Submitted:
30 October 2024
Posted:
31 October 2024
You are already at the latest version
Abstract
Poultry farming faces significant challenges due to viral diseases, which lead to high mortality and economic losses. Long noncoding RNAs (lncRNAs) have emerged as crucial regulators in the host immune response to viral infections. This review explores the multifaceted roles of lncRNAs in chicken viral diseases, including avian leukosis, Marek’s disease, infectious bursal disease, avian influenza, infectious bronchitis, and Newcastle disease. LncRNAs influence gene expression at various levels, such as chromatin remodeling, transcriptional regulation, and post-transcriptional modifications. They interact with viral and host genes, modulating immune responses and viral replication processes. Specific lncRNAs, such as ERL lncRNA, linc-GALMD3, and loc107051710, have been identified as key players in the pathogenesis and immune response to these viral infections. Understanding the mechanisms of lncRNA action provides insights into potential diagnostic and therapeutic strategies to enhance disease resistance in poultry. This review highlights the importance of lncRNAs in the complex interplay between host and virus, paving the way for future research and applications in poultry health management.
Keywords:
noncoding RNAs
; LncRNAs
; viral diseases
; poultry
; chicken
1. Introduction
Poultry meat has become the most consumed livestock commodity in the world especially in the developing countries. Due to the increase in demand for poultry meat, there has been an increase in global imports during the period from 2001 to 2021. Poultry meat import is going to be the most in the next ten years and most of the nations raised their domestic poultry production to match this need. Major exporters of poultry meat are Brazil, the United States, the European Union, and Thailand. The world’s leading poultry exporter, Brazil is going to remain in the top position through 2031 (USDA, 2022).
The main obstacle to modern poultry farming is viral diseases in the birds. Poultry birds are frequently the prey of viral diseases (some of which include zoonotic diseases), which result in high mortality and morbidity. These diseases are caused by a variety of factors, including the environment, stress, nutritional and immune-suppressive additives in poultry feed, antimicrobial resistance, irregular vaccination, and a lack of proper biosecurity. This in turn has an impact on the economics of poultry production, increasing losses and decreasing gains. Viruses are microorganisms that are inert particles and can only survive in living organisms. In order to avoid being recognized and eliminated by the host immune system, it must take advantage of the host immune system. However, in response to a virus's entry, the host immune system will use a variety of immunoregulation mechanisms to remove the virus from the host. Viruses create a variety of proteins that resemble crucial host-specific components in order to evade the host immune system (Mohandas et al., 2018). Reticulo-endotheliosis virus, Lymphoid Leukosis (LL), Infectious Bursal Disease Virus (IBDV), Avian Influenza Virus (AIV), Newcastle Disease Virus (NDV), Marek's Disease Virus (MDV), and Chicken Infectious Anaemia Virus (CIAV) are some of the avian viruses that significantly cause immunosuppression (Balamurugan and Kataria, 2006).Co-infection of one or more viruses can worsen immunosuppression and accelerate the progression of the disease in birds, resulting in symptoms such as decreased feed conversion ratio (FCR) and weight gain, incapability to eat, lethargy, and eventually death.
2. Long Non-Coding RNA (lncRNA)
The diseases caused by viruses pose a great challenge to human health, the development of which has been driven by the imbalanced host immune response. Host innate immunity is an evolutionary defense system that is critical for the elimination of the virus. The overactive innate immune response can also lead to inflammatory autoimmune diseases, which require precise control of innate antiviral response for maintaining immune homeostasis. Innate immunity is the first and most rapid line of defense against the invasion of microbial pathogens (Wang et al., 2020). Non-coding RNAs with a length of at least 200 nucleotides are referred to as lncRNAs (Braconiet al., 2011).The conventional lncRNAs are divided into five classes based on their position in relation to protein-coding genes (P-CGs): (i) long intergenic transcripts; (ii) intronic lncRNAs (located within the intron of P-CGs); (iii) bidirectional lncRNAs (transcribed in opposite directions with the promoter of P-CGs); (iv) antisense lncRNAs (transcribed across the exons of a P-CGs from the opposite direction); and (v) pseudogene-type lncRNAs (transcribed from a gene without the ability to produce proteins)(Zhang et al., 2016;Ma et al., 2013). Protein interactors of lncRNAs, such as conventional and non-conventional RNA-binding proteins (RBPs), are critical to the achievement of lncRNA functions (Yao et al., 2019; Zhang et al., 2016, Li et al., 2014; Chaudhury et al., 2020; Garcia- Moreno et al., 2018; Hentze et al., 2018; Lin et al., 2019). Many lncRNAs are gradually being recognised as key components of virus-host interaction, mainly through antiviral response-independent and -dependent mechanisms.
It is known that different transcript products of the genome, including enhancers, pseudogenes, intronic transcripts, antisense transcripts, intergenic transcripts, and retrotransposons, can give rise to lncRNA (Guttman et al., 2013; Beermann et al., 2016). Thus, there are antisense, divergent, intron, intergenic, enhancer, promoter, and transcription start site-associated lncRNAs in the cluster of lncRNAs, depending on the relative positions of the lncRNA and the encoded gene (Tahira et al., 2014). Normally, during pre-transcriptional, transcriptional, and post-transcriptional processing, lncRNAs function as cis-acting elements or trans-acting factors to control gene expression. LncRNA can take part in X chromosome inactivation through interactions with various proteins (Wang and Cheng, 2011),genomic imprinting (Lee, 2012; Chaumeil et al., 2006; Gebert et al., 2009; Brown et al., 1992), chromatin modifications (Werven et al., 2012; Redon et al., 2010; Martens et al., 2016), DNA methylation (Baniushin, 2005; Bao et al., 2014; O’Leary et al., 2005), mRNA degradation (Wilusz et al., 2009; Wang et al., 2013). LncRNA also plays the role of regulating innate immune response either by adsorbing miRNA through the sponge effect or directly binding to the component of innate immune molecules (Zeng et al., 2021). Furthermore, it controls mRNA splicing (Beltran et al., 2008). LncRNAs use transcriptional interference to control the splicing of genes (Latos et al., 2004), and some lncRNAs possess ORFs and use coding of one or more micro-peptides to demonstrate their unique regulatory roles (Jackson et al., 2004).
After more than ten years of research, scientists have connected the molecular mechanisms of lncRNAs to significant regulatory molecules that are functional in a range of biological and pathological processes (Smith and Chang, 2016). First, lncRNAs have the ability to control chromatin modifications. Rinn et al. (2007) found that lncRNA HOTAIR interacts directly with polycomb repressive complex 2 (PRC2) and further modifies chromatin silencing mediated by PRC2. Besides regulating transcription, lncRNAs can also interact with proteins to control crucial signalling cascades and post-transcriptional processes. Additionally, lncRNAs can function as ceRNAs to "sponge" or absorb miRNAs and stop the miRNAs' inhibitory effect on the genes they target. Wang et al. (2016), for instance, reported that lncRNA NRF could bind hsa-miR-873, upregulate RIPK1/RIPK3, and enhance necrosis. Finally, it is possible that lncRNAs can influence the stability and translation of mRNA in the cytoplasm and play a role in controlling processes that are dependent on cells, including apoptosis, migration/invasion, and growth (Wang et al., 2019; Zhao et al., 2018).
2.1. Role of lncRNAs in Gene Expression
RNA polymerase II (Pol II) is responsible for the transcription of lncRNAs, and they share a similar biogenesis with mRNAs due to their polyadenylation and 5'-cap formation. Although their exon count and splicing efficiency are typically lower than those of mRNAs, lncRNAs frequently undergo splicing(Zuckerman and Ulitsky, 2019; Tilgner, 2012; Mele et al., 2017; Hezroniet al., 2015). LncRNAs control gene expression on a number of levels. LncRNAs can influence chromatin structure and function, the transcription of nearby and distant genes, RNA splicing, stability, and translation, and more by interacting with proteins, DNA, and RNA. Moreover, lncRNAs have a role in the synthesis and control of nuclear condensates and organelles.
2.1.1. Chromatin Structure Regulation:
RNA-chromatin association detection and chromatin conformation capture techniques reveal complex regulation of chromatin architecture and gene expression in lncRNA. RNA has inherent regulatory potential, as its negative charge can neutralize histone tails, leading to chromatin de-compaction (Chu et al., 2011; Li et al., 2017; Bonetti et al., 2020). Nuclear lncRNAs interact with DNA to alter the chromatin environment, either indirectly through affinity for proteins or by binding to specific DNA sequence. Protein-assisted long-range chromatin interactions can facilitate direct lncRNA transcriptional effects on target genes (Saldana-Meyer et al., 2019). LncRNAs can create hybrid structures with DNA, influencing chromatin accessibility. These interactions can take the form of triple helices or R-loops, which are considered threats to genome stability (Tang Wong et al., 2019; Niehrs et al., 2020). However, recent findings suggest that this can be effective regulators of gene expression and coordinators of DNA repair, with lncRNAs interacting with these structures (Schmitz et al., 2010; Martianov et al., 2005; O’Leary et al., 2015; Mondal et al., 2015; Grope et al., 2013).
2.1.2. Regulation of Transcription by lncRNA:
One of the main factors influencing the regulatory interaction between a lncRNA and its neighbouring genes is their relative position. Given that bidirectional and extensive antisense lncRNA transcription has been shown to be evolutionarily conserved (Seila et al., 2008), the non-random genomic distribution of lncRNAs may indicate that genes have evolved to regulate their own expression in a context-specific way. For example, divergent lncRNAs' genomic organization is essential for cis-gene regulation. Two primary, non-exclusive methods can mediate this regulation: either the lncRNA transcript regulates the neighbouring loci, or the transcription or splicing process of the lncRNA results in a chromatin state or steric hindrance that affects the expression of neighbouring genes (Luo et al., 2016). By interfering with the transcription machinery, lncRNAs can decrease the expression of genes through modifying the recruitment of transcription factors or Pol II to the blocked promoter (Latos et al., 2012), altering histone modifications (Stojic et al., 2016), and decreasing chromatin accessibility(Thebault et al., 2011).The conserved lncRNA CHD2 adjacent, suppressive regulatory RNA (CHASERR), which is situated upstream of the chromatin remodeller Chd2 gene, exemplifies an additional way that lncRNAs can utilize to control extensive transcriptional inhibition. It was discovered that accessibility at the Chd2 promoter and numerous additional promoters—all of which were controlled by CHD2—was enhanced by Chaserr depletion (Rom et al., 2019). Membrane less RNA–protein compartments called nuclear condensates are essential for numerous biological activities. Various abundant lncRNAs have scaffolding or regulatory functions that are necessary for the building and operation of multiple nuclear condensates (Banani et al., 2017).
2.1.3. Role of lncRNA in Post-Transcriptional Regulation:
By binding to RNA sequence motifs or structures, proteins can be sequestered by lncRNAs and form specific lncRNA–protein complexes (lncRNPs). This results in an altered mRNA splicing and turnover, the modulation of signalling pathways in certain biological contexts. LncRNAs regulate many other aspects of gene expression in addition to their involvement in nuclear organisation and transcription regulation. Some lncRNAs are even translated into functional peptides (Hartford and Lal, 2020). trans-acting lncRNAs establish distinct structural motifs or engage in sequence-based interactions with RBPs. Pyrimidine-rich non-coding transcript (PNCTR) inhibits PTBP1-mediated mRNA splicing elsewhere in the nucleoplasm by confining pyrimidine tract-binding protein 1 (PTBP1) to the perinucleolar compartment (PNC) (Yap et al., 2018). Non-coding RNA triggered by DNA damage (NORAD), which is extensively produced in the cytoplasm after DNA damage, sequesters Pumilio (PUM) proteins to preserve genomic stability. PUM RBPs are sequestered in the cytoplasm by NORAD, which represses the translation and stability of the mRNAs it binds to (Lee et al., 2016; Tichon et al., 2018;Wang et al., 2002). Through base pairing, these trans-acting lncRNAs interface directly with other RNAs.
2.2. Role of lncRNA in Innate Antiviral Response
Proinflammatory activation and an antiviral response are brought on by the activity of host immune system, which plays a crucial role in the defense against viral infection (Altfeld and Gale, 2015; Takeuchi and Akira, 2009). Upon viral infection, pathogen-associated molecular patterns (PAMPs) on viruses are recognized by pathogen recognition receptors (PRRs), including toll-like receptors (TLRs), which in turn initiate the innate immune response (Xu and Zhong, 2016; Coccia and Battistini, 2016). According to Carnero et al., lncRNA EGOT induced by TLR4/TLR7 amplifies viral replication and antagonizes the antiviral response (Carnero et al, 2016). Furthermore, lncRNA EGOT may be involved in a number of processes during viral infection, given the critical roles played by TLR4 and TLR7 in host immunity and their crosstalk with transduction signalling pathways like NF-κB and IFN pathways (Zhang et al., 2007; Velloso et al., 2015; Schultheiss and Thimme, 2007). Further research is necessary to fully understand the precise downstream mechanisms of lncRNAs/TLRs in the antiviral immune response, even though the TLR-induced viral response causes the upregulation of lncRNAs that modulate the innate immunity.
Interferons are the primary immunomodulatory and pro-inflammatory cytokines in the antiviral immune response, and they also aid in the induction of lncRNA activity (Doyle et al., 2015). An example can be taken from the work of Kotzin, wherein he established that early on in both acute and chronic lymphocytic choriomeningitis virus infections, T-cell receptor (TCR) and type I IFN activation induce the lncRNA known as Morrbid. LncRNA Morrbid works by suppressing the PI3K/AKT pathway and inducing the pro-apoptotic gene BCL2L11, which in turn helps to regulate CD8 T-cell survival and differentiation, thus helping in immune response (Kotzin et al., 2019).
Not only are lncRNAs stimulated by the viral immune system but can also modulate the immune response by regulating important immune molecules like cytokines. According to Li et al. (2014), TNFα and hnRNPL related immune-regulatory lincRNA THRIL is necessary for TNFα expression. THRIL regulates TNFα downstream targets and interacts with hnRNPL directly. THRIL induces other cytokines and chemokines, including IL-8, CXCL10, and CSF1, but further research is needed to determine the mechanisms.
According to newly available research, lncRNAs have the ability to transcriptionally control gene expression, which can alter viral replication and the immune response (Zhou et al., 2019; Ouyang et al., 2017;Salerno et al., 2016). First, the transcription of innate immune genes like IFN and ISGs can be activated or repressed by lncRNAs through the recruitment of transcription factors (TFs) (Liu and Ding, 2017).It was demonstrated by Ma and colleagues that the Hantaan virus (HTNV) induces the lncRNA NEAT1. The SFPQ is relocated to paraspeckles by induced NEAT1, which also reverses the transcriptional inhibition of RIG-I and DDX60. The IFN response induced by RIG-I is facilitated and endogenous RIG-I expression is promoted by restored DDX60. As a result, NEAT1 has the ability to influence the innate immune response and activate IFN signalling, providing negative feedback against the HTNV viral infection (Ma et al., 2017).
dsRNA-binding proteins are important components of the antiviral innate immune system and play a crucial role in repressing viral replication by causing various changes in cellular and viral RNA processes (Cadena and Hur, 2017). Exogenous circRNAs can initiate an innate immune response that provides protection against viral infection, according to research by Chen et al.(2017). The authors found that in various cell lines, circRNAs potently induced the expression of multiple innate immune system regulatory genes, such as retinoic acid-inducible gene-I (RIG-I), protein kinase R (PKR), melanoma-differentiation-associated gene 5 (MDA5), 2′-5′ oligoadenylate synthase 1 (OAS1), and OAS-like protein (OASL).
2.3. Role of lncRNA In Virus Pathogenesis
Given how pleiotropic their functions are, it is not surprising that lncRNAs could play a role in virus replication. It has been discovered that viruses with low coding capacity control the expression and function of both host and viral genes by means of cellular lncRNA. Various animal viruses have been demonstrated to dysregulate host lncRNA expression, including avian leukemia virus (Tam et al., 1997), herpes simplex virus (Sonkoly et al., 2005), Marek's disease virus (Ahanda et al., 2006), human immunodeficiency virus (HIV) (Zhang et al., 2013), hepatitis B virus (HBV) (Braconi et al., 2011) and severe acute respiratory syndrome coronavirus (SARS-CoV) (Peng et al., 2010). Subsequent research has demonstrated that viral genes control the level of cellular lncRNAs, which in turn controls the expression of genes encoding proteins and ultimately promotes viral infection (Yi et al., 2019).
2.3.1. LncRNA In Viral Gene Expression:
The viral genome controls the expression of "late" proteins, which are required to assemble the capsid and package the viral genome, and "early" proteins, which allow genome replication, once the virus enters the proper cell compartment. Moon et al. (2012) reported that sub-genomic flavivirus RNA (sfRNA), a lncRNA partially degraded from the viral genomic RNA, most likely by the cellular 5'–3' exoribonuclease XRN1, inhibits XRN1 activity and modifies host mRNA stability in cells infected with the dengue or Kunjin viruses. This effect might interfere with the control of host cell gene expression and help stabilize viral transcripts. According to a study by Rosetto et al. (2013), polyadenylated nuclear (PAN) RNA, a lncRNA encoded by the genome of the Kaposi's sarcoma-associated herpesvirus (KSHV), can physically interact with the KSHV genome to transcriptionally activate the expression of the KSHV gene. Another study conducted in 2013 by Lisnic and colleagues demonstrated that PAN RNA can alleviate gene suppression by binding to the host poly(A)-binding protein C1 (PABPC1) to regulate mRNA stability and translation efficiency. This serves as a molecular scaffold for chromatin modifying enzymes to remove the H3K27me3 mark, which is necessary for the production of late viral proteins.
2.3.2. LncRNA In Viral Replication:
Whereas RNA viruses copy their genomes directly to RNA, DNA viruses copy their genomes straight to DNA. On the other hand, certain RNA viruses and certain DNA viruses use DNA and RNA intermediates to copy their genomes (Wang et al., 2017).
RNA-RNA interaction and RNA-protein interaction are two ways that virus-encoded lncRNAs control viral replication. Numerous secondary stem-loop II structures can be found in the 3' untranslated region (UTR) of flavivirus RNA genome. These structures prevent the nuclease XRN1 from breaking them down, which produces the functional lncRNA, i.e., sfRNA (Funk et al., 2010). It was reported that the regulation of the efficiency of flavivirus genome replication was significantly influenced by sfRNA. The host's innate immune response has the ability to produce miRNA and eliminate the viral genomic RNA during a flavivirus infection. But in order to protect viral genomic RNA, sfRNA bound to the miRNAs and caused their degradation, which allowed the virus to replicate in the host cell (Pijlman et al., 2008).
2.3.3. LncRNA In Viral Assembly and Release:
Once the newly generated viral genomes and proteins are adequately prepared within the host cells, they are packaged, assembled, and released. Additionally, these processes involve HBoV1 BocaSR viral lncRNAs (Wang et al., 2017). According to findings, the Japanese Encephalitis virus (JEV) sfRNA is a trans-acting riboswitch that promotes genomic RNA synthesis, packaging, and virion release while inhibiting the translation of JAV and host antigenomic genes. It is found in the late stages of the viral replication cycle (Pijlman et al., 2008). Thus, these results indicate that lncRNAs produced by viruses are probably involved in controlling the entire viral life cycle, which includes viral genome replication, gene expression, assembly, and release of virions.
2.4. Methods of Detecting lncRNA
Based on sequence similarity and the existence of long open reading frames (ORFs), it is possible to anticipate protein-coding gene transcripts and several families of non-coding RNAs (tRNAs and rRNAs) with a fair degree of accuracy (Lander et al., 2001). The following is a list of some of the techniques.
2.4.1. Full- Length cDNA Sequencing:
Creating a whole cDNA sequence is the best way to create precise transcript models. The Functional Annotation of The Mammalian Genome project (FANTOM) was the first effort to characterize the entire coding capacity of the mammalian genome. Cap-analysis of gene expression (CAGE) tag sequences (FANTOM3) was used to identify transcriptional start sites (TSSs) for future FANTOM data releases. Using this technique, approximately 30 percent of all transcripts analysed were categorized as non-coding RNAs (Carninci et al., 2005).
2.4.2. Chromatin State Maps:
Histone alterations are conserved among mammalian species, and their associations with transcriptional activity justify their use as indicators of various regulatory components [Bernstein et al., 2005]. Particularly, extensive investigations using chromatin immunoprecipitation in conjunction with hybridization to microarray tiling arrays (ChIP-chip) have demonstrated that certain histone marks are suggestive of particular regulatory elements (Bernstein et al., 2005; Heintzmann et al., 2007). Guttman and colleagues found around 1250 potential lncRNA loci in various mouse cell lines using the K4–K36 mark (Guttmann et al., 2009). After analysing expression data from four mouse cell lines, embryonic development over time, and multiple adult tissues, the researchers concluded that these identified lncRNAs may play a role in various biological processes, such as ESC pluripotency, neuronal development, and immune function. This study successfully uncovered numerous lncRNAs by leveraging knowledge of the connection between chromatin state and transcriptional activity, although it had some limitations.
2.4.3. RNA Sequencing:
RNA-seq deep sequencing provides a more straightforward method for evaluating transcriptomes compared to the analysis of chromatin markers. By sequencing short cDNA fragments, it is possible to effectively reconstruct the entire transcriptome, delivering a broader dynamic range than microarrays. This technique also enables the identification of novel loci and transcripts, in addition to quantifying the prevalence of alternatively spliced variants. (Doerr, 2012). The lack of evidence for transcripts that extend across adjacent protein-coding genes and lncRNA loci serves as an additional criterion for differentiating the two. Consequently, researchers aiming to identify functional lncRNAs within their specific systems consider RNA-seq to be highly beneficial (Nicholas and Ponting, 2013) as shown in Figure 1.
The subsequent computer reconstruction of transcriptomes has made it possible to identify a number of lncRNA primary characteristics. For instance, the locations of lncRNA loci and protein-coding genes are not always the same; their transcripts can be in intergenic sequence, within the introns of protein-coding genes, or overlap them on the same or opposite strand. Only RNA-seq (or cDNA sequencing) can provide the possibility of classifying lncRNAs other than intergenic non-coding RNAs. This observation was based on research conducted in 2013 by Pauli and his colleagues using RNA-seq during the embryogenesis to identify long noncoding RNAs expressed in zebrafish embryo in a systematic manner. They identified 1133 noncoding multi-exonic transcripts expressed during development. These consist of precursors for small RNAs (sRNAs), intronic overlapping lncRNAs, exonic antisense overlapping lncRNAs, and long intergenic ncRNAs (lincRNAs). They noticed that the temporal expression profile of lncRNAs disclosed two novel characteristics: they were selectively enriched in early-stage embryos and were expressed within smaller time windows compared to protein-coding genes. Moreover, a number of lncRNAs exhibited unique subcellular localization patterns and tissue-specific expression.
2.5. Role of lncRNAs in Specific Chicken Viral Diseases
Long non-coding RNAs have a significant impact on regulating gene expression in addition to genes, which are essential in determining the host immune response. Extended non-coding RNAs impact genes at various phases, such as chromatin remodelling-induced activation or inactivation, transcription activation or suppression through binding to transcription factors, translation inhibition and splicing modulation through transcript binding, and mRNA degradation through microRNA regulation. It has also been reported that long non-coding RNAs have an impact at the protein level (Statello et al., 2021).
2.5.1. Avian Leucosis:
Infection with the avian leukosis virus J (ALVJ) causes hemangioma and hematopoietic malignancy in myeloid leukemia in chickens. On the other hand, nothing is known about the mechanisms underlying the distinct pathophysiology of ALVJ. Research on the differentially expressed lncRNAs (DE lncRNAs) between non-infected and infected tissues (Lan et al., 2017; Qiu et al., 2018), chick embryo fibroblast cells (Hue et al., 2018), or primary monocyte-derived macrophages from chickens (Dai et al., 2019) was found to have the potential to interact with immune-related miRNAs and genes to demonstrate the role of DElncRNAs in disease processes. A number of lncRNAs (XLOC_672329, ALDBGALG0000001429, XLOC_016500, and ALDBGALG0000000253) were hypothesized to cis-regulate cholesterol 25-hydroxylase [(CH25H)/cytokine inducible SH2 containing protein (CISH)/interleukin 1 beta (IL-1β)/CD80 molecule (CD80)] in order to take part in host antiviral responses, according to Dai et al. (2019).
Hu and colleagues (2018) utilised RNA-Seq to conduct a systematic review of the lncRNAs in CEF cells and analyzed the gene expression profiles of ALVJ-infected chicken cells. Target prediction analysis showed that five lncRNAs- MG066601, MG066617, MG066602, MG066618 and MG066603 may act in cis or trans and affect the expression of genes involved in the anti-viral innate immune responses. They found 36 differentially expressed lncRNAs and 91 genes, and the results showed that the JAK-STAT signalling pathway, toll-like receptor, RIG-I receptor, and NOD-like receptor were all enriched.
2.5.2. Marek’s Disease:
MDV is a member of the α-Herpesvirinae family of Herpesviruses, with linear dsDNA and which causes tissue hyperplasia and neoplasia along with a highly contagious phymatosis. The pathophysiological mechanism of MD is prompted by proteins encoded by the virulent genes of the MDV. There are three serotypes of MDV; namely serum type 1 (MDV-1) or Gallid herpesvirus (GaHV-2), serum type 2 (MDV-2) or Gallid herpesvirus 3 (GaHV-3), and serum type 3 (MDV-3), that is also known as Meleagrid herpesvirus I (MeHV-1) (Davison et al., 2009). Three different types of lncRNAs—ERL lncRNA, linc-GALMD3, and linc-stab1 are involved in the MDV infectivity process.
Figueroa et al. (2016) reported that the ERL lncRNA is a naturally occurring antisense transcript of the MDV carcinogen, with a length of 7.5 kbp. It is connected to the adenosine to inosine acting on RNA (ADAR1) protein. During the lytic and latent phases of viral infection and reactivation, ERL lncRNA is expressed. In chickens, the JAK/STAT (Janus kinase/signal transducer and activator of transcription) and IFN-response pathway controls the expression of ADAR1 via an inducible promoter containing IFN-stimulated response elements.
According to research by Burnside et al. (2008) and Zhao et al. (2015), the unedited ERL lncRNA has a long half-life because stable introns have been removed and the expression of most of the miRNA MDV-miR-M4 and lytic-associated miRNA MDV-miR-M1 has been inhibited.
gga-miR-223 is a downstream non-coding RNA whose expression is regulated by the long intergenic non-coding RNA linc-GALMD3. The expression of the gga-miR-223 gene was markedly reduced by a lincGALMD3 knockdown in the MDCC-MSB1 cells, while the expression of other genes was increased (Zhang et al., 2021). According to Han and colleagues (2017), linc-GLAMD3 trans-regulates the expression of other genes in the chicken genome and cis-regulates the expression of gga-miR-223. Using RNA-Seq and qRT-PCR, they discovered and confirmed that MDV-infected CD4+T cells had high expression of linc-GALMD3. After linc-GALMD3 function was lost by shRNA, RNA-Seq analysis in MDCC-MSB1 cells revealed that linc-GALMD3 could positively cis-regulate the expression of its downstream gga-miR-223 gene. Although there is not any experimental support for the gga-miR-223, it targets the insulin-like growth factor 1 receptor (IGF1R), which controls MD lymphoma (Yao et al., 2009).
A particular kind of lincRNA called linc-SATB1 controls the MD resistance gene SATB1. It stimulates the growth and metastasis of cancer cells and has a positive correlation with SATB1 (He et al., 2015). In addition, SATB1 activates cell-mediated immunity and immune response genes to kill MDV-infected cells. SATB1 is also regulated by miR-21-5p, according to Zhao et al. (2018). Further research is necessary to fully understand the other roles that linc-satb1 plays.
LncRNA may interact with both host MDV susceptible and resistant genes to actually participate in MDV. According to Bacon (2002), model animals for the investigation of responsible biomarkers or clinical diagnosis during MDV infection are chicken MD resistant line 63 and chicken MD susceptible line 72. Chickens with and without MDV infection had linc-GALMD1, a DE lincRNA. The expression of immunoglobulin lambda-like polypeptide 1 (IGLL1) varies from line 63 to line 72. Following MDV infection, it expresses more in line 72 chickens than in line 63 chickens, suggesting that IGLL1 may be a line-specific or susceptible gene in response to MD (He et al., 2019).
2.5.3. Infectious Bursal Disease (Gumboro Disease):
Within the family Birnaviridae, the non-enveloped double-stranded RNA virus known as Infectious Bursal Disease Virus (IBDV) can cause acute, highly contagious, and immunosuppressive disease in chickens as early as 3–6 weeks of age. This can result in a significant economic increase in mortality as well as losses. The profound immunosuppression of broilers and egg-laying hens, as well as their heightened susceptibility to other illnesses and vaccination failure, are the primary causes of indirect losses in Gumboro disease (Sharma, 2000).
Studies have been conducted to analyse the effect the of lncRNAs on IBDV infection, during which the host innate immune system produces dendritic cells (DCs), which play a unique role in both the innate and acquired immune systems during virus infection (Banchereau and Steinman, 1998). A microarray study conducted by Lin et al. (2016) on DCs stimulated with IBDV and non-stimulated DCs showed that 965 mRNA, 18 miRNAs, and 114 lncRNAs were expressed differently after IBDV stimulation. They also came to the conclusion that functional annotation of the DE lncRNA genes revealed relationships with the RNA biosynthesis process, protein localization, cellular response to starvation, and other concepts. According to a pathway analysis, these were involved in the JAK- STAT/MAPK/mTOR/neurotrophin/CCR5/Interleukin-17 (IL-17) signalling pathways.
A study on the antiviral activity of long non-coding RNA loc107051710 during infectious bursal disease virus infection owing to increased interferon production was conducted by Huang and his colleagues in 2019. Following an investigation of the relationship between the loc107051710 and IRF8, type I IFNs, STATs, and ISGs, IBDV infection was induced in cultured DF-1 cells. RNA-seq was used to analyse the expression of mRNAs and lncRNAs in order to determine the antiviral activity of loc107051710. The findings demonstrated that loc107051710 positively controls IRF8 expression. Furthermore, the researchers discovered that the locus107051710 functions as a positive transcriptional regulator of the antiviral genes Mx1, PKR, STAT1, STAT2, IFN-α, and IFN-β.
2.5.4. Avian Influenza:
Over the past ten years, there has been a notable surge in lncRNA research, leading to an exploration of the critical roles played by lncRNAs in the regulation of influenza virus infection. Few lncRNAs have been found to interact with viral components, while the majority of lncRNAs have been found to regulate type I IFN signalling and IFN-stimulated genes (ISGs) (Wang and Cen, 2020). Taking an example is the identification and analysis of lncRNAs in duck (Anas platyrhynchos) in response to H5N1 influenza viruses was done by Lu et al. (2019). After examining the characteristics of 62,447 lncRNAs from zebrafish, chicken, human, mouse, and elegans, the researchers created a pipeline to identify lncRNAs by incorporating the features with transcriptomic data. They annotated 4094 duck lncRNAs using 151,970 assembled transcripts from RNA-Seq data of 21 individuals across three tissues. Results indicated that 619 lncRNAs and 3586 (87.6%) lncRNAs located in intergenic regions had differential expression in H5N1-infected ducks when compared to duck protein-coding transcripts. Researchers verified that eight lncRNAs, following H5N1 virus infection, exhibited remarkably different expression in vitro (in duck embryo fibroblast cells, DEF cells) and in vivo (in duck individuals), suggesting that lncRNAsmay have significant roles in the antiviral immune response to influenza A virus infection.
By blocking various stages of the host immune response, lncRNAs can also function as positive regulators of viral infection (Ma et al.,2016). One study by Ouyang and colleagues (2014) that demonstrated how lncRNA NRAV is significantly downregulated during IAV infection shows evidence. NRAV likely inhibits the initial transcription of various significant ISGs, such as MxA and IFITM3, by altering the histone modifications H3K4me3 and H3K27me3 in these genes. This, in turn, can promote IAV replication. Few such examples, with lncRNA acting as positive regulator for viral replication, are mentioned in the Table 1 (Wang and Cen, 2020).
2.5.5. Infectious Bronchitis:
The avian infectious bronchitis virus (IBV), a member of the Gamma-coronavirus genus, affects the kidneys, reproductive system, and upper respiratory tract of chickens (Cavanagh et al., 1997).
According to research published in 2019 by An and his co-workers, the coronavirus infectious bronchitis virus (IBV) causes virus-infected cells to produce a unique non-coding RNA (ncRNA). With the exception of a poly(A) tail from the 3′ untranslated region of the IBV genome and a 63-nt terminal leader sequence from the 5′ end of the viral genome, this ncRNA was composed of 563 nucleotides. This non-coding RNA was discovered to be a sub-genomic RNA produced by a discontinuous transcription mechanism, and it was revealed through the use of mutagenesis and reverse genetics techniques. In the IBV-infected cells, they have discovered the presence of a novel sgRNA that is primarily derived from the 3′ UTR of IBV. A truncated core sequence, 27104UAACA27108, which was identical to nucleotides 3–7 of the IBV core sequence, facilitated the synthesis of this sgRNA. which contributes at least three nucleotides (A27105, A27106, and C27107) to the efficient production of sgRNAs, further supporting the idea that non-canonical transcriptional signals are employed in the synthesis of coronavirus sgRNAs. Using mRNA, miRNA, and lncRNA microarray analysis, Lin et al. (2019) investigated the host responses against IBV. The study offered details on the molecular pathophysiology and interactions between the virus and the host, avian bone marrow-derived dendritic cells (BMDCs).
2.5.6. Newcastle Disease:
The differentially expressed genes and lncRNAs during NDV challenge were discovered by Vanamamalai and his colleagues in 2023, and these findings may have some bearing on the observed differential resistance pattern. It was discovered that there were 1580 lncRNAs and 552 genes with differential expression. 52 annotated genes and a greater number of positively correlating lncRNAs were found to be downregulated, according to pathway analysis and gene ontology.
Sha et al. (2024) utilized RNA sequencing to explore the transcriptional profiles of the visceral tissues of chicken embryos (CEVTs) subjected to infection by either the virulent NA-1 strain or the avirulent LaSota strain, assessing samples at 24 and 36 hours post-infection. The analysis of long non-coding RNAs (lncRNAs) indicated that the significant pathological alterations and clinical manifestations resulting from virulent Newcastle disease virus (NDV) infection could be, in part, linked to associated target genes that are modulated by differentially expressed lncRNAs, including MSTRG.1545.5, MSTRG.14601.6, MSTRG.7150.1, and MSTRG.4481.1. The findings reveal that virulent NDV infection capitalizes on the host's metabolic resources and modifies the host's metabolic functions, which is associated with a heightened activation of the immune response.
Conclusion
One of the spin-offs of the ENCODE project is the identification of non-coding RNAs, especially lncRNAs. Since then, several lncRNAs have been reported as the regulators of gene expressions, viral diseases, cancer development, and many more pathological conditions. The abovementioned discussion on lncNRAs in chicken viral diseases has provided an assessment of the current state of knowledge regarding their locations, functions, and mechanisms. Host lncRNAs are considered to have a role in protecting the host from viral invasion. On the other hand, the viruses utilize some lncRNAs to infect and replicate itself. When combined, these results have strongly suggested that lncRNAs play a crucial role in viral pathogenicity and function as important regulators of host–virus interaction. Although the reported lncRNAs in conjunction with the chicken viral diseases are less it is evident that these lncRNAs have been involved in the regulation of host antiviral responses. These findings pave the way for using the serum from the chickens to use it as a diagnostic tool to assess the viral disease progression in a flock. Moreover, the role of lncRNAs in modulating the host's innate immune response to viral infections may lead to future research toward developing tools such as genome editing to increase host disease resistance. LncRNAs have a lot of potential for usage as therapeutic targets and indicators to achieve an antiviral state because of their proviral and antiviral properties. Hence, more evidence on lncRNAs in regulating viral diseases may help to identify potential disease diagnostic tools and disease control strategies in the future.
References
- Funk, A.; Truong, K.; Nagasaki, T.; Torres, S.; Floden, N.; Balmori Melian, E.; Edmonds, J.; Dong, H.; Shi, P.-Y.; Khromykh, A.A. RNA Structures Required for Production of Subgenomic Flavivirus RNA. J. Virol. 2010, 84, 11407–11417. [Google Scholar] [CrossRef] [PubMed]
- Altfeld, M.; Gale, M Jr. Innate immunity against HIV-1 infection. Nat Immunol. 2015, 16, 554–562. [Google Scholar] [CrossRef] [PubMed]
- An, H.; Cai, Z.; Yang, Y.; Wang, Z.; Liu, D.X.; Fang, S. Identification and formation mechanism of a novel noncoding RNA produced by avian infectious bronchitis virus. Virology 2019, 528, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Bacon, L.D. The National Registry of Genetically Unique Animal Populations: USDA-ADOL Chicken Genetic Lines. National Animal Germplasm Program: East Lansing, MI, USA,2002.
- Bamunuarachchi, G.; Pushparaj, S.; Liu, L. Interplay between host non-coding RNAs and influenza viruses. RNA Biol. 2021, 18, 767–784. [Google Scholar] [CrossRef]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular condensates: organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Baniushin, B.F. Methylation of adenine residues in DNA of eukaryotes. Mol. Biol. 2005, 39, 557–66. [Google Scholar]
- Bao, X.; Wu, H.; Zhu, X.; Guo, X.; Hutchins, A.P.; Luo, Z.; Song, H.; Chen, Y.; Lai, K.; Yin, M.; et al. The p53-induced lincRNA-p21 derails somatic cell reprogramming by sustaining H3K9me3 and CpG methylation at pluripotency gene promoters. Cell Res. 2014, 25, 80–92. [Google Scholar] [CrossRef]
- Beermann, J.; Piccoli, M.-T.; Viereck, J.; Thum, T. Non-coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef]
- Beltran, M.; Puig, I.; Pena, C.; Garcia, J.M.; Alvarez, A.B.; Pena, R.; Bonilla, F.; de Herreros, A.G. A natural antisense transcript regulates Zeb2/Sip1 gene expression during Snail1-induced epithelial-mesenchymal transition. Genes Dev. 2008, 22, 756–769. [Google Scholar] [CrossRef]
- Bernstein, B.E.; Kamal, M.; Lindblad-Toh, K.; Bekiranov, S.; Bailey, D.K.; Huebert, D.J.; McMahon, S.; Karlsson, E.K.; Kulbokas, E.J.; Gingeras, T.R.; et al. Genomic Maps and Comparative Analysis of Histone Modifications in Human and Mouse. Cell 2005, 120, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Bonetti, A.; et al. RADICL-seq identifies general and cell type-specific principles of genome-wideRNA–chromatin interactions. Nat. Commun. 2020, 11, 1018. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.J.; Hendrich, B.D.; Rupert, J.L.; Lafrenière, R.G.; Xing, Y.; Lawrence, J.; Willard, H.F. The human XIST gene: Analysis of a 17 kb inactive X-specific RNA that contains conserved repeats and is highly localized within the nucleus. Cell 1992, 71, 527–542. [Google Scholar] [CrossRef] [PubMed]
- Burnside, J.; Ouyang, M.; Anderson, A.; Bernberg, E.; Lu, C.; Meyers, B.C.; Green, P.J.; Markis, M.; Isaacs, G.; Huang, E.; et al. Deep Sequencing of Chicken microRNAs. BMC Genom. 2008, 9, 185–185. [Google Scholar] [CrossRef]
- Braconi, C.; Kogure, T.; Valeri, N.; Huang, N.; Nuovo, G.; Costinean, S.; Negrini, M.; Miotto, E.; Croce, C.M.; Patel, T. microRNA-29 can regulate expression of the long non-coding RNA gene MEG3 in hepatocellular cancer. Oncogene 2011, 30, 4750–4756. [Google Scholar] [CrossRef]
- Rossetto, C.C.; Pari, G. KSHV PAN RNA Associates with Demethylases UTX and JMJD3 to Activate Lytic Replication through a Physical Interaction with the Virus Genome. PLOS Pathog. 2012, 8, e1002680. [Google Scholar] [CrossRef]
- Rossetto, C.C.; Tarrant-Elorza, M.; Verma, S.; Purushothaman, P.; Pari, G.S. Regulation of viral and cellular gene expression by Kaposi's sarcoma-associated herpesvirus polyadenylated nuclear RNA. J. Virol. 2013, 87, 5540e5553. [Google Scholar] [CrossRef]
- Cadena, C.; Hur, S. Antiviral Immunity and Circular RNA: No End in Sight. Mol. Cell 2017, 67, 163–164. [Google Scholar] [CrossRef]
- Carnero, E.; Barriocanal, M.; Prior, C.; Pablo Unfried, J.; Segura, V.; Guruceaga, E.; Enguita, M.; Smerdou, C.; Gastaminza, P.; Fortes, P. Long noncoding RNA EGOT negatively affects the antiviral response and favors HCV replication. EMBO Rep. 2016, 17, 1013–1028. [Google Scholar] [CrossRef]
- Carninci, P.; Kasukawa, T.; Katayama, S.; Gough, J.; Frith, M.C.; Maeda, N.; Oyama, R.; Ravasi, T.; Lenhard, B.; Wells, C.; et al. The Transcriptional Landscape of the Mammalian Genome. Science 2005, 309, 1559–1563. [Google Scholar] [CrossRef]
- Cavanagh, D.; Elus, M. M.; Cook, J. K. A. Relationship between sequence variation in the S1 spike protein of infectious bronchitis virus and the extent of cross-protection in vivo. Avian pathology 1997, 26, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Charley, P.A.; Wilusz, J. Standing your ground to exoribonucleases: Function of Flavivirus long non-coding RNAs. Virus Res. 2015, 212, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, R.; Singh, A.; Singh, P.K.; Teja, E.S.; Varshney, R. Dynamics of Marek’s disease in poultry industry. Pharma Innov. 2021, 10, 80–83. [Google Scholar] [CrossRef]
- Chaumeil, J.; Le Baccon, P.; Wutz, A.; Heard, E. A novel role for Xist RNA in the formation of a repressive nuclear compartment into which genes are recruited when silenced. Genes Dev. 2006, 20, 2223–2237. [Google Scholar] [CrossRef]
- Chen, Y.G.; Kim, M.V.; Chen, X.; Batista, P.J.; Aoyama, S.; Wilusz, J.E.; Iwasaki, A.; Chang, H.Y. Sensing Self and Foreign Circular RNAs by Intron Identity. Mol. Cell 2017, 67, 228–238. [Google Scholar] [CrossRef]
- Choudhury, N.R.; Heikel, G.; Michlewski, G. TRIM25 and its emerging RNA-binding roles in antiviral defense. Wiley Interdiscip. Rev. RNA 2020, 11, e1588. [Google Scholar] [CrossRef]
- Chu, C.; Qu, K.; Zhong, F.L.; Artandi, S.E.; Chang, H.Y. Genomic Maps of Long Noncoding RNA Occupancy Reveal Principles of RNA-Chromatin Interactions. Mol. Cell 2011, 44, 667–678. [Google Scholar] [CrossRef]
- Coccia, E.M.; Battistini, A. Early IFN type I response: Learning from microbial evasion strategies. Semin. Immunol. 2015, 27, 85–101. [Google Scholar] [CrossRef]
- Dai, M.; Feng, M.; Xie, T.; Zhang, X. Long non-coding RNA and MicroRNA profiling provides comprehensive insight into non-coding RNA involved host immune responses in ALV-J-infected chicken primary macrophage. Dev. Comp. Immunol. 2019, 100, 103414. [Google Scholar] [CrossRef]
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2008, 154, 171–177. [Google Scholar] [CrossRef]
- Doerr, A. Predicting PPIs. Nature Methods 2012, 9, 1139. [Google Scholar] [CrossRef] [PubMed]
- Doyle, T.; Goujon, C.; Malim, M.H. HIV-1 and interferons: who's interfering with whom? Nat Rev Microbiol. 2015, 13, 13–403. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Bata-Csorgo, Z.; Pivarcsi, A.; Polyanka, H.; Kenderessy-Szabo, A.; Molnar, G.; Szentpali, K.; Bari, L.; Megyeri, K.; Mandi, Y.; et al. Identification and Characterization of a Novel, Psoriasis Susceptibility-related Noncoding RNA gene, PRINS. J. Biol. Chem. 2005, 280, 24159–24167. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, T.; Boumart, I.; Coupeau, D.; Rasschaert, D. Hyperediting by ADAR1 of a new herpesvirus lncRNA during the lytic phase of the oncogenic Marek’s disease virus. J. Gen. Virol. 2016, 97, 2973–2988. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Delgado, S.; Gas, M.-E.; Carbonell, A.; Molina, D.; Gago, S.; De la Peña, M. Viroids: the minimal non-coding RNAs with autonomous replication. FEBS Lett. 2004, 567, 42–48. [Google Scholar] [CrossRef]
- Fortes, P.; Morris, K.V. Long noncoding RNAs in viral infections. Virus Res. 2016, 212, 1–11. [Google Scholar] [CrossRef]
- Pijlman, G.P.; Funk, A.; Kondratieva, N.; Leung, J.; Torres, S.; van der Aa, L.; Liu, W.J.; Palmenberg, A.C.; Shi, P.-Y.; Hall, R.A.; et al. A Highly Structured, Nuclease-Resistant, Noncoding RNA Produced by Flaviviruses Is Required for Pathogenicity. Cell Host Microbe 2008, 4, 579–591. [Google Scholar] [CrossRef]
- Garcia-Moreno,M. ;Jarvelin, A.I.; Castello, A. Unconventional RNA-binding proteins step into the virus-host battlefront. WIREs RNA 2018, 9, e1498. [Google Scholar] [CrossRef]
- Gebert, C.; Kunkel, D.; Grinberg, A.; Pfeifer, K. H19 Imprinting Control Region Methylation Requires an Imprinted Environment Only in the Male Germ Line. Mol. Cell. Biol. 2010, 30, 1108–1115. [Google Scholar] [CrossRef]
- Grote, P.; Wittler, L.; Hendrix, D.; Koch, F.; Währisch, S.; Beisaw, A.; Macura, K.; Bläss, G.; Kellis, M.; Werber, M.; et al. The Tissue-Specific lncRNA Fendrr Is an Essential Regulator of Heart and Body Wall Development in the Mouse. Dev. Cell 2013, 24, 206–214. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Russell, P.; Ingolia, N.T.; Weissman, J.S.; Lander, E.S. Ribosome Profiling Provides Evidence that Large Noncoding RNAs Do Not Encode Proteins. Cell 2013, 154, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; He, Y.; Zhang, L.; Ding, Y.; Lian, L.; Zhao, C.; Song, J.; Yang, N. Long intergenic non-coding RNA GALMD3 in chicken Marek’s disease. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hartford, C.C.R.; Lal, A. When Long Noncoding Becomes Protein Coding. Mol. Cell. Biol. 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Li, M.; Wei, P.; Mo, M.-L.; Wei, T.-C.; Li, K.-R. Complete Genome Sequence of an Infectious Bronchitis Virus Chimera between Cocirculating Heterotypic Strains. J. Virol. 2012, 86, 13887–13888. [Google Scholar] [CrossRef]
- He, Y.; Han, B.; Ding, Y.; Zhang, H.; Chang, S.; Zhang, L.; Zhao, C.; Yang, N.; Song, J. Linc-GALMD1 Regulates Viral Gene Expression in the Chicken. Front. Genet. 2019, 10, 1122. [Google Scholar] [CrossRef]
- Heintzman, N.D.; Stuart, R.K.; Hon, G.; Fu, Y.; Ching, C.W.; Hawkins, R.D.; Barrera, L.O.; Van Calcar, S.; Qu, C.; Ching, K.A.; et al. Distinct and predictive chromatin signatures of transcriptional promoters and enhancers in the human genome. Nat. Genet. 2007, 39, 311–318. [Google Scholar] [CrossRef]
- Hentze, M.W.; Castello, A.; Schwarzl, T.; Preiss, T. A brave new world of RNA-binding proteins. Nat. Rev. Mol. Cell Biol. 2018, 19, 327–341. [Google Scholar] [CrossRef]
- Hezroni, H.; Koppstein, D.; Schwartz, M.G.; Avrutin, A.; Bartel, D.P.; Ulitsky, I. Principles of Long Noncoding RNA Evolution Derived from Direct Comparison of Transcriptomes in 17 Species. Cell Rep. 2015, 11, 1110–1122. [Google Scholar] [CrossRef]
- Hu, X.; Chen, S.; Jia, C.; Xue, S.; Dou, C.; Dai, Z.; Xu, H.; Sun, Z.; Geng, T.; Cui, H. Gene expression profile and long non-coding RNA analysis, using RNA-Seq, in chicken embryonic fibroblast cells infected by avian leukosis virus J. Arch. Virol. 2017, 163, 639–647. [Google Scholar] [CrossRef]
- Huang, X.; Xu, Y.; Lin, Q.; Guo, W.; Zhao, D.; Wang, C.; Wang, L.; Zhou, H.; Jiang, Y.; Cui, W.; et al. Determination of antiviral action of long non-coding RNA loc107051710 during infectious bursal disease virus infection due to enhancement of interferon production. Virulence 2019, 11, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Ilott, N.E.; Ponting, C.P. Predicting long non-coding RNAs using RNA sequencing. Methods 2013, 63, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.; Kroehling, L.; Khitun, A.; Bailis, W.; Jarret, A.; York, A.G.; Khan, O.M.; Brewer, J.R.; Skadow, M.H.; Duizer, C.; et al. The translation of non-canonical open reading frames controls mucosal immunity. Nature 2018, 564, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, S.; Yang, Z.; Lin, H.; Zhu, J.; Liu, L.; Wang, W.; Liu, S.; Liu, W.; Ma, Y.; et al. Self-Recognition of an Inducible Host lncRNA by RIG-I Feedback Restricts Innate Immune Response. Cell 2018, 173, 906–919. [Google Scholar] [CrossRef]
- Kanaujia, R.; Bora, I.; Ratho, R.K.; Thakur, V.; Mohi, G.K.; Thakur, P. Avian influenza revisited: concerns and constraints. VirusDisease 2022, 33, 456–465. [Google Scholar] [CrossRef]
- Kennedy, D.A.; Cairns, C.; Jones, M.J.; Bell,A. S.;Salathé, R.M.; Baigent, S.J. Industry-wide surveillance of Marek's disease virus on commercial poultry farms. Avian diseases 2017, 61, 153–164. [Google Scholar] [CrossRef]
- Kotzin, J.J.; Iseka, F.; Wright, J.; Basavappa, M.G.; Clark, M.L.; Ali, M.-A.; Abdel-Hakeem, M.S.; Robertson, T.F.; Mowel, W.K.; Joannas, L.; et al. The long noncoding RNA Morrbid regulates CD8 T cells in response to viral infection. Proc. Natl. Acad. Sci. 2019, 116, 11916–11925. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Lan, X.; Wang, Y.; Tian, K.; Ye, F.; Yin, H.; Zhao, X.; Xu, H.; Huang, Y.; Liu, H.; Hsieh, J.C.F.; et al. Integrated host and viral transcriptome analyses reveal pathology and inflammatory response mechanisms to ALV-J injection in SPF chickens. Sci. Rep. 2017, 7, 46156. [Google Scholar] [CrossRef]
- Latos, P.A.; Pauler, F.M.; Koerner, M.V.; ¸Senergin, H.B.; Hudson, Q.; Stocsits, R.R.; Allhoff, W.; Stricker, S.H.; Klement, R.M.; Warczok, K.E.; et al. Airn Transcriptional Overlap, But Not Its lncRNA Products, Induces Imprinted Igf2r Silencing. Science 2012, 338, 1469–1472. [Google Scholar] [CrossRef]
- Lee, S.; Kopp, F.; Chang, T.-C.; Sataluri, A.; Chen, B.; Sivakumar, S.; Yu, H.; Xie, Y.; Mendell, J.T. Noncoding RNA NORAD Regulates Genomic Stability by Sequestering PUMILIO Proteins. Cell 2015, 164, 69–80. [Google Scholar] [CrossRef]
- Lee, J.T. Epigenetic Regulation by Long Noncoding RNAs. Science 2012, 338, 1435–1439. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Liu, S.; Zheng, L.L.; Wu, J.; Sun, W.J.; Wang, Z.L.; et al. Discovery of Protein-lncRNA Interactions by Integrating Large-Scale CLIP-Seq and RNA-Seq Datasets. Front BioengBiotechnol. 2014, 2, 88. [Google Scholar] [CrossRef]
- Li, Z.; Chao, T.C.; Chang, K.Y.; et al. The long noncoding RNA THRIL regulates TNFalpha expression through its interaction with hnRNPL. Proc Natl Acad Sci U S A 2014, 111, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, M.; Wang, X.; Sun, M.; Ma, C.; Liang, W.; Gao, X.; Wei, L. Long Noncoding RNA NRAV Promotes Respiratory Syncytial Virus Replication by Targeting the MicroRNA miR-509-3p/Rab5c Axis To Regulate Vesicle Transportation. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Li, X.; Zhou, B.; Chen, L.; Gou, L.-T.; Li, H.; Fu, X.-D. GRID-seq reveals the global RNA–chromatin interactome. Nat. Biotechnol. 2017, 35, 940–950. [Google Scholar] [CrossRef]
- Lin, H.; Jiang, M.; Liu, L.; Yang, Z.; Ma, Z.; Liu, S.; Ma, Y.; Zhang, L.; Cao, X. The long noncoding RNA Lnczc3h7a promotes a TRIM25-mediated RIG-I antiviral innate immune response. Nat. Immunol. 2019, 20, 812–823. [Google Scholar] [CrossRef]
- Lin, J.; Xia, J.; Zhang, K.; Yang, Q. Genome-wide profiling of chicken dendritic cell response to infectious bursal disease. BMC Genom. 2016, 17, 1–12. [Google Scholar] [CrossRef]
- Liu, W.; Ding, C. Roles of LncRNAs in Viral Infections. Front. Cell. Infect. Microbiol. 2017, 7, 205. [Google Scholar] [CrossRef]
- Lu, C.; Xing, Y.; Cai, H.; Shi, Y.; Liu, J.; Huang, Y. Identification and analysis of long non-coding RNAs in response to H5N1 influenza viruses in duck (Anas platyrhynchos). BMC Genom. 2019, 20, 36. [Google Scholar] [CrossRef]
- Luo, S.; Lu, J.Y.; Liu, L.; Yin, Y.; Chen, C.; Han, X.; Wu, B.; Xu, R.; Liu, W.; Yan, P.; et al. Divergent lncRNAs Regulate Gene Expression and Lineage Differentiation in Pluripotent Cells. Cell Stem Cell 2016, 18, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Ahanda, M.-L.E.; Ruby, T.; Wittzell, H.; Bed’hom, B.; Chaussé, A.-M.; Morin, V.; Oudin, A.; Chevalier, C.; Young, J.R.; Zoorob, R. Non-coding RNAs revealed during identification of genes involved in chicken immune responses. Immunogenetics 2008, 61, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Han, P.; Ye, W.; et al. The long noncoding RNA NEAT1 exerts anti-hantaviral effects by acting as positive feedback for RIG-I signaling. J Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ouyang, J.; Wei, J.; Maarouf, M.; Chen, J.-L. Involvement of Host Non-Coding RNAs in the Pathogenesis of the Influenza Virus. Int. J. Mol. Sci. 2016, 18, 39. [Google Scholar] [CrossRef]
- Martens, J.A.; Laprade, L.; Winston, F. Intergenic transcription is required to repress the Saccharomyces cerevisiae SER3 gene. Nature 2004, 429, 571–574. [Google Scholar] [CrossRef]
- Martianov, I.; Ramadass, A.; Barros, A.S.; Chow, N.; Akoulitchev, A. Repression of the human dihydrofolate reductase gene by a non-coding interfering transcript. Nature 2007, 445, 666–670. [Google Scholar] [CrossRef]
- McFerran, J. B. "Infectious bursal disease." 1993, 51-56.
- Melé, M.; et al. Chromatin environment, transcriptional regulation, and splicing distinguishlincRNAs and mRNAs. Genome Res. 2017, 27, 27–37. [Google Scholar] [CrossRef]
- Mohandas, S.S.; Thakur, M.; Gangil, R. A Mini-Review on Immunosuppressive Viral Diseases and their Containment Methods at Commercial Poultry Farms. Veterinary Immunology & Biotechnology 2018, 1, 31. [Google Scholar]
- Mondal, T.; Subhash, S.; Vaid, R.; Enroth, S.; Uday, S.; Reinius, B.; Mitra, S.; Mohammed, A.; James, A.R.; Hoberg, E.; et al. MEG3 long noncoding RNA regulates the TGF-β pathway genes through formation of RNA–DNA triplex structures. Nat. Commun. 2015, 6, 7743. [Google Scholar] [CrossRef]
- Moon, S.L.; Anderson, J.R.; Kumagai, Y.; Wilusz, C.J.; Akira, S.; Khromykh, A.A.; Wilusz, J. A noncoding RNA produced by arthropod-borne flaviviruses inhibits the cellular exoribonuclease XRN1 and alters host mRNA stability. RNA 2012, 18, 2029–2040. [Google Scholar] [CrossRef] [PubMed]
- Niehrs, C.; Luke, B. Regulatory R-loops as facilitators of gene expression and genome stability. Nat. Rev. Mol. Cell Biol. 21, 167–178 (2020). Tan-Wong, S. M., Dhir, S. & Proudfoot, N. J. R-loops promote antisense transcription across the mammalian genome. Mol. Cell. 2016, 76, 600–616. [Google Scholar] [CrossRef]
- O’Leary, V.B.; Ovsepian, S.V.; Carrascosa, L.G.; Buske, F.A.; Radulovi´c, V.; Niyazi, M.; Mörtl, S.; Trau, M.; Atkinson, M.J.; Anastasov, N. PARTICLE, a triplex-forming long ncRNA, regulates locus-specific methylation in response to low-dose Irradiation. Cell Rep. 2015, 11, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Hu, J.; Chen, J. lncRNAs regulate the innate immune response to viral infection. Wiley Interdiscip. Rev. RNA 2015, 7, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Ouyang J, Zhu X, Chen Y, et al. NRAV, a long noncoding RNA, modulates antiviral responsesthrough suppression of interferon-stimulated gene transcription. Cell Host Microbe 2014, 16, 616–626. [Google Scholar] [CrossRef]
- Pauli, A.; Valen, E.; Lin, M.F.; Garber, M.; Vastenhouw, N.L.; Levin, J.Z.; Fan, L.; Sandelin, A.; Rinn, J.L.; Regev, A.; et al. Systematic identification of long noncoding RNAs expressed during zebrafish embryogenesis. Genome Res. 2011, 22, 577–591. [Google Scholar] [CrossRef]
- Prasad, A.; Prasad, M. Host-virus interactions mediated by long non-coding RNAs. Virus Res. 2021, 298, 198402. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, C.-Y.; Yedavalli, V.S.R.K.; Jeang, K.-T. NEAT1 Long Noncoding RNA and Paraspeckle Bodies Modulate HIV-1 Posttranscriptional Expression. mBio 2013, 4, e00596–12. [Google Scholar] [CrossRef]
- Qiu, L.; Chang, G.; Li, Z.; Bi, Y.; Liu, X.; Chen, G. Comprehensive Transcriptome Analysis Reveals Competing Endogenous RNA Networks During Avian Leukosis Virus, Subgroup J-Induced Tumorigenesis in Chickens. Front. Physiol. 2018, 9, 996. [Google Scholar] [CrossRef]
- Redon, S.; Reichenbach, P.; Lingner, J. The non-coding RNA TERRA is a natural ligand and direct inhibitor of human telomerase. Nucleic Acids Res. 2010, 38, 5797–5806. [Google Scholar] [CrossRef]
- Rigby, R.E.; Rehwinkel, J. RNA degradation in antiviral immunity and autoimmunity. Trends Immunol. 2015, 36, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional Demarcation of Active and Silent Chromatin Domains in Human HOX Loci by Noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Rom, A.; et al. Regulation of CHD2 expression by the Chaserrlong noncoding RNA gene is essential for viability. Nat. Commun. 2019, 10, 5092. [Google Scholar] [CrossRef] [PubMed]
- Saldaña-Meyer, R.; Rodriguez-Hernaez, J.; Escobar, T.; Nishana, M.; Jácome-López, K.; Nora, E.P.; Bruneau, B.G.; Tsirigos, A.; Furlan-Magaril, M.; Skok, J.; et al. RNA Interactions Are Essential for CTCF-Mediated Genome Organization. Mol. Cell 2019, 76, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Salerno, D.; Chiodo, L.; Alfano, V.; Floriot, O.; Cottone, G.; Paturel, A.; Pallocca, M.; Plissonnier, M.-L.; Jeddari, S.; Belloni, L.; et al. Hepatitis B protein HBx binds the DLEU2 lncRNA to sustain cccDNA and host cancer-related gene transcription. Gut 2020, 69, 2016–2024. [Google Scholar] [CrossRef]
- Schmitt, A.M.; Chang, H.Y. Long Noncoding RNAs in Cancer Pathways. Cancer Cell 2016, 29, 452–463. [Google Scholar] [CrossRef]
- Schmitz, K.-M.; Mayer, C.; Postepska, A.; Grummt, I. Interaction of noncoding RNA with the rDNA promoter mediates recruitment of DNMT3b and silencing of rRNA genes. Genes Dev. 2010, 24, 2264–2269. [Google Scholar] [CrossRef]
- Schultheiss,M. ;Thimme, R. Toll like receptor 7 and hepatitis C virus infection. J Hepatol. 2007, 47, 165–167. [Google Scholar] [CrossRef]
- Seila, A.C.; Calabrese, J.M.; Levine, S.S.; Yeo, G.W.; Rahl, P.B.; Flynn, R.A.; Young, R.A.; Sharp, P.A. Divergent Transcription from Active Promoters. Science 2008, 322, 1849–1851. [Google Scholar] [CrossRef]
- Sha, Y.; Liu, X.; Yan, W.; Wang, M.; Li, H.; Jiang, S.; Wang, S.; Ren, Y.; Zhang, K.; Yin, R. Long Non-Coding RNA Analysis: Severe Pathogenicity in Chicken Embryonic Visceral Tissues Infected with Highly Virulent Newcastle Disease Virus—A Comparison to the Avirulent Vaccine Virus. Microorganisms 2024, 12, 971. [Google Scholar] [CrossRef]
- Sharma, J.M.; Kim, I.-J.; Rautenschlein, S.; Yeh, H.-Y. Infectious bursal disease virus of chickens: pathogenesis and immunosuppression. Dev. Comp. Immunol. 2000, 24, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Shirahama, S.; Miki, A.; Kaburaki, T.; Akimitsu, N. Long Non-coding RNAs Involved in Pathogenic Infection. Front. Genet. 2020, 11, 454. [Google Scholar] [CrossRef] [PubMed]
- Stojic, L.; Niemczyk, M.; Orjalo, A.; Ito, Y.; Ruijter, A.E.M.; Uribe-Lewis, S.; Joseph, N.; Weston, S.; Menon, S.; Odom, D.T.; et al. Transcriptional silencing of long noncoding RNA GNG12-AS1 uncouples its transcriptional and product-related functions. Nat. Commun. 2016, 7, 10406. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, S.; Nair, A.J. Marek's disease: The never ending challenge - A review. International Journal of Pharma and Bio Sciences 2013, 4, B6–B11. [Google Scholar]
- Tahira, A.C.; Kubrusly, M.S.; Faria, M.F.; Dazzani, B.; Fonseca, R.S.; Maracaja-Coutinho, V.; Verjovski-Almeida, S.; Machado, M.C.; Reis, E.M. Long noncoding intronic RNAs are differentially expressed in primary and metastatic pancreatic cancer. Mol. Cancer 2011, 10, 141–141. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Innate immunity to virus infection. Immunol. Rev. 2009, 227, 75–86. [Google Scholar] [CrossRef]
- Thebault, P.; Boutin, G.; Bhat, W.; Rufiange, A.; Martens, J.; Nourani, A. Transcription Regulation by the Noncoding RNA SRG1 Requires Spt2-Dependent Chromatin Deposition in the Wake of RNA Polymerase II. Mol. Cell. Biol. 2011, 31, 1288–1300. [Google Scholar] [CrossRef]
- Tichon, A.; Perry, R.B.-T.; Stojic, L.; Ulitsky, I. SAM68 is required for regulation of Pumilio by the NORAD long noncoding RNA. Genes Dev. 2018, 32, 70–78. [Google Scholar] [CrossRef]
- Tilgner, H.; Knowles, D.G.; Johnson, R.; Davis, C.A.; Chakrabortty, S.; Djebali, S.; Curado, J.; Snyder, M.; Gingeras, T.R.; Guigó, R. Deep sequencing of subcellular RNA fractions shows splicing to be predominantly co-transcriptional in the human genome but inefficient for lncRNAs. Genome Res. 2012, 22, 1616–1625. [Google Scholar] [CrossRef]
- To, K.K.; Chan, J.F.; Chen, H.; Li, L.; Yuen, K.-Y. The emergence of influenza A H7N9 in human beings 16 years after influenza A H5N1: a tale of two cities. Lancet Infect. Dis. 2013, 13, 809–821. [Google Scholar] [CrossRef]
- Lisnic, V.J.; Cac, M.B.; Lisnic, B.; Trsan, T.; Mefferd, A.; Das Mukhopadhyay, C.; Cook, C.H.; Jonjic, S.; Trgovcich, J. Dual Analysis of the Murine Cytomegalovirus and Host Cell Transcriptomes Reveal New Aspects of the Virus-Host Cell Interface. PLOS Pathog. 2013, 9, e1003611. [Google Scholar] [CrossRef] [PubMed]
- van Berg, T.P.D. Acute infectious bursal disease in poultry: A review. Avian Pathol. 2000, 29, 175–194. [Google Scholar] [CrossRef] [PubMed]
- van Werven, F.J.; Neuert, G.; Hendrick, N.; Lardenois, A.; Buratowski, S.; van Oudenaarden, A.; Primig, M.; Amon, A. Transcription of Two Long Noncoding RNAs Mediates Mating-Type Control of Gametogenesis in Budding Yeast. Cell 2012, 150, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Velloso, L.A.; Folli, F.; Saad, M.J. TLR4 at the Crossroads of Nutrients, Gut Microbiota, and Metabolic Inflammation. Endocr. Rev. 2015, 36, 245–271. [Google Scholar] [CrossRef]
- Tam, W.; Ben-Yehuda, D.; Hayward, W.S. bic, a Novel Gene Activated by Proviral Insertions in Avian Leukosis Virus-Induced Lymphomas, Is Likely To Function through Its Noncoding RNA. Mol. Cell. Biol. 1997, 17, 1490–1502. [Google Scholar] [CrossRef]
- Wagari, A. A review on infectious bursal disease in poultry. Health Econ. Outcome Res. Open Access, 2021, 7, 167. [Google Scholar]
- Wang, A.; Bao, Y.; Wu, Z.; et al. Long noncoding RNA EGFR-AS1 promotes cell growth and metastasis via affecting HuR mediated mRNA stability of EGFR in renal cancer. Cell Death Dis. 2019, 10, 154. [Google Scholar] [CrossRef]
- Wang, J.; Cen, S. Roles of lncRNAs in influenza virus infection. Emerg Microbes Infect. 2020, 9, 1–22. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Li, Q.; et al. Influenza virus exploits an interferon-independent lncRNA to preserve viral RNA synthesis through stabilizing viral RNA polymerase PB1. Cell Rep. 2019, 27, 3295–3304. [Google Scholar] [CrossRef]
- Wang, K.; Liu, F.; Liu, C.Y.; et al. The long noncoding RNA NRF regulates programmed necrosis and myocardial injury during ischemia and reperfusion by targeting miR-873. Cell Death Differ. 2016, 23, 1394–1405. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, D.; Feng, W.; et al. Long noncoding RNA TSPOAP1 antisense RNA 1 negatively modulates type I IFN signaling to facilitate influenza A virus replication. J Med Virol. 2019, 94, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; et al. Human Bocavirus 1 Is a Novel Helper for Adeno-associated Virus Replication. J Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Zhou, R.; Zhao, J.; Zhang, Y.; Yi, D.; Li, Q.; Zhou, J.; Guo, F.; Liang, C.; et al. Host Long Noncoding RNA lncRNA-PAAN Regulates the Replication of Influenza A Virus. Viruses 2018, 10, 330. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gong, C.; Maquat, L.E. Control of myogenesis by rodent SINE-containing lncRNAs. Genes Dev. 2013, 27, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell. 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Wang, P. The Opening of Pandora’s Box: An Emerging Role of Long Noncoding RNA in Viral Infections. Frontiers in Immunology 2019, 9, 3138. [Google Scholar] [CrossRef]
- Wang, P.; Xu, J.; Wang, Y.; Cao, X. An interferon-independent lncRNA promotes viral replication by modulating cellular metabolism. Science 2017, 358, 1051–1055. [Google Scholar] [CrossRef]
- Wang, X.; McLachlan, J.; Zamore, P. D.; Hall, T. M. Modular recognition of RNA by a human Pumiliohomology domain. Cell 2002, 110, 501–512. [Google Scholar] [CrossRef]
- Wilusz, J.E.; Sunwoo, H.; Spector, D. Long noncoding RNAs: Functional surprises from the RNA world. Genes Dev. 2009, 23, 1494–1504. [Google Scholar] [CrossRef]
- Winterling, C.; Koch, M.; Koeppel, M.; Garcia-Alcalde, F.; Karlas, A.; Meyer, T.F. Evidence for a crucial role of a host non-coding RNA in influenza A virus replication. RNA Biol. 2013, 11, 66–75. [Google Scholar] [CrossRef]
- Peng, X.; Gralinski, L.; Armour, C.D.; Ferris, M.T.; Thomas, M.J.; Proll, S.; Bradel-Tretheway, B.G.; Korth, M.J.; Castle, J.C.; Biery, M.C.; et al. Unique Signatures of Long Noncoding RNA Expression in Response to Virus Infection and Altered Innate Immune Signaling. mBio 2010, 1, e00206–10. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.F.; Yin, Q.F.; Chen, T.; et al. Human colorectal cancer-specific CCAT1-L lncRNA regulates long-range chromatin interactions at the MYC locus. Cell Res. 2014, 24, 513–531. [Google Scholar] [CrossRef] [PubMed]
- Xu,Y,; Zhong, J. Innate immunity against hepatitis C virus. Curr Opin Immunol. 2016, 42, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Yao,R. W,; Wang, Y,; Chen, L.L. Cellular functions of long noncoding RNAs. Nat Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Zhao, Y.; Smith, L.P.; Lawrie, C.H.; Saunders, N.J.; Watson, M.; Nair, V. Differential expression of microRNAs in Marek’s disease virus-transformed T-lymphoma cell lines. J. Gen. Virol. 2009, 90, 1551–1559. [Google Scholar] [CrossRef]
- Yap, K.; et al. A short tandem repeat-enriched RNA assembles a nuclear compartment to control alternative splicing and promote cell survival. Mol. Cell. 2013, 72, 525–540. [Google Scholar] [CrossRef]
- Zeng, W.; Chu, Q.; Xu, T. The long noncoding RNA NARL regulates immune responses via microRNA-mediated NOD1 downregulation in teleost fish. J. Biol. Chem. 2021, 296, 100414. [Google Scholar]
- Zhang,S. Y.;Jouanguy, E.; Sancho-Shimizu, V.et al. Human Toll-like receptor-dependent induction of interferons in protective immunity to viruses. Immunol Rev. 2007, 220, 225–236. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, X. Long noncoding RNAs in innate immunity. Cell Mol Immunol. 2016, 13, 138–47. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, S.; Wang, G.; Feng, S.; Han, K.; Han, L.; Han, L. Role of microRNA and long non-coding RNA in Marek's disease tumorigenesis in chicken. Research in Veterinary Science 2021, 135, 134–142. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, Y.; Lin, L.; et al. The lncRNA MACC1-AS1 promotes gastric cancer cell metabolic plasticity via AMPK/Lin28 mediated mRNA stability of MACC1. Mol Canc. 2018, 17, 69. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Li, X.; Han, B.; Qu, L.; Liu, C.; Song, J.; Lian, L.; Yang, N. Gga-miR-130b- 3p inhibits MSB1 cell proliferation, migration, invasion, and its downregulation in MD tumor is attributed to hypermethylation. Oncotarget 2018, 9, 24187–24198. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Li, X.J.; Teng, M.; Dang, L.; Yu, Z.H.; Chi, J.Q.; Su, J.W.; Zhang, G.P.; Luo, J. In vivo expression patterns of microRNAs of Gallid herpesvirus 2 (GaHV-2) during the virus life cycle and development of Marek’s disease lymphomas. Virus Genes 2015, 50, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B. , Qi, F.; Wu, F.et al. Endogenous retrovirus-derived long noncoding RNA enhances innate immune responses via derepressing RELA expression. mBio.
- Zuckerman, B.; Ulitsky, I. Predictive models of subcellular localization of long RNAs. RNA 2019, 25, 557–572. [Google Scholar] [CrossRef]
Figure 1.
Protocol of RNA sequencing technique (N.E. Ilott, C.P. Ponting, Methods,2013).
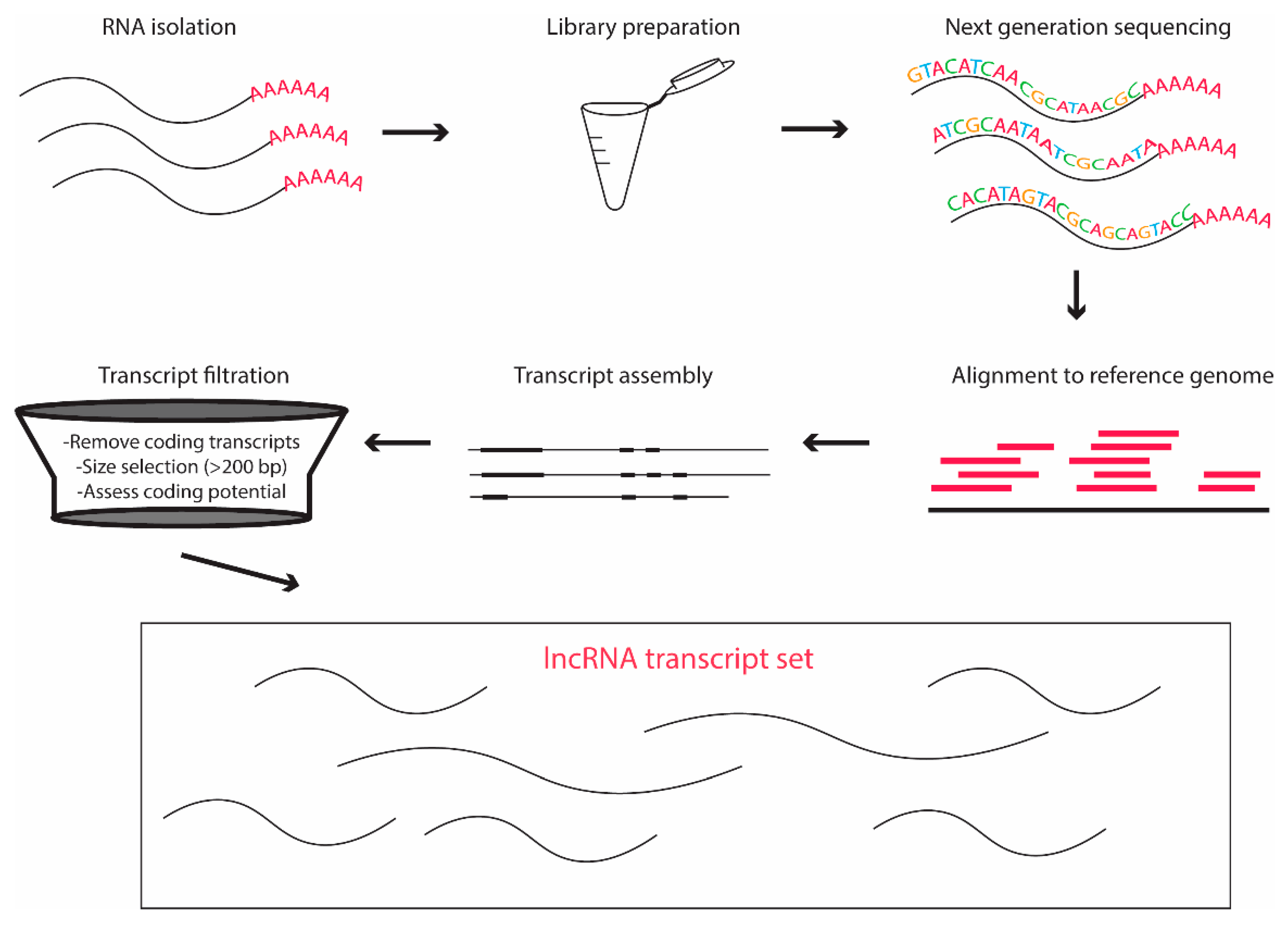

Table 1.
LncRNAs positively regulation viral replication.
| LncRNA | Influenza strain | Mechanism | Subcellular localization | References |
|---|---|---|---|---|
| NRAV | A/WSN/1933 (H1N1) | Suppresses the initial transcription of a number of important ISGs, including MxA, IFITM3, OASL, IFIT2, and IFIT3. | Nucleus | Ouyang et al. (2014) |
| TSPOAP1-AS1 | A/Puerto Rico/8/1934 (H1N1) | OASL, ISG20, IFIT1, IFITM1, and other anti-IAV ISGs are negatively regulated, which suppresses IAV-triggered type I IFN signalling. | Nucleus | WangQet al. (2019) |
| Lnc-Lsm3b | Blocks the overproduction of type Ά IFNs and inhibits RIG-I activation by competing with viral RNAs for the binding of RIG-I monomers. | Cytoplasm | Jiang et al. (2018) | |
| IPAN | A/WSN/1933 (H1N1) | Enhances the stability of the viral PB1 by forming an association that promotes IAV transcription and replication. | Cytoplasm/ Nucleus Cytoplasm/ Nucleus | Wang et al. (2019) |
| LncRNA-PAAN | A/WSN/1933 (H1N1) | Increases viral RNA polymerase activity by facilitating the assembly of the RdRp complex. | Nucleus Nucleus | Wang J et al. (2018) |
| LncRNA-ACOD1 | A/Puerto Rico/8/1934 (H1N1) | Increases Increases the synthesis of metabolites and the catalytic activity of GOT2. | Cytoplasm Cytoplasm | Wang P et al. (2017) |
| VIN | A/WSN/1933 (H1N1) | Unknown Unknown | Nucleus Nucleus | Winterling et al. (2014) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



