
Submitted:
20 December 2024
Posted:
20 December 2024
You are already at the latest version
Abstract
The neurobiology of sex differences in response to chronic stress is a topic that has been extensively discussed in the scientific literature, but remains poorly understood. In our research, we found that chronic unpredictable mild stress (CUMS) experienced during adolescence induces different behavioural patterns in adult males и females. Similarly, immunohistochemical analysis of the CA1 field of the dorsal and ventral hippocampus revealed no changes in quantitative and morphological changes in astrocytes at long-term period after CUMS. Real-time PCR analysis showed no increase in SIGMAR1 expression level relative to individual housekeeping genes after CUMS. Analysis of mouse cerebral cortex homogenates showed that IL-1β levels decreased after stress modelling only in males. However, Sigma R1 levels were significantly higher in the CUMS group than in the controls in both sexes. It can be concluded that biological sex and age do indeed influence the response to CUMS, although not in all cases. There is a need for further research in order to understand the effects of chronic stress on males and females. This is of crucial importance because men and women are exposed to different risks in relation to stress and mental health.
Keywords:
Brain sexual dimorphism
; chronic unpredictable mild stress
; SigmaR1
; behavior
; immunohistochemistry
1. Introduction
Psycho-emotional tension, known as stress, is considered by modern biomedical science to be a catalyst for increasing the frequency and severity of various pathologies. Particular attention is now being paid to the problems of stress in childhood and adolescence, when the foundations for proper adaptation and handling of stressors are being laid in the family and educational contexts. Stress is a complex neuroimmunoendocrine response involving many organs and systems. The main players in this process are the immune, nervous and endocrine systems.
Research has shown that chronic unpredictable mild stress (CUMS) affects brain structure [1] and function [2]. Comprehensive approaches are needed to understand how stress experienced early in life can reprogramme the brain’s cognitive and emotional networks, affect neurodevelopment, and increase the risk of psychopathology. These approaches must take into account the multifaceted nature of stress and its impact on nervous system development.
The behavioural changes induced by CUMS are associated with dopaminergic hyperfunction and serotonergic hypofunction [3]. Interestingly, the stress experienced in childhood leads to different behavioural strategies in male and female individuals. This may be partly due to the different effects of sex steroids on the hypothalamic-pituitary-adrenal (HPA) axis activity through their effects on gene expression, protein synthesis, and cellular excitability via androgen and estrogen receptors [4].
Furthermore, it has been shown that during puberty, sex steroid hormones including progesterone, androgens and estrogens can further modulate brain development during adolescence in ways that have long-term consequences for behavior in adulthood [5]. Because sex steroids are involved in the regulation of behavioural and neuroendocrine responses to stress [6], they are able to modulate dopamine levels, with testosterone, for example, having a positive effect on dopamine levels and estrogens having the opposite effect. It is known that dopamine levels increase under acute stress, and under chronic stress, low dopamine levels lead to maladaptive stress-associated responses [7]
Indeed, there is a marked sexual dimorphism in the function and structure of the dopaminergic system between males and females in normals [8]. After CUMS, there is an increase in the levels of dopamine, its metabolites, and tyrosine hydroxylase (TH) protein in the nucleus accumbens (NAc) [9]. Tyrosine hydroxylase plays a key role in the synthesis of catecholamines such as dopamine, norepinephrine, and adrenaline [8]. At the same time, unpredictable mild stress leads to decreased levels of serotonin and tryptophan hydroxylase levels in the hippocampus and prefrontal cortex [9]. Changes in the prefrontal cortex affect higher cognitive functions such as decision-making, attention, and social behaviour, while changes in the hippocampus affect emotional status and memory. Thus, stress has opposing effects on different neurotransmitter systems in the brain. SIGMAR1 knockout mice [10] show dysfunction of this system and also show signs of behavioural depression.
Unpredictable mild stress is known to increase in inflammatory markers such as IL-6, IL-1β, TNF-α, and activate peripheral inflammatory pathways [11,12]. Emotional responses to stress may differ according to gender [13]. For example, women who react negatively to everyday stress have higher levels of inflammatory markers. Therefore, it is important to consider biological sex when investigating the mechanisms of exposure to unpredictable mild stress.
Furthermore, exposure to stress via cytokine signalling reliably leads to microglial responses in the hippocampus and prefrontal cortical regions [14],which in turn may lead to structural and functional changes in the brain following CUMS suffered at an early age.
However, mechanisms involving various types of proteins in the nervous system have been identified that contribute to the suppression of neuroinflammatory responses and the polarisation of microglia into the M1 type. For example, activation of the opioid receptor subtype triggers antidepressant, anti-inflammatory, and neuroprotective responses. This process is accomplished through the regulation of mitochondrial function and the interaction of the endoplasmic reticulum with mitochondria [15,16].
Alterations in the mitochondrial axis are increasingly considered to underlie the pathogenesis of several diseases. Mitochondria as a scaffold are responsible not only for mitochondrial quality control, but also for various types of signalling, including neuroinflammation and neurodegeneration [17]. A number of protein components of the mitochondrial outer membrane have been identified that affect mitochondrial dynamics and distribution by modulating fission-fusion mediators. These molecules include the proteins mitofusin 1 (MFN1), MARCH5 and SIGMAR1. Mfn1 is an outer mitochondrial membrane protein required for mitochondrial fusion. Mitochondrial ubiquitin ligase, also known as MARCH5, is an E3 ubiquitin ligase. It is also one of two subtypes of sigma receptors, a chaperone protein in the endoplasmic reticulum (ER) that modulates calcium signalling through the IP3 receptor. [18].
However, very little is known about how mild, unpredictable stress during adolescence may affect mitochondrial signalling, inflammatory processes and the structural and functional features of the nervous system, in general, in female and male animals.
2. Results
2.1. Behavioural Phenotyping
2.1.1. Increased Anxiety in Females After Stressful Experiences
Behavioural phenotyping showed that CUMS (Figure 1) females exhibited higher levels of anxiety in the open field test compared to control females and males in both groups, and spent significantly less time in the central area of the arena than the control group (p=0.068) (Figure2. (A)). The increased post-stress anxiety in females is also evidenced by the reduced number of entries into the central zone of the open field arena (Figure2. (B)) compared to control and CUMS males. However, there was no significant increase in the duration of grooming acts, which is also a marker of anxiety, and there was no significant increase in this parameter (Figure2. (C)).
2.1.2. Decreased Locomotor Activity After Stress Is Characteristic of Both Sexes
In addition, the level of locomotor activity was significantly different between CUMS and control groups. Distance travelled (Figure 2. (D)) was reduced both in CUMS females (p=0.0088) compared to intact controls and in CUMS males (p=0.0026) compared to males that did not experience CUMS during early ontogeny. CUMS females showed a significant increase in resting time (Figure 2. (E)) compared to both intact females (p=0.0027) and males. Simultaneously, there was an effect of stress on the reduction of fast movements (Figure 2. (F)) in males (p=0.0410) and females (p=0.0081) in the experimental groups, as well as a reduction in the average movement speed (Figure 2. (G)) across the Open Field arena in females (p=0.0088) and males after CUMS (p=0.0026).
2.1.3. Differences in Exploratory Activity in Males and Females
Analysis of exploratory activity in the Open Field test showed a statistically significant reduction in the number of rearing acts (Figure 2. (H)) in CUMS females (p = 0.0403) in the remote period after stress modelling. In contrast, CUMS males showed an increase in exploratory activity and the number of rearing acts was significantly higher in experimental animals (p=0.0107) than in the control group. And the total number of sniffing acts (Figure 2. (I)) did not differ between groups. At the same time, there is a tendency for this index of exploratory activity to increase in CUMS males compared to both control males and control females.
2.1.4. Decreased Hippocampus-Dependent Working Memory During the Developing of the Conditioned Passive Avoidance Reflex in Females and Decreased Hippocampus-Dependent Object Memory in Males After Stress
The study of memory in the passive avoidance test (Figure 3. (A)) revealed a decrease in working memory in the female CUMS group (p=0.0143), but no significant differences in males in the remote period after the simulation of stress. Percentage analysis of animal learning in the Novel Object Recognition test (Figure3. (D)) revealed that chronic stress experienced early in life caused a significant decrease in interaction time with a novel object (p=0.0134) in CUMS males, but not in females.
2.1.5. Increased Social Activity in Females After Stress
Social activity (Figure3. (B)) among females increased dramatically in the CUMS group, as the interaction time with an unfamiliar animal in the first session of the three-chamber Crowley test was significantly higher than that of the control group (p=0.0452). Moreover, females from the experimental group showed a desire for social novelty (Figure3. (C)), which was expressed by favoring an unfamiliar animal over a previously familiar one (p=0.0326). At the same time, there were no significant differences among males in this criterion. However, the group of control males showed a tendency to increase social activity compared to the experimental group.
2.1.6. CUMS Does Not Lead to the Development of Despair Behaviour in the Remote Period
When analyzing the development of depression-like behaviour in the Porsolt forced swim test as a result of CUMS, it was found (Figure 4. (A)) that the time to first immobilization was significantly longer for the CUMS female group than for control (p=0.0436). CUMS males had significantly less active swimming time (Figure4. (D)) (p=0.0026) and significantly longer climbing time (Figure 4. (C)) (p=0.0171) compared to intact males and CUMS females. At the same time, no significant differences in immobility time were found in any group (Figure 4. (B)).
2.1.7. Behavioural tests demonstrated fairly significant behavioural differences between males and females, particularly in learning ability, social behaviour, and the emotional component of behaviour. Memory and reflex development is mostly performed by the dorsal hippocampus, whereas social activity in mice is primarily mediated by ventral hippocampal CA1 [19] and CA2 regions [20] as well as their projections to other brain regions such as the contiguous nucleus and medial prefrontal cortex [21]. Accordingly, we decided to perform a general morphometric analysis of CA1 of the dorsal and ventral hippocampus.
2.2. Immunohistochemical Study
2.2.1. MAP2 Intensity in Entorhinal Cortex and Hippocampus Does Not Correlate with Behavioural Phenotype
MAP2 immunostaining is effective in detecting and assessing brain damage in cases of acute ischaemia [22] and synaptic rearrangement [23]. Based on the results of MAP2 fluorescence intensity analysis, no statistically significant differences were found in the CA1 field of the dorsal and ventral hippocampus in the entorhinal cortex (Figure 5). However, the variability in the entorhinal cortex is higher than in the hippocampus. It appears that the effects observed in the behavioural phenotype are not related to the rearrangement of the cytoskeleton of dendritic outgrowths.
2.2.2. CUMS in Infantile Age Does Not Induce Changes in Quantitative and Morphologic Changes in Astrocytes in CA1 Dorsal and Ventral Hippocampal Regions
Microglial and astrocytic cells in the CA1 field of the dorsal (Figure 6) and ventral (Figure 7) hippocampus were analysed quantitatively. Although the number of glial cells in the CA1 stratum radiatum is significantly higher in the ventral hippocampus, the number of astroglial cells and microglia per mm² does not change in the different experimental groups. The distribution of microglia within the hippocampal layers is uniform in all experimental groups. No sexual dimorphism was found.
2.3. Enzyme Immunoassay of Cortex Homogenates
2.3.1. CUMS Induces a Decrease in Male IL-1 Beta Concentrations in Cortical Homogenate
IL-1beta levels decreased after stress modelling only in males. No statistically significant differences were found when analysing IL-10 and TNFα (Figure 8).
2.3.2. CUMS Causes an Increase in Hippocampal Sigma R1 Protein Levels Regardless of Biological Sex
When mouse cortical homogenates were analysed, females in the CUMS group had significantly higher levels of Sigma R1 than controls (P = 0.016). Sigma R1 levels were also high in the male group exposed to CUMS, with significant differences from the control group (P = 0.004). Biological sex did not influence these dynamics, but it is worth noting that males have more significant differences.
2.4. Real-Time PCR of Hippocampal Homogenates
2.4.1. CUMS Does Not Induce Changes in Cortical SIGMAR1 Gene Expression Regardless of Biological Sex
Experiments did not show statistically significant changes of gene expression of SIGMAR1 gene relative to geometric mean of all used house-keeping genes (ActB, B2m, Gapdh, Gusb, Pum1, Rpl1) (Figure 9 A).
The experiments showed a statistically significant difference: 1) between a control group of males and a group of males exposed to mild unpredictable stress (11% decrease in Sigmar1 expression, p = 0.0483) when Actb was used as a housekeeping gene; 2) between a control group of males and a group of males exposed to mild unpredictable stress (35% decrease in Sigmar1 expression, p = 0.0453) when B2m was used as the housekeeping gene; 3) between a group of males exposed to mild unpredictable stress and a group of females exposed to mild unpredictable stress (129% increase in SIGMAR1 expression, p = 0.0197) when Pum1 was used as the housekeeping gene. However, no statistically significant differences were found when using Gapdh, Rpl19 and Gusb as housekeeping genes individually (Figure 9B).
Also, there were not found any significant difference of IL6 gene expression changes both relative to geometric mean of all used housekeeping genes (ActB, B2m, Gapdh, Gusb, Pum1, Rpl1) (Figure 9C) and relative to individual ones (not presented here).
3. Discussion
Functional and structural brain characteristics associated with biological sex and age may influence responses to stress-associated events in early life, but the neural networks involved are not fully understood. However, the existence of these differences is critical for both fundamental science and medicine, as men and women are known to be at different risk for psychopathology [24].
Indeed in our study we found a number of features, some of which correlated with biological sex. For example, only females with CUMS showed increased anxiety. This may be related to the sustained activation of the HPA axis caused by chronic stress and hormonal dysregulation triggered by the fact that during puberty the adrenal volume increases in females more than in males [25]. In addition, females showed more significantly increased social activity after stress, as well as decreased exploratory activity and impaired working memory in the remote period after CUMS compared to males. At the same time, decreased motor activity in the remote period after CUMS is characteristic of both sexes. These responses may be due to differential activation of the brain’s dopaminergic and serotonergic systems. For example, the dopaminergic system has been shown to be more sensitive to early stressors in men, increasing the likelihood of developing conditions such as attention deficit hyperactivity disorder (ADHD) and schizophrenia. Conversely, females may show more pronounced changes in serotonergic pathways, which could potentially increase the risk of depression and anxiety disorders [26]. In addition, the brain and neuroendocrine system are sensitive to adrenal hormones during puberty [27], which may be critical in situations of chronic stress.
Various neural structures have been shown to be involved in the maintenance of social memory, including the ventral regions of the hippocampus [28], the medial prefrontal cortex (mPFC), the anterior cingulate cortex (ACC) [29], entorhinal cortical projections [30]. For example, the prefrontal cortex [31] and the anterior cingulate cortex [32], are cortical areas that have been implicated in anxiety anxiety. One of the aims of this study was to elucidate which biological targets are responsible for the divergent responses of males and females in the remote period after chronic unpredictable mild stress. To this end, the hippocampus and cortex were examined separately. From a morphological perspective, since females show increased social engagement and anxiety, accompanied by deficits in memory and learning, it was imperative to first examine both the ventral and dorsal hippocampus. In addition, studying the expression of genes associated with inflammation (specifically IL-6) and SIGMAR1 expression was identified as a promising objective, given their potential as targets for neurotropic and antidepressant action. It has been demonstrated that S1R agonists have been shown to facilitate neuronal activity in the hippocampus, thereby replicating the influx of intracellular calcium that is necessary for neuronal excitability and signalling [33]. It was expected that in males, given their increased exploratory activity, there would be notable changes in gene expression and morphology of the dorsal hippocampus. Similarly, changes in the ventral hippocampus were expected in females due to their increased social activity and anxiety. However, no changes were observed. The results did not show an increase in interleukin 6 (IL-6) expression in the hippocampus. However, some studies have reported an increase in IL-6 levels in various brain regions [34], including the hippocampus. In addition, no morphological correlates of memory decline and increased anxiety have been identified in females based on the number and shape of microglial and astrocytic cells in the hippocampus.
Like [35], our model did not show any change in the expression of the SIGMAR1 gene in the hippocampus. It is important to say that the housekeeping genes used to assess gene expression may not have had sufficient stability in expression for this study model of mild unpredictable stress, as proposed in some research papers [36] [37], the Actb and B2M genes are not stable, as demonstrated in this model. After normalisation for the aforementioned genes, statistically significant differences were observed in SigmaR1 in response to stress modelling. The absence of notable discrepancies in cell morphology, MAP2 staining intensity, and SigmaR1 expression does not support the claim of non-participation in the stress response. This is because SigmaR1 has the ability to affect cell signalling through changes in intracellular localisation from MAM [38], calcium signalling [39], and ion channels [40]. Therefore, future studies should investigate the intracellular localisation and interaction with other ion channels. The post-stress phenotype, which is expressed equally in females and males, is characterised by a decrease in locomotor activity. However, studies [41,42] have shown that mice lacking the Sigma-1 receptor exhibit locomotor deficits, muscle weakness, and loss of motor neurons [43].
In the cortical study, we analyzed the intensity of MAP2 immunostaining in different groups in the entorhinal cortex. Map2 intensity is typically used in the study of ischaemia [44] but may also be an early marker of neuronal damage. No statistically significant results were obtained; however, this parameter does not appear to be as stable as it is in the hippocampal CA1 region, and there is a trend towards a decrease in the intensity of immunostaining following CUMS modelling.
For cortex analysis, we examined protein levels measured by ELISA rather than gene expression. In our model, a statistically significant increase in SigmaR1 was found in brain homogenates after CUMS modelling, independent of biological sex, but to a greater extent in males.
In our model of CUMS, we found simultaneous decrease in motor activity and an increase in SigmaR1 protein levels. However, the analysis of pro-inflammatory cytokines (IL-1β, TNF-alpha) and anti-inflammatory cytokines (IL-10) in cortical homogenates didn’t show any statistical differences, except for IL-1β. For this interleukin, a statistically significant decrease was found among in a group of males after CUMS. IL-1β is involved in the process of stress-induced neuroinflammation, which can lead to anxiety-like behaviour [47]. In this work, we didn’t find a decrease in motor activity in males compared to females but we did find an increase in cognitive activity, which may be caused by a decreased level of this inflammatory cytokine in the male brain. It is well-known that elevated levels of IL-1β in the hippocampus correlate with disruption of LTP [48] and inhibition of GABA(A) receptor function, which can critically affect the learning process. It is possible that SigmaR1 plays a neuroprotective role here, with a small degree of sexual dimorphism that can be detected by comparing control groups. There is evidence from studies on the neuroprotective role of ligands of sigma1 receptors in an in vitro model [49], in a model of motor neuron degeneration. Thus, further investigations should focus on spatial transcriptomics of SIGMAR1, as different parts of the brain may respond differently to stress modelling. As there is a strong gender difference in response in the long-term after CUMS modelling, biological sex should be taken into account.
4. Materials and Methods
In vivo experiments were carried out in the SPF vivarium of the Central Research Laboratory of PRMU IFM. Females (F) (n=24) and males (M) (n=24) of the C57BL/6 strain were used in the study. Stress modelling was started on the 17th postnatal day. Animals maintained in individually ventilated cages at 20 ± 2°C and 60% humidity, with a 12 h/12 h light/dark cycle and constant access to food and water. The maintenance and care of laboratory rodents was approved by the Bioethics Committee “PRMU” of the Ministry of Health of the Russian Federation. The care and treatment of the animals was in accordance with the “Guide for the Care and Use of Laboratory Animals” (ILAR publication, 1996, National Academy Press) and “Guidelines for the maintenance and care of laboratory animals” (GOST 33216-2014).
The long-term effects of CUMS were studied when the animals reached adulthood at 68-74 days of postnatal development. Behavioural tests were performed according to the scheme (Figure 1. Schematic representation of the CUMS model and behavioural test battery, Table 1. Experimental stressors) and were ordered from the least stressful to the most stressful tests. Subsequently, the animals were sacrificed and brain tissues were collected, specifically cortex and hippocampus for morphological studies, hippocampus for gene expression, and cortex for quantitative protein analysis.
CUMS modelling. The experimental scheme is shown in Figure 1. CUMS modelling was performed from days 17 to 30 of mouse postnatal development. The detailed description of the stressors is presented in Table 1. On day 31 of postnatal development, mice were separated from their mothers and placed in new cages with individuals of the same sex and age. The long-term effects of СUMS were tested from days 68 to 74 of postnatal development, using behavioural tests as described in Figure 1.
Behavioural tests. The following tests have been used for the description of the behavioural phenotype of animals after CUMS:
Open Field Test. The activity of males and females exposed to CUMS was accused in the “Open Field” test. During the test, indicators of anxiety, locomotor activity, and exploratory activity were registered. The test was conducted in bright light on a 45x45x40 cm square Open Field arena (Panlab, Spain). The animal explored the arena for 5 min, which was divided into two zones (center and periphery). Animal movements were recorded using a video recording system and software Smart Video Tracking System v. 3.0 (Panlab, Spain), as well as behavioural acts such as grooming, freezing, rearing and sniffing.
The study of social behaviour in the Crowley test was conducted in dim light in the equipment, which consisted of a box measuring 60x42x22 cm, divided into three chambers by transparent partitions with doors (Panlab, Spain). The animal was allowed to move freely between compartments during both sessions. Cylinders containing unfamiliar animals of the same strain and sex were placed in the outermost chambers. The first session began after the habituation. At this stage, social activity was assessed. For this purpose, an unfamiliar animal was placed in the left cylinder while the right cylinder remained empty. In the next session, to assess social memory, another unfamiliar animal was placed in the right cylinder, while the animal in the left cylinder was considered already familiar. The test phases lasted 5 minutes each, with a 10-minute break between phases. Transitions of the experimental animal between chambers were recorded by camera and analyzed using Smart Video Tracking System software v. 3.0 (Panlab, Spain). The time spent by the animal interacting with the animals in the cylinders was recorded.
The Novel Object Recognition test was performed in the “Open Field” equipment (Panlab, Spain) in low light. After the habituation phase, the animal was placed in the equipment where mice were free to explore two identical objects for 7 min. After 1 hour in the next session, one of the objects was replaced by a new one that differed in shape and colour. The animal’s activity was recorded by camera and analysed using Smart Video Tracking System software v. 3.0 (Panlab, Spain). The total time spent with each object was recorded.
Passive avoidance (PA) test. The PA chamber (Panlab, Spain) consisted of brightly lit and dark compartments with a floor of metal bars separated by an automatic door. During the training phase, the animal was placed in the brightly lit compartment, then the animal moved to the dark compartment where it received an electrical stimulus through the metal bars of the floor (50 Hz, 1.5 mA for 1 sec.). After 24 h in the test phase, the animal was returned to the lighted compartment and the latency period of the transition to the dark compartment of the chamber was recorded. Stimulus delivery, the position of the door between compartments, and the time of transition from one compartment to another were recorded automatically with the Shutavoid v1.8.03 program (Panlab, Spain).
The Porsolt forced swimming test was performed in a glass cylinder with a height of 40 cm high and 20 cm in diameter, filled with water to the level of 25 cm. The water temperature was +25°C. During the test, the animal was placed in the cylinder for 5 min. The animal’s behaviour was recorded by camera and analysed using Smart Video Tracking System software v. 3.0 (Panlab, Spain). Animal strategies such as climbing, active swimming and immobility were recorded. At the end of the test, the mouse was removed from the water and excess water was removed from the coat with a paper towel. The animal was then placed on a heating pad to dry for 15 minutes before being returned to its home cage.
Immunohistochemical staining was performed on 12 mice. Transcardial perfusion with PFA was performed before brain dissection. Heparin solution in 0.1M PBS was injected for the first 15 seconds of perfusion, then the solution was changed to 4% PFA solution in 0.1M PBS in a volume of 50 ml, and the brain was extracted only at the end of this procedure. Each brain was sectioned along the midline into 2 hemispheres. The left hemisphere was then cut for frontal slices for further analysis of the dorsal hippocampus, while the right hemisphere was used for horizontal slices for further analysis of the ventral hippocampus, entorhinal cortex areas. Slices were prepared using a McIlwain Tissue Chopper (United Kingstone) to a total thickness of 300 µm. The prepared slices were fixed by incubation in 4% PFA solution at 4oC for 24 hours. In the next step, the sections were soaked in ethylene glycol and sucrose solution until complete immersion and then stored at -20oC until immunohistochemical processing. Sections were stained in 48-well plates. Tissues were washed twice from ethylene glycol with Tris-glycine buffer (0.1 M glycine normalised to pH 7.2-7.4 with 1 M Tris buffer), followed by incubation in this solution for 1 hour on an orbital shaker at 120 rpm. Unspecific binding was then blocked with a mixture of 5% goat serum, 0.5% Triton X-100 and 0.1% Tween 20 in PBS for 1 hour at RT using the same shaker characteristics. In the next step, each slice was transferred to a 48-well plate containing 300 uL of a mixture of chicken primary anti-MAP2 antibodies (Abcam, AB5392, 1:350), 5% goat serum, 0.5% Tween 20 in PBS. The plate was kept on an orbital shaker at 50 rpm and 4oC for 72 hours during antibody incubation. Samples were washed three times within 10 minutes with a wash buffer (0.1% Triton X-100, 0.1% Tween 20 in PBS) on an orbital shaker at 120 rpm. The next step was to incubate the sections with 400 µL of a mixture of secondary Alexa Fluor 488 anti-chicken IgY antibody (Invitrogen, A-11039, 1:500), 5% goat serum, 0.5% Triton X-100, 0.1% Tween 20 in PBS. The incubation was carried out for 1 hour at RT on an orbital shaker at 50 rpm. Following the same scheme, sections were incubated with rabbit polyclonal anti-Iba1 antibody (Fujifilm, 019-19741, 1:500) and Alexa Fluor 555 goat anti-rabbit IgG antibody (Invitrogen, A-21428, 1:500), guinea pig anti-GFAP antibody (Synaptic Systems, 173004, 1:350) and Alexa Fluor 647 goat anti-guinea pig IgG antibody (Invitrogen, A-21450, 1:500). After the final wash, sections were rinsed once with PBS and incubated in 500 uL Hoechst 33258 solution (1 µg/ml) for 10 minutes on an orbital shaker at 50 rpm for nuclear visualisation. After nuclear staining, sections were washed with PBS at RT for 10 minutes on 120 rpm orbital shaker and mounted with DAPI mounting medium (Abcam, ab104139). Final drying of samples was performed in a fridge at 4oC.
Morphometric analysis of glial cells was performed using NIS Elements software (Nikon). For general analysis, Z-stacks of the dorsal and ventral parts of the hippocampus with a total thickness of 24 um were prepared. Since MAP2 intensity in the brain slices can be correlated with structural dynamics of neuronal networks and dendritic plasticity, a detailed comparative analysis of intensity was prepared for the CA1 field in the ventral and dorsal hippocampus. For this purpose, three random regions of interest in the rad of CA1 field of the dorsal and ventral hippocampus and the entorhinal cortex were analysed with maximum pinhole width (optical thickness = 1 um). Mean intensity was then measured using the General Analysis3 utility. The quantitative cell ratio in the 24 um section was measured using a general image of the stratum radiatum of the CA1 field of the hippocampus. 2D mask was then created and parameters such as MinFeret, MaxFeret, Area, Circularity, ShapeFactor were analysed. Astrocytes were analysed using the same binary mask projection scheme. Statistical analysis of normality was performed using the Kolmogorov-Smirnov test. Statistical analysis carried out using the Mann-Whitney test, data visualisation includes median and 25, 75 percentiles.
ELISA. Brain tissue for ELISA was obtained after decapitation. The time from decapitation to freezing the brain tissue samples (hippocampus and cortex) in liquid nitrogen was less than 3 minutes. The samples were then stored at -80 oC. Prior to analysis, the mouse cortex was thawed, weighed and homogenised in a hand-held homogenizer using PBS 1:10 per weight, all procedures were performed on ice. It was centrifuged at 10000 xg for 15 min at 4°C. The supernatant was then snap frozen in liquid nitrogen. Samples were stored at -80°C for future use. In each group, n=6. Mann-Whitney test was used for statistical analysis, p-value < 0.05 was considered statistically significant. Median and 25th, 75th percentiles were selected for data visualisation. TNFa, Il-1b, Il-10 concentrations were tested using an ELISA kits (Cloud-Cone Corporation, #SEA133Mu, #MEA563Mu, #SEA056Mu) according to the manufacturer’s instructions. The ELISA analysis was based on the sandwich method using microtitre plates. Concentrations of MFN1, MARCH5 and Sigma R1 were determined using ELISA kits (BlueGene Biotech, #E03M1101, #E03M110, #E03S0058) according to the manufacturer’s instructions. The kits use a competitive enzyme-linked immunosorbent assay technique with anti-substrate and anti-conjugate antibodies. Microplate Washer APW-200 (China) was used for automatic washing of the plates. The optical density in each well was determined using a multifunctional automatic microplate photometer (Synergy™ MXT, BioTek, USA) set at 450 nm. A standard curve was generated using software (CurveExpert 1.4) from which the concentration for each sample was interpolated.
Real-time polymerase chain reaction. Total RNA extracted from hippocampal tissue was extracted using the analogue of TRIzol reagent (Evrogen, BC032) according to the manufacturer’s instructions. Synthesis of cDNA was performed with the MMLV RT kit (Evrogen, SK021S), using random deca-oligonucleotide primers. The PCR reaction was done using a master mix containing SYBR blue (Biolabmix, MHC030-2040). PCR amplification was performed using a CFX 96 instrument (Bio-Rad), with the annealing temperature of the primers set at 61 oC according to the previously experimentally determined efficiency value by titration method. The following housekeeping genes were used: Actb, Gapdh, Rpl19, B2m, Gusb and Pum1. The genes of interest were Il6, SIGMAR1. Primer sequence information and efficiency data are shown in Table 2.
Hellemans’ approach was used to evaluate gene expression relative to housekeeping genes [50]. Statistical assessment of normality of sample data was performed using the Kolmogorov-Smirnov criterion. Statistical significance was determined using the non-parametric Kruskal-Wallis criterion, data visualisation includes median and 25th, 75th percentiles.
Author Contributions
Conceptualization, O.M.S.; methodology, O.M.S., D.M.K., S.A.K., V.I.P., I.V.M.; validation, O.M.S., P.I.V., N.A.S and D.M.K.; formal analysis, O.M.S., N.A.S, P.I.V., O.G.Z. and D.M.K.; investigation, O.M.S., D.M.K., O.G.Z., EIK, S.A.K., V.I.P.; resources, O.M.S. and I.V.M.; data curation, O.M.S. and D.M.K.; writing—original draft preparation,O.M.S., O.G.Z., D.M.K.; writing—review and editing, N.A.S, P.I.V., V.I.P., S.A.K.; visualization, O.M.S..; supervision, O.M.S. and I.V.M.; project administration, O.M.S.; funding acquisition, O.M.S. and I.V.M. All authors have read and agreed to the published version of the manuscript.
Funding
The work was carried out with financial support from the Russian Science Foundation (project № 22-15-20043).
Institutional Review Board Statement
All procedures were performed in accordance with the European Communities Council Directive (November 24, 1986; 86/609/EEC) and the Declaration on humane treatment of animals. The Protocol of experiments was approved by the Commission on Bioethics of the Volga Region Research Medical University (No. 11 of 04.06.2024).
Data Availability Statement
The underlying data has been deposited in the Harvard Dataverse repository. Enzyme immunoassay data are available at Shirokova, Olesya; Shchelchkova Natalya; Kozlyaeva Elizaveta, 2024, “Data_ELISA_CUMS”, https://doi.org/10.7910/DVN/MKHVYU, Harvard Dataverse, V1 Confocal microscope images are available at Shirokova, Olesya; Korotchenko, Svetlana, 2024, “Images_CUMS”, https://doi.org/10.7910/DVN/8QTL1D, Harvard Dataverse, V1 Real-time PCR data are available at Shirokova, Olesya; Vasilchikov Petr; Pershin Vladimir, 2024, “Real-Time PCR of Mice after CUMS for F1000Research”, https://doi.org/10.7910/DVN/Q4SO0Z, Harvard Dataverse, V1 Data from behavioral tests are published in: Shirokova, Olesya; Daria Kuzmina; Olga Zaborskaya, 2024, “Behavior of mice after CUMS for the F1000 study”, https://doi.org/10.7910/DVN/4F9P6K, Harvard Dataverse, V1
Acknowledgments
Thanks to Chernov Yakov (PRMU) for help with sample preparation and illustration.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ACC | Anterior cingulate cortex |
| ActB | Actin beta |
| ADHD | Attention deficit hyperactivity disorder |
| B2m | Beta-2 microglobulin |
| CA | Cornu Ammonis |
| cDNA | Complementary deoxyribonucleic acid |
| CUMS | Chronic unpredictable mild stress |
| C57BL/6 | Common inbred strain of laboratory mouse |
| DAPI | 4′,6-diamidino-2-phenylindole |
| ELISA | Enzyme-linked immunosorbent assay |
| ER | Endoplasmic reticulum |
| GABA | Gamma-aminobutyric acid |
| Gapdh | Glyceraldehyde 3-phosphate dehydrogenase |
| GFAP | Glial fibrillary acidic protein |
| Gusb | Beta-Glucuronidase |
| HPA | Hypothalamic-pituitary-adrenal |
| IBA1 | Ionized calcium-binding adapter molecule 1 |
| IFM | Institute of Fundamental Medicine |
| IgG | Immunoglobulin G |
| IgY | Immunoglobulin Y |
| IL | Interleukin |
| IP3 | Inositol 1,4,5-trisphosphate |
| LTP | Long-term potentiation |
| MAM | Mitochondria-associated ER membrane |
| MAP2 | Microtubule-associated protein 2 |
| MARCH5 | Membrane-associated ring finger (C3HC4) 5 |
| MFN1 | Mitofusin-1 |
| MMLV | Murine leukemia virus |
| mPFC | Medial prefrontal cortex |
| mRNA | Messenger ribonucleic acid |
| NAc | Nucleus accumbens |
| PA | Passive avoidance |
| PBS | Phosphate-buffered saline |
| PCR | Polymerase chain reaction |
| PFA | Paraformaldehyde |
| PRMU | Privolzhsky Research Medical University |
| Pum1 | Pumilio homolog 1 |
| RNA | Ribonucleic acid |
| Rpl19 | Large ribosomal subunit 19 |
| RT | Room temperature |
| SIGMAR1 | Sigma-1 receptor |
| SPF | Specific-pathogen-free |
| TNF-α | Tumor necrosis factor |
| TH | Tyrosine hydroxylase |
References
- Csabai, D.; Sebők-Tornai, A.; Wiborg, O.; Czéh, B. A Preliminary Quantitative Electron Microscopic Analysis Reveals Reduced Number of Mitochondria in the Infralimbic Cortex of Rats Exposed to Chronic Mild Stress. Front. Behav. Neurosci. 2022, 16, 885849. [Google Scholar] [CrossRef]
- Hill, M.N.; Hellemans, K.G.C.; Verma, P.; Gorzalka, B.B.; Weinberg, J. Neurobiology of Chronic Mild Stress: Parallels to Major Depression. Neurosci. Biobehav. Rev. 2012, 36, 2085–2117. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Mouri, A.; Yang, Y.; Kunisawa, K.; Teshigawara, T.; Hirakawa, M.; Mori, Y.; Yamamoto, Y.; Libo, Z.; Nabeshima, T.; et al. Chronic Unpredictable Mild Stress-Induced Behavioral Changes Are Coupled with Dopaminergic Hyperfunction and Serotonergic Hypofunction in Mouse Models of Depression. Behav. Brain Res. 2019, 372, 112053. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Chen, Y.-X.; Hu, Y.-T.; Wu, X.-Y.; He, Y.; Wu, J.-L.; Huang, M.-L.; Mason, M.; Bao, A.-M. Sex Hormones Affect Acute and Chronic Stress Responses in Sexually Dimorphic Patterns: Consequences for Depression Models. Psychoneuroendocrinology 2018, 95, 34–42. [Google Scholar] [CrossRef]
- Pillerová, M.; Borbélyová, V.; Pastorek, M.; Riljak, V.; Hodosy, J.; Frick, K.M.; Tóthová, L. Molecular Actions of Sex Hormones in the Brain and Their Potential Treatment Use in Anxiety Disorders. Front. Psychiatry 2022, 13, 972158. [Google Scholar] [CrossRef]
- Sze, Y.; Brunton, P.J. Sex, Stress and Steroids. Eur. J. Neurosci. 2020, 52, 2487–2515. [Google Scholar] [CrossRef]
- Sheng, J.A.; Tan, S.M.L.; Hale, T.M.; Handa, R.J. Androgens and Their Role in Regulating Sex Differences in the Hypothalamic/Pituitary/Adrenal Axis Stress Response and Stress-Related Behaviors. Androg. Clin. Res. Ther. 2021, 2, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.Y.; Kim, D.W.; Nam, S.M.; Kim, J.W.; Chung, J.Y.; Won, M.-H.; Seong, J.K.; Yoon, Y.S.; Yoo, D.Y.; Hwang, I.K. Pyridoxine Improves Hippocampal Cognitive Function via Increases of Serotonin Turnover and Tyrosine Hydroxylase, and Its Association with CB1 Cannabinoid Receptor-Interacting Protein and the CB1 Cannabinoid Receptor Pathway. Biochim. Biophys. Acta BBA - Gen. Subj. 2017, 1861, 3142–3153. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Mouri, A.; Yang, Y.; Kunisawa, K.; Teshigawara, T.; Hirakawa, M.; Mori, Y.; Yamamoto, Y.; Libo, Z.; Nabeshima, T.; et al. Chronic Unpredictable Mild Stress-Induced Behavioral Changes Are Coupled with Dopaminergic Hyperfunction and Serotonergic Hypofunction in Mouse Models of Depression. Behav. Brain Res. 2019, 372, 112053. [Google Scholar] [CrossRef] [PubMed]
- Di, T.; Zhang, S.; Hong, J.; Zhang, T.; Chen, L. Hyperactivity of Hypothalamic-Pituitary-Adrenal Axis Due to Dysfunction of the Hypothalamic Glucocorticoid Receptor in Sigma-1 Receptor Knockout Mice. Front. Mol. Neurosci. 2017, 10, 287. [Google Scholar] [CrossRef]
- Rohleder, N. Stress and Inflammation – The Need to Address the Gap in the Transition between Acute and Chronic Stress Effects. Psychoneuroendocrinology 2019, 105, 164–171. [Google Scholar] [CrossRef]
- Avolio, E.; Fazzari, G.; Mele, M.; Alò, R.; Zizza, M.; Jiao, W.; Di Vito, A.; Barni, T.; Mandalà, M.; Canonaco, M. Unpredictable Chronic Mild Stress Paradigm Established Effects of Pro- and Anti-Inflammatory Cytokine on Neurodegeneration-Linked Depressive States in Hamsters with Brain Endothelial Damages. Mol. Neurobiol. 2017, 54, 6446–6458. [Google Scholar] [CrossRef]
- Sin, N.L.; Graham-Engeland, J.E.; Ong, A.D.; Almeida, D.M. Affective Reactivity to Daily Stressors Is Associated with Elevated Inflammation. Health Psychol. 2015, 34, 1154–1165. [Google Scholar] [CrossRef] [PubMed]
- Calcia, M.A.; Bonsall, D.R.; Bloomfield, P.S.; Selvaraj, S.; Barichello, T.; Howes, O.D. Stress and Neuroinflammation: A Systematic Review of the Effects of Stress on Microglia and the Implications for Mental Illness. Psychopharmacology (Berl.) 2016, 233, 1637–1650. [Google Scholar] [CrossRef] [PubMed]
- Ooi, K.; Hu, L.; Feng, Y.; Han, C.; Ren, X.; Qian, X.; Huang, H.; Chen, S.; Shi, Q.; Lin, H.; et al. Sigma-1 Receptor Activation Suppresses Microglia M1 Polarization via Regulating Endoplasmic Reticulum–Mitochondria Contact and Mitochondrial Functions in Stress-Induced Hypertension Rats. Mol. Neurobiol. 2021, 58, 6625–6646. [Google Scholar] [CrossRef]
- Hayashi, T. Conversion of Psychological Stress into Cellular Stress Response: Roles of the Sigma-1 Receptor in the Process. Psychiatry Clin. Neurosci. 2015, 69, 179–191. [Google Scholar] [CrossRef]
- Shiiba, I.; Takeda, K.; Nagashima, S.; Yanagi, S. Overview of Mitochondrial E3 Ubiquitin Ligase MITOL/MARCH5 from Molecular Mechanisms to Diseases. Int. J. Mol. Sci. 2020, 21, 3781. [Google Scholar] [CrossRef] [PubMed]
- Zorzano, A.; Pich, S. What Is the Biological Significance of the Two Mitofusin Proteins Present in the Outer Mitochondrial Membrane of Mammalian Cells? IUBMB Life Int. Union Biochem. Mol. Biol. Life 2006, 58, 441–443. [Google Scholar] [CrossRef]
- Okuyama, T.; Kitamura, T.; Roy, D.S.; Itohara, S.; Tonegawa, S. Ventral CA1 Neurons Store Social Memory. Science 2016, 353, 1536–1541. [Google Scholar] [CrossRef]
- Donegan, M.L.; Stefanini, F.; Meira, T.; Gordon, J.A.; Fusi, S.; Siegelbaum, S.A. Coding of Social Novelty in the Hippocampal CA2 Region and Its Disruption and Rescue in a 22q11.2 Microdeletion Mouse Model. Nat. Neurosci. 2020, 23, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.L.; Robinson, H.A.; Pozzo-Miller, L. Ventral Hippocampal Projections to the Medial Prefrontal Cortex Regulate Social Memory. eLife 2019, 8, e44182. [Google Scholar] [CrossRef] [PubMed]
- Dawson, D.A.; Hallenbeck, J.M. Acute Focal Ischemia-Induced Alterations in MAP2 Immunostaining: Description of Temporal Changes and Utilization as a Marker for Volumetric Assessment of Acute Brain Injury. J. Cereb. Blood Flow Metab. 1996, 16, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Steward, O.; Halpain, S. Lamina-Specific Synaptic Activation Causes Domain-Specific Alterations in Dendritic Immunostaining for MAP2 and CAM Kinase II. J. Neurosci. 1999, 19, 7834–7845. [Google Scholar] [CrossRef]
- Goldfarb, E.V.; Seo, D.; Sinha, R. Sex Differences in Neural Stress Responses and Correlation with Subjective Stress and Stress Regulation. Neurobiol. Stress 2019, 11, 100177. [Google Scholar] [CrossRef] [PubMed]
- Heck, A.L.; Handa, R.J. Sex Differences in the Hypothalamic–Pituitary–Adrenal Axis’ Response to Stress: An Important Role for Gonadal Hormones. Neuropsychopharmacology 2019, 44, 45–58. [Google Scholar] [CrossRef]
- Wellman, C.L.; Bangasser, D.A.; Bollinger, J.L.; Coutellier, L.; Logrip, M.L.; Moench, K.M.; Urban, K.R. Sex Differences in Risk and Resilience: Stress Effects on the Neural Substrates of Emotion and Motivation. J. Neurosci. 2018, 38, 9423–9432. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.R.; Spencer, K.A. Steroid Hormones, Stress and the Adolescent Brain: A Comparative Perspective. Neuroscience 2013, 249, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, T.; Kitamura, T.; Roy, D.S.; Itohara, S.; Tonegawa, S. Ventral CA1 Neurons Store Social Memory. Science 2016, 353, 1536–1541. [Google Scholar] [CrossRef]
- Tanimizu, T.; Kenney, J.W.; Okano, E.; Kadoma, K.; Frankland, P.W.; Kida, S. Functional Connectivity of Multiple Brain Regions Required for the Consolidation of Social Recognition Memory. J. Neurosci. 2017, 37, 4103–4116. [Google Scholar] [CrossRef]
- Leung, C.; Cao, F.; Nguyen, R.; Joshi, K.; Aqrabawi, A.J.; Xia, S.; Cortez, M.A.; Snead, O.C.; Kim, J.C.; Jia, Z. Activation of Entorhinal Cortical Projections to the Dentate Gyrus Underlies Social Memory Retrieval. Cell Rep. 2018, 23, 2379–2391. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Eiland, L.; Hunter, R.G.; Miller, M.M. Stress and Anxiety: Structural Plasticity and Epigenetic Regulation as a Consequence of Stress. Neuropharmacology 2012, 62, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Holzschneider, K.; Mulert, C. Neuroimaging in Anxiety Disorders. Dialogues Clin. Neurosci. 2011, 13, 453–461. [Google Scholar] [CrossRef]
- Monnet, F.P.; Morin-Surun, M.P.; Leger, J.; Combettes, L. Protein Kinase C-Dependent Potentiation of Intracellular Calcium Influx by σ1 Receptor Agonists in Rat Hippocampal Neurons. J. Pharmacol. Exp. Ther. 2003, 307, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Aniszewska, A.; Chłodzińska, N.; Bartkowska, K.; Winnicka, M.M.; Turlejski, K.; Djavadian, R.L. The Expression of Interleukin-6 and Its Receptor in Various Brain Regions and Their Roles in Exploratory Behavior and Stress Responses. J. Neuroimmunol. 2015, 284, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.-L.; Peng, J.-B.; Fu, C.-H.; Tong, L.; Wang, Z.-Y. Sigma-1 Receptor Activation Ameliorates Anxiety-like Behavior through NR2A-CREB-BDNF Signaling Pathway in a Rat Model Submitted to Single-Prolonged Stress. Mol. Med. Rep. 2017, 16, 4987–4993. [Google Scholar] [CrossRef]
- Timaru-Kast, R.; Herbig, E.L.; Luh, C.; Engelhard, K.; Thal, S.C. Influence of Age on Cerebral Housekeeping Gene Expression for Normalization of Quantitative Polymerase Chain Reaction after Acute Brain Injury in Mice. J. Neurotrauma 2015, 32, 1777–1788. [Google Scholar] [CrossRef]
- Kovalenko, A.A.; Zakharova, M.V.; Schwarz, A.P.; Zubareva, O.E.; Zaitsev, A.V. Identification of Reliable Reference Genes for Use in Gene Expression Studies in Rat Febrile Seizure Model. Int. J. Mol. Sci. 2024, 25, 11125. [Google Scholar] [CrossRef] [PubMed]
- Kourrich, S.; Su, T.-P.; Fujimoto, M.; Bonci, A. The Sigma-1 Receptor: Roles in Neuronal Plasticity and Disease. Trends Neurosci. 2012, 35, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Su, T.-P.; Hayashi, T. Understanding the Molecular Mechanism of Sigma-1 Receptors: Towards A Hypothesis That Sigma-1 Receptors Are Intracellular Amplifiers for Signal Transduction. Curr. Med. Chem. 2003, 10, 2073–2080. [Google Scholar] [CrossRef]
- Zhang, K.; Zhao, Z.; Lan, L.; Wei, X.; Wang, L.; Liu, X.; Yan, H.; Zheng, J. Sigma-1 Receptor Plays a Negative Modulation on N-Type Calcium Channel. Front. Pharmacol. 2017, 8, 302. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Wang, L.; Zhang, T.; Zhang, B.; Chen, L. Sigma-1 Receptor Knockout Increases α-Synuclein Aggregation and Phosphorylation with Loss of Dopaminergic Neurons in Substantia Nigra. Neurobiol. Aging 2017, 59, 171–183. [Google Scholar] [CrossRef]
- Bernard-Marissal, N.; Médard, J.-J.; Azzedine, H.; Chrast, R. Dysfunction in Endoplasmic Reticulum-Mitochondria Crosstalk Underlies SIGMAR1 Loss of Function Mediated Motor Neuron Degeneration. Brain 2015, 138, 875–890. [Google Scholar] [CrossRef]
- Hong, J.; Wang, L.; Zhang, T.; Zhang, B.; Chen, L. Sigma-1 Receptor Knockout Increases α-Synuclein Aggregation and Phosphorylation with Loss of Dopaminergic Neurons in Substantia Nigra. Neurobiol. Aging 2017, 59, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Dawson, D.A.; Hallenbeck, J.M. Acute Focal Ischemia-Induced Alterations in MAP2 Immunostaining: Description of Temporal Changes and Utilization as a Marker for Volumetric Assessment of Acute Brain Injury. J. Cereb. Blood Flow Metab. 1996, 16, 170–174. [Google Scholar] [CrossRef]
- Crouzier, L.; Couly, S.; Roques, C.; Peter, C.; Belkhiter, R.; Arguel Jacquemin, M.; Bonetto, A.; Delprat, B.; Maurice, T. Sigma-1 (Σ1) Receptor Activity Is Necessary for Physiological Brain Plasticity in Mice. Eur. Neuropsychopharmacol. 2020, 39, 29–45. [Google Scholar] [CrossRef]
- Shi, J.-J.; Jiang, Q.-H.; Zhang, T.-N.; Sun, H.; Shi, W.-W.; Gunosewoyo, H.; Yang, F.; Tang, J.; Pang, T.; Yu, L.-F. Sigma-1 Receptor Agonist TS-157 Improves Motor Functional Recovery by Promoting Neurite Outgrowth and pERK in Rats with Focal Cerebral Ischemia. Molecules 2021, 26, 1212. [Google Scholar] [CrossRef]
- Takao, T.; Culp, S.G.; De Souza, E.B. Reciprocal Modulation of Interleukin-1 Beta (IL-1 Beta) and IL-1 Receptors by Lipopolysaccharide (Endotoxin) Treatment in the Mouse Brain-Endocrine-Immune Axis. Endocrinology 1993, 132, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.A.; Lynch, M.A. Evidence That Increased Hippocampal Expression of the Cytokine Interleukin-1β Is a Common Trigger for Age- and Stress-Induced Impairments in Long-Term Potentiation. J. Neurosci. 1998, 18, 2974–2981. [Google Scholar] [CrossRef]
- Christmann, U.; Garriga, L.; Llorente, A.V.; Díaz, J.L.; Pascual, R.; Bordas, M.; Dordal, A.; Porras, M.; Yeste, S.; Reinoso, R.F.; et al. WLB-87848, a Selective σ1 Receptor Agonist, with an Unusually Positioned NH Group as Positive Ionizable Moiety and Showing Neuroprotective Activity. J. Med. Chem. 2024, 67, 9150–9164. [Google Scholar] [CrossRef]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase Relative Quantification Framework and Software for Management and Automated Analysis of Real-Time Quantitative PCR Data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of CUMS modelling and behavioural test battery.
Figure 2.
Effect of CUMS on male and female mice in the Open Field test. (A) Time spent in the central area during the 5-minute test. (B) Total number of entries to the central zone of the Open Field arena. (C) Total time of grooming acts performed in the test. (D) Distance travelled in the Open Field. (E) Total time spent on resing. (F) Total duration of fast locomotion. (G) Mean walking speed of mouse in the 5-minute test period. (H) Number of vertical stands performed in the test. (I) Total number of acts of sniffing while exploring the Open Field. *p-values were calculated using the two-way repeated-measures ANOVA.
Figure 2.
Effect of CUMS on male and female mice in the Open Field test. (A) Time spent in the central area during the 5-minute test. (B) Total number of entries to the central zone of the Open Field arena. (C) Total time of grooming acts performed in the test. (D) Distance travelled in the Open Field. (E) Total time spent on resing. (F) Total duration of fast locomotion. (G) Mean walking speed of mouse in the 5-minute test period. (H) Number of vertical stands performed in the test. (I) Total number of acts of sniffing while exploring the Open Field. *p-values were calculated using the two-way repeated-measures ANOVA.
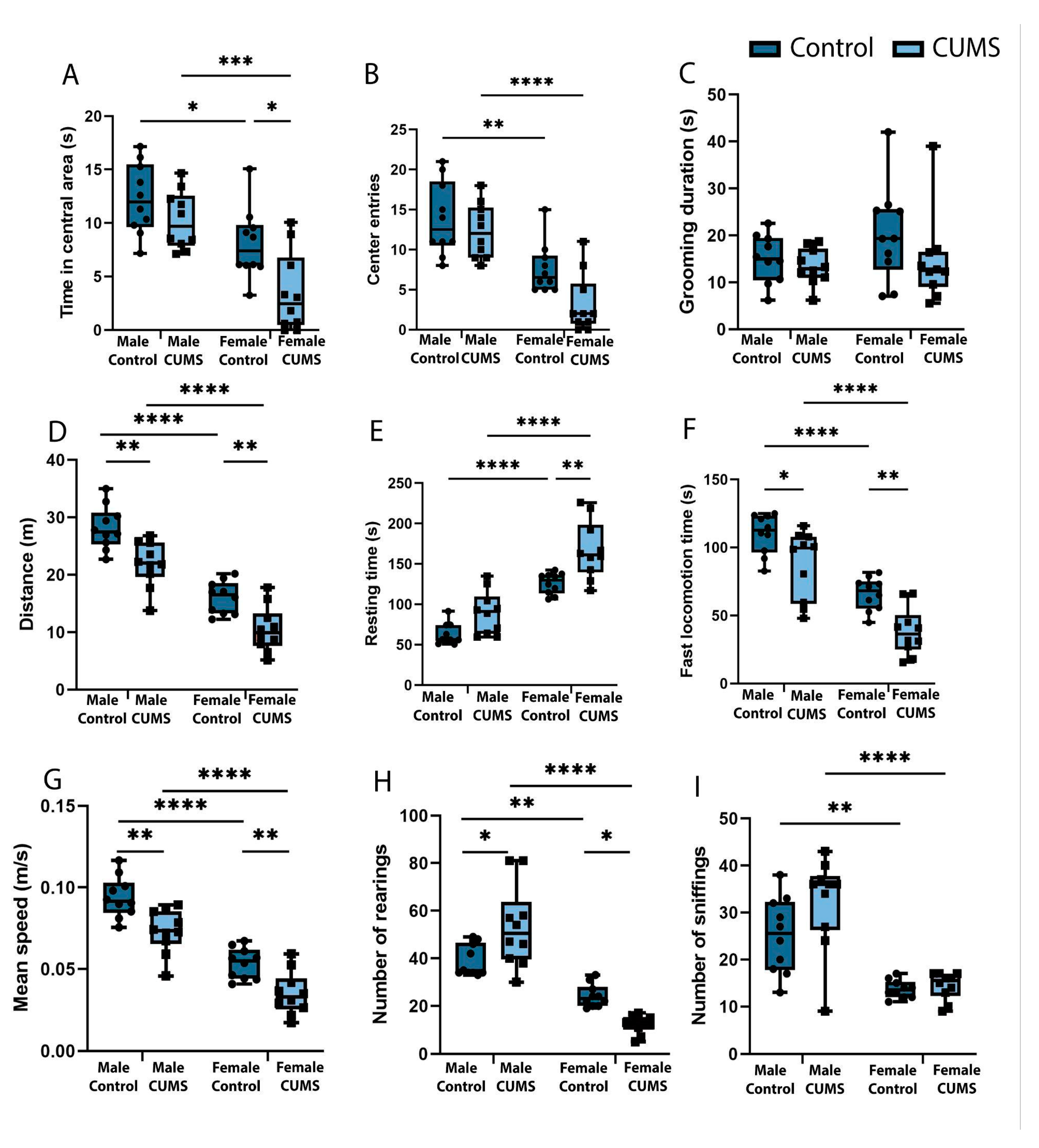
Figure 3.
Effect of CUMS on learning and memory of male and female mice. (A) Latency to enter the dark chamber in the passive avoidance test. (B) The social activity of mice in the three-chamber Crowley test. (C) Proportion (percentage) of time spent with an unfamiliar animal in the three-chamber Crowley test. (D) Proportion (percentage) of time spent with new object in the Novel Object Recognition test. *p-values were calculated using the two-way repeated-measures ANOVA.
Figure 3.
Effect of CUMS on learning and memory of male and female mice. (A) Latency to enter the dark chamber in the passive avoidance test. (B) The social activity of mice in the three-chamber Crowley test. (C) Proportion (percentage) of time spent with an unfamiliar animal in the three-chamber Crowley test. (D) Proportion (percentage) of time spent with new object in the Novel Object Recognition test. *p-values were calculated using the two-way repeated-measures ANOVA.
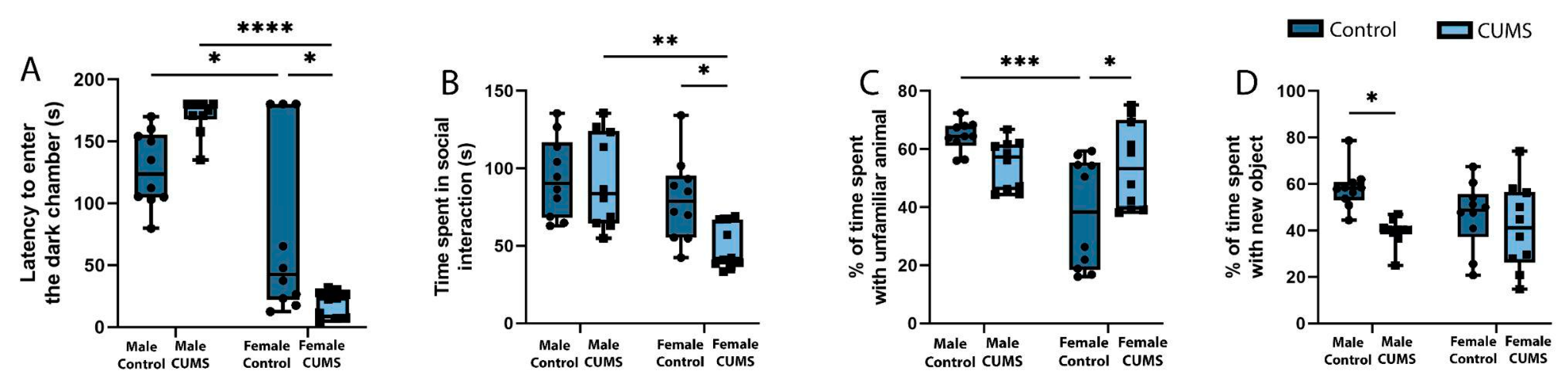
Figure 4.
Depressive-like behaviour of male and female mice in the Porsolt forced swim test after CUMS. (A) Time before the first immobilization. (B) Duration of immobilization. (С) Total climbing time. (D) Time spent on active swimming. *p-values were calculated using the two-way repeated-measures ANOVA.
Figure 4.
Depressive-like behaviour of male and female mice in the Porsolt forced swim test after CUMS. (A) Time before the first immobilization. (B) Duration of immobilization. (С) Total climbing time. (D) Time spent on active swimming. *p-values were calculated using the two-way repeated-measures ANOVA.
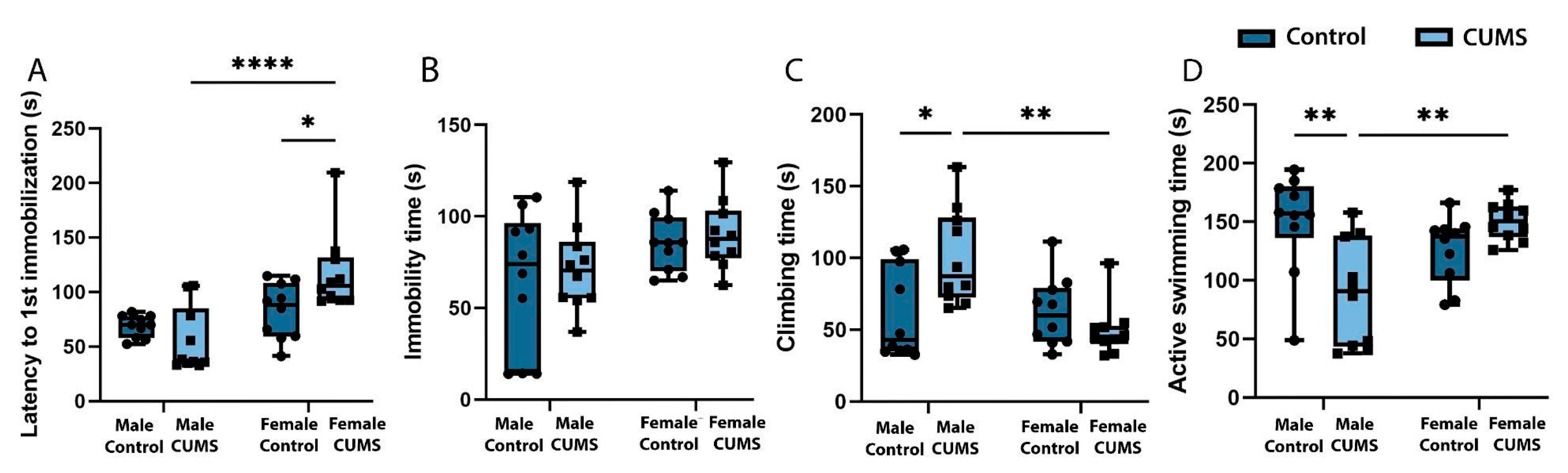
Figure 5.
Immunofluorescent analysis of different hippocampal and entorhinal cortex sections in each experimental group. A - Representative images of MAP2 labeling of the dorsal and ventral hippocampus the CA1 stratum radiatum and the enteric cortex. Scale bar: 10 µm. B - Mean intensity in different experimental groups.
Figure 5.
Immunofluorescent analysis of different hippocampal and entorhinal cortex sections in each experimental group. A - Representative images of MAP2 labeling of the dorsal and ventral hippocampus the CA1 stratum radiatum and the enteric cortex. Scale bar: 10 µm. B - Mean intensity in different experimental groups.
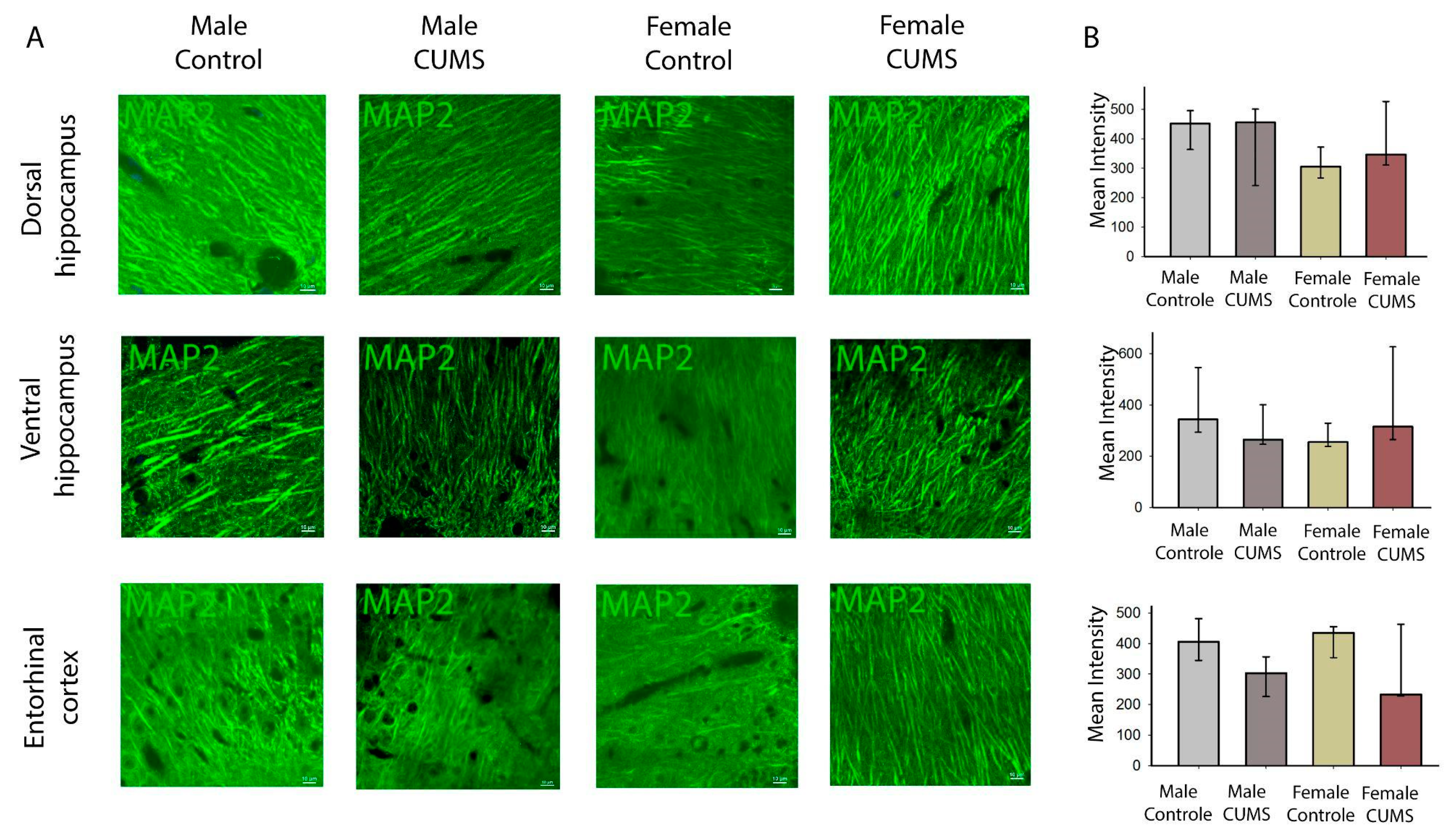
Figure 6.
Immunofluorescent analysis of the dorsal hippocampus. A - Triple labeling for MAP2, Iba1 and GFAP in the dorsal hippocampus. В-Representative image of the dorsal hippocampus on a frontal slice of the mouse brain. С-Number of astroglia (GFAP positive) and microglia (Iba positive). D- Morphometric analysis of astrocytes in the CA1 field of the hippocampus using ShapeFactor, Area, Circularity. Scale bar: 100 µm.
Figure 6.
Immunofluorescent analysis of the dorsal hippocampus. A - Triple labeling for MAP2, Iba1 and GFAP in the dorsal hippocampus. В-Representative image of the dorsal hippocampus on a frontal slice of the mouse brain. С-Number of astroglia (GFAP positive) and microglia (Iba positive). D- Morphometric analysis of astrocytes in the CA1 field of the hippocampus using ShapeFactor, Area, Circularity. Scale bar: 100 µm.
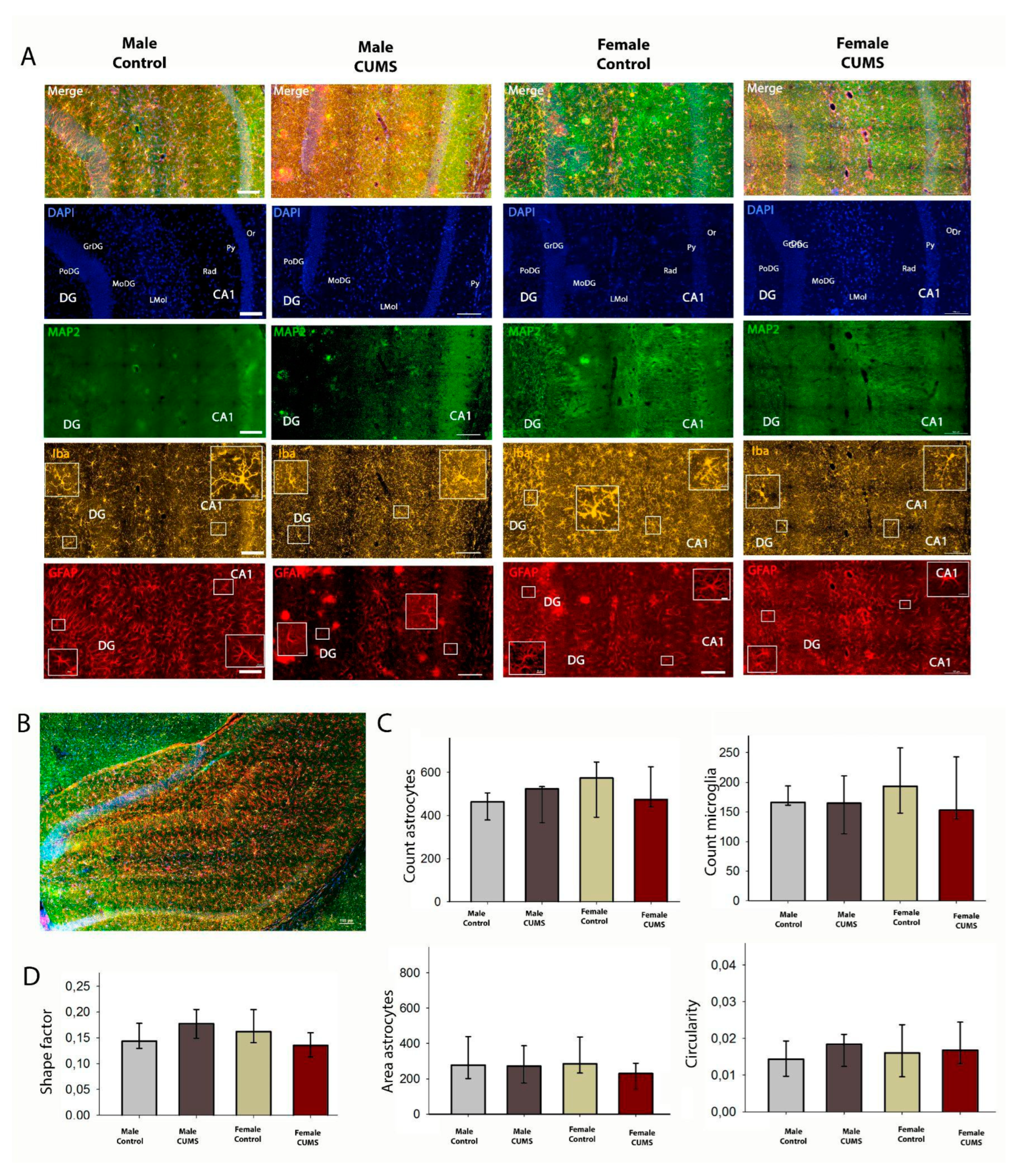
Figure 7.
Immunofluorescent analysis of ventral hippocampus. A - Triple labeling for MAP2, Iba1 and GFAP of ventral hippocampus. Scale bar: 100 µm. B - Representative image of the ventral hippocampus on a horizontal slice of the mouse brain, С - number of astroglial cells (GFAP-positive and microglial cells (iba-positive), D - morphometric analysis of astrocytes of the CA1 field of the hippocampus using ShapeFactor, Area, Circularity.
Figure 7.
Immunofluorescent analysis of ventral hippocampus. A - Triple labeling for MAP2, Iba1 and GFAP of ventral hippocampus. Scale bar: 100 µm. B - Representative image of the ventral hippocampus on a horizontal slice of the mouse brain, С - number of astroglial cells (GFAP-positive and microglial cells (iba-positive), D - morphometric analysis of astrocytes of the CA1 field of the hippocampus using ShapeFactor, Area, Circularity.
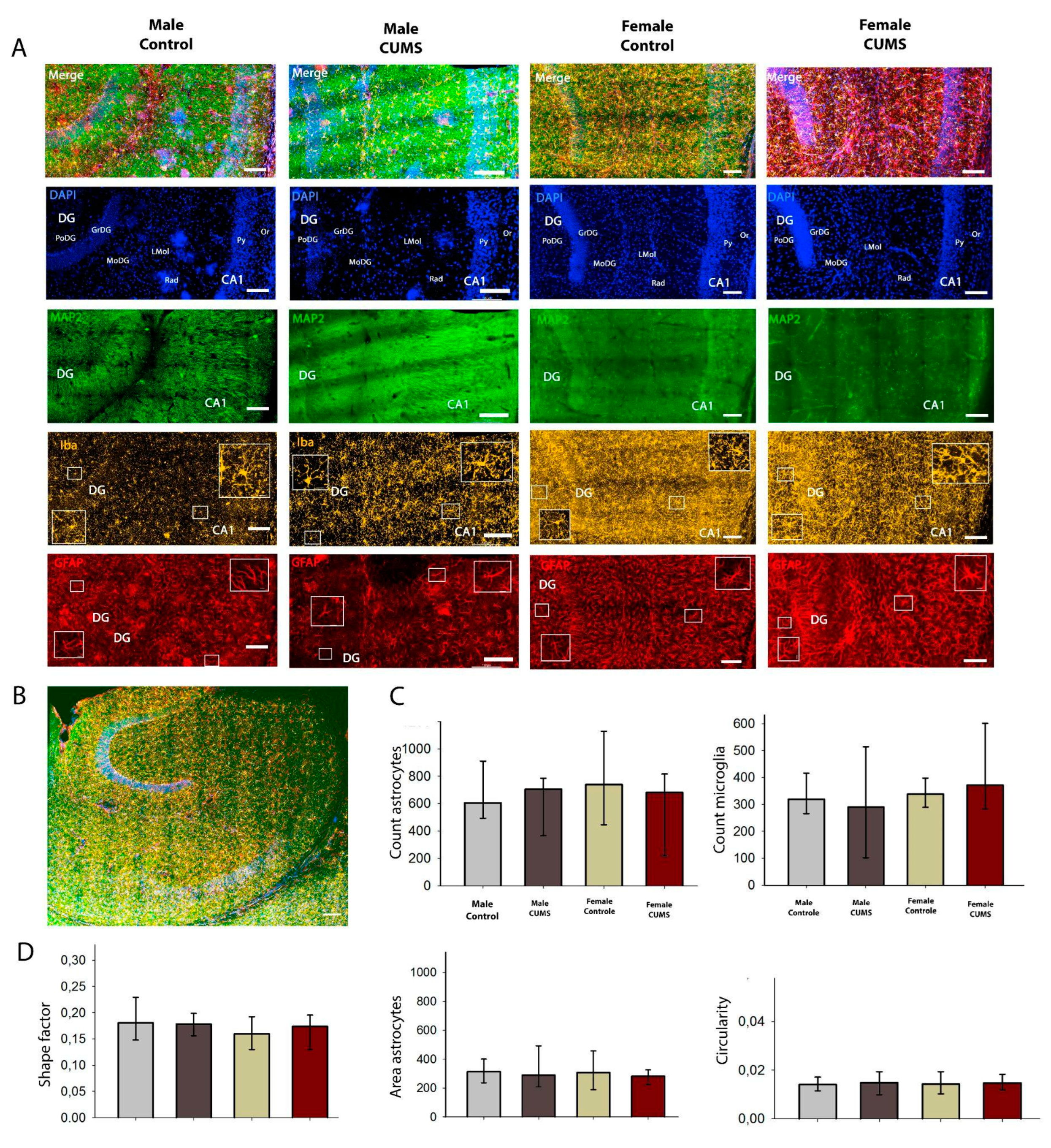
Figure 8.
Results of immunoenzymatic analysis of proteins of interest in the cerebral cortex of mice in different experimental groups. A-Concentration of pro-inflammatory IL-10. B-Concentration of anti-inflammatory factors IL-1b. C-Concentration of pro-inflammatory TNFa. E-F- Concentration of mitochondrial proteins MARCH5 and MFN.
Figure 8.
Results of immunoenzymatic analysis of proteins of interest in the cerebral cortex of mice in different experimental groups. A-Concentration of pro-inflammatory IL-10. B-Concentration of anti-inflammatory factors IL-1b. C-Concentration of pro-inflammatory TNFa. E-F- Concentration of mitochondrial proteins MARCH5 and MFN.
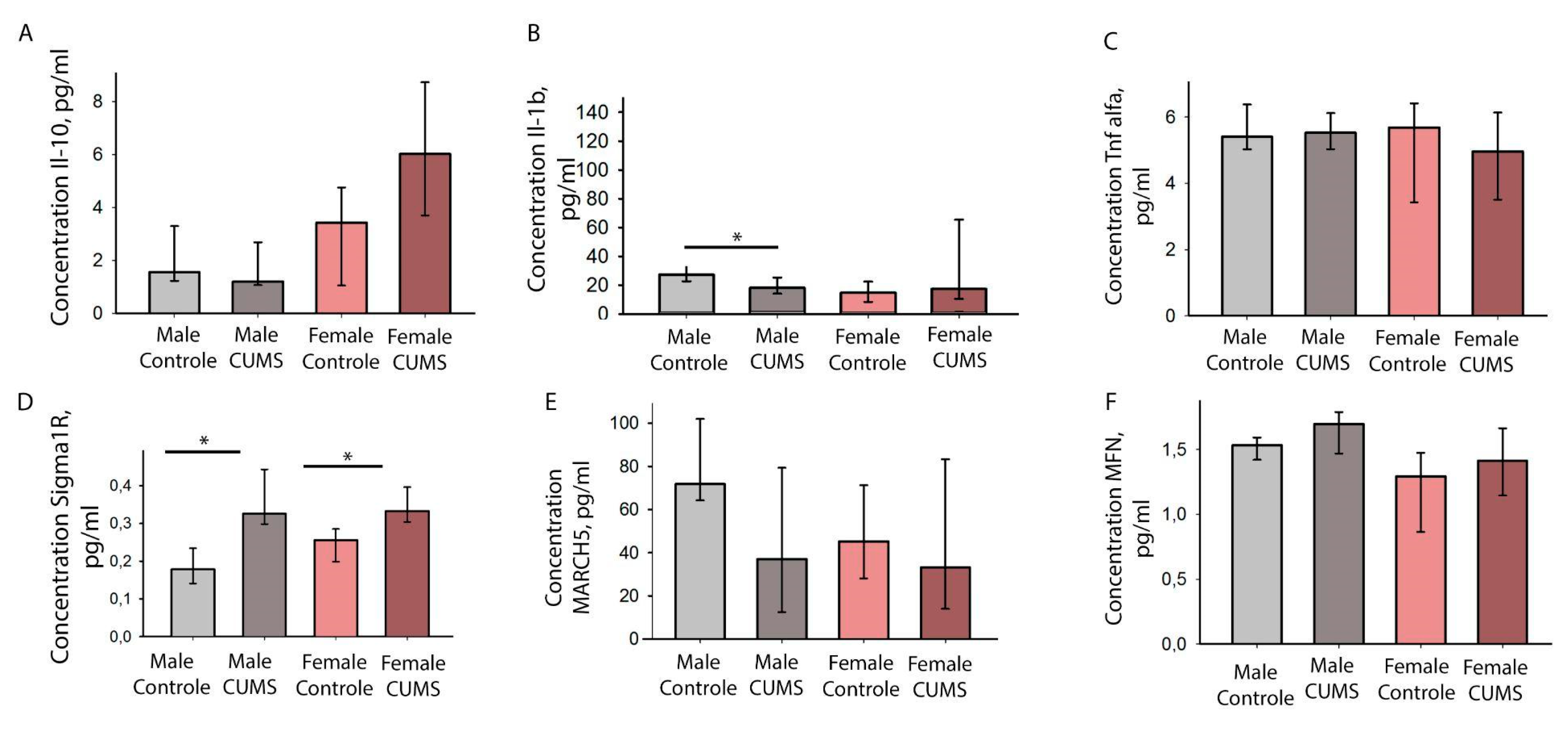
Figure 9.
Relative expression of SIGMAR1 and IL6 in the hippocampus. A - the expression level of SIGMAR1 relative to 6 housekeeping genes. В - the expression level of SIGMAR1 relative to individual housekeeping genes (ActB, B2m, Gapdh, Gusb, Pum1, Rpl1). C - the level of expression of IL-6 in relation to 6 housekeeping genes.
Figure 9.
Relative expression of SIGMAR1 and IL6 in the hippocampus. A - the expression level of SIGMAR1 relative to 6 housekeeping genes. В - the expression level of SIGMAR1 relative to individual housekeeping genes (ActB, B2m, Gapdh, Gusb, Pum1, Rpl1). C - the level of expression of IL-6 in relation to 6 housekeeping genes.
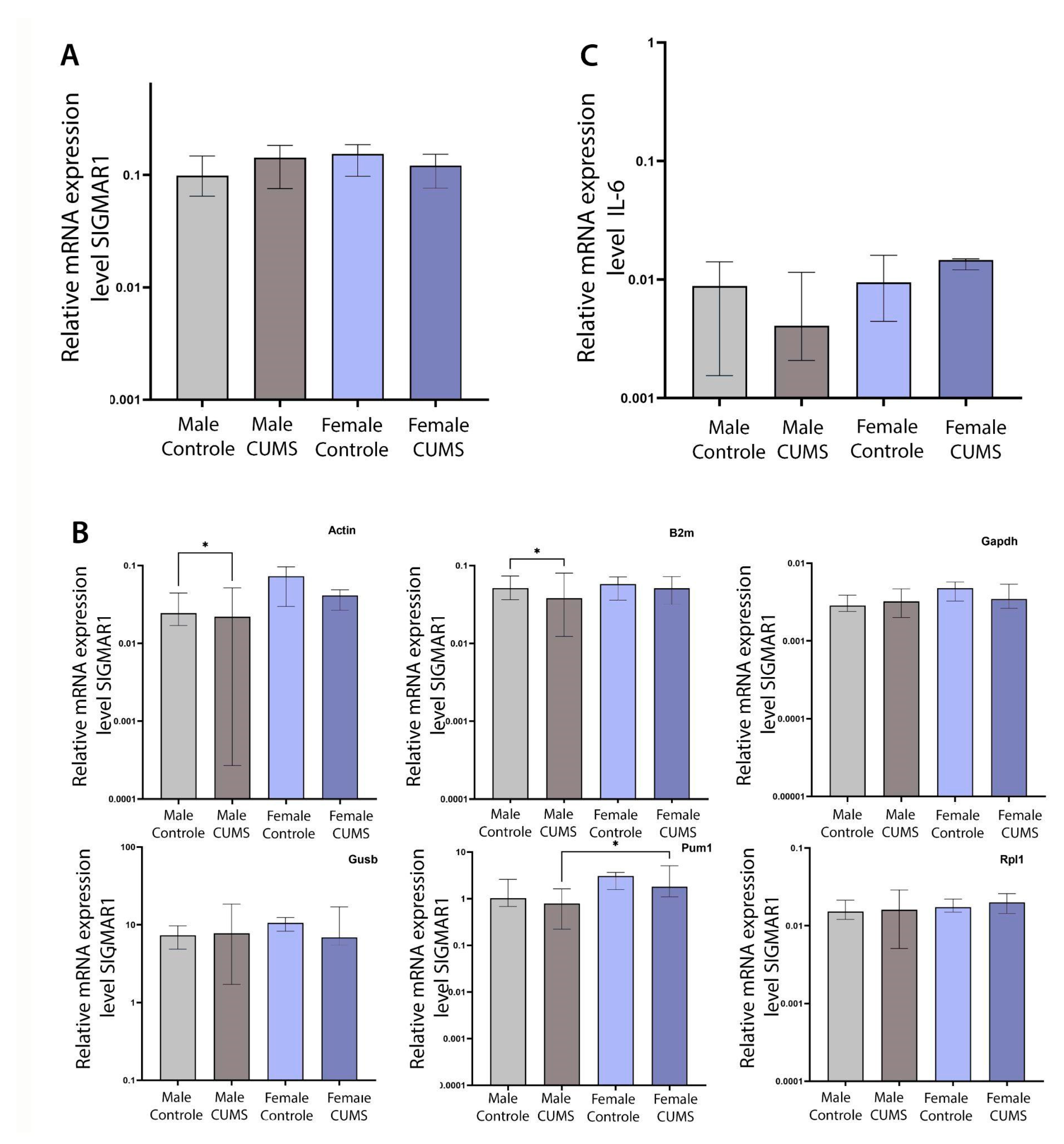

Table 1.
Experimental stressors.
| Stressor | Description |
|---|---|
| White noise | White noise (80 dB) is played through a loudspeaker near the homecages during 3 h |
| Tilted cage | Homecages were tilted in a 30° angle during 21 h |
| Water deprivation | Deprivation of water during 9 h |
| Food deprivation | Deprivation of food during 19 h |
| Overnight illumination | Mice were exposed to regular room light during 21 h |
| Foreign object | Plastic lego piece placed to the cage for 21 h |
| Shaking | Mice were placed in a plastic box container and placed in an orbital shaker for 3 h at 100 rpm |
| Foreign sawdust | The sawdust in homecages is changed with sawdust from other mice for 21 h |
| Rat sawdust | About 50 ml of rat sawdust is deposited in the homecage for a period during 24 h. |
| Damp sawdust | 40 ml water is placed in each cage for 5 |
Table 2.
List of primers and their characteristics in terms of efficiency.
| Name | 5’→3’ | Efficiency, % | Name | 5’→3’ | Efficiency, % |
|---|---|---|---|---|---|
| ACTB_R | GCCGGACTCATCGTACTCC | 109 | SIGMAR F_3 | CCTCTTTGGCCAAGACTCCTGA | 96 |
| ACTB_F | GTGACGTTGACATCCGTAAAGA | SIGMAR _3 | GCATGGTATACGCTGCTGTCTGA | ||
| GAPDH_F | CCCACTCTTCCACCTTCGATG | 101 | IL6_F | GAGACTTCCATCCAGTTGCCTTC | 107 |
| GAPDH_R | GTCCACCACCCTGTTGCTGTAG | IL6_R | GAACATGTGTAATTAAGCCTCCGAC | ||
| GusB_F | AGGACGTACTTCAGCTCTGTGAC | 114 | RPL19 F | TCATCCGCAAGCCTGTACTGT | 96 |
| GusB_R | TGCCGAAGTGACTCGTTGCCAA | RPL19 R | ACCTTCTCAGGCATCCGAGCAT | ||
| PUM F | ACAGCCTGCCAACACGTCCTTG | 118 | B2M F | ACAGTTCCACCCGCCTCACATT | 98 |
| PUM R | CCACTGCCAGTGTTGGAGTTTG | B2M R | TAGAAAGACCAGTCCTTGCTGAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



