Submitted:
20 December 2024
Posted:
23 December 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Mycoplasma is the smallest, independently replicating microbe that lacks a cell wall and resembles a liposome because of its sterol-rich cell membrane. Under pathological circumstances, Mycoplasmas cause respiratory and urogenital infections in humans.
Human immunodeficiency virus (HIV) is known to be associated with Mycoplasma genitalium, suggesting that viruses may commonly utilize Mycoplasmas to enhance infec-tivity. Along this line, COVID-19 critical illness was documented in individuals with con-comitant Mycoplasma infection, raising the possibility of a symbiotic relationship or binary biological weapon.
Binary biological weapons are comprised of two components that are safe to handle separately but, in combination, be-come lethal. For example, concomitant infection with hepatitis D and B is deadly because the D virus takes advantage of the proteins expressed by the B virus, turning hepatitis B into a fatal disease.
Mycoplasmas have been known for their neuropsychiatric manifestations, including psychosis, anxiety, depressive dis-orders, cognitive impairment, and schizophrenia. This is relevant as mycoplasmas, like many antipsychotic drugs, enter host cells via clathrin-dependent endocytic pathways, often dormant in endosomes. Moreover, Mycoplasmas express aryl hydrocarbon receptor (AhR), a transcription factor activated by many ligands relevant to neuropsychiatry, including dopamine, serotonin, melatonin, clozapine, carbidopa, tryptophan, and microbial metabolites. Mycoplasma AhR, like its human counterpart, senses environmental stimuli, including viruses, indicating that it may play a role in the Mycoplas-ma/SARS-CoV-2 symbiosis.
This perspective article examines Mycoplasma and its relationship with COVID-19, premature cellular senescence, and major depressive disorder. We also discuss neuropathology potentially associated with designer pathogens and some therapeutic strategies.
Keywords:
binary bioweapons
; symbiosis mycoplasma
; interventions
; mycoplasma in psychiatry
Introduction
Mycoplasma is the smallest, independently replicating microbe discovered by Pasteur in the eighteenth century. It can be parasitic or saprophytic and is found in plants, animals, insects, humus, water, and sewage.
Detection of Mycoplasma in Golf War Illness (GWI) veterans, patients with Human Immunodeficiency Virus (HIV) and COVID-19 critical illness has brought this bacterium to the attention of researchers and clinicians as a possible bioweapon [1,2]. This is further substantiated by the fact that Mycoplasma was first synthesized in a US military laboratory in 1993.
The recent COVID-19 pandemic has highlighted the disastrous impact a pathogen can have on the population and economy of the entire planet. Non-kinetic means for waging wars, often called fifth-generation warfare, involve information, economy, healthcare, education, and other aspects of life that form the tapestry of human society. Since battles are often won before ever fought, influencing the enemy may start years or even decades before the open hostilities. Making drug abuse available to the enemy population leads to addictions, chronic diseases, disability, depression, suicide, and plummeting birth rates. All these deplete the military-age population, influencing the outcome of present or future wars, while providing plausible deniability of planned interference in the internal affairs of other nations
The chronology of man-made Mycoplasma: Shyh-Ching Lo, chief of the Division of Molecular Pathology at the Armed Forces Institute of Pathology in Washington, built the first synthetic microorganism, Mycoplasma fermentans incognitus (Lo’s Mycoplasma) in 1993 (patent number 5,242,820). Interestingly, this Mycoplasma was detected in 45% of patients with GWI, linking this pathogen to fatiguing disorders. Along this line, Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS) and Fibromyalgia (FM) have been linked to Mycoplasma infection, indicating a predilection for chronic illnesses with muscle pain [3]. In addition, Mycoplasma fermentans incognitus strain enhances tissue plasminogen activator (tPA), which converts plasminogen to the biologically active plasmin, a protein that cleaves the S antigen of the SARS-CoV-2 virus at the S1/S2 site, indicating a symbiotic relationship between this virus and Mycoplasma, a cooperation reminiscent of Mycoplasma genitalium and Human Immunodeficiency Virus (HIV) [4,5].
The next step in Mycoplasma weaponization occurred in 1995, when the Institute for Genome Research in Rockville, Maryland, sequenced the genome of Mycoplasma genitalium, opening the door for synthetic biology in warfare [6].
The story of artificial Mycoplasma continues. In 2010, a completely synthetic Mycoplasma mycoides JCVI-syn1.0 was created in the laboratory, contributing further to the weaponization of this pathogen. After this, Mycoplasma JCVI-syn2.0 and 3.0 strains were developed by minimizing the genome to the smallest size compatible with survival [7,8].
Mycoplasmas are members of the Mollicutes class of bacteria, the most straightforward and smallest known self-replicating microorganisms [9]. They can be commensal or pathogenic, the former dwelling on the skin or oral cavity, while the latter are usually found in the intracellular compartment [10].
COVID-19 is characterized by variable clinical outcomes, ranging from few or no symptoms to critical illness and death. Several novel studies have found that coinfection with Mycoplasma, could be the primary driver of unfavorable COVID-19 outcomes [11,12]. Indeed, epidemiological studies found 47% comorbidity between SARS-CoV-2 and Mycoplasma, further indicating a symbiotic relationship. Mycoplasmas benefit viruses as they disrupt host immunity by blocking host antiviral antibodies. Moreover, viruses may also use Mycoplasma as a vehicle for entry into host cells [13].
Several Mycoplasmas have been known for their neuropsychiatric manifestations, including psychosis, anxiety, depressive disorders, cognitive impairment, and schizophrenia (SCZ) [14]. Interestingly, Mycoplasmas express aryl hydrocarbon receptor (AhR), a transcription factor activated by many ligands relevant to neuropsychiatry, including dopamine, serotonin, melatonin, clozapine, carbidopa, tryptophan, and microbial metabolites. The Mycoplasma AhR, like its human counterpart, senses environmental stimuli, including viruses, indicating that it may play a role in the Mycoplasma/SARS-CoV-2 symbiosis.
SARS-CoV-2 infection altered tryptophan catabolism, generating excessive kynurenine, a primary AhR activator [15]. Conversely, AhR inhibitors could block or disrupt Mycoplasma’s role in binary weapons by various actions, including altering the integrin motif, which connects pathogens with the host ECM [16,17]. ECM disruption can lead to increased blood-brain barrier (BBB) and gut barrier permeability, a characteristic of neuropsychiatric illnesses and inflammatory bowel disease (IBD) [18]. Furthermore, lipid-associated membrane proteins (LAMPs) of Mycoplasma fermentans and Mycoplasma hominis have been shown to upregulate cortisol secretion, connecting this bacterium to dysfunctional biological barriers, post-traumatic stress disorder (PTSD, and chronic fatigue [19].
Mycoplasmas destabilize the genome and exhibit carcinogenic potential. Host cells respond by activating cellular senescence, an established antitumor defense mechanism [20]. However, cellular senescence has been associated with several neuropsychiatric conditions, including MDD [21,22,23,24]. Virus-induced senescence (VIS) refers to the capability of many viruses, including SARS-CoV-2, to induce premature cellular/neuronal senescence and subsequent depression. Indeed, the prevalence of depression and suicide increased dramatically during the COVID-19 pandemic [25,26].
Biological Warfare, Synthetic Microbiology, and Virology
Biological warfare involves pathogens such as bacteria, viruses, and fungi that cause disease, death, or environmental damage [27]. As many of these agents enter the brain and trigger pathology, this definition also includes neuropsychiatric illness.
Biological warfare was used for thousands of years by military commanders throughout time, and civilizations realized that individuals who died of infectious diseases could be used as weapons if their bodies were left to decompose in the enemy water or food supply. Moreover, scarves from people with leprosy were often sold in enemy territory to ignite epidemics prior to war.
Synthetic biology started with Lo’s Mycoplasma in 1993 and has developed rapidly. Whether synthetic microorganisms affect the brain is currently unknown. Extinct bacteria or viruses were brought back to life without considering their interaction with modern microbes and humans and the possibility of triggering epidemics.
In 1997, Taubenberger and coworkers recovered viral RNA from the 1918 Spanish Flu in frozen tissues and bodies preserved in permafrost and rebuilt the virus despite the risk of an outbreak [28].
In 2002, Cello’s team constructed a synthetic poliovirus using original cDNA and RNA, officially establishing the discipline of synthetic virology. Moreover, ancient viruses that infected our ancestors, such as human endogenous retrovirus (HERVs) activation, were implemented in 2002-2003 regardless of the risks. The same reasoning was given for building synthetic SARS-CoV viruses using RNA from that year’s outbreak [29].
Binary bioweapons were unknown to the Western military establishment until 1992 when a Soviet Army biologist defected with this information. He later published a chapter, “Next Generation Bioweapons: Genetic Engineering and Biological Warfare,” in a 2004 book entitled “The Gathering Biological Warfare Storm.” In this work, Temple 23 (defector’s code name) described several Soviet binary bioweapon projects, including a nonvirulent plague (Yersinia pestis) that could be handled safely. Still, it became deadly and antibiotic-resistant in combination with a virus (classified).
Centers for Disease Control and Prevention (CDC) have categorized pathogens and toxins in three groups based on their potential use in developing biological weapons. Group A includes significant pathogens, such as Anthrax, Botulism, Plague, etc. Group B (biological modulators) include binary partners, plasmids, and microbial antigens. Group C consists of emerging pathogens, including synthetic bacteria and viruses, with a high potential for use in bioterrorism. It is noteworthy that both Mycoplasma and SARS-CoV-2 are enumerated as potential bioweapons: http://www.selectagents.gov/SelectAgentsandToxinsList.html and https://www.niaid.nih.gov/topics/biodefenserelated/biodefense/pages/cata.aspx.
Mycoplasmas, the Ubiquitous and Secretive Slow Killers
Mycoplasmas belong to the Mollicutes family, microbes without a cell wall, surrounded by a sterol-rich plasma membrane, engendering a liposome-like structure. Sterols and their principal representative, cholesterol, are components of the lipid rafts and, therefore, crucial for SARS-CoV-2 infectivity [30]. For example, people with altered raft zymosterol, 24-dehydrolathosterol, or desmosterol have been associated with poor COVID-19 prognosis, emphasizing the antiviral role of these lipids [31]. Indeed, impaired cholesterol synthesis enhances viral infection, suggesting that the SARS-CoV-2 thrives by usurping host cholesterol metabolism.
The clinical picture of viruses and Mycoplasma infection overlap at many points, further complicating the differential diagnosis [32,33]. Moreover, diagnostic tests, including Mycoplasma species serology, culture, and even nucleic acid amplification, such as PCR, are marred by numerous limitations [34]. In this regard, false-positive and --negative COVID-19 serological test results have been reported in many patients with Mycoplasma pneumoniae, highlighting the limitation of these assays [35]. The next-generation sequencing by shotgun methodology appears promising for differentiating Mycoplasma from SARS-CoV-2 and may help diagnose long COVID [36]. However, leukopenia, lymphocytopenia, thrombocytopenia, and thromboembolism were documented in SARS-CoV-2 and Mycoplasma infections, further highlighting their intertwined etiopathogenesis [37]. Furthermore, specific anti-microbial treatments, such as azithromycin or tetracyclines, were beneficial for Mycoplasma and COVID-19, suggesting a likely silent partnership between these quite different infections [38]. Another enhancer of the Mycoplasma/SARS-CoV-2 symbiosis, the RGD integrin motif, is expressed by both pathogens, suggesting that, independently of ACE-2, the integrin pathway enables viral entry [39]. Other ECM proteins, including reelin and AhR, have been implicated in severe mental illness, including SCZ. In addition, as both Mycoplasma and SARS-CoV-2 express fibronectin (FBN), these pathogens may fuse, engendering combined pathology [39]. It was suggested that the SARS-CoV-2 virus could be a bacteriophage “hiding” inside Mycoplasma thus, averting host immune defenses [41].
Integrin originates in the cytoskeleton, exits the cell through the cholesterol-rich lipid rafts, and changes length as it connects to intra and extracellular proteins. The dimensional shift of the integrin molecule acts as a switch, initiating or terminating the contact with other ECM proteins. Depending on the location of the ligand attachment, contact between intra and extracellular proteins may be activated or inhibited [42]. For example, when a ligand binds to the cytoplasmic domain of the integrin molecule, the extracellular integrin portion elongates, establishing contact with the ECM components [43,44]. Conversely, when a ligand binds to the extracellular portion, the integrin shortens, terminating the interaction with ECM proteins [45]. Integrins’ ability to influence signaling between intra and extracellular molecular assemblies, especially in neurons, highlights integrin’s transistor-like qualities, including access to Boolean logic gates, the building blocks of computation. This indicates that the SARS-CoV-2/Mycoplasma complex affects information processing by targeting integrins, probably accounting for cognitive changes or “brain fog” experienced by many COVID-19 patients [45,46]. This may also explain the role of ECM proteins, including integrin, reelin, and AhR, in psychosis, including SCZ. Interestingly, new-onset psychosis has been associated with integrins, AhR, and reelin.
Figure 1.
Integrins originate in the actin filaments of the cellular cytoskeleton, cross the cell membrane, and interact with the extracellular matrix (ECM) molecules. Uniting intra- and extracellular molecular assemblies is believed to support cognition and perhaps awareness. Ligand-binding to integrins lengthens or shortens the molecule, connecting and disconnecting intra- and extracellular molecular assemblies. This may explain the difference in information processing during wakefulness vs. sleep [47].
Figure 1.
Integrins originate in the actin filaments of the cellular cytoskeleton, cross the cell membrane, and interact with the extracellular matrix (ECM) molecules. Uniting intra- and extracellular molecular assemblies is believed to support cognition and perhaps awareness. Ligand-binding to integrins lengthens or shortens the molecule, connecting and disconnecting intra- and extracellular molecular assemblies. This may explain the difference in information processing during wakefulness vs. sleep [47].
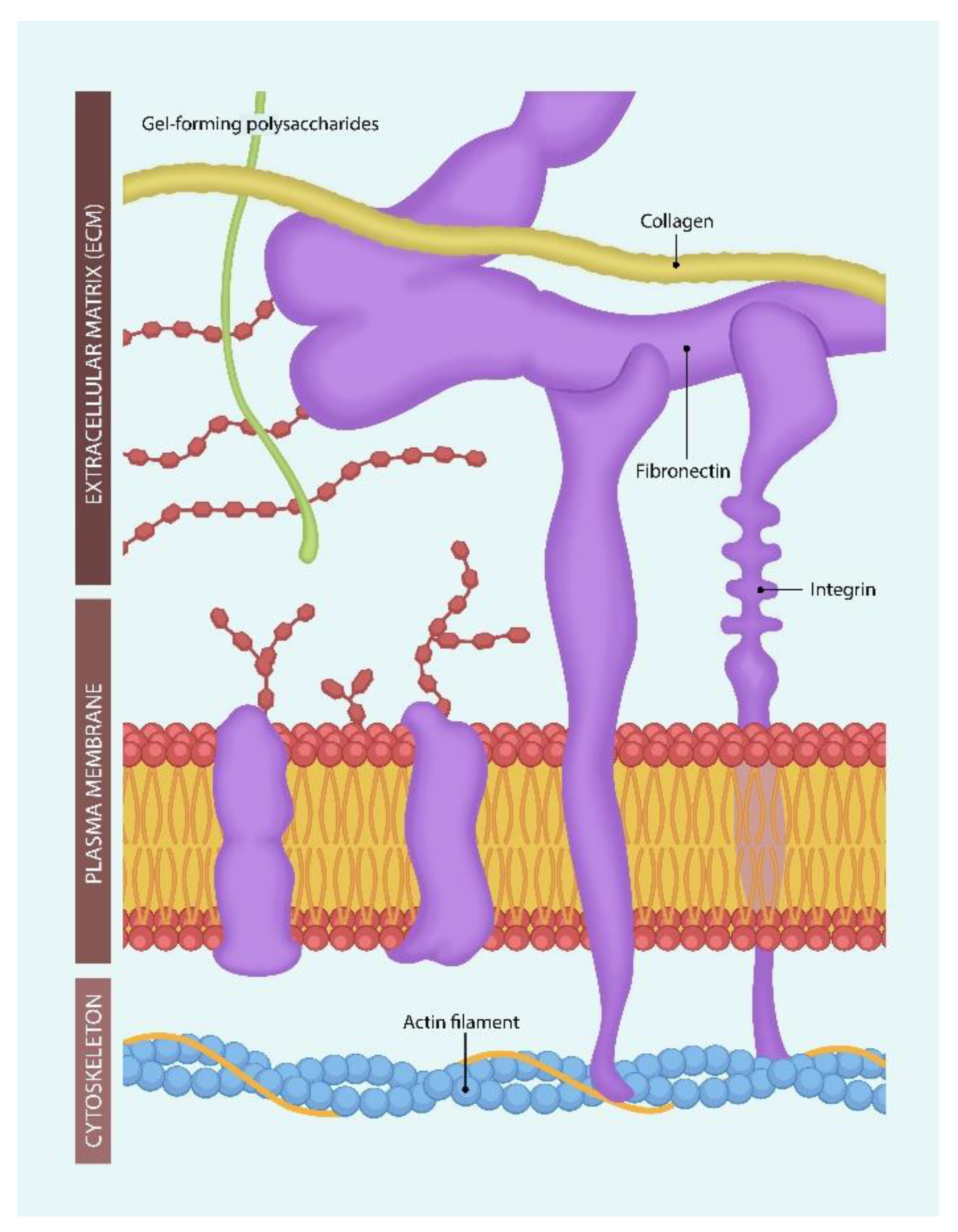
Putting it all together, Mycoplasma and SARS-CoV-2 bind to ECM proteins, disrupting the function of integrins in somatic cells and neurons along with information processing. Due to its liposome resemblance, Mycoplasma can be utilized as a virus-transporting vehicle, facilitating the development of binary bioweapons.
SARS-CoV-2 and Mycoplasma-Induced Premature Endothelial Senescence
In 1961, Leonard Hayflick discovered that human cells undergo 40-60 division cycles, after which they enter a state of proliferative arrest marked by an active and rewired metabolism that generates toxic molecules, known generically as senescence-associated secretory phenotype (SASP) [48]. Due to their intimate contact with the circulatory system, senescent endothelial cells (EC) release SASP directly into the systemic circulation, spreading senescence throughout the body. For this reason, many viruses, including SARS-CoV-2, usurp ECs, facilitating viral infection by activating VIS. Since depression and other neuropsychiatric disorders are associated with upregulated senescent markers such as p16, p53, SASP, major depressive disorder (MDD) (is 21) [49,50].
Senescent cells provide a hospitable environment for viral progeny due to the abundance of intracellular iron, which can be up to 40 times higher in old than in young cells [51]. In this regard, both the SARS-CoV-2 virus and Mycoplasma possess the capability of inducing premature cellular senescence, a feature exploited by the manufacturers of biological weapons. Indeed, the accelerated aging of the enemy population, affecting combat readiness, is highly desirable for population reduction and disseminating depression, docility, and reluctance for combat. Furthermore, depression treatment, especially SSRIs (selective serotonin reuptake inhibitors) may cause judgment errors, such as accepting unfair outcomes to avoid confrontation [52].
Angiotensin II (ANG II,) upregulated by the SARS-CoV-2 virus, induces cellular senescence, further contributing to organismal aging [53]. Indeed, combining VIS with other lethal means, such as drugs, makes much sense from foreign military planners’ standpoint because it reduces the number of combat-ready troops. Therefore, releasing a virus before a military conflict can be part of the overall planning,
At the cellular level, the accumulation of aging cells can overwhelm efferocytosis (elimination of damaged cells), exacerbating inflammation and promoting chronic diseases [54].
The key drivers of senescence-associated with SARS-CoV-2 and Mycoplasma are disturbances of cholesterol metabolism and the renin-angiotensin system (RAS).
Under normal circumstances, cholesterol synthesis in the liver regulates cellular senescence, while oxidated cholesterol likely disrupts this process, leading to premature senescence, as observed in COVID-19 and HIV [55,56]. Oxidized cholesterol affects the renin-angiotensin system (RAS) by increasing the activity of angiotensin II (ANG II), an endogenous toxin hydrolyzed by angiotensin-converting enzyme-2 (ACE-2) (Singh BM). Mycoplasma infection can increase ANG II levels, while some Mycoplasma strains possess specific ANG II binding sites, suggesting a direct interaction between this bacterium and RAS [56,57].
COVID-19 has been associated with dysfunctional RAS, as the SARS-CoV-2 virus attaching to ACE-2 disables this enzyme, leading to unopposed accumulation of ANG II and incapacitation of the entire protective RAS branch (Figure 2). The MasR axis has been implicated in the pathophysiology of depression and anxiety. At the same time, high levels of Ang-[1,2,3,4,5,6,7] exert antidepressant properties [59].
ANG II was associated with premature cellular senescence marked by telomere attrition, a common finding in severe COVID-19 and children with Hutchinson-Gilford progeria, a congenital syndrome of accelerated aging [60,61].
Putting it all in perspective, accelerated aging may have been the aim of the COVID/Mycoplasma binary complex. Premature cellular senescence and accelerated aging are a strategy to lower military combat readiness. Cellular senescence and subsequent depression and other neuropsychiatric conditions affect motivation, energy, and the level of alertness necessary for successful combat. Fatigue added to these symptoms contributes to weakness and lowers the combatants’ morale.
Potential Interventions
Combating the effects of the SARS-CoV-2/Mycoplasma complex requires reversing or preventing the onset of the senescent phenotype. Until recently, cellular senescence was considered permanent and irreversible. Currently, there is growing evidence that cellular senescence may be reversed. Moreover, senolytic drugs selectively eliminate senescent and damaged cells, while senomorphic compounds delete senescent markers, such as SASP, p16, or p21. Interestingly, these markers are directly related to MDD. For example, SASP is considered by some researchers as a biomarker for mood disorders [24]. Moreover, as the integrin motif connects Mycoplasma with the virus, anti-integrin drugs might be an option to prevent Mycoplasma/SARS-CoV-2 interaction (Figure 3).
Phosphoinositide-Dependent Kinase 1 (PDK-1) Inhibitors
Novel data have shown that inhibitors of phosphoinositide-dependent kinase 1 (PDK-1), such as kaempferol (also an AhR antagonist) may reverse the senescent phenotype [63]. On the other hand, PDK1 activation of protein kinase B (Akt) and glycogen synthase kinase 3 beta (GSK-3β) contribute to the pathogenesis of severe neuropsychiatric disorders [64]. In contrast, PDK-1 inhibitors may exert antipsychotic and antidepressant properties by GSK-3β deactivation without the typical adverse effects of conventional psychotropics.
AhR Antagonists
The antipsychotic properties of AhR antagonists comprise the proof of concept that excessive AhR activation triggers neuropathology. The following natural and synthetic AhR inhibitors were found therapeutic in severe mental illness, including affective disorders.
Quercetin is a natural flavonoid, a plant pigment found in fruits, and exhibits anti-inflammatory, antioxidant, and anticancer activities. In the CNS, quercetin is a negative allosteric modulator of gamma-aminobutyric A (GABA-A) receptors and lowers gray matter loss [65].
Luteolin, a natural antipsychotic that reduces microglial inflammation, is currently in clinical trials for SCZ (NCT05204407) [66].
Alstonine is an indole alkaloid with antidepressant and antipsychotic properties that does not affect dopaminergic pathways but increases serotonergic transmission [67].
Apigenin is a plant-based remedy extract from Elsholtzia rugulosa used by traditional African psychiatrists to treat mental illnesses. Aside from antagonizing AhR, apigenin exhibits vasorelaxant and antioxidant properties [68].
Synthetic AhR Antagonists
Salicylamide is an analgesic drug that exerts a potent and long-lasting inhibition of AhR-induced Cyp1a enzyme. In preclinical studies, it has shown psychotropic properties [69].
IK-175, structure undisclosed, was shown by preclinical studies to block ligand-stimulated AhR activation of Cyp1a [70]
HBU651 are novel synthetic AhR antagonists developed primarily for cancer but appear to have a favorable profile for neuropsychiatric disorders [71].
Senotherapeutics
It is currently established that severe mental illness is associated with cellular/neuronal senescence, indicating that endogenous or exogenous toxins may play a key role in this pathology [72,73]. For example, viral or bacterial infections induce premature aging in brain cells, which may trigger neuropsychiatric symptomatology [74].
Senotherapeutics are natural or synthetic compounds that delay, prevent, or reverse cellular/neuronal senescence. Senotherapeutics comprise senolytics agents that facilitate the elimination of senescent cells and senomorphic compounds capable of deleting senescence markers, including SASP and Senescence-associated beta-galactosidase (SA-β-gal) [75]. While it has been thought that cellular senescence cannot be reversed in the past, newer studies found that inhibiting 3–phosphoinositide-dependent protein kinase 1 (PDK1) can revert cellular senescence in humans [63].
Senolytic antibiotics belong to a distinct class of agents, which include azithromycin, minocycline, and roxithromycin, and possess neuroprotective, anti-inflammatory, and senolytic properties. For example, it has been known for some time that minocycline may be beneficial for neuropathology, suggesting that senolytics have a place in psychiatry [76].
A senolytic vaccine, recently tested in progeroid mice, may usher in a new era in neuropsychiatry, raising the possibility of vaccination or serum treatment for neuropsychiatric disorders [77]. Another immunological intervention, an antibody-drug conjugate against a membrane senescence marker, was demonstrated to clear senescent, damaged, or infected cells, emphasizing a new therapeutic strategy.
Mitochondrial Transfer and Transplantation
Mitochondria are former bacteria that retain the ability to “talk” with microbes and influence their behavior. The endosymbiotic theory proposes that mitochondria originated from bacteria and archaea and formed intracellular organelles that benefit both partners. Mitochondria generate energy in the form of ATP and counteract the “low energy” symptoms of depression and fatiguing disorders [78].
Cell-free mitochondrial DNA (cf-mtDNA) is a marker of IBD and psychological stress [79,80]. For instance, loneliness and socialization deficit increased cf-mtDNA/TLR9 signaling, an inflammatory pathway. Furthermore, loneliness and isolation-upregulated cf-mtDNA acts as a damage-associated molecular pattern (DAMP), igniting “sterile” inflammation implicated in psychological stress [81,82].
Mitochondrial transplantation experiments started in the 1980s when naked organelles were co-incubated with various cell types to facilitate mitochondrial internalization. Successful mitochondrial transplantation is currently possible on several cell types, including cardiomyocytes [83].
Mitochondrial transplantation for rescuing neurons from apoptosis has been performed successfully in animals and humans [85]. Nevertheless, to the best of our knowledge, it has not been attempted in mental illness.
Under physiological or pathological conditions, intercellular mitochondrial transfer can occur via tunneling nanotubes (TNTs) or extracellular vesicles (EVs).
Non-canonical mitochondrial transfer can occur through cell-cell fusion, synaptosomes, or dendritic networks. Cell-cell fusion can occur in senescent neurons that reenter the cell cycle but cannot complete replication, remaining indefinitely in a fused state.
Besides its role in the intracellular compartment, a growing body of evidence indicates that under physiological or pathological conditions, mitochondria can be secreted into the extracellular space, which plays a vital role in regulating metabolism and immunity [85]
Preclinical research has shown that mitochondrial transplantation decreases LPS-induced depression, emphasizing a possible therapeutic application for PTSD or MDD [86].
Conclusions
Approximately 47% of patients with COVID-19 have Mycoplasma as a comorbidity. Mycoplasma is difficult to identify and a frequent contaminant. Because of these characteristics, if detected in a sample, Mycoplasma would be deemed contamination, rather than a clear sign of pathology.
Biological weapons target physical and mental readiness for combat in enemy troops. Designer bacteria and viruses, unknown to the human immune system could trigger unpredictable responses and cause unknown pathology and chronic illnesses. In other words, the immune system may not recognize the acute phase of illnesses, allowing chronicity to settle in. Cellular senescence can be attenuated by mitochondrial transplantation, senolytic, senomorphic drugs, PDK-1 Inhibitors, and AhR antagonists.
References
- Michael J. Ainscough. Next Generation Bioweapons: Genetic Engineering and BWUS Air Force. Counterproliferation Center. Future Warfare Series No. 14.
- Nicolson GL, Nasralla MY, Haier J, Pomfret J. High frequency of systemic mycoplasmal infections in Gulf War veterans and civilians with Amyotrophic Lateral Sclerosis (ALS). J Clin Neurosci. 2002 Sep;9(5):525-9. [CrossRef]
- Donta, S.T.; Engel, C.C.; Collins, J.F.; Baseman, J.B.; Dever, L.L.; Taylor, T.; Boardman, K.D.; Kazis, L.E.; Martin, S.E.; Horney, R.A.; et al. Benefits and harms of doxycycline treatment for Gulf War veterans’ illnesses: A randomized, double-blind, placebo-controlled trial. Ann. Intern. Med. 2004, 141, 85–94. [Google Scholar] [CrossRef]
- arshis M, Morag B, Mayer M. Mycoplasma cells stimulate in vitro activation of plasminogen by purified tissue-type plasminogen activator. FEMS Microbiol Lett. 1993 Jan 15;106(2):201-4.
- Yavlovich A, Higazi AA, Rottem S. Plasminogen binding and activation by Mycoplasma fermentans. Infect Immun. 2001 Apr;69(4):1977-82. [CrossRef]
- Elizabeth Pennisi,Synthetic Genome Brings New Life to Bacterium.Science328,958-959(2010). [CrossRef]
- Sleator RD. JCVI-syn3.0—A synthetic genome stripped bare! Bioengineered. 2016 Apr 2;7(2):53-6. [CrossRef]
- Hutchison C.A., 3rd, Chuang R.Y., Noskov V.N., Assad-Garcia N., Deerinck T.J., Ellisman M.H., Gill J., Kannan K., Karas B.J., Ma L., et al. Design and synthesis of a minimal bacterial genome. Science. 2016;351:aad6253. [CrossRef]
- Citti C, Dordet-Frisoni E, Nouvel LX, Kuo CH, Baranowski E. Horizontal Gene Transfers in Mycoplasmas (Mollicutes). Curr Issues Mol Biol. 2018;29:3-22. [CrossRef]
- Rosengarten R, Citti C, Glew M, Lischewski A, Droesse M, Much P, Winner F, Brank M, Spergser J. Host-pathogen interactions in mycoplasma pathogenesis: virulence and survival strategies of minimalist prokaryotes. Int J Med Microbiol. 2000 Mar;290(1):15-25. [CrossRef]
- Lai C-C, Wang C-Y, Hsueh P-R. Co-infections among patients with COVID-19: the need for combination therapy with non-anti-SARS-CoV-2 agents? J Microbiol Immunol Infect. 2020;53:505–512. [CrossRef]
- Chaudhry R, Sreenath K, Vinayaraj EV, Sahoo B, Vishnu Narayanan MR, Kiran KVPS, Batra P, Rathor N, Singh S, Mohan A, Bhatnagar S. Mycoplasma pneumoniae co-infection with SARS-CoV-2: A case report. Access Microbiol. 2021 Mar 10;3(3):000212. [CrossRef]
- Arfi Y, Lartigue C, Sirand-Pugnet P, Blanchard A. Beware of Mycoplasma Anti-immunoglobulin Strategies. mBio. 2021 Dec 21;12(6):e0197421. [CrossRef]
- Bransfield RC, Mao C, Greenberg R. Microbes and Mental Illness: Past, Present, and Future. Healthcare (Basel). 2023 Dec 29;12(1):83. [CrossRef]
- Dehhaghi M, Heydari M, Panahi HKS, Lewin SR, Heng B, Brew BJ, Guillemin GJ. The roles of the kynurenine pathway in COVID-19 neuropathogenesis. Infection. 2024 Oct;52(5):2043-2059. [CrossRef]
- Johnson MDL, Younis US, Menghani SV, Addison KJ, Whalen M, Pilon AL, Cress AE, Polverino F, Romanoski CE, Kraft M, Martinez FD, Guerra S, Ledford JG. CC16 Binding to α4β1 Integrin Protects against Mycoplasma pneumoniae Infection. Am J Respir Crit Care Med. 2021 Jun 1;203(11):1410-1418. [CrossRef]
- Zimmermann L, Peterhans E, Frey J. RGD motif of lipoprotein T, involved in adhesion of Mycoplasma conjunctivae to lamb synovial tissue cells. J Bacteriol. 2010 Jul;192(14):3773-9. [CrossRef]
- Vilardi, A., Przyborski, S., Mobbs, C. et al. Current understanding of the interplay between extracellular matrix remodelling and gut permeability in health and disease. Cell Death Discov. 10, 258 (2024). [CrossRef]
- Iyama, K., Zhang, S. & Lo, SC. Effects of Mycoplasmal LAMPs on Receptor Responses to Steroid Hormones in Mammalian Cells. Curr Microbiol 43, 163–169 (2001). [CrossRef]
- Benedetti F, Curreli S, Zella D. Mycoplasmas-Host Interaction: Mechanisms of Inflammation and Association with Cellular Transformation. Microorganisms. 2020 Sep 4;8(9):1351. [CrossRef]
- Seitz-Holland, J., Mulsant, B.H., Reynolds III, C.F. et al. Major depression, physical health and molecular senescence markers abnormalities. Nat. Mental Health 1, 200–209 (2023). [CrossRef]
- Yan B, Liao P, Han Z, Zhao J, Gao H, Liu Y, Chen F, Lei P. Association of aging related genes and immune microenvironment with major depressive disorder. J Affect Disord. 2025 Jan 15;369:706-717. [CrossRef]
- Chen J, Xie X, Lin M, Han H, Wang T, Lei Q, He R. Genes associated with cellular senescence as diagnostic markers of major depressive disorder and their correlations with immune infiltration. Front Psychiatry. 2024 May 31;15:1372386. [CrossRef]
- Diniz, B.S., Reynolds III, C.F., Sibille, E. et al. Major depression and enhanced molecular senescence abnormalities in young and middle-aged adults. Transl Psychiatry 9, 198 (2019). [CrossRef]
- Farooq S, Tunmore J, Wajid Ali M, Ayub M. Suicide, self-harm and suicidal ideation during COVID-19: A systematic review. Psychiatry Res. 2021 Dec;306:114228. [CrossRef]
- Pathirathna, M.L., Nandasena, H.M.R.K.G., Atapattu, A.M.M.P. et al. Impact of the COVID-19 pandemic on suicidal attempts and death rates: a systematic review. BMC Psychiatry 22, 506 (2022). [CrossRef]
- Busl KM, Bleck TP. Treatment of neuroterrorism. Neurotherapeutics. 2012;9(1):139–157. [CrossRef]
- Taubenberger JK, Reid AH, Krafft AE, Bijwaard KE, Fanning TG. Initial genetic characterization of the 1918 “Spanish” influenza virus. Science. 1997 Mar 21;275(5307):1793-6. [CrossRef]
- Becker MM, Graham RL, Donaldson EF, Rockx B, Sims AC, Sheahan T, Pickles RJ, Corti D, Johnston RE, Baric RS, Denison MR. Synthetic recombinant bat SARS-like coronavirus is infectious in cultured cells and in mice. Proc Natl Acad Sci U S A. 2008 Dec 16;105(50):19944-9. [CrossRef]
- Radenkovic D, Chawla S, Pirro M, Sahebkar A, Banach M. Cholesterol in Relation to COVID-19: Should We Care about It? J Clin Med. 2020 Jun 18;9(6):1909. [CrossRef]
- Kočar E, Katz S, Pušnik Ž, Bogovič P, Turel G, Skubic C, Režen T, Strle F, Martins Dos Santos VAP, Mraz M, Moškon M, Rozman D. COVID-19 and cholesterol biosynthesis: Towards innovative decision support systems. iScience. 2023 Aug 31;26(10):107799. [CrossRef]
- Guarnera A, Podda P, Santini E, Paolantonio P, Laghi A. Differential diagnoses of COVID-19 pneumonia: the current challenge for the radiologist-a pictorial essay. Insights Imaging. 2021 Mar 11;12(1):34. PMID: 33704615. [CrossRef]
- Miyashita N, Nakamori Y, Ogata M, Fukuda N, Yamura A, Ishiura Y, Nomura S. Clinical Differences between Community-Acquired Mycoplasma pneumoniae Pneumonia and COVID-19 Pneumonia. J Clin Med. 2022 Feb 12;11(4):964. [CrossRef]
- Waites KB, Talkington DF. Mycoplasma pneumoniae and its role as a human pathogen. Clin Microbiol Rev. 2004 Oct;17(4):697-728, table of contents. [CrossRef]
- Halbedel S, Stülke J. Tools for the genetic analysis of Mycoplasma. Int J Med Microbiol. 2007 Feb;297(1):37-44. [CrossRef]
- Liu Y, Ma Y. Clinical applications of metagenomics next-generation sequencing in infectious diseases. J Zhejiang Univ Sci B. 2024 May 17;25(6):471-484. [CrossRef]
- Fan BE, Lim KGE, Chong VCL, Chan SSW, Ong KH, Kuperan P. COVID-19 and mycoplasma pneumoniae coinfection. Am J Hematol. 2020 Jun;95(6):723-724. Epub 2020 Apr 3. [CrossRef]
- Cheng, Y., Fang, Q. F., & Chen, B. Q. (2024). Clinical Efficacy of Azithromycin in the Treatment of Pediatric Mycoplasma pneumoniae Pneumonia and Its Impact on Platelet Count and D-Dimer Levels. Infection and Drug Resistance, 17, 5195–5202. [CrossRef]
- Liu J, Lu F, Chen Y, Plow E, Qin J. Integrin mediates cell entry of the SARS-CoV-2 virus independent of cellular receptor ACE2. J Biol Chem. 2022 Mar;298(3):101710. [CrossRef]
- Lemańska-Perek A, Krzyżanowska-Gołąb D, Dragan B, Tyszko M, Adamik B. Fibronectin as a Marker of Disease Severity in Critically Ill COVID-19 Patients. Cells. 2022 May 6;11(9):1566. [CrossRef]
- Brogna C, Brogna B, Bisaccia DR, Lauritano F, Marino G, Montano L, Cristoni S, Prisco M, Piscopo M. Could SARS-CoV-2 Have Bacteriophage Behavior or Induce the Activity of Other Bacteriophages? Vaccines (Basel). 2022 Apr 29;10(5):708. [CrossRef]
- Lietha D, Izard T. Roles of Membrane Domains in Integrin-Mediated Cell Adhesion. Int J Mol Sci. 2020 Aug 1;21(15):5531. [CrossRef]
- Campbell ID, Humphries MJ. Integrin structure, activation, and interactions. Cold Spring Harb Perspect Biol. 2011 Mar 1;3(3):a004994. [CrossRef]
- Chastney, M.R., Kaivola, J., Leppänen, VM. et al. The role and regulation of integrins in cell migration and invasion. Nat Rev Mol Cell Biol (2024). [CrossRef]
- Harburger DS, Calderwood DA. Integrin signalling at a glance. J Cell Sci. 2009 Jan 15;122(Pt 2):159-63. Erratum in: J Cell Sci. 2009 May 1;122(Pt 9):1472. [CrossRef]
- Arnaout MA, Goodman SL, Xiong JP. Structure and mechanics of integrin-based cell adhesion. Curr Opin Cell Biol. 2007 Oct;19(5):495-507. Epub 2007 Oct 24. PMID: 17928215. [CrossRef]
- Agnati LF, Guidolin D, Carone C, Dam M, Genedani S, Fuxe K. Understanding neuronal molecular networks builds on neuronal cellular network architecture. Brain Res Rev. 2008 Aug;58(2):379-99. [CrossRef]
- Shay JW, Wright WE. Hayflick, his limit, and cellular ageing. Nat Rev Mol Cell Biol. 2000 Oct;1(1):72-6. [CrossRef] [PubMed]
- Rentscher KE, Carroll JE, Polsky LR, Lamkin DM. Chronic stress increases transcriptomic indicators of biological aging in mouse bone marrow leukocytes. Brain Behav Immun Health. 2022 Apr 12;22:100461. [CrossRef]
- Lorenzo EC, Figueroa JE, Demirci DA, El-Tayyeb F, Huggins BJ, Illindala M, Bartley JM, Haynes L, Diniz BS. Unraveling the association between major depressive disorder and senescent biomarkers in immune cells of older adults: a single-cell phenotypic analysis. Front Aging. 2024 Apr 11;5:1376086. [CrossRef]
- Masaldan, S., Clatworthy, S.A., Gamell, C., Meggyesy, P.M., Rigopoulos, A., Haupt, S., Haupt, Y., Denoyer, D., Adlard, P.A., Bush, A.I., & Cater, M.A. (2017). Iron accumulation in senescent cells is coupled with impaired ferritinophagy and inhibition of ferroptosis. Redox Biology, 14, 100–115.
- Crockett MJ, Clark L, Hauser MD, Robbins TW. Serotonin selectively influences moral judgment and behavior through effects on harm aversion. Proc Natl Acad Sci U S A. 2010 Oct 5;107(40):17433-8. [CrossRef]
- Kunieda T, Minamino T, Nishi J, Tateno K, Oyama T, Katsuno T, Miyauchi H, Orimo M, Okada S, Takamura M, Nagai T, Kaneko S, Komuro I. Angiotensin II induces premature senescence of vascular smooth muscle cells and accelerates the development of atherosclerosis via a p21-dependent pathway. Circulation. 2006 Aug 29;114(9):953-60. [CrossRef]
- Logunov DY, Scheblyakov DV, Zubkova OV, Shmarov MM, Rakovskaya IV, Gurova KV, Tararova ND, Burdelya LG, Naroditsky BS, Ginzburg AL, Gudkov AV. Mycoplasma infection suppresses p53, activates NF-kappaB and cooperates with oncogenic Ras in rodent fibroblast transformation. Oncogene. 2008 Jul 31;27(33):4521-31. [CrossRef]
- Schmitt CA, Tchkonia T, Niedernhofer LJ, Robbins PD, Kirkland JL, Lee S. COVID-19 and cellular senescence. Nat Rev Immunol. 2023 Apr;23(4):251-263. Epub 2022 Oct 5. [CrossRef]
- Lefèvre C, Auclair M, Boccara F, Bastard JP, Capeau J, Vigouroux C, Caron-Debarle M. Premature senescence of vascular cells is induced by HIV protease inhibitors: implication of prelamin A and reversion by statin. Arterioscler Thromb Vasc Biol. 2010 Dec;30(12):2611-20. [CrossRef]
- Collins KL, Younis US, Tanyaratsrisakul S, Polt R, Hay M, Mansour HM, Ledford JG. Angiotensin-(1-7) Peptide Hormone Reduces Inflammation and Pathogen Burden during Mycoplasma pneumoniae Infection in Mice. Pharmaceutics. 2021 Oct 4;13(10):1614. [CrossRef]
- Servant G, Escher E, Guillemette G. The angiotensin II binding site on Mycoplasma hyorhynis is structurally distinct from mammalian AT1 and AT2 receptors. Regul Pept. 1998 Jan 2;73(1):35-41. [CrossRef] [PubMed]
- Gong S, Deng F. Renin-angiotensin system: The underlying mechanisms and promising therapeutical target for depression and anxiety. Front Immunol. 2023 Jan 24;13:1053136. [CrossRef]
- Herbert KE, Mistry Y, Hastings R, Poolman T, Niklason L, Williams B. Angiotensin II-mediated oxidative DNA damage accelerates cellular senescence in cultured human vascular smooth muscle cells via telomere-dependent and independent pathways. Circ Res. 2008 Feb 1;102(2):201-8. [CrossRef]
- Sinha JK, Ghosh S, Raghunath M. Progeria: a rare genetic premature ageing disorder. Indian J Med Res. 2014 May;139(5):667-74. PMID: 25027075; PMCID: PMC4140030. [PubMed]
- Harhouri K, Frankel D, Bartoli C, Roll P, De Sandre-Giovannoli A, Lévy N. An overview of treatment strategies for Hutchinson-Gilford Progeria syndrome. Nucleus. 2018 Jan 1;9(1):246-257. [CrossRef]
- S. An, S. Cho, J. Kang, S. Lee, H. Kim, D. Min, E. Son, K. Cho, Inhibition of 3-phosphoinositide–dependent protein kinase 1 (PDK1) can revert cellular senescence in human dermal fibroblasts, Proc. Natl. Acad. Sci. U.S.A.117 (49) 31535-31546. (2020). [CrossRef]
- Sonnenburg, Erica Dutil et al. The Phosphoinositide-dependent Kinase, PDK-1, Phosphorylates Conventional Protein Kinase C Isozymes by a Mechanism That Is Independent of Phosphoinositide 3-Kinase. Journal of Biological Chemistry, Volume 276, Issue 48, 45289–45297.
- Moghbelinejad S, Alizadeh S, Mohammadi G, Khodabandehloo F, Rashvand Z, Najafipour R, Nassiri-Asl M. The effects of quercetin on the gene expression of the GABAA receptor α5 subunit gene in a mouse model of kainic acid-induced seizure. J Physiol Sci. 2017 Mar;67(2):339-343. [CrossRef]
- Saleem A, Qurat-Ul-Ain, Akhtar MF. Alternative Therapy of Psychosis: Potential Phytochemicals and Drug Targets in the Management of Schizophrenia. Front Pharmacol. 2022 May 17;13:895668. [CrossRef]
- Elisabetsky E, Costa-Campos L. The alkaloid alstonine: a review of its pharmacological properties. Evid Based Complement Alternat Med. 2006 Mar;3(1):39-48. [CrossRef]
- Olayinka JN, Akawa OB, Ogbu EK, Eduviere AT, Ozolua RI, Soliman M. Apigenin attenuates depressive-like behavior via modulating monoamine oxidase A enzyme activity in chronically stressed mice. Curr Res Pharmacol Drug Discov. 2023 Jul 11;5:100161. [CrossRef]
- Żmudzka E, Lustyk K, Głuch-Lutwin M, Wolak M, Jaśkowska J, Kołaczkowski M, Sapa J, Pytka K. Novel Multimodal Salicylamide Derivative with Antidepressant-like, Anxiolytic-like, Antipsychotic-like, and Anti-Amnesic Activity in Mice. Pharmaceuticals (Basel). 2023 Jan 24;16(2):175. [CrossRef]
- McGovern K, Castro AC, Cavanaugh J, Coma S, Walsh M, Tchaicha J, Syed S, Natarajan P, Manfredi M, Zhang XM, Ecsedy J. Discovery and Characterization of a Novel Aryl Hydrocarbon Receptor Inhibitor, IK-175, and Its Inhibitory Activity on Tumor Immune -7163.MCT-21-0984.
- Kang, S.; Lee, A.G.; Im, S.; Oh, S.J.; Yoon, H.J.; Park, J.H.; Pak, Y.K. A Novel Aryl Hydrocarbon Receptor Antagonist HBU651 Ameliorates Peripheral and Hypothalamic Inflammation in High-Fat Diet-Induced Obese Mice. Int. J. Mol. Sci. 2022, 23, 14871. [Google Scholar] [CrossRef]
- Yan L., Li H., Qian Y., Zhang J., Cong S., Zhang X., Wu L., Wang Y., Wang M., Yu T. Transcutaneous vagus nerve stimulation: A new strategy for Alzheimer’s disease intervention through the brain-gut-microbiota axis? Front. Aging Neurosci. 2024;16:1334887. [CrossRef]
- Karpiński P., Żebrowska-Różańska P., Kujawa D., Łaczmański Ł., Samochowiec J., Jabłoński M., Plichta P., Piotrowski P., Bielawski T., Misiak B. Gut microbiota alterations in schizophrenia might be related to stress exposure: Findings from the machine learning analysis. Psychoneuroendocrinology. 2023;155:106335. [CrossRef]
- Seeman, M.V. Subjective Overview of Accelerated Aging in Schizophrenia. Int. J. Environ. Res. Public Health 2023, 20, 737. [Google Scholar] [CrossRef] [PubMed]
- Kozato N., Mishra M., Firdosi M. New-onset psychosis due to COVID-19. BMJ Case Rep. 2021;14:e242538. [CrossRef]
- Solis G.M., Kardakaris R., Valentine E.R., Bar-Peled L., Chen A.L., Blewett M.M., McCormick M.A., Williamson J.R., Kennedy B., Cravatt B.F., et al. Translation attenuation by minocycline enhances longevity and proteostasis in old post-stress-responsive organisms. eLife. 2018;7:e40314. [CrossRef]
- Suda, M., Shimizu, I., Katsuumi, G. et al. Senolytic vaccination improves normal and pathological age-related phenotypes and increases lifespan in progeroid mice. Nat Aging 1, 1117–1126 (2021). [CrossRef]
- Liesa M, Shirihai OS. Mitochondrial dynamics in the regulation of nutrient utilization and energy expenditure. Cell Metab. 2013 Apr 2;17(4):491-506. [CrossRef]
- Trumpff C, Michelson J, Lagranha CJ, Taleon V, Karan KR, Sturm G, Lindqvist D, Fernström J, Moser D, Kaufman BA, Picard M. Stress and circulating cell-free mitochondrial DNA: A systematic review of human studies, physiological considerations, and technical recommendations. Mitochondrion. 2021 Jul;59:225-245. [CrossRef]
- Boyapati RK, Dorward DA, Tamborska A, Kalla R, Ventham NT, Doherty MK, Whitfield PD, Gray M, Loane J, Rossi AG, Satsangi J, Ho GT. Mitochondrial DNA Is a Pro-Inflammatory Damage-Associated Molecular Pattern Released During Active IBD. Inflamm Bowel Dis. 2018 Sep 15;24(10):2113-2122. [CrossRef]
- Konaka H, Kato Y, Hirano T, Tsujimoto K, Park J, Koba T, Aoki W, Matsuzaki Y, Taki M, Koyama S, Itotagawa E, Jo T, Hirayama T, Kawai T, Ishii KJ, Ueda M, Yamaguchi S, Akira S, Morita T, Maeda Y, Nishide M, Nishida S, Shima Y, Narazaki M, Takamatsu H, Kumanogoh A. Secretion of mitochondrial DNA via exosomes promotes inflammation in Behçet’s syndrome. EMBO J. 2023 Oct 16;42(20):e112573. [CrossRef]
- Van Bogart K, Engeland CG, Sliwinski MJ, Harrington KD, Knight EL, Zhaoyang R, Scott SB, Graham-Engeland JE. The Association Between Loneliness and Inflammation: Findings From an Older Adult Sample. Front Behav Neurosci. 2022 Jan 11;15:801746. [CrossRef]
- Ali Pour P, Hosseinian S, Kheradvar A. Mitochondrial transplantation in cardiomyocytes: foundation, methods, and outcomes. Am J Physiol Cell Physiol. 2021 Sep 1;321(3):C489-C503. Epub 2021 Jun 30. PMID: 34191626. [CrossRef]
- Fairley LH, Grimm A, Eckert A. Mitochondria Transfer in Brain Injury and Disease. Cells. 2022 Nov 14;11(22):3603. [CrossRef]
- Stefano GB, Büttiker P, Weissenberger S, Esch T, Anders M, Raboch J, Kream RM, Ptacek R. Independent and sensory human mitochondrial functions reflecting symbiotic evolution. Front Cell Infect Microbiol. 2023 Jun 14;13:1130197. [CrossRef]
- Kmita H, Pinna G, Lushchak VI. Potential oxidative stress related targets of mitochondria-focused therapy of PTSD. Front Physiol. 2023 Nov 13;14:1266575. [CrossRef]
Figure 2.
The vertical and horizontal RAS branches. The ACE-2/SARS-CoV-2 binding disables the horizontal (protective) branch, deleting NO and upregulating the toxic peroxynitrate. As angiotensin-converting 2 enzyme (ACE-2) is “occupied” by the virus, Angiotensin II (ANG II) levels increase, inducing premature cellular senescence.
Figure 2.
The vertical and horizontal RAS branches. The ACE-2/SARS-CoV-2 binding disables the horizontal (protective) branch, deleting NO and upregulating the toxic peroxynitrate. As angiotensin-converting 2 enzyme (ACE-2) is “occupied” by the virus, Angiotensin II (ANG II) levels increase, inducing premature cellular senescence.
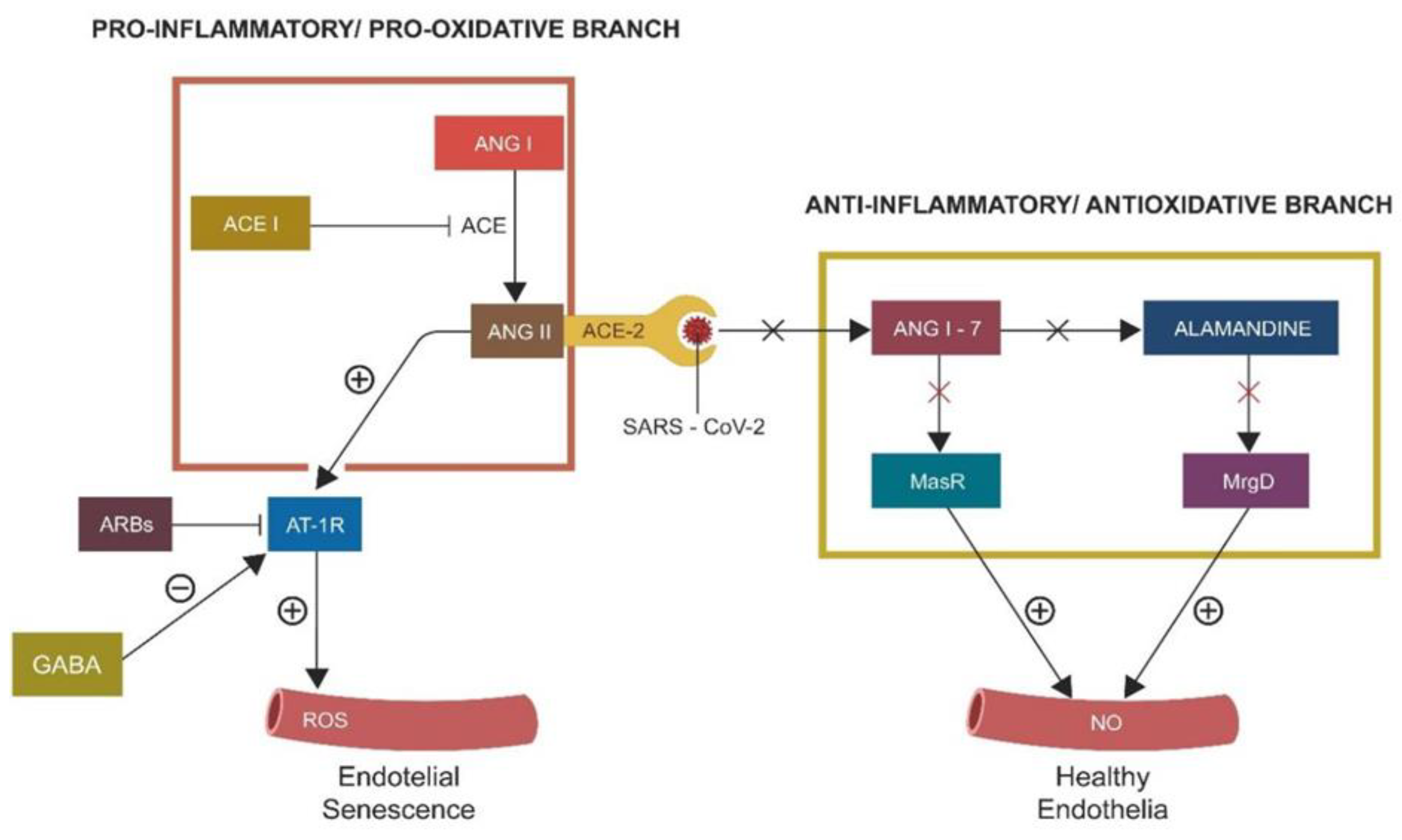
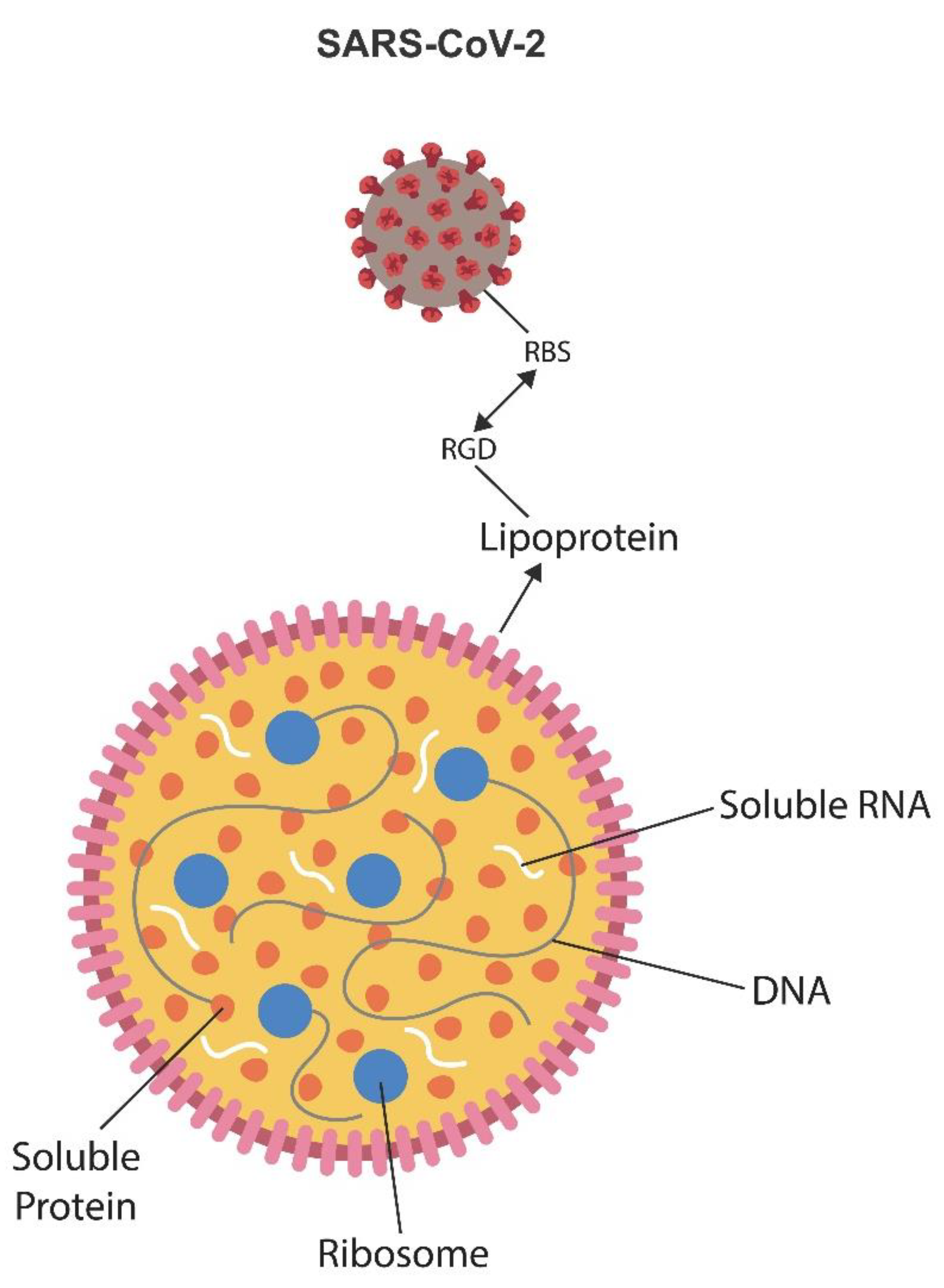
Figure 3.
Mycoplasma lacks a cell wall but contains a membrane comprised of lipoproteins and sterols. This bacterium resembles a liposome or lipid nanoparticle with the SARS-CoV-2 virus as a cargo. SARS-CoV-2 and Mycoplasma connect via the integrin (RGD) motif that attaches on the viral side to the viral receptor-binding site (RBS) and to lipoprotein on the Mycoplasma site. Major Mycoplasma components, including RNA, DNA, ribosomes, and proteins, are shown. The Mycoplasma/SARS-CoV-2 complex attaches to host ECM proteins via integrin αVβ3, which the virus exploits to induce senescence. Anti-integrin drugs may dissociate the virus from Mycoplasma, interfering with the symbiosis.
Figure 3.
Mycoplasma lacks a cell wall but contains a membrane comprised of lipoproteins and sterols. This bacterium resembles a liposome or lipid nanoparticle with the SARS-CoV-2 virus as a cargo. SARS-CoV-2 and Mycoplasma connect via the integrin (RGD) motif that attaches on the viral side to the viral receptor-binding site (RBS) and to lipoprotein on the Mycoplasma site. Major Mycoplasma components, including RNA, DNA, ribosomes, and proteins, are shown. The Mycoplasma/SARS-CoV-2 complex attaches to host ECM proteins via integrin αVβ3, which the virus exploits to induce senescence. Anti-integrin drugs may dissociate the virus from Mycoplasma, interfering with the symbiosis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



