Submitted:
16 January 2025
Posted:
16 January 2025
You are already at the latest version
Abstract
Palmer amaranth (Amaranthus palmeri S. Watson), native to North America, is one of the most prominent invasive weed species in agricultural land. Acetolactate synthase (ALS)-resistant A. palmeri is widespread, but its resistance pattern and molecular basis have not yet been studied in China. In 2019, an A. palmeri population that survived the recommended rate of imazethapyr was collected in Shandong Province, China. The objective of this research was to investigate the resistant mechanism and pattern of A. palmeri to imazethapyr. Dose-response assay showed that the resistant (R) population displayed a high resistance level (292.5-fold) to imazethapyr compared with the susceptible (S) population. Sequence analysis of the ALS gene revealed that nucleotide mutations resulted in three resistance-conferring amino acid substitutions, Pro-197-Ile, Trp-574-Leu, and Ser-653-Asp, in the individual plants of the R population. An in vitro enzyme assay indicated that the ALS was relatively unsusceptible to imazethapyr in the R population, showing a resistance index of 88.6-fold. ALS gene expression and copy number did not confer resistance to imazethapyr in the R population. Pro-197-Ile is the first reported amino acid substitution conferring ALS-resistant in A. palmeri. This is the first case of an herbicide-resistant A. palmeri biotype in China.
Keywords:
Amaranthus. palmeri
; Resistance
; Imazethapyr
; ALS gene mutation
; ALS gene copy number
; ALS gene expression
1. Introduction
Palmer amaranth (Amaranthus palmeri S. Wats), an annual broadleaf weed species native to the area encompassing the western United States to northern Mexico, started to spread beyond its original habitat in the early 20th century [1]. Nowadays, it has infested many crop fields and caused significant yield losses. Yield reduction was up to 68% in soybean [Glycine Max (L.) Merrill] fields due to A. palmeri interference at a density of 10 plants m⁻¹ of row [2]. Similarly, researchers reported a reduction of more than 22% in cotton (Gossypium herbaceum L.) yield and 11 to 91% in corn (Zea mays L.) yield from A. palmeri competition at the density of 0.9 plants m-1 of row [3, 4] and 0.5 to 8 plants m-1 of row [5, 6], respectively.
In China, Li and Che first observed A. palmeri in Changping District, Beijing [7, 8], in 1985. At present, ten provinces (e.g., Shandong, Tianjin, Henan) have reported the presence of A. palmeri [9, 10]. Due to the environmental adaption and strong competitive capacity, it can grow in cultivated fields, wasteland, ditches, garbage dumps, and feedlots [7]. Recently, an A. palmeri population has colonized crop fields and orchards in China [11].
A wide range of pre- and post-emergence herbicides are available for A. palmeri control. Among them, acetolactate synthase (ALS)-inhibiting herbicides are used worldwide due to their low cost and low mammalian toxicit. The ALS inhibitors are classified into five chemical families, including sulfonylureas (SUs) , imidazolinones (IMIs), pyrimidinylthiobenzoates (PTBs), triazolopyrimidines (TPs), and sulfonylaminocarbonyltriazolinones (SACTs) [12-16]. IMIs, such as imazethapyr, imazaquin, imazapic, imazameth, are traditionally used for weed control in legumes and noncultivated land. Long-term use of the herbicides has resulted in the evolution of ALS-resistant populations in A. palmeri. Since the first case of imazethapyr resistance in A. palmeri in the United States [17], 17 cases have been reported to be resistant to IMIs herbicides worldwide [18].
Currently, target-site and non-target-site resistance are the reported resistance mechanisms to ALS-inhibiting herbicides. However, target-site is the predominant mechanism conferring resistance to ALS-inhibiting herbicides [19]. Generally, target-site resistance is associated with one of several amino acid substitutions in ALS (Yu and Powles, 2014). To date, ten amino acid substitutions (Ala-122-Ser, Ala-122-Thr, Pro-197-ser, Pro-197-Ala, Pro-197-Thr, Pro-197-Asp, Ala-282-Asp, Asp-376-Glu, Trp-574-Leu, and Ser-653-Asn) at six positions in ALS confer resistance to ALS-inhibiting herbicides in A. palmeri globally [20-24], including the cases in Argentina, Brazil, Spain, and the United States [25-28]. In addition, target gene overexpression and gene copy number amplification have also caused the presence of the resistance of A. palmeri to the herbicides in other groups [29,30]. For example, the copy number of 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) genes and the expression level were correlated with the resistance of a population of A. palmeri to glyphosate in Georgia in the United States [31]. Nevertheless, it is still unknown whether copy number amplification and duplication of target genes contribute to some cases of ALS herbicide resistance in A. palmeri [20].
To our knowledge, there are no relevant reports on herbicide-resistant A. palmeri in China. In 2019, a population of A. palmeri in Shandong, China, survived following imazethapyr applications at the recommended rate in our yearly survey. The objective of this research was to characterize the level of imazethapyr resistance in this A. palmeri population and to identify the biochemical and molecular bases of the herbicide resistance.
2. Results
2.1. Response of A. palmeri to imazethapyr
Increasing imazethapyr doses caused shoot mass reduction compared to the nontreated control for both R and S populations. However, the S population of A. palmeri exhibited substantially greater susceptibility to imazethapyr than the R population, as shown in Figure 1. From the regression analysis, GR50 values measured 4124.6 g ai. ha-1 and 14.1 g ai. ha-1 for the R and S populations, respectively, indicating that the R population had a 292.5-fold higher resistance to imazethapyr than the S population. Therefore, these findings suggested that the R population of A. palmeri exhibited a high level of resistance to imazethapyr.
2.2. Sequencing of ALS gene
ALS gene sequencing from 100 individual plants of the R population in A. palmeri revealed that the population possessed three different known ALS resistance mutations, tryptophane (TGG) to leucine (TTG) at position 574 (Trp-574-Leu), serine (AGC) to aspartic acid (AAC) at position 653 (Ser-653-Asp), and proline (CCC) to isoleucine (ATC) at position 197 (Pro-197-lle) (Figure 2). No other known ALS resistance mutations were found. Nevertheless, it is worth mentioning that Pro-197-lle mutation had not been previously reported in A. palmeri.
Of the 100 analyzed individual plants from the R population, 19 plants had Pro-197-Ile mutation, and 20 individuals had Ser-653-Asp mutation, whereas only 13 plants carried the Trp-574-Leu mutation. Additionally, many individuals displayed more than one ALS gene mutation, with 45% of individuals having two mutations (197+574, 197+653, 574+653), 2 plants with all three resistance mutations (197+574+653), and 1 plant did not contain any of the known ALS gene mutations. Trp-574-Leu and Ser-653-Asp mutations were more common than Pro-197-lle mutations (Table 2).
2.3. In vitro ALS assay
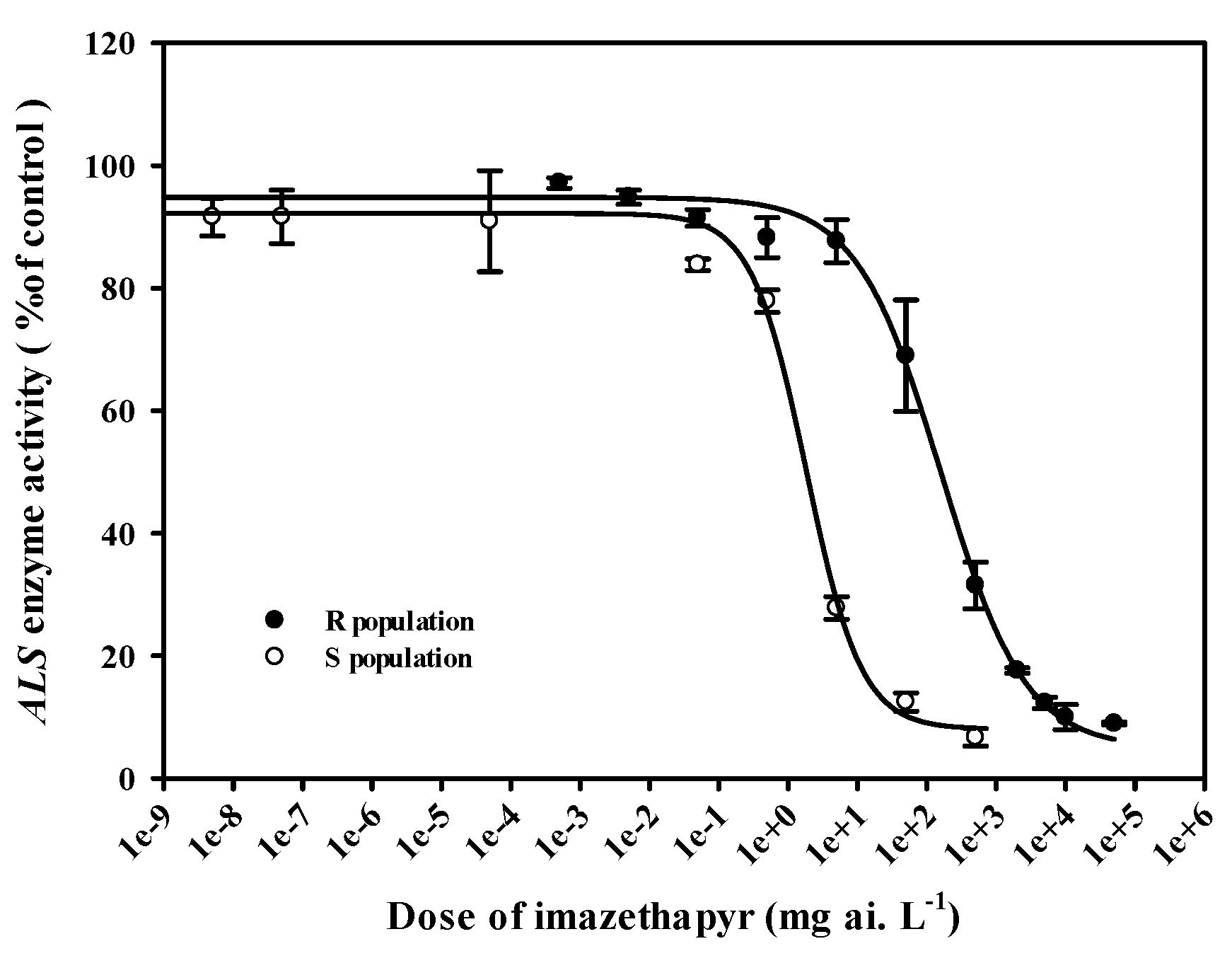
As the herbicide dose increased, the ALS enzyme activity relative to the nontreated control decreased for both S and R populations in A. palmeri (Figure 3). Nevertheless, the ALS enzyme activity of the S population sharply declined and showed substantially greater sensitivity to imazethapyr than the R population. The IC50 value of the S population was 1.8 mg ai. L-1, while the IC50 value of the R population was 159.5 mg ai. L-1 when calculated based on acetoin (acetolactate) produced per mg of fresh weight. The IC50 of the R population was 88.6-fold greater than that of the S population. Thus, the result suggested that the resistance to imazethapyr in the R population is associated with the target-site resistance mechanism.
Figure 3.
In vitro ALS activity assays in response to imazethapyr were performed using protein extracts of plants derived from the resistant A. palmeri population (●) and the susceptible population (○). Results were pooled over experimental runs. Vertical bars represent standard error of the mean (n=3).
Figure 3.
In vitro ALS activity assays in response to imazethapyr were performed using protein extracts of plants derived from the resistant A. palmeri population (●) and the susceptible population (○). Results were pooled over experimental runs. Vertical bars represent standard error of the mean (n=3).
2.4. ALS gene copy number and ALS gene expression
There were no significant differences in the ALS gene copy numbers between the S and R populations. The ALS gene copy number relative to internal reference gene A36 for both S and R populations ranged from 1.0 to 2.3. Similarly, the ALS gene expression levels in the R population did not significantly differ from the S population. Without imazethapyr treatment, the ALS gene expression level in the R population ranged from 1.6 to 2.3, while the level in the S population ranged from 0.9 to 1.8. These findings indicated that the ALS gene copy numbers and gene expression levels are unlikely associated with the resistance in the R population.
Figure 4.
ALS copy number relative to A36 for the S and R populations; ALS gene expression relative to A36 for the S and R populations not treated with imazethapyr. Vertical bars represent the mean ± standard deviation; ns represent non-significant.
Figure 4.
ALS copy number relative to A36 for the S and R populations; ALS gene expression relative to A36 for the S and R populations not treated with imazethapyr. Vertical bars represent the mean ± standard deviation; ns represent non-significant.
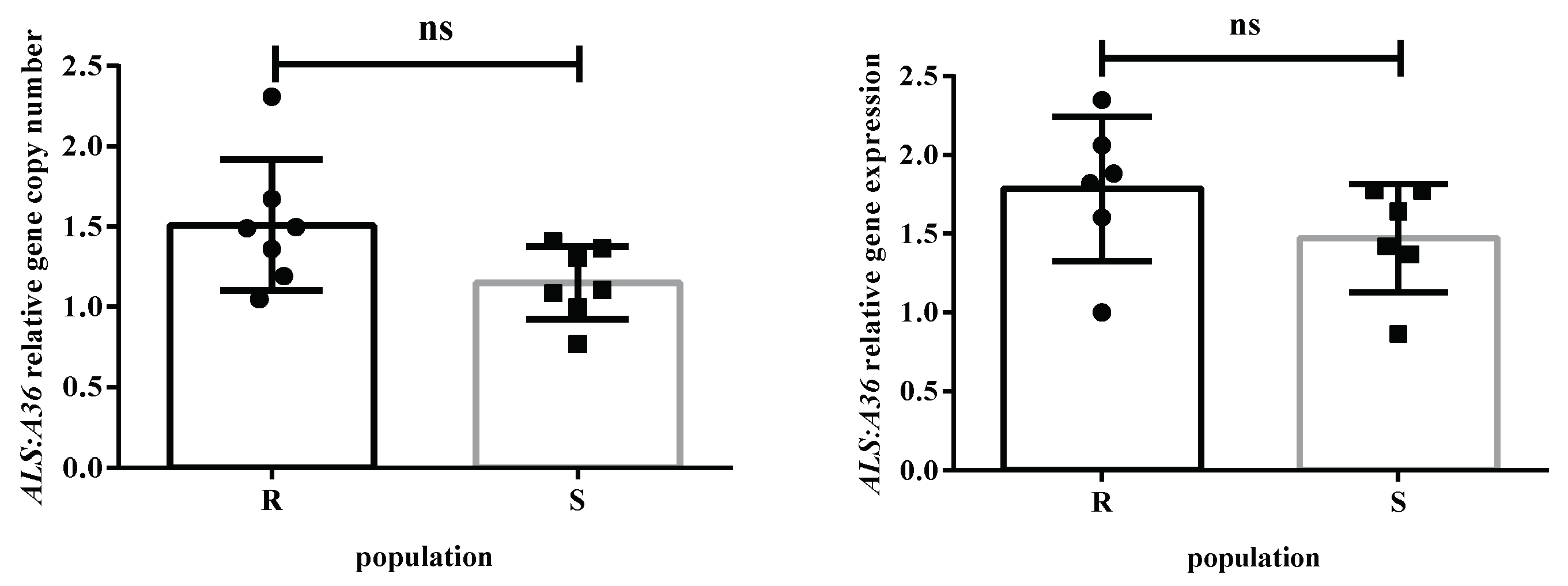
3. Discussion
Amaranthus species have the most cases of resistance to various herbicide modes of action because of their high genetic diversity, high seed production, and continuous emergence [32, 33]. For example, a prostrate pigweed (Amaranthus blitoides S. Watson) population demonstrated 790-fold resistance to ALS inhibitors [34], and a smooth pigweed (Amaranthus hybridus L.) population exhibited up to 537-fold resistance to imazethapyr [35]. An early investigation conducted in the United States showed that resistant A. palmeri biotypes had greater than 2800-fold resistance to imazethapyr than the susceptible biotypes [36]. In the present research, the R population displayed resistance to imazethapyr (292.5-fold) at the whole plant level, and the IC50 value at the enzyme level of the R population was 88.6-fold greater than that of the S population. These findings suggested the R population of A. palmeri investigated in the present study has evolved a high level of resistance to imazethapyr.
In this study, ALS gene sequencing identified three amino acid substitutions, including Pro197, Trp574, and Ser653, in the R population. These mutations agree with the previous reports in A. palmeri populations [18]. However, it should be noted that Pro-197-lle mutation has not been previously reported in A. palmeri in the literature. To date, the substitution (Pro-197-Ile) has been found only in mustard (Sisymbrium orientale L.) [18]. Importantly, many individual plants of the R population in the present study displayed more than one ALS gene mutation, with 45% of individuals possessing two resistance mutations (197+574,197+653 and 574+653) and 2% of the plants carrying all three mutations (197+574+653). Multiple gene mutations may be related to the fact that A. palmeri is an obligate outcrossing, dioecious species. A plant with no mutation detected in the ALS gene sequencing may be due to the accumulation of a pair of recessive genes during the reproduction of the population.
It is worth mentioning that a single ALS gene has two or more ALS mutations (double or multiple mutations) at the allele level may complicate the resistance situation to ALS-inhibiting herbicides [19]. However, the epistatic (multiplicative, additive, synergistic, and compensatory) effect of multiple resistance alleles on plant fitness cost is not yet clear when individual plants accumulate multiple ALS resistance alleles [19]. In the present study, the proportion of double or triple mutations in all plants tested was nearly 50%. Thus, an additional study should be conducted to confirm the contribution of multiple ALS resistance alleles in different mutated biotypes in the R population for the resistance to imazethapyr.
Target gene amplification and overexpression have contributed to the herbicide resistance in some weed species [30, 31, 37]. In glyphosate-resistant plants, including Amaranthus spp., EPSPS gene amplification is the primary resistance mechanism [31, 38]). In the R population of goosegrass (Elusine indica L.), the expression of EPSPS was higher (13.8-fold) than that of the S population after glyphosate treatment [30]. Amaranthus spp. is a diploid plant that contains one copy of the ALS gene [39]. However, the intense selection pressure of ALS inhibitors may promote the genotypes with increased ALS gene copies, resulting in herbicide resistance [20]. In the present study, the ALS gene copy numbers were not significantly different between the R and S populations. Without imazethapyr treatment, the ALS gene expression level in the R and S populations did not exhibit a significant difference. These findings suggested that the slight change (elevation or depression) in relative ALS gene copy number and gene expression in this ALS-resistant A. palmeri population did not contribute to the resistance.
As an invasive weed in China, the development of herbicide-resistant A. palmeri will undoubtedly affect the ecological environment and agricultural production. In the United States, crop losses from invasive weeds have been estimated at approximately US $ 27,000 million per year [40]. In dioecious species, resistance genes can be exchanged or transferred between the plants [41]. Therefore, cross- or multiple-resistance is common in A. palmeri [18, 38]. Recently, the case of an A. palmeri population with multiple-resistance to 2,4-D, atrazine, chlorsulfuron, glyphosate, and mesotrione was confirmed in Kansas in the United States [42].
4. Materials and Methods
4.1. Plant materials and growth conditions
Matured A. palmeri seeds of a suspected resistant (R) population were collected on a roadside near a ditch in Shandong, China (N37°07′04″,119°05′45″), while a susceptible (S) population were collected in a corn field in Beijing, China (N40°25′41″, E116°24′20″).
Seeds of the R and S populations were planted in plastic pots (10-cm diameter and 10 cm height) containing commercial potting soil (50% peat, 25% pine bark, and 25% sand) (Beijing Kawin Technology Share-Holding Co., Ltd., China) and placed in a greenhouse at 30/25 ℃ day/night with natural sunlight. The plant seedlings were watered as needed. The seedlings were thinned to 5 plants per pot when they reached a 2- to 3-leaf stage.
4.2. Dose-response experiments
To determine the resistance level of the R population to imazethapyr, experiments were conducted in August and December 2020. Plants at the 4- to 6-leaf stage were treated with imazethapyr (Doushile®, 5% AS, Shandong Cynda Chemical Co., Ltd, China) in a spray cabinet (3WPSH-500D, Beijing Research Center for Information Technology in Agriculture, Beijing, China) equipped with a single moving Teejet XR 8003 flat fan nozzle and calibrated to deliver 367.5 L ha-1. Imazethapyr rates applied to the S population were based on 0, 0.125×, 0.25×, 0.5×, 1×, 2×, and 4× the recommended rate, while imazethapyr rates applied to R populations were based on 1×, 2×, 4×, 8×, 16×, 32×, 64×, 128×, and 256× the recommended rate (recommended field rate =75 g ai. ha-1). Shoots were harvested at 21 days after treatment (DAT), oven-dried at 80 ℃ for 72 h, and then weighed.
Experiments were designed as a randomized complete block with three replications and were conducted twice over time. Dose-response curves were generated by regression analysis using SigmaPlot v. 13 (Systat Software, Inc., San Jose, CA, USA). Data were regressed with a three-parameter log-logistic equation (Eq. 1):
where Y represents the shoot dry weight (percentage of nontreated control) at herbicide rate X, C is the lower limit, and D is the upper limit. Imazethapyr rate that caused 50% shoot mass reduction (GR50) was determined from the regression equation.
4.3. ALS gene sequencing and resistance mutation genotyping
Genomic DNA was extracted from young leaf tissues of 100 nontreated plants in the R population, in which seeds came from the reproduction of plants that survived the imazethapyr treatment in the previous study. Genomic DNA extraction was conducted according to the kit instruction (Tiangen Biotech Beijing Co., Ltd., China). Two primer pairs were designed based on the plant ALS gene sequence (KY781923.1) of A. palmeri and amplified the region containing the eight mutation sites that have been reported in other resistant plant species (Table 1). Three mutation sites (574, 653, and 654) were amplified with the primers (ALS-1199f/ALS-1199r: TGCCTAAACCCACTTATTCTGC; ATCTCCAACCAACTAATAAGCC). The sequence was amplified by the other primers (ALS-921f/ALS-921r: TTTGTTTCCCGATTTAGTCCT; AACAAATCGGCCTTATCAACC) containing five mutation sites (122, 197, 205, 376, and 377).
The PCR reaction consisted of 12.5 μL 2× Pfu PCR Master Mix (Tiangen Biotech Beijing Co., Ltd., China), 10.5 μL ddH2O, 0.5 μL the forward and reverse primers (Tiangen Biotech Beijing Co., Ltd., China), and 1 μL gDNA to make a 25-μL total volume. PCR was performed with the following conditions: initial denaturation at 95 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 30 sec, annealing at 55/53 °C for 30 sec, and extension at 72 °C for 1 min, and a final extension at 72 °C for 10 min. The PCR tubes were held at 4 °C until processing. PCR products were visualized on a 1% agarose gel to confirm the target fragment size. The PCR products were sequenced commercially (Beijing Sunbiotech Co., Ltd., China), and the sequencing results were analyzed using DNAMAN version 5.2.2 software (Lynnon Biosoft, Quebec, Canada).
4.4. In vitro ALS assay
Leaf materials at the 4- to 6-leaf stage were harvested. All samples were stored at -80 ℃ until use. ALS enzyme extraction and herbicide inhibition assays were conducted according to Yu [43]. A series of imazethapyr concentrations (0, 5×10-4, 5×10-3, 5×10-2, 0.5, 5, 50, 500, 2000, 5000, 10000, 25000, and 50000 mg ai. L-1 for the R population; 0, 5×10-8, 5×10-7, 5×10-6, 5×10-5, 5×10-4, 5×10-3, 5×10-2, 0.5, 5, 50, and 500 mg ai. L-1 for the S population) was used. The ALS enzyme activity assay was conducted twice over time with three replications per treatment. Data were regressed with the equation previously described in section 2.2.
4.5. ALS gene copy numbers and ALS gene expression
Leaf tissues for the R and S populations were collected from the nontreated plants, frozen immediately with liquid nitrogen, and stored at -80 ℃ until processing. Genomic DNA was extracted using the methods previously described in section 2.3. The primers for the internal reference genes A36 (A36_F244 (5′-TTGGAACTGTCAGAGCAACC-3′) and A36_R363 (5′-GAACCCACTTCCACCAAAAC-3′) were designed by Singh [44]. For the ALS gene, the primers ALS-F2 (5’-GCAATTCCTCCGCAATACGCC-3’) and ALS-R2 (5’-CAAACCCCATAGCCCCCAAAC-3’) were designed with Oligo version 7 software and based on the GenBank entry KY781922.1 for use in quantitative PCR on genomic DNA and cDNA. ALS primers were designed based on conserved regions in published plant ALS gene sequences. Efficiency curves were made for each primer set using a 0.0625×, 0.125×, 0.25×, 0.5×, and 1× dilution series of genomic DNA from the R population.
The relative ALS gene copy numbers were investigated using qPCR. A 25 μl reaction solution was prepared using 10 μl of Bester SybrGreen Master Mix (DBI® Bioscience), 0.5 μl of forward and reverse primers (Beijing Sunbiotech Co., Ltd., China), 8 μl of RNase-free water, and 0.04 μl of 50× ROX Reference Dye (DBI® Bioscience). qPCR was performed with the following steps: initial denaturation at 95 °C for 4 min, followed by 40 cycles of denaturation at 95 °C for 20 sec, and annealing at 60 °C for 1 min. This program was followed by a melt-curve analysis of 81 cycles of 55 °C for 30 sec. The negative control contained no template in the reaction. No amplification products were detected in the negative control. Data were analyzed using a modification of the 2−ΔΔCt method to determine the genomic copy number of ALS relative to A36 as ΔCt = (Ct, A36 − Ct, ALS), and the relative increase in the genomic ALS copy number was expressed as 2ΔCt. Each reaction had three technical replicates.
The nontreated plant leaf tissues from the R and S populations were collected, immediately frozen with liquid nitrogen, and then stored at −80 ℃ until processing. Total RNA was extracted using an RNA-prep Pure Plant Kit (Tiangen Biotech Beijing CO., LTD). First-strand complementary DNA was synthesized using a FastQuant RT Kit (Tiangen Biotech Beijing CO., LTD). Quantitative real-time PCR was performed in a 25 µL reaction on a PCR machine (7500 Fast Real-Time, Thermo Fisher Scientific, Shanghai, China) under the following conditions: 10 min at 95 ℃, 40 cycles of 95 ℃ for 20 sec and 60 ℃ for 1 min, and then increasing the temperature by 0.5 ℃ every 5 sec to obtain the product melt curve. Relative quantification of ALS was calculated using the 2−ΔΔCt method and Ct=− [(Ct, ALS − Ct, A36) R − (Ct, ALS − Ct, A36) S]. The results are expressed as the fold increase in ALS expression level relative to A36.
5. Conclusions
In summary, this study reported the first case of ALS-resistant A. palmeri in China. Various mutations in ALS caused a high level of resistance to imazethapyr, and a new amino acid substitution (Pro-197-Ile) was detected in the R population. Additional studies need to be conducted to confirm the resistance levels of different mutation types and the cross- and multi-resistance patterns in the R population. Integrated weed management should be implemented to reduce the risk of further genetic evolution and the spread of resistant ALS gene in A. palmeri in China.
Author Contributions
Xiangju Li designed the research; Meijing Ji, performed most of the experiments; Haiyan Yu, Hailan Cui and Jingchao Chen helped the data analysis ; Jialin Yu revised the manuscript. All authors read and approved of this manuscript.
Funding
This work was financially supported by China Agriculture Research System (CARS25)、023DXZD0001 and STUAQ03.
Data Availability Statement
The sequence of A. palmeri ALS gene has been deposited in the NCBI with GenBank accession number of KY781923.1 and KY781922.1.
Acknowledgments
We thank the AJE Language Editing Services for the professional English language editing. And appreciate Prof. Xianquan Wang for helping to collect the seed.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ward, S.M.; Webster, T.M.; Steckel, L.E. Palmer amaranth (Amaranthus. palmeri): a review. Weed Technol. 2013, 27, 12–27. [Google Scholar] [CrossRef]
- Klingaman, T.E.; Oliver, L.R. Palmer amaranth (Amaranthus. palmeri) interference in soybeans (Glycine max). Weed Sci. 1994, 42, 523–527. [Google Scholar] [CrossRef]
- Rowland, M.W.; Murray, D.S.; Verhalen, L.M. Full-season Palmer amaranth (Amaranthus. palmeri) interference with cotton (Gossypium hirsutum). Weed Sci. 1999, 47, 305–309. [Google Scholar] [CrossRef]
- Smith, D.T.; Baker, R.V.; Steele, G.L. Palmer Amaranth (Amaranthus palmeri) Impacts on Yield, Harvesting, and Ginning in Dryland Cotton (Gossypium hirsutum). Weed Technol. 2000, 14, 122–126. [Google Scholar] [CrossRef]
- Massinga, R.A.; Currie, R.S.; Horak, M.J.; Boyer, J. Interference of Palmer amaranth in corn. Weed Sci. 2001, 49, 202–208. [Google Scholar] [CrossRef]
- Massinga, R.A.; Currie, R.S. Impact of Palmer amaranth (Amaranthus. palmeri) on corn (Zea mays) grain yield and yield and quality of forage. Weed Technol. 2002, 16, 532–536. [Google Scholar] [CrossRef]
- Li, Z.Y. Amaranthus. palmeri S Watson, A newly naturalized species in China. Botanical bulletin 2003, 20, 734–735. (In Chinese) [Google Scholar]
- Che, J.D. Invasive weed- Amaranthus. palmeri S Watson. Weed Science 2008, 26, 58–60. (In Chinese) [Google Scholar]
- Mo, X.Q.; Meng, W.Q.; Li, H.Y. New distribution records of three species of exotic plants in Tianjin: Amaranthus palmeri. Ipomoea lacunosa and Aster subulatu. Journal of Tianjin Normal University (Natural Science Edition) 2017, 37, 36–38. (In Chinese) [Google Scholar]
- Li, Y.G.; Cao, J.J.; Wang, R. Risk assessment based on analytic hierarchy process of an invasive alien weed,Amaranthus. palmeri,in northern Henan Province. Plant Quarantine 2021, 35, 60–65. (In Chinese) [Google Scholar]
- Zhang, M. Study on ecological effects of two amaranth invasive plants on the functional traits of native plants; Nankai university: Tianjin, 2020. [Google Scholar]
- Chaleff, R.S.; Mauvais, C.J. Acetolactate synthase is the site of action of two sulfonylurea herbicides in higher plants. Science. 1984, 224, 1443–1445. [Google Scholar] [CrossRef] [PubMed]
- Shaner, D.L.; Anderson, P.C.; Stidham, M.A. Imidazolinones: potential inhibitors of acetohydroxyacid synthase. Plant Physiol. 1984, 76, 545–546. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, B.C.; Subramanian, M.V.; Loney-Gallant, V.I.; Chandler, D.P. Mechanism of action of the 1, 2, 4-triazolo [1,5-a] pyrimidines. J. Pestic. Sci. 1990, 29, 357–364. [Google Scholar] [CrossRef]
- Stidham, M.A. Herbicides that inhibit acetohydroxyacid synthase. Weed Sci. 1991, 39, 428–434. [Google Scholar] [CrossRef]
- Santel, H.J.; Bowden, B.A.; Sorensen, V.M.; Mueller, K.H.; Reynolds, J. Flucarbazone-sodium: a new herbicide for grass control in wheat. Weed Science Society of America Abstract. 1999, 39, 7. [Google Scholar]
- Horak, M.; Peterson, D. Biotypes of Palmer Amaranth (Amaranthus palmeri) and common waterhemp (Amaranthus rudis) are resistant to imazethapyr and thifensulfuron. Weed Technol. 1995, 9, 192–195. [Google Scholar] [CrossRef]
- Heap, I. The international survey of herbicide resistant weeds. Available online: https://www.weedscience.org/ (accessed on 10 April 2024).
- Yu, Q.; Powles, S. Resistance to AHAS inhibitor herbicides: current understanding. Pest Manag Sci. 2014, 70, 1340–1350. [Google Scholar] [CrossRef]
- Singh, S.; Singh, V.; Salas-Perez, R.A.; Bagavathiannan, M.V.; Lawton-Rauh, A.; Roma-Burgos, N. Target-site mutation accumulation among ALS inhibitor-resistant Palmer amaranth. Pest Manag Sci. 2019, 75, 1131–1139. [Google Scholar] [CrossRef]
- Kohrt, J.R.; Sprague, C.L.; Swathi, N.; Douches, D. Confirmation of a three-way (glyphosate, als, and atrazine) herbicide-resistant population of Palmer amaranth (Amaranthus. palmeri) in Michigan. Weed Sci. 2017, 65, 327–338. [Google Scholar] [CrossRef]
- Chaudhari, S.; Varanasi, V.K.; Nakka, S.; Bhowmik, P.C.; Thompson, C.R.; Peterson, D.E.; Currie, R.S.; Jugulam, M. Evolution of target and non-target based multiple herbicide resistance in a single Palmer amaranth (Amaranthus palmeri) population from Kansas. Weed Technol. 2020, 34, 447–453. [Google Scholar] [CrossRef]
- Mahoney, D.J.; Jordan, D.L.; Roma-Burgos, N.; Jennings, K.M.; Leon, R.G.; Vann, M.C.; Everman, W.J.; Cahoon, C.W. Susceptibility of Palmer amaranth (Amaranthus palmeri) to herbicides in accessions collected from the North Carolina Coastal Plain. Weed Sci. 2020, 68, 582–593. [Google Scholar] [CrossRef]
- Manicardi, A.; Scarabel, L.; Llenes, J.M.; Montull, J.M.; Osuna, M.D.; Farré, J.T.; Milani, A. Genetic basis and origin of resistance to acetolactate synthase inhibitors in Amaranthus palmeri from Spain and Italy. Pest Manag Sci. 2023, 79, 4886–4896. [Google Scholar] [CrossRef] [PubMed]
- Foes, M.J.; Tranel, P.J.; Wax, L.M.; Stoller, E.W. A biotype of common waterhemp (Amaranthus. rudis) resistant to triazine and ALS herbicides. Weed Sci. 1998, 5, 514–520. [Google Scholar] [CrossRef]
- Larran, A.S.; Palmieri, V.E.; Perotti, V.E.; Lieber, L.; Tuesca, D.; Permingeat, H.R. Target-site resistance to acetolactate synthase (ALS)-inhibiting herbicides in from Argentina. Pest Manag Sci. 2017, 73, 2578–2584. [Google Scholar] [CrossRef] [PubMed]
- Küpper, A.; Borgato, E.A.; Patterson, E.L.; Gonçalves Netto, A.; Nicolai, M.; Carvalho, S.J.P.d.; Nissen, S.J.; Gaines, T.A.; Christoffoleti, P.J. Multiple Resistance to Glyphosate and Acetolactate Synthase Inhibitors in Palmer Amaranth (Amaranthus palmeri) Identified in Brazil. Weed Sci. 2017, 65, 317–326. [Google Scholar] [CrossRef]
- Torra, J.; Royo-Esnal, A.; Romano, Y.; Osuna, M.D.; León, R.G.; Recasens, J. Amaranthus palmeri a New Invasive Weed in Spain with Herbicide Resistant Biotypes. Agronomy. 2020, 10, 993. [Google Scholar] [CrossRef]
- Nakka, S.; Godar, A.S.; Wanti, P.S.; Thompson, C.R.; Peterson, D.E.; Roelofs, J.; Jugulam, M. Physiological and molecular characterization of hydroxyphenylpyruvate dioxygenase (HPPD)-inhibitor resistance in Palmer Amaranth (Amaranthus. palmeri S. Wats) . Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Chen, J.C.; Huang, H.J.; Zhang, C.X.; Wei, S.H.; Huang, Z. F.; Chen, J. Y.; Wang, X. Mutations and amplification of EPSPS gene confer resistance to glyphosate in goosegrass (Eleusine indica). Planta. 2015, 242, 859–868. [Google Scholar] [CrossRef]
- Gaines, T.A.; Zhang, W.L.; Wang, D.F.; Bukun, B.; Chisholm, S.T.; Shaner, D.L.; Westra, P. Gene amplification confers glyphosate resistance in Amaranthus palmeri. Proc. Natl. Acad. Sci. USA. 2010, 107, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, S.; Jordan, D.; York, A.; Jennings, K.M.; Cahoon, C.W.; Inman, A.; Chandi, M.D. Biology and management of glyphosate-resistant and glyphosate-susceptible Palmer amaranth (Amaranthus. palmeri) phenotypes from a segregating population. Weed Sci. 2007, 65, 755–768. [Google Scholar] [CrossRef]
- Molin, W.T.; Nandula, V.K.; Wright, A.A.; Bond, J.A. Transfer and expression of ALS inhibitor resistance from Palmer amaranth (Amaranthus. palmeri) to an A. spinosus × A. palmeri hybrid. Weed Sci. 2016, 64, 240–247. [Google Scholar] [CrossRef]
- Sibony, M.; Rubin, B. Molecular basis for multiple resistance to acetolactate synthase-inhibiting herbicides and atrazine in Amaranthus blitoides (prostrate pigweed). Planta. 2003, 216, 1022–1027. [Google Scholar] [CrossRef]
- Whaley, C.A.; Wilson, H.P.; Westwood, J.H. ALS resistance in several smooth pigweed (Amaranthus hybridus) biotypes. Weed Sci. 2006, 54, 828–832. [Google Scholar] [CrossRef]
- Sprague, C.L.; Stoller, E.W.; Wax, L.M.; Horak, M.J. Palmer amaranth (Amaranthus. palmeri) and common water hemp (Amaranthus rudis) resistance to selected ALS-inhibiting herbicides. Weed Sci. 1997, 45, 192–197. [Google Scholar] [CrossRef]
- Yu, J.; Mccullough, P.E.; Mcelroy, J.S.; Jespersen, D.; Shilling, D.G. Gene expression and target-site mutations are associated with resistance to ALS inhibitors in annual sedge (Cyperus compressus) biotypes from Georgia. Weed Sci. 2020, 68, 460–466. [Google Scholar] [CrossRef]
- Singh, S.; Burgos, N.R.; Singh, V.; Alcober, E.A.L.; Salaa-Perez, R.; Shivrain, V. Differential response of Arkansas Palmer amaranth (Amaranthus. palmeri) to glyphosate and mesotrione. Weed Technol 2018, 32, 579–585. [Google Scholar] [CrossRef]
- Ferguson, G.M.; Hamill, A.S.; Tardif, F.J. ALS inhibitor resistance in populations of Powell amaranth and redroot pigweed. Weed Sci. 2001, 49, 448–453. [Google Scholar] [CrossRef]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Steckel, L.E. The dioecious Amaranthus spp.: here to stay. Weed Technol. 2007, 21, 567–570. [Google Scholar] [CrossRef]
- Kumar, V.; Liu, R.; Boyer, G.; Stahlman, P.W. Confirmation of 2,4-D resistance and identification of multiple resistance in a Kansas Palmer amaranth (Amaranthus. palmeri) population. Pest Manag Sci. 2019, 75, 2925–2933. [Google Scholar] [CrossRef]
- Yu, Q.; Shane Friesen, L.J.; Zhang, X.Q.; Powles, S.B. Tolerance to acetolactate synthase and acetyl-coenzyme A carboxylase inhibiting herbicides in Vulpia bromoides is conferred by two co-existing resistance mechanisms. Pestic. Biochem. Physiol. 2004, 78, 21–30. [Google Scholar] [CrossRef]
- Singh, S.; Singh, V.; Lawton-Rauh, A.; Bagavathiannan, M.V.; Roma-Burgos, N. EPSPS gene amplification primarily confers glyphosate resistance among Arkansas Palmer amaranth (Amaranthus. palmeri) populations. Weed Sci. 2018, 66, 293–300. [Google Scholar] [CrossRef]
Figure 1.
Dry shoot mass reduction of the susceptible (S) and the resistant A.palmeri population (R) at 21 d after treatment with imazethapyr in two growth chamber experiments. Results were pooled over experimental runs. Vertical bars represent the standard error of the mean (n = 6).
Figure 1.
Dry shoot mass reduction of the susceptible (S) and the resistant A.palmeri population (R) at 21 d after treatment with imazethapyr in two growth chamber experiments. Results were pooled over experimental runs. Vertical bars represent the standard error of the mean (n = 6).
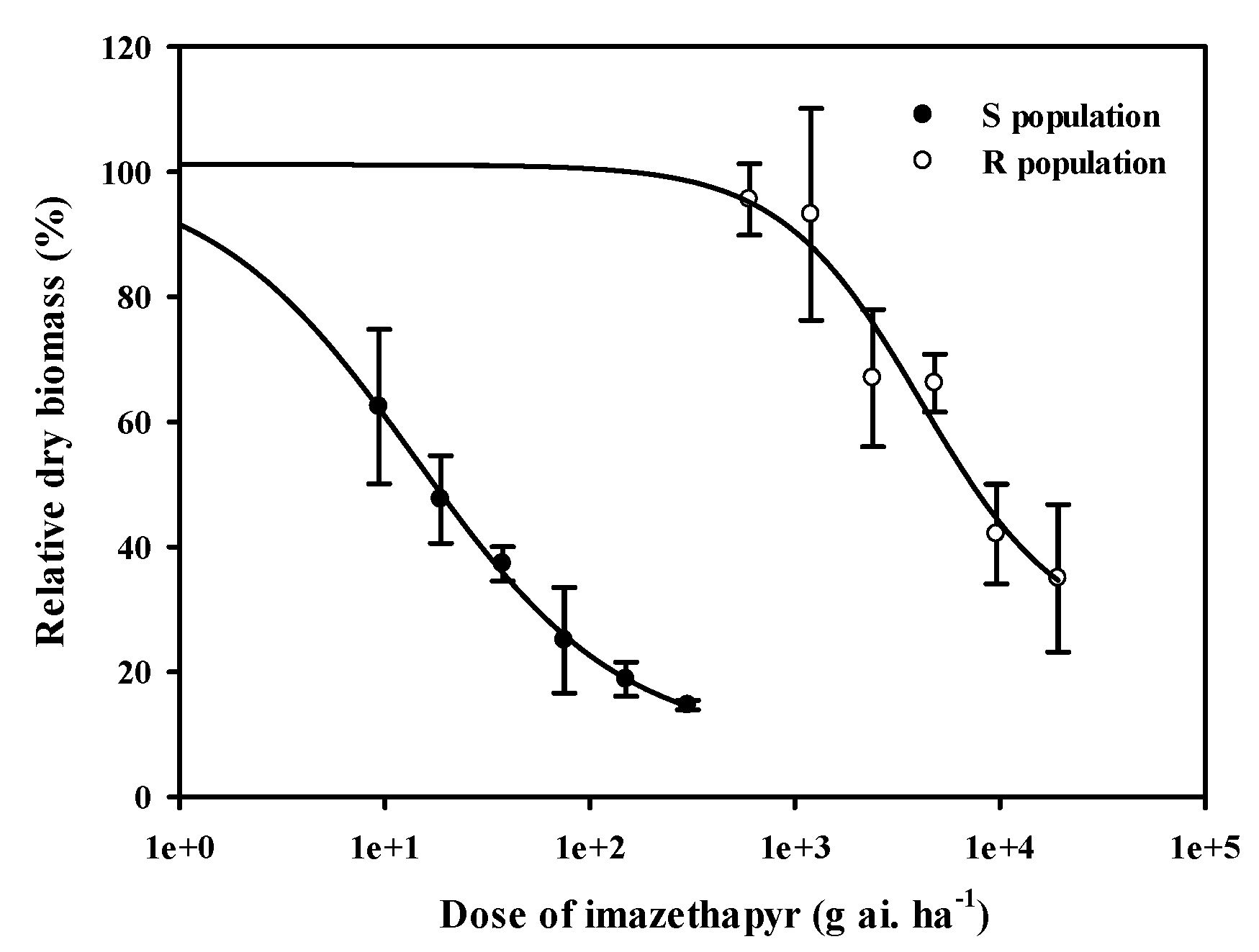
Figure 2.
Target-site mutations in the ALS gene conferring herbicide resistance in the R population: (a) Pro-197-Ile (CCC mutated ATC), (b) Trp-574-Leu (TGG mutated TTG), and (c) Ser-653-Asp (AGC mutated AAC) in the ALS gene sequence.
Figure 2.
Target-site mutations in the ALS gene conferring herbicide resistance in the R population: (a) Pro-197-Ile (CCC mutated ATC), (b) Trp-574-Leu (TGG mutated TTG), and (c) Ser-653-Asp (AGC mutated AAC) in the ALS gene sequence.
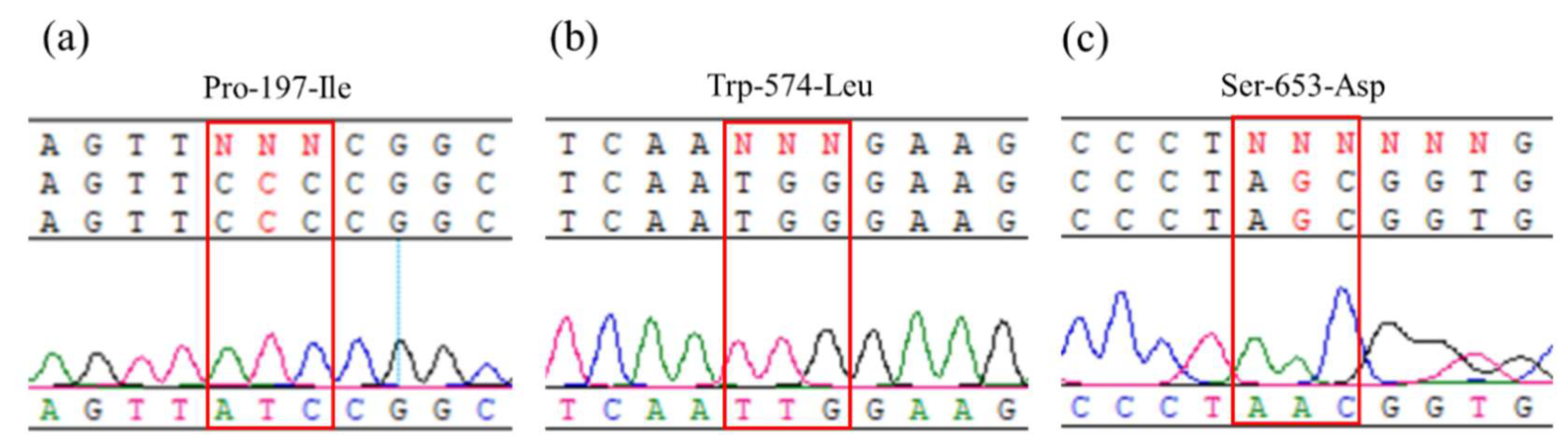

Table 2.
ALS domain sequencing results show one, two, or three different mutations in 100 individual plants of the R population.
Table 2.
ALS domain sequencing results show one, two, or three different mutations in 100 individual plants of the R population.
| Gene mutation types | Number of individuALS |
| Pro-197-lle | 19 |
| Trp-574-Leu | 13 |
| Ser-653-Asp | 20 |
| Pro-197-lle+ Trp-574-Leu | 11 |
| Pro-197-lle+ Ser-653-Asp | 9 |
| Trp-574-Leu+ Ser-653-Asp | 25 |
| Pro-197-lle+ Trp-574-Leu+ Ser-653-Asp | 2 |
| Mutation not detected | 1 |
Abbreviation: ALS, acetolactate synthase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



