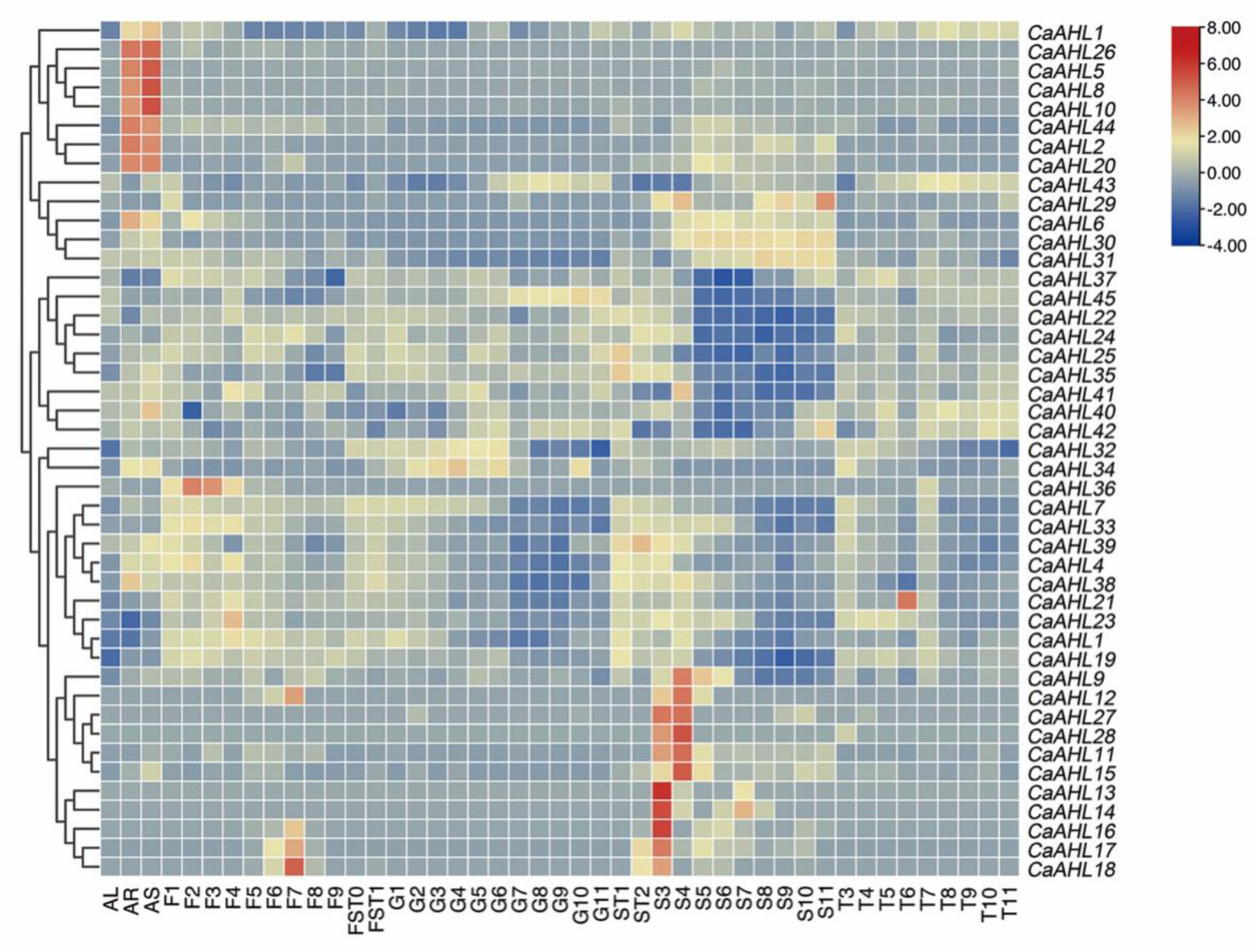
Figure 7.
Subcellular localization (a) and yeast one-hybrid assay (b) of CaAHL23.
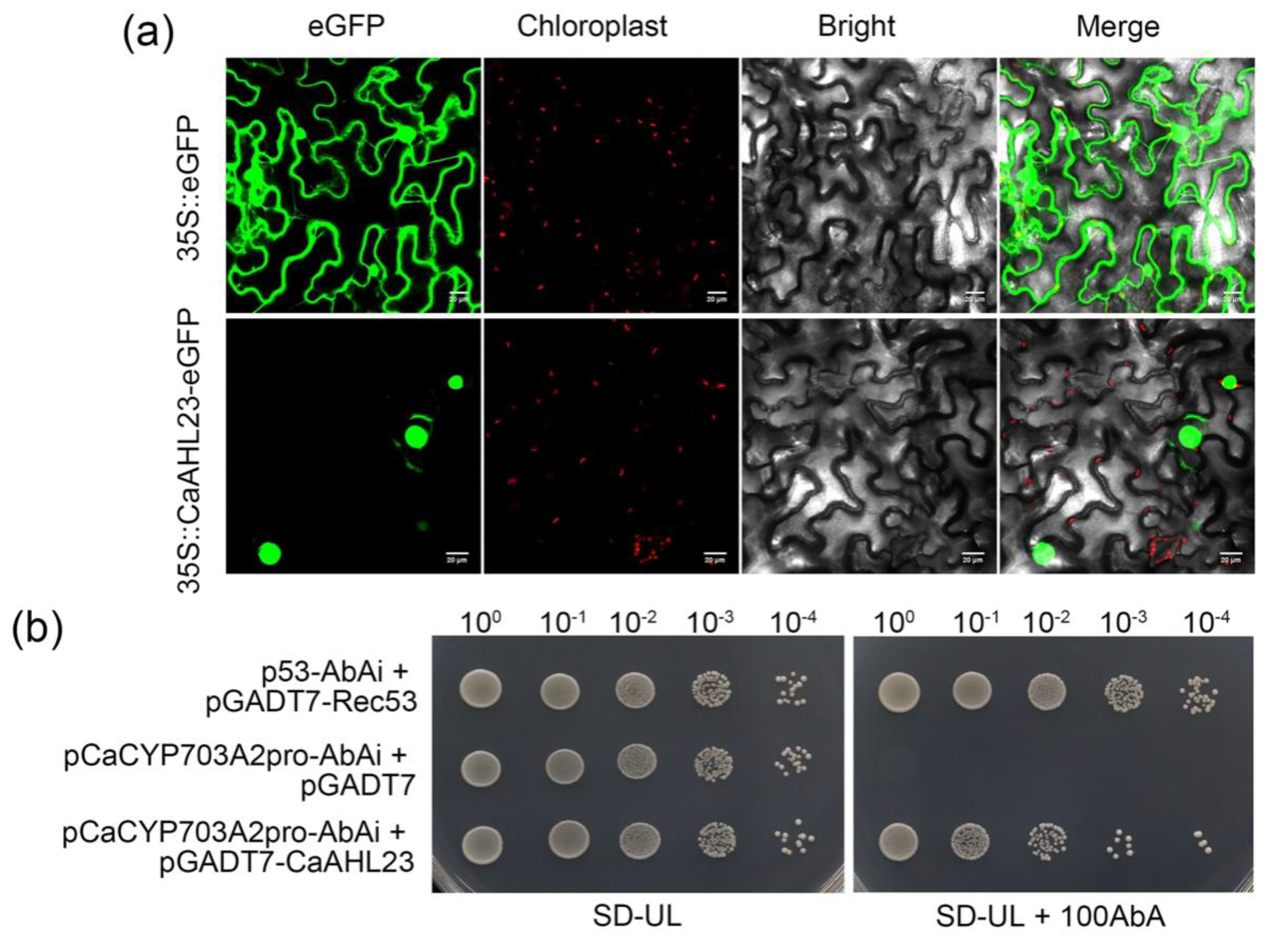
Figure 8.
Expression level analysis of CaCYP703A2 during flower development. Floral buds were sampled at 0.25, 0.35, 0.5, 0.8, 1.0, 1.2, and 1.7 cm, and marked correspondingly as F1, F2, F3, F4, F5, F6, F7, F8, and F9.
Figure 8.
Expression level analysis of CaCYP703A2 during flower development. Floral buds were sampled at 0.25, 0.35, 0.5, 0.8, 1.0, 1.2, and 1.7 cm, and marked correspondingly as F1, F2, F3, F4, F5, F6, F7, F8, and F9.
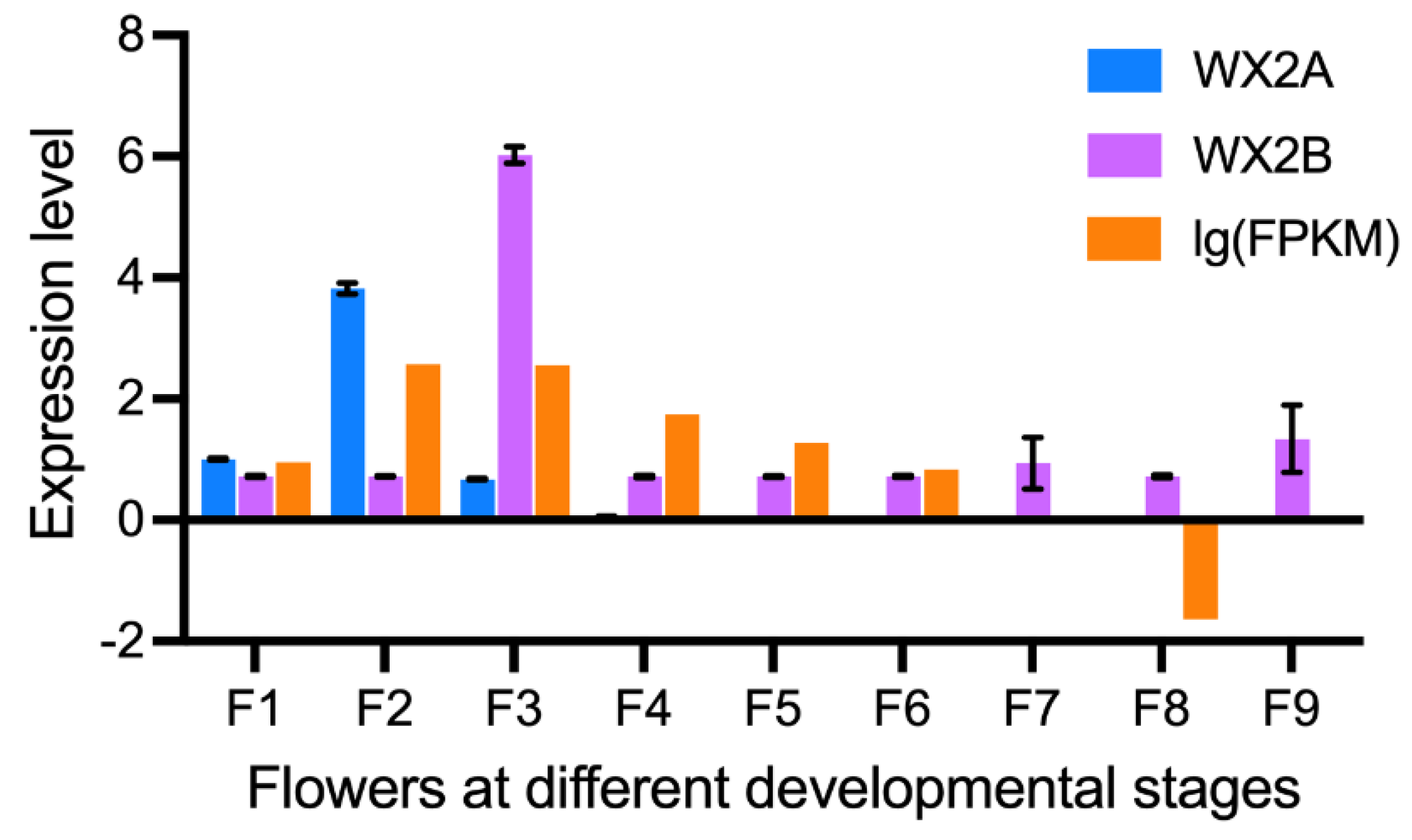

Table 1.
Basic information of AHL genes identified in pepper.
| Name | Gene ID | Chromosome location | Protein length | Molecular weight (kDa) | Theoretical pI | Instability index | Grand average of hydropathicity | Subcellular localization1 |
|---|---|---|---|---|---|---|---|---|
| CaAHL1 | Caz01g03310.1 | Chr01:5691094-5697294 | 347 | 35.27 | 9.62 | 38.59 | -0.33 | nucl |
| CaAHL2 | Caz01g04670.1 | Chr01:9114966-9116603 | 295 | 31.17 | 5.79 | 52.67 | -0.54 | nucl |
| CaAHL3 | Caz01g04810.1 | Chr01:9383031-9386809 | 322 | 33.18 | 8.74 | 50.79 | -0.17 | chlo |
| CaAHL4 | Caz01g08370.1 | Chr01:20063956-20073612 | 418 | 43.66 | 8.82 | 60.17 | -0.76 | nucl |
| CaAHL5 | Caz01g08400.1 | Chr01:20111560-20112474 | 304 | 31.69 | 5.75 | 51.83 | -0.27 | nucl |
| CaAHL6 | Caz01g13980.1 | Chr01:55893363-55894391 | 314 | 33.10 | 6.83 | 53.54 | -0.60 | nucl |
| CaAHL7 | Caz01g14080.1 | Chr01:56075436-56079692 | 324 | 33.72 | 9.57 | 47.83 | -0.25 | chlo |
| CaAHL8 | Caz01g31040.1 | Chr01:251924071-251924907 | 278 | 29.43 | 6.54 | 50.25 | -0.47 | chlo |
| CaAHL9 | Caz01g35190.1 | Chr01:276409484-276418256 | 123 | 13.45 | 5.87 | 47.09 | -0.20 | cyto |
| CaAHL10 | Caz01g40510.1 | Chr01:326409380-326414186 | 265 | 28.25 | 5.96 | 44.58 | -0.44 | nucl |
| CaAHL11 | Caz01g40520.1 | Chr01:326434617-326442536 | 331 | 35.20 | 9.51 | 49.49 | -0.46 | chlo |
| CaAHL12 | Caz01g40530.1 | Chr01:326492721-326618469 | 316 | 33.80 | 10.04 | 51.86 | -0.37 | nucl |
| CaAHL13 | Caz01g40540.1 | Chr01:326621531-326626277 | 207 | 22.30 | 6.96 | 44.75 | -0.13 | nucl |
| CaAHL14 | Caz01g40550.1 | Chr01:326687022-326691770 | 207 | 22.34 | 7.84 | 44.38 | -0.14 | chlo |
| CaAHL15 | Caz01g40640.1 | Chr01:327024609-327031508 | 329 | 34.72 | 9.65 | 48.09 | -0.47 | chlo |
| CaAHL16 | Caz01g40680.1 | Chr01:327253871-327258557 | 288 | 30.70 | 8.76 | 47.03 | -0.45 | chlo |
| CaAHL17 | Caz01g40710.1 | Chr01:327378313-327383011 | 273 | 29.34 | 7.79 | 47.92 | -0.53 | chlo |
| CaAHL18 | Caz01g40720.1 | Chr01:327501110-327505809 | 273 | 29.30 | 7.79 | 51.24 | -0.53 | chlo |
| CaAHL19 | Caz01g41290.1 | Chr01:329720440-329727085 | 332 | 34.13 | 9.54 | 44.92 | -0.10 | chlo |
| CaAHL20 | Caz01g41410.1 | Chr01:329900501-329901727 | 316 | 33.79 | 6.05 | 59.8 | -0.63 | chlo |
| CaAHL21 | Caz01g41740.1 | Chr01:331181444-331186914 | 352 | 35.75 | 9.44 | 49.49 | -0.27 | nucl |
| CaAHL22 | Caz02g02600.1 | Chr02:44359225-44368231 | 345 | 34.69 | 8.97 | 55.44 | -0.11 | E.R. |
| CaAHL23 | Caz02g20690.1 | Chr02:156842236-156848762 | 331 | 33.33 | 9.99 | 50.43 | -0.23 | nucl |
| CaAHL24 | Caz03g00210.1 | Chr03:551605-558146 | 111 | 12.02 | 6.39 | 30.43 | 0.01 | chlo |
| CaAHL25 | Caz03g21400.1 | Chr03:72579177-72587401 | 346 | 36.17 | 6.33 | 45.59 | -0.38 | nucl |
| CaAHL26 | Caz03g34730.1 | Chr03:267518820-267519650 | 276 | 28.85 | 5.45 | 57.84 | -0.47 | cyto |
| CaAHL27 | Caz03g36660.1 | Chr03:273808903-273810928 | 122 | 12.87 | 5.19 | 34.02 | 0.18 | cyto |
| CaAHL28 | Caz03g36670.1 | Chr03:273824211-273825851 | 113 | 12.01 | 6.01 | 47.05 | -0.25 | nucl |
| CaAHL29 | Caz03g36680.1 | Chr03:273829352-273850851 | 273 | 28.76 | 9.64 | 49.08 | -0.30 | vacu |
| CaAHL30 | Caz04g00390.1 | Chr04:713251-718475 | 267 | 26.36 | 6.42 | 48.45 | -0.13 | nucl |
| CaAHL31 | Caz04g08200.1 | Chr04:20375417-20376564 | 352 | 37.79 | 7.05 | 63.62 | -0.66 | nucl |
| CaAHL32 | Caz05g17580.1 | Chr05:235719249-235723745 | 294 | 31.56 | 5.35 | 52.68 | -0.61 | nucl |
| CaAHL33 | Caz06g17080.1 | Chr06:50064634-50076684 | 349 | 36.50 | 9.34 | 53.47 | -0.51 | nucl |
| CaAHL34 | Caz06g17990.1 | Chr06:57415662-57416120 | 152 | 15.59 | 4.44 | 51.44 | -0.03 | nucl |
| CaAHL35 | Caz06g24080.1 | Chr06:182665460-182672828 | 341 | 35.70 | 7.02 | 40.07 | -0.30 | plas |
| CaAHL36 | Caz07g19270.1 | Chr07:251735483-251736292 | 269 | 28.24 | 9.24 | 33.67 | -0.13 | cyto |
| CaAHL37 | Caz08g07460.1 | Chr08:137932240-137938844 | 341 | 34.99 | 10.26 | 58.12 | -0.22 | nucl |
| CaAHL38 | Caz09g21720.1 | Chr09:274809109-274813725 | 438 | 45.01 | 9.38 | 50.94 | -0.34 | nucl |
| CaAHL39 | Caz12g05920.1 | Chr12:13682382-13697371 | 578 | 60.51 | 7.72 | 52.36 | -0.30 | chlo |
| CaAHL40 | Caz12g05950.1 | Chr12:13811174-13813071 | 115 | 11.71 | 11.25 | 59.27 | -0.01 | chlo |
| CaAHL41 | Caz12g06070.1 | Chr12:14037114-14041618 | 177 | 18.10 | 4.89 | 59.53 | -0.20 | chlo |
| CaAHL42 | Caz12g06080.1 | Chr12:14056732-14062760 | 438 | 47.47 | 9.27 | 50.27 | -0.21 | chlo |
| CaAHL43 | Caz12g08880.1 | Chr12:32039622-32040395 | 257 | 28.05 | 7.83 | 49.44 | -0.42 | nucl |
| CaAHL44 | Caz12g18220.1 | Chr12:219722263-219725994 | 293 | 29.60 | 6.16 | 46.17 | -0.33 | nucl |
| CaAHL45 | Caz12g18510.1 | Chr12:221530027-221540499 | 358 | 37.13 | 9.57 | 50.03 | -0.37 | cyto |
1Note: nucl, nucleus; chlo, chloroplast; cyto, cytoplasm; plas, plasma membrane; vacu, vacuole membrane; E.R., endoplasmic reticulum.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



