Submitted:
06 January 2025
Posted:
07 January 2025
You are already at the latest version
Abstract
Staphylococcus aureus is a versatile zoonotic pathogen capable of causing a wide range of infections. Due to the organism’s ability to persist, recalcitrant and recurring infections are a major concern for public and animal health. This study investigated the establishment of persistence using two S. aureus strains—ATCC 29740, a bovine mastitis isolate, and USA300, a human clinical isolate—under substrate depletion. This stress established a persistence phenotype where the strains remained persistent for >120 days at notable concentrations and developed persister-like growth, including small colony variant formations. With qRT-PCR we found the cell density was higher than represented by the plate count while the intracellular ATP remained constant during the persistence phase. These findings indicate that S. aureus has complex survival strategies to support its persistent state, providing a host-specific perspective when addressing recurrent infections in human and animal infectious disease.
Keywords:
persistence
; persisters
; small colony variants
; nutritional stress
; bovine mastitis
; MRSA
; non-culturable
1. Introduction
Staphylococcus aureus, a ubiquitous gram-positive bacterium, is a highly adaptable, versatile pathogen [1]. While S. aureus is a common commensal of the human and animal microbiome, it can cause a wide spectrum of infections in a diverse range of hosts, including livestock, wildlife, and humans [2,3]. In humans, S. aureus can range from mild skin and soft tissue infections to severe, life-threatening diseases such as pneumonia, endocarditis, sepsis, and toxic shock [4]. It is one of the leading causes of nosocomial infections and a major contributor to mortality in hospitals [4] due to the wave of methicillin-resistant S. aureus (MRSA) [5,6], which in part is due to its capacity to form biofilms [7]. In veterinary medicine, S. aureus causes great economic losses in livestock production, most prominently in dairy production, being the leading cause of bovine mastitis worldwide [2,8]. Infections are also widespread in other animals including swine, poultry, equine, small ruminants, and companion animals [2,9]. Due to the plethora of animal hosts this zoonotic pathogen infects, host-switching events are a common and significant issue at the human/animal interface [9]. For example, more recent zoonotic concerns are being raised as cases of MRSA strains, such as the MRSA ST398 type, can be transmit between humans and animals [2,10,11,12]. Zoonotic events entail the acquisition and shedding of mobile genetic elements and the emergence of host-specific mutations to adapt to the new host, further emphasizing S. aureus' remarkable capacity to evolve and adapt to new host-specific environmental challenges [9]. Such adaptation can be driven by overuse of antibiotics in agricultural practices, providing a selective environment for endemic strains that may be associated with risk of human-adapted antibiotic-resistant transmission [11].
This opportunistic pathogen is difficult to prevent and treat due to its arsenal of virulence factors that may allow it to survive for prolonged periods of time in stressful conditions. S. aureus is well recognized for its robust adhesion and biofilm development. When incorporated into a biofilm, the extracellular matrix acts as a protector from environmental stresses, including antibiotics and the host immune system; thereby, allowing strains that are antimicrobial sensitive to survive and persist within a host [13]. These traits highlight the ability of S. aureus to be a highly adaptive pathogen that quickly acquires antibiotic resistance mechanisms through mobile genetic elements or chromosomal mutations, as witnessed with the development and spread of MRSA strains into communities worldwide and leading to the serious threat of vancomycin-resistant S. aureus (VRSA) [14,15,16]. Studies exploring S. aureus-host interactions found that development and employment of host-specific immune evasion molecules permit this organism to survive phagocytosis [17]. Even if treatment is successful against this adaptive pathogen, S. aureus produces heat- and proteolytic-resistant exotoxins that enable further disease to spread after cell death. A noteworthy example being the pyrogenic toxin superantigens that cause toxic shock syndrome and Staphylococcus-associated food-borne illness [18]. While these virulence factors are familiar in the field, recently recognized and understudied persistence mechanisms are found in S. aureus, including the phenotypic subpopulations of persisters and small colony variants (SCVs).
The persister phenotype is comprised of cellular sub-populations that reduce growth and exhibit antibiotic tolerance beyond that found in biofilms. Most antibiotics target active growth mechanisms of microbes, so the inhibition of growth in persisters bypasses the functional mechanisms of the antibiotic. Growth resumes once this stress is removed, and these persisters return to active growth and are indistinguishable from wild-type, suggesting that alternative approaches are needed to understand antimicrobial resistance in concert with persistence. While it is understood that persisters have an altered metabolism and altered ATP production, the mechanisms to enter this antibiotic-tolerant persister state are not well understood [19,20,21]. Within the broad umbrella of persistence are SCVs, being a specific type of persister with an altered colony morphology. SCVs are phenotypically distinct varients that become slow growing and demonstrate increased tolerance to antibiotics and multiple environmental stresses. The colonies appear 1/10 the size of the parent colony with a rougher texture, where development is associated with an intracellular persistence strategy to reduce expression of virulence factors and invade host cells, enabling protection from antibiotic therapy and become hiden to the host innate defense system. SCVs of S. aureus can persist within host cells for years, acting as reservoirs for chronic infections and being difficult to detect. SCVs are considered part of the normal life cycle of bacteria but often revert rapidly to a wild-type phenotype to cause infection [22,23,24,25,26]. Mounting clinical evidence is increasingly correlating chronic infections to persisters and SCVs sub-populations, which has led to a surge of research focused on understanding the underlying mechanisms behind their induction and survival strategies and development of anti-persister drugs [19,20,23,27]. The importance of SCV infections is also mounting for linkage to chronic conditions that include cystic fibrosis, osteomyelitis, and device-related infections in humans, having an incidence in clinical specimens between <1% to >30% [22]. Bovine mastitis suffers from S. aureus persistent infections that are characterized by biofilms and antibiotic tolerance [8], and there are cases of SCVs and persisters being linked with chronic bovine mastitis [28,29].
As S. aureus continues to be a significant concern to public and animal health, understanding the growth patterns and persistence of S. aureus strains from different hosts under stress conditions is important for the development of targeted strategies to prevent, manage, and treat infections in a zoonotic pathogen. The aim of this work was to examine the growth patterns and persistence of two strains under nutritional duress: one isolated from bovine mastitis and another from a human clinical infection. We hypothesized that the two isolates would have different characteristics leading to and during persistence.
2. Materials and Methods
2.1. Strains and Culture Conditions
S. aureus ATCC 29740 and USA300 are a bovine mastitis and human clinical isolate, respectively. Both strains were cultured from stock onto BHI plates and incubated at 37°C for 24 hours. One colony of the overnight stock was inoculated into 10 mL of BHI broth (Difco) and incubated overnight at 37°C afterwhich the strains were normalized to OD595=0.1-0.3 with sterile saline before inoculation.
2.2. Growth Curves in Different Media
Overnight cultures were inoculated at 1% v/v into BHI broth (Difco, Sparks, MD), CDM [30] with 0.5% lactose, CDM with 0.5% glucose, and DMEM (Cytiva, Logan, UT). With three technical replicates, 200 μL aliquoted into a 96 well plate in three biological replicates. Growth was monitored for 72 hours in a Molecular Devices DTX 880 Multimode Detector (Beckman Coulter, Brea, CA) with readings taken every 30 minutes in three biological replicates.
2.3. Nutritional Deprivation and Cell Enumeration
Samples (10 mL) were grown were incubacted were centrifuged (5,000xg 5 minutes) and supernatant was discarded and replaced with an equal volume of PBS followed by inoculation of 2 mL into 200 mL of CDM with 0.5% lactose in three biological replicates and incubation at 37°C. At each time point for sample collection, 100 μL was taken from each bottle for analysis. Collections occurred at the same time each day and serial dilutions were done with sterile PBS. BHI plates were divided in half, plating 10 μL of a diluted sample per side of the plate. Growth plating assays were incubated at 37°C and colonies were counted at 24 and 48 hours of incubation. This was done at multiple times (T) 0, 0.5, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 14, 16, 21, 23, 50, 70, 87, and 120 days.
2.4. Quantitative PCR and ATP Assays
Cell pellets at T87 and T120 were collected (1 mL) for qRT-PCR to measure cell density and ATP assays followed by centrifugation (5,000xg 5 minutes). The cell pellet was resuspended in an equal volume of PBS for ATP measurement or 750 μL of Trizol LS for qRT-PCR analysis. ATP was measured using a BacTiter-Glo Microbial Cell Viability Assay kit (Promega, Madison, WI) according to manufacturer’s instructions.
RNA was extracted as described previously using TRIzol LS (Invitrogen, Carlsbad, CA) [31] and checked for quality and concentration using a NanoDrop (Thermo Scientific, Waltham, MA). From the RNA sample, cDNA was produced using the First Strand cDNA Synthesis (Quick Protocol) (New England Biolabs, Ipswich, MA).
The cDNA was used for RT-qPCR quantification of viable cells using a CFX 96 Real Time System (BioRad, 311 Hercules, CA) according to previously established methods [32]. iQ Sybr Green Mastermix (Biorad, Hercules, CA) was used along with 100 nM of forward (F) and reverse (R) PCR primers for the EF-TU gene (F: 5’- ACG CGG 313 TAT CAT CAA AGT GG - 3’; R: 5’ - ATC GGG TGG ATC AGG GTA AC - 3’) to quanitfy the CFU/mL of cells at T87 and T120. The parameters for both primers was done using a denaturation step at 95°C for 5 min, 317 followed by 40 cycles of denaturation, annealing, and extension at 95˚C for 15 s, 56˚C for 30 s, 72˚C for 318 30 s, respectively, and a final extension at 72°C for 1 min. The amplified product was verified using 319 melt curve analysis from 50˚C to 95˚C with a transition rate of 0.2˚C/s.
2.5. Statistical Analysis
Data are represented as the mean ± SEM of three biological replicates. A Mann-Whitney U Test was used to identify statistically significant differences between the strain or treatment, using R (version 4.0.5) for computations. Two tailed t-tests with unequal variances were done using GraphPad Prism Version 10 (Graphpad, Menlo Park, CA). Growth rate determination was done with GraphPad Prism Version 10 (Graphpad, Menlo Park, CA) while lag phase calculations were made with the Microbial Lag Phase Duration Calculator [33]. All figures were visualized with the programs GraphPad Prism Version 10 (Graphpad) and Biorender (Biorender, Toronto, Canada).
3. Results
3.1. Persistence Establishes within 72 Hours in Nutrient Deficient Conditions
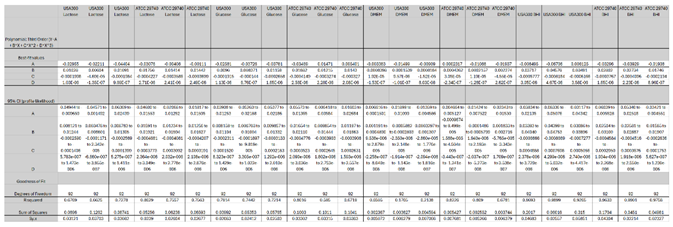
To differentiate between the transition to stationary phase or persistence, S. aureus ATCC 29740 and USA300 initial growth curves were determined in varying nutrient-availability media (Figure 1). BHI was used as the positive control for growth analysis, noting that each strain had significantly different growth characteristics that were used to determine individual growth differences within each strain. Stationary phase was reached at similar densities within CDM but began to decline within hours, suggesting the induction of persistence very quickly in the growth period. Grouping differences were seen between strains rather than between sugar composition. S. aureus ATCC 29740 had a shorter lag phases compared to S. aureus USA300, initiating growth a few hours sooner. While growth rates were similar (Table A1 and Table A2) across strains and media, lag phase was significant between strains in the same media with lactose sugar (p=0.0033) while glucose sugar was bordering significance (p=0.0871). Insignificant growth was seen when cultured in DMEM.
3.2. Persistence Continues for Over 120 Days When Challenged with Nutritional Stress and Induces Persister-Like Growth Patterns
Active replication was determined using plate counts on BHI agar, where persistence was observed over a 120-day incubation (Figure 2). The growth rate of the two strains was not significantly different (p>0.05) within the initial 5 hours. Both strains maintained a similar replicating population up to ~T7. However, a significant difference (p<0.05) of the replicating population was observed differentiate between the strains during T7 to T10. With continued incubation, a differential decline in the replicating population was observed.
The replicating population continued to differentiate after T15 that included initiation of three growth characteristics and multiple colony types. The first and most common colony type was ‘wild-type’ growth, with colonies appearing within 24 hours with the expected colony morphology of S. aureus. The second colony type had a smaller colony morphology and appeared within 24 hours, exhibiting SCV characteristics. The third observed growth variation was marked by delayed visibility, requiring 48 hours of incubation to produce visible colonies, indicating persister-like growth. These slow-growing colonies would appear dispersed among wild-type colonies with continued incubation on the plates. Only a few instances were observed where the entire plated population had persister-like growth. Despite the initial time-consuming appearance, these colonies would resume growth and become indistinguishable from the wild-type morphology after 48 hours (Figure 3). Although there were multiple biological replicates per strain, there were divergences within replicates regarding the occurrence of these three growth patterns.
3.3. Culture-Based Methods of Detection Do Not Accurately Reflect Viability
To verify the cell viability of the population, qRT-PCR and ATP assays were done at various incubation times during persistence (Figure 4). Results from the qPCR revealed an increase in cell density (Log10 CFU/mL) between the two timepoints with both strains. This result is divergent from the estimate determined using plate count, suggesting there are two populations in the broth: 1) one that is persistent and 2) one that is non-culturable on plates but intact and active. There was a statistical difference seen within USA300 between days T87 and T120 but no difference seen within ATCC 29740 or between strains. To determine if the amount of energy is different between the populations, we used intracellular ATP assays, which remained consistent for both timepoints. There were no statistical differences between strains or between days.
4. Discussion
In this study, we aimed to characterize the persistence and adaptability of Staphylococcus aureus strains under conditions of nutritional stress, with significant implications for understanding the mechanisms behind chronic and recalcitrant infections. Furthermore, adopting a multi-host perspective captures the diversity of a pathogen that displays host specificity and specialization, an important factor when developing targeted strategies to prevent and manage persistent infections in both human and veterinary contexts.
When first determining initial growth patterns and the divergence toward persistence, persistence induction remained universal in both strains, becoming steady in the stationary phase within 72 hours. This highlights the swift response and adaption to environmental stresses this pathogen is capable of. Considering the short time-period between inoculation to induction of persistence, persistence is not an acquired genetic resistance but an innate phenotypic change conserved across S. aureus strains in response to stressors. Additionally, persistence within 72 hours implies a limited window for effective antimicrobial intervention. Treatments may need to be aggressive and rapid to prevent the formation of persistence phenotypes.
The difference in lag phase between the strains could elude to accustomed metabolism plasticity and switching based on the host environment. S. aureus derived from bovine mastitis has undergone gene acquisition, loss, and diversification in order to distinctively thrive in the mammary gland [8], expanding its nutrient utilization to incorporate lactose uptake, a common sugar found in dairy milk [34]. The human host environment may encourage catabolite repression, where resource optimization would prioritize glucose over lactose [35], accounting for slight differences of growth between the sugars in S. aureus USA300. Lack of growth in DMEM implies that this media lacks key nutrients required for both S. aureus growth and persistence, highlighting the auxotrophic nature of the bacteria and its need for specific amino acids, vitamins, and other nutrients found abundantly when housed in its host. Overall, despite known host-switching and host specializations, persistence is a maintained virulence factor, indicating its necessity for infection. Based on this information, we decided to use lactose as our sugar for our extended persistence experiment due to it being the largest difference in persistence induction between the two strains.
When investigating the persistence of this pathogen beyond 72 hours, S. aureus remained actively replicating for over 120 days. While differences were noticed in initial growth patterns, both human clinical and bovine mastitis S. aureus isolates shared similar persistence activities. Our findings align with previous studies describing the starvation response of S. aureus, who similarly found persistence lasting for months and the development of SCVs when grown in glucose- and multi-nutrient-limited media [36,37]. The increases in variance over time between biological replicated suggests persistence is influenced by microenvironmental factors, causing stochastic fluctuations in population. These variations may additionally be heightened by the small sample size.
Staphylococcus commonly experience nutrient-deficient conditions in terrestrial, clinical, and host environments [37]. In addition to the immune system and other harsh host environments, infecting and persisting in a host specifically requires overcoming challenges like nutritional deprivation, such as nutritional immunity [38]. Frequent exposures to stresses like this can motivate the development of persistence factors that address nutrient availability or the lack thereof, such as persisters and SCVs.
Despite antibiotic treatment until the infection is seemingly cleared and symptoms are eliminated, once the antibiotic stressor is removed, persisters will resume growth and cause a relapse in infection [19]. Persister resuscitation is an underdeveloped but advancing issue being addressed in the field. Revival of persisters is triggered through chemotaxis systems, sensing nutrient availability within the environment, such as essential amino acids. Secondary messenger proteins like cAMP that are in high concentrations in persisters are reduced, leading to ribosomal resuscitation and following protein synthesis and growth [39]. Furthermore, the slow growth and altered characteristics of SCVs, such as reduced metabolism and increased antibiotic tolerance [40], allows them to evade the immune system and antibiotic treatment and persist longer in nutrient depleted environments, contributing to chronic and recurrent infections [22]. The growth patterns observed throughout this experiment resemble that of persisters and SCVs. However, further testing would be required to confirm these phenotypes [26]. The simultaneous occurrence of persister-like colonies among wild-type and the resumed growth after 48 hours of incubation could indicate a reversion back to the wild-type phenotype when introduced back to a nutrient-rich environment. The appearance of persister-like colonies at different timepoints across biological replicates and their subsequent reversion is expected, given the unstable and often transient nature of SCVs [24]. Furthermore, SCVs are difficult to culture, contributing to the challenge of researching this persister, but as this study presents and others have suspected, the strategy of continuous culturing is proving to be an effective strategy for selection of persisters like SCVs [41].
Confirmation of viability using multiple assays revealed inconsistencies within culture-based detection methods. Culture-based methods of detection are considered the “gold standard” when testing for food contamination and for food analysis [42]. However, while the results from the plating and qPCR were similar on T=87, divergences occurred on T=120, where the qPCR determined a larger viable population than the culture-based method. With the development of persisters, culture-based methods for detection become unreliable due to the slow growth of these persisters. For example, SCVS may take 2-3 days to become visible on a plate [43], not appearing within the incubation period used within the experiment and therefore seemingly having a lower viable population. Additionally, the ATP assay revealed a steady level of intracellular ATP between both strains on both days, confirming viability and active metabolism. When the integrity of the membrane is compromised, ATP production is halted and ATPases will deplete any remaining intracellular ATP [44]. Based on the maintained ATP presence within the cells on both timepoints, the population may have reached a favored ATP concentration to sustain a persister state, enough to support this chemotaxis sensing necessary for resuscitation. The 48-hour incubations in combination with the small sample size and limited sample collections provide a restricted perspective of the true viability of the population, but it is clear that local conditions are causing stochastic fluctuations in plate counts, persister-like growth, and viability within these assays, further complicating the phenomenon of persistence.
Our objective was to annotate the persistence mechanisms involved during nutritional stress in a subset of different host-derived isolates, with a specific interest in the induction of persisters. These findings highlight the broader implications of S. aureus persistence mechanisms, especially when considering health risks, food safety, and zoonotic potential. By revealing how S. aureus adapts and survives under prolonged nutritional stress, this research underscores the capacity of this pathogen to persist in livestock and human settings alike, contributing to recalcitrant and recurring infections. Such adaptability not only complicates treatment but also poses a risk for cross-species transmission, which can amplify the spread of resilient strains and potentially contribute to outbreaks. Understanding these persistence strategies is essential for informing effective therapeutic and preventative measures aimed at minimizing the impact of S. aureus on both human and veterinary health. Further investigation into the molecular mechanisms driving SCV formation, persistence, and resuscitation when considering environmental stressors and host-specific factors is essential for improving therapeutic approaches to prevent and manage these challenging infections in human and veterinary medicine.
Author Contributions
Conceptualization, KR, CS, and BCW; methodology, KR, CS, and BCW formal analysis, KR, and CS; resources, BCW; data curation, KR, and CS; writing—original draft preparation, KR; writing—review and editing, KR, CS, and BCW; visualization, KR and CS; project administration, BCW; funding acquisition, BCW. All authors have read and greed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| SCV | Small Colony Variant |
Appendix A

Table A1.
Two tailed t-tests with unequal variances of growth rates in different medias.
| Between Strain | Lactose | 0.14370278 | |||
| Glucose | 0.684679703 | ||||
| DMEM | 0.902200382 | ||||
| BHI | 0.396304143 | ||||
| USA300 | Lactose | Glucose | DMEM | BHI | |
| Lactose | NA | ||||
| Glucose | 0.474397606 | NA | |||
| DMEM | 0.459986984 | 0.404311317 | NA | ||
| BHI | 0.275856074 | 0.171969557 | 0.648222506 | NA | |
| ATCC 29740 | Lactose | Glucose | DMEM | BHI | |
| Lactose | NA | ||||
| Glucose | 0.686956269 | NA | |||
| DMEM | 0.284479808 | 0.07761123 | NA | ||
| BHI | 0.07761123 | 0.158564251 | 0.372151583 | NA |
Table A2.
Polynomials for growth rate determination in different medias.
References
- Lowy, F.D. Staphylococcus aureus Infections. New England Journal of Medicine 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Peton, V.; le Loir, Y. Staphylococcus aureus in Veterinary Medicine. Infection, Genetics and Evolution 2014, 21, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Peres, A.G.; Madrenas, J. The broad landscape of immune interactions with Staphylococcus aureus: From commensalism to lethal infections. Burns 2013, 39, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Gnanamani, A.; Hariharan, P.; Paul- Satyaseela, M.; Gnanamani, A.; Hariharan, P.; Paul- Satyaseela, M. Staphylococcus aureus: Overview of Bacteriology, Clinical Diseases, Epidemiology, Antibiotic Resistance and Therapeutic Approach. Frontiers in Staphylococcus aureus 2017. [Google Scholar] [CrossRef]
- Thompson, R.L.; Cabezudo, I.; Wenzel, R.P. Epidemiology of Nosocomial Infections Caused by Methicillin-Resistant Staphylococcus aureus. Ann Intern Med 1982, 97, 309–317. [Google Scholar] [CrossRef]
- Sharma, N.K.; Garg, R.; Baliga, S.; Bhat, K.G. Nosocomial Infections and Drug Susceptibility Patterns in Methicillin Sensitive and Methicillin Resistant Staphylococcus aureus. J Clin Diagn Res 2013, 7, 2178–80. [Google Scholar] [CrossRef]
- Bhattacharya, M.; Wozniak, D.J.; Stoodley, P.; Hall-Stoodley, L. Prevention and treatment of Staphylococcus aureus biofilms. Expert Review of Anti-Infective Therapy 2015, 13, 1499–1516. [Google Scholar] [CrossRef]
- Campos, B.; Pickering, A.C.; Rocha, L.L.; Aguilar, A.P.; Fabres-Klein, M.H.; Mendes, T.A.D.O.; Fitzgerald, J.R.; Ribon, A.D.O.B. Diversity and pathogenesis of Staphylococcus aureus from bovine mastitis: Current understanding and future perspectives. BioMed Central 2022, 18. [Google Scholar] [CrossRef] [PubMed]
- Haag, A.F.; Fitzgerald, J.R.; Penadés, J.R. Staphylococcus aureus in Animals. Microbiol Spectr 2019, 7. [Google Scholar] [CrossRef]
- Jangra, S.; Khunger, S.; Chattopadhya, D. Antimicrobial Resistance in Staphylococci Special Emphasis on Methicillin Resistance among Companion Livestock and Its Impact on Human Health in Rural India. IntechOpen 2022. [Google Scholar] [CrossRef]
- Park, S.; Ronholm, J. Staphylococcus aureus in Agriculture: Lessons in Evolution from a Multispecies Pathogen. Clin Microbiol Rev 2021, 34. [Google Scholar] [CrossRef] [PubMed]
- Leonard, F.C.; Markey, B.K. Methicillin-resistant Staphylococcus aureus in animals: A Review. The Veterinary Journal 2008, 175, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Parastan, R.; Kargar, M.; Solhjoo, K.; Kafilzadeh, F. Staphylococcus aureus biofilms: Structures, antibiotic resistance, inhibition, and vaccines. Gene Reports 2020, 20, 100739. [Google Scholar] [CrossRef]
- Pantosti, A.; Sanchini, A.; Monaco, M. Mechanisms of Antibiotic Resistance in Staphylococcus Aureus. Future Microbiology 2007, 2, 323–334. [Google Scholar] [CrossRef]
- Appelbaum, P.C. Microbiology of Antibiotic Resistance in Staphylococcus aureus. Clinical Infectious Diseases 2007, 45, S165–S170. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; Deleo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef]
- Howden, B.P.; Giulieri, S.G.; Wong Fok Lung, T.; Baines, S.L.; Sharkey, L.K.; Lee, J.Y.H.; Hachani, A.; Monk, I.R.; Stinear, T.P. Staphylococcus aureus Host Interactions and Adaptation. Nat Rev Microbiol 2023, 21, 380–395. [Google Scholar] [CrossRef]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin Microbiol Rev 2000, 13. [Google Scholar] [CrossRef] [PubMed]
- Conlon, B.P. Staphylococcus aureus chronic and relapsing infections: Evidence of a role for Persister Cells. BioEssays 2014, 36, 991–996. [Google Scholar] [CrossRef]
- Chang, J.; Lee, R.E.; Lee, W. A pursuit of Staphylococcus aureus continues: A role of persister cells. Arch. Pharm. Res 2020, 43, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister Cells: Molecular Mechanisms Related to Antibiotic Tolerance. Handbook of Experimental Pharmacology 2012, 211, 121–133. [Google Scholar] [CrossRef]
- Proctor, R.; von Eiff, C.; Kahl, B.; Becker, K.; McNamara, P.; Merrmann, M.; Peters, G. Small colony variants: A pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat Rev Microbiol 2006, 4, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.A.; Kriegeskorte, A.; Kahl, B.C.; Becker, K.; Löffler, B.; Peters, G. Staphylococcus aureus Small Colony Variants (SCVs): A road map for the metabolic pathways involved in persistent infections. Front Cell Infect Microbiol 2014, 4. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Heitmann, V.; Hussain, M.; Viemann, D.; Roth, J.; von Eiff, C.; Peters, G.; Becker, K.; Löffler, B. Staphylococcus aureus Small-Colony Variants Are Adapted Phenotypes for Intracellular Persistence. The Journal of Infectious Diseases 2010, 202, 1031–1040. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Medina, E.; Hussain, M.; Völker, W.; Heitmann, V.; Niemann, S.; Holzinger, D.; Roth, J.; Proctor, R.A.; Becker, K.; Peters, G.; Löffler, B. Staphylococcus aureus phenotype switching: An effective bacterial strategy to escape host immune response and establish a chronic infection. EMBO Mol Med 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Becker, K. Detection, Identification and Diagnostic Characterization of the Staphylococcal Small Colony-Variant (SCV) Phenotype. Antibiotics (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Rowe, S.E.; Beam, J.E.; Conlon, B.P. Recalcitrant Staphylococcus aureus Infections: Obstacles and Solutions. Infect Immun 2021, 8. [Google Scholar] [CrossRef]
- Zaatout, N.; Ayachi, A.; Kecha, M. Staphylococcus aureus persistence properties associated with bovine mastitis and alternative therapeutic modalities. Journal of Applied Microbiology 2020, 129, 1102–1119. [Google Scholar] [CrossRef]
- Lin, X.-Q.; Liu, Z.-Z.; Zhou, C.-K.; Zhang, L.; Gao, Y.; Luo, X.-Y.; Zhang, J.-G.; Chen, W.; Yang, Y.-J. Trained immunity in recurrent Staphylococcus aureus infection promotes bacterial persistence. PLoS Pathog 2024, 20. [Google Scholar] [CrossRef]
- Gao, S.; Oh, D.H.; Broadbent, J.R.; Johnson, M.E.; Weimer, B.C.; Steele, J.L. Aromatic amino acid catabolism by lactococci. Lait 1997, 77, 371–381. [Google Scholar] [CrossRef]
- Shaw, C.; Weimer, B.C.; Gann, R.; Desai, P.T.; Shah, J.D. The Yin and Yang of pathogens and probiotics: Interplay between Salmonella enterica sv. Typhimurium and Bifidobacterium infantis during co-infection. Front. Microbiol. 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Arabyan, N.; Park, D.; Foutouhi, S.; et al. Salmonella Degrades the Host Glycocalyx Leading to Altered Infection and Glycan Remodeling. Sci Rep 2016, 6, 29525. [Google Scholar] [CrossRef] [PubMed]
- Smug, B. J.; Opalek, M.; Necki, M.; Wloch-Salamon, D. Microbial lag calculator: A shiny-based application and an R package for calculating the duration of microbial lag phase. Methods in Ecology and Evolution 2023, 15, 301–307. [Google Scholar] [CrossRef]
- Richardson, E.J.; Bacigalupe, R.; Harrison, E.M.; et al. Gene exchange drives the ecological success of a multi-host bacterial pathogen. Nat Ecol Evol 2018, 2, 1468–1478. [Google Scholar] [CrossRef] [PubMed]
- Stülke, J.; Hillen, W. Carbon catabolite repression in bacteria. Current Opinion in Microbiology 1999, 2(2), 195–201. [Google Scholar] [CrossRef]
- Watson, S.P.; Clements, M.O.; Foster, S.J. Characterization of the Starvation-Survival Response of Staphylococcus aureus. J Bacteriol 1998, 180. [Google Scholar] [CrossRef]
- Onyango, L.A.; Alreshidi, M.M. Adaptive metabolism in staphylococci: Survival and persistence in environmental and clinical settings. J Patho. 2018, 2018, 1092632. [Google Scholar] [CrossRef]
- Cassat, J.E.; Skaar, E.P. Metal ion acquisition in Staphylococcus aureus: Overcoming nutritional immunity. Semin Immunopathol 2012, 34, 215–235. [Google Scholar] [CrossRef]
- Wainwright, J.; Hobbs, G.; Nakouti, I. Persister cells: Formation, resuscitation and combative therapies. Arch Microbiol 2021, 203, 5899–5906. [Google Scholar] [CrossRef]
- Eiff, C.V. Staphylococcus aureus small colony variants: A challenge to microbiologists and clinicians. Elsevier BV 2008, 31, 507–510. [Google Scholar] [CrossRef]
- Lee, J.; Zilm, P.S.; Kidd, S.P. Novel Research Models for Staphylococcus aureus Small Colony Variants (SCV) Development: Co-pathogenesis and Growth Rate. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Foddai, A.C.G.; Grant, I.R. Methods for detection of viable foodborne pathogens: Current state-of-art and future prospects. Appl Microbiol Biotechnol 2020, 104, 4281–4288. [Google Scholar] [CrossRef] [PubMed]
- Becker, K. Detection, Identification and Diagnostic Characterization of the Staphylococcal Small Colony-Variant (SCV) Phenotype. Antibiotics (Basel) 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Riss, T. L.; Moravec, R. A.; Niles, A. L.; Duellman, S.; Benink, H. A.; Worzella, T. J.; Minor, L. Cell viability assays. In Assay guidance manual, 2016.
Figure 1.
S. aureus growth curves in different media. Similar cell densities were reached for both strains when not grown in BHI. The growth curve in DMEM was maintained density of about 0..
Figure 1.
S. aureus growth curves in different media. Similar cell densities were reached for both strains when not grown in BHI. The growth curve in DMEM was maintained density of about 0..
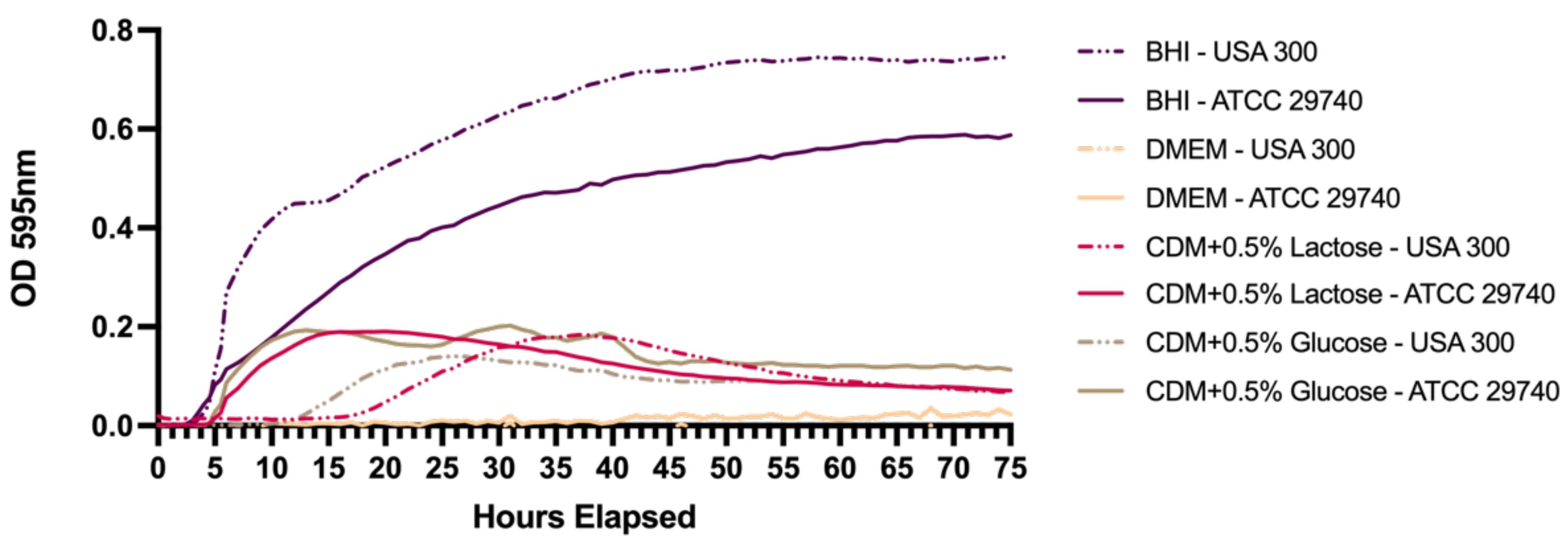
Figure 2.
Plate count enumeration during extended incubation in CDM containing 0.5% lactose. Plate counts were averaged over three biological replicates and three technical replicates. Error bars represent ± SEM.
Figure 2.
Plate count enumeration during extended incubation in CDM containing 0.5% lactose. Plate counts were averaged over three biological replicates and three technical replicates. Error bars represent ± SEM.
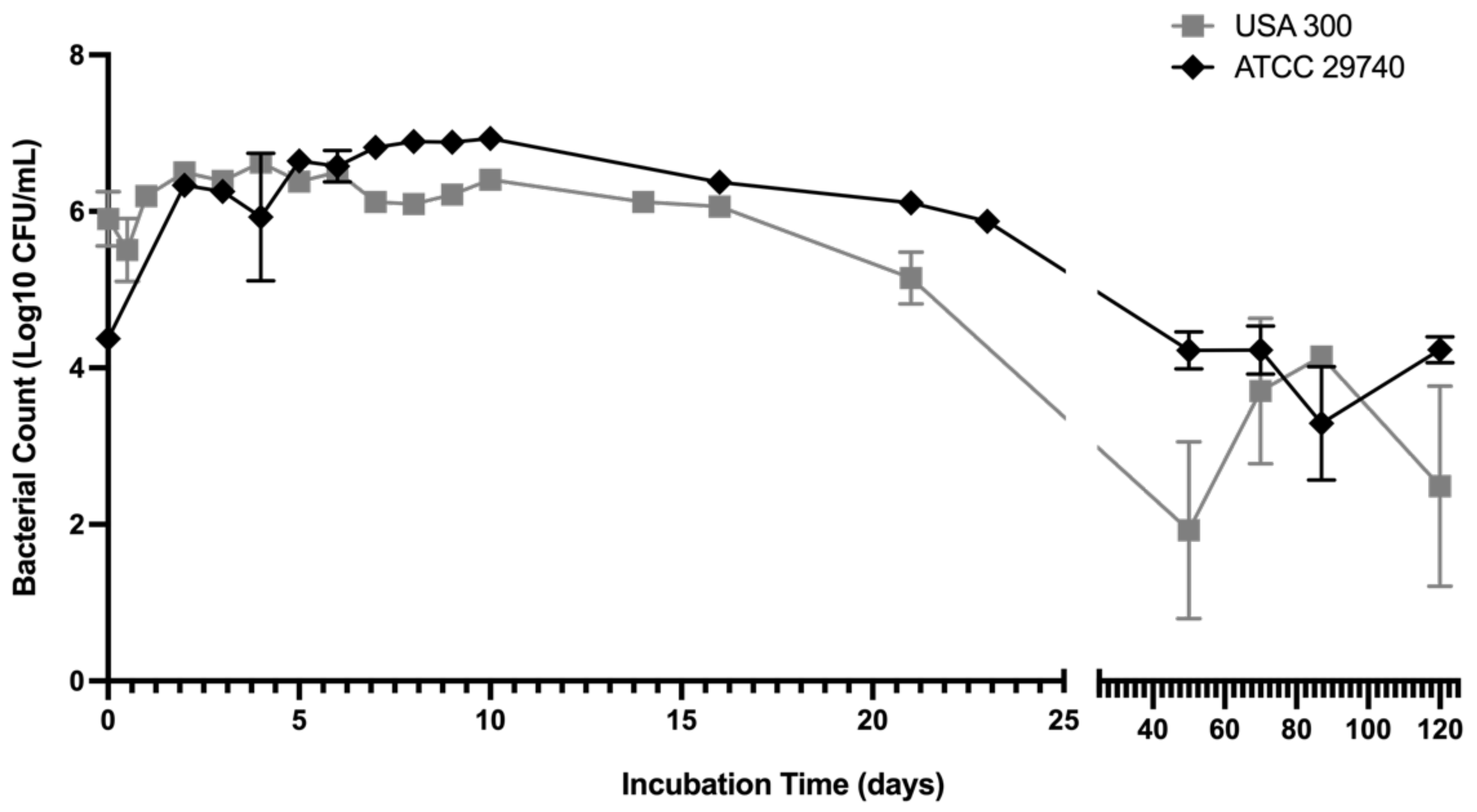
Figure 3.
Colony morphology within the culturable population during growth on plating. (a) S. aureus ATCC 297490 A at T120 after 24 h of incubation on the plate. There are miniscule colonies appearring but are too small to retrieve an accurate count. This characterizes the entire plated population. (b) The plate from Figure 3A after 48 hours of incubation on the plate. Colonies now have reverted and display wild-type features. (c) S. aureus USA300 B at T87 after 24 hours of incubation on the plate that contained two phenotypic colonies: wild-type and small morphology. The dashed arrows indicate SCV-like colonies while the solid arrow indicate wild-type colonies. .
Figure 3.
Colony morphology within the culturable population during growth on plating. (a) S. aureus ATCC 297490 A at T120 after 24 h of incubation on the plate. There are miniscule colonies appearring but are too small to retrieve an accurate count. This characterizes the entire plated population. (b) The plate from Figure 3A after 48 hours of incubation on the plate. Colonies now have reverted and display wild-type features. (c) S. aureus USA300 B at T87 after 24 hours of incubation on the plate that contained two phenotypic colonies: wild-type and small morphology. The dashed arrows indicate SCV-like colonies while the solid arrow indicate wild-type colonies. .

Figure 4.
Cell density and intracellular energy during the persistence phase. (a) qRT-PCR at T87 and T120, illustrating an increase in population over these two day. Signicant difference was seen between USA300 between T87 and T120 at p=0.0011. (b) ATP assay on T87 and T120, indicating a maintained composition level of intracellular ATP.
Figure 4.
Cell density and intracellular energy during the persistence phase. (a) qRT-PCR at T87 and T120, illustrating an increase in population over these two day. Signicant difference was seen between USA300 between T87 and T120 at p=0.0011. (b) ATP assay on T87 and T120, indicating a maintained composition level of intracellular ATP.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



