Submitted:
14 November 2024
Posted:
14 November 2024
You are already at the latest version
Abstract
Terminalia bellirica, and Terminalia chebula are significant botanicals in ancient Ayurvedic medicine. They are renowned for their therapeutic properties, notably in addressing gastrointestinal (GI) diseases. These plants have undergone thorough examination related to their antibacterial, anti-inflammatory, and antioxidant properties, which make them highly efficient natural treatments for controlling gastrointestinal infections. The current research demonstrated the antibacterial efficacy of fruit extracts of Terminalia bellirica, and Terminalia chebula against Bacillus cereus, Shigella sonnei, Shigella flexneri, and Salmonella typhimurium. We performed disc diffusion and liquid microdilution experiments to evaluate the antibacterial efficacy. All extracts of Terminalia bellirica and Terminalia chebula showed good antibacterial effects against B. cereus and S. flexneri. The minimum inhibitory concentration (MIC) values ranged from 94 µg/mL to 556 µg/mL. The methanol extracts from both plants also showed noteworthy antibacterial activity against S. sonnei and S. typhimurium, with MIC values of 755 µg/mL for both. Fractional inhibitory concentration studies revealed additive interactions between the extracts and conventional antibiotics when used concurrently. The phytochemical composition of T. bellirica and T. chebula extracts was analysed using liquid chromatography-mass spectrometry (LC-MS), which identified several tannins, including methyl gallate, propyl gallate, gallic acid, and ellagic acid. Lethality assays conducted using Artemia franciscana Kellogg nauplii indicated that all plant extracts are non-toxic. The antibacterial properties and absence of toxicity in T. bellirica and T. chebula fruit extracts indicate their potential for antibiotic development, warranting additional mechanistic and phytochemical studies.
Keywords:
Medicinal plants
; plant-derived antimicrobials
; gallotannins
; gastrointestinal pathogens
; herbal antidiarrheals
; combinational therapies
; LC-MS
1. Introduction
Diarrhoeal disease is the third most common cause of mortality among children aged five and below, causing more than 443,000 deaths annually [1]. Diarrhoea can persist for multiple days and deplete the body of vital water and minerals essential for survival. Historically, severe dehydration and fluid depletion were the primary factors leading to fatalities related with diarrhoea. Diarrhoea often arises as a manifestation of an infection in the gastrointestinal tract, which can be triggered by a range of bacterial, viral, and parasite microorganisms [1].
Common bacterial pathogens involved in the development of diarrhoeal infections include Escherichia coli, Salmonella spp., Shigella spp., Campylobacter spp., Clostridium difficile, Yersinia spp. etc [1]. Notably, gastrointestinal bacterial infections are increasingly contributing to the overall mortality associated with diarrhoea. The individuals most vulnerable to life-threatening diarrhoea include children and the elderly, people with malnutrition or weakened immune systems, as well as individuals living with HIV. The transmission of GI infections occurs through the consumption of contaminated food or drinking-water, or through person-to-person contact due to inadequate hygiene practices [1]. The global economic burden of diarrheal diseases is substantial, costing an estimated $12 billion annually in healthcare expenses, productivity losses, and long-term disability [2]. This significant financial impact underscores the need for improved prevention and treatment strategies.
Bacillary dysentery is a condition that is marked by severe diarrhoea, and S. flexneri and S. sonnei are two of the most important pathogens that are linked with it [3]. These pathogens are highly contagious, especially in areas with inadequate sanitation. Due to their vulnerability, these infections pose a substantial public health issue, particularly the elderly and young children [3]. In addition, S. typhimurium is a prevalent cause of foodborne illness, spreading through contaminated poultry and eggs, resulting in symptoms ranging from mild gastroenteritis to more severe forms of the disease [4]. Furthermore, B. cereus produces toxins that can result in food poisoning, leading to vomiting and diarrhoea [5]. This condition is often linked to inadequate food storage or the reheating of starchy foods. Additionally, Bacillus species pose an additional challenge due to their psychotropic characteristics, allowing them to survive and grow at low temperatures. This reduces the effectiveness of refrigeration as a food preservation method, necessitating alternative strategies to address contamination and spoilage. Bacillus cereus infections generally resolve spontaneously, although they pose a considerable risk to vulnerable populations, including children and the elderly [5].
Multiple strains of S. flexneri and S. sonnei have recently developed increased resistance to a range of antibiotics including fluoroquinolones, cephalosporins, azithromycin, sulphonamides, tetracycline, and streptomycin [6,7]. Similarly, some S. typhimurium strains have developed resistance to ampicillin, chloramphenicol, streptomycin, sulphonamides and tetracycline [8]. Additionally, some B. cereus strains exhibit significant resistance to β-lactam antibiotics such as penicillin G and cefotaxime [9,10].
Traditional herbal treatments may be useful in developing novel therapies to combat antimicrobial resistant gastrointestinal infections. Secondary metabolites of herbal plants, such as alkaloids, flavonoids, phenolic acids, tannins, and terpenoids play crucial role in a plant’s defence system and protect from infections [11]. Ayurvedic medicinal plants are a promising target for developing novel antibiotic chemotherapy treatments due to their easy accessibility, cost effectiveness, and expanding evidence-based efficacy [11]. A previous study from our group highlighted the role of traditionally used Ayurvedic medicinal plants in the treatment and management of dysentery, diarrhoea, and other gastrointestinal infections [12]. In particular, Terminalia bellirica (Gaertn.) Roxb. and Terminalia chebula Retz fruit decoctions, infusions or pulp were highlighted for their traditional uses to treat dysentery and chronic diarrhoea. Numerous other medicinal plants, including Croton lechleri Linn., Mentha piperita Linn., Camellia sinensis (L.) Kuntze, Emblica officinalis Linn., Phyllanthus niruri Linn., and Azadirachta indica A. Juss. have also been reported to possess antidiarrhoeal activity [12,13]. Furthermore, several constituents of these plants s, including apigenin, friedelin, 1,8-cineole, eriosematin E, and stachysrosane (1) and (2), have been found to exhibit antidiarrhoeal activity in a castor oil-induced diarrhoea model [13].
Potent antibacterial activity has previously been reported for aqueous and ethanolic fruit extracts of T. bellirica and T. chebula, against S. flexneri and S. sonnei, with MIC values ranging from 0.01 to 100 µg/mL [14]. However, the methodology and preparation of the extracts was not clearly stated, making these results difficult to duplicate. In addition, no toxicity studies were conducted, and the antibacterial assays were not performed in duplicates or triplicates to confirm the results. In contrast, other studies reported very low antibacterial activity for aqueous T. bellirica fruit extracts against S. flexneri, with MIC values = 12,500 µg/mL [15]. Similarly, another study showed low antibacterial activity of aqueous and alcoholic T. bellirica fruit extracts in agar well diffusion method against S. typhimurium (MIC = 12,500 µg/mL) [16]. Aqueous T. bellirica fruit extracts have also been reported to reduce bacterial loads in a S. typhimurium infected animal model [17]. In a previous investigation from our group, we noted that the aqueous and methanolic fruit extracts of T. bellirica and T. chebula exhibit low to moderate antibacterial activity against S. typhimurium and S. sonnei [18]. In contrast, the methanolic fruit extracts of both species exhibited high antibacterial activity against S. flexneri, with MIC values of 287 µg/mL and 276 µg/mL respectively.
Our research study examined antibacterial activities and the phytochemical constituents of the aqueous, methanolic and ethyl acetate extracts of T. bellirica and T. chebula. Shigella flexneri, Shigella sonnei, Salmonella typhimurium, and Bacillus cereus were used for the antibacterial analyses. Liquid chromatography – mass spectrometry (LC-MS) was conducted to examine the phytochemical composition of T. bellirica and T. chebula extracts, with a focus on identifying noteworthy flavonoids, tannins and terpenoid compounds. Interactions between any active plant extracts and selected reference antibiotics were investigated, alongside extract toxicity determinations using Artemia franciscana Kellogg nauplii lethality assays.
2. Materials and Methods
2.1. Sources of Plant Samples
Terminalia bellirica fruit powder (batch no: BNFP/01) was manufactured by Organic Prime and purchased from Navafresh, Australia. The Terminalia chebula fruit powder (batch no: HRP1020) was formulated by Aarshaveda (Sattvic, Australia). The supplier's website listed the traditional Ayurvedic names for T. bellirica (Bhitaki, Baheda) and T. chebula (Haritaki, Harad) and also confirmed the authenticity and quality of the plant materials. Voucher specimens of the T. bellirica fruit (NBG-TB0220GU) and T. chebula fruit (NBG-TC0220GU) are stored at Griffith University's Gold Coast campus in the School of Pharmacy and Medical Sciences.
2.2. Preparation of Extracts
Individual 1 g masses of either T. bellirica or T. chebula fruit powders were added to six 50 mL Falcon tubes. Sterile deionized water, methanol (AR grade) or ethyl acetate (AR grade) were then added separately to individual tubes to make a total volume of 50 mL [18,19]. The organic solvents methanol and ethyl acetate were supplied by ChemSupply (Gillman, Australia). The extraction tubes were thoroughly mixed at ambient temperature for 24 hours using continuous rolling (30 rpm). Subsequently, the samples were filtered using Whatman No. 54 filter paper (Sigma-Aldrich, Melbourne, Australia) into pre-weighed 50 mL tubes under negative vacuum pressure. The aqueous samples were subjected to lyophilization using an Alpha 1-4 LSC plus benchtop freeze dryer (Martin Christ, Osterode am Harz, Germany) for three days to freeze-dried. The extracts containing organic solvent samples were evaporated at 40°C for a duration of two days, or until the evaporation process was completed. To determine the final yields, the weight of all dried extracts was measured. The extracts were then reconstituted in 10 mL of a 1% solution of dimethyl sulfoxide (DMSO; Merck, Macquarie Park, Australia) and subjected to three rounds of sonication. Each round consisted of 20-second pulses from a probe sonicator set at 1kHz, with a 30-second break between pulses. The extracts were sterilised by filtration through 0.22 µm filters (Sarstedt, Mawson Lakes, Australia) and then stored at -20 °C until they were used.
2.3. Antibiotics and Bacterial Strains
The following antibiotic powders were purchased from Sigma-Aldrich (Melbourne, Australia): Penicillin G (potency range of 1440-1680 µg/mg), erythromycin (potency ≥850 µg/mg), tetracycline (purity ≥95% as determined by HPLC), chloramphenicol (≥98% as determined by HPLC), ciprofloxacin (purity ≥98% as determined by HPLC), polymyxin B (purity ≥90%), oxacillin (purity ≥95% as determined by TLC), amoxycillin (potency of 900 µg/mg), gentamicin (purity ≥98% as determined by HPLC), and vancomycin (potency ≥900 μg/mg). Stock solutions of antibiotics at a concentration of 1 mg/mL were prepared for use in broth microdilution experiments and stored at a temperature of -20 °C until they were needed. Preloaded standard antibiotic susceptibility testing discs were purchased from Oxoid Ltd. (Thebarton, Australia) and included penicillin G (10 IU), erythromycin (10 µg), tetracycline (30 µg), chloramphenicol (30 µg), ciprofloxacin (1 µg), polymyxin B (300IU), oxacillin (1 µg), gentamicin (10 µg), vancomycin (30 µg), cefoxitin (30 µg), or Augmentin® (15 µg). Additionally, 10 µL of an amoxycillin solution (0.01 mg/mL) was infused into sterile filter paper discs and used as an additional antibiotic control. Broth microdilution experiments were performed using liquid solutions of all reference antibiotics, except for Augmentin® and cefoxitin, which were unavailable.
Bacterial species were purchased from the American Type Culture Collection (ATCC; Manassas, VA, USA) and included Shigella sonnei (ATCC 25931), Salmonella typhimurium (Salmonella enterica serovar Typhimurium; ATCC 14028), Shigella flexneri (ATCC 12022), and Bacillus cereus (ATCC 14579). Oxoid Ltd. (Australia) supplied the Mueller–Hinton (MH) agar and broth (Oxoid Ltd., Australia) to sustain bacterial growth. Each MH agar plate was prepared according to manufacturers’ instructions at an agar depth of 4 mm.
2.4. Antibacterial Susceptibility Screening
The antibacterial activity of all plant extracts in MH agar was examined using a modified Kirby–Bauer disc diffusion technique [19]. Briefly, individual colonies were isolated from streaked MH agar plates and subsequently inoculated in 40 mL of fresh MH broth. The bacteria were cultivated at 37 °C for 18–24 hours. McFarland 0.5 standards were prepared for each strain using the individual bacterial cultures and 100 µL of the 0.5 McFarland standards were individually distributed evenly on MH agar plates. Sterile forceps were used to attach 6 mm diameter Whatman sterile filter paper discs to MH agar surface. Then, 10 µL of the extracts (dissolved in 1% DMSO) were added to individual discs. Each bacterial strain was inoculated on MH agar, and all reference antibiotic discs were inserted. The extract and reference antibiotic plates were incubated at 37 °C until confluent growth was achieved (18-24 hours). The samples were analysed in triplicate. The diameter of the inhibition zones (ZOI) surrounding each disc was measured to the nearest whole millimetre to assess the inhibition of bacterial growth. Samples lacking antibacterial activity were deemed to have a ZOI of 6 mm (equal to the diameter of the test disc).
2.5. Minimum Inhibitory and Fractional Ihibitory Concentrations
The extracts and most of the reference antibiotics were subjected to broth microdilution assays in order to obtain minimum inhibitory concentration (MIC) values, as described previously [18,19]. Values ≥10,000 μg/mL were categorised as inactive, whereas those falling within the range of 2000 to 10,000 μg/mL were deemed to possess low activity. MIC values ranging from 1000 to 2000 μg/mL were categorised as moderate activity, values between 400 and 1000 μg/mL were considered noteworthy activity, and values between 100 and 400 μg/mL were classified as good activity. Any MIC values that were less than 100 μg/mL were considered to have a high level of activity.
Plant extracts and reference antibiotics that showed antibacterial activity with MIC ≤ 3000 µg/mL and ≤2.5 µg/mL respectively were combined in equal ratios (50:50) and retested using the MIC methodology described above. The sum of the fractional inhibitory concentrations (ΣFIC) for each combination was determined using the equations shown below (a = extracts; b = antibiotics):
FIC(a) = MIC (a in combination with b)/MIC (a independently)
FIC(b) = MIC (b in combination with a)/MIC (b independently).
The sum of the fractional inhibitory concentrations (∑FIC) was determined by:
∑FIC = FIC(a) + FIC(b).
The classifications of the interactions included synergistic (∑FIC ≤ 0.5), additive (∑FIC > 0.5–≤ 1.0), indifferent (∑FIC > 1.0–≤ 4.0), or antagonistic (∑FIC > 4.0) [20].
2.7. Toxicity Assays
Extracts and control samples were subjected to Artemia franciscana Kellogg nauplii lethality assays (ALA) [21] as an indicator of their toxicities. Extracts were diluted to 2 mg/mL in artificial seawater and 400 µL of these samples or 400 µL of artificial seawater (Red Sea Pty. Ltd.) were added to sterile 48-well plates followed by the addition of approximately 50 newly hatched A. franciscana nauplii. A 400 µL volume of the negative (32 g/L artificial seawater; Red Sea) and positive controls (10 mg/mL sodium azide) were also added to all plates for comparison. The plates were incubated at 25 °C ± 1 °C for 24 hours and then the viability of the brine shrimp nauplii were assessed. Probit analysis was used to calculate the LC50 values, which indicate the concentration of extract or control needed to cause the death of 50% of A. franciscana nauplii in individual wells. The LC50 values were determined by graphically calculating the mean percentage of mortality from three separate experiments, which represents the concentration that caused 50% mortality.
2.8. Non-Targeted Headspace LC-MS Workflow for Quantitative Analysis
The Vanquish Ultra High-Performance Liquid Chromatography (UHPLC) system (Thermo Fisher Scientific in Waltham, MA, USA) was used to perform a non-targeted headspace metabolic profile analysis on all plant samples [19]. Individual phytochemical components were separated using an Accucore ™ RP-MS column (100 mm × 2.1 mm), with a particle size of 2.6 μm, connected to an Orbitrap Exploris 120 mass spectrometer (Thermo Fisher Scientific). The injection concentration of plant samples was 1mg/mL. The UHPLC system is equipped with a quaternary pump at a flow rate of 0.6 mL/min. Compound elution was achieved using the following elution solvents: (A) 0.1% volume/volume (v/v) formic acid dissolved in ultrapure water; and (B) acetonitrile (MeCN) containing 0.1% v/v formic acid. The Xcalibur acquisition programme (version 2.0) was employed to generate the following gradient profile: 5% solvent B for 5 minutes; Then, there was a gradual increase 5% to 30% solvent B over 5 minutes; This is followed by a 3-minute isocratic step at 30% solvent B; Next, there is a linear gradient from 30% to 90% solvent B over 4 minutes; An isocratic elution step at 90% solvent B was for 4 minutes was then used to clean the column of any nonpolar compounds.
The mass spectra of the eluted compounds were analysed using the Orbitrap Exploris 120 mass spectrometer in information-dependent acquisition (IDA) mode. The Orbitrap system employed electrospray ionisation (ESI) in negative ionisation mode, utilising a vaporiser temperature of 350 °C, sheath gas pressure of 60 psi, auxiliary gas pressure of 15 psi, sweep gas pressure of 2 psi, and a spray voltage ranging from 2.5 kV to 5 kV. Data were examined via Compound DiscovererTM 3.3. Background absorbance (determined by the blank file) was subtracted from each test chromatogram to make a list of identified extract compoounds. Utilising Mz Cloud, ChemSpider, Predicted Compositions, and MassList, Compound Discoverer correlated mass signals with established compounds. Compound Discoverer 3.3 generated Excel files for each extract. The files were subsequently examined to identify potential speculative compounds. The relative abundance of the identified compounds was determined as a percentage of the total area of the chromatogram.
2.9. Statistical Analyses
Three independent experiments were conducted to generate the ZOI results to generate means ± standard error of the mean (± SEM). Using one-way analysis of variance (ANOVA), the statistical significance between the treatment and negative control groups was investigated; p-values <0.001 were considered statistically extremely significant, and p-values <0.01 were considered statistically very highly significant.
3. Results
3.1. Antibacterial Assays
The extract concentrations for the two plants were comparable, with the highest yields achieved using methanol as a solvent. In contrast, ethyl acetate extraction produced significantly lower yields for both plant species. The extract yields of T. bellirica with water, methanol, and ethyl acetate were 50.2, 48.3, and 3.6 mg/mL respectively. In contrast, T. chebula yielded 35.6, 48.3, and 4.9 mg/mL for the same solvents.
The extracts and reference antibiotics were the subjected to agar disc diffusion assays (Figure 1) and broth microdilution assays (Table 1). These assays revealed that S. flexneri, S. sonnei, and S. typhimurium were resistant to penicillin G, oxacillin, amoxycillin and erythromycin. The extracts of both plants demonstrated similar results against B. cereus in agar disc diffusion assays, exhibiting ZOIs ranging from 9 to 13 mm. However, the T. bellirica ethyl acetate extract was inactive on agar against that bacterium. The aqueous, methanolic, and ethyl acetate extracts of T. bellirica produced MIC values of 392, 94, and 450 µg/mL, respectively against B. cereus. In contrast, MIC values of 556, 377, and 306 µg/mL were determined for the aqueous, methanol, and ethyl acetate T. chebula extracts against B. cereus respectively.
All T. bellirica (except for ethyl acetate) and T. chebula extracts exhibited substantial antibacterial activity against S. flexneri in the agar diffusion assays, with ZOIs ranging from 10 to 13 mm. This antibacterial activity is promising and comparable to the reference antibiotics such as polymyxin B. In broth microdilution assays, all extracts from both plant species demonstrated significant antibacterial activity against S. flexneri, with MIC values between 278 and 450 µg/mL in the broth microdilution assays.
In the disc diffusion assays, only the aqueous and methanol extracts of both plants demonstrated antibacterial activity against S. sonnei, with ZOIs of 8-9 mm. Aqueous and methanol extracts of T. bellirica demonstrated antibacterial activity against S. sonnei, with MIC values of 1569 and 755 µg/mL, respectively. In contrast, all T. chebula extracts exhibited low to moderate antibacterial activity against S. sonnei in broth dilution assays, with MIC values ranging from 1225 to 2225 µg/mL.
The aqueous and methanolic extracts of T. bellirica and T. chebula displayed antibacterial activity against S. typhimurium in the disc diffusion assay, with ZOIs ranging from 7.5 to 11.5 mm. Only the methanolic extract of T. bellirica exhibited a statistically significant ZOI (11.5 mm), comparable to polymyxin B, gentamicin, and Augmentin. The broth microdilution assays revealed low antibacterial activities of T. bellirica (aqueous) and T. chebula (aqueous and methanol), with MIC values ranging from 2225 to 3138 µg/mL. However, the methanolic extract of T. bellirica exhibited noteworthy antibacterial activity against S. typhimurium, with MIC of 755 µg/mL.
3.2. Combination Assays: Sum of Fractional Inhibitory Concentration (ƩFIC) Determinations
The antibiotics could be combined with the T. bellirica and T. chebula extracts in some cases to identify combinational effects between the two components against the bacterial strains tested in this study (Table 2). The extract and the antibiotic must both demonstrate efficacy against the bacterial species under investigation in order to be evaluated in combination. This is because MIC values for each component are required to calculate ƩFIC values. No synergistic interactions were detected in any of the combinations analysed. Thirty-five combinations resulted in additive effects, whilst seventy-nine combinations exhibited noninteraction. Notably, seventeen combinations exhibited antagonistic effects, particularly bin combinations containing polymyxin B as the antibiotic component.
3.3. Compound Identification by LC-MS Metabolomics Profiling
LC-MS profiling analysis was utilised to examine the metabolomic profiles of all extracts, emphasising the flavonoid, tannin, and terpenoid compounds. The majority of the extract compounds eluted during the 30% and 90% acetonitrile gradient elution phase of the chromatogram [22], indicating that most extract components exhibited relatively high polarity [23]. In contract, the organic acid and amine components exhibit an enhanced tendency to elute in polar environments, resulting in their early appearance in the chromatogram. The lipophilic substances and hydrocarbons, which demonstrate a greater affinity for the non-polar stationary phase, eluted later as the gradient continues. To construct a prospective inventory of all the discovered phytochemicals, only compounds that were fully matched in any of the databases (and not present in the blank controls) were selected [22]. In this investigation, we have concentrated on the phenolic acid, tannin, flavonoid, and terpenoid compounds (Table 3). These compounds are important because of their various biological functions. They have antioxidant, anti-inflammatory, and antibacterial characteristics, making them useful in health, medicine, and nutrition. Their activities in plant defence also provide natural sources for drug research and therapeutic uses, which are beneficial to human health and disease prevention.
% Relative Abundance is the ratio of a single compound's signal intensity to the total signal intensity of all detected compounds in a chromatogram. The percentage represents the compound's contribution to the overall chromatographic profile. However, the percentage relative abundance does not represent the actual quantity of the compound in the sample; rather, it shows its proportion based on signal strength, which might vary depending on the detector's response to different compounds. Compounds identified in both plants extracts are in black; Compounds identified only in Terminalia bellirica (TB) extracts are in red; Compounds identified only in Terminalia chebula (TCh) extracts are in green; Compounds less than 0.01% of the total area were considered as trace amounts and denoted as T.
3.4. Toxicity Quantification
Plant extract toxicity was assessed by Artemia franciscana Kellogg nauplii lethality assays (ALA). The extracts were categorised as toxic if they produced LC50 values ≤1000 µg/mL after a 24-hour exposure period [18]. The findings for all of the extracts were similar to those of the negative control (artificial seawater) since they failed to induce ≥50% mortality at 1000 µg/mL and were thus deemed nontoxic.
4. Discussion
Our study showed that significant growth inhibition of four different gastrointestinal bacterial pathogens was elicited by the aqueous and methanolic extracts of T. bellirica and T. chebula fruit extracts. The methanolic extracts exhibited the highest antibacterial efficacy in both the disc diffusion and MIC assays against all pathogens. The ethyl acetate T. bellirica extracts exhibited noteworthy antibacterial activity against B. cereus and S. flexneri, although only in the liquid microdilution assays. Notably, the T. chebula ethyl acetate extracts exhibited good antibacterial activities against B. cereus and S. flexneri in both the disc diffusion and liquid microdilution assays. Furthermore, the T. chebula ethyl acetate extracts also exhibited moderate antibacterial activities against S. sonnei in liquid microdilution assays, but not in the agar diffusion assays. The variations in net yields and the concentrations of phytochemicals extracted by the different solvents may explain these differences in potency. The polarity of methanol and water is far greater than ethyl acetate, tus permitting the extraction of greater quantities of phytochemicals that are within the mid-to high polarity ranges. In contrast, ethyl acetate only extracts compounds with mid to low polarity, resulting in substantially less complexity and lower compound yields [24].
The variations in phytochemical content among the extracts may contribute to the distinct antibacterial growth inhibition effects observed in the disc diffusion and liquid dilution studies. Phytochemicals with lower polarity or larger size exhibit reduced diffusion rates through solid agar, leading to a diminished apparent antibacterial effectiveness in disc diffusion assays [25]. Previous studies have shown that the depth and uniformity of agar in petri dishes can influence the size of zones of inhibition (ZOIs) in agar diffusion experiments [26]. The solubility of these phytochemicals in broth is influenced by their polarity [27], which affects their dissolution and may result in inaccurate MIC values.
Notably, all T. bellirica and T. chebula extracts exhibited antibacterial activity against B. cereus and S. flexneri, with MIC values ranging from 94 to 556 µg/mL. In contrast, only the T. bellirica methanolic extract showed noteworthy antibacterial activity against S. sonnei and S. typhimurium, with an MIC value of 755 µg/mL. The B. cereus, S. sonnei, S. flexneri, and S. typhimurium bacterial strains exhibited substantial susceptibility to the standard antibiotics. Subsequent research must broaden the scope of bacterial strains tested to assess the efficacy of plant extracts on additional antibiotic-resistant strains. This is essential for tackling bacterial antibiotic resistance and developing novel treatment alternatives. Our research findings shown that S. flexneri, S. sonnei, and S. typhimurium were resistant to penicillin G, oxacillin, amoxycillin and erythromycin. These bacteria are significant causal agents of gastrointestinal infections. Shigella species induce shigellosis, which is characterised by diarrhoea and fever. Salmonella typhimurium causes foodborne gastroenteritis, and is associated with contaminated poultry and eggs, resulting in considerable global public health concerns.
The effectiveness of the extracts against bacterial pathogens may be ascribed to their levels of specific phytochemicals, including flavonoids, tannins, and terpenoids. These compounds may exert their antibacterial mechanisms by disrupting bacterial cell walls and membranes, altering internal pH levels, inhibiting enzymatic activity, and interfering with DNA replication [12]. Furthermore, the combined effect of these compounds improves their overall antibacterial effectiveness compared to the individual components in isolation. Furthermore, plant extracts may inhibit toxin production and quorum sensing system, thereby diminishing the virulence of the bacterial pathogens and its capacity to form biofilms [28,29,30,31,32].
All reference antibiotics (except polymyxin B) showed varying levels of antibacterial activity against B. cereus. Polymyxin B was ineffective against B. cereus as it specifically targets Gram-negative bacteria by binding to lipopolysaccharides in their outer membrane [33], which are absent in Gram-positive bacteria such as B. cereus. This bacterium possesses a thick peptidoglycan layer and lacks the requisite binding sites [34], which renders polymyxin B ineffective and results in intrinsic resistance. The reference antibiotics tetracycline, chloramphenicol, ciprofloxacin, polymyxin B and gentamicin exhibited varying levels of antibacterial activity against S. flexneri, S. sonnei and S. typhimurium.
The ineffectiveness of penicillin G, erythromycin, oxacillin, amoxycillin, and vancomycin against S. flexneri, S. sonnei, and S. typhimurium can be attributed to multiple factors. These bacteria are Gram-negative and have an outer membrane that serves as a barrier [30,32], inhibiting the penetration of various antibiotics, including penicillin G and vancomycin, which primarily target Gram-positive bacteria. Shigella and Salmonella species frequently produce β-lactamase enzymes [35,36], which degrade β-lactam antibiotics including penicillin G, oxacillin and amoxycillin, thereby rendering those antibiotics ineffective. The presence of efflux pumps and genetic mutations in these bacterial pathogens may further diminish the effectiveness of conventional antibiotics [37,38]. Additionally, vancomycin targets the peptidoglycan layer [39], which is not accessible in Gram-negative bacteria, thereby restricting its efficacy. The presence of these combined resistance mechanisms requires the implementation of alternative treatments capable of surmounting these challenges.
Our study also investigated into the application of T. bellirica and T. chebula extracts in conjunction with conventional antibiotics. Combinational therapies have considerable potential for creating novel antibiotic chemotherapies as many bacteria have evolved resistance to traditional antibiotics. Plant-derived compounds may provide novel strategies to inhibit or block these resistance mechanisms [40]. By combining antibiotics with plant extracts, we hoped to improve their efficiency and to possibly overcome the bacterial resistance mechanisms. Augmentin®, a combination of amoxycillin and clavulanic acid, is a well-known example of how combining drugs can improve treatment outcomes [41]. Clavulanic acid inhibits β-lactamase enzymes by irreversibly attaching to their active site, thereby preventing antibiotic degradation. This allows the amoxycillin component of the Augmentin® combination therapy to function with greater potency, even in bacteria otherwise resistant to its effects.
Our investigation demonstrated that penicillin G, oxacillin, and amoxycillin exhibited additive effects against B. cereus when combined with any of the T. chebula extracts, or the aqueous T. bellirica extract. This may be due to the presence of phytochemicals capable of inhibiting β-lactamase enzyme(s). [42]. [43] Phytochemicals in the extracts may interact with β-lactamase enzymes via a mechanism similar to that of clavulanic acid, although this requires confirmation. This interaction (irrespective of the mechanism) aids in safeguarding antibiotics from degradation by bacterial enzymes and enhances their efficacy against bacterial pathogens. Similarly, aqueous T. bellirica extracts showed additive effects against B. cereus in combination with vancomycin. Vancomycin inhibits peptidoglycan synthesis, thereby targeting the bacterial cell wall in Gram-positive bacteria such as B. cereus. The combination of plant extracts with vancomycin may enhance its efficacy. The flavonoids, tannins, and phenolic acids may directly target bacterial structures, disrupt membrane permeability, or inhibit bacterial enzymes associated with resistance [43]. Plant extracts may therefore provide treatment alternatives and could provide a safer and less harmful option for addressing antibiotic resistance [40]. Adverse effects can also be avoided when natural compounds derived from plants are used, whilst helping to minimise the development of AMR as compared to conventional antibiotics [12]. Furthermore, the potential for additional antibiotic resistance development may be diminished by the diverse range of phytochemicals found in plant extracts, which may function by targeting different aspects of bacterial cell growth and survival.
Interestingly, the combination of tetracycline with the Terminalia spp. extracts produced additive interactions against B. cereus, S. flexneri, S. sonnei, and S. typhimurium. The aqueous T. bellirica extract. The methanolic T. chebula extract demonstrated additive interactions with tetracycline against B. cereus, whilst the ethyl acetate extracts produced from both plant species had similar effects against S. flexneri. Additionally, the aqueous and methanolic extracts produced from both plants demonstrated additive interactions when combined with tetracycline against S. sonnei and S. typhimurium. Tetracycline resistance is primarily associated with tetracycline-specific efflux pumps [44]. Thus, the observed additive effect may indicate that T. bellirica and T. chebula extracts may have impaired the activity of these efflux pumps. Inhibition of efflux would facilitate prolonged retention of tetracycline within cells, resulting in higher intracellular antibiotic concentrations, thereby enhancing the efficacy of tetracycline. Alterations in ribosomal structure may also contribute to tetracycline resistance. However, this pathway is substantially less prevalent [44]. The present research indicates that some phytochemicals present in the T. bellirica and T. chebula extracts may have broad-spectrum antibacterial activity against multiple bacterial strains and/or, that the extracts contain antibiotic potentiating agents.
The T. chebula extracts showed additive effects against B. cereus and S. sonnei when used in conjunction with chloramphenicol, whilst the aqueous and ethyl acetate T. bellirica extracts displayed additive antibacterial effects against S. sonnei and S. flexneri. Chloramphenicol binds to the 50S ribosomal subunit in the bacterial ribosome, specifically interacting with the 23S rRNA component [45]. This binding inhibits peptidyl transferase activity, an essential mechanism for peptide bond formation in protein synthesis. Chloramphenicol therefore inhibits bacterial protein synthesis by obstructing peptide bond formation [45], whilst the plant extract may affect various bacterial targets, including cell wall synthesis, membrane integrity, and/or metabolic processes [12,43]. The plant extracts may also influence additional mechanisms related to bacterial resistance, including the disruption of efflux pumps and modification of cell wall structures [43], potentially increasing bacterial susceptibility to chloramphenicol. Additional studies are necessary to identify which mechanism(s) is/are influenced by the Terminalia spp. extracts. Alternatively, the T. bellirica and T. chebula extracts may contain potentiating compounds that enhance the antibacterial effects of chloramphenicol. Similarly, the combination of aqueous and methanolic T. bellirica extracts with gentamicin produced additive effects against S. flexneri, S. sonnei, and S. typhimurium. Similarly, the aqueous and methanolic T. chebula extracts exhibited additive effects when combined with gentamicin against S. sonnei and B. cereus. The aqueous and methanolic Terminalia spp. extracts may include compounds that hinder bacterial resistance mechanisms, or alternatively, that interact with alternative targets within the bacteria, thereby enhancing gentamicin’s mechanism of action (disruption of protein synthesis) [46].
The T. bellirica ethyl acetate extract had antagonistic effects against B. cereus and S. sonnei when tested in conjunction with gentamicin. In contrast, the T. chebula ethyl acetate extract antagonised the effects of gentamicin against S. flexneri and S. sonnei. Compounds in the ethyl acetate extract may inhibit the uptake of gentamicin into bacterial cells, or alternatively may disrupt its binding to ribosomal subunits, thereby diminishing its effectiveness, although these possibilities remain to be tested.
Significantly, combinations containing polymyxin B and the T. bellirica or T. chebula extracts demonstrated considerable antagonistic effects against S. flexneri and S. sonnei. Alterations in broth pH levels may impact the antagonistic interaction of polymyxin B and the plant extracts. Polymyxin B disrupts bacterial cell membranes by binding to lipopolysaccharides. Notably, polymyxin B is pH sensitive, with significantly lower efficacy in acidic or alkaline environments [47]. The incorporation of plant extracts may alter the pH of the broth, thereby impacting the efficacy of both polymyxin B and the phytochemicals, which could lead to a reduction in total antibacterial activity, although this requires verification through additional investigation. Alterations in pH may also influence bacterial physiology, thereby diminishing bacterial susceptibility to the combined effects of the polymyxin B and the plant extracts. Alternatively, the antagonistic interactions between the extracts and polymyxin B may be attributed to the binding of bioactive phytochemicals to polymyxin B, which may hinder its absorption into the target bacterial cells [48].
LC-MS metabolomics analysis of the T. bellirica and T. chebula fruit extracts revealed the presence of flavonoids, tannins, terpenoids, and phenolic acid compounds (Table 3). The full lists of phytochemicals found detected in the individual plant extracts are provided in the supplementary files of our previous study [22]. Some important phytochemicals identified in both plant extracts include methyl gallate (Figure 2A), propyl gallate (Figure 2B), gallic acid (Figure 2C), ellagic acid (Figure 2D), hamamelitannin (Figure 2E), pyrogallol (Figure 2F), quercetin (Figure 2G), 6-galloylglucose (Figure 2H), gallic acid 3-O-(6-galloylglucoside) (Figure 2I), 1,2,6-trigalloyl-β-D-glucopyranose (Figure 2J), 1,3,6-tri-O-galloyl-β-D-glucose (Figure 2K), 1,6-bis-O-(3,4,5-trihydroxybenzoyl) hexopyranose (Figure 2L), chebulic acid (Figure M), and chebuloside II (Figure N). Notably, gallic acid, ellagic acid, chebulic acid, chebuloside II, methyl gallate, propyl gallate, ethyl gallate, phloroglucinol, pyrogallol, quercetin, kaempferol, and various other compounds detected in our study have also previously been reported in T. bellirica and T. chebula fruit extracts [49,50,51]. In the prior studies, qualitative GC-MS headspace analysis was performed on T. bellirica and T. chebula aqueous, methanolic and ethyl acetate fruit extracts. The previous phytochemical analyses also putatively identified several significant volatile terpenoids, including endo-borneol, linalool, terpinene-4-ol, methoxy citronellal, pinocarveol, eucalyptol, carvone, camphor, L-fenchone, hyscylene, patchoulane, p-cumic aldehyde, and phenylbutanal [18].
Interestingly, combinations of methyl gallate (30 µg/mL) and marbofloxacin (0.015 µg/mL) inhibit adhesion and invasion of S. typhimurium in macrophage cell lines (RAW 264.7 cells) by 70 % and 67%, respectively [52]. Moreover, this combination downregulated the sdiA, srgE, and rck quorum sensing genes of S. typhimurium by 53%, 62% and 22 %, respectively. Methyl gallate has been isolated and identified from the fruit extracts of T. chebula and has been reported to have antibacterial activity against S. flexneri and S. sonnei, with MIC values 128 to 256 µg/mL respectively [53]. That study also reported bacterial membrane-damaging activity for methyl gallate, which was determined by membrane perturbation assays and transmission electron microscopy (TEM). Furthermore, previous studies have also reported that high levels of tetracycline and methyl gallate are accumulated in HeLa cells infected with Shigella dysenteriae [53]. This suggests that methyl gallate may be inhibiting tetracycline efflux pumps in S. dysenteriae. However, the combinational interactions of reference antibiotics and methyl gallate was not investigated in that study. Additionally, another study demonstrated that methyl gallate has weak β-lactamase inhibition activity compared to that of clavulanic acid [54]. This suggest that methyl gallate-related antibacterial activity result from cell membrane disruption, inhibition of DNA synthesis, or targeting metabolic pathways. Indeed, methyl gallate has been reported to have substantial antibacterial activity against clinical isolates of S. typhimurium, with MIC values between 3.9 and 15.6 µg/mL [55]. That study also demonstrated that methyl gallate, when used in conjunction with ATPase inhibitors such as sodium azide (NaN3) and dicyclohexylcarbodiimide (DCCD), reduced S. typhimurium by approximately 40%. Furthermore, oral administration of methyl gallate significantly reduces S. typhimurium infections in BALB/c mice, with no observed clinical symptoms [55]. Another study also reported that methyl gallate (125 µg/mL) reduced the MIC of nalidixic acid from 500 µg/mL to 31.25 µg/mL against nalidixic acid resistant S. typhimurium pathogenic strain, indicating that methyl gallate may negate the bacteria’s resistance mechanisms towards nalidixic acid [56].
Kang et al. (2018) demonstrated the notable inhibitory effect of gallic acid on S. flexneri biofilm formation through the regulation of the mdoH gene and the OpgH protein [57]. Gallic acid exhibits weak antibacterial activity against S. flexneri, with a MIC of 2000 µg/mL. However, that study did not examine the combination of gallic acid with conventional antibiotics. Another study reported antibacterial activities of gallic acid and hamamelitannin against multidrug resistant strains of S. typhimurium, with MIC values of 256 µg/mL and 512–1024 µg/mL respectively [58]. Fractional inhibitory combinational experiments demonstrated the additive effects of gallic acid with ampicillin, ceftiofur, and thiamphenicol, whilst hamamelitannin showed additive interactions with ceftiofur, cefotaxime, and marbofloxacin against S. typhimurium (ATCC14028). Additionally, the vitality of S. typhimurium biofilm formation was entirely suppressed by the combination of gallic acid and ceftiofur. Another study reported good antibacterial activity for ellagic acid against a panel of Salmonella species, including S. paratyphi, S. choleraesuis subsp., and S. enteridis, with MIC values of 20, 10, and 15 µg/mL respectively [59]. A further study reported antibacterial efficacy for 3,3,4-tri-O-methylellagic acid and 3,4-di-O-methylellagic acid against S. flexneri, each with MICs of 5 µg/mL [60]. Both compounds also demonstrated antibacterial activity against B. cereus, with MIC of 39 and 19 µg/mL, respectively. Notably, studies are deficient regarding the combinational interactions between ellagic acid and conventional antibiotics, and substantially more work is required.
The synergistic effect of pyrogallol in conjunction with marbofloxacin against S. typhimurium has also been reported [61]. Pyrogallol exhibited MIC values of 128 µg/mL for all S. typhimurium isolates tested. The gentamicin protection assay demonstrated that pyrogallol (30 μg/mL), both independently and in conjunction with sub-MIC concentrations of marbofloxacin (0.015 μg/mL for susceptible strains and 0.25 μg/mL for marbofloxacin-resistant strains), inhibited 73% and 76% of the invading bacteria in Caco-2 cells, respectively [61]. Fluoroquinolone accumulation assays indicated that pyrogallol (at 30 μg/mL and 100 μg/mL concentrations) enhanced ciprofloxacin accumulation within S. typhimurium by 15% and 35%, respectively [61].
Hirudkar et al. (2020) reported substantial antidiarrhoeal activity for quercetin (50 mg/kg) in rats induced with diarrhoea by S. flexneri [62]. Furthermore, the authors also reported substantially decreased expression of proinflammatory cytokines (interleukin-6 and tumour necrosis factor-α), and a reduction in the density of S. flexneri in the faeces of the animals studied. Furthermore, docking simulation studies revealed that quercetin effectively inhibits the protease activity of the S. flexneri protein (SepA), which is responsible for damaging epithelial tissue during infection. A further investigation demonstrated the antibacterial efficacy of quercetin against S. typhimurium (MIC 250 µg/mL) [63]. Moreover, field emission scanning electron microscopy revealed that quercetin inhibited the biofilm formation of S. typhimurium and diminished the expression of virulence (rpoS, avrA) and quorum-sensing (luxS) genes. However, neither study explored the combinatorial interactions of quercetin and conventional antibiotics in relation to the bacterial pathogens examined.
Our study identified several galloyl glucose derivatives within the T. bellirica and T. chebula extracts, including 6-galloylglucose, gallic acid 3-O-(6-galloylglucoside), 1,2,6-trigalloyl-β-D-glucopyranose, 1,3,6-tri-O-galloyl-β-D-glucose, and 1,6-bis-O-(3,4,5-trihydroxybenzoyl) hexopyranose. The presence of these compounds in the plant extracts may contribute to the noteworthy antibacterial activity of these extracts against the bacterial pathogens examined herein. A previous study demonstrated the antibacterial efficacy of 1,2,3,4,6-penta-O-galloyl-β-D-glucose against B. cereus and S. typhimurium, reporting MIC values of 32 µg/mL and 128 µg/mL respectively [64]. However, studies examining the combinatorial interactions of galloyl glucose derivatives with conventional antibiotics are largely lacking.
Our analysis did not reveal any synergistic enhancement of antibacterial activity between either the T. bellirica and T. chebula extracts and the selected antibiotics against the bacterial pathogens studied herein. Nevertheless, conventional antibiotics, including penicillin G, tetracycline, chloramphenicol, oxacillin, amoxycillin, gentamicin, and vancomycin, have been reported to exhibit additive interactions with plant extracts against these bacterial pathogens. The phytochemicals in T. bellirica and T. chebula extracts may exhibit inhibitory properties against β-lactamase and/or efflux pumps. Additional research is necessary to examine the impact of these extracts on the resistance mechanisms.
Artemia nauplii bioassays indicated that all T. bellirica and T. chebula extracts are nontoxic, supporting their safety as for use as antibacterial agents. However, additional research with a panel of mammalian cell lines should be performed in future studies to comprehensively evaluate whether these extracts are safe for medicinal use. Taken together, our findings suggest that T. bellirica and T. chebula fruit extracts could be an important source of antimicrobial compounds for future research into antibiotic development in the fight against bacterial gastrointestinal infections.
5. Conclusions
Traditionally, T. bellirica and T. chebula fruit extracts have shown promising results in the treatment of gastrointestinal infections because to their high concentration of bioactive flavonoids, tannins, polyphenols and terpenoids. Many of these phytochemicals have antibacterial, anti-inflammatory, and antioxidant activities that can reduce pathogenic microorganisms and improve gut health. Plant-based antimicrobials provide a natural, sustainable alternative to conventional antibiotics, lowering the danger of antibiotic resistance. The findings of our study indicate that active extracts from T. bellirica and T. chebula can effectively inhibit the growth of bacterial pathogens that cause gastrointestinal infections. This indicates that the extracts may have distinct antibacterial mechanisms that necessitate further investigation. Furthermore, all extracts improved the efficacy of several conventional antibiotics, particularly β-lactam antibiotics and tetracycline. It is possible that antibiotic reactivation may be occurring even in bacterial strains that are typically resistant to their effects. The method of potentiation remains to be determined, although specific extract components may inhibit bacterial extended spectrum β-lactamase enzymes and efflux pumps, resulting in elevated antibiotic concentrations within the cells. Further research is necessary to validate these mechanisms and to explore additional possible enhancement pathways. Several phytochemicals found in the extracts may contribute to these activities, indicating that they could be promising targets for the development of novel antibacterial agents, although additional research is required. An assessment of the activities of the pure phytochemicals and their abilities to enhance conventional antibiotic activities should be the topic of future study.
Author Contributions
Conceptualization, investigation, resources, M.J.C. and I.E.C.; methodology, G.T. and M.J.C.; validation, formal analysis, data curation, G.T.; writing—original draft preparation, G.T.; writing—review and editing, supervision, M.J.C. and I.E.C.; project administration and funding acquisition, I.E.C. and M.J.C. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support for this work was provided by a PhD Scholarship, Griffith University, Australia.
Data Availability Statement
Data are either presented within the manuscript or are available from the corresponding author upon reasonable request.
Acknowledgments
The authors wish to thank Muhammad Jawad Zai for assistance with compound identification.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- World Health Organization. Diarrhoeal disease. 2024. Available online: https://www.who.int/news-room/fact-sheets/detail/diarrhoeal-disease (accessed on 31 July 2024).
- Troeger, C.; Blacker, B.F.; Khalil, I.A.; Rao, P.C.; Cao, S.; Zimsen, S.R.; Albertson, S.B.; Stanaway, J.D.; Deshpande, A.; Abebe, Z.; et al. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of diarrhoea in 195 countries: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1211–1228. [Google Scholar] [CrossRef] [PubMed]
- Raso, M.M.; Arato, V.; Gasperini, G.; Micoli, F. Toward a Shigella vaccine: opportunities and challenges to fight an antimicrobial-resistant pathogen. Int. J. Mol. Sci. 2023, 24, 4649. [Google Scholar] [CrossRef] [PubMed]
- Wotzka, S.Y.; Nguyen, B.D.; Hardt, W.D. Salmonella Typhimurium diarrhea reveals basic principles of enteropathogen infection and disease-promoted DNA exchange. Cell Host Microbe 2017, 21, 443–454. [Google Scholar] [CrossRef] [PubMed]
- El-Arabi, T.F.; Griffiths, M.W. Bacillus cereus. Foodborne Infections and Intoxications 2021, 431–437. [Google Scholar] [CrossRef]
- Puzari, M.; Sharma, M.; Chetia, P. Emergence of antibiotic resistant Shigella species: A matter of concern. J. Infect. Public Health 2018, 11, 451–454. [Google Scholar] [CrossRef]
- Chung The, H.; Bodhidatta, L.; Pham, D.T.; Mason, C.J.; Ha Thanh, T.; Voong Vinh, P.; Turner, P.; Hem, S.; Dance, D.A.B.; Newton, P.N.; et al. Evolutionary histories and antimicrobial resistance in Shigella flexneri and Shigella sonnei in Southeast Asia. Commun. Biol. 2021, 4, 1–12. [Google Scholar] [CrossRef]
- Wang, X.; Biswas, S.; Paudyal, N.; Pan, H.; Li, X.; Fang, W.; Yue, M. Antibiotic resistance in Salmonella Typhimurium isolates recovered from the food chain through national antimicrobial resistance monitoring system between 1996 and 2016. Front. Microbiol. 2019, 10, 450730. [Google Scholar] [CrossRef]
- Fiedler, G.; Schneider, C.; Igbinosa, E.O.; Kabisch, J.; Brinks, E.; Becker, B.; Stoll, D.A.; Cho, G.S.; Huch, M.; Franz, C.M.A.P. Antibiotics resistance and toxin profiles of Bacillus cereus-group isolates from fresh vegetables from German retail markets. BMC Microbiol. 2019, 19, 1–13. [Google Scholar] [CrossRef]
- Chon, J.W.; Kim, J.H.; Lee, S.J.; Hyeon, J.Y.; Seo, K.H. Toxin profile, antibiotic resistance, and phenotypic and molecular characterization of Bacillus cereus in Sunsik. Food Microbiol. 2012, 32, 217–222. [Google Scholar] [CrossRef]
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.P.; Nautiyal, A.R. Antimicrobial resistance and the alternative resources with special emphasis on plant-based antimicrobials—A review. Plants 2017, 6, 16. [Google Scholar] [CrossRef]
- Tiwana, G.; Cock, I.E.; Cheesman, M.J. A Review of Ayurvedic principles and the use of Ayurvedic plants to control diarrhoea and gastrointestinal infections. Pharmacogn. Commun. 2023, 13, 152–162. [Google Scholar] [CrossRef]
- Rawat, P.; Singh, P.K.; Kumar, V. Evidence based traditional anti-diarrheal medicinal plants and their phytocompounds. Biomed. Pharmacother. 2017, 96, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- Srikumar, R.; Jeya Parthasarathy, N.; Shankar, E.M.; Manikandan, S.; Vijayakumar, R.; Thangaraj, R.; Vijayananth, K.; Sheeladevi, R.; Rao, U.A. Evaluation of the growth inhibitory activities of Triphala against common bacterial isolates from HIV infected patients. Phytother. Res. 2007, 21, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Mani, P.; Devi, P.N.; Kaleeswari, S.; Poonkothai, M. Antimicrobial activity and phytochemical analysis of fruit extracts of Terminalia Bellerica. Int. J. Pharm. Pharm. Sci. 2014, 43, 364. [Google Scholar]
- Madani, A.; Jain, S.K. Anti-Salmonella activity of Terminalia belerica: In vitro and in vivo studies. Indian J. Exp. Biol. 2008, 46, 817–821. [Google Scholar]
- Kong, Q.; Shang, Z.; Liu, Y.; Fakhar-e-Alam Kulyar, M.; Suo-lang, S.; Xu, Y.; Tan, Z.; Li, J.; Liu, S. Preventive effect of Terminalia bellirica (Gaertn.) Roxb. extract on mice infected with Salmonella Typhimurium. Front. Cell. Infect. Microbiol. 2022, 12. [Google Scholar] [CrossRef]
- Tiwana, G.; Cock, I.E.; White, A.; Cheesman, M.J. Use of specific combinations of the triphala plant component extracts to potentiate the inhibition of gastrointestinal bacterial growth. J. Ethnopharmacol. 2020, 260, 112937. [Google Scholar] [CrossRef]
- Tiwana, G.; Cock, I.E.; Cheesman, M.J. Phyllanthus niruri Linn.: Antibacterial activity, phytochemistry, and enhanced antibiotic combinatorial strategies. Antibiotics 2024, 13, 654. [Google Scholar] [CrossRef]
- Doern, C.D. When does 2 plus 2 equal 5? A review of antimicrobial synergy testing. J. Clin. Microbiol. 2014, 52, 4124–4128. [Google Scholar] [CrossRef]
- Ruebhart, D.R.; Wickramasinghe, W.; Cock, I.E. Protective efficacy of the antioxidants vitamin E and Trolox against Microcystis aeruginosa and microcystin-LR in Artemia franciscana Nauplii. J. Toxicol. Environ. Health A 2009, 72, 1567–1575. [Google Scholar] [CrossRef]
- Tiwana, G.; Cock, I.E.; Cheesman, M.J. Combinations of Terminalia bellirica (Gaertn.) Roxb. and Terminalia chebula Retz. extracts with selected antibiotics against antibiotic-resistant bacteria: Bioactivity and phytochemistry. Antibiotics 2024, 13(10), 994. [Google Scholar] [CrossRef] [PubMed]
- Cajka, T.; Hricko, J.; Rudl Kulhava, L.; Paucova, M.; Novakova, M.; Kuda, O. Optimization of mobile phase modifiers for fast LC-MS-based untargeted metabolomics and lipidomics. Int. J. Mol. Sci. 2023, 24, 31987. [Google Scholar] [CrossRef] [PubMed]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.G.; Lightfoot, D.A. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Bonev, B.; Hooper, J.; Parisot, J. Principles of assessing bacterial susceptibility to antibiotics using the agar diffusion method. J. Antimicrob. Chemother. 2008, 61, 1295–1301. [Google Scholar] [CrossRef]
- Flanagan, J.N.; Steck, T.R. The relationship between agar thickness and antimicrobial susceptibility testing. Indian J. Microbiol. 2017, 57, 503–506. [Google Scholar] [CrossRef]
- Jain, S.; Patel, N.; Lin, S. Solubility and dissolution enhancement strategies: Current understanding and recent trends. Drug Dev. Ind. Pharm. 2015, 41, 875–887. [Google Scholar] [CrossRef]
- Rocha, J.; Flores, V.; Cabrera, R.; Soto-Guzmán, A.; Granados, G.; Juaristi, E.; Guarneros, G.; De La Torre, M. Evolution and some functions of the NprR-NprRB quorum-sensing system in the Bacillus cereus group. Appl. Microbiol. Biotechnol. 2012, 94, 1069–1078. [Google Scholar] [CrossRef]
- Ceuppens, S.; Rajkovic, A.; Heyndrickx, M.; Tsilia, V.; Van De Wiele, T.; Boon, N.; Uyttendaele, M. Regulation of toxin production by Bacillus cereus and its food safety implications. Crit. Rev. Microbiol. 2011, 37, 188–213. [Google Scholar] [CrossRef]
- Stårsta, M.; Hammarlof, D.L.; Waneskog, M.; Schlegel, S.; Xu, F.; Gynnå, A.H.; Borg, M.; Herschend, S.; Koskiniemi, S. RHS-elements function as type II toxin-antitoxin modules that regulate intra-macrophage replication of Salmonella Typhimurium. PLoS Genet. 2020, 16, e1008607. [Google Scholar] [CrossRef]
- Pakbin, B.; Amani, Z.; Allahyari, S.; Mousavi, S.; Mahmoudi, R.; Brück, W.M.; Peymani, A. Genetic diversity and antibiotic resistance of Shigella spp. isolates from food products. Food Sci. Nutr. 2021, 9, 6362–6371. [Google Scholar] [CrossRef]
- Puzari, M.; Sharma, M.; Chetia, P. Emergence of antibiotic resistant Shigella species: A matter of concern. J. Infect. Public Health 2018, 11, 451–454. [Google Scholar] [CrossRef] [PubMed]
- Slingerland, C.J.; Kotsogianni, I.; Wesseling, C.M.J.; Martin, N.I. Polymyxin stereochemistry and its role in antibacterial activity and outer membrane disruption. ACS Infect. Dis. 2022, 8, 2396–2404. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Koehler, T.M.; Lereclus, D. The Bacillus cereus group: Bacillus species with pathogenic potential. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Berbers, B.; Vanneste, K.; Roosens, N.H.C.J.; Marchal, K.; Ceyssens, P.J.; De Keersmaecker, S.C.J. Using a combination of short-and long-read sequencing to investigate the diversity in plasmid and chromosomally encoded extended-spectrum beta-lactamases (ESBLs) in clinical Shigella and Salmonella isolates in Belgium. Microb. Genom 2023, 9, 000925. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. Epidemiology of β-lactamase-producing pathogens. Clin. Microbiol. Rev. 2020, 33. [Google Scholar] [CrossRef]
- Auda, I.G.; Ali Salman, I.M.; Odah, J.G. Efflux pumps of gram-negative bacteria in brief. Gene Rep. 2020, 20, 100666. [Google Scholar] [CrossRef]
- Alenazy, R. Antibiotic Resistance in Salmonella: Targeting multidrug resistance by understanding efflux pumps, regulators, and the inhibitors. J. King Saud Univ. Sci. 2022, 34, 102275. [Google Scholar] [CrossRef]
- Stogios, P.J.; Savchenko, A. Molecular mechanisms of vancomycin resistance. Protein Sci. 2020, 29, 654–669. [Google Scholar] [CrossRef]
- Cheesman, M.; Ilanko, A.; Blonk, B.; Cock, I.E. Developing new antimicrobial therapies: Are synergistic combinations of plant extracts/compounds with Conventional antibiotics the solution? Pharmacogn. Rev. 2017, 11, 57. [Google Scholar] [CrossRef]
- Huttner, A.; Bielicki, J.; Clements, M.N.; Frimodt-Møller, N.; Muller, A.E.; Paccaud, J.P.; Mouton, J.W. Oral amoxicillin and amoxicillin–clavulanic Acid: Properties, indications, and usage. Clin. Microbiol. Infect. 2020, 26, 871–879. [Google Scholar] [CrossRef]
- Oelschlaeger, P. β-Lactamases: Sequence, structure, function, and inhibition. Biomolecules 2021, 11, 986. [Google Scholar] [CrossRef] [PubMed]
- Khameneh, B.; Eskin, N.A.M.; Iranshahy, M.; Fazly Bazzaz, B.S. Phytochemicals: A promising weapon in the arsenal against antibiotic-resistant bacteria. Antibiotics 2021, 10, 1044. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gupta, V.K.; Pathania, R. Efflux pump inhibitors for bacterial pathogens: From bench to bedside. Indian J. Med. Res. 2019, 149, 129. [Google Scholar] [CrossRef] [PubMed]
- Kehrenberg, C.; Schwarz, S.; Jacobsen, L.; Hansen, L.H.; Vester, B. A new mechanism for chloramphenicol, florfenicol and clindamycin resistance: Methylation of 23S ribosomal RNA at A2503. Mol. Microbiol. 2005, 57, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; He, J.; Bai, L.; Ruan, S.; Yang, T.; Luo, Y. Ribosome-targeting antibacterial agents: Advances, challenges, and opportunities. Med. Res. Rev. 2021, 41, 1855–1889. [Google Scholar] [CrossRef]
- Cai, X.; Javor, S.; Gan, B.H.; Köhler, T.; Reymond, J.L. The antibacterial activity of peptide dendrimers and polymyxin B increases sharply above pH 7.4. Chem. Commun. 2021, 57, 5654–5657. [Google Scholar] [CrossRef]
- Zavascki, A.P.; Goldani, L.Z.; Li, J.; Nation, R.L. Polymyxin B for the treatment of multidrug-resistant pathogens: A critical review. J. Antimicrob. Chemother. 2007, 60, 1206–1215. [Google Scholar] [CrossRef]
- Cock, I.E. The medicinal properties and phytochemistry of plants of the genus Terminalia (Combretaceae). Inflammopharmacology 2015, 23, 203–229. [Google Scholar] [CrossRef]
- Kumari, S.; Krishna, M.J.; Joshi, A.B.; Gurav, S.; Bhandarkar, A. V; Agarwal, A.; Mythili Krishna, C.J. A pharmacognostic, phytochemical and pharmacological review of Terminalia bellerica. J. Pharmacogn. Phytochem. 2017, 6, 368–376. [Google Scholar]
- Bag, A.; Bhattacharyya, S.K.; Chattopadhyay, R.R. The development of Terminalia chebula Retz. (Combretaceae) in clinical research. Asian Pac. J. Trop. Biomed. 2013, 3, 244. [Google Scholar] [CrossRef]
- Birhanu, B.T.; Park, N.H.; Lee, S.J.; Hossain, M.A.; Park, S.C. Inhibition of Salmonella Typhimurium adhesion, invasion, and intracellular survival via treatment with methyl gallate alone and in combination with marbofloxacin. Vet. Res. 2018, 49, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Acharyya, S.; Sarkar, P.; Saha, D.R.; Patra, A.; Ramamurthy, T.; Bag, P.K. Intracellular and membrane-damaging activities of methyl gallate isolated from Terminalia chebula against multidrug-resistant Shigella spp. J. Med. Microbiol. 2015, 64, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Jiamboonsri, P.; Eurtivong, C.; Wanwong, S. Assessing the potential of gallic acid and methyl gallate to enhance the efficacy of β-lactam antibiotics against methicillin-resistant Staphylococcus aureus by targeting β-lactamase: In silico and in vitro studies. Antibiotics 2023, 12, 1622. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Mun, S.H.; Chahar, H.S.; Bharaj, P.; Kang, O.H.; Kim, S.G.; Shin, D.W.; Kwon, D.Y. Methyl gallate from Galla rhois successfully controls clinical isolates of Salmonella infection in both in vitro and in vivo systems. PLoS ONE 2014, 9, e102697. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Kang, O.H.; Lee, Y.S.; et al. Antibacterial activity of methyl gallate isolated from Galla Rhois or carvacrol combined with nalidixic acid against nalidixic acid resistant bacteria. Molecules 2009, 14, 1773–1780. [Google Scholar] [CrossRef]
- Kang, J.; Liu, L.; Liu, M.; Wu, X.; Li, J. Antibacterial activity of gallic acid against Shigella flexneri and its effect on biofilm formation by repressing mdoH gene expression. Food Control 2018, 94, 147–154. [Google Scholar] [CrossRef]
- Hossain, M.A.; Park, H.C.; Lee, K.J.; Park, S.W.; Park, S.C.; Kang, J. In vitro synergistic potentials of novel antibacterial combination therapies against Salmonella enterica serovar Typhimurium. BMC Microbiol. 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Abuelsaad, A.S.A.; Mohamed, I.; Allam, G.; Al-Solumani, A.A. Antimicrobial and immunomodulating activities of hesperidin and ellagic acid against diarrheic Aeromonas hydrophila in a murine model. Life Sci. 2013, 93, 714–722. [Google Scholar] [CrossRef]
- Kuete, V.; Wabo, G.F.; Ngameni, B.; Mbaveng, A.T.; Metuno, R.; Etoa, F.X.; Ngadjui, B.T.; Beng, V.P.; Meyer, J.J.M.; Lall, N. Antimicrobial activity of the methanolic extract, fractions and compounds from the stem bark of Irvingia gabonensis (Ixonanthaceae). J. Ethnopharmacol. 2007, 114, 54–60. [Google Scholar] [CrossRef]
- Birhanu, B.T.; Lee, E.B.; Lee, S.J.; Park, S.C. Targeting Salmonella Typhimurium invasion and intracellular survival using pyrogallol. Front. Microbiol. 2021, 12, 631426. [Google Scholar] [CrossRef]
- Hirudkar, J.R.; Parmar, K.M.; Prasad, R.S.; Sinha, S.K.; Jogi, M.S.; Itankar, P.R.; Prasad, S.K. Quercetin a major biomarker of Psidium guajava L. inhibits SepA protease activity of Shigella flexneri in treatment of infectious diarrhoea. Microb. Pathog. 2020, 138, 103807. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.K.; Song, M.G.; Park, S.Y. Impact of quercetin against Salmonella Typhimurium biofilm formation on food–contact surfaces and molecular mechanism pattern. Foods 2022, 11, 977. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Sohn, M.J.; Lee, J.; Kim, W.G. Isolation and identification of pentagalloylglucose with broad-spectrum antibacterial activity from Rhus trichocarpa Miquel. Food Chem. 2010, 123, 501–506. [Google Scholar] [CrossRef]
Figure 1.
Antimicrobial activity of T. bellirica and T. chebula fruit extracts in the disc diffusion assays against (A) B. cereus, (B) S. flexneri, (C) S. sonnei, (D) S. typhimurium. TB-AQ = T. bellirica aqueous; TB-MeOH = T. bellirica methanolic; TB-EtOAc = T. bellirica ethyl acetate. TCh-AQ = T. chebula aqueous; TCh-MeOH = T. chebula methanol; TCh-EtOAc = T. chebula ethyl acetate. Negative controls = 1% DMSO and Blank = sterile water. Reference antibiotics: PEN G = penicillin G (10 IU), ERY = erythromycin (10 µg), TET = tetracycline (30 µg), CHL = chloramphenicol (30 µg), CIP = ciprofloxacin (1 µg), POL B = polymyxin B (300IU), OXA = oxacillin (1 µg), AMX = amoxycillin (10 µL of 0.01mg/mL stock solution), GEN = gentamicin (10 µg), VAN = vancomycin (30 µg), AUG = Augmentin® (15 µg), CEF = cefoxitin (30 µg). Horizontal red lines on the y-axis at 6 mm indicates the disc diameter used in the assay. Mean values (±SEM) are reported from three independent studies. p-values < 0.01 are represented with a single asterisk symbol (*), while p-values < 0.001 are represented with a double asterisk symbol (**).
Figure 1.
Antimicrobial activity of T. bellirica and T. chebula fruit extracts in the disc diffusion assays against (A) B. cereus, (B) S. flexneri, (C) S. sonnei, (D) S. typhimurium. TB-AQ = T. bellirica aqueous; TB-MeOH = T. bellirica methanolic; TB-EtOAc = T. bellirica ethyl acetate. TCh-AQ = T. chebula aqueous; TCh-MeOH = T. chebula methanol; TCh-EtOAc = T. chebula ethyl acetate. Negative controls = 1% DMSO and Blank = sterile water. Reference antibiotics: PEN G = penicillin G (10 IU), ERY = erythromycin (10 µg), TET = tetracycline (30 µg), CHL = chloramphenicol (30 µg), CIP = ciprofloxacin (1 µg), POL B = polymyxin B (300IU), OXA = oxacillin (1 µg), AMX = amoxycillin (10 µL of 0.01mg/mL stock solution), GEN = gentamicin (10 µg), VAN = vancomycin (30 µg), AUG = Augmentin® (15 µg), CEF = cefoxitin (30 µg). Horizontal red lines on the y-axis at 6 mm indicates the disc diameter used in the assay. Mean values (±SEM) are reported from three independent studies. p-values < 0.01 are represented with a single asterisk symbol (*), while p-values < 0.001 are represented with a double asterisk symbol (**).
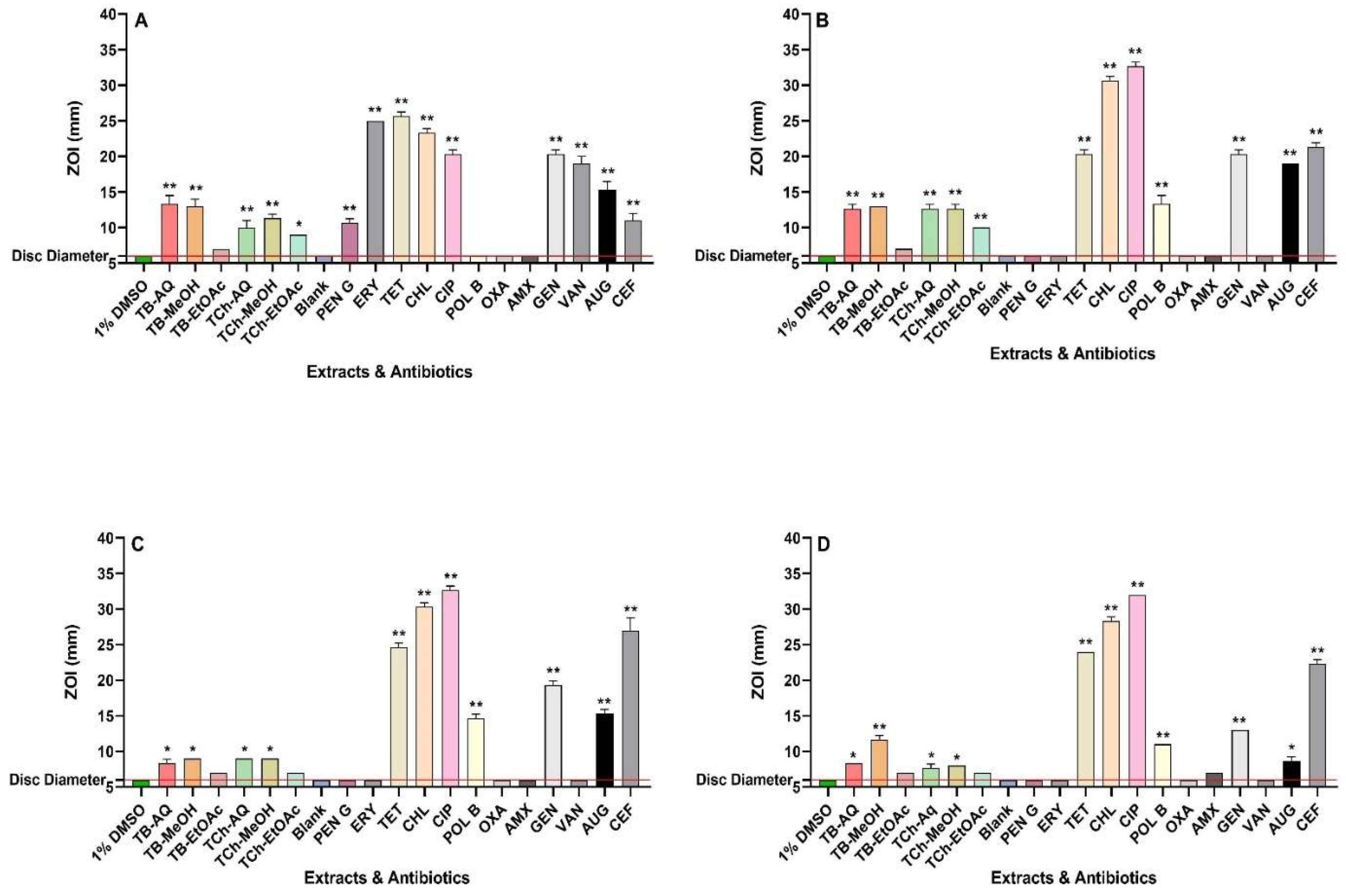
Figure 2.
Structures of noteworthy compounds identified in the fruit extracts of T. bellirica and T. chebula. Methyl gallate (A), propyl gallate (B), gallic acid (C), ellagic acid (D), hamamelitannin (E), pyrogallol (F), quercetin (G), 6-galloylglucose (H), gallic acid 3-O-(6-galloylglucoside) (I), 1,2,6-trigalloyl-β-D-glucopyranose (J), 1,3,6-tri-O-galloyl-β-D-glucose (K), 1,6-bis-O-(3,4,5-trihydroxybenzoyl) hexopyranose (L), chebulic acid (M), and chebuloside II (N).
Figure 2.
Structures of noteworthy compounds identified in the fruit extracts of T. bellirica and T. chebula. Methyl gallate (A), propyl gallate (B), gallic acid (C), ellagic acid (D), hamamelitannin (E), pyrogallol (F), quercetin (G), 6-galloylglucose (H), gallic acid 3-O-(6-galloylglucoside) (I), 1,2,6-trigalloyl-β-D-glucopyranose (J), 1,3,6-tri-O-galloyl-β-D-glucose (K), 1,6-bis-O-(3,4,5-trihydroxybenzoyl) hexopyranose (L), chebulic acid (M), and chebuloside II (N).
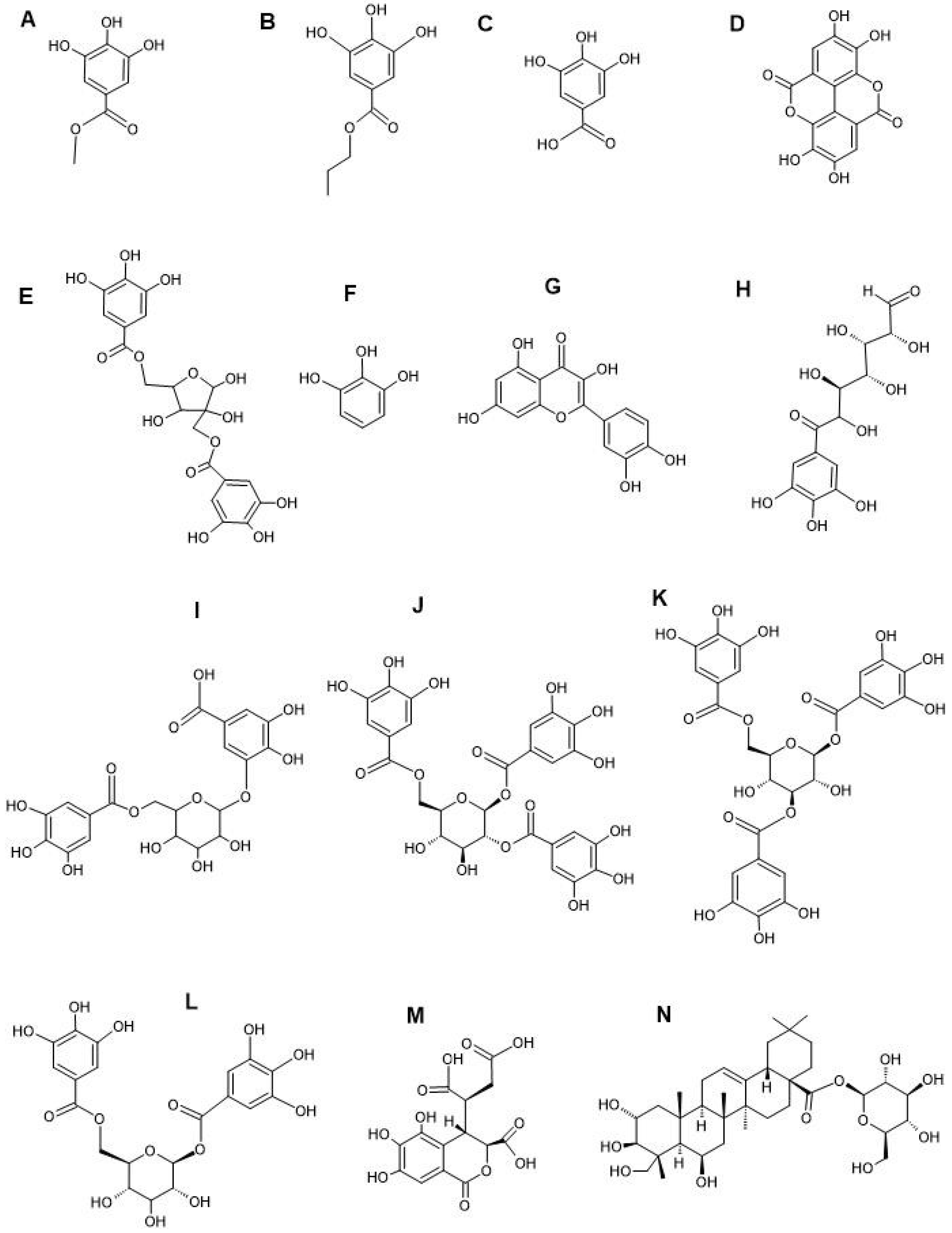

Table 1.
MIC values (µg/mL) for the T. bellirica and T. chebula aqueous, methanolic, and ethyl acetate extracts, as well as the positive control antibiotics, against the bacterial species tested in this study.
Table 1.
MIC values (µg/mL) for the T. bellirica and T. chebula aqueous, methanolic, and ethyl acetate extracts, as well as the positive control antibiotics, against the bacterial species tested in this study.
| Extract type or Antibiotic | Bacterial species & MIC (µg/mL) | |||
|---|---|---|---|---|
| B. cereus | S. flexneri | S. sonnei | S. typhimurium | |
| TB-AQ | 392 | 392 | 1569 | 3138 |
| TB-MeOH | 94 | 377 | 755 | 755 |
| TB-EtOAc | 450 | 450 | >10000 | >10000 |
| TCh-AQ | 556 | 278 | 2225 | 2225 |
| TCh-MeOH | 377 | 377 | 1509 | 3019 |
| TCh-EtOAC | 306 | 306 | 1225 | >10000 |
| PENG | 0.625 | >2.5 | >2.5 | >2.5 |
| ERY | 0.08 | >2.5 | >2.5 | >2.5 |
| TET | 0.08 | 0.625 | 0.31 | 0.625 |
| CHL | 1.25 | 1.25 | 2.5 | 2.5 |
| CIP | 0.08 | 0.02 | 0.02 | 0.02 |
| POL | >2.5 | 0.31 | 0.31 | 0.31 |
| OXA | 1.25 | >2.5 | >2.5 | >2.5 |
| AMX | 0.625 | >2.5 | >2.5 | >2.5 |
| GEN | 0.16 | 0.625 | 2.5 | 0.625 |
| VAN | 0.625 | >2.5 | >2.5 | >2.5 |
MIC values are categorized as inactive (MIC > 10,000 µg/mL), low activity (2000–5000 µg/mL; indicated in bold), moderate activity (1000–2000 µg/mL; indicated in blue), noteworthy activity (400–1000 µg/mL; indicated in red), good activity (100–400 µg/mL; indicated in green), or potent activity (<100 µg/mL; highlighted in yellow). The antibiotics included PENG (penicillin G), ERY (erythromycin), TET (tetracycline), CHL (chloramphenicol), CIP (ciprofloxacin), POLB (polymyxin B), OXA (oxacillin), AMX (amoxycillin), VAN (vancomycin) and GEN (gentamicin). Values for active antibiotics are shown in bold, while those lacking antibiotic activity in this study possessed a MICs greater than 2.5 µg/mL.
Table 2.
∑FIC values determined for the relevant combinations of T. bellirica or T. chebula extracts with conventional antibiotics.
Table 2.
∑FIC values determined for the relevant combinations of T. bellirica or T. chebula extracts with conventional antibiotics.
| Bacteria | Extract | PENG | ERY | TET | CHL | CIP | POLB | OXA | AMX | GEN | VAN |
| B. cereus | TB-AQ | 0.56 | 2.00 | 1.00 | 1.06 | 2.00 | - | 1.03 | 0.56 | 1.50 | 0.56 |
| TB-MeOH | 1.03 | 1.25 | 1.25 | 1.02 | 1.25 | - | 1.01 | 1.03 | 1.13 | 2.07 | |
| TB-EtOAc | 1.50 | 2.12 | 2.12 | 2.00 | 1.06 | - | 1.50 | 1.50 | 8.81 | 1.50 | |
| TCh-AQ | 0.56 | 1.50 | 1.50 | 1.13 | 1.50 | - | 0.53 | 0.81 | 2.00 | 1.26 | |
| TCh-MeOH | 0.78 | 2.00 | 1.00 | 0.53 | 2.00 | - | 0.52 | 0.56 | 0.75 | 1.13 | |
| TCh-EtOAc | 0.51 | 2.25 | 2.25 | 1.50 | 2.25 | - | 0.62 | 1.00 | 2.44 | 1.50 | |
| S. flexneri | TB-AQ | - | - | 1.13 | 1.06 | 2.00 | 5.00 | - | - | 0.56 | - |
| TB-MeOH | - | - | 1.13 | 1.06 | 1.50 | 5.00 | - | - | 1.13 | - | |
| TB-EtOAc | - | - | 0.75 | 1.00 | 1.06 | 15.75 | - | - | 3.00 | - | |
| TCh-AQ | - | - | 1.13 | 1.06 | 1.50 | 5.00 | - | - | 1.13 | - | |
| TCh-MeOH | - | - | 1.13 | 1.06 | 2.00 | 5.00 | - | - | 1.13 | - | |
| TCh-EtOAc | - | - | 1.00 | 1.50 | 2.25 | 16.00 | - | - | 4.00 | - | |
| S. sonnei | TB-AQ | - | - | 1.00 | 0.56 | 1.25 | 8.50 | - | - | 1.12 | - |
| TB-MeOH | - | - | 0.75 | 1.06 | 3.00 | 9.00 | - | - | 0.53 | - | |
| TB-EtOAc | - | - | 3.00 | 3.00 | 3.00 | 4 | - | - | 4 | - | |
| TCh-AQ | - | - | 0.75 | 0.62 | 1.12 | 32.25 | - | - | 0.62 | - | |
| TCh-MeOH | - | - | 1.00 | 1.12 | 2.50 | 16.50 | - | - | 0.56 | - | |
| TCh-EtOAc | - | - | 1.12 | 1.00 | 2.06 | 63.00 | - | - | 4 | - | |
| S. typhimurium | TB-AQ | - | - | 1.00 | 1.25 | 2.44 | 3.00 | - | - | 1.50 | - |
| TB-MeOH | - | - | 0.75 | 1.12 | 2.00 | 2.00 | - | - | 0.75 | - | |
| TCh-AQ | - | - | 1.00 | 1.25 | 2.44 | 6.03 | - | - | 2.00 | - | |
| TCh-MeOH | - | - | 1.00 | 1.25 | 2.44 | 3.00 | - | - | 2.00 | - |
The T. bellirica and T. chebula aqueous extracts (TB-AQ, TCh-AQ), methanolic extracts (TB-MeOH, TCh-MeOH) and ethyl acetate extracts (TB-EtOAc, TCh-EtOAc) were included. Additive interactions are represented by ∑FIC values of >0.5≤1.00; indifferent interactions by >1.01–≤4.00; and antagonistic interaction at >4.0. - indicates the combination could not be tested since an MIC could not be determined for either the extract or the antibiotic. PENG = penicillin G, ERY = erythromycin, TET = tetracycline, CHL = chloramphenicol, CIP = ciprofloxacin, POL B = polymyxin B, OXA = oxacillin, AMX = amoxycillin, GEN = gentamicin, and VAN = vancomycin.
Table 3.
The putative identification and % relative abundance of phytochemicals identified using LC-MS in the fruit extracts of Terminalia bellirica (TB) and Terminalia chebula (TCh).
Table 3.
The putative identification and % relative abundance of phytochemicals identified using LC-MS in the fruit extracts of Terminalia bellirica (TB) and Terminalia chebula (TCh).
| Retention Time (min) | Molecular Weight | Empirical Formula | Putative Compounds | % Relative Abundance (TB) | % Relative Abundance (TCh) | ||||
|---|---|---|---|---|---|---|---|---|---|
| AQ | MeOH | EtOAc | AQ | MeOH | EtOAc | ||||
| 1.496 | 192.06305 | C7H12O6 | Quinic acid | 3.13 | 0.33 | 0.19 | 0.39 | - | 1.53 |
| 1.573 | 344.07409 | C14H16O10 | Theogallin | - | 3.47 | - | - | 0.03 | 0.02 |
| 1.784 | 302.06328 | C12H14O9 | Pyrogallol-2-O-glucuronide | - | 0.21 | - | 0.07 | - | - |
| 2.045 | 294.03749 | C13H10O8 | Banksiamarin B | - | - | 0.15 | 0.17 | 0.05 | 0.13 |
| 2.058 | 174.05268 | C7H10O5 | Shikimic acid | - | 0.41 | 0.67 | 2.64 | 3.14 | 4.80 |
| 2.237 | 154.02646 | C7H6O4 | Gentisic acid | 0.02 | 0.05 | - | 0.01 | - | 0.09 |
| 2.296 | 448.15769 | C19H28O12 | 8-O-Acetyl shanzhiside methyl ester | - | - | - | 0.19 | 0.18 | - |
| 2.356 | 332.07411 | C13H16O10 | 6-Galloylglucose | 6.06 | 3.97 | 3.39 | 3.74 | 0.68 | - |
| 2.444 | 448.06386 | C20H16O12 | Ellagic acid 2-rhamnoside | 0.11 | 1.44 | 0.69 | 0.01 | 0.21 | 0.33 |
| 2.51 | 292.02177 | C13H8O8 | Brevifolincarboxylic acid | 0.09 | - | - | 0.11 | 0.03 | 0.61 |
| 2.56 | 288.0844 | C12H16O8 | Phlorin | - | 0.02 | - | 0.03 | 0.02 | - |
| 3.11 | 484.0848 | C20H20O14 | Gallic acid 3-O-(6-galloylglucoside) | 0.74 | - | - | - | - | - |
| 3.158 | 212.06824 | C10H12O5 | Propyl gallate | - | - | - | 0.30 | 0.35 | 0.12 |
| 4.4 | 277.05821 | C13H11NO6 | Salfredin C1 | - | - | - | 0.14 | - | 0.03 |
| 4.847 | 184.03703 | C8H8O5 | Methyl gallate | 0.25 | 11.39 | - | 0.04 | 1.05 | - |
| 8.44 | 484.0855 | C20H20O14 | 1,6-Bis-O-(3,4,5-trihydroxybenzoyl) hexopyranose | 5.13 | 5.19 | 2.52 | 6.52 | 2.16 | 3.78 |
| 9.316 | 470.01176 | C21H10O13 | Sanguisorbic acid dilactone | - | 0.08 | - | 0.56 | 0.61 | 0.19 |
| 9.567 | 634.08082 | C27H22O18 | Sanguiin H4 | 0.84 | - | 0.07 | - | 5.63 | 0.54 |
| 9.811 | 478.07452 | C21H18O13 | Miquelianin | 0.04 | 0.42 | - | 0.37 | 0.60 | 0.39 |
| 9.902 | 152.04703 | C8H8O3 | Vanillin | - | - | - | 0.16 | 0.17 | 0.17 |
| 9.921 | 126.03146 | C6H6O3 | Phloroglucinol | 0.10 | 1.93 | - | 0.07 | 0.24 | 0.27 |
| 10.043 | 636.09614 | C27H24O18 | 1,2,6-Trigalloyl-β-D-glucopyranose | 0.09 | 5.44 | 2.85 | 0.11 | 1.42 | 2.22 |
| 10.419 | 484.08522 | C20H20O14 | Hamamelitannin | 2.10 | 0.02 | 0.62 | 0.02 | 0.11 | - |
| 10.421 | 126.0316 | C6H6O3 | Pyrogallol | 6.28 | 5.28 | 2.19 | 6.29 | 3.82 | 4.87 |
| 10.422 | 296.05248 | C13H12O8 | cis-Coutaric acid | 0.33 | 0.86 | 0.19 | 0.31 | 0.93 | 1.05 |
| 10.55 | 601.99658 | C28H10O16 | Diellagilactone | - | - | - | 1.13 | 0.60 | - |
| 10.703 | 372.1055 | C16H20O10 | Veranisatin C | 0.02 | 0.04 | - | - | - | - |
| 10.754 | 636.09608 | C27H24O18 | 1,3,6-Tri-O-galloyl-β-D-glucose | - | 0.25 | 0.93 | 0.64 | - | - |
| 11.305 | 170.02138 | C7H6O5 | Phloroglucinic acid | 0.98 | 2.80 | 4.33 | 0.98 | 0.83 | 1.24 |
| 11.091 | 610.1535 | C27H30O16 | Rutin | - | 0.14 | 0.10 | - | 0.03 | 0.08 |
| 11.176 | 432.10539 | C21H20O10 | Vitexin | - | - | - | - | 0.05 | - |
| 11.448 | 302.04213 | C15H10O7 | Quercetin | 0.11 | 0.33 | 0.17 | 0.02 | - | - |
| 11.473 | 464.0951 | C21H20O12 | Myricitrin | 0.05 | 0.13 | 0.19 | - | - | - |
| 11.589 | 170.02147 | C7H6O5 | Gallic acid | 26.23 | 8.64 | 2.60 | 23.90 | 1.90 | 1.56 |
| 11.822 | 462.07961 | C21H18O12 | Aureusidin 6-glucuronide | - | - | 2.52 | - | - | 0.04 |
| 11.905 | 286.04729 | C15H10O6 | Maritimetin | - | - | - | T | 0.01 | 0.01 |
| 12.183 | 448.0996 | C21H20O11 | Maritimein | - | 0.01 | 0.02 | - | - | - |
| 12.185 | 302.00627 | C14H6O8 | Ellagic acid | 2.45 | 10.74 | 33.49 | 8.39 | 10.46 | 11.96 |
| 12.233 | 262.0476 | C13H10O6 | Maclurin | - | 0.02 | - | 0.04 | - | - |
| 12.254 | 148.05223 | C9H8O2 | trans-Cinnamic acid | - | - | - | 0.76 | 0.96 | 0.87 |
| 12.503 | 192.07847 | C11H12O3 | (R)-Shinanolone | - | - | - | 0.04 | 0.03 | 0.05 |
| 12.509 | 610.1894 | C28H34O15 | Neohesperidin | - | - | 0.16 | - | - | - |
| 12.584 | 498.1741 | C23H30O12 | Eucaglobulin | - | 0.02 | - | - | - | - |
| 12.593 | 436.13688 | C21H24O10 | Nothofagin | - | - | - | - | - | 0.12 |
| 12.721 | 680.37692 | C36H56O12 | Tenuifolin | - | - | - | - | 0.11 | 0.37 |
| 12.8 | 486.33397 | C30H46O5 | Bassic acid | - | - | - | - | - | 0.05 |
| 12.844 | 518.1785 | C26H30O11 | Phellodensin E | - | - | 0.05 | - | - | - |
| 12.889 | 346.1052 | C18H18O7 | Hamilcone | - | - | - | 0.02 | - | 0.01 |
| 12.914 | 190.13544 | C13H18O | β-Damascenone | - | - | - | - | 0.02 | 0.06 |
| 12.915 | 666.39739 | C36H58O11 | Chebuloside II | - | - | - | - | 3.85 | - |
| 12.924 | 302.1152 | C17H18O5 | Lusianin | 0.01 | - | - | - | - | - |
| 12.929 | 356.03763 | C14H12O11 | (+)-Chebulic acid | 0.54 | - | - | 3.98 | 1.40 | 2.88 |
| 12.969 | 504.34417 | C30H48O6 | Madecassic acid | - | - | - | - | - | 1.42 |
| 12.971 | 202.13522 | C14H18O | (±)-Anisoxide | - | - | - | - | - | 0.05 |
| 12.973 | 244.14583 | C16H20O2 | Lahorenoic acid C | - | - | - | - | - | 0.02 |
| 13.208 | 288.06318 | C15H12O6 | Eriodictyol | - | - | - | - | - | 0.27 |
| 13.428 | 442.1992 | C25H30O7 | Exiguaflavanone M | - | - | 0.13 | - | - | - |
| 13.451 | 274.08371 | C15H14O5 | Phloretin | - | - | - | 0.01 | 0.02 | 0.01 |
| 13.536 | 462.11641 | C22H22O11 | Leptosin | - | 0.01 | - | 0.01 | 0.06 | - |
| 13.59 | 302.04255 | C15H10O7 | Bracteatin | - | - | - | 0.01 | 0.01 | - |
| 13.593 | 502.1836 | C26H30O10 | Flavaprin | - | - | 0.07 | - | - | - |
| 13.678 | 568.12178 | C28H24O13 | Isoorientin 2"-p-hydroxybenzoate | - | - | - | - | - | 0.04 |
| 13.703 | 280.13067 | C15H20O5 | Artabsinolide A | - | - | - | - | - | 0.02 |
| 13.805 | 444.10499 | C22H20O10 | 3'-O-Methylderhamnosylmaysin | - | - | - | 0.04 | 0.12 | 0.08 |
| 13.805 | 292.09447 | C15H16O6 | (S)-Angelicain | - | - | - | 0.05 | - | - |
| 14.024 | 500.1674 | C26H28O10 | Ikarisoside A | - | - | 0.03 | - | - | - |
| 14.078 | 534.28285 | C29H42O9 | Corchoroside A | - | - | - | - | - | 0.02 |
| 14.358 | 272.06832 | C15H12O5 | Naringenin | - | - | - | - | - | 0.18 |
| 14.367 | 470.33907 | C30H46O4 | Glycyrrhetinic acid | - | - | - | - | 0.08 | 0.01 |
| 14.783 | 234.16167 | C15H22O2 | Valerenic acid | - | - | - | - | - | 0.13 |
| 14.928 | 202.17175 | C15H22 | Rulepidadiene B | - | - | - | - | - | 0.08 |
| 14.93 | 222.16153 | C14H22O2 | Rishitin | - | - | - | - | - | 0.04 |
| 15.046 | 426.09477 | C22H18O9 | Epiafzelechin 3-O-gallate | - | - | - | - | - | 0.01 |
| 15.454 | 650.4026 | C36H58O10 | Pedunculoside | - | - | - | - | 1.18 | 2.61 |
| 16.257 | 738.41926 | C39H62O13 | Isonuatigenin 3-[rhamnosyl-(1->2)-glucoside] | - | - | - | - | 0.04 | 0.12 |
| 16.301 | 470.1941 | C26H30O8 | Limonin | - | - | 0.14 | - | - | - |
| 16.478 | 540.1663 | C25H32O11S | Sumalarin B | - | - | 0.02 | - | - | - |
| 16.532 | 316.1308 | C18H20O5 | Methylodoratol | - | - | 0.02 | - | - | - |
| 16.787 | 472.2097 | C26H32O8 | Kushenol H | - | - | 0.07 | - | - | - |
| 16.844 | 344.0532 | C17H12O8 | 3,4,3'-Tri-O-methylellagic acid | - | - | 0.01 | - | - | - |
| 16.876 | 252.20856 | C16H28O2 | Isoambrettolide | - | - | - | - | - | 0.32 |
| 17.064 | 440.1828 | C25H28O7 | Lonchocarpol E | - | - | 0.15 | - | - | - |
| 17.209 | 544.2668 | C30H40O9 | Physagulin F | - | - | 0.10 | - | - | - |
| 17.276 | 180.11471 | C11H16O2 | Jasmolone | - | 0.61 | 1.48 | 0.05 | - | - |
| 17.31 | 504.34409 | C30H48O6 | Protobassic acid | - | - | - | 1.26 | 0.02 | 0.01 |
| 17.33 | 468.32326 | C30H44O4 | Glabrolide | - | - | - | 0.87 | - | 3.19 |
| 17.33 | 502.32964 | C30H46O6 | Medicagenic acid | - | - | - | - | T | 0.07 |
| 17.331 | 200.15605 | C15H20 | (S)-gamma-Calacorene | - | - | - | 0.02 | - | 0.29 |
| 17.363 | 696.40877 | C37H60O12 | Momordicoside E | - | - | - | - | 0.01 | 0.04 |
| 17.626 | 286.08357 | C16H14O5 | Homobutein | - | T | 0.01 | - | - | 0.01 |
| 17.635 | 282.12541 | C18H18O3 | Ohobanin | 0.01 | T | 0.09 | T | - | 0.06 |
| 17.671 | 652.27302 | C32H44O14 | Dicrocin | - | - | - | - | - | 0.03 |
| 17.902 | 424.1881 | C25H28O6 | Paratocarpin G | T | - | - | - | - | - |
| 17.949 | 456.2144 | C26H32O7 | Antiarone J | - | 0.54 | - | - | - | - |
| 18.124 | 372.2509 | C20H36O6 | Sterebin Q4 | - | - | 0.01 | - | - | - |
| 18.237 | 428.1831 | C24H28O7 | Heteroflavanone B | - | - | 0.05 | - | - | - |
| 18.237 | 312.13615 | C19H20O4 | Desmosdumotin C | T | - | 0.02 | T | - | 0.02 |
| 18.292 | 546.35535 | C32H50O7 | Hovenidulcigenin B | - | - | - | 0.04 | - | - |
| 18.339 | 236.1776 | C15H24O2 | Capsidiol | - | - | - | - | - | 0.02 |
| 18.346 | 268.13069 | C14H20O5 | Kamahine C | 0.02 | 0.01 | - | - | 0.01 | 0.15 |
| 18.363 | 336.0994 | C20H16O5 | Ciliatin A | - | - | - | - | 0.01 | - |
| 18.498 | 340.09447 | C19H16O6 | Ambanol | - | - | - | - | 0.01 | - |
| 18.507 | 484.31803 | C30H44O5 | Liquoric acid | - | - | - | T | 0.01 | 0.04 |
| 18.623 | 488.35017 | C30H48O5 | Pitheduloside I | - | - | - | T | 0.06 | 0.11 |
| 19.641 | 226.09903 | C15H14O2 | 7-Hydroxyflavan | - | 0.84 | - | 0.03 | - | 0.05 |
| 18.748 | 208.1096 | C12H16O3 | Isoelemicin | 0.01 | - | 0.19 | - | - | - |
| 18.767 | 526.2565 | C30H38O8 | Kosamol A | - | 0.52 | - | - | - | - |
| 18.987 | 372.1208 | C20H20O7 | Tangeretin | - | - | 0.03 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



