
Submitted:
25 November 2024
Posted:
26 November 2024
You are already at the latest version
Abstract
V-ATPases are ubiquitous and evolutionarily conserved rotatory proton pumps, which are crucial for maintaining various biological functions. Previous investigations have shown that a V-ATPase is present in the support cells of moth trichoid sensilla and influences their olfactory sensory neuron performance. Generally, V-ATPases are thought to regulate the pH value within the sensillum lymph, and aid K+ homeostasis within the sensillum. This, in turn, could influence various mechanisms involved within the support cells like maintaining the receptor membrane potential (receptor current), nutrient and ion transport, odorant solubility and various signaling mechanisms. In this study, we identify V-ATPase expression and localization in the Drosophila melanogaster antenna using bioinformatics and immunohistochemistry. Elucidating an olfactory V-ATPase function will improve our current understanding of how support cells contribute to Drosophila's sense of smell.
Keywords:
Proton transmembrane transport
; olfaction
; antenna
; OSN
; support cell
; sensillum lymph
; H+ homeostasis
1. Introduction
Homeostasis, or the maintenance of stable internal conditions independent of changing external factors, is a fundamental requirement for living organisms. Insects, for example, need to regulate their sensory sensitivity, respiration, body temperature and movement to thrive in a constantly changing environment [1]. An important organelle that contributes to homeostasis at cellular level is the mitochondrion, which generates adenosine triphosphate (ATP) through oxidative phosphorylation to provide energy for various cellular processes [2]. A molecule essential for cellular homeostasis is the H+ vacuolar-type ATPase (V-ATPase), an evolutionarily conserved protein that functions as a primary proton pump derived from a common ancestral enzyme [3,4]. V-ATPases create an electrochemical proton gradient across cell membranes, acidifying intracellular vesicles and organelles [5,6,7,8,9]. They also balance the cellular pH and contribute to the transport of ions and nutrients across membranes, thereby also regulating protein sorting and degradation [10,11,12,13,14]. Impairments in V-ATPase function disturbs the degradation process within the lysosome, which may lead to neurodegenerative disorders such as Alzheimer's disease [11,15,16]. Furthermore, disruption of V-ATPase-mediated acidification has shown to cause aberrant signaling during D. melanogaster and Caenorhabditis elegans embryogenesis [17,18]. Notably, these proton pumps are found in the apical region of various eukaryotic cell types such as in intercalated kidney cells, epithelial cells in the nasal mucosa, support cells in the inner ear, as well as in many other cell types including insect epithelia [3,19,20]. They are involved in signaling pathways such as mTOR, Notch and Wnt [21] and also play a critical role in neurotransmitter uptake and release [8].
In the 1950s, 1980s and 1990s researchers identified a so called ‘K+ pump’ in various insect tissues which is now known as V-ATPase [22,23,24,25,26]. V-ATPases are expressed in insect organs like salivary glands of the blowfly Calliphora, Malpighian tubules of the blood-sucking bug Rhodnius, the midgut of moths, and various insect sensory organs [19,20]. V-ATPases also occur in endomembrane and apical plasma membranes of insect epithelia [20]. For example, V-ATPase monoclonal antibodies (antibody 86-3 and antibody 47-5) have been employed to detect V-ATPase protein in an extremely folded apical plasma membrane region of support cells of moth sensilla [27,28].
Earlier, it was shown that these electrogenic potassium pumps (H+ V-ATPase) were responsible for driving receptor current upon ligand-receptor interaction and acts as an energy source for the transepithelial voltage [29]. These pumps create a proton gradient across the membrane by pumping H+ ions into cells. Another protein, the K+/H+ antiporter uses this H+ gradient to transport K+ into the cell [19,20,26,30]. The K+ transport mechanism has been observed in many insect tissues including Malpighian tubules of yellow fever mosquito Aedes aegypti and also in the support cells of mechanosensory sensilla of the cockroach Periplaneta americana [31,32]. Several genes coding for V-ATPase subunits have since been identified in the Drosophila genome with high expression levels in Malpighian tubules, rectum, antennal palps, and uterus [17,19,33].
The present study asks whether a V-ATPase might play a role in Drosophila olfaction. D. melanogaster possesses two olfactory appendages on their heads: the antennae and the maxillary palps [34,35,36,37,38]. The third antennal segment is covered with chemosensory hair-like structures termed sensilla [34,36,37]. These sensilla house the olfactory sensory neurons (OSNs) that are enveloped by three different types of supporting cells, the thecogen cells (first layer), the trichogen cells (second layer), and the tormogen cells (outermost layer) [34,39]. OSN dendrites are equipped with olfactory receptors such as odorant receptors (ORs) which are composed of an odor-specific protein and a coreceptor protein (Orco) thereby forming an odor-gated ion channel [40,41]. OSN dendrites are surrounded by a K+ rich sensillum lymph [29]. A previous study has indicated an active role of support cells in D. melanogaster odor processing [39]. Specifically, thecogen cells were indicated to take up K+ previously released by OSNs according to enhanced activity after OR stimulation. As V-ATPases and K+/H+ antiporters are plausible to form a K+ uptake system together, we asked whether V-ATPases might occur in D. melanogaster olfactory cells in the antenna. Supporting this idea, a previous study documented abundant V-ATPase expression in tormogen cells of trichoid sensilla of male moths [27,42]. This study aims to update our understanding of antennal V-ATPases using molecular methodologies to elucidate where V-ATPase is expressed in the antenna of the D. melanogaster model organism using bioinformatics and immunohistochemistry.
2. Materials and Methods
2.1. Bioinformatic Analysis
36 genes composing Drosophila V-ATPase subunits (FlyBase gene group FBgg0000111) were shortlisted for gene expression comparison with other members annotated with the Gene Ontology term “proton transmembrane transport” (GO:1902600), as a functionally related control set. Eight antennal reference genes (Orco, Ir8a, Ir25a, elav, repo, nompA, Su(H), sv) were selected based on involvement as common antennal cell type markers (as three coreceptors demarcating various OSN classes, and as neuronal, glial, thecogen-, tormogen-specific and support cell-demarcating markers, respectively). Ubiquitous, pancellular housekeeping genes were selected further as transcriptional references for involvement in common cytoskeletal, metabolic, physiological, and enzymatic functions (Act5C, Gapdh1, Cam and eEF1β, respectively). For bioinformatic analysis, we consulted two separate bulk antennal tissue RNA-seq transcriptomic datasets. One compares adult male and female D. melanogaster, sampling 1,200 antennae pairs for each sex in Canton-S flies aged > 1 day old post-eclosion [43]. The second RNA-seq dataset pools 300 mixed sex D. melanogaster of 5-12 day old age, for wildtype Canton-S flies and homozygous atonal (ato) mutants [44].
Fragments/reads per kilobase per million mapped fragments/reads (FPKM or RPKM) values for all genes detectable in the antenna of each fly cohort were ordered, plotted, labeled and color-coded using R. For all studies, all (outdated, synonymous or alternative) gene names/symbols were converted to the FlyBase standard gene symbol for parity, and checked manually and automatically with a custom script. All gene nomenclature follows FlyBase gene symbol naming convention. Gene transcripts are italicized and references to protein counterparts are unitalicized in line with FlyBase nomenclature guidelines.
For single-cell data, we consulted the antennal “10x stringent” dataset of the Fly Cell Atlas derived using a microfluidic droplet-based cell capture methodology [45]. Differential expression analysis was performed comparing cell group classifications on specific gene expression using non-parametric Wilcoxon rank-sum statistical testing (via Seurat) using the Automated Single-cell Analysis Pipeline (ASAP) portal asap.epfl.ch [46] , with the following parametrization: minimum % of cells with gene > 0 = 0.1 (10%); min%diff = NULL; max cells per group = NULL; foldchange cutoff was not used to prevent any data subsetting. Explicit parametrization settings are available in the data repository associated with this study. A false detection rate (FDR) adjusted p-value of < 0.05 was used to differentiate significantly up- or downregulated genes for each cell type class, which is defined based on the grouping “annotation_broad”.
2.2. Immunohistochemistry
For immunohistochemistry, we used 6-8 day old transgenic D. melanogaster flies: nompA-GAL4 and ASE5-GAL4 line (kindly provided by Craig Montell (University of California, Santa Barbara)), N-GFP-Orco flies [47] and UAS-DenMark flies (made in this lab) to perform immunostaining of antennae and confocal imaging thereof. Dissected antennae were mounted in an optimal cutting temperature (OCT) mounting media (Lot: 03816567; VWR Chemicals, Leuven, Belgium). Microm HM 560 was used to cut thin antennal sections on FisherSuperFrost Plus slides. Sections were then immediately fixed using 4% paraformaldehyde for 10 mins in a humidified chamber. Afterwards, slides were washed twice using phosphate-buffered saline (PBS) for 10 mins on a shaker at room temperature. Slides were then kept in blocking solution (5% normal goat serum in PBS) for 30 mins and covered with a coverslip (#1). Blocking solution was then removed and primary monoclonal antibody was added prior to covering slides with a coverslip (#1), and left for overnight incubation at 4°C. Next day, slides were washed four times with PBS, 10 mins each, and then blocked with blocking solution for 30 minutes. Following, secondary fluorescent antibodies were added to each slide and incubated for two hours at room temperature in the dark. Later, slides were washed four times, five mins per wash, in PBS on a shaker in the dark. Finally, Vectashield (Vector Laboratories, Newark, CA, USA) was added onto the slides, and covered with a coverslip (#0) for storage at 4°C. Primary antibodies used: 47-5, which binds to the G/E subunits of the V1 complex of V-ATPase (made in mouse) (kindly gifted by Hans Merzendorfer and Helmut Wieczorek) (3:25), chicken anti-GFP (1:500). Secondary fluorescent antibodies: goat anti-mouse Alexa 546 (1:250), goat anti-chicken Alexa 488 (1:250), goat anti-rabbit Alexa 488 (1:250), goat anti-mouse Alexa 633. Imaging of antennal sections was performed using a Zeiss cLSM880 confocal microscope (Carl Zeiss, Oberkochen, Germany) with a 10× and 40×/1.20 water immersion objective (C-Apochromat, Carl Zeiss). Samples were excited with 488 nm, 543 nm and 633 nm wavelengths for different experiments. Laser and detector gain for two channels were optimized to avoid saturation and all the images were acquired with the same confocal settings for contrast and brightness. Z-stack images were captured at 2048 × 2048-pixel resolution at 8-bit color depth and presented as maximum intensity projections.
3. Results & Discussions
To first gauge whether V-ATPase is expressed in the Drosophila antenna, we consulted two independent antennal transcriptomes of D. melanogaster comparing expression of all known Drosophila V-ATPase subunit transcripts to a reference panel of antennal and pancellular housekeeping genes, across two separate studies comparing female and male antennae [43], and antennae of the wildtype Canton-S strain and atonal (ato) mutants lacking coeloconic sensilla constituting the IR olfactory subsystem [44]. Here, we considered all 36 known genes involved in the V0 and V1 (both catalytic and non-catalytic) domains of V-ATPase, and its accessory subunits [17,48,49]. We found that a large majority of these subunit transcripts are highly expressed in both datasets, with 20 of 36 genes at degrees of expression comparable to reference gene levels, in both studies, including all structural subunit types (Figure 1).
Next, to understand how abundant V-ATPase subunits are among antennal genes, we plotted the expression of all antennal genes detected within each antennal transcriptome, ordered by expression level abundance. In transcriptomes from both studies, we found that 20 common V-ATPase subunits rank among the most abundant antennal genes by expression (83rd-100th or 76th-100th percentile range in each respective study) (Figure 2). We found expression levels and percentile rankings of these abundant V-ATPase subunits to corroborate well between both studies. Depending on experimental condition, a remaining 13 V-ATPase subunits were detectably expressed in one dataset (within the 0th-40th percentile ranges) (Figure 2A), and 5 and 9 subunits in the other (0th-38th or 0th-25th percentile range for ato or Canton-S fly cohorts, respectively) (Figure 2B). Furthermore, we found no sexual dimorphic (sex-biased) expression for any gene involved in proton transmembrane transport (FDR < 0.05) [43].Only minor differences were found between ato mutant vs. wildtype flies regarding V-ATPase subunit expression: a 1.44 to 1.71-fold depletion exists for VhaSFD, VhaM9.7-b, Vha26, Vha44, Vha13, Vha16-1 and Vha100-2 above the detection threshold (FDR < 0.05) [44]. Though this initially might suggest enrichment of V-ATPase subunits within a particular (coeloconic) sensillum type, data from a similar study comparing antennal expression between wildtype flies and absent MD neurons and olfactory sensilla (amos) mutants that lack basiconic and trichoid sensilla reveals that 15 subunits (including all six aforementioned subunits) also appear depleted in the amos mutant antennal transcriptome at the same FDR threshold [50]. This likely suggests that loss of any sensillum type or olfactory subsystem contributes to V-ATPase subunit transcript depletion, and thus that V-ATPase subunits are generic components of antennal sensilla, rather than reside elsewhere in the antennal organ.
Taken together, the bulk transcriptomic analyses thus suggest most Drosophila V-ATPase subunits are present in the antenna, that a majority subset of these subunits is highly abundant in this tissue, likely constituent within sensilla, and that their expression is not sex- nor olfactory subsystem-biased.
We next wanted to infer the localization of V-ATPases bioinformatically, by assessing whether any V-ATPase subunit transcripts segregate differentially among antennal cell types. Here we consulted the antennal single-cell transcriptomes of the Fly Cell Atlas [45] and performed a differential expression analysis checking whether any V-ATPase subunits appear preferentially enriched or depleted in certain annotated classes (i.e. antennal cell type groupings). We recorded significant detections (FDR < 0.05 threshold) using a bubble plot showing magnitude of depletion/enrichment for all V-ATPase subunit gene transcripts, across all cell type classes. Here we found a general pattern wherein many antennal V-ATPase subunits were depleted in neuronal cell types, but not in non-neuronal and unannotated classes (Figure 3), the latter largely consisting of non-neuronal cells. On the other hand, though fewer, we found several cases of ATPase subunit enrichment in non-neuronal cell classes (Figure 3), indicating that V-ATPases are likelier found within non-neuronal cell populations of the antenna. For comparative purposes, inverse to this pattern, a single proton transmembrane transporter Otopetrin-like c (OtopLc), not constituent of the V-ATPase complex [51], was found specifically enriched in neuronal classes and depleted in non-neuronal cell classes (Figure 3). We therefore conclude V-ATPase subunits, specifically among proton transmembrane transporters, are likely to be localized to non-neuronal cells of the Drosophila antenna.
Subsequently, at the protein level, we wanted to check if V-ATPase might colocalize with Orco labeling Drosophila antennal OSNs, in preparations of cryosectioned antennae. For this, we used transgenic N-GFP-Orco flies expressing GFP conjugated to Orco chimerically, shown previously to label Orco-positive OSNs [47], along with the 47-5 antibody staining V-ATPase [27,28]. Following immunohistochemical preparation, we observed some co-localization of native N-GFP-Orco and V-ATPase signal in some OSNs (Figure 4). However, we suspected this might be an artifact since a vast majority of OSNs did not exhibit co-localization with V-ATPase. To clarify this, we generated DenMark flies that natively label somata and outer dendrites of OSNs. We then performed a further immunohistochemistry experiment using antibodies raised against Orco and V-ATPase in antennae of flies where OSNs are labeled with a native DenMark signal. Our results showed no co-localization of V-ATPase with either Orco or DenMark (Figure 5), though we cannot definitively rule out that V-ATPase may be closely associated with OSNs at the inner dendrite or ciliary constriction junction.
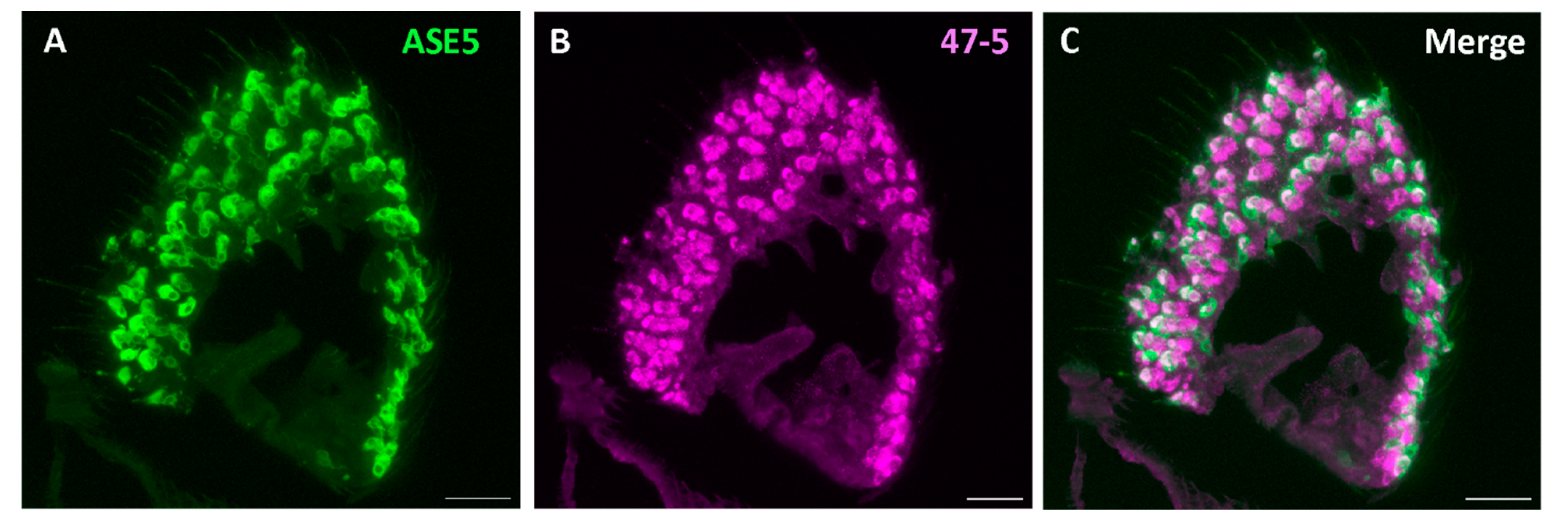
We next checked if V-ATPase is present in the non-neuronal antennal support cells. For this, we employed the available specific labeling of two of three of the support cells of the sensillum: thecogen and tormogen cells. First, a nompA-GAL4 transgenic fly line was used, driving the expression of membrane-bound GFP specifically in thecogen cells [39,52]. Though few intact thecogen cells were present, we observed a typical co-localization with the V-ATPase antibody in these thecogen cells (Figure 6). Given the appearance of other cells labeled with the V-ATPase antibody, we next used the ASE5-GAL4 line to drive membrane-bound GFP expression in tormogen cells specifically [39,53]. Here, we found more pronounced co-localization of V-ATPase protein in most observed tormogen cells (Figure 7). We also observed intermediate labelling between the thecogen and tormogen cells, which may suggest V-ATPase localization to trichogen cells, which are located between these two cell types. Because no specific method currently exists for labeling trichogen cells, we can only extrapolate that V-ATPase might be present in trichogen cells of antennal sensilla (Figure 6 and Figure 7).
In conclusion, we show that a specific majority of transcripts encoding functional subunits of Drosophila V-ATPase are highly abundantly expressed in the Drosophila antenna, in various bulk and single-cell antennal transcriptomes. Furthermore, results from immunohistochemical co-labeling of V-ATPase and antennal cell types confirm that V-ATPase is restricted to the non-neuronal support cells of the antenna, and may be entirely absent from OSNs. Though V-ATPase appears to localize to thecogen and tormogen support cells, it remains unknown if V-ATPase is explicitly expressed in trichogen, glial or epithelial cells also found in the antennal appendage. Therefore, to understand the role of V-ATPase in Drosophila olfactory sensilla, further functional physiological investigations and expression studies are necessary. Given that V-ATPase is a ubiquitous proton pump essential for maintaining acidic environments within cellular compartments [20] and likely plays crucial roles in ion homeostasis at the level of the sensillum, we purport that V-ATPase activity specifically within the support cells plays a crucial regulatory role in odor processing.
Author Contributions
Conceptualization, D.W., K.J., and S.P.; Investigation, S.P. and K.J.; Bioinformatics, S.P.; Formal Analysis, S.P.; Immunohistochemistry, K.J.; Writing – Original Draft Preparation, S.P. and K.J.; Writing – Review & Editing, S.P., K.J., B.S.H., and D.W.; Supervision, B.S.H. and D.W.
Funding
This study was supported by the Max Planck Society and the International Max Planck Research School (IMPRS) at the Max Planck Institute for Chemical Ecology in Jena, Germany.
Data Availability Statement
Original data used in this study are archived in the online data repository EDMOND at the following link: https://doi.org/10.17617/3.XVJJ8O.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Flavell, S.W. , et al., The emergence and influence of internal states. Neuron, 2022. 110(16): p. 2545-2570.
- Brand, M.D. , et al., The role of mitochondrial function and cellular bioenergetics in ageing and disease. Br J Dermatol, 2013. 169 Suppl 2(0 2): p. 1-8.
- Eaton, A.F. M. Merkulova, and D. Brown, The H(+)-ATPase (V-ATPase): from proton pump to signaling complex in health and disease. Am J Physiol Cell Physiol, 2021. 320(3): p. C392-c414. [CrossRef]
- Muench, S.P., J. Trinick, and M.A. Harrison, Structural divergence of the rotary ATPases. Q Rev Biophys, 2011. 44(3): p. 311-56.
- Kellokumpu, S. , Golgi pH, ion and redox homeostasis: how much do they really matter? Frontiers in cell and developmental biology, 2019. 7: p. 93.
- Colacurcio, D.J. and R.A. Nixon, Disorders of lysosomal acidification—The emerging role of v-ATPase in aging and neurodegenerative disease. Ageing Research Reviews, 2016. 32: p. 75-88.
- Mindell, J.A. , Lysosomal Acidification Mechanisms*. Annual Review of Physiology, 2012. 74(Volume 74, 2012): p. 69-86.
- Hnasko, T.S. and R.H. Edwards, Neurotransmitter Corelease: Mechanism and Physiological Role. Annual Review of Physiology, 2012. 74(Volume 74, 2012): p. 225-243.
- Breton, S. and D. Brown, Regulation of Luminal Acidification by the V-ATPase. Physiology, 2013. 28(5): p. 318-329. [CrossRef]
- Casey, J.R., S. Grinstein, and J. Orlowski, Sensors and regulators of intracellular pH. Nature Reviews Molecular Cell Biology, 2010. 11(1): p. 50-61.
- Paroutis, P., N. Touret, and S. Grinstein, The pH of the secretory pathway: Measurement, determinants, and regulation. Physiology, 2004. 19(4): p. 207-215.
- Maxfield, F.R. and T.E. McGraw, Endocytic recycling. Nature Reviews Molecular Cell Biology, 2004. 5(2): p. 121-132.
- Naslavsky, N. and S. Caplan, The enigmatic endosome - Sorting the ins and outs of endocytic trafficking. Journal of Cell Science, 2018. 131(13).
- Forgac, M. , Vacuolar ATPases: rotary proton pumps in physiology and pathophysiology. Nature Reviews Molecular Cell Biology, 2007. 8(11): p. 917-929.
- Williamson, W.R. and P.R. Hiesinger, On the role of v-ATPase V0a1-dependent degradation in Alzheimer disease. Commun Integr Biol, 2010. 3(6): p. 604-7.
- Bagh, M.B. , et al., Misrouting of v-ATPase subunit V0a1 dysregulates lysosomal acidification in a neurodegenerative lysosomal storage disease model. Nature Communications, 2017. 8.
- Dow, J.A. , The multifunctional Drosophila melanogaster V-ATPase is encoded by a multigene family. J Bioenerg Biomembr, 1999. 31(1): p. 75-83.
- Lee, S.-K. , et al., Vacuolar (H+)-ATPases in Caenorhabditis elegans: What can we learn about giant H+ pumps from tiny worms? Biochimica et Biophysica Acta (BBA) - Bioenergetics, 2010. 1797(10): p. 1687-1695.
- Dow, J. , et al., Molecular genetic analysis of V-ATPase function in Drosophila Melanogaster. The Journal of experimental biology, 1997. 200: p. 237-45.
- Wieczorek, H. , et al., Vacuolar-type proton pumps in insect epithelia. Journal of Experimental Biology, 2009. 212(11): p. 1611-1619.
- Sun-Wada, G.-H. and Y. Wada, Role of vacuolar-type proton ATPase in signal transduction. Biochimica et Biophysica Acta (BBA) - Bioenergetics, 2015. 1847(10): p. 1166-1172.
- Ramsay, J.A. , Active Transport of Potassium by the Malpighian Tubules of Insects. Journal of Experimental Biology, 1953. 30(3): p. 358-369.
- Wolfersberger, M.G., W. R. Harvey, and M. Cioffi, Chapter 6 Transepithelial Potassium Transport in Insect Midgut by an Electrogenic Alkali Metal Ion Pump, in Current Topics in Membranes and Transport, A. Kleinzeller, F. Bronner, and C.L. Slayman, Editors. 1982, Academic Press. p. 109-133.
- Harvey, W.R., M. Cioffi, and M.G. Wolfersberger, Chemiosmotic potassium ion pump of insect epithelia. Am J Physiol, 1983. 244(2): p. R163-75.
- Harvey, W.R. , et al., H + V-ATPases Energize Animal Plasma Membranes for Secretion and Absorption of Ions and Fluids'. Integrative and Comparative Biology, 1998. 38: p. 426-441.
- Wieczorek, H. , et al., A vacuolar-type proton pump energizes K+/H+ antiport in an animal plasma membrane. J Biol Chem, 1991. 266(23): p. 15340-7.
- Klein, U. and B. Zimmermann, The vacuolar-type ATPase from insect plasma membrane: immunocytochemical localization in insect sensilla. Cell and Tissue Research, 1991. 266(2): p. 265-273.
- Klein, U., G. Löffelmann, and H. Wieczorek, The Midgut as a Model System for Insect K+-Transporting Epithelia: Immunocytochemical Localization of a Vacuolar-Type H+ Pump. Journal of Experimental Biology, 1991. 161(1): p. 61-75.
- Thurm, U. and J. Küppers, Epithelial physiology of insect sensilla, in Insect biology in the future. 1980, Elsevier. p. 735-763.
- Wieczorek, H. , et al., A Vacuolar-type Proton Pump in a Vesicle Fraction Enriched with Potassium Transporting Plasma Membranes from Tobacco Hornworm Midgut. Journal of Biological Chemistry, 1989. 264(19): p. 11143-11148.
- Küppers, J. and I. Bunse, A Primary Cation Transport by a V-Type Atpase of Low Specificity. Journal of Experimental Biology, 1996. 199(6): p. 1327-1334.
- Weng, X.-H. , et al., The V-type H+-ATPase in Malpighian tubules of Aedes aegypti: localization and activity. Journal of Experimental Biology, 2003. 206: p. 2211 - 2219.
- Davies, S.A. , et al., Analysis and inactivation of vha55, the gene encoding the vacuolar ATPase B-subunit in Drosophila melanogaster reveals a larval lethal phenotype. Journal of Biological Chemistry, 1996. 271(48): p. 30677-30684.
- Nava Gonzales, C. , et al., Systematic morphological and morphometric analysis of identified olfactory receptor neurons in Drosophila melanogaster. eLife, 2021. 10: p. e69896.
- Shanbhag, S.R., B. Müller, and R.A. Steinbrecht, Atlas of olfactory organs of Drosophila melanogaster: 1. Types, external organization, innervation and distribution of olfactory sensilla. International Journal of Insect Morphology and Embryology, 1999. 28(4): p. 377-397.
- Shanbhag, S.R., B. Müller, and R.A. Steinbrecht, Atlas of olfactory organs of Drosophila melanogaster: 2. Internal organization and cellular architecture of olfactory sensilla. Arthropod Structure & Development, 2000. 29(3): p. 211-229.
- Shanbhag, S.R., B. Müller, and R.A. Steinbrecht, Atlas of olfactory organs of Drosophila melanogaster 2. Internal organization and cellular architecture of olfactory sensilla. Arthropod Struct Dev, 2000. 29(3): p. 211-29.
- Vosshall, L.B. and R.F. Stocker, Molecular architecture of smell and taste in Drosophila. Annu Rev Neurosci, 2007. 30: p. 505-33.
- Prelic, S. , et al., Functional Interaction Between Drosophila Olfactory Sensory Neurons and Their Support Cells. Front Cell Neurosci, 2021. 15: p. 789086.
- Neuhaus, E.M. , et al., Odorant receptor heterodimerization in the olfactory system of Drosophila melanogaster. Nature Neuroscience, 2005. 8(1): p. 15-17.
- Vosshall, L.B. and B.S. Hansson, A Unified Nomenclature System for the Insect Olfactory Coreceptor. Chemical Senses, 2011. 36(6): p. 497-498.
- Dow, J.A.T. , V-ATPases in Insects, in Organellar Proton-ATPases. 1995, Springer Berlin Heidelberg: Berlin, Heidelberg. p. 75-102.
- Shiao, M.-S. , et al., Transcriptional profiling of adult Drosophila antennae by high-throughput sequencing. Zoological Studies, 2013. 52(1): p. 42.
- Menuz, K. , et al., An RNA-Seq Screen of the Drosophila Antenna Identifies a Transporter Necessary for Ammonia Detection. PLOS Genetics, 2014. 10(11): p. e1004810.
- Li, H. , et al., Fly Cell Atlas: A single-nucleus transcriptomic atlas of the adult fruit fly. Science, 2022. 375(6584): p. eabk2432.
- Chen, W. , et al., Profiling of Single-Cell Transcriptomes. Current Protocols in Mouse Biology, 2017. 7(3): p. 145-175.
- Jain, K. , et al., A new Drosophila melanogaster fly that expresses GFP-tagged Orco. Frontiers in Ecology and Evolution, 2023. 11.
- Chintapalli, V.R. , et al., Data-mining the FlyAtlas online resource to identify core functional motifs across transporting epithelia. BMC Genomics, 2013. 14(1): p. 518.
- Allan, A.K. , et al., Genome-wide survey of V-ATPase genes in Drosophila reveals a conserved renal phenotype for lethal alleles. Physiological Genomics, 2005. 22(2): p. 128-138.
- Mohapatra, P. and K. Menuz, Molecular Profiling of the Drosophila Antenna Reveals Conserved Genes Underlying Olfaction in Insects. G3 Genes|Genomes|Genetics, 2019. 9(11): p. 3753-3771.
- Ganguly, A. , et al., Requirement for an Otopetrin-like protein for acid taste in Drosophila. Proceedings of the National Academy of Sciences, 2021. 118(51): p. e2110641118.
- Chung, Y.D. , et al., nompA Encodes a PNS-Specific, ZP Domain Protein Required to Connect Mechanosensory Dendrites to Sensory Structures. Neuron, 2001. 29(2): p. 415-428.
- Barolo, S. , et al., A Notch-Independent Activity of Suppressor of Hairless Is Required for Normal Mechanoreceptor Physiology. Cell, 2000. 103(6): p. 957-970.
Figure 1.
Bulk antennal tissue expression of Drosophila V-ATPase subunit and other genes with known involvement in proton transmembrane transport, alongside antennal and housekeeping reference genes. Myriad V-ATPase genes are highly abundant in expression in various antennal tissues, involving all functional subunits. Two antennal RNA-seq datasets are shown contrasting expression across fly sex [43] and wildtype Canton-S vs. ato mutant flies lacking coeloconic sensilla of the IR olfactory subsystem [44].
Figure 1.
Bulk antennal tissue expression of Drosophila V-ATPase subunit and other genes with known involvement in proton transmembrane transport, alongside antennal and housekeeping reference genes. Myriad V-ATPase genes are highly abundant in expression in various antennal tissues, involving all functional subunits. Two antennal RNA-seq datasets are shown contrasting expression across fly sex [43] and wildtype Canton-S vs. ato mutant flies lacking coeloconic sensilla of the IR olfactory subsystem [44].
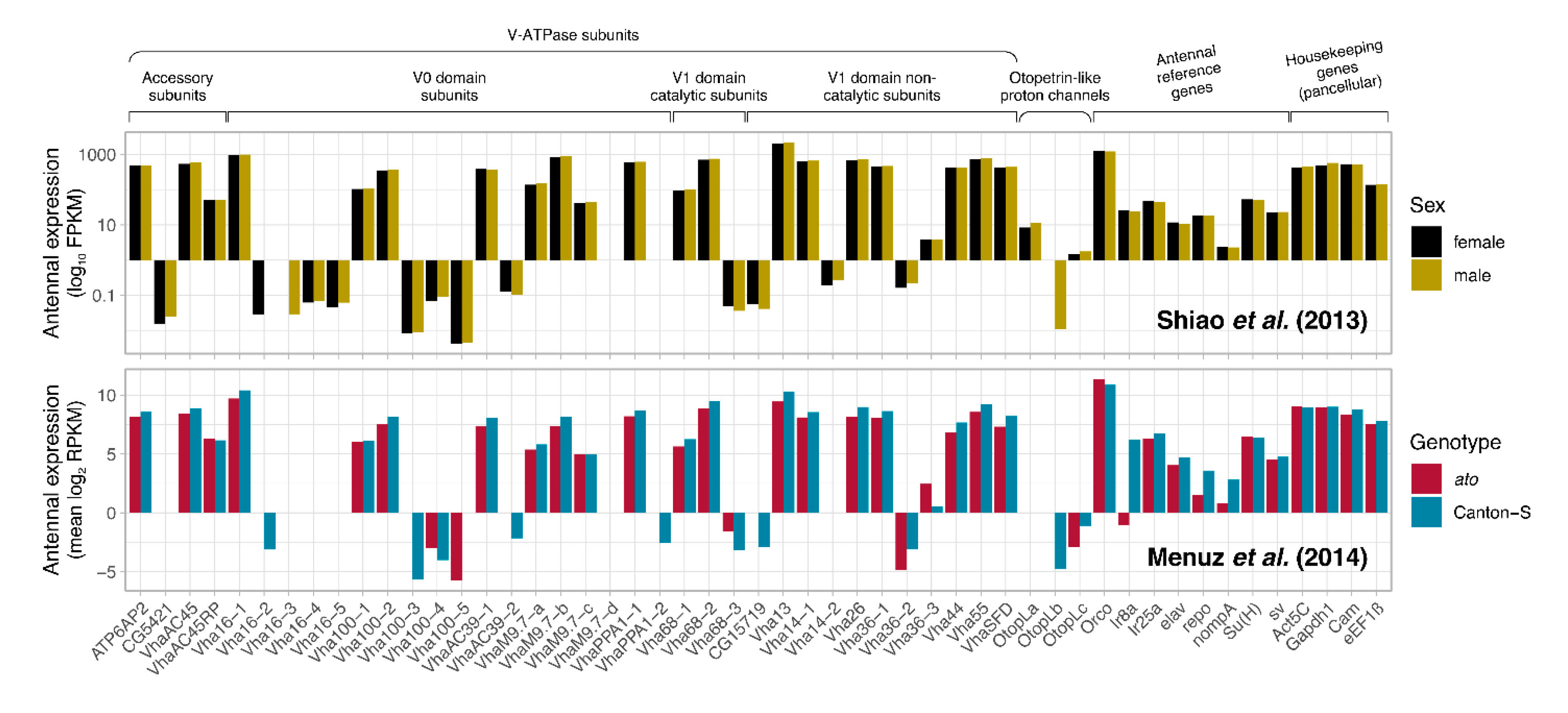
Figure 2.
V-ATPase subunit expression abundance in various antennal transcriptomes. Expression of all genes detected in the Drosophila antennal datasets [43,44] is plotted, colored and ordered low-to-high by its expression abundance. All known genes involved in proton transmembrane transport (including all V-ATPase subunits) are labeled, whereby labels are vertically displaced from where the gene appears. V-ATPase subunit expression is compared across fly sex (A) and between fly cohorts with intact or absent coeloconic sensilla constituent of the IR olfactory subsystem (B).
Figure 2.
V-ATPase subunit expression abundance in various antennal transcriptomes. Expression of all genes detected in the Drosophila antennal datasets [43,44] is plotted, colored and ordered low-to-high by its expression abundance. All known genes involved in proton transmembrane transport (including all V-ATPase subunits) are labeled, whereby labels are vertically displaced from where the gene appears. V-ATPase subunit expression is compared across fly sex (A) and between fly cohorts with intact or absent coeloconic sensilla constituent of the IR olfactory subsystem (B).
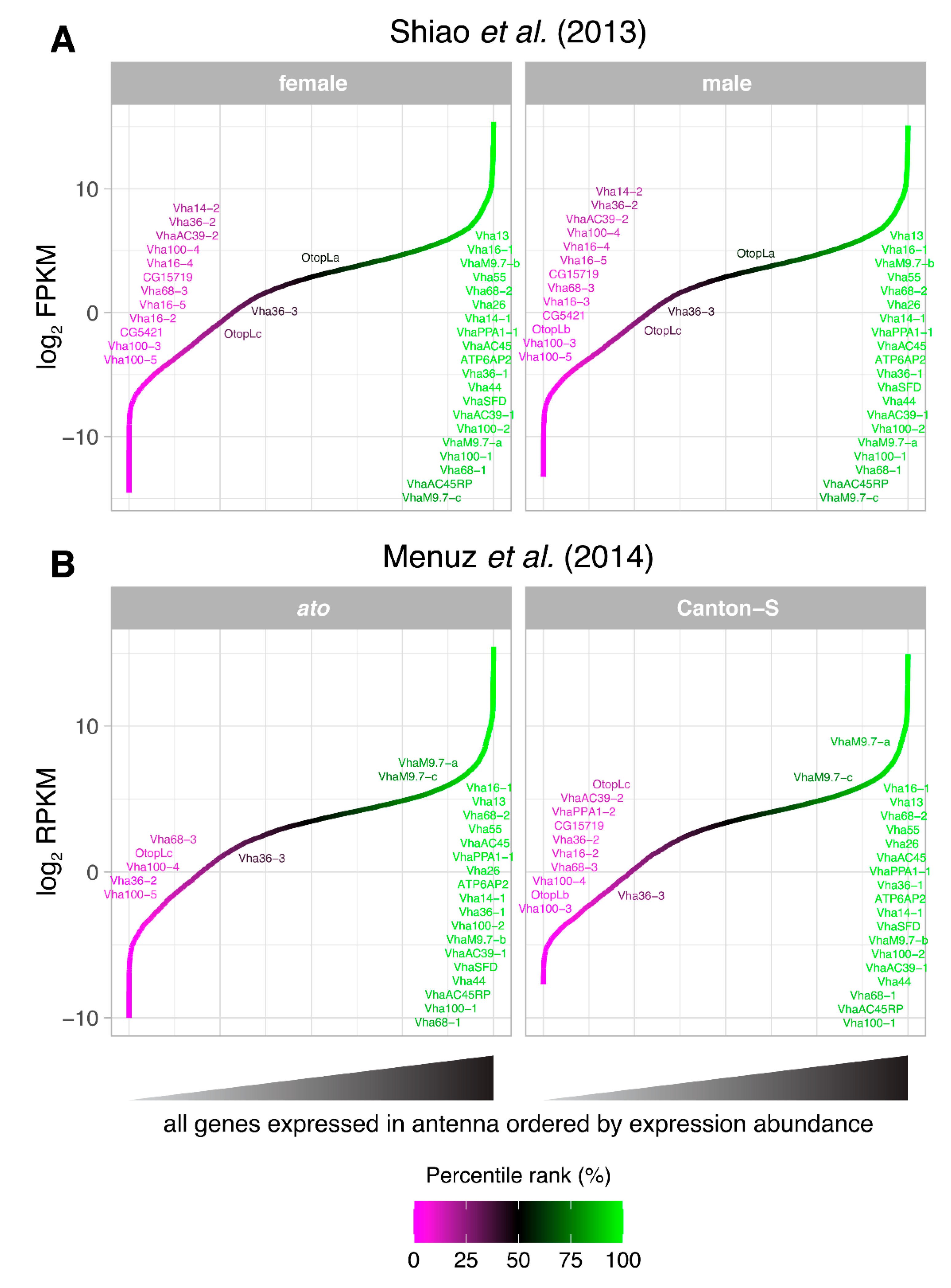
Figure 3.
Differential expression of all Drosophila genes with known involvement in proton transmembrane transport (including all V-ATPase subunits) in the Fly Cell Atlas single-cell antennal transcriptome. Gene expression for each annotated cell type class is compared with all other antennal cells using a Wilcoxon test. Circles are plotted only where significant differential gene expression is detected for a cell type class (FDR < 0.05), colored by relative up- (blue) or downregulation (red) and sized based on absolute expression difference magnitude. Circles are filled only when gene expression difference is larger than a two-fold difference in expression (|log2FC| > 1). Genes in bold have been identified prior as highly abundant in antennal tissue. FC: foldchange.
Figure 3.
Differential expression of all Drosophila genes with known involvement in proton transmembrane transport (including all V-ATPase subunits) in the Fly Cell Atlas single-cell antennal transcriptome. Gene expression for each annotated cell type class is compared with all other antennal cells using a Wilcoxon test. Circles are plotted only where significant differential gene expression is detected for a cell type class (FDR < 0.05), colored by relative up- (blue) or downregulation (red) and sized based on absolute expression difference magnitude. Circles are filled only when gene expression difference is larger than a two-fold difference in expression (|log2FC| > 1). Genes in bold have been identified prior as highly abundant in antennal tissue. FC: foldchange.
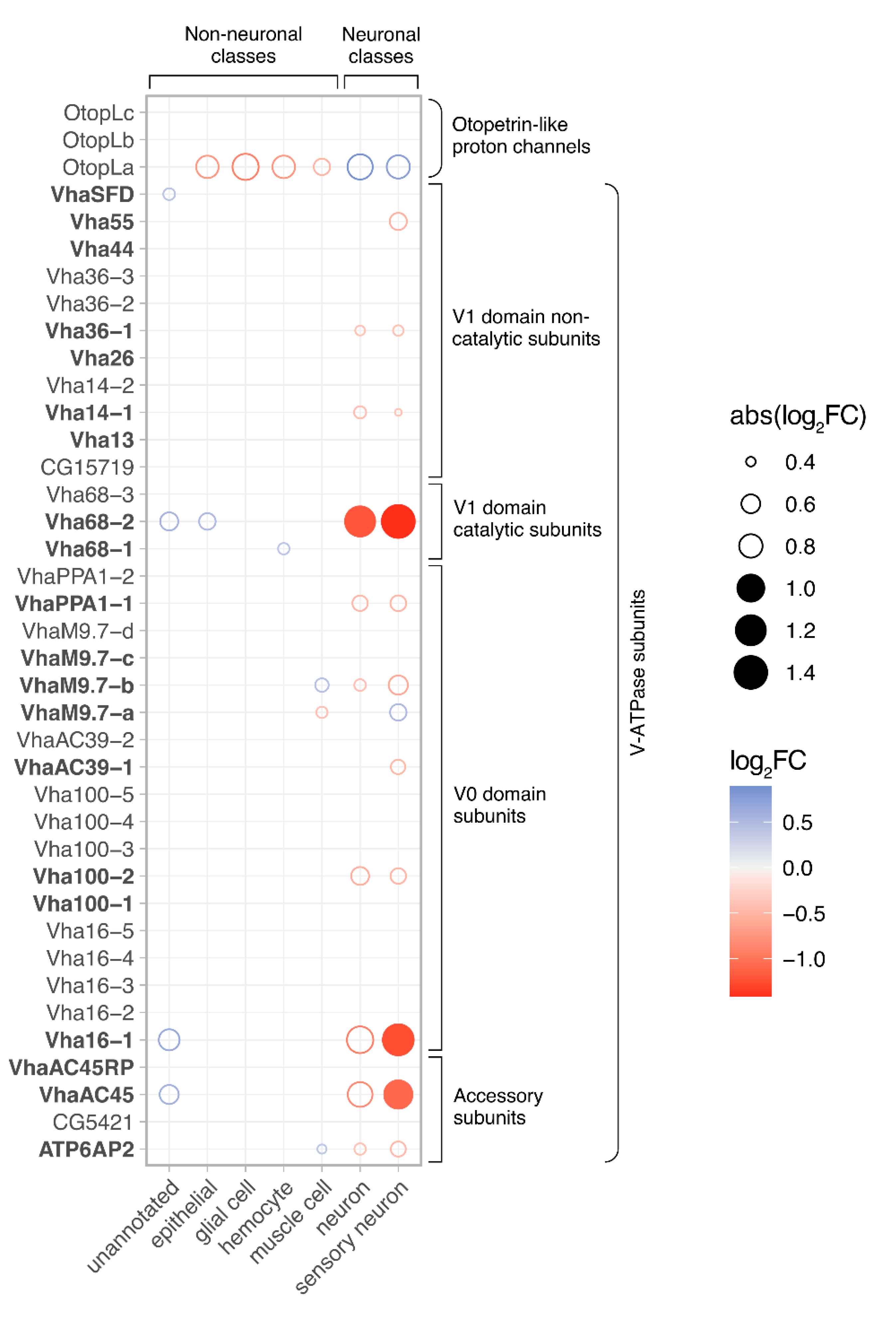
Figure 4.
Expression pattern of V-ATPase in the antennal cross sections of N-GFP-Orco D. melanogaster fly line. (A) Native GFP signal from the N-GFP-Orco fly. (B) Immunofluorescence micrographs of V-ATPase (47-5, magenta) staining of the same antennal cross sections as shown in (A). (C) No colocalization of GFP and V-ATPase observed in compartments of Drosophila OSNs. Scale bar: 17 µm.
Figure 4.
Expression pattern of V-ATPase in the antennal cross sections of N-GFP-Orco D. melanogaster fly line. (A) Native GFP signal from the N-GFP-Orco fly. (B) Immunofluorescence micrographs of V-ATPase (47-5, magenta) staining of the same antennal cross sections as shown in (A). (C) No colocalization of GFP and V-ATPase observed in compartments of Drosophila OSNs. Scale bar: 17 µm.
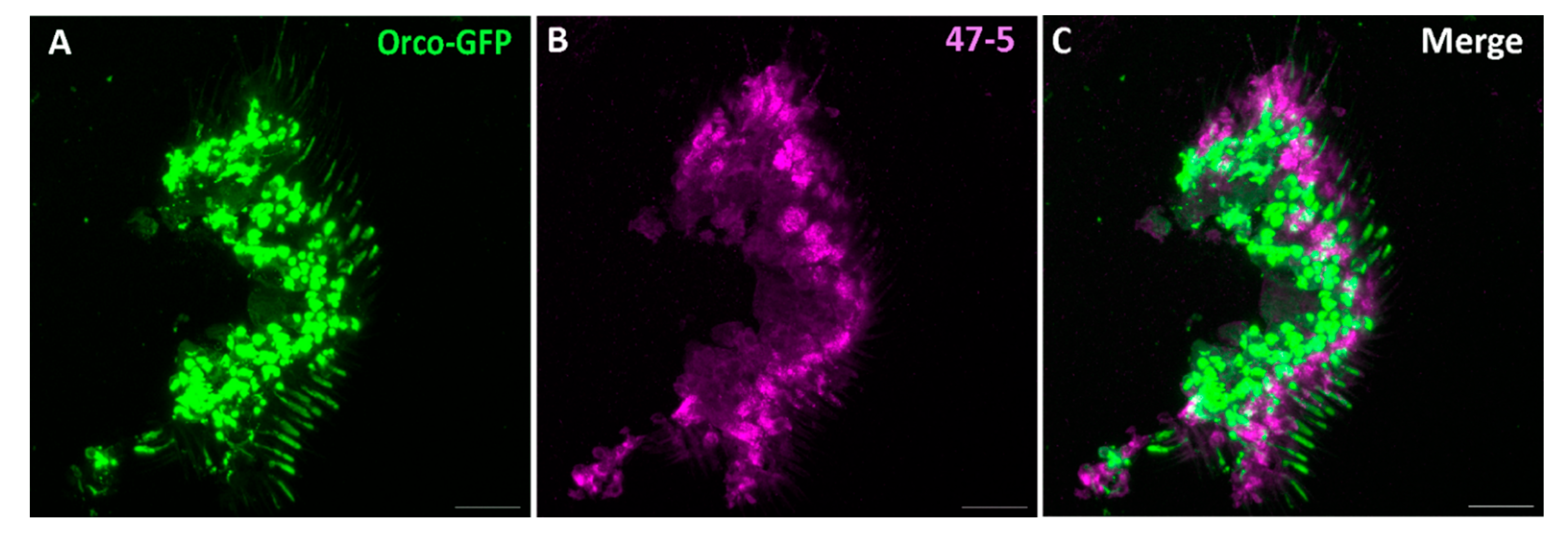
Figure 5.
Expression pattern of V-ATPase in the antennal cross sections of DenMark expressing D. melanogaster fly line. Immunofluorescence staining of DenMark (red) (A), Orco-positive OSNs (green) (B), V-ATPase protein (47-5, magenta) (C). (D) Absence of V-ATPase in olfactory sensory neurons and colocalization of DenMark and Anti-Orco in these OSNs. Scale bar: 17 µm.
Figure 5.
Expression pattern of V-ATPase in the antennal cross sections of DenMark expressing D. melanogaster fly line. Immunofluorescence staining of DenMark (red) (A), Orco-positive OSNs (green) (B), V-ATPase protein (47-5, magenta) (C). (D) Absence of V-ATPase in olfactory sensory neurons and colocalization of DenMark and Anti-Orco in these OSNs. Scale bar: 17 µm.
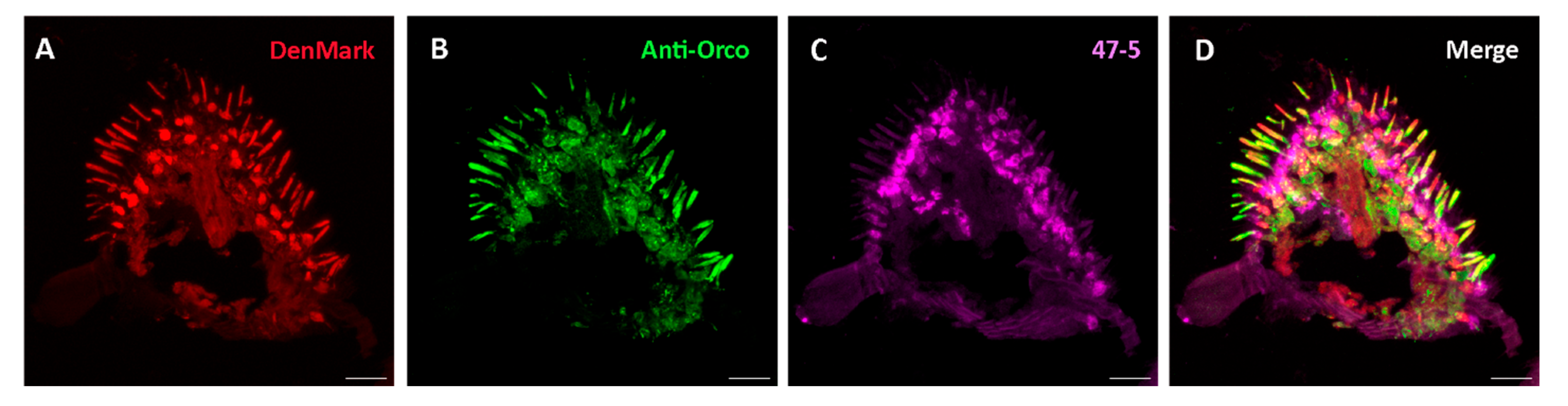
Figure 6.
Expression pattern of V-ATPase in the thecogen cells of D. melanogaster antennal cross sections. (A) Immunofluorescence micrographs of mCD8::GFP (α-GFP, green) and V-ATPase (47-5, magenta). (B) from the antennal cross sections of nompA GAL4 fly line labelling thecogen cells. (C) Observation of partial colocalization of mCD8::GFP and V-ATPase in the thecogen cells. Scale bar: 17 µm.
Figure 6.
Expression pattern of V-ATPase in the thecogen cells of D. melanogaster antennal cross sections. (A) Immunofluorescence micrographs of mCD8::GFP (α-GFP, green) and V-ATPase (47-5, magenta). (B) from the antennal cross sections of nompA GAL4 fly line labelling thecogen cells. (C) Observation of partial colocalization of mCD8::GFP and V-ATPase in the thecogen cells. Scale bar: 17 µm.

Figure 7.
Expression pattern of V-ATPase in the tormogen cells of D. melanogaster antennal cross-sections. (A) Immunofluorescence image of mCD8::GFP (α-GFP, green) and V-ATPase (47-5, magenta). (B) from the antennal cross sections of ASE5-GAL4 fly line labelling tormogen cells. (C) Observation of major colocalization of mCD8::GFP and V-ATPase in the tormogen cells. Scale bar: 17 µm.
Figure 7.
Expression pattern of V-ATPase in the tormogen cells of D. melanogaster antennal cross-sections. (A) Immunofluorescence image of mCD8::GFP (α-GFP, green) and V-ATPase (47-5, magenta). (B) from the antennal cross sections of ASE5-GAL4 fly line labelling tormogen cells. (C) Observation of major colocalization of mCD8::GFP and V-ATPase in the tormogen cells. Scale bar: 17 µm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



