
Submitted:
09 December 2024
Posted:
10 December 2024
You are already at the latest version
Abstract
Adeno-associated viruses (AAVs) are non-pathogenic, replication-deficient viruses that have gained widespread attention for their application as gene therapy vectors. While these vectors offer high transduction efficiency and long-term gene expression, the host immune response poses a significant challenge to their clinical success. This review focuses on the obstacles of evaluating humoral response to AAVs. We discuss the problems with the validation of in vitro tests, and the possible approaches to overcome them. Using published data of neutralizing titers of AAV serotypes, we built the first antigenic maps of AAV in order to visualize the antigenic relationships between varying serotypes.
Keywords:
AAV
; neutralizing antibodies
; antigenic cartography
; anti-AAV antibodies
; humanized models
; NHP models
; mice models
1. Introduction
Adeno-associated viral vectors are thought to be a perspective delivery platform for gene therapies due to their high transduction and long-term expression efficiency in different cell types, non-pathogenic nature, and low immune responses in humans [1,2]. AAVs are unknown to induce any human diseases [3], however, they naturally circulate in certain host animals. People may be spontaneously infected by AAVs during life, which can lead to increased levels of AAV-specific serum antibodies [4,5]. As a result, up to 80% of the human population has neutralizing antibodies to AAVs [6], with humoral response to AAV2 being most prevalent [7]. T-cell-mediated immune responses to AAV are rarely detected in treatment-naive individuals because of low sensitivity tests or other reasons [8,9,10,11,12]. However, this review is focused on humoral responses to AAV and methods of evaluating antibody titers to AAV.
Pre-existing humoral immune response is thought to be a deleterious factor for AAV-based gene therapy because of the neutralization vector caused by specific serum antibodies [13,14,15]. A high level of pre-existing anti-AAV antibodies may promote phagocytosis and complement activation [14]. Phagocytized vectors are not processed to transgene expression [16]. As a result, a higher dose of drug is required to achieve therapeutic goals. According to the FDA, high titers of anti-AAV antibodies are an exclusion criteria in the gene therapy clinical trials in which systemic administration is used [17].
Over 30 clinical studies of AAV-based gene therapy (GT) that reported anti-AAV serum antibody titers were analyzed in this study. We found that cutoff titer values varied from study to study, regardless of local or systemic vector administration, the method of measuring titers, or the serotype selected for GT delivery. However, in order to fairly compare cutoff titers from different clinical trials, antibody titer tests need to have been conducted in similar conditions. We found that the protocols of these assays are often not detailed enough in descriptions of clinical trials or scientific papers. The differences in providing these assays have direct influence on the titer values.
The correct determination of anti-AAV titer is important not only for definition of the exclusion criteria for GT, but also when comparing humoral responses to different serotypes of AAV. A serotype-switching approach by using low cross-reactive wild-type AAVs or synthetic AAVs may be used to overcome a preexisting humoral response [18]. In such cases, direct comparison of sera antibody activity against serotypes of interest is paramount. ELISA and in vitro neutralization assays with permissive cell lines are the most common methods used to determine the level of anti-AAV antibody response [19]. Whereas ELISA is relatively easy to standardize, in vitro neutralization assays have limitations for AAV. Here, we discuss the theoretical basement of the methodological problems of modern in vitro assays. Particularly, we show why in in vitro neutralization assays the ratio of full and empty capsids is important, how the ratio of serum antibodies and used viral particles influences the result, and why the differences in transduction efficiency of cell lines between AAV serotypes are a crucial problem with this assay [20,21]. Some recommendations to overcome these problems are proposed.
The other approach to determining humoral response to AAV is to use animal models with a humanized immune system. Regardless of the obvious difficulties in the usage of animals, there is a possibility for AAVs to transduce different cell types in the whole organism and interact with specific antibodies or immune cells [22]. In this paper, we have reviewed animal models that are used to study human-like humoral responses to AAVs with subsequent in vitro neutralization assays. Moreover, we describe an animal model approach that allows the complete avoidance of in vitro neutralization assays for the determination of human sera neutralization titers to AAV.
Although animal models have several advanced compared to in vitro assays, the last one is more suitable for screening assays and more easy to standardize [23]. The previously listed disadvantages of most current in vitro assays influence the level of transduced target cells, which are used to evaluate the neutralization titers. As a result, different neutralization titers reflect not the optimized conditions of the assay, but the antigenic characteristics of different serotypes. However, if antibody titers to different AAV serotypes are obtained in similar conditions, their antigenic characteristics can be evaluated and visualized with various methods, such as antigenic cartography [24]. In the case of AAV serotypes, such maps may be useful in order to reveal the direction in which novel synthetic AAV synthesis should be taken to create AAV that will antigenically differ from the wild type, to which pre-existing immunity is often present. As far as we know, no AAV antigenic maps have been created yet. Here, we constructed an AAV antigenic map using previously published neutralization titer data.
The present review discusses the pros and cons of current in vitro assays and animal models to detect human humoral immune responses to AAV and proposes possible improvements. Overcoming limitations of current test systems will allow for better analysis and comparison of data in order to overcome pre-existing humoral responses. This makes the review relevant to researchers and clinicians seeking to enhance the safety and efficacy of gene therapies.
2. Anti-AAV Abs Generation
AAVs transduce a wide spectrum of target cells, including antigen-presenting cells (APCs) [25,26]. In in vitro experiments, AAVs were uptaken predominantly by monocyte-derived dendritic cells (moDCs) and by monocytes, conventional and plasmacytoid DCs, and neutrophils to a lesser degree [14]. AAV proposed PAMPs, such as capsid proteins, and CpG in viral genomes [27], interact with PRRs (TLR2, endosomal TLR9, respectively) on or within the cells that trigger an inflammatory reaction and maturation of the dendritic cells, their migration to the lymph nodes [8] and presentation to naive CD8+ or CD4+ T cells. Jamie L. Shirley et al. provided some insights into this process. They showed that activation of TLR9 on pDCs, in which it is highly expressed, led to type 1 interferon production that, in combination with costimulation of CD40L, activated maturation of conventional DCs (cDCs) and presentation capsid antigens in their MHC I [28]. After endocytosis by target cells or APCs, AAVs undergo proteasomal degradation following endosomal escape [29], capsid phosphorylation, and ubiquitination [28]. As a result, capsid antigens are presented onto MHC class I or cross-presented onto MHC II. Activated APCs trigger specific T cell response. Activated CD4+ Tfh cells induce B cell activation and antibody secretion. Also, in in vitro and in mouse model, it was shown that AAV capsid triggers IL-1β– and IL-6–dependent B cell differentiation and specific IgM and IgGs secretion [30].
In fact, the process of humoral response to AAVs induction is not well studied. There are certain challenges with studying this process due to the absence of an acute inflammation stage after infection with AAVs in humans. However, the presence of a humoral response to AAVs was repeatedly shown, and in adults the level of this response is higher than in children [5]. The level of pre-existing anti-AAV Abs is crucial in AAV-based gene therapies. For example, recently, it was proven in vitro that high titers of anti-AAV NAbs trigger complement activation that may determine thrombotic microangiopathy [14].
3. Anti-AAV Antibody Titers as Exclusion Criteria in Clinical Trials
Pre-existing AAV-specific antibodies hamper efficient transduction of AAV and potentially lead to decreased delivery of the gene therapy candidate [6]. No correlation has been shown between anti-AAV Ab levels and adverse events in gene therapy (GT) studies [31]. However, that may be explained by the fact that, in at least 45 percent of the U.S. Food and Drug Administration (FDA)-approved GTs with systemic injection of AAV, patients with pre-existing Abs to AAV are often excluded from clinical trials [32]. The enrollment criteria have been set at either <1:10 anti-AAV NAb titer or <1:100 total anti-AAV-IgG titer [16].
Exclusion of patients from clinical trials with high levels of anti-AAV antibodies greatly depends on the therapeutic area. The deleterious impact of pre-existing antibodies on the therapeutic effect is expected to be more significant when the administration is systemic, as compared to local administration [32]. A meta-analysis by Hau Kiu Edna Au et. al. (2022) showed that almost 90% of blood disease studies exclude patients with pre-existing antibodies, while only <10% of eye and 21% of CNS disease studies exclude such patients [18]. However, FDA guidance recommends exclusion of seropositive patients for both local and systemic administration [33,34,35,36].
Titer exclusion criteria differ between studies. In Table 1, we summarized this data from various clinical studies. Trials were extracted from the U.S National Library of Medicine database (ClinicalTrials.gov), the largest clinical trials database to date, following the keyword search for “AAV,” using a cut-off of 23 July 2021. Only studies with exclusion criteria for the presence of Nabs in AAV were selected. It should be noted that these titers seemingly do not show absolute titers of ADA (Anti-Drug Antibody).
Table 1 shows heterogeneity in the approaches to anti-AAV determination: some studies use ELISA to measure binding antibodies, while others use neutralization tests with cell cultures to measure NAbs or transduction inhibition tests. Titer values vary from 1:2 to 1:20 for neutralizing antibody titers and from 1:50 to 1:1600 for binding antibody titers, which confirms the lack of uniform standards for this parameter. The insufficiently precise wording in exclusion criteria in a number of clinical studies is also questionable.
In most clinical trials presented in Table 1, there is no information on how titers were defined. Apparently, it is a serum dilution that exceeds the cut-point value, which was also not defined. By the recommendation of the FDA, the cut-point of the level of ADA should be determined using samples from treatment-naïve subjects [37]. Bioanalytical laboratories establish the cut-point for antibody assessment if 1-10% of the tested treatment-naive samples are false-positive [31].
Detection of binding or neutralizing antibodies before and after drug injection is required to understand the immunogenicity, safety, and efficacy of gene therapy products based on AAV. It would be beneficial to have standard protocols in order to compare the data obtained from different studies of the immunogenicity of AAV vectors.
As Table 1 shows, ELISA and in vitro neutralization tests are most used in clinical trials to determine anti-AAV Ab titers. Although some studies show that binding and neutralizing AAV-specific IgGs correlate well [11], measuring NAbs is thought to be more desirable. Currently used assays for detecting anti-AAV NAbs include in vitro assays, in vivo models, and ex situ assays (Figure 1). In vitro assays are suitable for screening assays due to their simplicity and quickness. However, in scientific areas where more precise characteristics of a vector are required, in vivo models and ex situ assays may be used to obtain more information about the vector of interest.
We discuss below the advantages and troubles of the current assays used for evaluating serum NAbs to AAV.
4. In Vitro Neutralization Assay
The NAb titer to AAV is one of the most important criteria used to exclude patients for clinical trials and gene therapy treatment. Most clinical trials use in vitro assays to test NAb levels in patients’ sera, because they are easy to set up and give consistent results [38].
The choice of target cell line is one of the important factors having influence on the results of the in vitro virus-neutralization assay (VNA). For SARS coronaviruses, the key cell receptor (ACE-2) is well-characterized [39]. Since the beginning of the COVID-19 pandemic, it has been relatively quickly shown that ACE-2 is a key cell receptor for the novel SARS-CoV-2 virus [40]. It enabled the rapid development and validation of a good in vitro model to detect anti-SARS-CoV-2 NAbs. Engineered cells with stable expression of ACE-2 are target cells in this assay [41]. Then, expression of TMPRSS was added to target cell lines [42] and now widely used in scientific studies and clinical trials.
Unlike SARS-CoV-2, there are no well-defined key cell receptors that are responsible for AAV transduction to the cells. Thus, glycan receptors are suggested to be low-specificity attachment factors, but they are not ‘primary receptors’ [43]. The potential key role of AAVR is also discussive [44]. Because of it, different cell lines are used in in vitro neutralization assays of AAVs. НЕК293, Hela, COS-7, GM16095, and Huh7 cell lines are most used to determine the level of anti-AAV NAbs in vitro today, although transduction efficiencies of these cell lines vary by up to 5 orders of magnitude among different AAV serotypes [45,46]. Low transduction efficiency of several serotypes makes it impossible to determine NAb titers, due to the limit of detection. Higher multiplicity of infections (MOIs) can be used to overcome this problem, but it leads to underestimating NAb titers [38]. Moreover, lower MOIs in terms of in vitro VNAs enable cost reduction of the experiment because they require a lower number of AAV. As a result, more permissive cell lines suitable for transduction by most serotypes are highly desirable.
To improve transduction efficiency, target cells may be infected with adenovirus 5 before the neutralization assay [4,47,48]. 2V6.11, a cell line obtained from HEK293, overexpresses the adenovirus gene, E4 ORF, under the control of the ecdysone-inducible promoter. It was shown [19] that AAV6, 8, and 9 transduce this cell line more effectively than Huh7, HEK293, and HeLa, respectively. Hoi Yee Chow et al. established a Hela cell line with stable overexpression of AAVR and showed increased sensitivity of NAbs detection for different AAV serotypes [46].
The choice of cell line for in vitro neutralization assays is not the only important choice. Typically, VNAs consist of 2 steps: incubating viral particles with diluted serum and adding this mixture to target cells. Choosing the optimal dose of viral vector is pivotal when the goal is to compare neutralization titers against different serotypes. 3 VNT methods, and their limitations, are described below. Ultimately, good principles have been proposed. These principles are applied to VNA in general, in which other viruses are used.
The first and most wide-spread approach is to use a similar number of viral particles (measured by viral genomes, for example). The first limitation of this approach is that it does not consider the number of full and empty capsids. Empty particles compete with full AAV particles for binding to serum NAbs [47] and reduce the number of NAbs, blocking functional (full) AAVs. As a result, a large quantity of full AAVs can transduce target cells and express the reporter transgene, which will influence titration curves and neutralization titers.
Figure 2.
A figure illustrating the importance of considering the number of full and empty capsids in neutralization tests. Serotype A and serotype B are taken in equal numbers of viral particles measured by viral genomes. However, the serotype B sample contains a lower percentage of full capsids than serotype A. AAV NAbs bind not only full capsids, but also empty capsids, so the number of cells transduced by serotype B is higher.
Figure 2.
A figure illustrating the importance of considering the number of full and empty capsids in neutralization tests. Serotype A and serotype B are taken in equal numbers of viral particles measured by viral genomes. However, the serotype B sample contains a lower percentage of full capsids than serotype A. AAV NAbs bind not only full capsids, but also empty capsids, so the number of cells transduced by serotype B is higher.
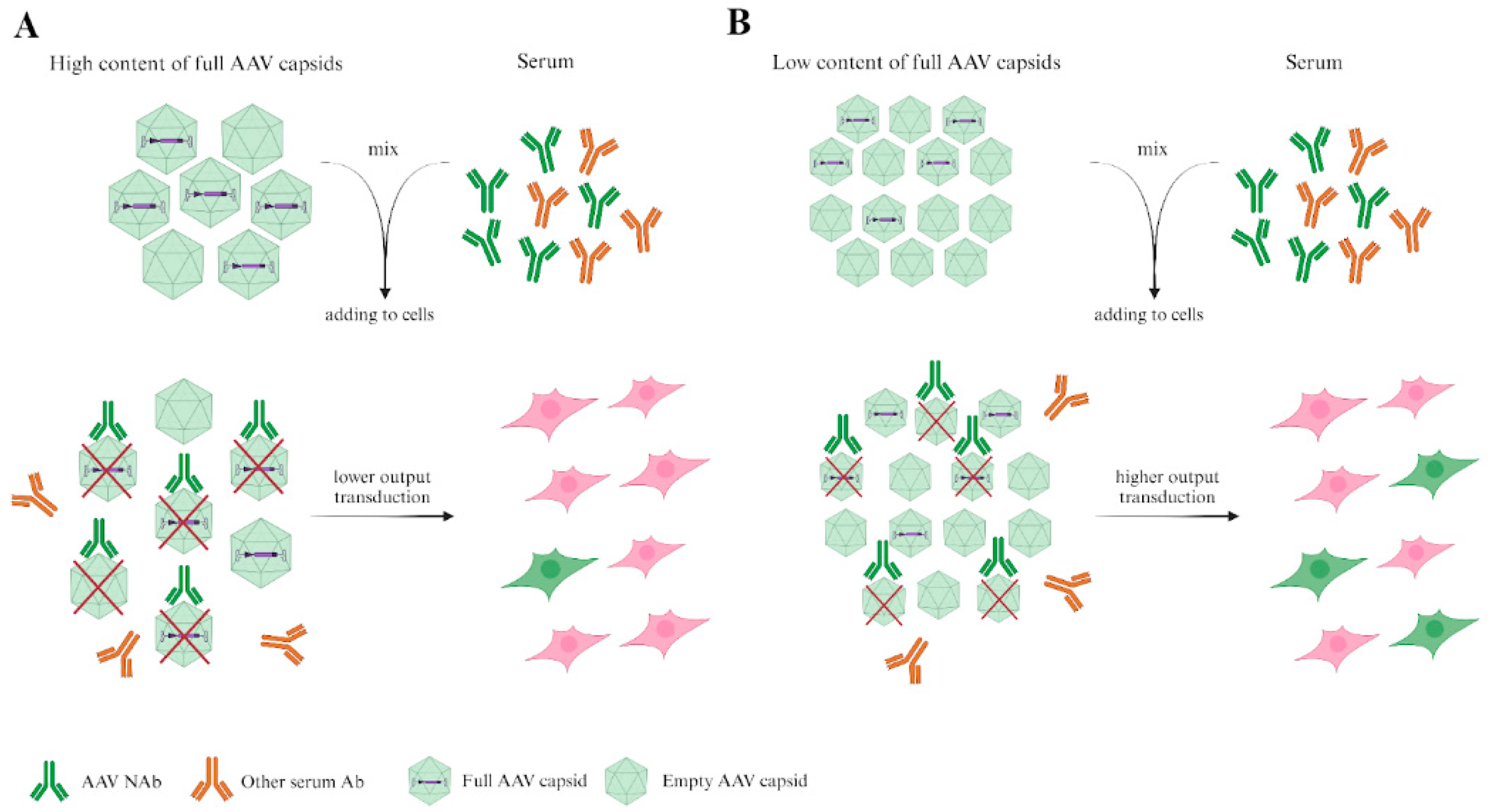
For this reason, we propose that quality control tests of virus particles (VPs) are required. Tests of the ratio of full/empty capsids and the presence of VP aggregates may be appropriate. Taking viruses with a similar ratio of full/empty capsids is a good practice.
The second approach is to use the number of viral particles (VPs) that transduce a similar percentage of target cells [49]. This approach is limited by the varying ability of different serotypes to infect cells. Thus, in order to achieve the same level of transduction for different serotypes, it is necessary to select a number of VPs, which, in some cases, may differ by an order of magnitude. It is very important for chimeric AAV, which are often purposefully constructed to improve the abilities of transduction target cells [50,51,52]. Sera samples are traditionally serially 2-fold diluted and mixed with the chosen viral dosage. However, if the chosen dosages of viruses are dramatically distinguished, the ratio of serum antibodies and viral particles will be dramatically different. The lower number of VPs will require a lower number of NAbs to be neutralized, which will also have an influence on neutralization titers against different serotypes.
Thus, our recommendation is to use an equal ratio between the number of VPs and the volume of serum samples. The limitation that has affected both of the above approaches is the differing transduction levels of AAV serotypes. In our opinion, this limitation is the most difficult to overcome. If an equal number of AAV is used in an experiment for different serotypes, the transduction level may vary from 0 to 100% for different serotypes. Low transduction levels (about 10%) increase the significance of mistakes related to measurement, which will have an influence on titration curves and neutralization titers. If the number of transduced cells is close to 100%, it is not always clear which number of AAVs transduce cells in abundance. The appropriate transduction level for different serotypes may not always be achieved. Alternatively, if transduction level is used as a metric, then the same transduction level may be achieved by vastly different levels of VPs, with variances as high as in degrees of magnitude.
Figure 3.
A figure illustrating the importance of the variance in the MOIs of AAV serotypes. Chosen doses of serotype A and serotype B transduce an equal number of target cells in the absence of serum antibodies (not shown in figure). However, in the presence of equal volume/number of specific AAV antibodies from serum, the levels of transduced target cells are different, which reflects the lower obtained viral dosage of serotype A, rather than the difference in antigenic characteristics between the two serotypes.
Figure 3.
A figure illustrating the importance of the variance in the MOIs of AAV serotypes. Chosen doses of serotype A and serotype B transduce an equal number of target cells in the absence of serum antibodies (not shown in figure). However, in the presence of equal volume/number of specific AAV antibodies from serum, the levels of transduced target cells are different, which reflects the lower obtained viral dosage of serotype A, rather than the difference in antigenic characteristics between the two serotypes.
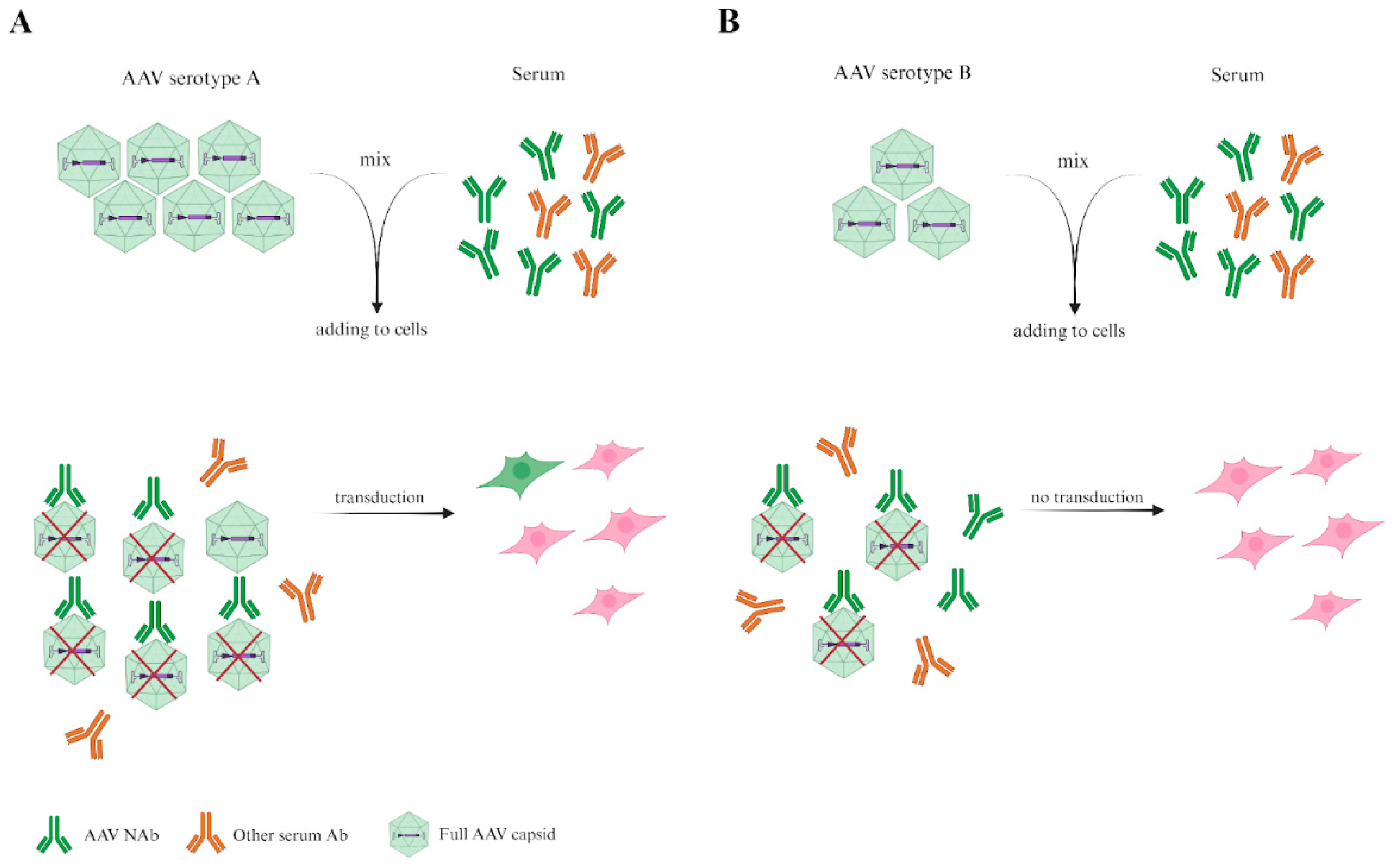
As previously discussed, incubation with an equal ratio of VPs and serum volume is needed. Thus, we suggest incubating VPs with serum and transferring the volume of this mixture, which, in the absence of serum, would transduce the necessary percentage of target cells. However, following this recommendation becomes challenging when MOIs differ by more than three orders of magnitude.
5. Determination of Immune Response to AAVs in Animal Models
As discussed above, the most difficult limitation to overcome in in vitro assay is the different transduction efficiency of cell lines. It should be underlined that this limitation is only relevant when the goal is to compare NAb responses to different AAV serotypes. The usage of animal models may provide extra insights in studying NAb response to AAVs and expand the methods used to determine the level of these NAbs.
A study by Mingozzi et al. (2018) demonstrated that, despite promising outcomes in animal models, human trials of AAV gene therapy have encountered unexpected immune reactions [55]. These differences highlight a challenge in translating findings about immune responses to AAVs from animal models to humans [56].
Mice and non-human primates (NHPs) are the most widespread animal models that are used to study immune response to AAV. However, the composition and role of immune cells differ between humans and mice [57]. Mestas and Hughes (2004) highlighted that human immune systems contain unique immune cell subsets not found in mice (CXCR1, CD58 (LFA-3), CD40 on EC, etc.), which can affect the study of viral infections and the immune response to AAV vectors [57]. NHP models have demonstrated the potential for unexpected immune reactions, including the activation of memory T cells and antibody responses that were not fully predicted by rodent models [58]. Despite their advantages, NHPs are not without limitations [59]. Rhesus macaques display a higher number of certain immune cells, such as CD4+/CD8+ double-positive T cells, than humans [60]. They may react differently to AAV vectors compared to cynomolgus macaques, leading to variability in study outcomes [59,61,62]. Furthermore, in vaccine studies, while NHPs like macaques often develop strong NAb responses, their immune system doesn’t always mimic human long-term immunity or antibody decay rates, as seen in COVID-19 vaccine research [63]. Thus, there is a need to develop more appropriate systems to test human or human-like immune responses to AAVs.
Humanized mouse models would be valuable in studying the immune response to AAVs. These models aim to address the limitations of traditional animal models, offering more accurate predictions of human immune responses. Studies using these mice have shown that, while humanized models can replicate some aspects of the human immune response, such as T cell activation and antibody production, they may not fully recapitulate the complexity of human immune reactions to viral infections, particularly those involving regulatory T cells and innate immune cells like plasmacytoid dendritic cells (pDCs) [64].
The breakthrough mouse model with a complete, functional human immune system and human-like gut microbiome was created by the Paolo Casali group [65]. The most interesting result demonstrated in this model, in terms of detection of human humoral response, is that these (truly humanized) THX mice showed mature neutralizing antibody responses to Salmonella Typhimurium and SARS-CoV-2 virus after vaccination with Salmonella flagellin and the Pfizer COVID-19 mRNA vaccine, respectively. It would be desirable to test NAb formation that will reflect the human humoral response in these mice to other viruses, including AAVs.
Knock-in mice engineered to express human immune components offer a controlled system for studying, including AAV interactions. For instance, different studies used knock-in mice with human immunoglobulin loci that provided further insight into human-specific immune responses [66,67,68].
To summarize, the previously described animal models reflect human immune responses and develop the humoral response to AAV after vector injection to varying extents. However, induced humoral response to AAV is usually measured by in vitro assays, the limitations of which were also discussed above. Next, we describe animal models that allow the measurement of human-like NAb responses, fully avoiding the in vitro neutralization tests.
This approach involves inducing a human-like humoral immune system in animals, followed by AAV injection. The effectiveness of vector neutralization is compared to the same line of mice that weren’t humanized.
In a study by Lan Sun et al. [69], researchers generated a mouse model of passive immunity to quantitatively assess anti-AAV8 NAb titers. C57BL/6 mice were first injected with rhesus macaque sera followed by scAAV2/8.CB.hAAT. The level of anti-hAAT Abs was measured by ELISA and inversely corresponded, according to the authors’ study design, to the level of anti-AAV NAbs. This study showed that this in vivo NAb assay was more sensitive than an in vitro NAb assay; serum samples with low titers <1:5 NAbs were detected as positive to the anti-AAV8 response only by in vivo assay. Lili Wang et al. introduced this approach, but they transferred monkey sera to C57BL/6 mice prior to injection of AAVs with cFIX as a [70]. cFIX expression levels in mouse plasma were indicators of AAV neutralization.
Thus, in the first example, the level of NAbs to AAV was evaluated relative to the anti-transgene antibody level, whereas in the second example, it was relative to transgene expression. The level of viral DNA and/or transgene RNA may be included in the study design when this approach is used.
To summarize, animal models may be used in two ways. The first way is to use animals with a modified immune system or animals with an immune system close to human, inject AAV and measure NAbs in in vitro tests. In other words, this method may be used without the necessity for in vitro neutralization tests, which expands the possibilities when it comes to the detection of AAV NAbs.
6. Ex Situ Assays with Whole Perfused Explants
The whole human explant offers another valuable perspective for evaluating the level of NAbs to AAVs. In this context, Marti Cabanes-Creus et al. (2024) introduced an ex situ system of normothermic human liver explants. One of them was perfused with human blood in the presence of anti-AAV NAbs that contained an AAV library [71]. The other liver explant was perfused with the AAV library only. The level of AAV neutralization corresponded to differences between two explants in the level of viral DNA and transgene cDNA, as determined by NGS. This method did not require in vitro neutralization assays.
This model is characterized by the ability to preserve the structure of human tissue, deliver the drug through a natural network of capillaries, and observe the neutralization of AAV in the presence of the resident macrophages, Kupfer cells. This allows plasma to be passed through the perfusion system under conditions close to physiological ones. The authors showed that, in the presence of human plasma containing anti-AAV NAbs, vectors derived from AAV2/AAV3b were extensively neutralized, unlike AAV8-derived variants. The authors noticed that AAV were rapidly cleared from perfusate if the explant contained NAb to the serotype. This may have been mediated by Kuppfer cells and could not be represented in vitro.
In general, the use of ex situ normothermal perfusion of human organs opens up prospects for studying the neutralization of AAVs in an environment containing different tissues and cell types, including the resident immune cells of the organ. Besides the liver, models of isolated hearts perfused with AAV already exist [72,73], and may potentially be used for detecting neutralizing antibody activity if human plasma is added to the perfusing solution.
Ex situ perfusion of human organs with NAbs and AAVs has several limitations. The availability of donor material is both generally limited, then further restricted by exclusion criteria for transplantation. The maintenance of the explants in a viable state for several days is also a difficult methodological task. Upon connection to the perfusion unit, donor organs may undergo reperfusion damage and fragmentation, compromising the capillary structure and, subsequently, the virus’ distribution pathway. It is shown that, after six days of perfusion, markers of necrosis in the liver explant increased and organ functionality declined [74]. Moreover, the normothermic perfusion explant model is limited by the absence of circulating immune cells, which may potentially be involved in AAV response. The advantages of this model include that it does not require in vitro neutralization tests and partially reflects the interaction of AAVs with cells with immune function as it occurs in the whole organism.
7. Antigenic Map of AAVs Created on Published Data
The surface capsid proteins of different variants (strains, serotypes, isolates) of the same virus always have a certain homology in sequence and structure [43,75,76]. Such homology, which is closely linked with evolutionary relationships, may result in similarity of the physicochemical properties of the protein surface. This leads to cross-reactivity, when antibodies formed by the host in response to one variant of the virus neutralize another variant.
Despite several attempts to predict cross-reactivity in silico [77,78,79], the gold standard of antigenic similarity estimation is mass assessment of neutralizing properties of diverse serum samples. The serum samples can be taken from model animals immunized with a specific variant. However, as previously discussed, animals can develop similar, but not identical humoral responses to humans, which leads to unpredictable deleterious immune responses in AAV-based GTs clinical trials. Sera from conditionally healthy (not infected) individuals or who have been infected by a known virus serotype are preferable [80], but in the case of AAVs, this is not achievable. Titer neutralization data obtained from individuals who have been vaccinated with an AAV GT drug based on a known serotype provide sufficient data about the formation of cross-reactive antibodies.
Animal or human serum neutralization capacity is usually tested in vitro, the advantages and limitations of which were discussed above. If these limitations are overcome or compromises are found, the reliable comparison of humoral responses to different AAV serotypes becomes possible. It is highly preferred in studies in which, for example, chimeric AAVs are developed with the purpose of evasion of pre-existing humoral responses [45,81,82,83].
Over the past decades, many different studies have been conducted that have assessed the prevalence of neutralizing antibodies against various AAV serotypes in diverse human populations [4,7,84]. Such investigations not only provide critical information about the percentage of seropositive individuals (and hence the potential for clinical use of a particular virus variant), but also allow the assessment of antigenic similarity between different AAV serotypes. However, this data is often segmental and difficult to visualize and analyze.
Previously, this problem appeared when cross-reactive immune response to influenza virus strains or SARS-CoV-2 variants and was solved by the antigenic cartography approach developed by Dereck [85,86,87,88,89,90]. This approach visualizes a large dataset of titers for several serotypes on a map. The closer the serotypes from each other are located on the antigenic maps (represented by areas), the more antigenically similar they are. Sera from various patients, represented by circles, are closer to the variants that they better neutralize. One grid line on the map corresponds to a twofold dilution in the virus neutralization assay and is called an “antigenic unit”.
Here, we collected data from three publications in which neutralization titers of sera were performed [91,92]. The description of cohorts and AAV serotypes for which neutralization titers are performed in each of these studies is summarized in Table 2.
Titer data was analyzed using the Racmacs R package [93] and plotted on an antigenic map (Figure 4). Areas consider experimental and statistical uncertainty of their location determined by both pairwise similarity (ability to neutralize the same antigens) and proximity to related antigens.
The map, in general, represents some expected patterns. For example, AAV2 is located in the center of the sera samples cloud as the variant with the most common neutralization in the human population. AAV5 and AAV2 are distant in the antigenic map. It is known that these two variants have low sequence homology [94]. Here, we show that they may also be antigenically distinct.
Figure 4.
Antigenic map of AAVs constructed on published titer neutralization data. AAV serotypes are represented by areas, sera by circles. Areas consider experimental and statistical uncertainty of their location. One grid line corresponds to one antigenic unit.
Figure 4.
Antigenic map of AAVs constructed on published titer neutralization data. AAV serotypes are represented by areas, sera by circles. Areas consider experimental and statistical uncertainty of their location. One grid line corresponds to one antigenic unit.
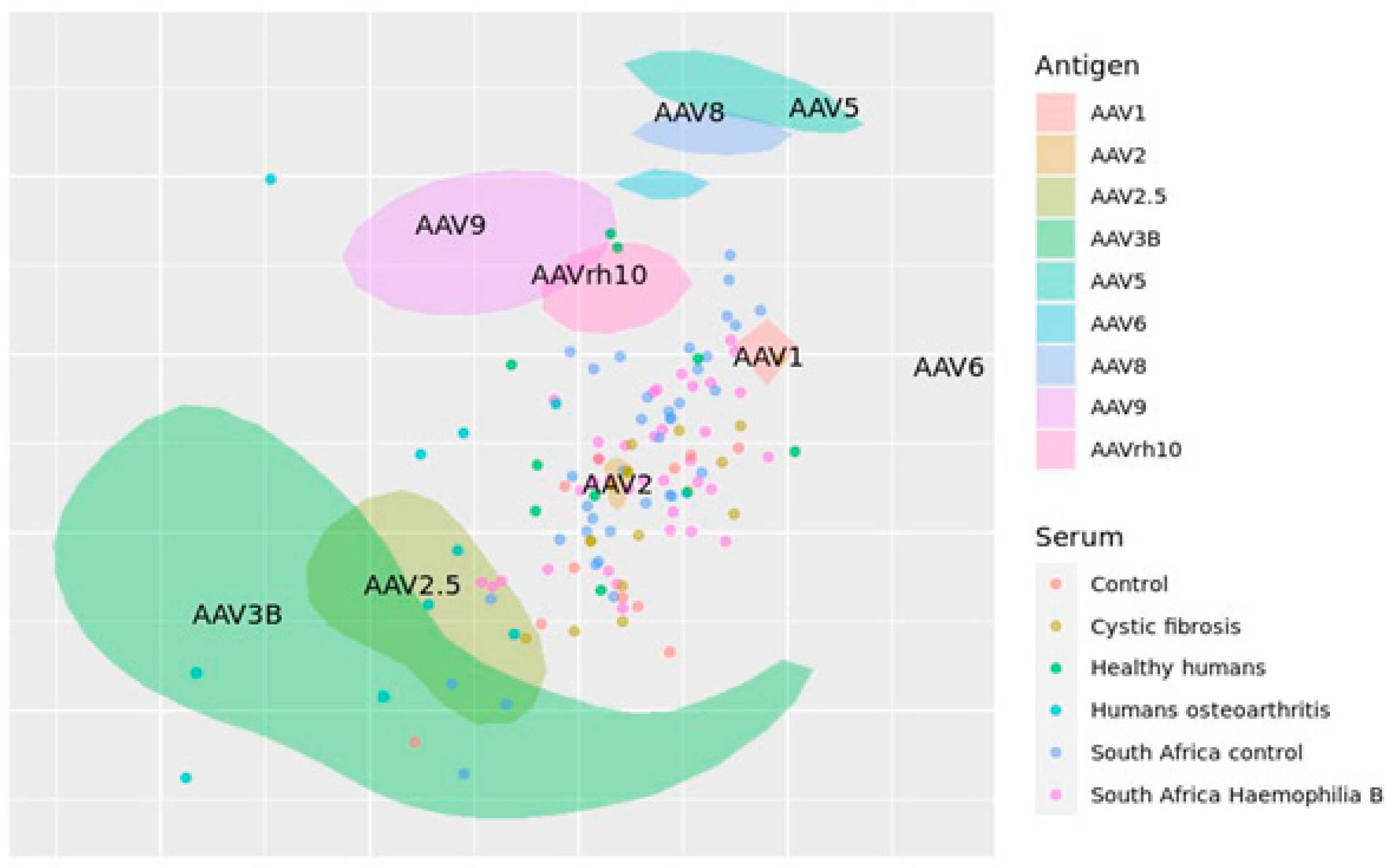
It should be noted that the represented antigenic map has several limitations. Firstlyl, the used titers were obtained in different conditions in different laboratories. The number of studies we included in our analysis is low due to the limited availability of the raw data of neutralization titers in publications. In a study by Temilola Abdul et al, individuals with autoimmune disease (osteoarthritis) were included, although the humoral response may differ from healthy donors [92].
Taken together, for robust and reliable determination of the antigenic relationship between AAV serotypes, a systematic study is needed. This study must include many AAV variants (ideally, all variants of clinical importance) and a large homogeneous cohort of healthy individuals that are tested simultaneously. The limitations of neutralization assays should be reduced to a minimum with the recommendations we previously discussed. Such a study would provide valuable information for subsequent capsid design to evade neutralization by pre-existing antibodies.
8. Prospects and Future Directions
Pre-existing humoral immune response to AAV is a well-known limitation of GT application. The measurement AAV NAb titers and the determination of the threshold level, acceptable in GT, remain discussive. Here, we have shown the differences in antibody titers that are exclusion criteria for GTs. They may be explained by the absence of standardization of the assays used for determination of specific anti-AAV Abs. We focused on the details of these assays in the context of AAV, and reviewed alternative approaches to NAbs titer assessment, aside from the most popular in vitro assays. A more precise description of methods used in anti-AAV NAbs titer assessment and following good practice recommendations that we introduced here are necessary to increase the relevance and homogeneity of studies. The reliable comparison of titers to different AAV serotypes could help researchers design more effective vectors that avoid pre-existing immunity. The antigenic mapping approach is suitable for visualizing antigenic relationships between AAV serotypes. However, more reliable titer data are required to construct a more precise antigenic map.
Author Contributions
Conceptualization, D.A.N., O.N.M. and P.Y.V.; writing—original draft preparation D.A.N., T.K., D.M., E.A.A.; visualization, D.M. and E.A.A.; writing—review and editing, D.A.N., E.A.A., O.N.M. and P.Y.V. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by the Ministry of Science and Higher Education of the Russian Federation (agreement # № 075-03-2022-107/10); 2. This work was supported by the Russian Science Foundation (Grant № 23-64-00002).
Conflicts of Interest
The authors declare no conflicts of interest
References
- Kotterman, M.A.; Schaffer, D. V. Engineering Adeno-Associated Viruses for Clinical Gene Therapy. Nat. Rev. Genet. 2014, 15, 445–451. [CrossRef]
- Gaj, T.; Epstein, B.E.; Schaffer, D. V. Genome Engineering Using Adeno-Associated Virus: Basic and Clinical Research Applications. Mol. Ther. 2016, 24, 458–464. [CrossRef]
- Flotte, T.R.; Carter, B.J. 6 Adeno-Associated Viral Vectors. In Gene Therapy Technologies, Applications and Regulations; Anthony Meager, Ed.; 1999; pp. 109–125 ISBN 0471976709.
- Kruzik, A.; Fetahagic, D.; Hartlieb, B.; Dorn, S.; Koppensteiner, H.; Horling, F.M.; Scheiflinger, F.; Reipert, B.M.; de la Rosa, M. Prevalence of Anti-Adeno-Associated Virus Immune Responses in International Cohorts of Healthy Donors. Mol. Ther. Methods Clin. Dev. 2019, 14, 126–133. [CrossRef]
- Li, C.; Narkbunnam, N.; Samulski, R.J.; Asokan, A.; Hu, G.; Jacobson, L.J.; Manco-Johnson, M.L.; Monahan, P.E.; Riske, B.; Kilcoyne, R.; et al. Neutralizing Antibodies against Adeno-Associated Virus Examined Prospectively in Pediatric Patients with Hemophilia. Gene Ther. 2012, 19, 288–294. [CrossRef]
- Louis Jeune, V.; Joergensen, J.A.; Hajjar, R.J.; Weber, T. Pre-Existing Anti-Adeno-Associated Virus Antibodies as a Challenge in AAV Gene Therapy. Hum. Gene Ther. Methods 2013, 24, 59–67. [CrossRef]
- Calcedo, R.; Vandenberghe, L.H.; Gao, G.; Lin, J.; Wilson, J.M. Worldwide Epidemiology of Neutralizing Antibodies to Adeno-Associated Viruses. J. Infect. Dis. 2009, 199, 381–390. [CrossRef]
- Ertl, H.C.J. T Cell-Mediated Immune Responses to AAV and AAV Vectors. Front. Immunol. 2021, 12, 1–11. [CrossRef]
- Hui, D.J.; Edmonson, S.C.; Podsakoff, G.M.; Pien, G.C.; Ivanciu, L.; Camire, R.M.; Ertl, H.; Mingozzi, F.; High, K.A.; Basner-Tschakarjan, E. AAV Capsid CD8+ T-Cell Epitopes Are Highly Conserved across AAV Serotypes. Mol. Ther. Methods Clin. Dev. 2015, 2, 15029. [CrossRef]
- Chirmule, N.; Propert, K.J.; Magosin, S.A.; Qian, Y.; Qian, R.; Wilson, J.M. Immune Responses to Adenovirus and Adeno-Associated Virus in Humans. Gene Ther. 1999, 6, 1574–1583. [CrossRef]
- Veron, P.; Leborgne, C.; Monteilhet, V.; Boutin, S.; Martin, S.; Moullier, P.; Masurier, C. Humoral and Cellular Capsid-Specific Immune Responses to Adeno-Associated Virus Type 1 in Randomized Healthy Donors. J. Immunol. 2012, 188, 6418–6424. [CrossRef]
- Patton, K.S.; Harrison, M.T.; Long, B.R.; Lau, K.; Holcomb, J.; Owen, R.; Kasprzyk, T.; Janetzki, S.; Zoog, S.J.; Vettermann, C. Monitoring Cell-Mediated Immune Responses in AAV Gene Therapy Clinical Trials Using a Validated IFN-γ ELISpot Method. Mol. Ther. Methods Clin. Dev. 2021, 22, 183–195. [CrossRef]
- Scallan, C.D.; Jiang, H.; Liu, T.; Patarroyo-White, S.; Sommer, J.M.; Zhou, S.; Couto, L.B.; Pierce, G.F. Human Immunoglobulin Inhibits Liver Transduction ByAAV Vectors at Low AAV2 Neutralizing Titers in SCID Mice. Blood 2006, 107, 1810–1817. [CrossRef]
- Smith, C.J.; Ross, N.; Kamal, A.; Kim, K.Y.; Kropf, E.; Deschatelets, P.; Francois, C.; Quinn, W.J.; Singh, I.; Majowicz, A.; et al. Pre-Existing Humoral Immunity and Complement Pathway Contribute to Immunogenicity of Adeno-Associated Virus (AAV) Vector in Human Blood. Front. Immunol. 2022, 13, 1–14. [CrossRef]
- Calcedo, R.; Franco, J.; Qin, Q.; Richardson, D.W.; Mason, J.B.; Boyd, S.; Wilson, J.M. Preexisting Neutralizing Antibodies to Adeno-Associated Virus Capsids in Large Animals Other Than Monkeys May Confound in Vivo Gene Therapy Studies. Hum. Gene Ther. Methods 2015, 26, 103–105. [CrossRef]
- Meadows, A.S.; Pineda, R.J.; Goodchild, L.; Bobo, T.A.; Fu, H. Threshold for Pre-Existing Antibody Levels Limiting Transduction Efficiency of Systemic RAAV9 Gene Delivery: Relevance for Translation. Mol. Ther. Methods Clin. Dev. 2019, 13, 453–462. [CrossRef]
- Weber, T. Anti-AAV Antibodies in AAV Gene Therapy: Current Challenges and Possible Solutions. Front. Immunol. 2021, 12, 1–5. [CrossRef]
- Au, H.K.E.; Isalan, M.; Mielcarek, M. Gene Therapy Advances: A Meta-Analysis of AAV Usage in Clinical Settings. Front. Med. 2022, 8, 1–14. [CrossRef]
- Meliani, A.; Leborgne, C.; Triffault, S.; Jeanson-Leh, L.; Veron, P.; Mingozzi, F. Determination of Anti-Adeno-Associated Virus Vector Neutralizing Antibody Titer with an in Vitro Reporter System. Hum. Gene Ther. Methods 2015, 26, 45–53. [CrossRef]
- Ellis, B.L.; Hirsch, M.L.; Barker, J.C.; Connelly, J.P.; Steininger, R.J.; Porteus, M.H. A Survey of Ex Vivo/in Vitro Transduction Efficiency of Mammalian Primary Cells and Cell Lines with Nine Natural Adeno-Associated Virus (AAV1-9) and One Engineered Adeno-Associated Virus Serotype. Virol. J. 2013, 10, 1–10. [CrossRef]
- Ryals, R.C.; Boye, S.L.; Dinculescu, A.; Hauswirth, W.W.; Boye, S.E. Quantifying Transduction Efficiencies of Unmodified and Tyrosine Capsid Mutant AAV Vectors in Vitro Using Two Ocular Cell Lines. Mol. Vis. 2011, 17, 1090–1102.
- Rapti, K.; Louis-Jeune, V.; Kohlbrenner, E.; Ishikawa, K.; Ladage, D.; Zolotukhin, S.; Hajjar, R.J.; Weber, T. Neutralizing Antibodies against AAV Serotypes 1, 2, 6, and 9 in Sera of Commonly Used Animal Models. Mol. Ther. 2012, 20, 73–83. [CrossRef]
- Cardoso, B.D.; Castanheira, E.M.S.; Lanceros-Méndez, S.; Cardoso, V.F. Recent Advances on Cell Culture Platforms for In Vitro Drug Screening and Cell Therapies: From Conventional to Microfluidic Strategies. Adv. Healthc. Mater. 2023, 12, 1–30. [CrossRef]
- Smith, D.J.; Lapedes, A.S.; De Jong, J.C.; Bestebroer, T.M.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Mapping the Antigenic and Genetic Evolution of Influenza Virus. Science (80-. ). 2004, 305, 371–376. [CrossRef]
- Nonnenmacher, M.; Weber, T. Intracellular Transport of Recombinant Adeno-Associated Virus Vectors. Gene Ther. 2012, 19, 649–658. [CrossRef]
- Berry, G.E.; Asokan, A. Cellular Transduction Mechanisms of Adeno-Associated Viral Vectors. Curr. Opin. Virol. 2016, 21, 54–60. [CrossRef]
- Costa-Verdera, H.; Unzu, C.; Valeri, E.; Adriouch, S.; Aseguinolaza, G.G.; Mingozzi, F.; Kajaste-Rudnitski, A. Understanding and Tackling Immune Responses to Adeno-Associated Viral Vectors. Hum. Gene Ther. 2023, 34, 836–852. [CrossRef]
- Shirley, J.L.; Keeler, G.D.; Sherman, A.; Zolotukhin, I.; Markusic, D.M.; Hoffman, B.E.; Morel, L.M.; Wallet, M.A.; Terhorst, C.; Herzog, R.W. Type I IFN Sensing by CDCs and CD4+ T Cell Help Are Both Requisite for Cross-Priming of AAV Capsid-Specific CD8+ T Cells. Mol. Ther. 2020, 28, 758–770. [CrossRef]
- Li, C.; He, Y.; Nicolson, S.; Hirsch, M.; Weinberg, M.S.; Zhang, P.; Kafri, T.; Samulski, R.J. Adeno-Associated Virus Capsid Antigen Presentation Is Dependent on Endosomal Escape. J. Clin. Invest. 2013, 123, 1390–1401. [CrossRef]
- Kuranda, K.; Jean-Alphonse, P.; Leborgne, C.; Hardet, R.; Collaud, F.; Marmier, S.; Verdera, H.C.; Ronzitti, G.; Veron, P.; Mingozzi, F. Exposure to Wild-Type AAV Drives Distinct Capsid Immunity Profiles in Humans. J. Clin. Invest. 2018, 128, 5267–5279. [CrossRef]
- Braun, M.; Lange, C.; Schatz, P.; Long, B.; Stanta, J.; Gorovits, B.; Tarcsa, E.; Jawa, V.; Yang, T.Y.; Lembke, W.; et al. Preexisting Antibody Assays for Gene Therapy: Considerations on Patient Selection Cutoffs and Companion Diagnostic Requirements. Mol. Ther. Methods Clin. Dev. 2024, 32, 101217. [CrossRef]
- Naso, M.F.; Tomkowicz, B.; Perry, W.L.; Strohl, W.R. Adeno-Associated Virus (AAV) as a Vector for Gene Therapy. BioDrugs 2017, 31, 317–334. [CrossRef]
- Human Gene Therapy for Retinal Disorders. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/human-gene-therapy-retinal-disorders (accessed on 5 December 2024).
- Human Gene Therapy for Neurodegenerative Diseases. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/human-gene-therapy-neurodegenerative-diseases (accessed on 5 December 2024).
- Human Gene Therapy for Hemophilia. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/human-gene-therapy-hemophilia (accessed on 5 December 2024).
- Human Gene Therapy for Rare Diseases. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/human-gene-therapy-rare-diseases (accessed on 5 December 2024).
- Developing and Validating Assays for Anti-Drug Antibody Detection. Available online: https://www.fda.gov/media/119788/download (accessed on 5 December 2024).
- Masat, E., Pavani, G., & Mingozzi, F. Humoral Immunity to AAV Vectors in Gene Therapy: Challenges and Potential Solutions. Discov. Med. 2013, 15, 379–389.
- Tanonaka, K.; Marunouchi, T. Angiotensin-Converting Enzyme 2. Folia Pharmacol. Jpn. 2016, 147, 120–121. [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271-280.e8. [CrossRef]
- Shang, Y.; Chen, F.; Li, S.; Song, L.; Gao, Y.; Yu, X.; Zheng, J. Investigation of Interaction between the Spike Protein of SARS-CoV-2 and ACE2-Expressing Cells Using an In Vitro Cell Capturing System. Biol. Proced. Online 2021, 23, 1–9. [CrossRef]
- Hyseni, I.; Molesti, E.; Benincasa, L.; Piu, P.; Casa, E.; Temperton, N.J.; Manenti, A.; Montomoli, E. Characterisation of SARS-CoV-2 Lentiviral Pseudotypes and Correlation between Pseudotype-Based Neutralisation Assays and Live Virus-Based Micro Neutralisation Assays. Viruses 2020, 12. [CrossRef]
- Meyer, N.L.; Chapman, M.S. Adeno-Associated Virus (AAV) Cell Entry: Structural Insights. Trends Microbiol. 2022, 30, 432–451. [CrossRef]
- Large, E.E.; Chapman, M.S. Adeno-Associated Virus Receptor Complexes and Implications for Adeno-Associated Virus Immune Neutralization. Front. Microbiol. 2023, 14. [CrossRef]
- Grimm, D.; Lee, J.S.; Wang, L.; Desai, T.; Akache, B.; Storm, T.A.; Kay, M.A. In Vitro and In Vivo Gene Therapy Vector Evolution via Multispecies Interbreeding and Retargeting of Adeno-Associated Viruses. J. Virol. 2008, 82, 5887–5911. [CrossRef]
- Zheng, Z.; Ye, J.; Leng, M.; Gan, C.; Tang, N.; Li, W.; Valencia, C.A.; Dong, B.; Chow, H.Y. Enhanced Sensitivity of Neutralizing Antibody Detection for Different AAV Serotypes Using HeLa Cells with Overexpressed AAVR. Front. Pharmacol. 2023, 14, 1–11. [CrossRef]
- Calcedo, R.; Chichester, J.A.; Wilson, J.M. Assessment of Humoral, Innate, and T-Cell Immune Responses to Adeno-Associated Virus Vectors. Hum. Gene Ther. Methods 2018, 29, 86–95. [CrossRef]
- Qian, R.; Xiao, B.; Li, J.; Xiao, X. Directed Evolution of AAV Serotype 5 for Increased Hepatocyte Transduction and Retained Low Humoral Seroreactivity. Mol. Ther. Methods Clin. Dev. 2021, 20, 122–132. [CrossRef]
- Perocheau, D.P.; Cunningham, S.; Lee, J.; Antinao Diaz, J.; Waddington, S.N.; Gilmour, K.; Eaglestone, S.; Lisowski, L.; Thrasher, A.J.; Alexander, I.E.; et al. Age-Related Seroprevalence of Antibodies Against AAV-LK03 in a UK Population Cohort. Hum. Gene Ther. 2019, 30, 79–87. [CrossRef]
- Li, C.; Samulski, R.J. Engineering Adeno-Associated Virus Vectors for Gene Therapy. Nat. Rev. Genet. 2020, 21, 255–272. [CrossRef]
- Korneyenkov, M.A.; Zamyatnin, A.A. Next Step in Gene Delivery: Modern Approaches and Further Perspectives of Aav Tropism Modification. Pharmaceutics 2021, 13, 1–16. [CrossRef]
- Carneiro, A.; Schaffer, D. Engineering Novel Adeno-Associated Viruses (AAVs) for Improved Delivery in the Nervous System. Curr. Opin. Chem. Biol. 2024, 83. [CrossRef]
- Gardner, M.R.; Mendes, D.E.; Muniz, C.P.; Martinez-Navio, J.M.; Fuchs, S.P.; Gao, G.; Desrosiers, R.C. High Concordance of ELISA and Neutralization Assays Allows for the Detection of Antibodies to Individual AAV Serotypes. Mol. Ther. Methods Clin. Dev. 2022, 24, 199–206. [CrossRef]
- Halbert, C.L.; Miller, A.D.; McNamara, S.; Emerson, J.; Gibson, R.L.; Ramsey, B.; Aitken, M.L. Prevalence of Neutralizing Antibodies against Adeno-Associated Virus (AAV) Types 2, 5, and 6 in Cystic Fibrosis and Normal Populations: Implications for Gene Therapy Using AAV Vectors. Hum. Gene Ther. 2006, 17, 440–447. [CrossRef]
- Colella, P.; Ronzitti, G.; Mingozzi, F. Emerging Issues in AAV-Mediated In Vivo Gene Therapy. Mol. Ther. Methods Clin. Dev. 2018, 8, 87–104. [CrossRef]
- Mingozzi, F.; Hasbrouck, N.C.; Basner-Tschakarjan, E.; Edmonson, S.A.; Hui, D.J.; Sabatino, D.E.; Zhou, S.; Wright, J.F.; Jiang, H.; Pierce, G.F.; et al. Modulation of Tolerance to the Transgene Product in a Nonhuman Primate Model of AAV-Mediated Gene Transfer to Liver. Blood 2007, 110, 2334–2341. [CrossRef]
- Mestas, J.; Hughes, C.C.W. Of Mice and Not Men: Differences between Mouse and Human Immunology. J. Immunol. 2004, 172, 2731–2738. [CrossRef]
- Mingozzi, F.; High, K.A. Immune Responses to AAV Vectors: Overcoming Barriers to Successful Gene Therapy. Blood 2013, 122, 23–36. [CrossRef]
- Bjornson-Hooper, Z.B.; Fragiadakis, G.K.; Spitzer, M.H.; Chen, H.; Madhireddy, D.; Hu, K.; Lundsten, K.; McIlwain, D.R.; Nolan, G.P. A Comprehensive Atlas of Immunological Differences Between Humans, Mice, and Non-Human Primates. Front. Immunol. 2022, 13, 1–17. [CrossRef]
- Crowley, A.R.; Ackerman, M.E. Mind the Gap: How Interspecies Variability in IgG and Its Receptors May Complicate Comparisons of Human and Non-Human Primate Effector Function. Front. Immunol. 2019, 10, 1–19. [CrossRef]
- Sibley, L.; Daykin-Pont, O.; Sarfas, C.; Pascoe, J.; White, A.D.; Sharpe, S. Differences in Host Immune Populations between Rhesus Macaques and Cynomolgus Macaque Subspecies in Relation to Susceptibility to Mycobacterium Tuberculosis Infection. Sci. Rep. 2021, 11, 1–9. [CrossRef]
- Goertsen, D.; Flytzanis, N.C.; Goeden, N.; Chuapoco, M.R.; Cummins, A.; Chen, Y.; Fan, Y.; Zhang, Q.; Sharma, J.; Duan, Y.; et al. AAV Capsid Variants with Brain-Wide Transgene Expression and Decreased Liver Targeting after Intravenous Delivery in Mouse and Marmoset. Nat. Neurosci. 2022, 25, 106–115. [CrossRef]
- Albrecht, L.; Bishop, E.; Jay, B.; Lafoux, B.; Minoves, M.; Passaes, C. Covid-19 Research: Lessons from Non-Human Primate Models. Vaccines 2021, 9, 1–26. [CrossRef]
- Wong, M.X.Y.; Chen, Q. Editorial: Humanized Mouse Models to Study Immune Responses to Human Infectious Organisms. Front. Immunol. 2022, 13, 1–2. [CrossRef]
- Chupp, D.P.; Rivera, C.E.; Zhou, Y.; Xu, Y.; Ramsey, P.S.; Xu, Z.; Zan, H.; Casali, P. A Humanized Mouse That Mounts Mature Class-Switched, Hypermutated and Neutralizing Antibody Responses. Nat. Immunol. 2024, 25, 1489–1506. [CrossRef]
- Verkoczy, L.; Alt, F.W.; Tian, M. Human Ig Knockin Mice to Study the Development and Regulation of HIV-1 Broadly Neutralizing Antibodies. Immunol. Rev. 2017, 275, 89–107. [CrossRef]
- Lee, E.C.; Liang, Q.; Ali, H.; Bayliss, L.; Beasley, A.; Bloomfield-Gerdes, T.; Bonoli, L.; Brown, R.; Campbell, J.; Carpenter, A.; et al. Complete Humanization of the Mouse Immunoglobulin Loci Enables Efficient Therapeutic Antibody Discovery. Nat. Biotechnol. 2014, 32, 356–363. [CrossRef]
- Mian, S.A.; Anjos-Afonso, F.; Bonnet, D. Advances in Human Immune System Mouse Models for Studying Human Hematopoiesis and Cancer Immunotherapy. Front. Immunol. 2021, 11, 1–16. [CrossRef]
- Sun, L.; Tu, L.; Gao, G.; Sun, X.; Duan, J.; Lu, Y. Assessment of a Passive Immunity Mouse Model to Quantitatively Analyze the Impact of Neutralizing Antibodies on Adeno-Associated Virus-Mediated Gene Transfer. J. Immunol. Methods 2013, 387, 114–120. [CrossRef]
- Wang, L.; Calcedo, R.; Wang, H.; Bell, P.; Grant, R.; Vandenberghe, L.H.; Sanmiguel, J.; Morizono, H.; Batshaw, M.L.; Wilson, J.M. The Pleiotropic Effects of Natural AAV Infections on Liver-Directed Gene Transfer in Macaques. Mol. Ther. 2010, 18, 126–134. [CrossRef]
- Cabanes-Creus, M.; Liao, S.H.Y.; Gale Navarro, R.; Knight, M.; Nazareth, D.; Lau, N.S.; Ly, M.; Zhu, E.; Roca-Pinilla, R.; Bugallo Delgado, R.; et al. Harnessing Whole Human Liver Ex Situ Normothermic Perfusion for Preclinical AAV Vector Evaluation. Nat. Commun. 2024, 15. [CrossRef]
- Chiang, Y.P.; Bishawi, M.; Wang, C.; Watson, M.J.; Lee, F.H.; Roki, A.; Doty, A.; Lewis, A.; Kemplay, C.; Tatum, J.; et al. Successful Gene Delivery to Cardiac Allograft with Adeno-Associated Viral Vector Using Ex Vivo Storage Perfusion Platform. J. Hear. Lung Transplant. 2020, 39, S360–S361. [CrossRef]
- Vervoorn, M.T.; Amelink, J.J.G.J.; Ballan, E.M.; Doevendans, P.A.; Sluijter, J.P.G.; Mishra, M.; Boink, G.J.J.; Bowles, D.E.; van der Kaaij, N.P. Gene Therapy during Ex Situ Heart Perfusion: A New Frontier in Cardiac Regenerative Medicine? Front. Cardiovasc. Med. 2023, 10, 1–14. [CrossRef]
- Lau, N.S.; Ly, M.; Dennis, C.; Jacques, A.; Cabanes-Creus, M.; Toomath, S.; Huang, J.; Mestrovic, N.; Yousif, P.; Chanda, S.; et al. Long-Term Ex Situ Normothermic Perfusion of Human Split Livers for More than 1 Week. Nat. Commun. 2023, 14. [CrossRef]
- Ghafoori, S.M.; Petersen, G.F.; Conrady, D.G.; Calhoun, B.M.; Stigliano, M.Z.Z.; Baydo, R.O.; Grice, R.; Abendroth, J.; Lorimer, D.D.; Edwards, T.E.; et al. Structural Characterisation of Hemagglutinin from Seven Influenza A H1N1 Strains Reveal Diversity in the C05 Antibody Recognition Site. Sci. Rep. 2023, 13, 1–9. [CrossRef]
- Solanki, K.; Rajpoot, S.; Kumar, A.; Zhang, K.Y.J.; Ohishi, T.; Hirani, N.; Wadhonkar, K.; Patidar, P.; Pan, Q.; Baig, M.S. Structural Analysis of Spike Proteins from SARS-CoV-2 Variants of Concern Highlighting Their Functional Alterations. Future Virol. 2022, 17, 723–732. [CrossRef]
- Qiu, T.; Yang, Y.; Qiu, J.; Huang, Y.; Xu, T.; Xiao, H.; Wu, D.; Zhang, Q.; Zhou, C.; Zhang, X.; et al. CE-BLAST Makes It Possible to Compute Antigenic Similarity for Newly Emerging Pathogens. Nat. Commun. 2018, 9. [CrossRef]
- Bergamaschi, G.; Fassi, E.M.A.; Romanato, A.; D’Annessa, I.; Odinolfi, M.T.; Brambilla, D.; Damin, F.; Chiari, M.; Gori, A.; Colombo, G.; et al. Computational Analysis of Dengue Virus Envelope Protein (E) Reveals an Epitope with Flavivirus Immunodiagnostic Potential in Peptide Microarrays. Int. J. Mol. Sci. 2019, 20, 1–13. [CrossRef]
- Shah, S.A.W.; Palomar, D.P.; Barr, I.; Poon, L.L.M.; Quadeer, A.A.; McKay, M.R. Seasonal Antigenic Prediction of Influenza A H3N2 Using Machine Learning. Nat. Commun. 2024, 15, 1–12. [CrossRef]
- Wang, W.; Lusvarghi, S.; Subramanian, R.; Epsi, N.J.; Wang, R.; Goguet, E.; Fries, A.C.; Echegaray, F.; Vassell, R.; Coggins, S.A.; et al. Antigenic Cartography of Well-Characterized Human Sera Shows SARS-CoV-2 Neutralization Differences Based on Infection and Vaccination History. Cell Host Microbe 2022, 30, 1745-1758.e7. [CrossRef]
- Ito, M.; Takino, N.; Nomura, T.; Kan, A.; Muramatsu, S. ichi Engineered Adeno-Associated Virus 3 Vector with Reduced Reactivity to Serum Antibodies. Sci. Rep. 2021, 11, 1–10. [CrossRef]
- Havlik, L.P.; Simon, K.E.; Smith, J.K.; Klinc, K.A.; Tse, L. V.; Oh, D.K.; Fanous, M.M.; Meganck, R.M.; Mietzsch, M.; Kleinschmidt, J.; et al. Coevolution of Adeno-Associated Virus Capsid Antigenicity and Tropism through a Structure-Guided Approach. J. Virol. 2020, 94. [CrossRef]
- Maheshri, N.; Koerber, J.T.; Kaspar, B.K.; Schaffer, D. V. Directed Evolution of Adeno-Associated Virus Yields Enhanced Gene Delivery Vectors. Nat. Biotechnol. 2006, 24, 198–204. [CrossRef]
- Monteilhet, V.; Veron, P.; Leborgne, C.; Benveniste, O. Prevalence of Serum IgG and Neutralizing Factors and 9 in the Healthy Population : Implications for Gene Therapy Using AAV Vectors. Hum. Gene Ther. 2010, 712, 704–712.
- Andrews, S.F.; Kaur, K.; Pauli, N.T.; Huang, M.; Huang, Y.; Wilson, P.C. High Preexisting Serological Antibody Levels Correlate with Diversification of the Influenza Vaccine Response. J. Virol. 2015, 89, 3308–3317. [CrossRef]
- Chen, Y.Q.; Wohlbold, T.J.; Zheng, N.Y.; Huang, M.; Huang, Y.; Neu, K.E.; Lee, J.; Wan, H.; Rojas, K.T.; Kirkpatrick, E.; et al. Influenza Infection in Humans Induces Broadly Cross-Reactive and Protective Neuraminidase-Reactive Antibodies. Cell 2018, 173, 417-429.e10. [CrossRef]
- Garcia-Beltran, W.F.; Lam, E.C.; St. Denis, K.; Nitido, A.D.; Garcia, Z.H.; Hauser, B.M.; Feldman, J.; Pavlovic, M.N.; Gregory, D.J.; Poznansky, M.C.; et al. Multiple SARS-CoV-2 Variants Escape Neutralization by Vaccine-Induced Humoral Immunity. Cell 2021, 184, 2372-2383.e9. [CrossRef]
- Miyamoto, S.; Arashiro, T.; Adachi, Y.; Moriyama, S.; Kinoshita, H.; Kanno, T.; Saito, S.; Katano, H.; Iida, S.; Ainai, A.; et al. Vaccination-Infection Interval Determines Cross-Neutralization Potency to SARS-CoV-2 Omicron after Breakthrough Infection by Other Variants. Med 2022, 3, 249-261.e4. [CrossRef]
- Zhang, G.F.; Meng, W.; Chen, L.; Ding, L.; Feng, J.; Perez, J.; Ali, A.; Sun, S.; Liu, Z.; Huang, Y.; et al. Neutralizing Antibodies to SARS-CoV-2 Variants of Concern Including Delta and Omicron in Subjects Receiving MRNA-1273, BNT162b2, and Ad26.COV2.S Vaccines. J. Med. Virol. 2022, 94, 5678–5690. [CrossRef]
- Chi, X.; Guo, Y.; Zhang, G.; Sun, H.; Zhang, J.; Li, M.; Chen, Z.; Han, J.; Zhang, Y.; Zhang, X.; et al. Broadly Neutralizing Antibodies against Omicron-Included SARS-CoV-2 Variants Induced by Vaccination. Signal Transduct. Target. Ther. 2022, 7, 1–11. [CrossRef]
- Majowicz, A.; Ncete, N.; van Waes, F.; Timmer, N.; van Deventer, S.J.; Mahlangu, J.N.; Ferreira, V. Seroprevalence of Pre-Existing Nabs Against AAV1, 2, 5, 6 and 8 in South African Hemophilia B Patient Population. Blood 2019, 134, 3353–3353. [CrossRef]
- Abdul, T.Y.; Hawse, G.P.; Smith, J.; Sellon, J.L.; Abdel, M.P.; Wells, J.W.; Coenen, M.J.; Evans, C.H.; De La Vega, R.E. Prevalence of AAV2.5 Neutralizing Antibodies in Synovial Fluid and Serum of Patients with Osteoarthritis. Gene Ther. 2023, 30, 587–591. [CrossRef]
- Racmacs. Available online: https://acorg.github.io/Racmacs/ (accessed on 5 December 2024).
- Gao, G.; Vandenberghe, L.H.; Alvira, M.R.; Lu, Y.; Calcedo, R.; Zhou, X.; Wilson, J.M. Clades of Adeno-Associated Viruses Are Widely Disseminated in Human Tissues. J. Virol. 2004, 78, 6381–6388. [CrossRef]
Figure 1.
The present test systems for detecting anti-AAV Nabs.
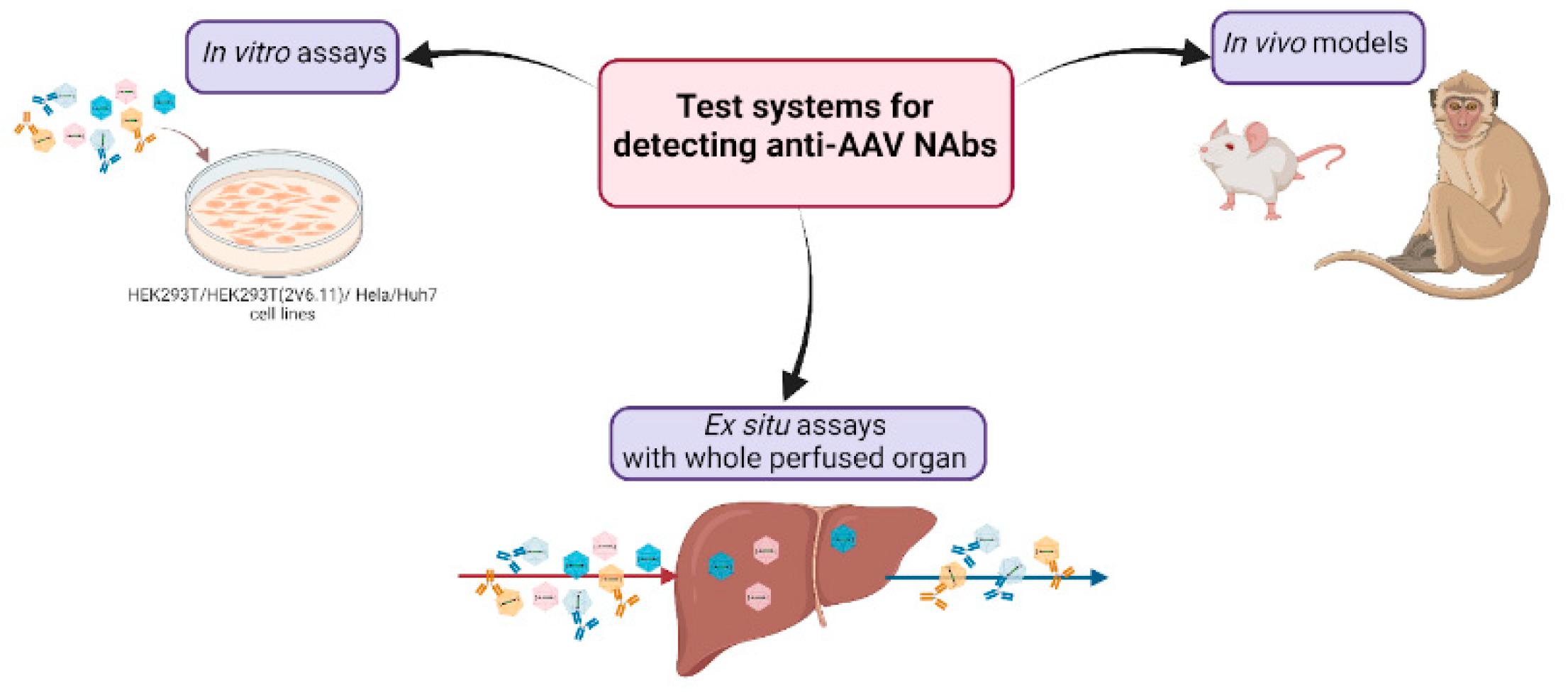

Table 1.
Selected examples of over 30 clinical studies employing AAV.
| Disease | Status | Phase | Exclusion Criteria and Method of Detection Anti-AAV Abs | Administration | Sponsor | ClinicalTrials.gov ID |
|---|---|---|---|---|---|---|
| AAV9 | ||||||
| Muscular Atrophy Type 1 | COMPLETED | Phase 1 | binding antibody titers >1:50, ELISA | Intravenous | Novartis (Novartis Gene Therapies) | NCT02122952 |
| MPS IIIA | ACTIVE, NOT RECRUITING | Phase 2 Phase 3 |
binding antibody titers ≥ 1:100, ELISA | Intravenous | Ultragenyx Pharmaceutical Inc | NCT02716246 |
| Mucopolysaccharidosis (MPS) IIIB (MPSIIIB) | TERMINATED | Phase 1 Phase 2 |
binding antibody titers ≥ 1:100, ELISA | Intravenous | Abeona Therapeutics, Inc | NCT03315182 |
| Late Infantile Neuronal Ceroid Lipofuscinosis 6 (vLINCL6) | COMPLETED | Phase 1 Phase 2 |
binding antibody titers > 1:50, ELISA | Intrathecally into the lumbar spinal cord region | Amicus Therapeutics | NCT02725580 |
| AAVrh10 | ||||||
| Late Infantile Krabbe Disease Treated Previously With HSCT (REKLAIM) | RECRUITING | Phase 1 Phase 2 |
binding antibody titers >1:100, ELISA *This criteria will not apply to children screened before they have received HSCT or for children who sign the inform consent within 60 days from HSCT. |
Intravenous | Forge Biologics, Inc | NCT05739643 |
| Alpha-1 Antitrypsin (A1AT) | COMPLETED | Phase 1 Phase 2 |
neutralizing antibody titer ≥ 1:5, neutralizing Ab | Intravenous or intrapleural | Adverum Biotechnologies, Inc. | NCT02168686 |
| Hemophilia B | TERMINATED | Phase 1 Phase 2 |
neutralizing antibody titer > 1:5, neutralizing Ab | Intravenous | Ultragenyx Pharmaceutical Inc | NCT02618915 |
| AAV2 | ||||||
| Advanced Parkinson s Disease | COMPLETED | Phase 1 | total antibody titer >1000, ELISA | Intracerebral (Bilateral Stereotactic Convection-Enhanced Delivery) | National Institute of Neurological Disorders and Stroke (NINDS) | NCT01621581 |
| Aromatic L-amino Acid Decarboxylase (AADC) Deficiency | COMPLETED | Phase 2 | neutralizing antibody titer over 1,200 folds or an ELISA OD over 1 cannot be recruited into this trial. | Intracerebral | National Taiwan University Hospital | NCT02926066 |
| Leber congenital amaurosis (LCA) | ACTIVE, NOT RECRUITING | Phase 1 | AAV antibody titers greater than two standard deviations above normal at baseline; | Subretinal | University of Pennsylvania | NCT00481546 |
| Hemophilia B | TERMINATED | Phase 1 | Presence of neutralizing antibodies AAV2/6 vector | Intravenous | Sangamo Therapeutics | NCT02695160 |
| AAV8 | ||||||
| Hemophilia B | TERMINATED | Phase 1 | neutralizing antibody titer > 1:5 | Intravenous | Spark Therapeutics, Inc. | NCT01620801 |
| Late Onset Pompe Disease (FORTIS) | RECRUITING | Phase 1 Phase 2 |
neutralizing antibody titer > 1:20 | Intravenous | Astellas Gene Therapies | NCT04174105 |
| Homozygous Familial Hypercholesterolemia (HoFH) | TERMINATED | Phase 1 Phase 2 |
neutralizing antibody titer > 1:10 | Intravenous | REGENXBIO Inc | NCT02651675 |
| Hemophilia B | TERMINATED | Phase 1 Phase 2 |
neutralizing antibody titers ≥ 1:5 | Intravenous | Baxalta now part of Shire | NCT04394286 |
| Hemophilia B | ACTIVE, NOT RECRUITING | Phase 1 | Detectable antibodies reactive with AAV8 | Intravenous | St. Jude Children’s Research Hospital | NCT00979238 |
| cocaine use disorder | RECRUITING | Phase 1 | Patientss how detectable pre-existing immunity to the AAV8 capsid as measured by AAV8 transduction inhibition and AAV8 total antibodies. | Intravenous | W. Michael Hooten | NCT04884594 |
| The Human Immunodeficiency Virus (HIV) | ACTIVE, NOT RECRUITING | Phase 1 | titer of pre-existing antibodies to capsid > 1:90 | Intramuscular | National Institute of Allergy and Infectious Diseases (NIAID) | NCT03374202 |
| Hemophilia B | TERMINATED | Phase 1 | neutralizing antibody titer > 1:5 | Intravenous | Spark Therapeutics, Inc. | NCT01620801 |
| AAVrh74 | ||||||
| Limb-Girdle Muscular Dystrophy, Type 2D (LGMD2D) | COMPLETED | Phase 1 Phase 2 |
binding antibody titers ≥ 1:50, ELISA | Isolated limb infusion (ILI) | Sarepta Therapeutics, Inc. | NCT01976091 |
| Dysferlinopathies | COMPLETED | Phase 1 | binding antibody titers > 1:50, ELISA | Extensor digitorum brevis muscle (EDB) | Sarepta Therapeutics, Inc. | NCT02710500 |
| Duchenne Muscular Dystrophy | COMPLETED | Phase 1 | binding antibody titers ≥ 1:50, ELISA | Extensor Digitorum Brevis (EDB) muscle | Jerry R. Mendell | NCT02376816 |
| AAV5 | ||||||
| Hemophilia A | ACTIVE, NOT RECRUITING | Phase 1 Phase 2 |
Absence of pre-existing antibodies against the AAV5 vector capsid, measured by total AAV5 antibody ELISA | Intravenous | BioMarin Pharmaceutical | NCT03520712 |
| Hemophilia B | COMPLETED | Phase 1 Phase 2 |
Neutralizing antibodies against AAV5 at Visit 1 | Intravenous | CSL Behring | NCT02396342 |
| Arthritis | UNKNOWN STATUS | Phase 1 | Presence of neutralising antibody (Nab) titers against adeno-associated virus type 5 (AAV5) and/or hIFN-β. | Intra-articular | Arthrogen | NCT02727764 |
| Haemophilia A | ACTIVE, NOT RECRUITING | Phase 1 Phase 2 |
Detectable pre-existing immunity to the AAV5 capsid as measured by AAV5 transduction inhibition or AAV5 total antibodies | Intravenous | BioMarin Pharmaceutical | NCT02576795 |
| AAV1 | ||||||
| CMT1A | SUSPENDED Vector has not been produced |
Phase 1 Phase 2 |
binding antibody titers ≥ 1:50, ELISA | Intramuscular | Nationwide Children’s Hospital | NCT03520751 |
| Duchenne Muscular Dystrophy | COMPLETED | Phase 1 Phase 2 |
binding antibody titers > 1:50, ELISA | Intramuscular | Jerry R. Mendell | NCT02354781 |
| Heart Failure | TERMINATED | Phase 2 | neutralizing antibody titers ≥ 1:2 | Intracoronary | Assistance Publique - Hôpitaux de Paris | NCT01966887 |
| Advanced Heart Failure | COMPLETED | Phase 2 | neutralizing antibody titers ≥ 1:2 | Intracoronary | Celladon Corporation | NCT01643330 |
| Limb Girdle Muscular Dystrophy Type 2C | COMPLETED | Phase 1 | Pre-injection neutralizing anti-AAV1 antibodies titer superior or equal to 1/800. | Intramuscular | Genethon | NCT01344798 |
| Becker Muscular Dystrophy and Sporadic Inclusion Body | COMPLETED | Phase 1 | neutralizing antibody titers ≥ 1:1600, ELISA | Intramuscular | Nationwide Children’s Hospital | NCT01519349 |
| Heart Failure | RECRUITING | Phase 1 | Anti-AAV1 neutralizing antibodies | Antegrade epicardial coronary artery infusion | Sardocor Corp. | NCT06061549 |
| Others | ||||||
| Hemophilia A | ACTIVE, NOT RECRUITING | Phase 3 | Anti-AAV6 neutralizing antibodies | Intravenous | Pfizer | NCT04370054 |
| Hemophilia B | COMPLETED | Phase 2 | Neutralizing antibodies reactive with AAV-Spark100 above and/or below a defined titre | Intravenous | Pfizer | NCT02484092 |
| Hemophilia A | ACTIVE, NOT RECRUITING | Phase 1 Phase 2 |
Detectable antibodies reactive with AAVhu37capsid. | Intravenous | Bayer | NCT03588299 |
| Hemophilia A | COMPLETED | Phase 1 Phase 2 |
Detectable antibodies reactive with AAV-Spark200 capsid | Intravenous | Spark Therapeutics, Inc. | NCT03003533 |
| Hemophilia A | COMPLETED | Phase 1 Phase 2 |
Detectable antibodies reactive with AAV-Spark capsid | Intravenous | Spark Therapeutics, Inc. | NCT03734588 |
| Fabry Disease | ACTIVE, NOT RECRUITING | Phase 1 Phase 2 |
Presence of high titer neutralizing antibody to 4D-310 capsid, or presence of high antibody titer to AGA | Intravenous | 4D Molecular Therapeutics | NCT04519749 |
Table 2.
Summary of data used to construct antigenic map.
| Group | Cohort Description |
In Vitro Neutralization Test Features | Reference | AAV Serotypes Used | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AAV1 | AAV2 | AAV2.5 | AAV3B | AAV5 | AAV6 | AAV8 | AAV9 | AAVrh10 | ||||
| 1 | Osteoarthritis patient population | 7.5 × 10^6 transducing units of AAV-GFP + 56 µL diluted synovial fluid dilution, 5 × 10^4 HIG-82 cells/well, GFP expression analyzed by FACS. The Nab titer is given as the dilution of synovial fluid required to obtain 50% inhibition of transduction by AAV.GFP, as compared to cells incubated with AAV.GFP alone. | Abdul T. Y. et al. 2023 | ☑ | ☑ | ☑ | ||||||
| 2 | Normal human donors | 10^9 AAV-LacZ vg/well + 2-fold serial diluted serum samples (initial dilution, 1:20), 10^5 Huh7 cells/well, luciferase activity was measured by microplate luminometer. The NAb titer was reported as the highest serum dilution that inhibited AAV transduction by 50%, compared with the mouse serum control. | Giles A. R. et 2020 | ☑ | ☑ | ☑ | ☑ | ☑ | ||||
| 3 | Healthy and hemophilia B patient populations | 70 ul of virus + serial diluted serum samples (initial dilution, 1:20), HEK293T cells, luciferase activity was measured by microplate luminometer. NAB titer (IC50) is the dilution at which antibodies inhibit Hek293T cell transduction with AAV-LUC by 50%. | Majowicz A. et al. 2019 | ☑ | ☑ | ☑ | ☑ | ☑ | ||||
| 4 | Healthy and cystic fibrosis patient populations | 10^8 AAV-AP vg/well + 100 ul diluted serum samples (initial dilution, 1:20), HTX cells, AP-positive focusforming units were measured. The highest dilution of serum that inhibited AAV transduction by 50% or more compared with the untreated vector was defined as the neutralizing titer. | Halbert C. L. et al 2006 | ☑ | ☑ | ☑ | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



