
Submitted:
13 December 2024
Posted:
13 December 2024
You are already at the latest version
Abstract
Background: Cancer remains a leading cause of mortality globally. Conventional treatment modalities, includ-ing radiation and chemotherapy, often fall short of achieving complete remission, highlighting the critical need for novel therapeutic strategies. One promising approach involves the oncolytic potential of Group A Streptococcus (GAS) strains for tumor treatment. This study aimed to investigate the oncolytic efficacy of S. pyogenes GUR and its M protein knockout mutant, S. pyogenes strain GURSA1, which has been genetically con-structed to minimize overall toxicity, against mouse hepatoma 22A, pancreatic cancer PANC02, and human glioma U251 cells, both in vitro and in vivo, using the C57BL/6 mouse model.
Methods: The in vitro oncolytic cytotoxic activity of GAS strains was studied against human glioma U251, pancreatic cancer PANC02, murine hepatoma 22a, and normal skin fibroblast cells using the MTT assay and the real-time xCELLigence system. A syngeneic mouse model of hepatoma and pancreatic cancer was used to evaluate the in vivo oncolytic effect of GAS strains. Statistical analysis was conducted using the Student’s t-test and Mann-Whitney U-test with GraphPad Prism software.
Results: The in vitro model showed that the live S. pyogenes GUR strain had a strong cytotoxic effect (67.4±1.9%) against pancreatic cancer PANC02 cells. This strain exhibited moderate (38.0±1.8%) and weak (16.3±5.4%) oncolytic activities against glioma and hepatoma cells, respectively. In contrast, the S. pyogenes GURSA1 strain demonstrated strong (86.5±1.6%) and moderate (36.5±1.8%) oncolytic activities against glioma and hepatoma cells. Additionally, the S. pyogenes GURSA1 strain did not exhibit cytotoxic activity against healthy skin fibroblast cells (cell viability 104.2±1.3%, p=0.2542). We demonstrated that tumor treatment with S. pyogenes GURSA1 significantly increased the lifespan of C57BL/6 mice with hepatoma (34 days, p=0.040) and pancreatic cancer (32 days, p=0.039) relative to the control groups (24 and 28 days, respectively). Increased lifespan was accompanied by a slowdown in tumor progression, as evidenced by a reduction in the growth of hepatoma and pancreatic cancer tumors under treatment with GAS strains in mice
Conclusions: Both S. pyogenes GUR and S. pyogenes GURSA1 strains demonstrated strong oncolytic activity against murine hepatoma 22a, pancreatic cancer PANC02 and human U251 glioma cells in vitro. In contrast, S. pyogenes GUR and GURSA1 didn`t show a toxicity against human normal skin fibroblasts. The overall survival rate and lifespan of mice treated with S. pyogenes GURSA1, a strain lacking the M protein on its surface, were significantly higher compared to the control and S. pyogenes GUR strain groups.
Keywords:
Streptococcus pyogenes
; recombinant M-protein
; human glioma U251
; pancreatic cancer PANC02
; mice hepatoma 22a
; cytotoxicity
; mice survival rate
1. Introduction
It is obvious that in recent years significant progress has been achieved in the field of cancer therapy, however, it continues to be one of the most common causes of death in developed countries. According to the forecasts of the World Health Organization (WHO), by 2040 the number of oncological diseases in the world will increase to 29.5 million people against 18.1 million in 2018 [1,2]. Conventional methods, such as radiation and chemotherapy, do not provide a complete cure, which necessitates the search for new alternative approaches, one of which is the tumors` treatment using microorganisms.
The study and use of bacterial strains with the ability to inhibit of cancer development has a fairly long history. Even long before radiation and chemotherapy, practicing physicians noticed that spontaneous healing of cancer cases often occurred against the background of a bacterial or viral infection accompanied by fever. Largely with the participation of research by American surgeon William Coley, it was proved that a number of bacteria, primarily Streptococci, have pronounced oncolytic properties. During his research, Coley managed to treat more than 1500 terminal patients with various cancer types with a high success rate [3,4].
The ability to cure cancer or make it a treatable chronic disease is a force that drives discovery and innovation in oncological microbial therapy. Bacterial-mediated cancer therapy (BMCT) may be considered as a new direction of antitumor therapy. Microbial therapy has the potential to solve many clinical problems that cannot be solved with current oncological methods. For example, BMCT may be efficient for the treatment of resistant metastatic cancers, and for diminishing the immunosuppressive tumor cells` effect. A significant advantage of microbial therapy is its specific effect on cancer cells and tissues. Thus, microbial therapy is well suited for the treatment of metastatic diseases, the main death cause from cancer [5]. For example, anaerobic strains of Salmonella sp., Bifidobacterium sp., Pseudomonas aeruginosa, Clostridium sp. and Streptococcus sp. are able to selectively migrate and colonize of tumor hypoxic niche [6,7]. In addition, these microorganisms can be genetically modified to reduce systemic toxicity and increase anticancer efficacy [8]. The use of such genetically modified bacteria can modify the expression of genes and proteins in cancer cells, which allows for a strong it dose-response dependence, increasing safety and simultaneously enhancing bacterial cells` anticancer activity [9].
To date, many preclinical studies on bacterial microbial therapy have shown a slowdown in cancer growth and an increase in animal survival [10,11]. А complete cancer regression has been achieved with oncolytic bacteria in immunocompetent animals with syngeneic cancers and in companion dogs with spontaneous tumors [12,13,14]. Attenuated live bacteria have satisfactory safety profiles in both healthy and tumors animals [15,16]. These microbial therapy studies led to the approval in 2015 by the US Food and Drug Administration (FDA) the first oncolytic drug (Imylgic T-VEC) based on herpes simplex virus-1 [17]. Imlygic's approval has fueled renewed interest in microbial therapy. In this regard, pharmacological and toxicological clinical studies in oncology patients have begun to be conducted intensively with bacterial strains [18,19].
Despite the successful use of bacteria, including group A Streptococci (GAS), for cancer treatment, the anticancer mechanisms of oncolytic bacteria are still not well understood [20]. These bacteria consume nutrients and secrete toxins and cytolytic agents (such as lipases and proteases), inducing apoptosis or autophagy in cancer cells [21]. In addition, bacteria stimulate leukocytes and T-lymphocytes to secrete immunomodulatory cytokines and chemokines that suppress the proliferation of cancer cells and prevent tumor recurrence. For example, bacteria and their lipopolysaccharide and flagellin components activate of toll-like receptor 4, CD14 or connexin 43 expression on cancer cells which contributes to it recognition by migrating dendritic cells, neutrophils and macrophages [21,22]. The flagellin of Salmonella sp. can activate cancer cell death through the production of IFN-γ by cytotoxic T-lymphocytes [23]. Salmonella typhimurium showed an anticancer effect against MDA-MB-435, MDA-MB-361, MDA-MB-231, 4T1, MCF-7 breast cancer, B16-F10 melanoma, U87MG glioblastoma, ASPC-1 pancreatic cancer, WiDr, CT26 colorectal carcinoma cells [24,25]. In addition, bacteria can serve as delivery tools for anticancer drugs into tumors [21]. Motile bacteria loaded with carrier cells had been designed to penetrate cancer areas inaccessible to standard drugs and small molecules [26].
It has also been shown that some streptococcal proteins, in particular, the enzymes arginine deiminase or streptokinase, can significantly slow down tumor cells` proliferation [27,28,29]. To date, the anticancer effect of bovicin, HC5 secreted by Streptococcus bovis, have been established in the context of milk gland adenocarcinoma and liver carcinoma cells [30]. The hyaluronidase of S. pyogenes H4489A bacteriophage (HylP) has an anticancer effect in MCF-7, MDA-MB-231, Hs578T breast cancer cells [31]. The OK-432 (picibanil), drug based on S. pyogenes, induces the death of lymphangioma cells [32]. Meanwhile, the anticancer activity of GAS strains remains unstudied in human colorectal, pancreatobiliary and brain tumors cells. Thus, GAS strains represent promising candidates for investigating cytotoxic mechanisms and effects against cancer cells, as well as for developing cellular vaccines and innovative approaches to anticancer therapy.
The current study explores the potential of oncolytic therapy using GAS strains, focusing on the effect of S. pyogenes GUR and its M-protein knockout mutant, S. pyogenes GURSA1, in a syngeneic mouse models of hepatoma and pancreatic carcinoma.
2. Materials and Methods
2.1. Streptococcus Pyogenes Strains
For the study four GAS strains with previously discovered oncolytic activities were chosen. S. pyogenes GUR (type emm111) is a throat isolate from a scarlet fever patient, which has been used clinically to treat cancer patients in the former Soviet Union for more than 20 years [50]. S. pyogenes GUR was kindly provided by prof. Chereshnev V.A., Perm State University (Perm, Russia). S. pyogenes GURSA1 is derivative of S. pyogenes GUR strain with an inactivated M-protein gene [29]. S. pyogenes GUR and S. pyogenes GURSA1 strains showed a cytotoxic activity against murine cancer cells [27]. The strains were cultivated in Todd-Hewitt broth (Condalab, Spain) for 16 h at 37 0C from single colonies. For all experiments 10⁶ CFU of S. pyogenes in Dulbecco's modified eagle medium (DMEM) or PBS was used. The optical density of overnight cultures was measured at 600 nm to estimate the bacterial cell count, based on previously established calibration curves. The cultures were then centrifuged at 6000× g for 5 minutes, washed once with PBS, and resuspended in DMEM or PBS to achieve a uniform bacterial concentration.
2.2. Obtaining M Protein Gene Knockout Mutant of S. Pyogenes GUR
To generate a knockout of the M protein gene (emm) in S. pyogenes GUR, we designed an integrative plasmid for homologous recombination. The plasmid was constructed using the pT7ermB vector, which contains part of emm gene necessary for integration into the S. pyogenes GUR chromosome. By using modified primers MF_Hind (acagcagaggcttaggcggaggatcattg) and MR_EcoR (atttcttaagatgctgccatagctt), the emm gene fragment was successfully amplified. The amplicons and the pT7ermB vector were digested with HindIII and EcoRI, and then ligated into the pT7ermB(emm111) vector. The resulting vector was used to transform S. pyogenes GUR. S. pyogenes GUR was grown to mid-log phase in Todd-Hewitt (ТН) broth (Condalab, Spain). The cells were made electrocompetent by washing twice with ice-cold 10% glycerol and resuspending them in 1/40 (by volume) of the same solution. Approximately 1 µg of the integrative (nonreplicative) plasmid was electroporated into the competent S. pyogenes cells using a standard electroporation protocol (1.8 kV, 25 µF, 200 Ω). Following electroporation, the cells were recovered in 1 mL of TH broth and incubated at 37°C for 1 hour to allow for plasmid integration in the bacterial chromosome. After recovery, the transformed cells were plated on selective media containing 2.5 µg/mL erythromycin to select for the integrants. Recombinants were allowed to grow for 24–48 hours at 37°C. Colonies that appeared on the selective media were picked and grown in TH broth with antibiotic selection for further analysis. To confirm the successful integration of the plasmid and knockout of the M protein gene PCR confirmation were conducted using primers ERM1 (gggcccaaaatttgtttga), ERM2 (tcggcagcgactcatagaat), and EMM (agtcgtaggagcagggttagc).
2.3. Cell Cultures
Human glioma U251, pancreas cancer PANC-02, and murine hepatoma MH-22a was obtained from the Russian collection of cell cultures (Saint-Petersburg, Russia). These cells were cultured at 1.0×106/mL in 25 cm2 flasks (T25 Nunclon, ThermoFisher Scientific Inc, USA) in RPMI-1640 and DMEM (Sigma-Aldrich, USA) containing 10% fetal bovine serum (FBS, Sigma-Aldrich, USA), 2mM glutamine (Sigma-Aldrich, USA) at 37 °C, 95% humidity and 5% CO2 for 1-2 days [33,34].
Normal human skin fibroblasts (NHF) were isolated following an aseptic surgical procedure. The skin sample was transferred to a 100 mm cell culture Petri dish using sterile forceps. The epidermis was oriented upwards, and the sample was gently spread using curved forceps. The skin was cut into small pieces and placed into a 50 mL tube. Then, 2.5 mL of dispase solution was added, and the mixture was incubated overnight at 4°C. Afterward, the dermis and epidermis were carefully separated using two curved forceps, and the dermis was incubated in collagenase solution for 60 minutes at 37°C in a 5% CO₂ atmosphere, with regular shaking. The digested tissue was then mixed with 15 mL of DMEM medium supplemented with 10% FBS in a 50 mL Falcon tube. The NHF suspension was centrifuged at 200 g for 5 minutes at room temperature. The resulting pellet was resuspended in 10 mL of complete DMEM medium, stained with trypan blue, and the cell count was determined using an automated cell counter. Approximately 1 × 10⁶ cells were seeded into an uncoated 25 cm² tissue culture flask in DMEM medium with 10% FBS and incubated at 37°C in a 5% CO₂ atmosphere and 95% humidity. Subculture was performed when cells reached 70–90% confluence [35].
2.4. МТТ Assay
The oncolytic activity of GAS against human glioma U251, pancreatic cancer PANC02, murine hepatoma MH-22a, and normal skin fibroblast cells was assessed using the MTT assay [36]. For this purpose, 1 × 10⁴ cells/well, suspended in 50 μL of RPMI-1640 or DMEM medium with 10% FBS, were seeded into the wells of 96-well flat-bottom plates (TPP, Switzerland) 1 day before GAS treatment and incubated for 24 hours at 37°C in a 5% CO₂ atmosphere. The following day, 10 μL of the GAS samples (GUR, GURSA1, and strains 7, 21 at 1.2 × 10⁶ and 2.5 × 10⁶ cells/mL) were added to 90 μL of medium in each well. Three replicates were performed for each GAS concentration. As a positive control, 100 μL of DMEM medium was added to the wells containing tumor cells instead of GAS. Negative controls were included, with 10 μL of Triton X-100 added to the cells, or empty wells. The final sample volume in each well was 100 μL. The plates were incubated for 20 hours in a CO₂ incubator. Then, 50 μL of resazurin dye was added to each well, and incubation continued until a color difference was observed between the positive and negative controls. The optical density (OD) of the solution in the wells was measured at a wavelength of 570 nm, subtracting the OD at 690 nm as the background, using a SpectraMax 250 plate reader (Molecular Devices, USA). Experimental data were collected using SoftMax Pro 5.2 software (Molecular Devices, USA). Based on the collected data, the cytotoxicity of GAS samples was determined. The percentage of dead cells was calculated by comparing the OD of the samples with the positive (100% viable cells) and negative (0% viable cells) controls.
2.5. Real-Time Cytotoxicity Analysis by Using RT xCELLigence System
For real-time detection of GAS oncolytic activity against pancreatic cancer PANC02, glioma U251, MH-22a, and NHF cells, the RT xCELLigence system (Agilent, USA) with E-Plate L8 was used [38]. The RT xCELLigence system records changes in the electronic impedance (resistance) of a specially designed plate containing embedded gold micro-electrodes. The change in resistance over time allows the generation of a graph showing the functional dependence of the cell index on time. On Day 0, 150 µL of medium and 150 µL of a cell suspension containing 50,000–200,000 cells per well (U251, PANC02, and MH-22a) were added to each well of the E-Plate L8. The method was standardized by measuring the OD of the cell suspension (OD = 0.200 ± 0.010) using a spectrophotometer at a wavelength of 600 nm. The loaded plates were incubated at 5% CO₂ in the RT xCELLigence system for 24 hours. After this period, overnight cultures of GAS strains (at 10⁶ CFU in 300 mL of DMEM) were added. Cancer cells were then cultured for an additional day, with the electrical resistance of the cell monolayer recorded every minute. The GAS strains (GUR, GURSA1, and strains 7, 21) were washed in PBS and resuspended in DMEM to obtain an equal final concentration.
2.6. Oncolytic Effects of S. pyogenes GUR and GURSA1 Strains on Hepatoma 22a and Pancreatic Cancer PANC02 Growth in Mouse Models
To establish a mouse hepatoma model, we used hepatoma 22A cells, which were harvested from the culture flask, resuspended in physiological solution at a concentration of 105 cells/mL, and subcutaneously inoculated into the right flank of C57BL/6 mice (n = 66, Rappolovo Nursery, Russia) under short-term ether anesthesia. The study used female C57BL/6 mice weighing 16-18 g. The animals were maintained under a 12-hour light/12-hour dark cycle and were provided with standard food (concentrated combined feed in the form of briquettes) and water ad libitum. Tumor size was measured 10 days post-injection, with an average size of 2.5 ± 0.5 mm³. The mice were then divided into three groups: GUR (n = 22) and GURSA1 (n = 22) groups received intratumoral injections of S. pyogenes GUR and S. pyogenes GURSA1 cells, respectively, at a dose of 106 cells in 50 μL of suspension. The Control group (n = 22) received no treatment.
Similarly, a C57BL/6 mouse model with PANC02 cancer cells was established according to the protocol described above. When the tumors reached a size of 5 ± 0.5 mm³ (18 days post-injection), the mice were divided into the same groups: GUR (n = 8), GURSA1 (n = 8), and Control (n = 7). The survival of all animals was monitored for 40 days. At the end of the experiment, the animals were euthanized by an overdose of ether anesthesia.
2.7. Studying of GAS Migration in PANC02 Cancer C57BL/6 Mouse Model
Using the PANC02 cancer C57BL/6 mouse model described above, we studied the ability of S. pyogenes GURSA1 to migrate into the tumor site following intraperitoneal administration. On day 18 after tumor cell inoculation, when tumors had reached a size of 5 ± 0.5 mm³, one group (n = 6) was intraperitoneally infected with 106 CFU of S. pyogenes GURSA1 in a volume of 50 μL dissolved in PBS. Animals that received an equivalent volume of PBS alone served as controls (n = 6). Mice were sacrificed one day after administration of S. pyogenes GURSA1. At the end of each experiment, tumor, spleen, and liver samples were homogenized in 500 μL of PBS using a vibrating ball mill (MM 400, Retsch, USA), and tenfold serial dilutions were plated on Petri dishes containing Columbia blood agar (Conda, Spain). The seeded dishes were incubated at 37°C for 24 hours. Colonies of microorganisms were identified using mass spectrometric analysis on a Bactoscreen device (Litech, Russia).
2.8. Acute oral Toxicity
The acute oral toxicity test was performed using C57BL/6 mice, aged 8-10 weeks, weighing between 18.6 and 22.7 g. The mice were allowed free access to food and water, except for 4 hours prior to the experiment. Four groups of female mice were divided by strain (S. pyogenes GUR and S. pyogenes GURSA) and administered doses (10⁶ or 10⁸ CFU/mouse). The GAS strains were administered intragastrically at a volume of 500 µl. Clinical signs, changes in body weight, and necropsy findings were investigated during the test period of 14 days.
2.9. Ethics Statement
All animal experiments were conducted in accordance with the “Rules of Laboratory Practice” (Ministry of Health of the Russian Federation, No. 708). The study was approved by the Local Ethics Committee for Animal Care and Use at the Institute of Experimental Medicine, Saint Petersburg, Russia.
2.10. Statistical Analysis
All experiments were performed in triplicate. Statistical significance between the means of different treatments and their respective control groups was determined using the Student's t-test. Data are presented as means ± standard deviation and were considered significant at p<0.05. For comparisons between two independent groups with a small sample size (n<30), the nonparametric Mann-Whitney U-test was used [39]. Descriptive statistics were performed using GraphPad Prism (version 6.01, 09.21.2012).
3. Results
3.1. Studying In vitro the Oncolytic Effects of GUR, GURSA1 Strains Against Hepatoma 22a, Pancreatic Cancer PANC02 and U251 Glioma Cells
The M protein of S. pyogenes is a major virulence factor, responsible for multiple functions such as adherence to host cells, immune evasion, and promoting bacterial survival in the host. The gene encoding the M protein is part of the emm gene family, and disruption or knockout of this gene significantly impacts the pathogenicity of S. pyogenes. Generating an M protein gene knockout mutant allowed the creation of a less virulent variant of S. pyogenes GUR, enabling the investigation of the role of this protein in the oncolytic activity of the S. pyogenes GUR strain. The M protein knockout mutant was successfully created according to the methodology described above and was named S. pyogenes GURSA1. The whole-genome sequences of S. pyogenes GUR and S. pyogenes GURSA1 are available from our previous work [29]. Interestingly, genomic study of the S. pyogenes GUR strain, belonging to M111 serotype, revealed a stop codon in the other gene of mga regulon – mrp, making the entire regulon nonfunctional.
As the first step of the study we tested the oncolytic activities of S. pyogenes GUR and S. pyogenes GURSA1 strains using MTT assay against murine hepatoma 22a, pancreatic cancer PANC02 and human U251 glioma cells (Figure 1).
The results in Figure 1 show the viability of cancer cells under exposure to live GAS strains, as measured by changes in metabolic activity according to the MTT assay. The live S. pyogenes GUR strain exerts a strong cytotoxic effect against pancreatic cancer PANC02 cells, resulting in a decrease in metabolic activity by up to 67.4±1.9%. However, against glioma and hepatoma cells, the GUR strain exhibited moderate (38.0±1.8%) and weak (16.3 ±5.4%) oncolytic activity, respectively. In contrast, the S. pyogenes GURSA1 strain exhibited strong (86.5±1.6%) and moderate (36.5±1.8%) oncolytic activity against glioma and hepatoma cells, respectively, but was ineffective against pancreatic cancer cells (5.5±2.1%). However, the oncolytic activity of S. pyogenes GURSA1 was significantly stronger than that of S. pyogenes GUR against U251 glioma (p=0.000004) and 22a hepatoma (p=0.003625) cells. In contrast, the oncolytic activity of S. pyogenes GUR was significantly stronger (p=0.000014) than that of S. pyogenes GURSA1 against PANC02 pancreatic cancer cells.
As the MTT assay measures cancer cell viability indirectly by assessing changes in metabolic activity, we also investigated the oncolytic activity of S. pyogenes GUR and S.pyogenes GURSA1 strains against 22a hepatoma, PANC02 pancreatic cancer, and U251 glioma cells using the RT xCelligence system. RT xCelligence system allows for the direct observation of the number of viable cells during GAS exposure. (Figure 2).
The real-time activity results in Figure 2 also show that S. pyogenes GUR exerted moderate to strong oncolytic effects against glioma (p=0.0495), hepatoma (p<0.01), and pancreatic cancer cells (p<0.01) over 48 hours. In addition, S. pyogenes GURSA1 exerted moderate to strong oncolytic effects against hepatoma (p<0.05), pancreatic cancer (p<0.05), and glioma cells (p=0.0080), respectively.
3.2. Studying In vitro the Effects of S. pyogenes GUR, S. pyogenes GURSA1 Strains Against Normal Skin Fibroblast Cells
In the next step, we investigated the oncolytic activity of S. pyogenes GUR and S. pyogenes GURSA1 on healthy skin fibroblast cells using the MTT assay and RT xCELLigence system (Figure 4).
The results of the MTT assay in Figure 4 indicate that the S. pyogenes GUR strain has a weak cytotoxic effect (6.9±3.8% cytotoxicity, cell viability 93.1±3.7%, p<0.05) on skin fibroblast cells. In contrast, the S. pyogenes GURSA1 strain does not exhibit cytotoxic activity (cell viability 104.2±1.3%, p=0.2542) against healthy fibroblast cells.
3.3. Studying In vivo the Oncolytic Effects of S. pyogenes GUR and GURSA1 Strains Against Hepatoma 22a, and Pancreatic cancer PANC02 Cells
We also investigated the oncolytic effect of S. pyogenes GUR and GURSA1 strains on the overall survival of mice with hepatoma 22a and PANC02 pancreatic cancer (Figure 5).
The results in Figure 5 show that the inoculation of the S.pyogenes GURSA1 significantly increased the lifespan of C57BL/6 mice with hepatoma (median survival 34 days) and pancreatic cancer (median survival 32 days) compared to the control groups (median survival 24 and 28 days, respectively). However, the application of S. pyogenes GUR shortened the lifespan of mice with both hepatoma (median survival 14 days) and pancreatic cancer (median survival 22 days) compared to the control group.
An increase in the lifespan of mice with cancers treated with S. pyogenes GUR and S. pyogenes GURSA1 strains was accompanied by slower tumor growth of hepatoma 22A and PANC02 pancreatic cancer compared to the control group (Figure 6).
Although treatment of tumors with S. pyogenes GUR led to a slowdown in tumor progression, which was more pronounced in the pancreatic cancer model, it did not result in an increase in the animals' lifespan. Moreover, it led to a reduction in the average lifespan of mice in the S. pyogenes GUR treatment groups in both models. In contrast, treatment with S. pyogenes GURSA1 significantly prolonged the animals' lifespan and inhibited tumor progression in both models.
Finally, using the C57BL/6 mice pancreatic cancer model, we studied the ability of S. pyogenes GURSA1 to migrate to the tumor site following intraperitoneal administration (Table 1). As a result, we demonstrated that the S. pyogenes GURSA1 strain selectively colonized tumors 24 hours after intraperitoneal administration (10⁶ CFU/mouse). Notably, S. pyogenes GURSA1 was completely absent from both the spleen and liver of the mice, highlighting its selective tumor targeting
The ability of oncolytic bacteria to selectively accumulate in tumors without colonizing normal tissues and organs is one of their most important characteristics. This property enables the development of genetic constructs based on oncolytic bacteria for targeted delivery of anticancer drugs to solid tumors.
3.4. Studying of Acute Oral Toxicity of S. pyogenes GUR and GURSA1 Strains
Acute oral toxicity was assessed using a two-dose acute oral toxicity test. The doses were selected based on the potential therapeutic dose — 106 CFU per mouse — and a 100-fold higher dose, 108 CFU per mouse. No fatalities or clinical signs were observed in the mice throughout the study. There were no significant changes in body weight in the rats administered S. pyogenes strains (Table 2). Furthermore, no abnormal findings were noted during necropsy. These results suggest that S. pyogenes GUR and S. pyogenes GURSA1 did not adversely affect the health of the mice.
4. Discussion
Using in vitro tools, we established that live S. pyogenes GUR and GURSA1 strains have a strong anticancer effect against human glioma U251, pancreatic cancer PANC02, and mouse hepatoma 22a cells by MTT assay and the RT xCelligence system. At the same time, it was determined that both strains exhibited low toxicity on human normal skin fibroblasts (Figure 4). Acute oral toxicity test also demonstrated low overall strains toxicity: administration of S. pyogenes GUR and S. pyogenes GURSA1 even in 100-fold therapeutic dose did not adversely affect the health of the mice. In our previous publication we also demonstrated that some GAS strains, especially S. pyogenes GURSA1, stimulated the survival of human perifocal brain tissue cells [37].
It is known that the M protein is the main surface-associated virulence factor of GAS and a key antigen target of host immunity [42]. The previously created M protein knockout mutant S. pyogenes GURSA1 was expected to have reduced oncolytic properties, but also lower overall toxicity compared to the wild-type S. pyogenes GUR strain. Nevertheless, the in vitro experiments showed that both strains exhibited comparable cytotoxic activity against tumor cells, which varied in degree across different cell lines.
In contrast to our expectations, in vivo experiments demonstrated that S. pyogenes GURSA1, but not S. pyogenes GUR, significantly increased the lifespan of C57BL/6 mice with hepatoma (34 days, p=0.040) and pancreatic cancer (32 days, p=0.039), compared to the control groups (24 and 28 days, respectively). This effect was accompanied by a slowdown in tumor progression in both hepatoma and pancreatic cancer models. The application of S. pyogenes GUR also delayed tumor development in mice, but it shortened the animals' lifespan compared to the control group in both tumor models.
5. Conclusions
Our results allow us to conclude that live S. pyogenes GUR and S. pyogenes GURSA1 strains exhibited strong oncolytic activity against murine hepatoma 22a, pancreatic cancer PANC02, and human U251 glioma cells in vitro. In contrast, both S. pyogenes GUR and S. pyogenes GURSA1 strains showed no toxicity against human normal skin fibroblasts, and no acute toxic effects were observed in in vivo model studies. The overall survival rate and lifespan of mice treated with S. pyogenes GURSA1 strain were significantly higher compared to both the control and S. pyogenes GUR groups. Using the C57BL/6 mouse model of pancreatic cancer, we demonstrated that the oncolytic S. pyogenes GURSA1 strain selectively colonized tumors 24 hours after intraperitoneal administration, while the strain was not detected in the spleens or livers of the animals. Further investigations are required to understand the role of the M-protein in the oncolytic activity of S. pyogenes. Future studies will focus on examining the unique features of the oncolytic GAS genomes, in contrast to those of bacteria lacking cytotoxic activity against cancer cells. Additionally, we aim to identify bacterial structures that contribute most significantly to the oncolytic activity of GAS strains.
Author Contributions
A.T., A.C. performed the investigations, wrote the manuscript; N.D., A.M., M.S. performed the investigations, manuscript corrections, K.B., T.F., E.E. performed the investigations; E.G. participated the investigations, statistical analysis, manuscript correction; A.S. manuscript correction, supervision and project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science and Higher Education of the Russian Federation, №075-15-2022-302 (20.04.2022), additional agreement 075-15-2022-302/9 (14.02.2024).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, LA.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (Globocan) World Health Organization. Available online: https://gco.iarc.fr/tomorrow/home (accessed on 18.09.2024).
- Nauts, H.C.; Fowler, G.A.; Bogatko, F.H. A review of the influence of bacterial infection and of bacterial products (Coley's toxins) on malignant tumors in man; a critical analysis of 30 inoperable cases treated by Coley's mixed toxins, in which diagnosis was confirmed by microscopic examination selected for special study. Acta Med Scand Suppl. 1953, 276, 1–103. [Google Scholar] [PubMed]
- Coley, W.B. Treatment of malignant tumors by repeated inoculation of erysipelas: with a report of 10 cases. Am. J. Med. Sci. 1893, 105, 487–564. [Google Scholar] [CrossRef]
- Forbes, N.S.; Coffin, R.S.; Deng, L.; Evgin, L. , Fiering, S., Giacalone, M.; Gravekamp, C.; Gulley, J.L.; Gunn, H.; R.M.; Hoffman, R.M. et al. White paper on microbial anti-cancer therapy and prevention. J. Immunotherapy Cancer 2018, 6, 78. [Google Scholar] [CrossRef]
- Yoshida, J.; Takamura, S.; Ishibashi, T.; Nishio, M. Antiproliferative and apoptosis-inducing effects of an antitumor glycoprotein from Streptococcus pyogenes. Anticancer Res 1999, 19, 1865–71. [Google Scholar]
- Fiedler, T.; Strauss, M.; Hering, S. , et al. Arginine deprivation by arginine deiminase of Streptococcus pyogenes controls primary glioblastoma growth in vitro and in vivo. Cancer Biol Ther. 2015, 16, 1047–55. [Google Scholar] [CrossRef]
- Bettegowda, C.; Huang, X.; Lin, J.; Cheong, I.; Kohl,i M. ; et al. The genome and transcriptomes of the anti-tumor agent Clostridium novyi-NT. Nat. Biotechnol 2006, 24, 1573–80. [Google Scholar] [CrossRef]
- Brown, J.M.; Wilson, W.R. Exploiting tumour hypoxia in cancer treatment. Nat Rev Cancer 2004, 4, 4,437–47. [Google Scholar] [CrossRef]
- Feng, X.; He, P.; Zeng, C.; Li, Y.H.; Das, S.K.; Li, B.; Yang, H.F.; Du,Y. Novel insights into the role of Clostridium novyi-NT related combination bacteriolytic therapy in solid tumors. Oncol Lett 2021, 21, 110. [Google Scholar] [CrossRef]
- Chen, J.; Yang, B.; Cheng, X.; Qiao, Y.; Tang, B.; Chen, G.; Wei, J.; Liu, X.; Cheng, W.; Du, P.; et al. Salmonella-mediated tumor-targeting TRAIL gene therapy significantly suppresses melanoma growth in mouse model. Cancer Sci 2012, 103, 325–33. [Google Scholar] [CrossRef]
- Roberts, N.J.; Zhang, L.; Janku, F.; Collins, A.; Bai, R.Y.; Staedtke, V.; Rusk, A.W.; Tung, D.; Miller, M.; Roix, J.; et al. Intratumoral injection of Clostridium novyi-NT spores induces antitumor responses. Sci Transl Med 2014, 6, 249ra111. [Google Scholar] [CrossRef] [PubMed]
- Gentschev, I.; Petrov, I.; Ye, M.; Kafuri Cifuentes, L.; Toews, R.; Cecil, A.; Oelschaeger,T. A.; Szalay, A.A. Tumor Colonization and Therapy by Escherichia coli Nissle 1917 Strain in Syngeneic Tumor-Bearing Mice Is Strongly Affected by the Gut Microbiome. Cancers (Basel) 2022, 14, 6033. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Lin, Y.; Zhao, Z.; Lai, Y.; Lu, M.; Shao, Z.; Mo, X.; Mu, Y.; Liang, Z.; Wang, X.; et al. Targeted deprivation of methionine with engineered Salmonella leads to oncolysis and suppression of metastasis in broad types of animal tumor models. Cell Rep Med 2023, 4, 101070. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, N.; Zhao, M.; Hoffman, R.M. Comparison of the selective targeting efficacy of Salmonella typhimurium A1-R and VNP20009 on the Lewis lung carcinoma in nude mice. Oncotarget 2015, 6, 14625–31. [Google Scholar] [CrossRef]
- Diaz, L.A.Jr.; Cheong, I.; Foss, C.A.; Zhang, X.; Peters, B.A.; Agrawal, N.; Bettegowda, C.; Karim, B.; Liu, G.; Khan, K.; et al. novyi-NT spores. Toxicol Sci 2005, 88, 562–75. [Google Scholar] [CrossRef]
- Pol, J.; Kroemer, G.; Galluzzia, L. First oncolytic virus approved for melanoma immunotherapy. Oncoimmunol. 2016, 5, e1115641. [Google Scholar] [CrossRef]
- Alexander, L.M.; van Pijkeren, J-P. Modes of therapeutic delivery in synthetic microbiology. Trends Microbiol 2023, 31, 197–211. [Google Scholar] [CrossRef]
- Janku, F.; Zhang, H.H.; Pezeshki, A.; Goel, S.; Murthy, R.; Wang-Gillam, A.; Shepard, D.R.; Helgason, T.; Masters, T.; Hong, D.S.; et al. Intratumoral Injection of Clostridium novyi-NT Spores in Patients with Treatment-refractory Advanced Solid Tumors. Clin Cancer Res 2021, 27, 96–106. [Google Scholar] [CrossRef]
- Marzhoseyni, Z.; Shojaie, L.; Tabatabaei, S.A.; Movahedpour, A.; Safari, M.; Esmaeili, D.; Mahjoubin-Tehran, M.; Jalili, A.; Morshedi, K.; Khan, H.; et al. Streptococcal bacterial components in cancer therapy. Cancer Gene Ther 2022, 29, 141–155. [Google Scholar] [CrossRef]
- Uchugonova, A.; Zhang, Y.; Salz, R.; Liu, F.; Suetsugu, A.; Zhang, L.; Koenig, K.; Hoffman, R.M.; Zhao, M. Imaging the different mechanisms of prostate cancer cell killing by tumor-targeting Salmonella typhimurium A1-R. Anticancer Res 2015, 35, 5225–9. [Google Scholar]
- Wang, D.; Wei, X.; Kalvakolanu, D.V.; Guo, B.; Zhang, L. Perspectives on Oncolytic Salmonella in Cancer Immunotherapy-A Promising Strategy. Front Immunol 2021, 12, 615930. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Hwang, I.; Lee, E.; Shin, S.J.; Lee, E.J.; Rhee, J.H.; Yu, J.W. Bacterial Outer Membrane Vesicle-Mediated Cytosolic Delivery of Flagellin Triggers Host NLRC4 Canonical Inflammasome Signaling. Front Immunol 2020, 11, 581165. [Google Scholar] [CrossRef] [PubMed]
- Mi, Z.; Feng, Z.-C.; Li, C.; Yang, X.; Ma, M.-T.; Rong, P.-F. Salmonella-Mediated Cancer Therapy: An Innovative Therapeutic Strategy. J Cancer 2019, 10, 4765–4776. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubi, A.; Khazaei, M.; Hasanian, S.M.; Avan, A.; Cho, W.C.; Soleimanpour, S. Bacteriotherapy in Breast Cancer. Int J Mol Sci 2019, 20, 5880. [Google Scholar] [CrossRef] [PubMed]
- Ganai, S.; Arenas, R.B.; Sauer, J.P.; Bentley, B.; Forbes, N.S. In tumors Salmonella migrate away from vasculature toward the transition zone and induce apoptosis. Cancer Gene Ther 2011, 18, 457–66. [Google Scholar] [CrossRef]
- Suvorova, M.A.; Kramskaya, T.A.; Duplik, N.V.; Chereshnev, V.A.; Grabovskaya, K.B.; Ermolenko, E.I.; et al. The effect of inactivation of the M-protein gene on the antitumor properties of live Streptococcus pyogenes in experiment. Voprosi oncologii 2017, 63, 803–807. [Google Scholar] [CrossRef]
- Freidlin, I.S.; Starikova, E.A.; Lebedeva, A. Overcoming the protective functions of macrophages by Streptococcus pyogenes virulence factors. Bulletin of Siberian Medicine 2019, 18, 109–118. [Google Scholar] [CrossRef]
- Suvorova, M.; Tsapieva, A.; Bak, E.; Chereshnev, V.; Kiseleva, E.; Suvorov, A.; Arumugam, М. Complete genome sequences of emm111 type Streptococcus pyogenes strain GUR with anti-tumor activity and it’s derivate strain GURSA1 with inactivated emm gene. Genome Announcements. 2017, 5, e00939–e17. [Google Scholar] [CrossRef]
- Huang, F.; Teng, K.; Liu, Y.; Cao Y, Wang T, Ma C, Zhang, J. , Zhong, J. Bacteriocins: Potential for Human Health. Oxid Med Cell Longev 2021, 2021, 5518825. [Google Scholar] [CrossRef]
- Lee, J.H.; Moore, L.; Kumar, S.; DeLucas, L.; Pritchard, D.; Ponnazhagan, S.; Deivanayagam, C. In vitro studies on anti-cancer effect of Streptococcus pyogenes phage hyaluronidase (HylP) on breast cancer. Cancer Res 2008, 68, 1506. [Google Scholar]
- Łukasiewicz, K.; Fol, M. Microorganisms in the treatment of cancer: advantages and limitations. J Immunol Res 2018, 2018, 2397808. [Google Scholar] [CrossRef] [PubMed]
- Freshney, R.I.; Griffiths, B.; Hay, R.J.; Reid, Y.A.; Carmiol, S.; Kunz-Schugart, L.; et al. Animal Cell Culture: a Practical Approach. Masters, J.R.W, editor. 3rd ed. London: Oxford Univ. Press, 2000.
- Schiedlauske, K.; Deipenbrock, A.; Pflieger, M.; Hamacher, A.; Hänsel, J.; Kassack, M.U.; Kurz, T.; Teusch, N.E. Novel Histone Deacetylase (HDAC) Inhibitor Induces Apoptosis and Suppresses Invasion via E-Cadherin Upregulation in Pancreatic Ductal Adenocarcinoma (PDAC). Pharmaceuticals (Basel). 2024, 17, 752. [Google Scholar] [CrossRef] [PubMed]
- Kisiel, M.A.; Klar, A.S. Isolation and Culture of Human Dermal Fibroblasts. In: Böttcher-Haberzeth, S., Biedermann, T. eds.; Skin Tissue Engineering. Methods in Molecular Biology, vol. 1993; Humana, New York, NY, 2019. [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Chernov, A.N.; Tsapieva, A.N.; Alaverdian, D.A.; Filatenkova, T.A.; Galimova, E.S.; Suvorova, M.; Shamova, O.V.; Suvorov, A.N. In vitro evaluation of cytotoxic effect of Streptococcus pyogenes strains, Protegrin PG-1, Cathelicidin LL-37, Nerve Growth Fac-tor and chemotherapy on C6 glioma cell line. Molecules 2022, 27, 569. [Google Scholar] [CrossRef]
- Junka, A.F.; Janczura, A.; Smutnicka, D.; Mączyńska, B.; Anna, S.; Nowicka, J.; Bartoszewicz, M.; Gościniak, G. Use of the Real Time xCelligence System for Purposes of Medical Microbiology. Pol J Microbiol 2012, 61, 191–197. [Google Scholar] [CrossRef]
- van Belle, G. , Fisher, L.D.; Heagerty, P.J., Lumley, T. Biostatistics: a methodology for the health sciences. Fisher, L.D., van Belle, G., Eds.; editors. Canada: Jonh Wiley and Sons Inc, 2004. [Google Scholar]
- Higuchi, Y. Antitumor Effect of Streptococcus pyogenes by Inducing Hydrogen Peroxide Production. Jpn J Cancer Res 1996, 87, 1271–1279. [Google Scholar] [CrossRef]
- Suvorova, M.; Tsapieva, A.; Suvorov, A.; Kiseleva, E. A99: The influence of live Streptococcus pyogenes on the growth of solid murine tumors. European J. of Cancer Supplements. 2015, 13, 59. [Google Scholar] [CrossRef]
- Ghosh, P. ; Variation, Indispensability, and Masking in the M protein. Trends Microbiol 2018, 26, 132–144. [Google Scholar] [CrossRef]
- McMillan, D.J.; Drèze, P.-A.; Vu, T.; Bessen, D.E.; Guglielmini, J.; Steer, A.C.; Carapetis, J.R.; Van Melderen, L.; Sriprakash, K.S.; Smeesters, P.R. Updated model of group A Streptococcus M proteins based on a comprehensive worldwide study. Clin Microbiol Infect 2013, 19, E222–9. [Google Scholar] [CrossRef]
- Facklam, R.F.; Martin, D.R.; Lovgren, M.; Johnson, D.R.; Efstratiou, A.; Thompson, T.A.; Gowan, S.; Kriz, P.; Tyrrell, G.J.; Kaplan. E.; Beall, B. Extension of the Lancefield classification for group A streptococci by addition of 22 new M protein gene sequence types from clinical isolates: emm103 to emm124. Clin Infect Dis 2002, 34, 28–38. [Google Scholar] [CrossRef]
- Lannergard, J.; Gustafsson, M.C.U.; Waldemarsson, J.; Norrby-Teglund, A.; Stålhammar-Carlemalm, M.; Lindahl, G. The hypervariable region of Streptococcus pyogenes M protein escapes antibody attack by antigenic variation and weak immunogenicity. Cell Host Microbe 2011, 10, 147–57. [Google Scholar] [CrossRef] [PubMed]
- Buffalo, C.Z.; Bahn-Suh, A.J.; Hirakis, S.P.; Biswas, T.; Amaro, R.E.; Nizet, V.; Ghosh, P. Conserved patterns hidden within group A Streptococcus M protein hypervariability recognize human C4b-binding protein. Nat Microbiol 2016, 1, 16155. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Takano, S.; Tomizawa, S.; Miyahara, Y.; Furukawa, K.; Takayashiki, T.; Kuboki, S.; Takada, M.; Ohtsuka, M. C4b-binding protein α-chain enhances antitumor immunity by facilitating the accumulation of tumor-infiltrating lymphocytes in the tumor microenvironment in pancreatic cancer. J Exp Clin Cancer Res 2021, 40, 212. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-H.; Yu,Y. P.; Zuo, Z.-H.; Nelson, J.B.; Michalopoulos, G.K.; Monga, S.; Liu, S.; Tseng, G.; Luo, J-H. Targeting genomic rearrangements in tumor cells through Cas9-mediated insertion of a suicide gene. Nat Biotechnol 2017, 35, 543–550. [Google Scholar] [CrossRef]
- Henderson, E.A.; Ivey, A.; Choi, S.; Santiago, S.; McNitt, D.; Liu, T.W.; Lukomski, S.; Boone, B.A. Group A Streptococcal Collagen-like Protein 1 Restricts Tumor Growth in Murine Pancreatic Adenocarcinoma and Inhibits Cancer-Promoting Neutrophil Extracellular Traps. BioRxiv. Preprint 2024, Jan 19. [CrossRef]
- Chereshnev, V.A.; Morova, A.A.; Ryamzina, I.N. Biological Laws and Human Viability: Method of Multifunctional Restorative Biotherapy. 2rd ed. Perm: Perm State Agricultural Academy Press, 2006, 215 p.
- Suvorov, A.N.; Polyakova, E.M.; McShan, W.M.; Ferretti, J.J. Bacteriophage content of M49 strains of Streptococcus pyogenes. FEMS Microbiol Lett. 2009, 294, 9–15. [Google Scholar] [CrossRef]
Figure 1.
The oncolytic activity of live S. pyogenes GUR, S. pyogenes GURSA1 against glioma U251, murine hepatoma 22a and pancreatic cancer PANC02 cells using MTT assay. **, **** statistically significant (p<0.01, p<0.0001) differences between the activities of the strains within each tumor cell type.
Figure 1.
The oncolytic activity of live S. pyogenes GUR, S. pyogenes GURSA1 against glioma U251, murine hepatoma 22a and pancreatic cancer PANC02 cells using MTT assay. **, **** statistically significant (p<0.01, p<0.0001) differences between the activities of the strains within each tumor cell type.
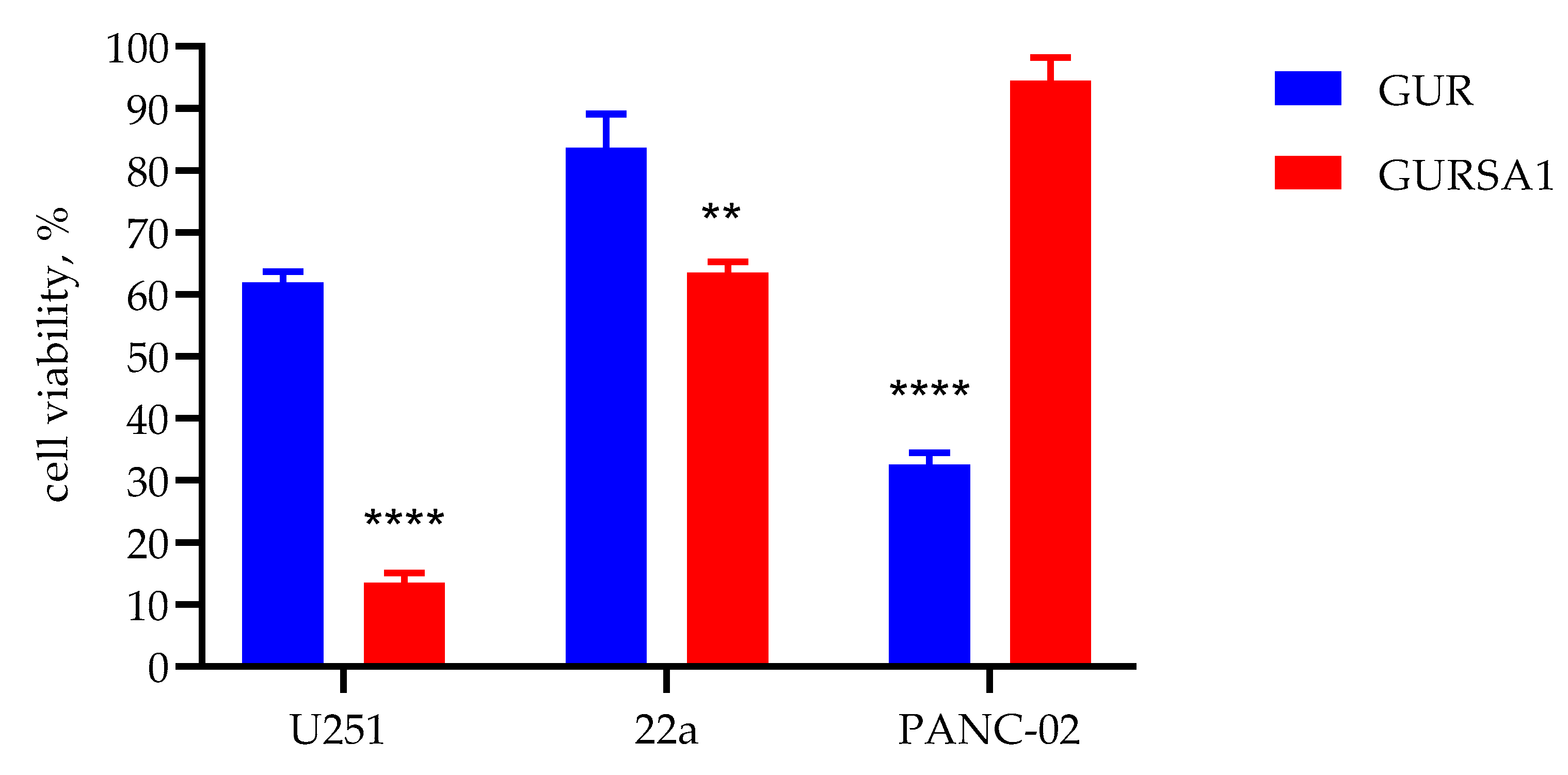
Figure 2.
The oncolytic activity of S. pyogenes GUR, S. pyogenes GURSA1 against (A) murine hepatoma 22a, (B) pancreatic cancer PANC02 and (C) U251 glioma cells using RT xCELLigence system. *, ** statistically significant (p<0.05, p<0.01) differences between the activities of the strains within each tumor cell type.
Figure 2.
The oncolytic activity of S. pyogenes GUR, S. pyogenes GURSA1 against (A) murine hepatoma 22a, (B) pancreatic cancer PANC02 and (C) U251 glioma cells using RT xCELLigence system. *, ** statistically significant (p<0.05, p<0.01) differences between the activities of the strains within each tumor cell type.
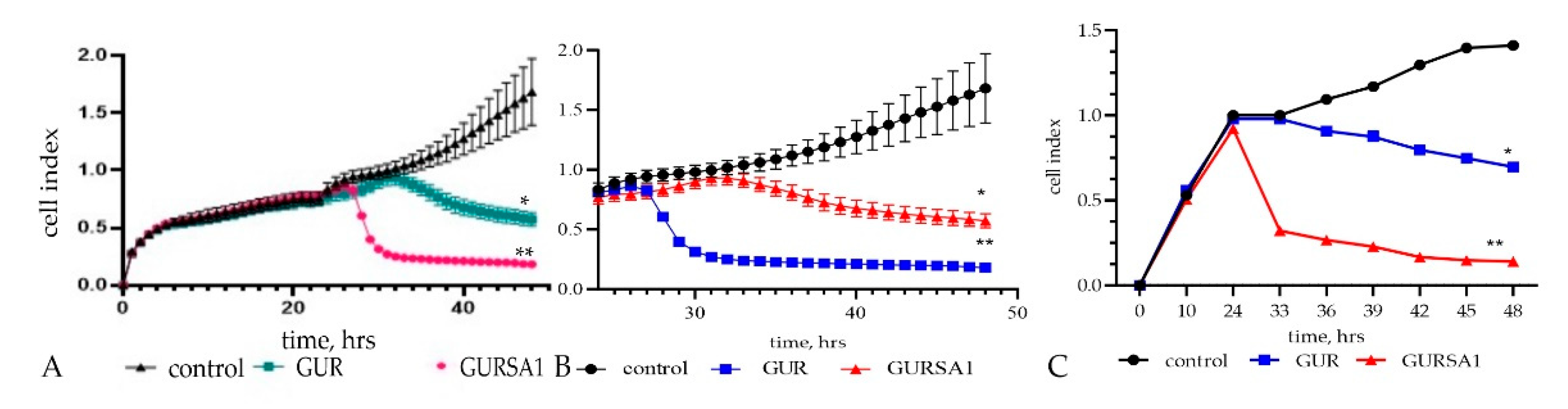
Figure 4.
The oncolytic activity of live S. pyogenes GUR, S. pyogenes GURSA1 against healthy skin fibroblast cells using (A) the MTT assay and (B) RT xCELLigence system. *statistically significant (p<0.05) difference between the activity of S. pyogenes GUR and control.
Figure 4.
The oncolytic activity of live S. pyogenes GUR, S. pyogenes GURSA1 against healthy skin fibroblast cells using (A) the MTT assay and (B) RT xCELLigence system. *statistically significant (p<0.05) difference between the activity of S. pyogenes GUR and control.
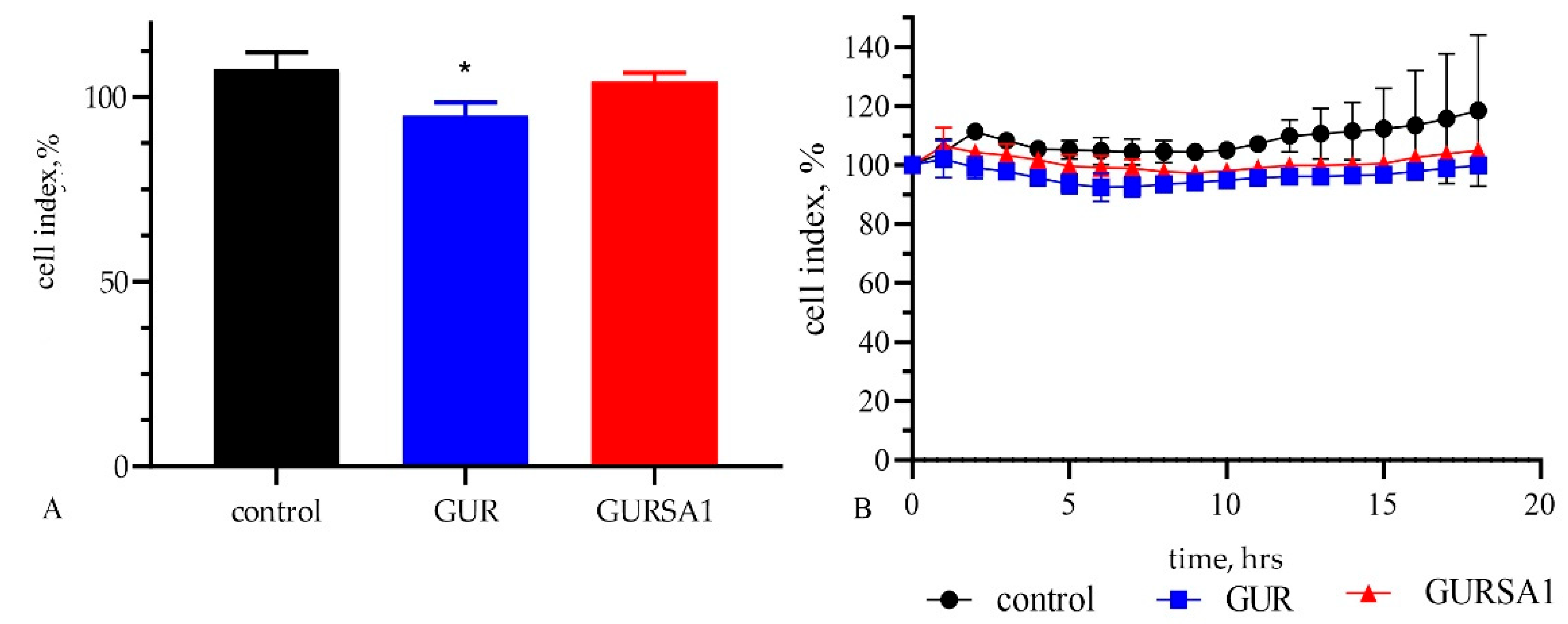
Figure 5.
The influence of live S. pyogenes GUR and S. pyogenes GURSA1 strains on C57BL/6 mice overall survival with (A) murine hepatoma, and (B) pancreatic cancer. 1- control, 2 - S. pyogenes GUR and 3 - S. pyogenes GURSA1. * statistically significant (p<0.05) increase in mice lifespan compared to control.
Figure 5.
The influence of live S. pyogenes GUR and S. pyogenes GURSA1 strains on C57BL/6 mice overall survival with (A) murine hepatoma, and (B) pancreatic cancer. 1- control, 2 - S. pyogenes GUR and 3 - S. pyogenes GURSA1. * statistically significant (p<0.05) increase in mice lifespan compared to control.
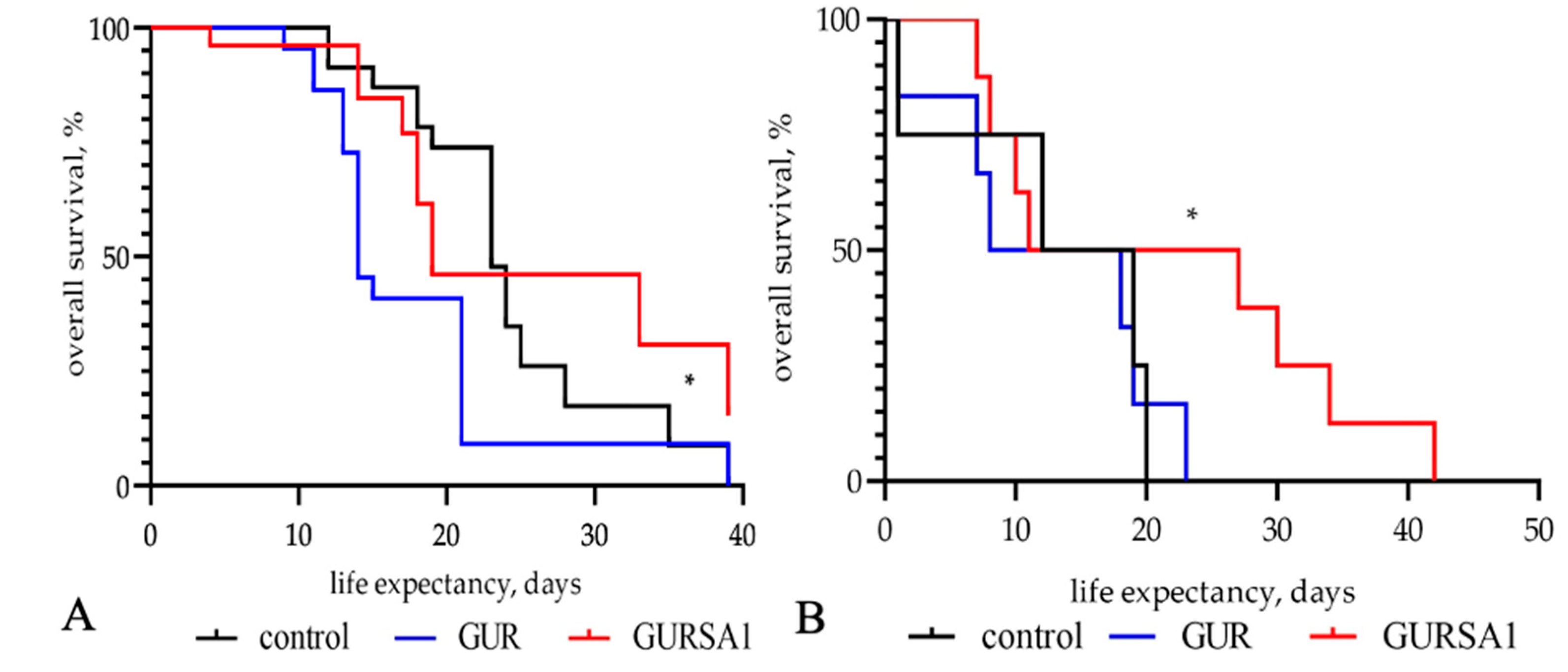
Figure 6.
The influence of S. pyogenes strains on tumor volume in C57BL/6 mice with (A) murine hepatoma and (B) pancreatic cancer was statistically significant (**p<0.01, **** p<0.0001) 10 days following inoculation with S. pyogenes GUR and S. pyogenes GURSA1.
Figure 6.
The influence of S. pyogenes strains on tumor volume in C57BL/6 mice with (A) murine hepatoma and (B) pancreatic cancer was statistically significant (**p<0.01, **** p<0.0001) 10 days following inoculation with S. pyogenes GUR and S. pyogenes GURSA1.
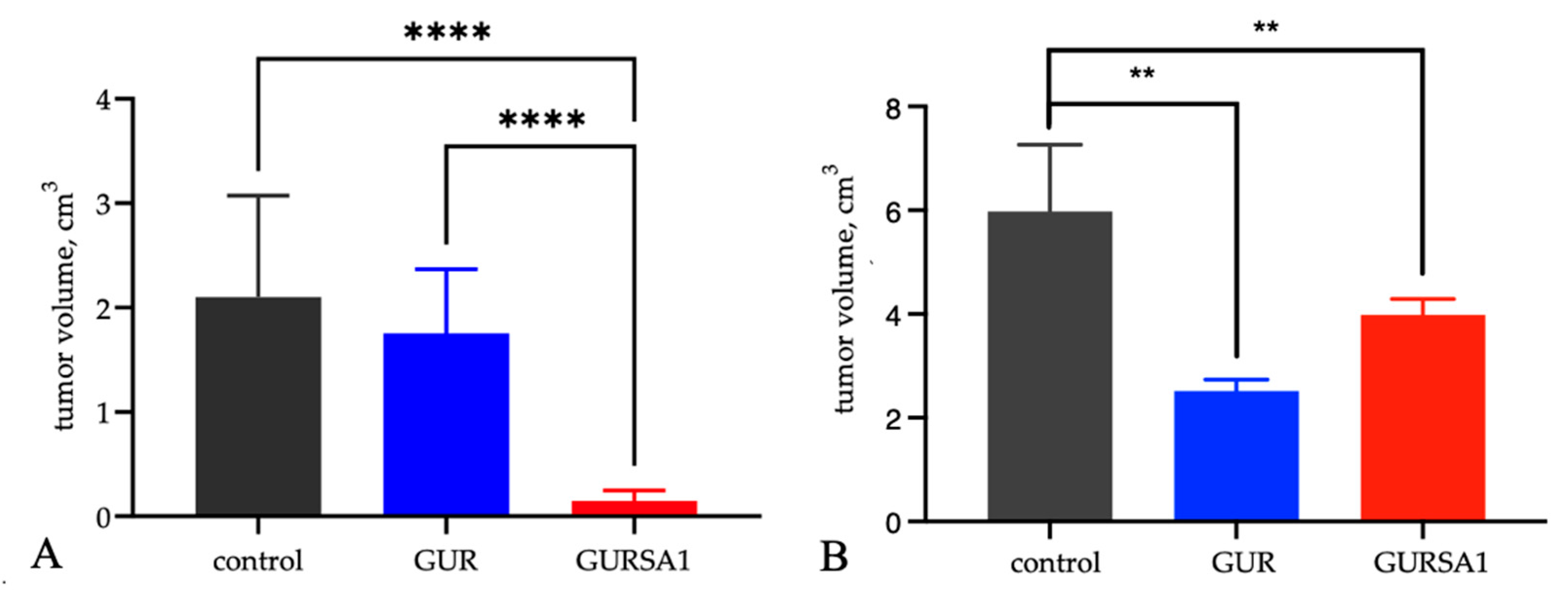

Table 1.
Contamination of mice tumors, livers, and spleens within one day after intraperitoneal injection of S. pyogenes GURSA1 strain.
Table 1.
Contamination of mice tumors, livers, and spleens within one day after intraperitoneal injection of S. pyogenes GURSA1 strain.
| Sample | Tumor, lgCFU | Spleen, lgCFU | Liver, lgCFU |
|---|---|---|---|
| GURSA1 (n=6) | 4,50±1,38 | 0 | 0 |
| Control (n=6) | 0 | 0 | 0 |
Table 7.
Body weight changes of mice administered the S. pyogenes strains.
| Strains | Control | S. pyogenes GURSA1 | S. pyogenes GUR | |||
|---|---|---|---|---|---|---|
| Dose, CFU | 0 | 106 | 108 | 106 | 108 | |
| Body weigth, g | Day 1 | 20,7±0,6 | 20,5±0,5 | 21±0,7 | 20,5±0,6 | 20,2±0,5 |
| Day 7 | 20,5±0,5 | 20,4±0,4 | 20,3±0,3 | 20,0±0,5 | 19,9±0,6 | |
| Day 14 | 20,8±0,5 | 20,6±0,5 | 20,6±0,9 | 20,0±0,1 | 20,4±1,4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



