
Submitted:
09 December 2024
Posted:
10 December 2024
Read the latest preprint version here
Abstract
Background/Objectives: Alzheimer’s disease (AD) remains a leading cause of cognitive decline and mortality worldwide, characterized by neurodegeneration, synaptic deficiencies, and neuroinflammation. Despite advancements in early detection, diagnosis, and treatment, AD presents substantial challenges due to its complex pathology, heterogeneity, and the limited efficacy of current therapies. Consequently, there is a pressing need for novel therapeutic agents to target the multifaceted aspects of AD pathology, enhance current treatments, and minimize adverse effects. AdipoRon, an adiponectin receptor agonist, has garnered interest for its potential neuroprotective effects, including reducing neuroinflammation, improving mitochondrial function, and mitigating tau hyperphosphorylation. Methods: This review aimed to evaluate the effects of AdipoRon-based adiponectin replacement therapy against AD using a comprehensive approach grounded in the PICO framework—Population, Intervention, Comparison, and Outcomes. Results: A total of six studies were reviewed, including in vitro and in vivo investigations, examining AdipoRon's impact on various AD models. These studies involved different cell lines and transgenic mouse models, assessing various outcomes such as cognitive function, neuroinflammation, tau phosphorylation, synaptic deficiencies, and relevant molecular pathways. Conclusions: By synthesizing data from these studies, our review provides a thorough understanding of AdipoRon's neuroprotective effects, its mechanisms of action, and its potential as a therapeutic agent for AD. This analysis aims to highlight the current state of knowledge, identify gaps in the research, and suggest directions for future studies and clinical applications.
Keywords:
AdipoRon
; Alzheimer’s Disease
; Adiponectin
; Cognitive Impairment
; Synaptic Defects
; Neurology
; Adiponectin-Replacement Therapy
; Adiponectin-Receptor Agonist
1. Introduction
Adipokines, secreted by the adipose tissue, are bioactive molecules that play essential roles in inflammation and metabolism regulation, energy homeostasis, cardiovascular function, immunity, and diseases such as cancer and dementia. However, as two-sided coins, adipokines may contribute to obesity-related disorders in scenarios of adipose tissue dysfunction. The paramount understanding of adipokines relies on the scenario of their release. In healthy adipose tissue individuals, adipokines are mainly anti-inflammatory. In dysfunctional adipose tissue subjects, adipokines are mostly pro-inflammatory. While anti-inflammatory adipokines decrease, the likelihood of cardiovascular and other inflammatory metabolic diseases increases [1,2,3,4]. Obesity induces adipokines imbalance. In obese individuals, “good” adipokines such as adiponectin and omentin-1 are downregulated. These adipokines maintain the survival and functional plasticity of mesenchymal stem cells. Conversely, “bad” adipokines such as leptin, visfatin, and resistin also promote mesenchymal stem cells to differentiate into adipocytes, impairing pleiotropic functions and accelerating senescence and cell death via apoptosis [5,6,7]. In the brain, receptors for adipokines are abundantly presented. Evidence has demonstrated that adipokines such as adiponectin cross the blood-brain barrier, and it is known that the neurotrophic and anti-inflammatory properties of adipokines drive their neuroprotective effects against neurodegeneration [8,9,10,11,12,13]. Investigations reported that dysregulated adipokine secretion underlines the co-occurrence of comorbidities such as metabolic syndrome and neurodegenerative diseases [14,15,16,17,18,19,20,21,22].
Alzheimer’s disease (AD) stands as one of the most pressing challenges in neurodegenerative disorders, characterized by a relentless decline in cognitive function and the accumulation of pathological proteins such as amyloid-beta (Aβ) plaques and hyperphosphorylated tau tangles [23,24,25,26,27,28]. This complex and progressive disease disrupts daily functioning and quality of life for millions worldwide, underscoring the urgent need for innovative and effective treatment strategies [29,30,31].
In recent years, AD research has increasingly focused on novel therapeutic approaches targeting underlying metabolic and neurodegenerative processes [32,33,34,35,36]. One promising strategy involves AdipoRon, a synthetic agonist that activates the adiponectin receptor [37,38,39,40]. Adiponectin is a hormone secreted by adipose tissue that is well-regarded for its role in regulating metabolic processes such as glucose homeostasis and fatty acid oxidation [41,42,43,44]. Beyond its metabolic functions, adiponectin has garnered attention for its potential neuroprotective effects, which have been linked to its ability to influence several critical pathways involved in neurodegeneration [45,46,47,48].
Adiponectin, AdipoQ, or ACRP30 has a 244 amino acid structure and a molecular weight of approximately 26 kDa. In terms of abundance, adiponectin is the second most secreted. Therefore, many studies have indicated that alterations in adiponectin secretion are related to adipose tissue dysregulation and obesity-related diseases. Regarding mechanisms of action, adiponectin influences metabolism, playing crucial roles in modulating energy homeostasis, inflammation, and other intricate cellular processes such as insulin sensitivity. Adiponectin possesses an N-terminal sequence, a C-terminal domain, a hypervariable domain, and 15 collagenous repeats. The high molecular weight is the most biologically active isoform, which is also the most encountered in plasma [31,49,50]. Adiponectin exerts its effects through various receptors, including Adiponectin Receptor 1 (AdipoR1) and Adiponectin Receptor 2 (AdipoR2), which are involved in modulating critical cellular processes [50,51]. These receptors activate downstream signaling pathways that regulate inflammation, oxidative stress, and mitochondrial function—crucial factors in neurodegenerative diseases like Alzheimer’s [52,53]. Adiponectin’s actions include enhancing insulin sensitivity [54,55,56,57], reducing inflammation [55,56,58,59,60,61], and promoting neuronal survival [62,63,64], all of which are beneficial for maintaining brain health and function [65,66,67,68,69]. Beyond AdipoR1 and AdipoR2, adiponectin also binds to T-cadherin. This third adiponectin receptor is a glycosyl phosphatidylinositol–anchored protein. T-cadherin is most highly expressed within the cardiovascular system, composed of a pro-peptide and five distinct extracellular cadherin domains. This protein lacks transmembrane and cytoplasmic regions. Besides its complex structure, T-cadherin only binds to the hexametric and high molecular weight adiponectin isoforms but not trimeric or spherical forms [70]. However, studies elucidated that adiponectin binding to T-cadherin protects against neointima proliferation and atherosclerosis, which protects individuals against dementia since atherosclerosis is intimately associated with the occurrence of AD [71,72].
AdipoRon (Figure 1), a synthetic adiponectin receptor agonist, was developed to replicate and amplify these beneficial effects [73,74,75,76,77,78]. AdipoRon is believed to harness its therapeutic potential for treating AD by mimicking the adiponectin action. Preclinical studies have shown that AdipoRon can activate similar signaling pathways as adiponectin, such as the Adenosine Monophosphate-Activated Protein Kinase (AMPK) pathway [74,77,79,80,81,82], which plays a role in regulating energy metabolism and cellular stress responses [83,84]. Additionally, AdipoRon has been observed to influence autophagy [82,85,86,87,88,89], a process critical for clearing neurotoxic proteins and maintaining cellular homeostasis [27,90].
This approach is auspicious due to the multifaceted role of adiponectin in neuroprotection. AdipoRon, by modulating critical pathways involved in cellular stress response and metabolism, presents a potential therapeutic strategy for addressing the complex pathology of AD [91,92]. By intervening in these fundamental mechanisms, AdipoRon holds the potential to slow disease progression, improve cognitive function, and support overall brain health. This makes AdipoRon a compelling option for advancing AD treatment and enhancing patient outcomes. Given these insights, this article presents a comprehensive systematic review of AdipoRon-based therapies for AD. To our knowledge, this is the only review that has thoroughly explored the full potential of AdipoRon in this context. We examine a broad array of studies using both in vitro and in vivo models to uncover the therapeutic benefits and mechanisms of AdipoRon treatment. Our review will detail experimental designs, treatment regimens, and outcomes from various studies, highlighting how AdipoRon addresses both cognitive and pathological features of AD. Additionally, we will discuss the possible clinical implications of each finding and outline future research endeavors needed to investigate and validate AdipoRon’s therapeutic potential further.
2. Results
Figure 2 illustrates the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) [93] Flow Diagram for the review process. Two experienced researchers (S.M.B. and L.F.L.) conducted the database search between 1st July 2024 and 1st November 2024. The search process yielded 208 records from databases and 24 records from registers. Following this initial identification, duplicate records were systematically removed, totaling 48. Additionally, 25 records were flagged as ineligible by automated tools, and a further 96 records were excluded for various reasons deemed appropriate. After these exclusions, 63 records remained for manual screening. This process resulted in the exclusion of 54 records that still needed to meet the criteria for further review. Subsequently, nine reports were requested for retrieval to gather more detailed information. Fortunately, all nine reports were successfully retrieved. Each of the nine successfully obtained reports was carefully assessed for eligibility. This assessment led to the exclusion of three reports: one was a non-experimental paper, one did not involve AdipoRon, and one was not based on an AD model. In the end, six studies were deemed eligible and included in the review. However, no additional reports related to these included studies were available.
Table 1 outlines the effects of AdipoRon-based adiponectin replacement therapy on Alzheimer’s disease, including the study types, experimental models, treatment regimens, outcomes, and involved pathways. It highlights the impact on cognitive function, amyloid-beta deposition, neuroinflammation, and the molecular mechanisms through which AdipoRon exerts its effects.
Figure 3 summarizes data from studies evaluating the effect of AdipoRon versus control on novel object preference. The pooled mean difference (MD) favors AdipoRon, though the overall effect does not reach statistical significance (MD=10.73, 95% CI [−1.17,22.64]; p=0.08). The heterogeneity is negligible (I2=0%). Figure 4 illustrates the results of studies comparing AdipoRon-treated groups and controls for the time spent in the novel arm during behavioral testing. The pooled MD significantly favors AdipoRon (MD=15.55, 95% CI [10.44,20.67]; p<0.00001, with no observed heterogeneity (I2=0%). Figure 5 analyses the effects of AdipoRon on the time spent in the target quadrant in behavioral tasks. The pooled MD significantly favors AdipoRon (MD=16.97, 95% CI [9.36,24.59]; p<0.0001), with minimal heterogeneity (I2=0%). Figure 6, which investigated total arm entries between AdipoRon and control groups, shows a significant overall effect favoring AdipoRon (MD=9.02, 95% CI [2.97,15.08]; p=0.003). Heterogeneity is low (I2=0%). Figure 7 shows escape latency results across studies. The pooled MD does not significantly favor either group (MD=−6.89, 95% CI [−15.74,1.96]; p=0.13), and high heterogeneity is observed (I2=82%). Figure 8 examines the swimming speed in behavioral testing. The overall MD suggests no significant difference between AdipoRon and control groups (MD=−1.12MD, 95% CI [−6.96,4.73]; p=0.71), with no heterogeneity (I2=0%). Figure 9 analyses the number of crossing times. AdipoRon shows a significant effect compared to the control (MD=2.32MD 95% CI [1.22,3.42]; p<0.0001, with no heterogeneity (I2=0%).
3. Discussion
To understand AdipoRon's potential in addressing AD, it is essential to systematically evaluate its effects through a comprehensive review of existing research. The following analysis aims to uncover the underlying logic behind AdipoRon's impact and provide a clear assessment of its efficacy and mechanisms of action.
3.1. Physicochemical Properties, Structural Characteristics, and Therapeutic Potential of AdipoRon: Progress and Obstacles
AdipoRon, with the molecular formula C27H28N2O3, has a molecular weight of 428.5 g/mol. Its exact mass is 428.20999276 g/mol, aligning with its monoisotopic mass. The compound exhibits high lipophilicity with an XLogP3-AA value of 4.7, indicating it is more soluble in fats than in water. It has one hydrogen bond donor and four hydrogen bond acceptors, suggesting its capacity for hydrogen bonding and potential interactions with other molecules. AdipoRon features eight rotatable bonds, reflecting its structural flexibility. Its topological polar surface area is 58.6 Ų, suggesting moderate polarity, influencing its ability to cross biological membranes. With a heavy atom count of 32, AdipoRon is relatively complex, and its complexity value of 582 further supports this. The compound is electrically neutral with a formal charge of 0 and lacks chiral centers, indicating no stereoisomerism. These properties highlight AdipoRon’s lipophilic nature, structural complexity, and potential behavior in biological contexts [100].
Since its discovery in 2013, the initially speculative therapeutic potential of AdipoRon has been confirmed through substantial research. After a thorough screening process at the Open Innovation Center for Drug Discovery (The University of Tokyo), AdipoRon has been identified as a pioneering orally active adiponectin receptor agonist. This recognition stems from its exceptional ability to activate AMPK and bind to AdipoR1 and AdipoR2, as demonstrated in C2C12 murine myoblast cells [101]. Enamine Ltd. (Kiev, Ukraine) developed AdipoRon via a multi-step synthesis, starting with the alkylation of hydroxybenzophenone using methyl chloroacetate. The resulting compound features three distinct functional groups: a 1-benzyl 4-substituted 6-membered cyclic amine moiety, a carbonyl group, and a terminal aromatic ring. Radioactive binding and Scatchard analysis have confirmed the specificity of AdipoRon in vitro, showing its affinity for AdipoR1 and AdipoR2 with dissociation constants (Kd) of approximately 1.8 μM and 3.1 μM, respectively—however, no randomized clinical trials involving AdipoRon in human subjects [73].
Although AdipoRon has been found to promote neuroprotection and restoration of synaptic functions in various models of brain disease in mice, caution is needed to overcome limitations associated with AdipoRon use during preclinical practice. Previous research noted that while low-dose AdipoRon treatment enhanced hippocampal cell proliferation while increasing serum adiponectin and brain-derived neurotrophic factor (BDNF) in mice with hippocampal defects, higher doses of AdipoRon treatments, conversely, suppressed hippocampal neurogenesis and influenced reductions in serum levels of adiponectin and BDNF. Thus, it is worth noting that negative feedback may occur by reducing adiponectin secretion while suppressing adipogenesis to overcome the hyperactivation of adiponectin receptors and their mediated cascades, impairing hippocampal neurogenesis. The authors potentially link AdipoRon hyper-activation of the AMPK signaling with the Mammalian Target of Rapamycin (mTOR) inhibition and BDNF level reduction, consequently leading to impaired hippocampal neurogenesis [102]. Given this scenario and AdipoRon is a lipophilic drug, dose- and time-dependent treatments must be necessarily and accurately described before AdipoRon can be included in translational medicine to treat neurodegenerative diseases. The blood-brain barrier (BBB) is a protective membrane naturally occurring in the central nervous system. Protecting the brain from toxins and pathogens in the blood, the BBB is crucial for developing the central nervous system and its well-functioning. However, the BBB also possesses its complications since it complicates pharmacotherapy for brain disorders [103]. AdipoRon, as a lipophilic drug candidate, has facilitated entering the central nervous system via the BBB. Lipophilic drugs often pass the BBB compartment by either passive diffusion or becoming solubilized into the lipid bilayer of the BBB blood vessels [104]. Although preoccupations such as insufficient drug delivery into the brain may not necessarily be a problem regarding AdipoRon administration in diseases like dementia due to its possible facilitated transport into the central nervous system, we must highlight that more pharmacodynamics and pharmacokinetics studies are necessary to fully realize the potential of AdipoRon in treating brain disorders. Since previous research denoted the toxicity of this synthetic peptide in the realm of hippocampal neurogenesis, caution is warranted in choosing the correct doses and treatment duration regimens.
3.2. Evaluating the Therapeutic Potential of AdipoRon in Alzheimer’s Disease
This study evaluates the efficacy of AdipoRon, an adiponectin receptor agonist, in addressing AD, cognitive impairment, and synaptic defects through various in vitro and in vivo models. The results indicate promising neuroprotective effects of AdipoRon, mediated through several molecular pathways. However, each study has its limitations, which must be considered when interpreting these findings.
Sun et al. [94] explored how AdipoRon affects autophagy in HT22 cells and APP/PS1 transgenic mice, revealing its potential to enhance Aβ clearance and improve cognitive function in AD. The study found that AdipoRon promotes Aβ clearance by activating neuronal autophagy through the AdipoR1/AMPK-dependent nuclear translocation of Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) and subsequent activation of Sirtuin1 (SIRT1). This interaction leads to the release of Deleted in Breast Cancer 1 (DBC1) and the activation of SIRT1, which induces autophagy. The inhibition of either GAPDH or SIRT1 counteracts these effects, underscoring the critical role of SIRT1 in AdipoRon’s mechanism of action. Clinically, these findings suggest that AdipoRon could be developed as a therapeutic agent targeting autophagic pathways to enhance Aβ clearance in Alzheimer’s patients, potentially modifying disease progression by targeting underlying pathophysiological mechanisms. This highlights its potential to alleviate symptoms and address the root cause of Aβ accumulation. However, the study's limitation lies in the reliance on specific mouse models, which may not fully represent the complexity of human AD, necessitating further validation in more diverse models and eventual clinical trials.
Wang et al. [95] conducted experiments using human neuroblastoma cells (SH-SY5Y) and P301S transgenic male mice. AdipoRon treatment reduced hyperphosphorylated tau accumulation, improved memory, synaptic function, mitochondrial fusion, and decreased neuroinflammation in both models. These effects were mediated through activating the AMPK/Glycogen Synthase Kinase 3 Beta (GSK3β) pathway and increased mitochondrial fusion proteins. Clinically, these findings suggest that AdipoRon could be developed as a therapeutic agent targeting tauopathies and mitochondrial dysfunction in AD. This highlights its potential to alleviate symptoms and modify disease progression by targeting underlying pathophysiological mechanisms. However, the limitation of this study lies in the long-term administration (4 months) in mice, which may not be feasible or directly translatable to human treatment regimens, raising concerns about adherence and long-term safety in a clinical setting. It is crucial to note that the long-term AdipoRon administration requires caution. Previous studies have indicated that long-term AdipoRon treatment (20 days) has negatively impacted whole-body insulin sensitivity, increasing IR through exacerbated adipose tissue lipolysis, increased hepatic gluconeogenesis, and impaired muscle tricarboxylic acid cycle [79,82,105,106,107,108]. For instance, increased IR increases the likelihood of AD occurrence [109,110,111,112,113,114]. Therefore, translating long-term AdipoRon treatments to humans warrants further attention. This implicates allometric scale studies and safety margins since AdipoRon is the first commercialized adiponectin receptor agonist for preclinical studies in significant proportions. Therefore, comparative approaches with similar drugs are not available. Additionally, human equivalent dose calculators for AdipoRon translation are also not accessible. Translating long-term AdipoRon regimens to humans would necessarily raise concerns about metabolic imbalances, hormonal disruptions, or unforeseen toxicities.
Khandelwal et al. [96] studied the effects of AdipoRon on Neuro2A cells and APP/PS1 transgenic male mice. The treatment enhanced glucose uptake and insulin sensitivity in vitro, reducing cognitive deficits, Aβ burden, and neuroinflammation in vivo. These outcomes were achieved by activating Glucose Transporter Protein Type-4 (GLUT4) translocation, Protein Kinase B (AKT), GSK3β, and AMPKα phosphorylation pathways. Clinically, this suggests that AdipoRon could mitigate insulin resistance (IR) and amyloid pathology in Alzheimer's patients, providing a dual therapeutic benefit by addressing both metabolic dysfunction and amyloid burden. However, using a specific mouse model may limit the applicability of these results to the broader AD population, as different genetic backgrounds and environmental factors in humans could influence treatment efficacy.
Ng et al. [97] explored the impact of AdipoRon on insulin-resistant HT-22 hippocampal cells and 5xFAD mice. The treatment improved neuronal insulin signaling and insulin sensitivity in vitro enhanced spatial memory synaptic function, and reduced neuroinflammation in vivo. These benefits were linked to AMPK activation and decreased Beta-Secretase 1 (BACE1) and Nuclear Factor Kappa B (NF-kB) levels. Clinically, AdipoRon holds potential as a therapy for IR-associated cognitive impairment in AD, suggesting a new avenue for treating AD patients with comorbid diabetes or prediabetic conditions. However, the study’s limitation is the relatively short treatment period (3 months), which may not capture the long-term effects and safety of the intervention, especially considering the chronic nature of AD.
He et al. [98] used N2a/APPswe cells and multiple mouse models (5xFAD, APP/PS1, and APN KO mice) to assess AdipoRon’s effects. The treatment reduced Aβ deposition, neuroinflammation, and cognitive impairment and enhanced microglia's autophagic activation and lysosomal activity. These effects were mediated through the AMPK-mTOR pathway and increased expression of AdipoR1 and Adaptor Protein Phosphotyrosine Interacting with PH Domain and Leucine Zipper 1 (APPL1). Clinically, AdipoRon could be valuable in reducing amyloid pathology and enhancing autophagy in AD patients, potentially slowing disease progression by promoting the clearance of toxic protein aggregates. The limitation here is the diverse genetic backgrounds of the mouse models used, which may lead to variability in responses that are not representative of the human condition, thereby necessitating further validation in more standardized models and eventual human trials.
Liu et al. [99] investigated the effects of AdipoRon on NE-4C cells and adult male APP/PS1 mice. The treatment improved cell viability and cognitive function and reduced Aβ deposition, mediated by AMPK/cAMP-Response Element Binding Protein (CREB) activation. Clinically, this indicates that AdipoRon could support neuronal survival and cognitive function in Alzheimer's patients by enhancing neuroplasticity and reducing amyloid toxicity. However, the limitation of this study is the short treatment duration (14 days) in vivo, which may not be sufficient to observe the full spectrum of therapeutic effects. Additionally, the mode of administration (injection) may not be the most practical for long-term treatment in humans, necessitating the development of alternative delivery methods.
The findings from the meta-analysis provide compelling insights into the effects of AdipoRon treatment. Across various behavioral tests, including novel object preference and time spent in the novel arm or target quadrant, AdipoRon significantly improved performance metrics. These outcomes suggest that AdipoRon positively influences spatial memory and cognitive function, supported by pooled mean differences. Results such as increased total arm entries and reduced escape latency in behavioral tasks highlight enhanced task-related motivation and mobility in the AdipoRon-treated groups. This indicates improved physical or neuromuscular performance. Most comparisons show negligible heterogeneity, reinforcing the robustness of the findings. However, high heterogeneity in specific metrics, like escape latency, warrants further investigation into methodological variations or population-specific responses.
Overall, the evidence indicates AdipoRon as a promising intervention to enhance cognitive, behavioral, and physical performance, though further research is essential to confirm specific effects and address variability. AdipoRon, a synthetic agonist of adiponectin receptors AdipoR1 and AdipoR2, has garnered significant attention for its potential therapeutic effects on cognitive function and behavioral outcomes [40]. Our meta-analysis provided offers valuable insights into these effects, particularly concerning enhanced cognitive and behavioral performance, as well as improved locomotor activity and task efficiency. Our findings reveal that AdipoRon administration significantly improves various cognitive and behavioral parameters. For instance, in tests assessing novel object preference and time spent in the novel arm or target quadrant, AdipoRon-treated groups exhibited superior performance compared to controls. These findings suggest that AdipoRon positively influences spatial memory and cognitive function. Such enhancements are crucial, especially in conditions characterized by cognitive deficits, such as AD and type 2 diabetes mellitus. Supporting these observations, Zhao et al. [115] demonstrated that AdipoRon ameliorates synaptic dysfunction and inhibits tau hyperphosphorylation through the Adiponectin Receptor (AdipoR)/AMPK/mTOR pathway in type 2 diabetes mellitus (T2DM) mice. The study reported significant restoration of cognitive deficits, including shorter escape latency and increased time spent in the target quadrant, aligning with the meta-analysis outcomes. Furthermore, Chu et al. [116] investigated the role of adiponectin deficiency in cognitive dysfunction among obese mice following sevoflurane exposure. The study found that AdipoRon administration partially prevented cognitive impairments, highlighting its neuroprotective properties. The meta-analysis also indicates that AdipoRon enhances locomotor activity and task efficiency. This is evidenced by increased total arm entries and reduced escape latency in behavioral tasks, suggesting heightened motivation and improved physical performance. Azizifar et al. [117] explored the effects of intranasal AdipoRon in a Parkinson's disease (PD) rat model. The study found that AdipoRon exhibited anxiolytic and antidepressant effects and improved motor function, as evidenced by performance in the open field and elevated plus maze tests. Additionally, Lee et al. [118] reported that chronic AdipoRon treatment mimics the effects of physical exercise by restoring hippocampal neuroplasticity in diabetic mice. The treatment increased progenitor cell proliferation and neuronal differentiation in the hippocampal dentate gyrus, improving cognitive and motor functions. The beneficial effects of AdipoRon on cognitive and behavioral functions are primarily mediated through activating the AdipoR/AMPK/mTOR signaling pathway. Activation of this pathway enhances synaptic plasticity, reduces neuroinflammation, and promotes neurogenesis, all essential for cognitive health [115]. Moreover, AdipoRon has been shown to modulate the expression of synaptic proteins such as Postsynaptic Density Protein 95 (PSD95), SYN, Growth-Associated Protein 43 (GAP43), and SYP, which are crucial for synaptic integrity and function. By preserving synaptic structure and function, AdipoRon contributes to improved cognitive and behavioral outcomes [115]. In summary, the meta-analysis, in conjunction with findings from recent studies, underscores the potential of AdipoRon as a therapeutic agent for enhancing cognitive and behavioral functions. Its ability to improve locomotor activity and task efficiency further highlights its multifaceted benefits. These effects are primarily attributed to the activation of the AdipoR/AMPK/mTOR pathway, leading to enhanced synaptic plasticity and neuroprotection. Future research should focus on elucidating the long-term effects of AdipoRon and its potential applications in clinical settings for the treatment of cognitive impairments associated with metabolic and neurodegenerative disorders.
It is essential to note that the included studies, besides similar, contained several differences. Sun et al. [94] and Khandelwal et al. [96] compared the effects of AdipoRon in the same mouse model of AD. However, they assessed different molecular targets, and Khandelwal et al. emphasized the impact of AdipoRon treatment against GLUT4 impairments, which are critical aspects of AD occurrence. Insulin is essential in long-term neuroprotection, and it is known that insulin absence leads to neurodegeneration. IR, which conditions cells to react poorly to insulin signaling, is a key feature of AD occurrence and a significant risk factor for the disease. IR leads to neural and cognitive abnormalities in the brain, predisposing to cognitive dysfunction through hyperinsulinemia, impaired insulin signaling, and altered Amyloid Precursor Protein (APP) metabolism [119,120,121,122]. Previous research has also indicated that fluctuations in Glucose Transporter Proteins (GLUT) are associated with brain disturbances linked to AD, which may be partially caused by a significant down-regulation of GLUT, most closely associated with IR in the brain. Some authors affirm that the first signal of AD is the lack of GLUT4 translocation [123,124,125]. Ng et al. [97] have also discussed insulin sensitivity in the context of AD and AdipoRon treatment. Their results were like the others, with AdipoRon being a fundamental factor for decreasing IR and augmenting insulin signaling and efficiency. However, these authors did not directly investigate GLUT translocation. Instead, they revealed information about neuroinflammation in the context of insulin signaling restoration. Brain IR causes synaptic dysregulation by impaired axonal transport, apoptosis, Aβ production, and neuroinflammation [126,127]. Particularly, hippocampal IR and inflammation are associated primarily with memory impairments and impaired executive functions [128,129]. This mechanism is proposed to be the primary mediator between peripheral IR and brain dysfunction during AD occurrence [130,131,132,133]. As confirmed by Ng et al., NF-kB levels are critical to AD occurrence. NF-kB facilitates the autocrine production of constitutive AD factors. NF-kB regulates the genetic signal for amyloid plaque production, neurofibrillary tangles occurring, neuronal apoptosis, and, principally, neuroinflammation development [134,135,136,137].
4. Literature Search Methodology
4.1. Database Search
A comprehensive literature review evaluated the effects of Adipo-Ron-based adiponectin replacement therapy against AD. Multiple databases, including PubMed, Scopus, Web of Science, and Google Scholar, were systematically searched using the PICO framework—Population, Intervention, Comparison, and Outcomes. Keywords included "AdipoRon," "adiponectin replacement therapy," "Alzheimer’s disease," "in vitro," "in vivo," "neuroinflammation," "tau hyperphosphorylation," "cognitive function," "mitochondrial fusion," and "autophagy." Boolean operators like AND, OR, and NOT were utilized to refine the search results; for example, "AdipoRon AND AD" or "AdipoRon AND neurodegeneration." Filters such as publication date (last decade), language (English), and document type (original research articles) were applied to ensure the relevance and quality of the studies included. The search strategy aimed to capture a broad range of studies focusing on AdipoRon-based therapies in AD treatment, providing a comprehensive review of the available evidence.
4.2. Inclusion Criteria
Due to the absence of clinical trials, the inclusion criteria for the studies were explicitly focused on preclinical research. These included studies involving either in vitro or in vivo models of AD that featured experimental groups treated with AdipoRon. We required that the studies reported outcomes related to cognitive function, neuroinflammation, tau phosphorylation, synaptic deficiencies, and relevant molecular pathways to ensure a comprehensive evaluation of AdipoRon’s effects. Studies were required to have clear and robust experimental designs and result reporting. They had to be published in peer-reviewed journals and provide detailed information on the intervention, control, endpoints, results, and conclusions. Only articles published in the last decade were considered to ensure the relevance of the findings to current research and advancements. Additionally, only studies published in English were included to maintain consistency and ensure accurate interpretation of the data. Discrepancies between the authors on what article must be included or not were resolved by the two authors with most publications on AdipoRon (L.F.L. and S.M.B.) through a critical and conjunct assessment of the study based on the inclusion criteria. These two authors have direct communication.
4.3. Exclusion Criteria
Exclusion criteria included reviews, meta-analyses, non-experimental papers, studies not involving AdipoRon as an intervention, and those not based on AD models. Studies with inadequate experimental designs or unclear reporting of results were also excluded.
4.4. Data Extraction
Data extraction was performed using a standardized form to capture essential information on experimental models, AdipoRon treatment details (concentration and duration), outcomes, and limitations.
4.5. Quality Assessment
The quality of the studies was assessed based on their experimental design, sample size, and clarity of result reporting, following established guidelines for scientific rigor. Initially, the methodological rigor of each study was evaluated based on the design and execution of experiments, the appropriateness of models and controls, and the clarity in defining interventions and endpoints. The sample size was examined to ensure it was adequate for drawing reliable conclusions, and the comprehensiveness of data reporting was assessed to verify that the results were presented with sufficient detail. Potential biases were identified, including variability in experimental conditions and short-term versus long-term outcomes. The studies' ability to translate preclinical findings into potential clinical implications received particular attention. A qualitative data synthesis was conducted to summarize the effects of AdipoRon on AD models, identify expected outcomes, and discuss limitations. The findings were organized to highlight the therapeutic effects of AdipoRon, its mechanisms of action, and its potential as a treatment for AD. By systematically applying these criteria, we aimed to ensure a thorough and objective evaluation of the studies' overall quality and relevance to advancing AdipoRon-based AD therapies. This review also aimed to identify gaps in current research and suggest directions for future studies and clinical applications.
4.6. Synthesis of Results and Summary Measures
After selecting all references, we evaluated the possibility of meta-analysis. The information required to construct the meta-analysis was from the post-intervention period. We adopted the criterion of extracting all data offered between groups in post-intervention. Heterogeneity was calculated via the I² statistic. We interpreted 0%-40%: not be important; 30%-60%: moderate heterogeneity; 50%-75%: substantial heterogeneity; 75%-100%: considerable heterogeneity [138,139]. For the "95% CI" and "Test for overall effect size" values, significant differences were assumed for p<0.05 (or <5%). If the studies did not provide dispersion values of change such as SD, 95% CI, standard errors, or p-values, the missing SDs of the changes (SDchanges) were calculated. Outcomes of the meta-analysis were reported in weighted MD, 95% CI, and p-value. p < 0.05 were considered statistically significant for the overall MD of the intervention group compared to the control group. The results were presented in forest plots. We imposed a random-effect model as this more conservative method permits the study heterogeneity to deviate beyond chance, providing further generalizable results [140]. All data were formed using the Review Manager Program (RevMan 5.4.1).
5. Conclusions
The collective body of research highlights that AdipoRon, an adiponectin receptor agonist, exhibits significant neuroprotective effects across various AD models (Figure 10). AdipoRon’s therapeutic potential appears multifaceted, engaging in several critical pathways that contribute to its efficacy. Specifically, AdipoRon has been shown to activate AMPK, a key regulator of cellular energy homeostasis, which can influence various downstream processes essential for maintaining neuronal health. The enhancement of glucose metabolism facilitated by AdipoRon also plays a crucial role in addressing the metabolic dysfunction commonly observed in AD, potentially improving neuronal energy availability and function. Additionally, AdipoRon’s effects on reducing tau phosphorylation—a hallmark of AD pathology—suggest that it may help mitigate one of the critical drivers of neurodegeneration. AdipoRon could help prevent tau aggregation and subsequent neurofibrillary tangle formation, central to cognitive decline in AD, by targeting tau hyperphosphorylation. Furthermore, AdipoRon’s ability to regulate autophagy is significant, as autophagic dysfunction is implicated in the accumulation of toxic protein aggregates and cellular debris, exacerbating neurodegenerative processes.
However, there are significant limitations in the current body of research that need to be addressed to validate AdipoRon’s therapeutic potential against AD fully. One such limitation is the reliance on specific animal models, which, while valuable, may not fully represent the complexity and variability of human AD. Differences in genetic backgrounds, disease progression, and comorbidities can affect how well these models translate to human outcomes [141,142,143,144,145,146]. Expanding. The duration of treatment in the existing studies also poses a limitation. Many studies have used relatively short treatment periods, which may not capture the long-term effects and safety of AdipoRon. Given the chronic nature of AD, it is crucial to investigate the long-term efficacy and safety of AdipoRon over extended periods to determine its potential for sustained benefit and to monitor for any delayed adverse effects. Additionally, some of the research has been conducted in vitro, which, while providing valuable insights into the cellular mechanisms, does not fully replicate the complexity of living organisms. In vivo studies and clinical trials are necessary to confirm the findings from in vitro experiments and to evaluate how AdipoRon performs in a more physiologically relevant context. Addressing these limitations through future research will be crucial for advancing the development of AdipoRon as a viable therapeutic option for AD. This includes expanding studies to encompass a broader range of animal models and patient populations, evaluating long-term treatment outcomes, and conducting robust clinical trials to assess the safety and efficacy of AdipoRon in diverse human populations. Such comprehensive research efforts will help establish AdipoRon’s role in AD treatment and contribute to its potential translation into clinical practice.
First, expanding studies to include a broader range of animal models and genetic backgrounds can help determine the generalizability of AdipoRon’s effects across different AD phenotypes and stages [67,68,147,148,149,150]. This would provide a more comprehensive understanding of how AdipoRon interacts with varying pathological features of the disease. Second, research should explore optimal dosing regimens and long-term treatment protocols to assess the durability and sustained efficacy of AdipoRon. Investigations into the pharmacokinetics and pharmacodynamics of AdipoRon could offer insights into effective dosing strategies and potential interactions with other medications commonly used in Alzheimer’s treatment. Third, given the promising preclinical results, there is a pressing need for well-designed clinical trials to evaluate the safety and efficacy of AdipoRon in human populations. These trials focus on assessing cognitive outcomes, biomarkers of neuroinflammation, tau phosphorylation, and autophagy in patients with different stages of AD. Moreover, clinical studies should consider diverse patient populations to understand the drug’s effects across genetic and environmental backgrounds. Additionally, future research could investigate the development of user-friendly delivery systems for AdipoRon, such as oral formulations or transdermal patches, which would improve patient adherence and reduce the treatment burden. Assessing the impact of such delivery methods on drug absorption, efficacy, and side effects is crucial for translating preclinical successes into clinical practice. Lastly, exploring the potential synergistic effects of AdipoRon in combination with other therapeutic agents, such as existing Alzheimer’s drugs or novel compounds, could enhance treatment outcomes. Investigating how AdipoRon interacts with other treatments may uncover new strategies for combination therapies that address multiple aspects of Alzheimer’s pathology.
In conclusion, the collective evidence from these studies underscores the potential of AdipoRon as a promising therapeutic agent for AD, highlighting its multifaceted benefits across various molecular pathways. The findings suggest that AdipoRon may offer significant neuroprotection by enhancing Aβ clearance, improving cognitive function, reducing tau hyperphosphorylation, and mitigating neuroinflammation. Despite the promising results, each study presents limitations, including the reliance on specific animal models and the need for validation in diverse and long-term human trials. Addressing these limitations through further research will be crucial in determining the clinical viability of AdipoRon and its potential to modify AD progression while addressing both symptomatic relief and underlying pathophysiological mechanisms.
In addition to the limitations discussed, other potential concerns warrant consideration. One significant aspect is the possibility of AdipoRon interacting with receptors beyond the adiponectin receptor [80,88,89,151,152]. Although the primary mechanism of action described involves AdipoR1/R2 and related pathways, AdipoRon may influence or interact with other cellular receptors or signaling pathways. For instance, interactions with insulin receptors or other metabolic signaling pathways could introduce unforeseen effects, particularly in complex diseases like Alzheimer's, where metabolic dysregulation is prevalent. Such interactions could either enhance therapeutic efficacy or contribute to unintended side effects, which may complicate clinical application. Moreover, the potential for off-target effects or interactions with other medications used in the treatment of Alzheimer's or related conditions must be carefully evaluated. As AdipoRon may affect multiple cellular processes, its use in combination with other drugs could lead to interactions that alter the efficacy or safety profile of the treatment. For example, concurrent use with anti-inflammatory or neuroprotective agents might result in synergistic effects or, conversely, create adverse interactions. Thus, a thorough investigation into potential drug-drug interactions is essential to ensure that AdipoRon can be safely and effectively integrated into existing therapeutic regimens.
Author Contributions
Conceptualization, L.F.L. (Lucas), S.M.B., and M.T.; methodology, L.F.L. (Lucas), B.d.O.Z., and S.M.B.; software, L.F.L. (Lucas); validation, L.F.L. (Lucas), S.M.B., and M.T.; formal analysis, L.F.L. (Lucas); investigation, L.F.L. (Lucas), R.M.S.d.S., L.G.C., V.B.F.N.J.d.M., M.E.B., C.R.P.D., J.F.d.S.H., V.M.C.S.C., S.M.B., and M.T.; resources, S.M.B.; data curation, L.F.L. (Lucas); writing—original draft preparation, L.F.L. (Lucas), R.M.S.d.S., J.d.S.C.O., L.F.L. (Lívia), V.D.R., and L.G.C.; writing—review and editing, S.M.B. and M.T.; visualization, L.F.L. (Lucas); supervision, S.M.B.; project administration, L.F.L. (Lucas); funding acquisition, S.M.B. and M.T. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing does not apply to this article.
Acknowledgments
We would like to express our sincere gratitude to Servier Medical Art for providing the medical figures used to produce the images for this article. Their high-quality illustrations significantly enhanced the clarity and visual appeal of our work. Servier Medical Art is licensed under CC BY 4.0, allowing for their use with proper attribution. We appreciate their support and contribution to the advancement of scientific communication.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Martínez-Guardado, I.; Navarro-Jiménez, E.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. The Role of Adipokines in Health and Disease. Biomedicines 2023, 11. [CrossRef]
- Arjunan, A.; Song, J. Pharmacological and physiological roles of adipokines and myokines in metabolic-related dementia. Biomedicine & Pharmacotherapy 2023, 163, 114847. [CrossRef]
- Díaz-Castro, F.; Morselli, E.; Claret, M. Interplay between the brain and adipose tissue: a metabolic conversation. EMBO Rep 2024. [CrossRef]
- Shi, X.; Jiang, A.; Qiu, Z.; Lin, A.; Liu, Z.; Zhu, L.; Mou, W.; Cheng, Q.; Zhang, J.; Miao, K.; et al. Novel perspectives on the link between obesity and cancer risk: from mechanisms to clinical implications. Front Med 2024. [CrossRef]
- Straub, R.H. Chapter II - Pathogenesis and Neuroendocrine Immunology. In The Origin of Chronic Inflammatory Systemic Diseases and their Sequelae, Straub, R.H., Ed.; Academic Press: 2015; pp. 59-129.
- Pham, D.-V.; Nguyen, T.-K.; Park, P.-H. Adipokines at the crossroads of obesity and mesenchymal stem cell therapy. Experimental & Molecular Medicine 2023, 55, 313-324. [CrossRef]
- Pestel, J.; Blangero, F.; Watson, J.; Pirola, L.; Eljaafari, A. Adipokines in obesity and metabolic-related-diseases. Biochimie 2023, 212, 48-59. [CrossRef]
- Jászberényi, M.; Thurzó, B.; Bagosi, Z.; Vécsei, L.; Tanaka, M. The Orexin/Hypocretin System, the Peptidergic Regulator of Vigilance, Orchestrates Adaptation to Stress. Biomedicines 2024, 12, 448.
- Duquenne, M.; Deligia, E.; Folgueira, C.; Bourouh, C.; Caron, E.; Pfrieger, F.; Schwaninger, M.; Nogueiras, R.; Annicotte, J.S.; Imbernon, M.; et al. Tanycytic transcytosis inhibition disrupts energy balance, glucose homeostasis and cognitive function in male mice. Mol Metab 2024, 87, 101996. [CrossRef]
- Engin, A. The Mechanism of Leptin Resistance in Obesity and Therapeutic Perspective. Adv Exp Med Biol 2024, 1460, 463-487. [CrossRef]
- Ghosh, P.; Fontanella, R.A.; Scisciola, L.; Taktaz, F.; Pesapane, A.; Basilicata, M.G.; Tortorella, G.; Matacchione, G.; Capuano, A.; Vietri, M.T.; et al. Obesity-induced neuronal senescence: Unraveling the pathophysiological links. Ageing Res Rev 2024, 101, 102533. [CrossRef]
- Höpfinger, A.; Behrendt, M.; Schmid, A.; Karrasch, T.; Schäffler, A.; Berghoff, M. A Cross-Sectional Study: Systematic Quantification of Chemerin in Human Cerebrospinal Fluid. Biomedicines 2024, 12. [CrossRef]
- Natarajan, D.; Ekambaram, S.; Tarantini, S.; Yelahanka Nagaraja, R.; Yabluchanskiy, A.; Hedrick, A.F.; Awasthi, V.; Subramanian, M.; Csiszar, A.; Balasubramanian, P. Chronic β3 adrenergic agonist treatment improves brain microvascular endothelial function and cognition in aged mice. bioRxiv 2024. [CrossRef]
- Lee, T.H.; Cheng, K.K.; Hoo, R.L.; Siu, P.M.; Yau, S.Y. The Novel Perspectives of Adipokines on Brain Health. Int J Mol Sci 2019, 20. [CrossRef]
- Charisis, S.; Short, M.I.; Bernal, R.; Kautz, T.F.; Treviño, H.A.; Mathews, J.; Dediós, A.G.V.; Muhammad, J.A.S.; Luckey, A.M.; Aslam, A.; et al. Leptin bioavailability and markers of brain atrophy and vascular injury in the middle age. Alzheimers Dement 2024, 20, 5849-5860. [CrossRef]
- Li, N.; Wang, X.; Lin, R.; Yang, F.; Chang, H.C.; Gu, X.; Shu, J.; Liu, G.; Yu, Y.; Wei, W.; et al. ANGPTL4-mediated microglial lipid droplet accumulation: Bridging Alzheimer's disease and obesity. Neurobiol Dis 2024, 106741. [CrossRef]
- Royall, D.R.; Palmer, R.F. Affliction class moderates the dementing impact of adipokines. Neuropsychology 2024. [CrossRef]
- Hedayati, M.; Valizadeh, M.; Abiri, B. Metabolic obesity phenotypes and thyroid cancer risk: A systematic exploration of the evidence. Obes Sci Pract 2024, 10, e70019. [CrossRef]
- Krienke, M.; Kralisch, S.; Wagner, L.; Tönjes, A.; Miehle, K. Serum Leucine-Rich Alpha-2 Glycoprotein 1 Levels in Patients with Lipodystrophy Syndromes. Biomolecules 2024, 14. [CrossRef]
- Malicka, A.; Ali, A.; MacCannell, A.D.V.; Roberts, L.D. Brown and beige adipose tissue-derived metabokine and lipokine inter-organ signalling in health and disease. Exp Physiol 2024. [CrossRef]
- Shao, X.; Pan, X.; Chen, T.; Chen, Z.; Li, Y.; Zhong, J.; Wang, R.; Yu, J.; Chen, J.; Chen, Y. Exploring the Role of Adipose Tissue Dysregulation in Vitiligo Pathogenesis: A Body Composition Analysis. Acta Derm Venereol 2024, 104, adv41018. [CrossRef]
- Weaver, K.D.; Simon, L.; Molina, P.E.; Souza-Smith, F. The Role of Lymph-Adipose Crosstalk in Alcohol-Induced Perilymphatic Adipose Tissue Dysfunction. Int J Mol Sci 2024, 25. [CrossRef]
- Abubakar, M.B.; Sanusi, K.O.; Ugusman, A.; Mohamed, W.; Kamal, H.; Ibrahim, N.H.; Khoo, C.S.; Kumar, J. Alzheimer's Disease: An Update and Insights Into Pathophysiology. Front Aging Neurosci 2022, 14, 742408. [CrossRef]
- Hampel, H.; Hardy, J.; Blennow, K.; Chen, C.; Perry, G.; Kim, S.H.; Villemagne, V.L.; Aisen, P.; Vendruscolo, M.; Iwatsubo, T.; et al. The Amyloid-β Pathway in Alzheimer's Disease. Mol Psychiatry 2021, 26, 5481-5503. [CrossRef]
- de Lima, E.P.; Tanaka, M.; Lamas, C.B.; Quesada, K.; Detregiachi, C.R.P.; Araújo, A.C.; Guiguer, E.L.; Catharin, V.; de Castro, M.V.M.; Junior, E.B.; et al. Vascular Impairment, Muscle Atrophy, and Cognitive Decline: Critical Age-Related Conditions. Biomedicines 2024, 12. [CrossRef]
- Laurindo, L.F.; de Carvalho, G.M.; de Oliveira Zanuso, B.; Figueira, M.E.; Direito, R.; de Alvares Goulart, R.; Buglio, D.S.; Barbalho, S.M. Curcumin-Based Nanomedicines in the Treatment of Inflammatory and Immunomodulated Diseases: An Evidence-Based Comprehensive Review. Pharmaceutics 2023, 15. [CrossRef]
- Nunes, Y.C.; Mendes, N.M.; Pereira de Lima, E.; Chehadi, A.C.; Lamas, C.B.; Haber, J.F.S.; Dos Santos Bueno, M.; Araújo, A.C.; Catharin, V.C.S.; Detregiachi, C.R.P.; et al. Curcumin: A Golden Approach to Healthy Aging: A Systematic Review of the Evidence. Nutrients 2024, 16. [CrossRef]
- Tanaka, M.; Vécsei, L. Revolutionizing our understanding of Parkinson’s disease: Dr. Heinz Reichmann’s pioneering research and future research direction. Journal of Neural Transmission 2024, 131, 1367-1387. [CrossRef]
- Passeri, E.; Elkhoury, K.; Morsink, M.; Broersen, K.; Linder, M.; Tamayol, A.; Malaplate, C.; Yen, F.T.; Arab-Tehrany, E. Alzheimer's Disease: Treatment Strategies and Their Limitations. Int J Mol Sci 2022, 23. [CrossRef]
- Buglio, D.S.; Marton, L.T.; Laurindo, L.F.; Guiguer, E.L.; Araújo, A.C.; Buchaim, R.L.; Goulart, R.A.; Rubira, C.J.; Barbalho, S.M. The Role of Resveratrol in Mild Cognitive Impairment and Alzheimer's Disease: A Systematic Review. J Med Food 2022, 25, 797-806. [CrossRef]
- Fornari Laurindo, L.; Minniti, G.; Dogani Rodrigues, V.; Fornari Laurindo, L.; Cavallari Strozze Catharin, V.M.; Federighi Baisi Chagas, E.; Dias Dos Anjos, V.; Vialogo Marques de Castro, M.; Baldi Júnior, E.; Cristina Ferraroni Sanches, R.; et al. Exploring the Logic and Conducting a Comprehensive Evaluation of the Adiponectin Receptor Agonists AdipoRon and AdipoAI's Impacts on Bone Metabolism and Repair-A Systematic Review. Curr Med Chem 2024. [CrossRef]
- de la Fuente, A.G.; Pelucchi, S.; Mertens, J.; Di Luca, M.; Mauceri, D.; Marcello, E. Novel therapeutic approaches to target neurodegeneration. Br J Pharmacol 2023, 180, 1651-1673. [CrossRef]
- Valotto Neto, L.J.; Reverete de Araujo, M.; Moretti Junior, R.C.; Mendes Machado, N.; Joshi, R.K.; Dos Santos Buglio, D.; Barbalho Lamas, C.; Direito, R.; Fornari Laurindo, L.; Tanaka, M.; et al. Investigating the Neuroprotective and Cognitive-Enhancing Effects of Bacopa monnieri: A Systematic Review Focused on Inflammation, Oxidative Stress, Mitochondrial Dysfunction, and Apoptosis. Antioxidants (Basel) 2024, 13. [CrossRef]
- Pagotto, G.L.O.; Santos, L.; Osman, N.; Lamas, C.B.; Laurindo, L.F.; Pomini, K.T.; Guissoni, L.M.; Lima, E.P.; Goulart, R.A.; Catharin, V.; et al. Ginkgo biloba: A Leaf of Hope in the Fight against Alzheimer's Dementia: Clinical Trial Systematic Review. Antioxidants (Basel) 2024, 13. [CrossRef]
- Tanaka, M.; Szabó, Á.; Vécsei, L. Redefining Roles: A Paradigm Shift in Tryptophan–Kynurenine Metabolism for Innovative Clinical Applications. International Journal of Molecular Sciences 2024, 25, 12767.
- Tanaka, M.; Vécsei, L. A Decade of Dedication: Pioneering Perspectives on Neurological Diseases and Mental Illnesses. Biomedicines 2024, 12, 1083.
- Laurindo, L.F.; Sosin, A.F.; Lamas, C.B.; de Alvares Goulart, R.; Dos Santos Haber, J.F.; Detregiachi, C.R.P.; Barbalho, S.M. Exploring the logic and conducting a comprehensive evaluation of AdipoRon-based adiponectin replacement therapy against hormone-related cancers-a systematic review. Naunyn Schmiedebergs Arch Pharmacol 2024, 397, 2067-2082. [CrossRef]
- Barbalho, S.M.; Méndez-Sánchez, N.; Fornari Laurindo, L. AdipoRon and ADP355, adiponectin receptor agonists, in Metabolic-associated Fatty Liver Disease (MAFLD) and Nonalcoholic Steatohepatitis (NASH): A systematic review. Biochem Pharmacol 2023, 218, 115871. [CrossRef]
- Laurindo, L.F.; Laurindo, L.F.; Rodrigues, V.D.; Catharin, V.; Simili, O.A.G.; Barboza, G.O.; Catharin, V.C.S.; Sloan, K.P.; Barbalho, S.M. Unraveling the rationale and conducting a comprehensive assessment of AdipoRon (adiponectin receptor agonist) as a candidate drug for diabetic nephropathy and cardiomyopathy prevention and intervention-a systematic review. Naunyn Schmiedebergs Arch Pharmacol 2024. [CrossRef]
- Laurindo, L.F.; Laurindo, L.F.; Rodrigues, V.D.; Chagas, E.F.B.; da Silva Camarinha Oliveira, J.; Catharin, V.; Barbalho, S.M. Mechanisms and effects of AdipoRon, an adiponectin receptor agonist, on ovarian granulosa cells-a systematic review. Naunyn Schmiedebergs Arch Pharmacol 2024. [CrossRef]
- Nguyen, T.M.D. Adiponectin: Role in Physiology and Pathophysiology. Int J Prev Med 2020, 11, 136. [CrossRef]
- Fadzil, M.A.M.; Abu Seman, N.; Abd Rashed, A. The Potential Therapeutic Use of Agarwood for Diabetes: A Scoping Review. Pharmaceuticals (Basel) 2024, 17. [CrossRef]
- Farella, I.; D'Amato, G.; Orellana-Manzano, A.; Segura, Y.; Vitale, R.; Clodoveo, M.L.; Corbo, F.; Faienza, M.F. "OMICS" in Human Milk: Focus on Biological Effects on Bone Homeostasis. Nutrients 2024, 16. [CrossRef]
- Guo, E.; Liu, D.; Zhu, Z. Phenotypic and functional disparities in perivascular adipose tissue. Front Physiol 2024, 15, 1499340. [CrossRef]
- Polito, R.; Di Meo, I.; Barbieri, M.; Daniele, A.; Paolisso, G.; Rizzo, M.R. Adiponectin Role in Neurodegenerative Diseases: Focus on Nutrition Review. Int J Mol Sci 2020, 21. [CrossRef]
- Bloemer, J.; Pinky, P.D.; Govindarajulu, M.; Hong, H.; Judd, R.; Amin, R.H.; Moore, T.; Dhanasekaran, M.; Reed, M.N.; Suppiramaniam, V. Role of Adiponectin in Central Nervous System Disorders. Neural Plast 2018, 2018, 4593530. [CrossRef]
- Laurindo, L.F.; Santos, A.; Carvalho, A.C.A.; Bechara, M.D.; Guiguer, E.L.; Goulart, R.A.; Vargas Sinatora, R.; Araújo, A.C.; Barbalho, S.M. Phytochemicals and Regulation of NF-kB in Inflammatory Bowel Diseases: An Overview of In Vitro and In Vivo Effects. Metabolites 2023, 13. [CrossRef]
- Nishikito, D.F.; Borges, A.C.A.; Laurindo, L.F.; Otoboni, A.; Direito, R.; Goulart, R.A.; Nicolau, C.C.T.; Fiorini, A.M.R.; Sinatora, R.V.; Barbalho, S.M. Anti-Inflammatory, Antioxidant, and Other Health Effects of Dragon Fruit and Potential Delivery Systems for Its Bioactive Compounds. Pharmaceutics 2023, 15. [CrossRef]
- Ramakrishnan, N.; Auger, K.; Rahimi, N.; Jialal, I. Biochemistry, Adiponectin. In StatPearls; StatPearls PublishingCopyright © 2024, StatPearls Publishing LLC.: Treasure Island (FL), 2024.
- Barbalho, S.M.; Minniti, G.; Miola, V.F.B.; Haber, J.; Bueno, P.; de Argollo Haber, L.S.; Girio, R.S.J.; Detregiachi, C.R.P.; Dall'Antonia, C.T.; Rodrigues, V.D.; et al. Organokines in COVID-19: A Systematic Review. Cells 2023, 12. [CrossRef]
- Lee, M.H.; Klein, R.L.; El-Shewy, H.M.; Luttrell, D.K.; Luttrell, L.M. The adiponectin receptors AdipoR1 and AdipoR2 activate ERK1/2 through a Src/Ras-dependent pathway and stimulate cell growth. Biochemistry 2008, 47, 11682-11692. [CrossRef]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13. [CrossRef]
- Santos, J.; Maio, M.C.; Lemes, M.A.; Laurindo, L.F.; Haber, J.; Bechara, M.D.; Prado, P.S.D., Jr.; Rauen, E.C.; Costa, F.; Pereira, B.C.A.; et al. Non-Alcoholic Steatohepatitis (NASH) and Organokines: What Is Now and What Will Be in the Future. Int J Mol Sci 2022, 23. [CrossRef]
- Lihn, A.S.; Pedersen, S.B.; Richelsen, B. Adiponectin: action, regulation and association to insulin sensitivity. Obes Rev 2005, 6, 13-21. [CrossRef]
- Panou, T.; Gouveri, E.; Papazoglou, D.; Papanas, N. The role of novel inflammation-associated biomarkers in diabetic peripheral neuropathy. Metabol Open 2024, 24, 100328. [CrossRef]
- Shang, D.; Zhao, S. Molecular mechanisms of obesity predisposes to atopic dermatitis. Front Immunol 2024, 15, 1473105. [CrossRef]
- Valencia-Ortega, J.; Castillo-Santos, A.; Molerés-Orduña, M.; Solis-Paredes, J.M.; Saucedo, R.; Estrada-Gutierrez, G.; Camacho-Arroyo, I. Influence of Maternal Adipokines on Anthropometry, Adiposity, and Neurodevelopmental Outcomes of the Offspring. Int J Mol Sci 2024, 25. [CrossRef]
- Ouchi, N.; Walsh, K. Adiponectin as an anti-inflammatory factor. Clin Chim Acta 2007, 380, 24-30. [CrossRef]
- Minniti, G.; Pescinini-Salzedas, L.M.; Minniti, G.; Laurindo, L.F.; Barbalho, S.M.; Vargas Sinatora, R.; Sloan, L.A.; Haber, R.S.A.; Araújo, A.C.; Quesada, K.; et al. Organokines, Sarcopenia, and Metabolic Repercussions: The Vicious Cycle and the Interplay with Exercise. Int J Mol Sci 2022, 23. [CrossRef]
- Tilg, H.; Ianiro, G.; Gasbarrini, A.; Adolph, T.E. Adipokines: masterminds of metabolic inflammation. Nat Rev Immunol 2024. [CrossRef]
- Wojciuk, B.; Frulenko, I.; Brodkiewicz, A.; Kita, D.; Baluta, M.; Jędrzejczyk, F.; Budkowska, M.; Turkiewicz, K.; Proia, P.; Ciechanowicz, A.; et al. The Complement System as a Part of Immunometabolic Post-Exercise Response in Adipose and Muscle Tissue. Int J Mol Sci 2024, 25. [CrossRef]
- Wu, X.; Luo, J.; Liu, H.; Cui, W.; Guo, W.; Zhao, L.; Guo, H.; Bai, H.; Guo, K.; Feng, D.; et al. Recombinant adiponectin peptide promotes neuronal survival after intracerebral haemorrhage by suppressing mitochondrial and ATF4-CHOP apoptosis pathways in diabetic mice via Smad3 signalling inhibition. Cell Prolif 2020, 53, e12759. [CrossRef]
- Laurindo, L.F.; de Maio, M.C.; Barbalho, S.M.; Guiguer, E.L.; Araújo, A.C.; de Alvares Goulart, R.; Flato, U.A.P.; Júnior, E.B.; Detregiachi, C.R.P.; Dos Santos Haber, J.F.; et al. Organokines in Rheumatoid Arthritis: A Critical Review. Int J Mol Sci 2022, 23. [CrossRef]
- Liu, Y.J.; Lee, C.W.; Liao, Y.C.; Huang, J.J.; Kuo, H.C.; Jih, K.Y.; Lee, Y.C.; Chern, Y. The role of adiponectin-AMPK axis in TDP-43 mislocalization and disease severity in ALS. Neurobiol Dis 2024, 202, 106715. [CrossRef]
- Fornari Laurindo, L.; Aparecido Dias, J.; Cressoni Araújo, A.; Torres Pomini, K.; Machado Galhardi, C.; Rucco Penteado Detregiachi, C.; Santos de Argollo Haber, L.; Donizeti Roque, D.; Dib Bechara, M.; Vialogo Marques de Castro, M.; et al. Immunological dimensions of neuroinflammation and microglial activation: exploring innovative immunomodulatory approaches to mitigate neuroinflammatory progression. Front Immunol 2023, 14, 1305933. [CrossRef]
- Alimohammadi, S.; Mohaddes, G.; Keyhanmanesh, R.; Athari, S.Z.; Azizifar, N.; Farajdokht, F. Intranasal AdipoRon mitigates motor and cognitive deficits in hemiparkinsonian rats through neuroprotective mechanisms against oxidative stress and synaptic dysfunction. Neuropharmacology 2025, 262, 110180. [CrossRef]
- Carbone, G.; Bencivenga, L.; Santoro, M.A.; De Lucia, N.; Palaia, M.E.; Ercolano, E.; Scognamiglio, F.; Edison, P.; Ferrara, N.; Vitale, D.F.; et al. Impact of serum leptin and adiponectin levels on brain infarcts in patients with mild cognitive impairment and Alzheimer's disease: a longitudinal analysis. Front Endocrinol (Lausanne) 2024, 15, 1389014. [CrossRef]
- Garcia-Garcia, I.; Kamal, F.; Donica, O.; Dadar, M. Plasma levels of adipokines and insulin are associated with markers of brain atrophy and cognitive decline in the spectrum of Alzheimer's Disease. Prog Neuropsychopharmacol Biol Psychiatry 2024, 134, 111077. [CrossRef]
- Nazzi, C.; Avenanti, A.; Battaglia, S. The Involvement of Antioxidants in Cognitive Decline and Neurodegeneration: Mens Sana in Corpore Sano. Antioxidants 2024, 13, 701.
- Pascolutti, R.; Erlandson, S.C.; Burri, D.J.; Zheng, S.; Kruse, A.C. Mapping and engineering the interaction between adiponectin and T-cadherin. J Biol Chem 2020, 295, 2749-2759. [CrossRef]
- Xie, B.; Shi, X.; Xing, Y.; Tang, Y. Association between atherosclerosis and Alzheimer's disease: A systematic review and meta-analysis. Brain Behav 2020, 10, e01601. [CrossRef]
- Fujishima, Y.; Maeda, N.; Matsuda, K.; Masuda, S.; Mori, T.; Fukuda, S.; Sekimoto, R.; Yamaoka, M.; Obata, Y.; Kita, S.; et al. Adiponectin association with T-cadherin protects against neointima proliferation and atherosclerosis. Faseb j 2017, 31, 1571-1583. [CrossRef]
- Nigro, E.; Daniele, A.; Salzillo, A.; Ragone, A.; Naviglio, S.; Sapio, L. AdipoRon and Other Adiponectin Receptor Agonists as Potential Candidates in Cancer Treatments. Int J Mol Sci 2021, 22. [CrossRef]
- Athari, S.Z.; Keyhanmanesh, R.; Farajdokht, F.; Karimipour, M.; Azizifar, N.; Alimohammadi, S.; Mohaddes, G. AdipoRon improves mitochondrial homeostasis and protects dopaminergic neurons through activation of the AMPK signaling pathway in the 6-OHDA-lesioned rats. Eur J Pharmacol 2024, 985, 177111. [CrossRef]
- Hunyenyiwa, T.; Kyi, P.; Scheer, M.; Joshi, M.; Gasparri, M.; Mammoto, T.; Mammoto, A. Inhibition of angiogenesis and regenerative lung growth in Lep(ob/ob) mice through adiponectin-VEGF/VEGFR2 signaling. Front Cardiovasc Med 2024, 11, 1491971. [CrossRef]
- Li, C.; Li, Q.; Wu, X.; Zhang, Z.; Li, J.; Jiang, R.; Li, G.; Liu, X.; Kang, X.; Li, Z.; et al. Localization and expression of C1QTNF6 in chicken follicles and its regulatory effect on follicular granulosa cells. Poult Sci 2024, 104, 104538. [CrossRef]
- Lin, F.; Masterson, E.; Gilbertson, T.A. Adiponectin Signaling Modulates Fat Taste Responsiveness in Mice. Nutrients 2024, 16. [CrossRef]
- Luo, Z.; Wu, A.; Robson, S.; Alper, S.; Yu, W. Adiponectin Signaling Regulates Urinary Bladder Function by Blunting Smooth Muscle Purinergic Contractility. bioRxiv 2024. [CrossRef]
- Samaha, M.M.; El-Desoky, M.M.; Hisham, F.A. AdipoRon, an adiponectin receptor agonist, modulates AMPK signaling pathway and alleviates ovalbumin-induced airway inflammation in a murine model of asthma. Int Immunopharmacol 2024, 136, 112395. [CrossRef]
- Feng, J.; Ma, X.; Liu, Y.; Shi, X.; Jin, L.; Le, Y.; Zhang, Q.; Wang, C. The Role of Human Adiponectin Receptor 1 in 2-Ethylhexyl Diphenyl Phosphate Induced Lipid Metabolic Disruption. Environ Sci Technol 2024, 58, 18190-18201. [CrossRef]
- Liu, Y.; Xu, Y.; Ji, H.; Gao, F.; Ge, R.; Zhou, D.; Fu, H.; Liu, X.; Ma, S. AdipoRon Alleviates Liver Injury by Protecting Hepatocytes from Mitochondrial Damage Caused by Ionizing Radiation. Int J Mol Sci 2024, 25. [CrossRef]
- Selvais, C.M.; Davis-López de Carrizosa, M.A.; Versele, R.; Dubuisson, N.; Noel, L.; Brichard, S.M.; Abou-Samra, M. Challenging Sarcopenia: Exploring AdipoRon in Aging Skeletal Muscle as a Healthspan-Extending Shield. Antioxidants (Basel) 2024, 13. [CrossRef]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol Cell 2017, 66, 789-800. [CrossRef]
- Chen, T.; Shuang, R.; Gao, T.; Ai, L.; Diao, J.; Yuan, X.; He, L.; Tao, W.; Huang, X. OPA1 Mediated Fatty Acid β-Oxidation in Hepatocyte: The Novel Insight for Melatonin Attenuated Apoptosis in Concanavalin A Induced Acute Liver Injury. J Pineal Res 2024, 76, e70010. [CrossRef]
- Duan, Z.X.; Tu, C.; Liu, Q.; Li, S.Q.; Li, Y.H.; Xie, P.; Li, Z.H. Adiponectin receptor agonist AdipoRon attenuates calcification of osteoarthritis chondrocytes by promoting autophagy. J Cell Biochem 2020, 121, 3333-3344. [CrossRef]
- Goli, S.H.; Lim, J.Y.; Basaran-Akgul, N.; Templeton, S.P. Adiponectin pathway activation dampens inflammation and enhances alveolar macrophage fungal killing via LC3-associated phagocytosis. bioRxiv 2024. [CrossRef]
- Laurindo, L.F.; Minniti, G.; Rodrigues, V.D.; Laurindo, L.F.; Catharin, V.; Chagas, E.F.B.; Anjos, V.D.; de Castro, M.V.M.; Baldi Júnior, E.; Sanches, R.C.F.; et al. Exploring the Logic and Conducting a Comprehensive Evaluation of the Adiponectin Receptor Agonists AdipoRon and AdipoAI's Impacts on Bone Metabolism and Repair-A Systematic Review. Curr Med Chem 2024. [CrossRef]
- Li, C.; Zhang, J.; Dionigi, G.; Liang, N.; Guan, H.; Sun, H. Uncovering the connection between obesity and thyroid cancer: the therapeutic potential of adiponectin receptor agonist in the AdipoR2-ULK axis. Cell Death Dis 2024, 15, 708. [CrossRef]
- Liu, Y.; Fu, X.; Sun, J.; Cui, R.; Yang, W. AdipoRon exerts an antidepressant effect by inhibiting NLRP3 inflammasome activation in microglia via promoting mitophagy. Int Immunopharmacol 2024, 141, 113011. [CrossRef]
- Gómez-Virgilio, L.; Silva-Lucero, M.D.; Flores-Morelos, D.S.; Gallardo-Nieto, J.; Lopez-Toledo, G.; Abarca-Fernandez, A.M.; Zacapala-Gómez, A.E.; Luna-Muñoz, J.; Montiel-Sosa, F.; Soto-Rojas, L.O.; et al. Autophagy: A Key Regulator of Homeostasis and Disease: An Overview of Molecular Mechanisms and Modulators. Cells 2022, 11. [CrossRef]
- Battaglia, S.; Nazzi, C.; Lonsdorf, T.B.; Thayer, J.F. Neuropsychobiology of fear-induced bradycardia in humans: progress and pitfalls. Molecular Psychiatry 2024, 29, 3826-3840. [CrossRef]
- Liu, B.; Liu, J.; Wang, J.-g.; Liu, C.-l.; Yan, H.-j. AdipoRon improves cognitive dysfunction of Alzheimer’s disease and rescues impaired neural stem cell proliferation through AdipoR1/AMPK pathway. Experimental Neurology 2020, 327, 113249. [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 2021, 372, n71-n71. [CrossRef]
- Sun, F.; Wang, J.; Meng, L.; Zhou, Z.; Xu, Y.; Yang, M.; Li, Y.; Jiang, T.; Liu, B.; Yan, H. AdipoRon promotes amyloid-β clearance through enhancing autophagy via nuclear GAPDH-induced sirtuin 1 activation in Alzheimer's disease. Br J Pharmacol 2024. [CrossRef]
- Wang, C.; Chang, Y.; Zhu, J.; Wu, Y.; Jiang, X.; Zheng, S.; Li, G.; Ma, R. AdipoRon mitigates tau pathology and restores mitochondrial dynamics via AMPK-related pathway in a mouse model of Alzheimer's disease. Exp Neurol 2023, 363, 114355. [CrossRef]
- Khandelwal, M.; Manglani, K.; Upadhyay, P.; Azad, M.; Gupta, S. AdipoRon induces AMPK activation and ameliorates Alzheimer's like pathologies and associated cognitive impairment in APP/PS1 mice. Neurobiol Dis 2022, 174, 105876. [CrossRef]
- Ng, R.C.; Jian, M.; Ma, O.K.; Bunting, M.; Kwan, J.S.; Zhou, G.J.; Senthilkumar, K.; Iyaswamy, A.; Chan, P.K.; Li, M.; et al. Chronic oral administration of adipoRon reverses cognitive impairments and ameliorates neuropathology in an Alzheimer's disease mouse model. Mol Psychiatry 2021, 26, 5669-5689. [CrossRef]
- He, K.; Nie, L.; Ali, T.; Wang, S.; Chen, X.; Liu, Z.; Li, W.; Zhang, K.; Xu, J.; Liu, J.; et al. Adiponectin alleviated Alzheimer-like pathologies via autophagy-lysosomal activation. Aging Cell 2021, 20, e13514. [CrossRef]
- Liu, B.; Liu, J.; Wang, J.G.; Liu, C.L.; Yan, H.J. AdipoRon improves cognitive dysfunction of Alzheimer's disease and rescues impaired neural stem cell proliferation through AdipoR1/AMPK pathway. Exp Neurol 2020, 327, 113249. [CrossRef]
- NLM, N.L.o.M.-N.I.o.H. AdipoRon. Available online: (accessed on 22 July, 2024).
- Okada-Iwabu, M.; Yamauchi, T.; Iwabu, M.; Honma, T.; Hamagami, K.; Matsuda, K.; Yamaguchi, M.; Tanabe, H.; Kimura-Someya, T.; Shirouzu, M.; et al. A small-molecule AdipoR agonist for type 2 diabetes and short life in obesity. Nature 2013, 503, 493-499. [CrossRef]
- Lee, T.H.; Christie, B.R.; van Praag, H.; Lin, K.; Siu, P.M.; Xu, A.; So, K.F.; Yau, S.Y. AdipoRon Treatment Induces a Dose-Dependent Response in Adult Hippocampal Neurogenesis. Int J Mol Sci 2021, 22. [CrossRef]
- Wu, D.; Chen, Q.; Chen, X.; Han, F.; Chen, Z.; Wang, Y. The blood–brain barrier: Structure, regulation and drug delivery. Signal Transduction and Targeted Therapy 2023, 8, 217. [CrossRef]
- Lawther, B.K.; Kumar, S.; Krovvidi, H. Blood–brain barrier. Continuing Education in Anaesthesia Critical Care & Pain 2011, 11, 128-132. [CrossRef]
- Wang, Y.; Liu, H.; Zhang, R.; Xiang, Y.; Lu, J.; Xia, B.; Peng, L.; Wu, J. AdipoRon exerts opposing effects on insulin sensitivity via fibroblast growth factor 21-mediated time-dependent mechanisms. J Biol Chem 2022, 298, 101641. [CrossRef]
- Formolo, D.A.; Lee, T.H.; Yu, J.; Lin, K.; Chen, G.; Kranz, G.S.; Yau, S.Y. Increasing Adiponectin Signaling by Sub-Chronic AdipoRon Treatment Elicits Antidepressant- and Anxiolytic-Like Effects Independent of Changes in Hippocampal Plasticity. Biomedicines 2023, 11. [CrossRef]
- Ma, O.K.; Ronsisvalle, S.; Basile, L.; Xiang, A.W.; Tomasella, C.; Sipala, F.; Pappalardo, M.; Chan, K.H.; Milardi, D.; Ng, R.C.; et al. Identification of a novel adiponectin receptor and opioid receptor dual acting agonist as a potential treatment for diabetic neuropathy. Biomed Pharmacother 2023, 158, 114141. [CrossRef]
- Selvais, C.M.; Davis-López de Carrizosa, M.A.; Nachit, M.; Versele, R.; Dubuisson, N.; Noel, L.; Gillard, J.; Leclercq, I.A.; Brichard, S.M.; Abou-Samra, M. AdipoRon enhances healthspan in middle-aged obese mice: striking alleviation of myosteatosis and muscle degenerative markers. J Cachexia Sarcopenia Muscle 2023, 14, 464-478. [CrossRef]
- Neth, B.J.; Craft, S. Insulin Resistance and Alzheimer’s Disease: Bioenergetic Linkages. Frontiers in Aging Neuroscience 2017, 9. [CrossRef]
- Abosharaf, H.A.; Elsonbaty, Y.; Tousson, E.; Mohamed, T.M. Metformin effectively alleviates the symptoms of Alzheimer in rats by lowering amyloid β deposition and enhancing the insulin signal. Metab Brain Dis 2024, 40, 41. [CrossRef]
- Ellis, D.; Watanabe, K.; Wilmanski, T.; Lustgarten, M.S.; Korat, A.V.A.; Glusman, G.; Hadlock, J.J.; Fiehn, O.; Sebastiani, P.; Price, N.D.; et al. APOE Genotype and Biological Age Impact Inter-Omic Associations Related to Bioenergetics. bioRxiv 2024. [CrossRef]
- Kciuk, M.; Kruczkowska, W.; Gałęziewska, J.; Wanke, K.; Kałuzińska-Kołat, Ż.; Aleksandrowicz, M.; Kontek, R. Alzheimer's Disease as Type 3 Diabetes: Understanding the Link and Implications. Int J Mol Sci 2024, 25. [CrossRef]
- Natale, F.; Spinelli, M.; Rinaudo, M.; Gulisano, W.; Nifo Sarrapochiello, I.; Aceto, G.; Puzzo, D.; Fusco, S.; Grassi, C. Inhibition of zDHHC7-driven protein S-palmitoylation prevents cognitive deficits in an experimental model of Alzheimer's disease. Proc Natl Acad Sci U S A 2024, 121, e2402604121. [CrossRef]
- Toledano, A.; Rodríguez-Casado, A.; Älvarez, M.I.; Toledano-Díaz, A. Alzheimer's Disease, Obesity, and Type 2 Diabetes: Focus on Common Neuroglial Dysfunctions (Critical Review and New Data on Human Brain and Models). Brain Sci 2024, 14. [CrossRef]
- Zhao, W.; Zhang, W.; Hu, Y.; Zhou, Y.; Zhao, J.; Li, Y.; Xu, Z. AdipoRon Ameliorates Synaptic Dysfunction and Inhibits tau Hyperphosphorylation through the AdipoR/AMPK/mTOR Pathway in T2DM Mice. Neurochem Res 2024, 49, 2075-2086. [CrossRef]
- Chu, J.M.T.; Chiu, S.P.W.; Wang, J.; Chang, R.C.C.; Wong, G.T.C. Adiponectin deficiency is a critical factor contributing to cognitive dysfunction in obese mice after sevoflurane exposure. Mol Med 2024, 30, 177. [CrossRef]
- Azizifar, N.; Mohaddes, G.; Keyhanmanesh, R.; Athari, S.Z.; Alimohammadi, S.; Farajdokht, F. Intranasal AdipoRon Mitigated Anxiety and Depression-Like Behaviors in 6-OHDA-Induced Parkinson 's Disease Rat Model: Going Beyond Motor Symptoms. Neurochem Res 2024, 49, 3030-3042. [CrossRef]
- Lee, T.H.; Ahadullah; Christie, B.R.; Lin, K.; Siu, P.M.; Zhang, L.; Yuan, T.F.; Komal, P.; Xu, A.; So, K.F.; et al. Chronic AdipoRon Treatment Mimics the Effects of Physical Exercise on Restoring Hippocampal Neuroplasticity in Diabetic Mice. Mol Neurobiol 2021, 58, 4666-4681. [CrossRef]
- Amin, A.M.; Mostafa, H.; Khojah, H.M.J. Insulin resistance in Alzheimer’s disease: The genetics and metabolomics links. Clinica Chimica Acta 2023, 539, 215-236. [CrossRef]
- Sędzikowska, A.; Szablewski, L. Insulin and Insulin Resistance in Alzheimer's Disease. Int J Mol Sci 2021, 22. [CrossRef]
- Yoon, J.H.; Hwang, J.; Son, S.U.; Choi, J.; You, S.W.; Park, H.; Cha, S.Y.; Maeng, S. How Can Insulin Resistance Cause Alzheimer's Disease? Int J Mol Sci 2023, 24. [CrossRef]
- Stoykovich, S.; Gibas, K. APOE ε4, the door to insulin-resistant dyslipidemia and brain fog? A case study. Alzheimer's & Dementia: Diagnosis, Assessment & Disease Monitoring 2019, 11, 264-269. [CrossRef]
- Albaik, M.; Sheikh Saleh, D.; Kauther, D.; Mohammed, H.; Alfarra, S.; Alghamdi, A.; Ghaboura, N.; Sindi, I.A. Bridging the gap: glucose transporters, Alzheimer’s, and future therapeutic prospects. Frontiers in Cell and Developmental Biology 2024, 12. [CrossRef]
- Kyrtata, N.; Emsley, H.C.A.; Sparasci, O.; Parkes, L.M.; Dickie, B.R. A Systematic Review of Glucose Transport Alterations in Alzheimer's Disease. Front Neurosci 2021, 15, 626636. [CrossRef]
- McNay, E.C.; Pearson-Leary, J. GluT4: A central player in hippocampal memory and brain insulin resistance. Exp Neurol 2020, 323, 113076. [CrossRef]
- Battaglia, S.; Avenanti, A.; Vécsei, L.; Tanaka, M. Neurodegeneration in Cognitive Impairment and Mood Disorders for Experimental, Clinical and Translational Neuropsychiatry. Biomedicines 2024, 12, 574.
- Battaglia, S.; Avenanti, A.; Vécsei, L.; Tanaka, M. Neural Correlates and Molecular Mechanisms of Memory and Learning. International Journal of Molecular Sciences 2024, 25, 2724.
- Panov, G.; Dyulgerova, S.; Panova, P. Cognition in Patients with Schizophrenia: Interplay between Working Memory, Disorganized Symptoms, Dissociation, and the Onset and Duration of Psychosis, as Well as Resistance to Treatment. Biomedicines 2023, 11, 3114.
- Shishkina, G.T.; Kalinina, T.S.; Gulyaeva, N.V.; Lanshakov, D.A.; Dygalo, N.N. Changes in Gene Expression and Neuroinflammation in the Hippocampus after Focal Brain Ischemia: Involvement in the Long-Term Cognitive and Mental Disorders. Biochemistry (Moscow) 2021, 86, 657-666. [CrossRef]
- Cabinio, M.; Saresella, M.; Piancone, F.; LaRosa, F.; Marventano, I.; Guerini, F.R.; Nemni, R.; Baglio, F.; Clerici, M. Association between Hippocampal Shape, Neuroinflammation, and Cognitive Decline in Alzheimer's Disease. J Alzheimers Dis 2018, 66, 1131-1144. [CrossRef]
- Komleva, Y.; Chernykh, A.; Lopatina, O.; Gorina, Y.; Lokteva, I.; Salmina, A.; Gollasch, M. Inflamm-Aging and Brain Insulin Resistance: New Insights and Role of Life-style Strategies on Cognitive and Social Determinants in Aging and Neurodegeneration. Frontiers in Neuroscience 2021, 14. [CrossRef]
- Rao, Y.L.; Ganaraja, B.; Murlimanju, B.V.; Joy, T.; Krishnamurthy, A.; Agrawal, A. Hippocampus and its involvement in Alzheimer's disease: a review. 3 Biotech 2022, 12, 55. [CrossRef]
- Vinuesa, A.; Pomilio, C.; Gregosa, A.; Bentivegna, M.; Presa, J.; Bellotto, M.; Saravia, F.; Beauquis, J. Inflammation and Insulin Resistance as Risk Factors and Potential Therapeutic Targets for Alzheimer's Disease. Front Neurosci 2021, 15, 653651. [CrossRef]
- Ju Hwang, C.; Choi, D.Y.; Park, M.H.; Hong, J.T. NF-κB as a Key Mediator of Brain Inflammation in Alzheimer's Disease. CNS Neurol Disord Drug Targets 2019, 18, 3-10. [CrossRef]
- Kaltschmidt, B.; Czaniera, N.J.; Schulten, W.; Kaltschmidt, C. NF-κB in Alzheimer's Disease: Friend or Foe? Opposite Functions in Neurons and Glial Cells. Int J Mol Sci 2024, 25. [CrossRef]
- Mishra, P.; Davies, D.A.; Albensi, B.C. The Interaction Between NF-κB and Estrogen in Alzheimer's Disease. Mol Neurobiol 2023, 60, 1515-1526. [CrossRef]
- Sun, E.; Motolani, A.; Campos, L.; Lu, T. The Pivotal Role of NF-kB in the Pathogenesis and Therapeutics of Alzheimer's Disease. Int J Mol Sci 2022, 23. [CrossRef]
- Higgins, J.P.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557-560. [CrossRef]
- Higgins, J.P.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat Med 2002, 21, 1539-1558. [CrossRef]
- Deeks, J.J.; Higgins, J.P.; Altman, D.G.; Group, o.b.o.t.C.S.M. Analysing data and undertaking meta-analyses. In Cochrane Handbook for Systematic Reviews of Interventions; 2019; pp. 241-284.
- Tanaka, M.; Battaglia, S.; Giménez-Llort, L.; Chen, C.; Hepsomali, P.; Avenanti, A.; Vécsei, L. Innovation at the Intersection: Emerging Translational Research in Neurology and Psychiatry. Cells 2024, 13, 790.
- Climacosa, F.M.M.; Anlacan, V.M.M.; Gordovez, F.J.A.; Reyes, J.C.B.; Tabios, I.K.B.; Manalo, R.V.M.; Cruz, J.M.C.; Asis, J.L.B.; Razal, R.B.; Abaca, M.J.M.; et al. Monitoring drug Efficacy through Multi-Omics Research initiative in Alzheimer's Disease (MEMORI-AD): A protocol for a multisite exploratory prospective cohort study on the drug response-related clinical, genetic, microbial and metabolomic signatures in Filipino patients with Alzheimer's disease. BMJ Open 2024, 14, e078660. [CrossRef]
- Dos Santos, J.R.; Catalão, C.H.R.; Vulczak, A.; Azzolini, A.; Alberici, L.C. Voluntary wheel running decreases amyloidogenic pathway and rescues cognition and mitochondrial energy metabolism in middle-aged female 3xTg-AD mouse model of Alzheimer's disease. J Alzheimers Dis 2024, 102, 424-436. [CrossRef]
- Jabeen, K.; Rehman, K.; Akash, M.S.H.; Hussain, A.; Shahid, M.; Sadaf, B. Biochemical Investigation of the Association of Apolipoprotein E Gene Allele Variations with Insulin Resistance and Amyloid-β Aggregation in Cardiovascular Disease. Clin Exp Pharmacol Physiol 2025, 52, e70007. [CrossRef]
- Permana, A.D.; Mahfud, M.A.S.; Munir, M.; Aries, A.; Rezka Putra, A.; Fikri, A.; Setiawan, H.; Mahendra, I.; Rizaludin, A.; Ramadhani Aziz, A.Y.; et al. A Combinatorial Approach with Microneedle Pretreatment and Thermosensitive Gel Loaded with Rivastigmine Lipid Nanoparticle Formulation Enables Brain Delivery via the Trigeminal Nerve. ACS Appl Mater Interfaces 2024. [CrossRef]
- Wang, J.; Wang, X.; An, Z.; Wang, X.; Wang, Y.; Lu, Y.; Qiu, M.; Liu, Z.; Tan, Z. Quantitative Proteomic Analysis of APP/PS1 Transgenic Mice. Curr Alzheimer Res 2024. [CrossRef]
- Tanaka, M.; Vécsei, L. From Lab to Life: Exploring Cutting-Edge Models for Neurological and Psychiatric Disorders. Biomedicines 2024, 12, 613.
- Kozhakhmetov, S.; Kaiyrlykyzy, A.; Jarmukhanov, Z.; Vinogradova, E.; Zholdasbekova, G.; Alzhanova, D.; Kunz, J.; Kushugulova, A.; Askarova, S. Inflammatory Manifestations Associated With Gut Dysbiosis in Alzheimer's Disease. Int J Alzheimers Dis 2024, 2024, 9741811. [CrossRef]
- Mahmoodi, M.; Mirzarazi Dahagi, E.; Nabavi, M.H.; Penalva, Y.C.M.; Gosaine, A.; Murshed, M.; Couldwell, S.; Munter, L.M.; Kaartinen, M.T. Circulating plasma fibronectin affects tissue insulin sensitivity, adipocyte differentiation, and transcriptional landscape of adipose tissue in mice. Physiol Rep 2024, 12, e16152. [CrossRef]
- Satizabal, C.L.; Himali, J.J.; Conner, S.C.; Beiser, A.S.; Maillard, P.; Vasan, R.S.; DeCarli, C.; Seshadri, S. Circulating adipokines and MRI markers of brain aging in middle-aged adults from the community. J Alzheimers Dis 2024, 102, 449-458. [CrossRef]
- Ma, B.; Zheng, Y.; Liu, S.; Qiu, Y.; Xing, X.; Gao, M.; Cao, Z.; Luan, X. Effect of goose-derived adiponectin peptide gADP3 on LPS-induced inflammatory injury in goose liver. Br Poult Sci 2024, 1-14. [CrossRef]
- Martínez-Cignoni, M.R.; González-Vicens, A.; Morán-Costoya, A.; Amengual-Cladera, E.; Gianotti, M.; Valle, A.; Proenza, A.M.; Lladó, I. Diabesity alters the protective effects of estrogens on endothelial function through adipose tissue secretome. Free Radic Biol Med 2024, 224, 574-587. [CrossRef]
Figure 1.
Molecular Structure of AdipoRon.
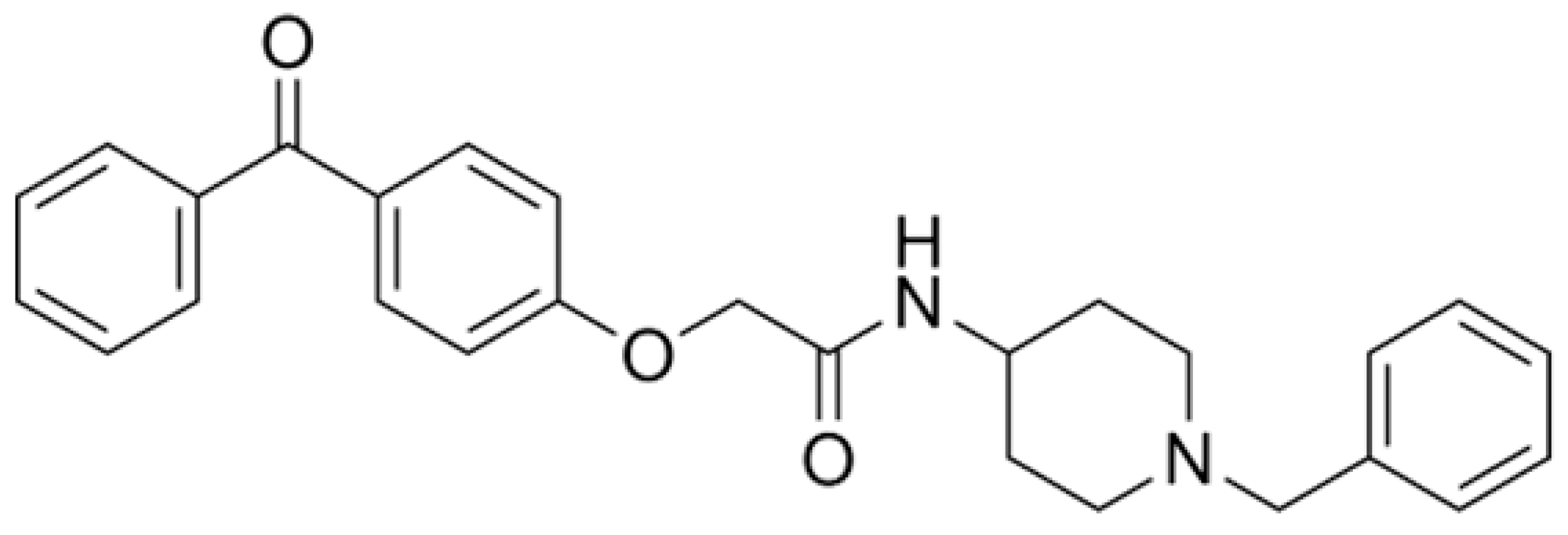
Figure 2.
Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) Flow Diagram Depicting the Systematic Review Process. AD, Alzheimer’s disease.
Figure 2.
Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) Flow Diagram Depicting the Systematic Review Process. AD, Alzheimer’s disease.
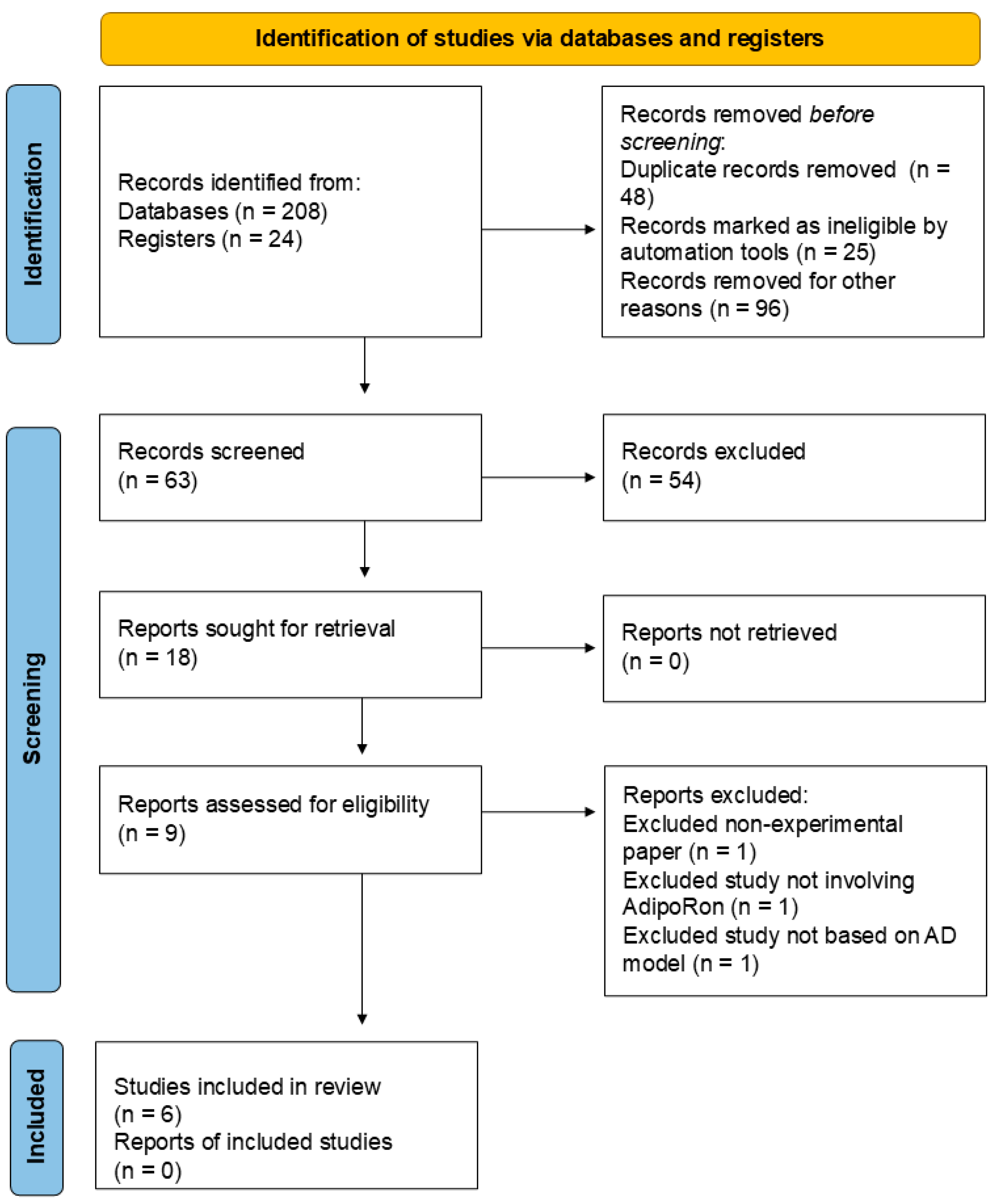
Figure 3.
Meta-analysis for overall effects of AdipoRon on Novel Object Preference.
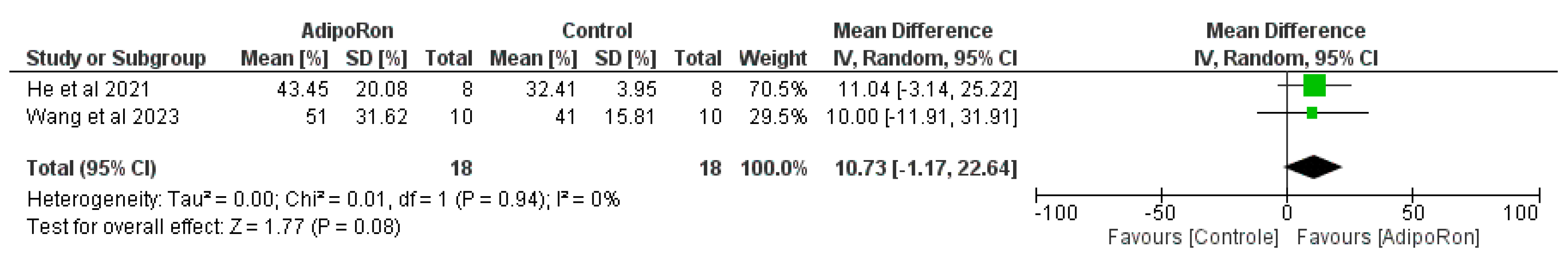
Figure 4.
Meta-analysis for overall effects of AdipoRon on Time Spent in the Novel Arm.
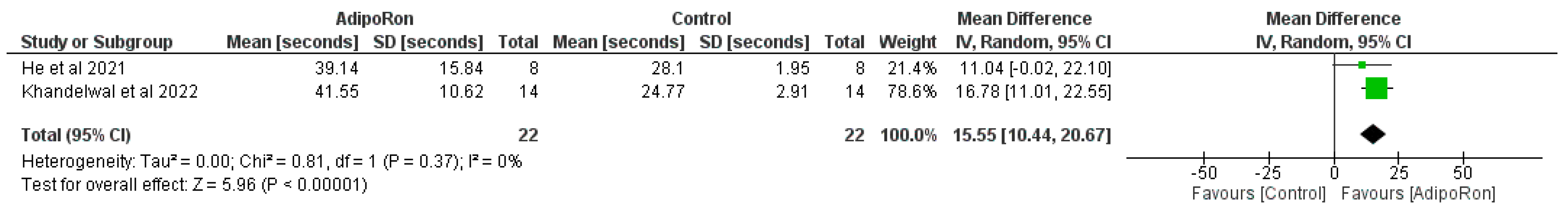
Figure 5.
Meta-analysis for overall effects of AdipoRon on Time Spent in the Target Quadrant.
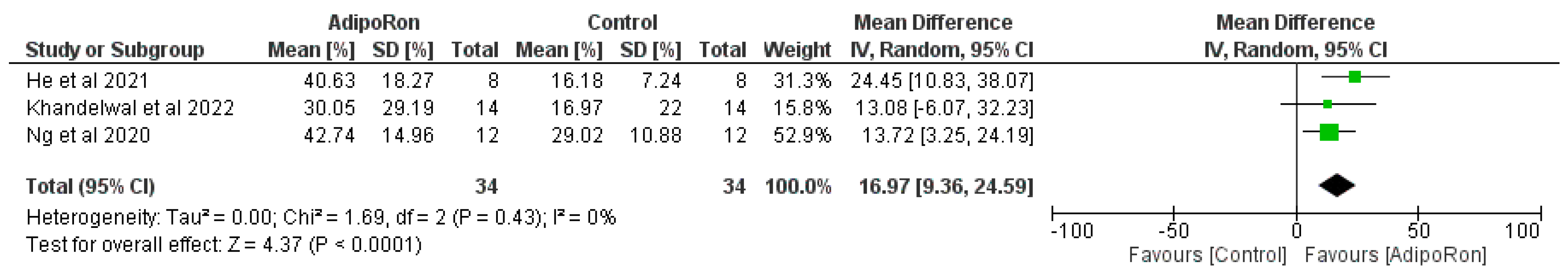
Figure 6.
Meta-analysis for overall effects of AdipoRon on Total Arm Entries.
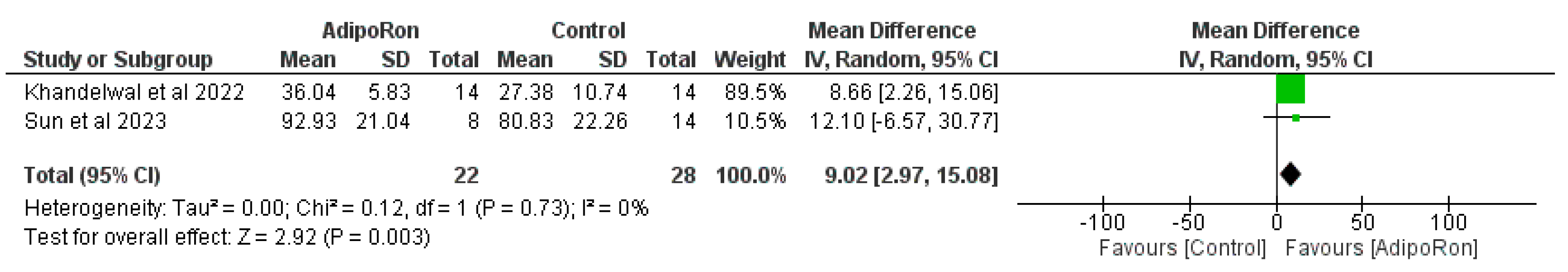
Figure 7.
Meta-analysis for overall effects of AdipoRon on Escape latency.
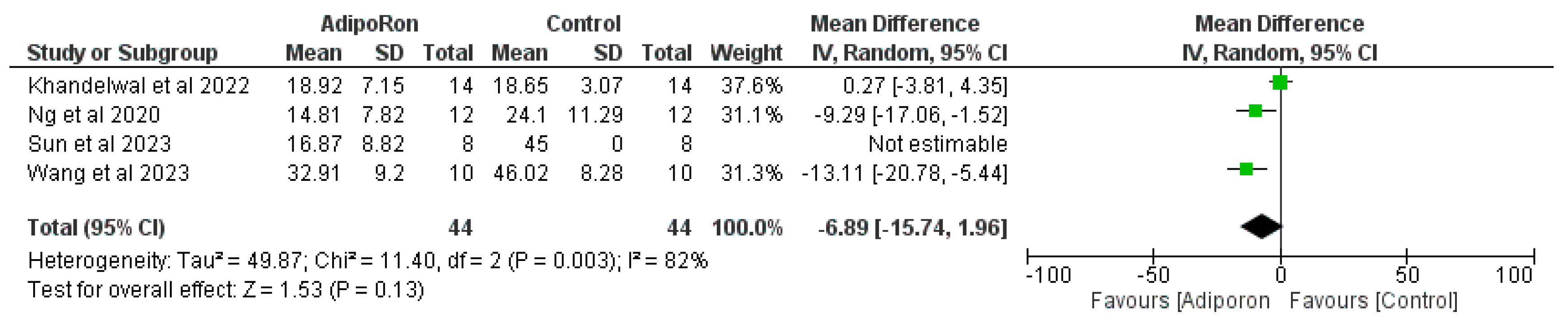
Figure 8.
Meta-analysis for overall effects of AdipoRon on Swimming speed.
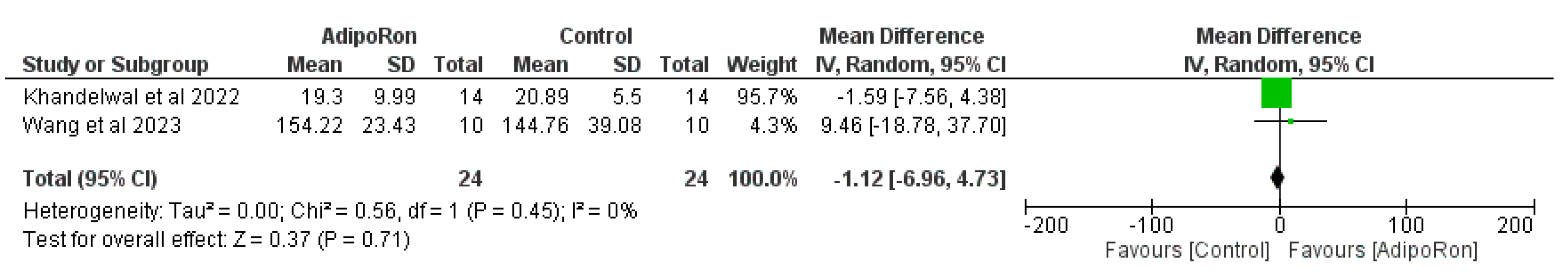
Figure 9.
Meta-analysis for overall effects of AdipoRon on Crossing times.
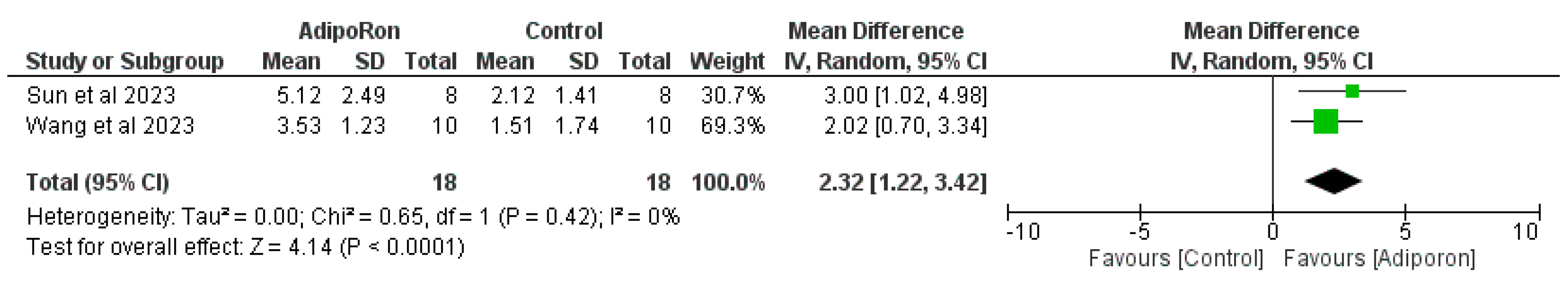
Figure 10.
Adiponectin synthesis and effects of AdipoRon against Alzheimer’s Disease.
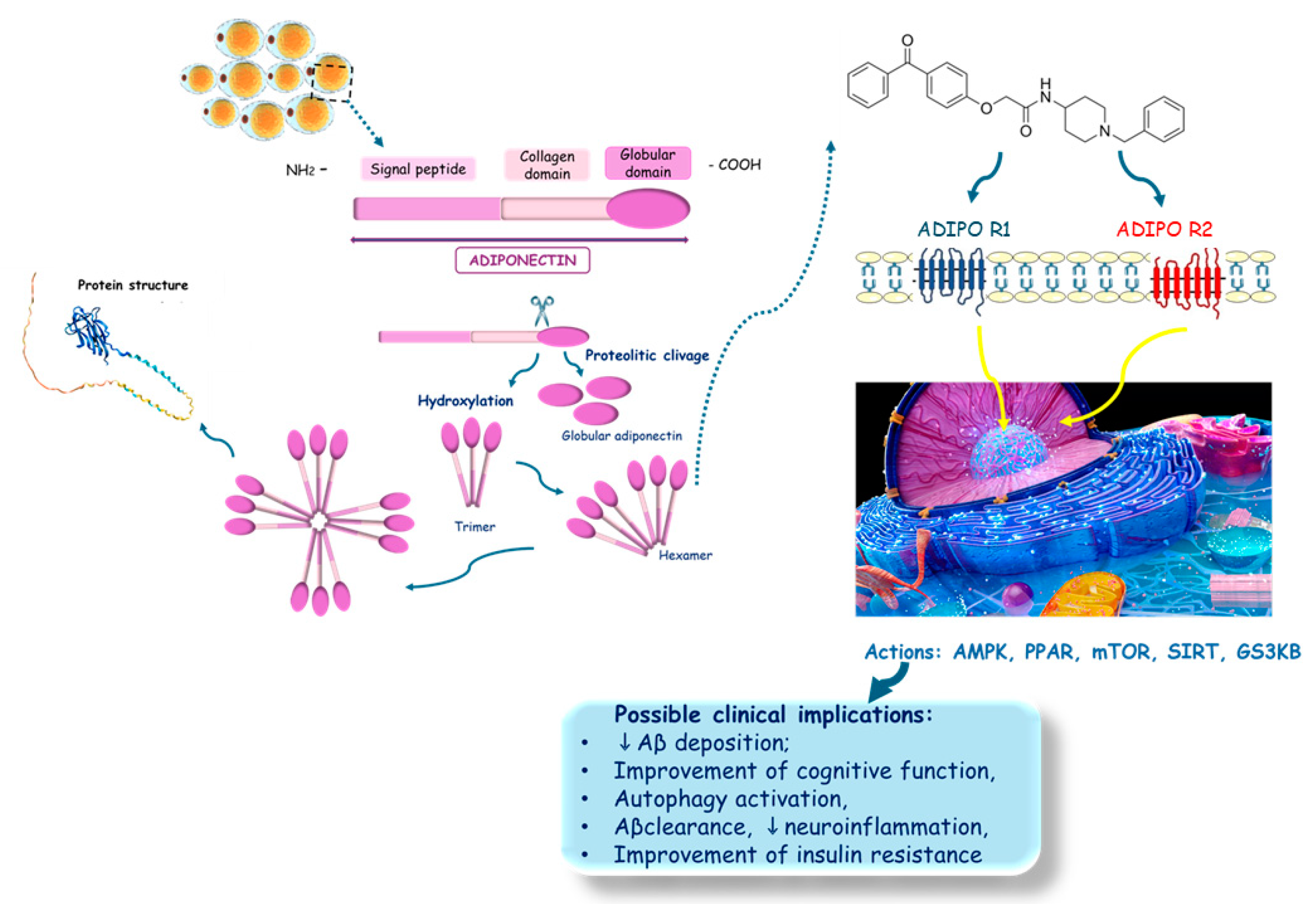

Table 1.
AdipoRon-Based Adiponectin Replacement Therapy Against Alzheimer’s Disease.
| Ref | Type of Study | Model (s) | Experimental Group (s) and Treatment’s Option and Duration | Intervention (s) (Concentration or Dose) | Outcome (s) | Pathway (s) |
|---|---|---|---|---|---|---|
| Sun et al., 2024 [94] | In vitro & In vivo | HT22 cells, APP/PS1 mice | In vitro: AdipoRon 48 hrs; In vivo: AdipoRon, 7 days | 1 mM in vitro; 1 μg in vivo | ↓Aβ deposition, ↑cognitive function, ↑autophagy, ↑Aβ clearance | ↑AdipoR1/AMPK-dependent nuclear translocation of GAPDH, ↑SIRT1 activation, ↓DBC1 interaction with SIRT1 |
| Wang et al., 2023 [95] | In vitro & In vivo | SH-SY5Y cells, P301S mice | In vitro: AdipoRon 48 hrs; In vivo: AdipoRon/corn oil, 4 months | 20 μM in vitro, 50 mg/kg in vivo | ↓Hyperphosphorylated tau, ↓memory deficits, ↓synaptic deficiencies, ↑mitochondrial fusion, ↓neuroinflammation | ↑AMPK/GSK3β activation, ↓tau (S396, T231) in vitro, ↑Mfn2, OPA1, ↑AMPK/SIRT3, ↑GSK3β in vivo |
| Khandelwal et al., 2022 [96] | In vitro & In vivo | Neuro2A cells, APP/PS1 mice | In vitro: AdipoRon 12 hrs; In vivo: AdipoRon/DMSO, 30 days | 10 μM in vitro, 50 mg/kg in vivo | ↑Glucose uptake, ↓insulin resistance, ↑insulin sensitivity, ↓cognitive deficits, ↓Aβ amyloid, ↓neuroinflammation | ↑GLUT4 translocation, ↑AKT, GSK3β, AMPKα phosphorylation in vitro, ↑GLUT1, GLUT4, ACCa, PGC1α, APOE, PSD-95, synaptophysin in vivo |
| Ng et al., 2021 [97] | In vitro & In vivo | HT-22 cells, 5xFAD mice | In vitro: AdipoRon 48 hrs; In vivo: AdipoRon/corn oil, 3 months | 5 μM in vitro, 50 mg/kg in vivo | ↑Neuronal insulin-signaling, ↑insulin sensitivity, ↑spatial/neuronal/synaptic memory, ↑learning, ↓neuroinflammation | ↑AMPK, ↓BACE1, NF-κB in vitro, ↑akts473, GSK3βS9, ↑akt, GSK3β phosphorylation, ↓IRS-1, Aβ, BACE1, sAPPβ, βCTF, IL-1β, TNF-α in vivo |
| He et al., 2021 [98] | In vitro & In vivo | N2a/APPswe cells, 5xFAD, APP/PS1, APN KO mice | In vitro: AdipoRon, 3-MA, CQ 24 hrs; In vivo: AdipoRon 2-4 months | 1 or 5 μM in vitro, 50 mg/kg in vivo | ↓Aβ deposition, ↓neuroinflammation, ↓cognitive impairment, ↓spatial memory deficits, ↑autophagy, ↑lysosomal activity | ↑AMPK-mTOR activation, ↑AdipoR1, APPL1 expression, ↓mTOR, TNF-α, IL-1β in vitro & in vivo |
| Liu et al., 2020 [99] | In vitro & In vivo | NE-4C cells, APP/PS1 mice | In vitro: AdipoRon 48 hrs; In vivo: AdipoRon/vehicle injection, 14 days | 5 μM in vitro, 1 μg in vivo | ↓Aβ-induced neuronal injury, ↓neuroinflammation, ↑cell viability, ↑cognitive function, ↓Aβ deposition | ↑NSC proliferation, ↑AMPK, CREB activation, ↑AMPK/CREB protein expression, ↓ΔΨm dissipation in vitro & in vivo |
Abbreviations: ↑, Increase; ↓, Decrease; 3-MA and CQ, Autophagy Inhibitors; ΔΨm, The Changes of Relative Mitochondrial Membrane Potential; βCTF, β-C-Terminal Fragment; ACC, Acetyl-CoA Carboxylase; Aβ, Amyloid Beta; AMPK, AMP-Activated Protein Kinase; AKT, Protein Kinase B; APOE, Apolipoprotein E; APPβ, Amyloid Precursor Protein Beta; APPL1, Adaptor Protein, Phosphotyrosine Interacting with PH Domain and Leucine Zipper 1; AR, AdipoRon; BACE1, Beta-Secretase 1; CREB, cAMP-Response Element Binding Protein; DMSO, Dimethyl Sulfoxide; GLUT1, Glucose Transporter 1; GLUT4, Glucose Transporter 4; GSK3β, Glycogen Synthase Kinase 3 Beta; Iba1, Ionized Calcium-Binding Adapter Molecule 1; IL-1β, Interleukin-1 Beta; IRS-1, Insulin Receptor Substrate 1; Mfn2, Mitofusin 2; mTOR, Mammalian Target Of Rapamycin; NF-κB, Nuclear Factor kappa B; NSC, Neural stem cells; OPA1, Optic atrophy 1; PGC1α, Peroxisome Proliferator-Activated Receptor-Gamma Coactivator 1; PSD-95, Postsynaptic Density 95; SIRT3, Sirtuin 3; Sp1, Specificity Protein 1; TNF-α, Tumor Necrosis Factor Alpha.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



