
Submitted:
26 December 2024
Posted:
27 December 2024
You are already at the latest version
Abstract
Rabies is a zoonotic viral disease preventable through vaccination. Control strategies should be based on the “One Health” concept, as addressing zoonotic pathogens at their animal source is the most effective and cost-efficient approach to protect human health. The aim of this study was to develop two third-generation anti-rabies vaccines based on non-replicative viral vectors, MVA and Ad5, expressing rabies virus (RABV) glycoprotein (MVA-RG and Ad-RG), and to evaluate their efficacy in an intracerebral RABV challenge model. MVA-RG was produced using a platform developed in our laboratory, while a commercial kit was used to produce Ad-RG. Protection against rabies was assessed in a mouse intracerebral challenge model. Our results demonstrated that both non-replicative viral vectors provided protection against RABV. The MVA-RG and Ad-RG vectors, administered in two homologous doses, conferred 60% and 60-100% protection against RABV challenge, respectively. The survival rate was influenced by the type of viral vector, the administered dose, and the immunization scheme applied. Remarkably, to our knowledge, our study is the first to report 100% protection against RABV IC challenge using non-replicative Ad5 in a homologous immunization scheme. These results encourage us to proceed with the evaluation of this vaccine candidate in target animals.
Keywords:
rabies
; MVA
; adenovirus
; Ad5
; vector-based vaccine
; viral vector
; glycoprotein
1. Introduction
Rabies virus (RABV) is the causative agent of an important viral zoonosis resulting in acute encephalitis and death in mammals. The RABV belongs to the genus Lyssavirus of the family Rhabdoviridae, they are enveloped viruses with a single-stranded negative-sense RNA genome [1]. The viral genome is 12,000 bp long and encodes the transmembrane glycoprotein (G), the primary antigen of the virus, the matrix protein (M) located on the inner side of virion, always is associated with the nucleoprotein (N, associated with genomic RNA), the phosphoprotein (P or NS), and the polymerase (L) [1]. The transmembrane glycoprotein is the only protein present on viral surface, contains epitopes recognized by CD4+ and CD8+ T cells and it is the major target of neutralizing antibodies (NA) [2,3].
Rabies is preventable through vaccination, and control strategies should be approached from the “One Health” concept. Indeed, the control of zoonotic pathogens at their animal source is the most effective and cost-efficient solution for protecting humans. Inactivated-RABV vaccines are widely used in humans, companion animals and livestock. They are safe and effective but require multiple doses to generate immunity. On the other hand, in a few countries, baits containing attenuated strains of RABV are used for the vaccination of wild animals (reviewed in [4]). However, it is crucial to demonstrate that they do not produce any adverse effects in the target and non-target species. To overcome these inconveniences, several recombinant viral vectored vaccines have been developed and assessed for companion animals, wildlife and livestock, with varying degrees of efficacy. However, only a few of them are licensed (reviewed in [4,5]).
Two replicative viral vectored rabies vaccines are commercially available. The first one was obtained over 30 years ago and is based on the Vaccinia virus (VACV) Copenhagen strain, which expresses the glycoprotein of the ERA strain of RABV [6]. The current commercial name of this vaccine is Raboral V-RG® (Boehringer Ingelheim Animal Health), and it is used for the prevention of wildlife rabies in North America and Europe (http://www.raboral.com/). Furthermore, there is a licensed adenovirus-vectored vaccine for use in wildlife. This vaccine is based on the replicative viral vector human adenovirus type 5 and carries the gene for the RABV G protein from the ERA strain (AdRG1.3, [7]). The commercial name of the vaccine is ONRAB® (Ontario Rabies Vaccine Bait, Artemis Technologies Inc., Canada), and it is currently used in Canada for the control of rabies in foxes, raccoons, and skunks (https://www.ontario.ca/page/wildlife-rabies-outbreaks-and-control-operations). Also, there is an anti-rabies vaccine for cats based on the non-replicative canary poxvirus (PUREVAX®, Boehringer Ingelheim).
Non-replicative viral vectored vaccines are chosen to develop biotechnological vaccines for humans, companion animals and livestock to ensure safety for the people. Modified vaccinia Ankara virus (MVA) constitutes an excellent platform to develop rationally designed viral vectored vaccines due to its safety profile, its capability to induce humoral and cellular immune responses and its intrinsic adjuvant capacities [8,9]. It has been extensively used as a viral vector for human and veterinary vaccines [10]. Furthermore, non-replicative adenoviruses (Ad) have been evaluated as viral vectors for the prevention of infectious diseases because they induce potent innate and adaptive immune responses (both antibodies and CD8+ T cell responses), there are well-established and simple techniques for constructing recombinant adenoviruses, and high viral titers can be achieved in vitro [11,12]. Even if MVA and Ad vectors have been widely assessed as vaccines in preclinical and clinical trials, the first viral vectored vaccines approved for humans was during Covid-19 pandemic and they were based on non-replicative Adenovirus 26 (Ad26), replication-defective human adenovirus type 5 (Ad5) or the modified chimpanzee adenovirus (ChAdOx1) [13].
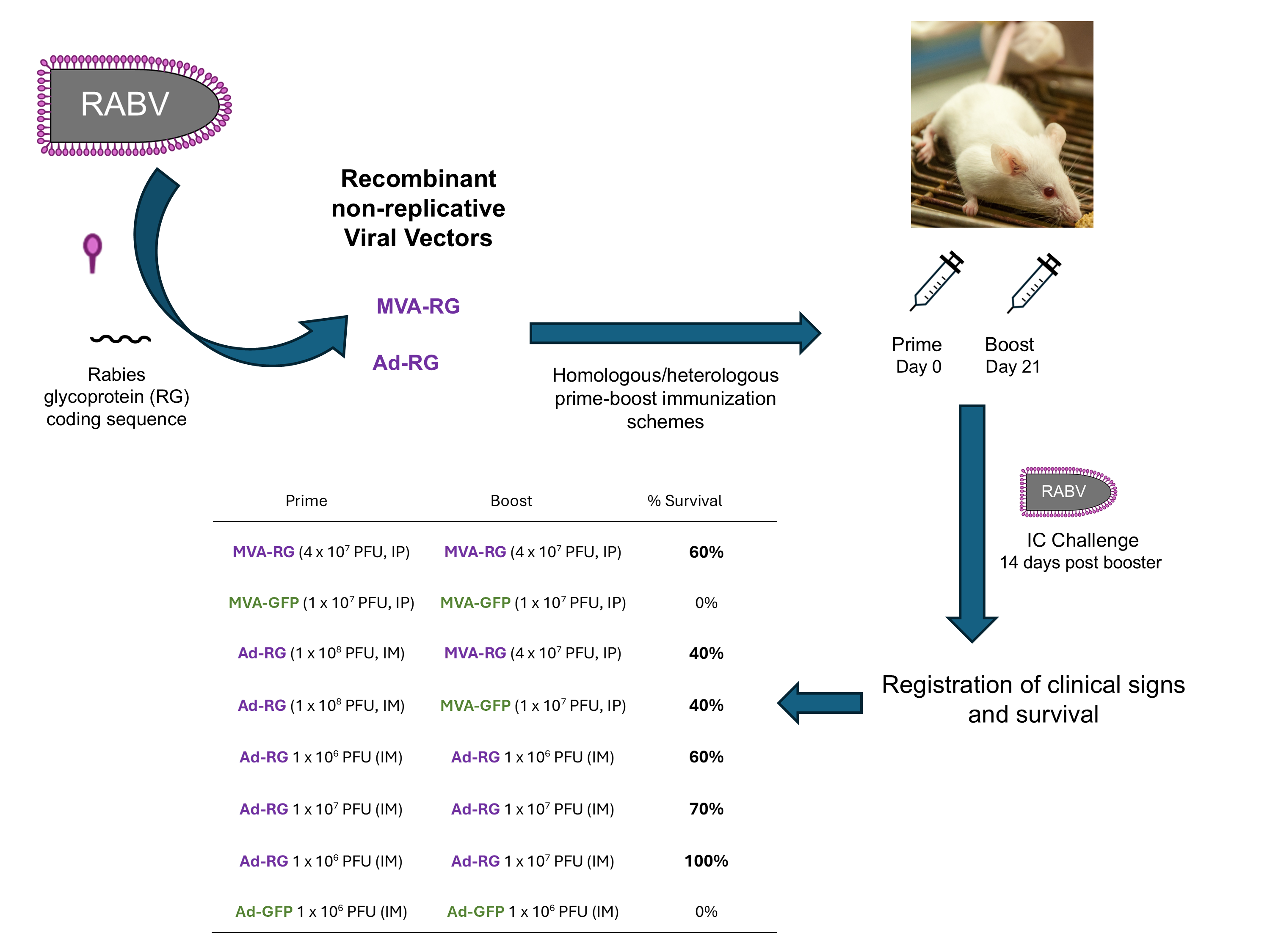
To obtain safe and effective third-generation rabies vaccines, in this study we generated two non-replicative viral vectors based on MVA and Ad5 expressing the RABV glycoprotein (RG). The ability of the recombinant viruses MVA-RG and Ad-RG to protect against RABV was tested in an intracerebral challenge model in mice.
2. Materials and Methods
2.1. Cell Lines and Viral Stocks
Modified vaccinia Ankara virus (MVA) and human adenovirus type 5 ΔE (Ad5) viruses were propagated in primary cultures of chicken embryo fibroblasts (CEFs) and human embryonic kidney cells transformed with the E1 region of human adenovirus type 5 (HEK293A), respectively. Viral immunization stocks were obtained by ultracentrifugation through 25% v/v sucrose cushion and titrated by plaque assay as described elsewhere [14]. Recombinant MVA carrying the gene encoding the green fluorescent protein (GFP, MVA-GFP), was obtained previously in our laboratory (unpublished results) and Ad expressing GFP (Ad-GFP) was provided by Dr. M. Pérez Filgueira (Instituto de Virología, IVIT, UEDD INTA-CONICET).
Rabies virus strain Challenge Virus Standard (RABV CVS), adapted for propagation in Vero cell culture, was provided by the Servicio de Vacuna Antirrábica (SVAR), ANLIS “Dr. Carlos G. Malbrán”.
2.2. Construction of Recombinant MVA-RG
MVA-RG was generated as described before [14]. Briefly, the complete coding sequence of RABV glycoprotein (hereafter named RG) from RABV strain CVS was subcloned into the transfer vector (TV) TV-MTK-GUS-EL [14]. This plasmid also codifies for the β- glucuronidase enzyme (GUS), that facilitates screening and plaque lysis purification of recombinant MVAs by using its substrate, and the viral genomic regions of the MVA086R gene [which codifies for the thymidine kinase (TK) enzyme]. TV-RG was transfected into CEFs previously infected with MVA, and pure recombinant virus (100% of blue viral plaques) was achieved after 15 plaque-purification rounds. The presence of the RG sequence was confirmed by PCR using primers RG1-RG4 [15].
Expression of RG sequence was evaluated at the transcriptional level by RT-PCR. Total RNA from MVA or MVA-RG infected cells was extracted using TRIzolTM (ThermoFisher Scientific) and reverse-transcribed with the reverse transcriptase M-MLVRT (200 U, Promega) using random hexamers. The cDNA obtained was used to amplify by PCR a product of approximately 1600 bp with primers RG1 and RG2 [15].
2.3. Generation of Recombinant Adenoviruses
The ViraPower™ Adenoviral Gateway Expression Kit™ (ThermoFisher Scientific) system was employed for the generation of recombinant adenoviruses. The complete sequence of RG was cloned into the entry vector (pCR8TM/GW/TOPOTM TA Cloning) and then an in vitro recombination assay was performed using this plasmid and the destination vector (pAd/CMV/V5-DEST™, containing replication incompetent Ad5 genome, ∆E1/E3) following the manufacturer’s instructions. Transfection of HEK293A cells was performed using the cationic lipid Lipofectamine 2000 (ThermoFisher Scientific) and incubated until characteristic cytopathic effect of adenovirus was observed (around 10-13 days post-transfection). The presence of the RG sequence was determined by PCR using universal primers T7-promoter (5’TAATACGACTCACTATAGGG) and V5-reverse (5’ACCGAGGAGAGGGTTAGGGAT), which hybridizes in sequences present in the destination plasmid. As described above, expression of RG sequence was evaluated at the transcriptional level by RT-PCR. Total RNA from uninfected or Ad-RG infected cells was extracted using TRIzolTM and reverse-transcribed with M-MLVRT using random hexamers. The cDNA obtained was used to amplify by PCR a product of approximately 900 bp with primers RG1 and RG4 [15].
2.4. Animals
Female BALB/c (H-2d) mice (6-8 weeks old), certified as specific pathogen-free, were purchased from Fundación Facultad de Ciencias Veterinarias (UNLP, La Plata, Argentina) or from the SVAR. Immunizations were performed in animal facilities at the Instituto de Biotecnología (IABIMO, INTA-CONICET) and animals were moved to the SVAR for the RABV intracerebral (IC) challenge. All experiments were carried out following international welfare guidelines with the approval of the Comité Institucional para el Cuidado y Uso de Animales de Experimentación, CICUAE-CNIA, INTA, Argentina, and with provision No. 6344/96 of ANMAT, Argentina (which regulates the animal facilities of laboratories that manufacture medicinal specialties and/or conduct analyses for third parties). To minimize mice suffering, isoflurane and CO2 inhalation were used as anesthetic and sacrifice methods, respectively.
2.5. Evaluation of Efficacy of Recombinant Viral Vectors in Homologous and Heterologous Immunization Schemes
Groups of 5-10 animals were immunized with the corresponding immunogen (MVA-RG or Ad-RG) on days 0 and 21 as detailed in Tables 1 and 2. Additionally, mice of control groups were injected with recombinant viruses encoding a non-related protein (MVA-GFP or Ad-GFP), PBS or the commercial rabies vaccine VeroRab (Sanofi Pasteur). On day 28-30 post vaccination, animals were moved to the SVAR to perform the RABV IC challenge (see below).
2.6. RABV Intracerebral Challenge
Fourteen days after the booster, all the animals were challenged with 30 μL of RABV CVS strain (20-50 LD50/0.03 mL) by the IC route. Mice clinical signs and survival were registered daily for 2 weeks and animals were euthanized when they developed clinical signs of rabies. Deaths that occurred in the first 4 days after the challenge were considered non-specific. Protection was calculated as the number of animals that survived out of the total number of animals.
3. Results
3.1. Construction and Molecular Characterization of Recombinant MVA and Ad5 Expressing Rabies Glycoprotein (RG)
Recombinant MVA containing the complete coding sequence of RABV glycoprotein interrupting the MVA086R viral gene (TK) were obtained by in vivo homologous recombination between the TV-RG and MVA genome (Figure 1A). Recombinant viruses (hereafter named MVA-RG) were isolated by plaque-purification based on its capability to produce blue lysis plaques in the presence of X-Gluc (substrate of the GUS marker enzyme). The presence of the RG sequence in MVA-RG was confirmed by PCR amplification of a 922 bp fragment (Figure 1B).
The recombinant Ad5 carrying the RG sequence, Ad-RG, was generated using the ViraPower™ Adenoviral Gateway Expression Kit™ (ThermoFisher Scientific) (Figure 1D). The inserted nucleotide sequence was confirmed by PCR using T7-promoter and V5-reverse primers that amplify a fragment of approximately 1700 bp (Figure 1E).
The expression of RG sequence by MVA-RG and Ad-RG was determined at the transcriptional level by a RT-PCR assay using RG1/RG2 or RG1/RG4 primers, respectively. Indeed, the amplified fragments, of approximately 1600 or 900 bp, were only observed in samples from cells infected with the recombinant viruses (Figure 1C,F).
3.2. Protection Induced by MVA-RG and Ad-RG Against Intracerebral RABV Challenge
In order to test the level of protection induced by the recombinant viruses in an RABV challenge model, mice were injected twice with the corresponding dose of the viral vector, following either homologous or heterologous immunization schemes, and were challenged intracerebrally (IC) fourteen days post-booster.
Firstly, we administered 4 x 107 PFU of MVA-RG twice via the IP route (homologous scheme) or 1 x 108 PFU of Ad-RG as the prime and 4 x 107 PFU of MVA-RG (or MVA-GFP) as the boost (heterologous scheme). Control groups immunized with MVA-GFP, PBS, or a commercial anti-rabies vaccine were included (Table 1).
As shown in Figure 2, the homologous immunization scheme induced 60% protection against the RABV IC challenge, whereas 40% of the animals from the heterologous immunization scheme were protected. Since groups that received Ad-RG as the prime were equally protected after the challenge regardless of whether MVA-RG or MVA-GFP was used as the boost, we suggest that survival was primarily due to Ad-RG immunization. As expected, all animals immunized with PBS or MVA-GFP died by day 7 post-challenge, while those vaccinated with the commercial vaccine survived.
Secondly, we evaluated the protection induced by Ad-RG in homologous prime-boost schemes using different doses of the recombinant viral vector (Table 2). Control groups immunized with PBS, a commercial anti-rabies vaccine or a recombinant adenovirus encoding an unrelated protein (Ad-GFP) were included.
As shown in Figure 3, two immunizations with either 1 x 106 (Group 1) or 1 x 107 PFU (Group 2) of Ad-RG induced 70% and 60% protection, respectively, whereas 100% survival was observed in mice injected with 1 x 106 PFU for the prime and 1 x 107 PFU for the boost (Group 3). Control groups responded as expected after the challenge (PBS and Ad-GFP groups died, while the commercial vaccine group had 100% survival).
These results demonstrate that the MVA-RG and Ad-RG viral vectors induced protection against RABV. The survival rate depends on the type of viral vector, the dose administered, and the immunization scheme applied.
4. Discussion
Rabies vaccines predominantly used worldwide are based on inactivated rabies virus. Additionally, two viral vectored vaccines are commercially available for wildlife, utilizing replicative strains of vaccinia virus and human adenovirus 5, and one non-replicative vaccine based on canarypox virus for cats [5]. While inactivated vaccines are effective, they require multiple doses to achieve protective immunity and necessitate specialized facilities for their production. In contrast, viral vectored vaccines are also effective against rabies, stimulate both branches of the immune response, and do not involve the manipulation of the rabies virus during manufacturing.
Modified vaccinia Ankara virus (MVA) and human adenovirus type 5 ∆E (Ad5) are non-replicative in mammals and have been extensively evaluated as viral platforms for vaccine development [16].
In this study, we obtained two non-replicative viral vectors expressing the rabies glycoprotein, MVA-RG and Ad-RG. We then investigated the protection induced by these vaccine candidates in a mouse RABV intracerebral challenge model, using both homologous and heterologous immunization schemes.
After administering two injections of 4x107 PFU of MVA-RG (homologous immunization scheme), we observed 60% protection against RABV challenge. To enhance these protection levels, three strategies are possible: increasing the immunogen dose, augmenting the time interval between immunizations or applying a heterologous vaccination scheme.
Regarding the first option, Weyer et al. [17] reported 10% protection in mice immunized once with 1 x 108 PFU of MVA-RG, while 70-80% survival was recorded when 1 x 109 PFU were administered. However, using high doses of recombinant viruses in a mouse model poses challenges for scaling up its production for mass use in target animals, such as dogs, cats, or livestock.
Another possibility is the time interval between priming and boosting, as applying the booster during the contraction phase of the immune response may prevent any observed increase in the response. Several studies using poxvirus-based vaccines have demonstrated that extending the time interval between the priming and boosting doses improves vaccine-induced immunity [8,18,19,20]. In this context, it would be interesting to test whether increasing the interval between MVA-RG immunizations enhances protection levels against RABV challenge. However, a recent study with the MVA-SARS-2-ST and MVA-SARS-2-S vaccine candidates showed similar immune response levels when comparing 21-day and 56-day intervals between doses [21]. In addition to that, since our model involves an intracerebral RABV challenge, technical difficulties may arise in piercing the skull of the animals, which could increase their suffering or lead to greater variability in inoculum delivery.
Concerning the use of a heterologous vaccination regimen, there is extensive literature, both in humans and animal models, showing that prime-boost schemes combining adenoviruses and poxviruses enhance immunogenicity and efficacy against pathogens [22,23,24,25,26,27,28]
Thus, we applied a heterologous immunization scheme, priming with Ad-RG and boosting with MVA-RG, which generated 40% protection against RABV IC challenge. Under the conditions tested in our study this strategy did not enhance the level of protection observed with the homologous scheme. One possible explanation is that the strong immune response elicited by the Ad-RG dose used for priming (1 x108 PFU) may have inhibited the effect of MVA-RG. Additionally, as mentioned above, increasing the time interval between immunizations could enhance the specific response. In this sense, Bruña-Romero et al. [25] reported complete protection against malaria and improved levels of cellular and humoral immune response by reducing the prime dose with a replication-defective recombinant adenovirus expressing the circumsporozoite (CS) protein of Plasmodium yoelii from 1x109 PFU to 1x108 PFU and extending the time interval for boost application with an attenuated recombinant vaccinia virus expressing the same malaria antigen, VacPyCS from 2 to 8 weeks. Unfortunately, this strategy cannot be easily extrapolated to our IC challenge model.
The 40% protection after RABV IC challenge observed in the heterologous immunization scheme could be attributed to the administration of a single dose of 1 x108 PFU of Ad-RG, as the same level of protection was noted after boosting with either MVA encoding the rabies glycoprotein or an unrelated protein (GFP). This protection level is lower than the reported by Zhao et al. [29] and Yan et al. [30]. Those authors describe that a single injection of 1 x 108 GFU (green fluorescent units) of recombinant Ad5, expressing both rabies glycoprotein and canine distemper virus hemagglutinin or severe fever with thrombocytopenia syndrome virus, induces 100% protection against RABV IM challenge. On the other hand, Kim et al. [2] reported 100% protection following IM immunization with 2 x 107 PFU of Ad5 expressing either the full-length or a truncated version of the RABV glycoprotein after IM challenge, but after IC challenge 0% to 40% protection was observed. These results suggest that the IC challenge route is more stringent than the IM route, emphasizing the importance of carefully analyzing efficacy results.
Continuing our study and considering that a single dose of Ad-RG induced 40% protection it was of great interest to evaluate the efficacy of Ad-RG in a homologous prime-boost scheme. We observed a similar percentage of protection against RABV with two injections of 1 x 106 or 1 x 107 PFU, achieving 70% and 60% protection, respectively. These levels of protection were higher than those induced by a single inoculation of 1 x 108 PFU. Notably, 100% survival was observed when a dose of 1 x 106 PFU was administered at the priming stage, followed by a dose of 1 x 107 PFU during the boost. Although we did not evaluate the profile of the immune response generated by Ad-RG, these results suggest that the anti-vector response induced by priming with 1x107 PFU might inhibit the booster effect on the anti-RABV specific response.
In this sense, one limitation of Ad-based vaccines is the induction of an anti-vector immune response. This immune response inhibits cell transduction, limits transgene expression, and ultimately diminishes the overall effectiveness of the vaccination. This “inhibitory” effect may vary depending on the types of adenoviruses, the dose applied and the route of administration. Pandey et al. [31] analyzed the interference of anti-vector immune responses with antigen-specific responses. They immunized BALB/c mice with different doses of wild-type human adenovirus serotype 5 via the intranasal (IN) route (the natural infection route for adenoviruses) or the IM route (commonly used in Ad-based vaccines). Four weeks later, the mice received a booster dose with different amounts of HAd-HA-NP (HAd expressing the hemagglutinin -HA- and nucleoprotein -NP- of A/Vietnam/1203/04, H5N1 influenza virus) and were subsequently challenged with a non-lethal reassortant influenza virus. The authors observed that Ad-neutralizing antibody responses increased with higher prime doses, and these responses influenced the efficacy induced by different booster doses. The best level of protection was achieved with a low dose at prime (1x107 PFU) followed by a higher dose at boost (1x108 PFU). In another study, anti-Ad5 immunity was induced administering different doses of a GFP-recombinant HAd5V (Ad5GFP) via IN or IM routes. Subsequently, mice were immunized subcutaneously (SC) with 1×109 PFU/animal of HAd5VCMV. The authors found that only the highest dose of Ad5GFP (1x107 PFU) administered via the IM route reduces the CD8+ T cells specific response against the heterologous antigen (CMV) by 50% [32].
Given the significance of NA against Ad5 in reducing its effectiveness for human vaccines development, focus has turned to the generation of vectors based on animal Ad. Among these, chimpanzee-derived Ad vectors (ChAd) are the most extensively studied. While NA against simian Ad vectors are relatively uncommon, the potential impact of cross-reactive pre-existing cellular immunity against Ad should not be underestimated (reviewed by [33]). There are several rabies vaccine candidates based on ChAd that have demonstrated efficacy against RABV in both mice and non-human primates [34,35]. In recent years, two of these candidates, ChAdOx2-RG and ChAd155-RG, have been evaluated in phase I human clinical trials showing acceptable anti-RABV NA titers but this response was lower (levels and duration) than the induced by a standard inactivated rabies virus vaccine [36,37].
Overall, our results suggest that performing a homologous prime-boost regimen with Ad5 should not be dismissed as a possible vaccine strategy against RABV, as the induction (or lack thereof) of an antigen-specific response will depend on the levels of anti-vector response caused by a prior vaccination.
In this study, we presented the development of two third-generation rabies vaccine candidates based on non-replicative viral vectors in mammals, MVA and Ad5. Both candidates, administered in homologous vaccination regimens, induced protection against RABV IC challenge. In the future, it would be interesting to investigate whether the protection conferred by MVA-RG can be improved by increasing the time interval between inoculations. Additionally, the results obtained with Ad-RG homologous prime (1x106 PFU) – boost (1x107 PFU) scheme encourage us to proceed with the evaluation of this vaccine candidate in target animals.
5. Conclusions
In the context of developing third-generation vaccines that do not contain rabies virus in their formulation and induce protective immune responses, viral vectors represent an excellent alternative. Moreover, the ability to implement homologous vaccination regimens simplifies their production in industrial systems. Besides, their safety profile makes them suitable for use in both humans and animals (companion and production animals), achieving high levels of protection without the risk of spreading either the infectious agent or the viral vector.
Author Contributions
Conceptualization: GC, OP, MDMZ. Methodology: DG, GC, MM, OP. Formal analysis: DG, MM. Investigation: DG, MM, ATL. Resources: GC, MDMZ, OP. Writing—original draft preparation: DG, MDMZ. Writing—review and editing: GC, MDMZ. Visualization: GC, MDMZ. Supervision: OP, GC. MDMZ. Project administration: GC, MDMZ. Funding acquisition: GC, MDMZ. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Instituto Nacional de Tecnología Agropecuaria (INTA) (grants numbers PNBIO 1131032, PD i116) and the Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT) (grants numbers PICT 2013-535; PICT 2020-1713). ATL holds a doctoral fellowship from CONICET.
Institutional Review Board Statement
The animal studies protocols were approved by the Experimental Animal Welfare Ethics Committee of INTA, Argentina. Protocol codes: 38/2017; 57/2017; 37/2021; 32/2022; 20/2023.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors would like to thank Mr. Silvio Diaz, Mrs. Elena Scarel and Mr. Guillermo Fernandez for his support in handling and care of animals and Mr. Diego Compaired, Mrs. Yesica Gonzalez and Dr. Osvaldo Zabal for preparing the primary chicken embryo fibroblasts and maintaining cell lines.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the study design, manuscript writing, or decision to publish the results.
References
- Wunner, W.H.; Conzelmann, K.-K. Rabies Virus. In Rabies (fourth edition); Fooks, A.R., Jackson, A.C., Eds.; Academic Press, 2020; pp. 43–81 ISBN 9780128187050.
- Kim, H.H.; Yang, D.K.; Nah, J.J.; Song, J.Y.; Cho, I.S. Comparison of the Protective Efficacy between Single and Combination of Recombinant Adenoviruses Expressing Complete and Truncated Glycoprotein, and Nucleoprotein of the Pathogenic Street Rabies Virus in Mice. Virol J 2017, 14. [CrossRef]
- Ertl, H.C.J. Next Generation of Rabies Vaccines. In Rabies; Fooks, A.R., Jackson, A.C., Eds.; Academic Press, 2020; pp. 509–526 ISBN 9780128187050.
- Rupprecht, C.E.; Buchanan, T.; Cliquet, F.; King, R.; Müller, T.; Yakobson, B.; Yang, D.K. A Global Perspective on Oral Vaccination of Wildlife against Rabies. J Wildl Dis 2024, 60, 241–284. [CrossRef]
- Del Médico Zajac, M.; Garanzini, D.; Pérez, O.; Calamante, G. Recombinant Veterinary Vaccines Against Rabies: State of Art and Perspectives. In Emerging and Reemerging Viral Pathogens; Ennaji, M.M., Ed.; Academic Press, 2020; pp. 225–242 ISBN 9780128149669.
- Wiktor, T.J.; Macfarlan, R.I.; Reagan, K.J.; Dietzschold, B.; Curtis, P.J.; Wunner, W.H.; Kieny, M.P.; Lathe, R.; Lecocq, J.P.; Mackett, M. Protection from Rabies by a Vaccinia Virus Recombinant Containing the Rabies Virus Glycoprotein Gene. Proc Natl Acad Sci U S A 1984, 81, 7194–7198. [CrossRef]
- Yarosh, O.K.; Wandeler, A.I.; Graham, F.L.; Campbell, J.B.; Prevec, L. Human Adenovirus Type 5 Vectors Expressing Rabies Glycoprotein. Vaccine 1996, 14, 1257–1264. [CrossRef]
- Altenburg, A.F.; Kreijtz, J.H.C.M.; de Vries, R.D.; Song, F.; Fux, R.; Rimmelzwaan, G.F.; Sutter, G.; Volz, A. Modified Vaccinia Virus Ankara (MVA) as Production Platform for Vaccines against Influenza and Other Viral Respiratory Diseases. Viruses 2014, 6, 2735–2761. [CrossRef]
- Volz, A.; Sutter, G. Modified Vaccinia Virus Ankara. In Advances in virus research; 2017; Vol. 97, pp. 187–243.
- Sánchez-Sampedro, L.; Perdiguero, B.; Mejías-Pérez, E.; García-Arriaza, J.; Di Pilato, M.; Esteban, M. The Evolution of Poxvirus Vaccines. Viruses 2015, 7, 1726–1803. [CrossRef]
- Fougeroux, C.; Holst, P.J. Future Prospects for the Development of Cost-Effective Adenovirus Vaccines. Int J Mol Sci 2017, 18. [CrossRef]
- Zhang, H.; Wang, H.; An, Y.; Chen, Z. Construction and Application of Adenoviral Vectors. Mol Ther Nucleic Acids 2023, 34. [CrossRef]
- Chavda, V.P.; Bezbaruah, R.; Valu, D.; Patel, B.; Kumar, A.; Prasad, S.; Kakoti, B.B.; Kaushik, A.; Jesawadawala, M. Adenoviral Vector-Based Vaccine Platform for COVID-19: Current Status. Vaccines (Basel) 2023, 11. [CrossRef]
- Ferrer, M.F.; Del Médico Zajac, M.P.; Zanetti, F.A.; Valera, A.R.; Zabal, O.; Calamante, G. Recombinant MVA Expressing Secreted Glycoprotein D of BoHV-1 Induces Systemic and Mucosal Immunity in Animal Models. Viral Immunol 2011, 24, 331–339. [CrossRef]
- Zanetti, F.A.; Rudak, L.; Micucci, M.; Conte Grand, D.; Luque, A.; Russo, S.; Taboga, O.; Perez, O.; Calamante, G. Obtención y Evaluación Preliminar de Un Virus Canarypox Recombinante Como Candidato a Vacuna Antirrábica. Rev Argent Microbiol 2012, 44, 75–84.
- Wang, S.; Liang, B.; Wang, W.; Li, L.; Feng, N.; Zhao, Y.; Wang, T.; Yan, F.; Yang, S.; Xia, X. Viral Vectored Vaccines: Design, Development, Preventive and Therapeutic Applications in Human Diseases. Signal Transduct Target Ther 2023, 8, 149. [CrossRef]
- Weyer, J.; Rupprecht, C.E.; Mansc, J.; Viljoen, G.J.; Nel, L.H. Generation and Evaluation of a Recombinant Modified Vaccinia Virus Ankara Vaccine for Rabies. Vaccine 2007, 25, 4213–4222. [CrossRef]
- Jackson, L.A.; Frey, S.E.; El Sahly, H.M.; Mulligan, M.J.; Winokur, P.L.; Kotloff, K.L.; Campbell, J.D.; Atmar, R.L.; Graham, I.; Anderson, E.J.; et al. Safety and Immunogenicity of a Modified Vaccinia Ankara Vaccine Using Three Immunization Schedules and Two Modes of Delivery: A Randomized Clinical Non-Inferiority Trial. Vaccine 2017, 35, 1675–1682. [CrossRef]
- Pitisuttithum, P.; Nitayaphan, S.; Chariyalertsak, S.; Kaewkungwal, J.; Dawson, P.; Dhitavat, J.; Phonrat, B.; Akapirat, S.; Karasavvas, N.; Wieczorek, L.; et al. Late Boosting of the RV144 Regimen with AIDSVAX B/E and ALVAC-HIV in HIV-Uninfected Thai Volunteers: A Double-Blind, Randomised Controlled Trial. Lancet HIV 2020, 7, e238–e248. [CrossRef]
- Palgen, J.L.; Tchitchek, N.; Rodriguez-Pozo, A.; Jouhault, Q.; Abdelhouahab, H.; Dereuddre-Bosquet, N.; Contreras, V.; Martinon, F.; Cosma, A.; Lévy, Y.; et al. Innate and Secondary Humoral Responses Are Improved by Increasing the Time between MVA Vaccine Immunizations. NPJ Vaccines 2020, 5. [CrossRef]
- Kalodimou, G.; Jany, S.; Freudenstein, A.; Schwarz, J.H.; Limpinsel, L.; Rohde, C.; Kupke, A.; Becker, S.; Volz, A.; Tscherne, A.; et al. Short- and Long-Interval Prime-Boost Vaccination with the Candidate Vaccines MVA-SARS-2-ST and MVA-SARS-2-S Induces Comparable Humoral and Cell-Mediated Immunity in Mice. Viruses 2023, 15. [CrossRef]
- Alharbi, N.K.; Padron-Regalado, E.; Thompson, C.P.; Kupke, A.; Wells, D.; Sloan, M.A.; Grehan, K.; Temperton, N.; Lambe, T.; Warimwe, G.; et al. ChAdOx1 and MVA Based Vaccine Candidates against MERS-CoV Elicit Neutralising Antibodies and Cellular Immune Responses in Mice. Vaccine 2017, 35, 3780–3788. [CrossRef]
- Kou, Y.; Wan, M.; Shi, W.; Liu, J.; Zhao, Z.; Xu, Y.; Wei, W.; Sun, B.; Gao, F.; Cai, L.; et al. Performance of Homologous and Heterologous Prime-Boost Immunization Regimens of Recombinant Adenovirus and Modified Vaccinia Virus Ankara Expressing an Ag85B-TB10.4 Fusion Protein against Mycobacterium Tuberculosis. J Microbiol Biotechnol 2018, 28, 1022–1029. [CrossRef]
- Kim, Y.C.; Dema, B.; Rodriguez-Garcia, R.; López-Camacho, C.; Leoratti, F.M.S.; Lall, A.; Remarque, E.J.; Kocken, C.H.M.; Reyes-Sandoval, A. Evaluation of Chimpanzee Adenovirus and MVA Expressing TRAP and CSP from Plasmodium Cynomolgi to Prevent Malaria Relapse in Nonhuman Primates. Vaccines (Basel) 2020, 8, 1–14. [CrossRef]
- Bruña-Romero, O.; Gonzá Lez-Aseguinolaza, G.; Hafalla, J.C.R.; Tsuji, M.; Nussenzweig, R.S. Complete, Long-Lasting Protection against Malaria of Mice Primed and Boosted with Two Distinct Viral Vectors Expressing the Same Plasmodial Antigen. Proc Natl Acad Sci U S A 2001, 98, 11491–11496. [CrossRef]
- Coughlan, L.; Sridhar, S.; Payne, R.; Edmans, M.; Milicic, A.; Venkatraman, N.; Lugonja, B.; Clifton, L.; Qi, C.; Folegatti, P.M.; et al. Heterologous Two-Dose Vaccination with Simian Adenovirus and Poxvirus Vectors Elicits Long-Lasting Cellular Immunity to Influenza Virus A in Healthy Adults. EBioMedicine 2018, 29, 146–154. [CrossRef]
- Paris, R.M.; Kim, J.H.; Robb, M.L.; Michael, N.L. Prime-Boost Immunization with Poxvirus or Adenovirus Vectors as a Strategy to Develop a Protective Vaccine for HIV-1. Expert Rev Vaccines 2010, 9, 1055–1069. [CrossRef]
- Folegatti, P.M.; Flaxman, A.; Jenkin, D.; Makinson, R.; Kingham-Page, L.; Bellamy, D.; Lopez, F.R.; Sheridan, J.; Poulton, I.; Aboagye, J.; et al. Safety and Immunogenicity of Adenovirus and Poxvirus Vectored Vaccines against a Mycobacterium Avium Complex Subspecies. Vaccines (Basel) 2021, 9. [CrossRef]
- Zhao, Z.; Zheng, W.; Yan, L.; Sun, P.; Xu, T.; Zhu, Y.; Liu, L.; Tian, L.; He, H.; Wei, Y.; et al. Recombinant Human Adenovirus Type 5 Co-Expressing RABV G and SFTSV Gn Induces Protective Immunity Against Rabies Virus and Severe Fever With Thrombocytopenia Syndrome Virus in Mice. Front Microbiol 2020, 11. [CrossRef]
- Yan, L.; Zhao, Z.; Xue, X.; Zheng, W.; Xu, T.; Liu, L.; Tian, L.; Wang, X.; He, H.; Zheng, X. A Bivalent Human Adenovirus Type 5 Vaccine Expressing the Rabies Virus Glycoprotein and Canine Distemper Virus Hemagglutinin Protein Confers Protective Immunity in Mice and Foxes. Front Microbiol 2020, 11. [CrossRef]
- Pandey, A.; Singh, N.; Vemula, S. V.; Couëtil, L.; Katz, J.M.; Donis, R.; Sambhara, S.; Mittal, S.K. Impact of Preexisting Adenovirus Vector Immunity on Immunogenicity and Protection Conferred with an Adenovirus-Based H5N1 Influenza Vaccine. PLoS One 2012, 7. [CrossRef]
- De Andrade Pereira, B.; Bouillet, L.E.M.; Dorigo, N.A.; Fraefel, C.; Bruna-Romero, O. Adenovirus Specific Pre-Immunity Induced by Natural Route of Infection Does Not Impair Transduction by Adenoviral Vaccine Vectors in Mice. PLoS One 2015, 10. [CrossRef]
- Fausther-Bovendo, H.; Kobinger, G.P. Pre-Existing Immunity against Ad Vectors: Humoral, Cellular, and Innate Response, What’s Important? Hum Vaccin Immunother 2014, 10, 2875–2884. [CrossRef]
- Xiang, Z.Q.; Greenberg, L.; Ertl, H.C.; Rupprecht, C.E. Protection of Non-Human Primates against Rabies with an Adenovirus Recombinant Vaccine. Virology 2014, 450–451, 243–249. [CrossRef]
- Napolitano, F.; Merone, R.; Abbate, A.; Ammendola, V.; Horncastle, E.; Lanzaro, F.; Esposito, M.; Contino, A.M.; Sbrocchi, R.; Sommella, A.; et al. A next Generation Vaccine against Human Rabies Based on a Single Dose of a Chimpanzee Adenovirus Vector Serotype c. PLoS Negl Trop Dis 2020, 14, 1–26. [CrossRef]
- Jenkin, D.; Ritchie, A.J.; Aboagye, J.; Fedosyuk, S.; Thorley, L.; Provstgaad-Morys, S.; Sanders, H.; Bellamy, D.; Makinson, R.; Xiang, Z.Q.; et al. Safety and Immunogenicity of a Simian-Adenovirus-Vectored Rabies Vaccine: An Open-Label, Non-Randomised, Dose-Escalation, First-in-Human, Single-Centre, Phase 1 Clinical Trial. Lancet Microbe 2022, 3, e663–e671. [CrossRef]
- Phadke, V.K.; Gromer, D.J.; Rebolledo, P.A.; Graciaa, D.S.; Wiley, Z.; Sherman, A.C.; Scherer, E.M.; Leary, M.; Girmay, T.; McCullough, M.P.; et al. Safety and Immunogenicity of a ChAd155-Vectored Rabies Vaccine Compared with Inactivated, Purified Chick Embryo Cell Rabies Vaccine in Healthy Adults. Vaccine 2024, 42. [CrossRef]
Figure 1.
Molecular characterization of MVA-RG and Ad-RG. Schematic representation recombinant MVA (A) or Ad-RG (D) genome. Primers used for molecular characterization are indicated with arrows. Presence of RG sequence was determined by PCR amplification using primers RG1 and RG4 for MVA (B) or T7-promoter and V5-reverse for Ad (E). (B) Line 1: 1 Kb DNA Ladder (Invitrogen, ThermoFisher); Line 2: DNA extracted from CEFs infected with MVA-RG; Line 3: DNA extracted from CEFs infected with MVA. (E) Line 1: DNA extracted from HEK293A infected with Ad-RG; Line 2: 1 Kb DNA Ladder (Invitrogen, ThermoFisher); Line 3: No-template control; Line 4: pAd/CMV/V5-DEST™-RG. Corroboration of expression of RG RNA by RT-PCR using primers RG1 and RG2 (C) or RG1 and RG4 (F). (C) Line 1: Total RNA extracted from MVA infected CEFs; Line 2: Total RNA extracted from MVA-RG infected CEFs; Line 3: 1 Kb Plus DNA Ladder (PB-L Productos Bio-Lógicos®).; Line 4: Positive control, TV-RG. (F) Line 1: 1 Kb Plus DNA Ladder (PB-L Productos Bio-Lógicos®).; Line 2: Total RNA extracted from non-infected HEK293A; Line 3: Total RNA extracted from Ad-RG infected HEK293A; Line 4: Positive control, pAd/CMV/V5-DEST™-RG.
Figure 1.
Molecular characterization of MVA-RG and Ad-RG. Schematic representation recombinant MVA (A) or Ad-RG (D) genome. Primers used for molecular characterization are indicated with arrows. Presence of RG sequence was determined by PCR amplification using primers RG1 and RG4 for MVA (B) or T7-promoter and V5-reverse for Ad (E). (B) Line 1: 1 Kb DNA Ladder (Invitrogen, ThermoFisher); Line 2: DNA extracted from CEFs infected with MVA-RG; Line 3: DNA extracted from CEFs infected with MVA. (E) Line 1: DNA extracted from HEK293A infected with Ad-RG; Line 2: 1 Kb DNA Ladder (Invitrogen, ThermoFisher); Line 3: No-template control; Line 4: pAd/CMV/V5-DEST™-RG. Corroboration of expression of RG RNA by RT-PCR using primers RG1 and RG2 (C) or RG1 and RG4 (F). (C) Line 1: Total RNA extracted from MVA infected CEFs; Line 2: Total RNA extracted from MVA-RG infected CEFs; Line 3: 1 Kb Plus DNA Ladder (PB-L Productos Bio-Lógicos®).; Line 4: Positive control, TV-RG. (F) Line 1: 1 Kb Plus DNA Ladder (PB-L Productos Bio-Lógicos®).; Line 2: Total RNA extracted from non-infected HEK293A; Line 3: Total RNA extracted from Ad-RG infected HEK293A; Line 4: Positive control, pAd/CMV/V5-DEST™-RG.
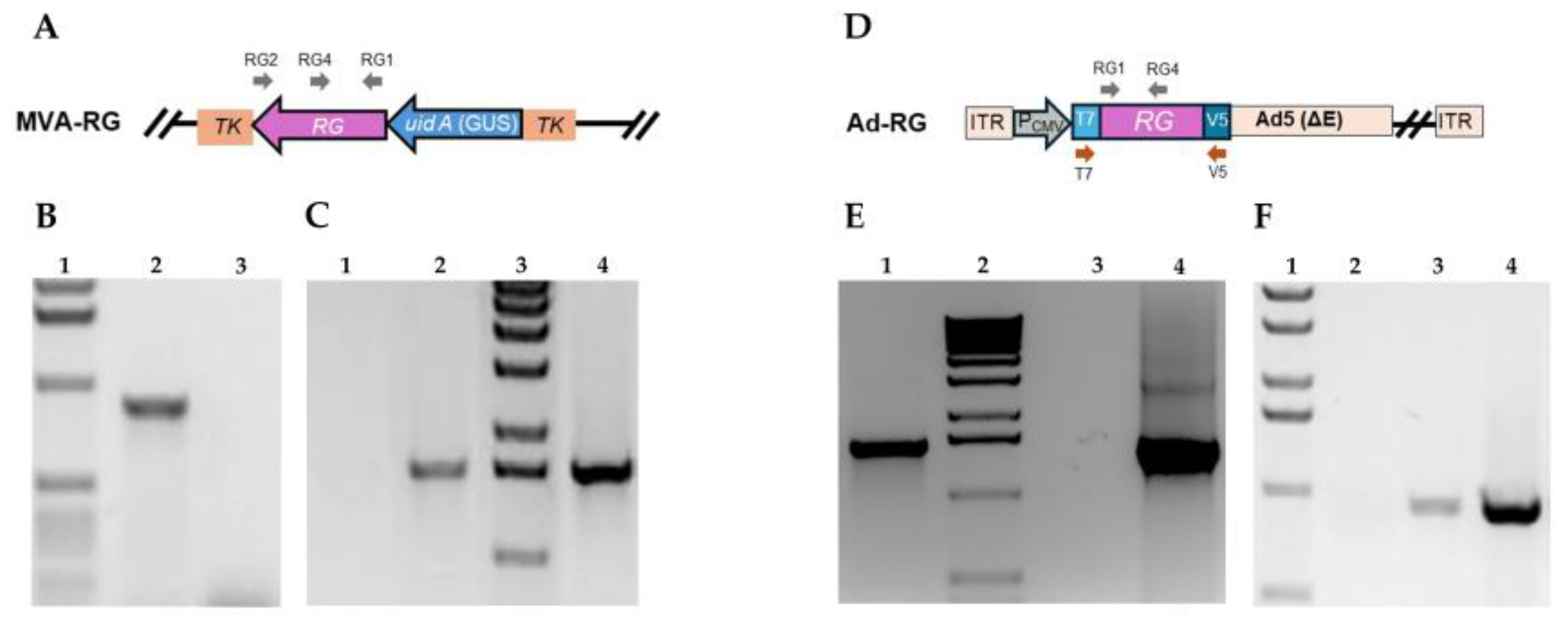
Figure 2.
Protection against RABV induced by MVA-RG in homologous or heterologous immunization schemes. Mice (n=5/10) were inoculated with MVA-RG as indicated in Table 1. Mice from control groups were inoculated with recombinant viruses expressing an unrelated protein (MVA-GFP and Ad-GFP), PBS or the commercial anti-rabies vaccine (Verorab®). Fourteen days post booster, all groups were IC challenged with 12,5-50 LD50 of RABV CVS strain. Survival of mice was checked daily for 2 weeks, deaths that occurred in the first four days were considered unspecific.
Figure 2.
Protection against RABV induced by MVA-RG in homologous or heterologous immunization schemes. Mice (n=5/10) were inoculated with MVA-RG as indicated in Table 1. Mice from control groups were inoculated with recombinant viruses expressing an unrelated protein (MVA-GFP and Ad-GFP), PBS or the commercial anti-rabies vaccine (Verorab®). Fourteen days post booster, all groups were IC challenged with 12,5-50 LD50 of RABV CVS strain. Survival of mice was checked daily for 2 weeks, deaths that occurred in the first four days were considered unspecific.
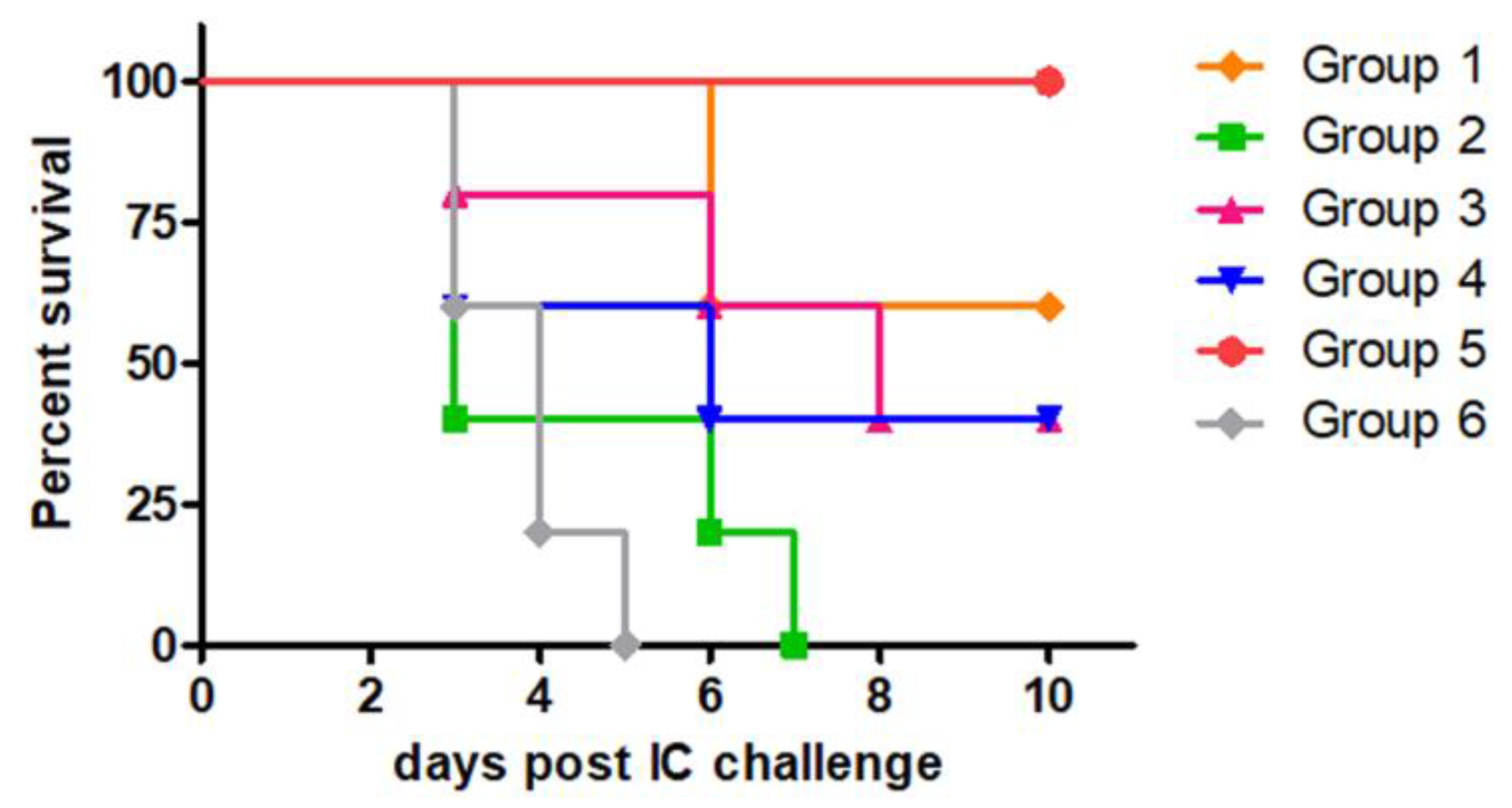
Figure 3.
Survival after RABV IC challenge induced by Ad-RG. Mice (n=5/10) were inoculated twice with different doses of Ad-RG as indicated in Table 2. Mice from control groups were inoculated with Ad-GFP, PBS or the commercial anti-rabies vaccine (Verorab®). Fourteen days post booster, all groups were IC challenged with 12,5-50 LD50 of RABV CVS strain. Survival of mice was checked daily for 2 weeks, deaths that occurred in the first four days were considered unspecific.
Figure 3.
Survival after RABV IC challenge induced by Ad-RG. Mice (n=5/10) were inoculated twice with different doses of Ad-RG as indicated in Table 2. Mice from control groups were inoculated with Ad-GFP, PBS or the commercial anti-rabies vaccine (Verorab®). Fourteen days post booster, all groups were IC challenged with 12,5-50 LD50 of RABV CVS strain. Survival of mice was checked daily for 2 weeks, deaths that occurred in the first four days were considered unspecific.
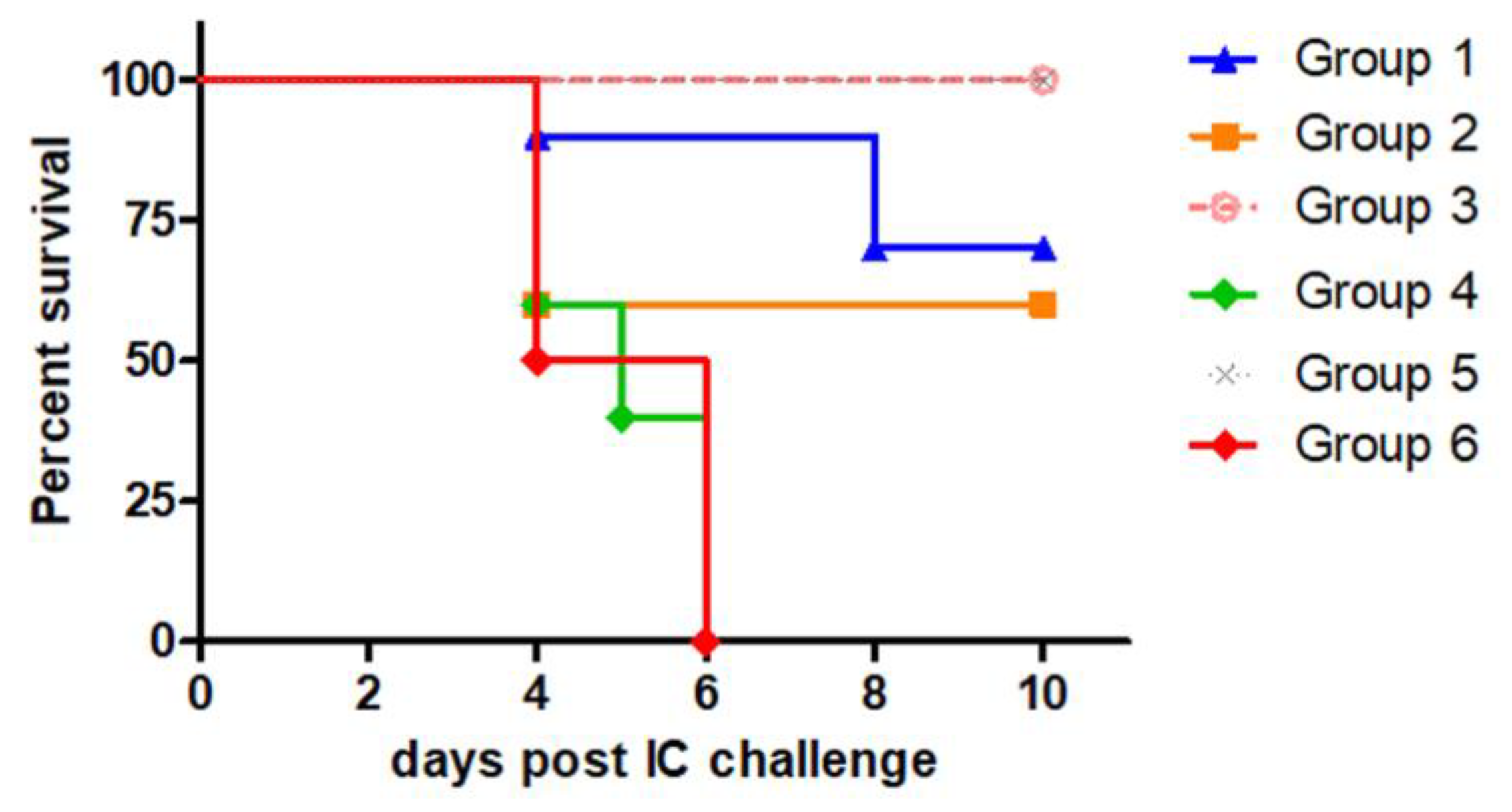

Table 1.
Immunogen and dose administered to mice in MVA-RG homologous and heterologous schemes.
| Prime | Boost | |
|---|---|---|
| Group 1 | MVA-RG (4 x 107 PFU, IP) | MVA-RG (4 x 107 PFU, IP) |
| Group 2 | MVA-GFP (1 x 107 PFU, IP) | MVA-GFP (1 x 107 PFU, IP) |
| Group 3 | Ad-RG (1 x 108 PFU, IM) | MVA-RG (4 x 107 PFU, IP) |
| Group 4 | Ad-RG (1 x 108 PFU, IM) | MVA-GFP (1 x 107 PFU, IP) |
| Group 5 | Commercial vaccine (IP) | Commercial vaccine (IP) |
| Group 6 | PBS (IP) | PBS (IP) |
Table 2.
Dose of Ad-RG administered to mice at the prime and the boost.
| Prime | Boost | |
|---|---|---|
| Group 1 | Ad-RG 1 x 106 PFU (IM) | Ad-RG 1 x 106 PFU (IM) |
| Group 2 | Ad-RG 1 x 107 PFU (IM) | Ad-RG 1 x 107 PFU (IM) |
| Group 3 | Ad-RG 1 x 106 PFU (IM) | Ad-RG 1 x 107 PFU (IM) |
| Group 4 | Ad-GFP 1 x 106 PFU (IM) | Ad-GFP 1 x 106 PFU (IM) |
| Group 5 | Commercial vaccine (IP) | Commercial vaccine (IP) |
| Group 6 | PBS (IP) | PBS (IP) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



