Submitted:
13 February 2025
Posted:
17 February 2025
You are already at the latest version
Abstract
Opioid analgesics have been used for more than 5,000 years and remain the main pain medications prescribed today. Although morphine is considered the gold standard of pain relief, this selective µ-opioid receptor (MOP) agonist provides only moderate relief for many chronic pain conditions, and produces a number of unwanted effects that can affect the patient’s quality-of-life, prevent adherence to treatment or lead to addiction. In addition to the lack of progress in developing better analgesics, there have been no significant breakthroughs to date in combating the above-mentioned side effects. Fortunately, a better understanding of opioid pharmacology has given renewed hope for the development of better and safer pain medications. In this review, we describe how clinically approved opioids were initially characterized as biased ligands and what impact this approach might have on clinical practice. We also looked at the preclinical and clinical development of biased MOP agonists, focusing on the history of oliceridine, the first specifically designed biased analgesic. In addition, we explore the discrepancies between ligands with low intrinsic efficacy and those with biased properties. Finally, we examine the rationale behind the development of biased ligands during the opioid crisis.
Keywords:
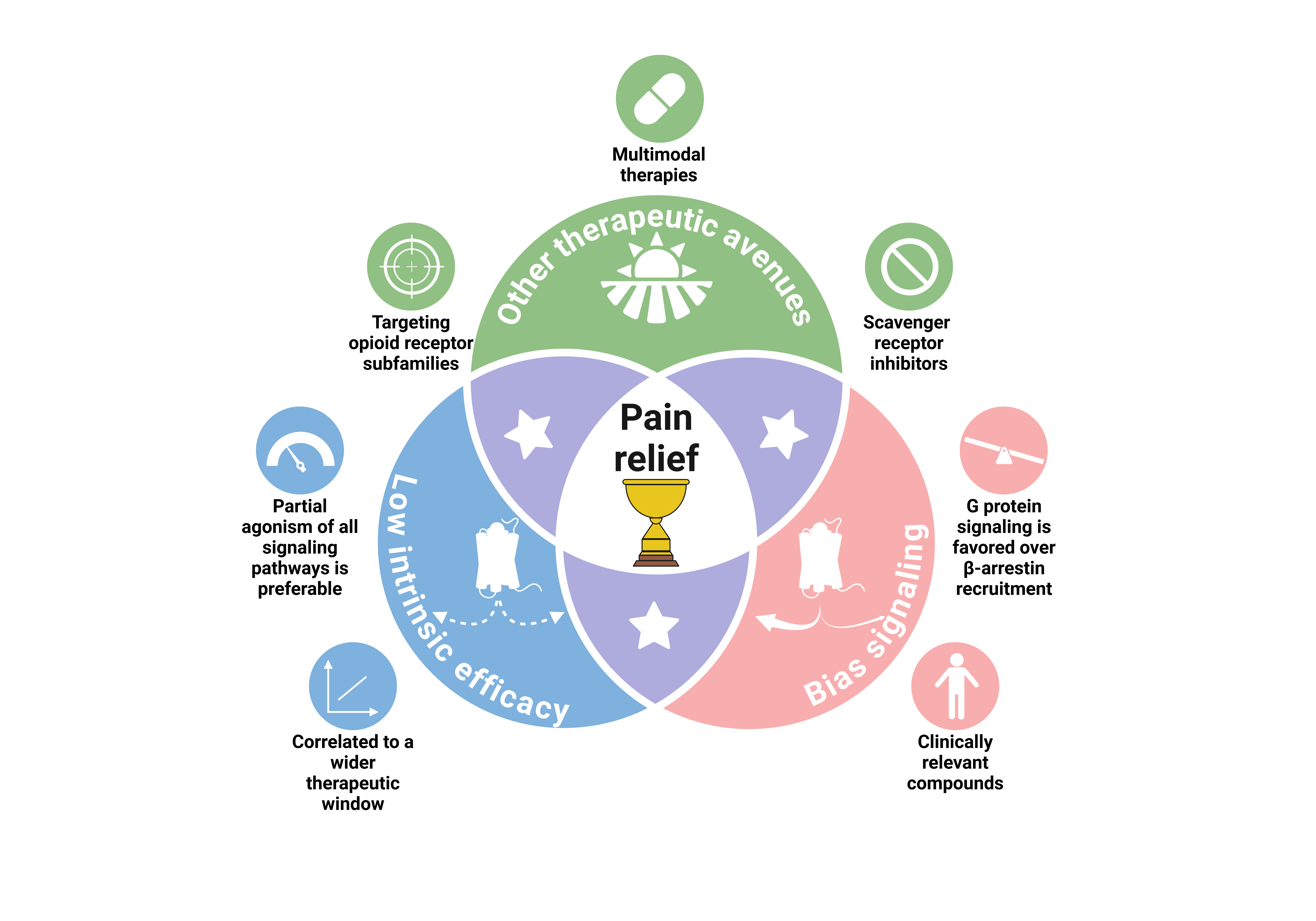
Graphical Abstract
1. Introduction
2. Understanding Opioid Pharmacology Design Better Analgesic Drugs
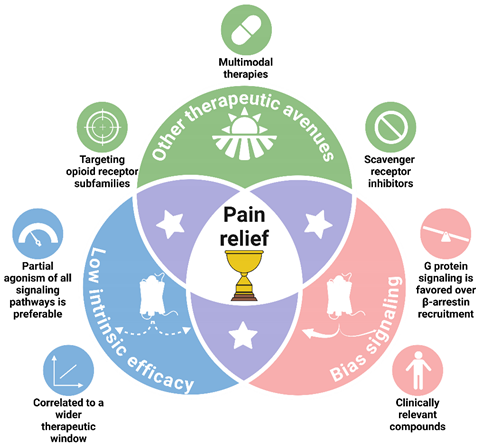
3. GPCRs: From Simple Switches to Integrated Signaling Processors
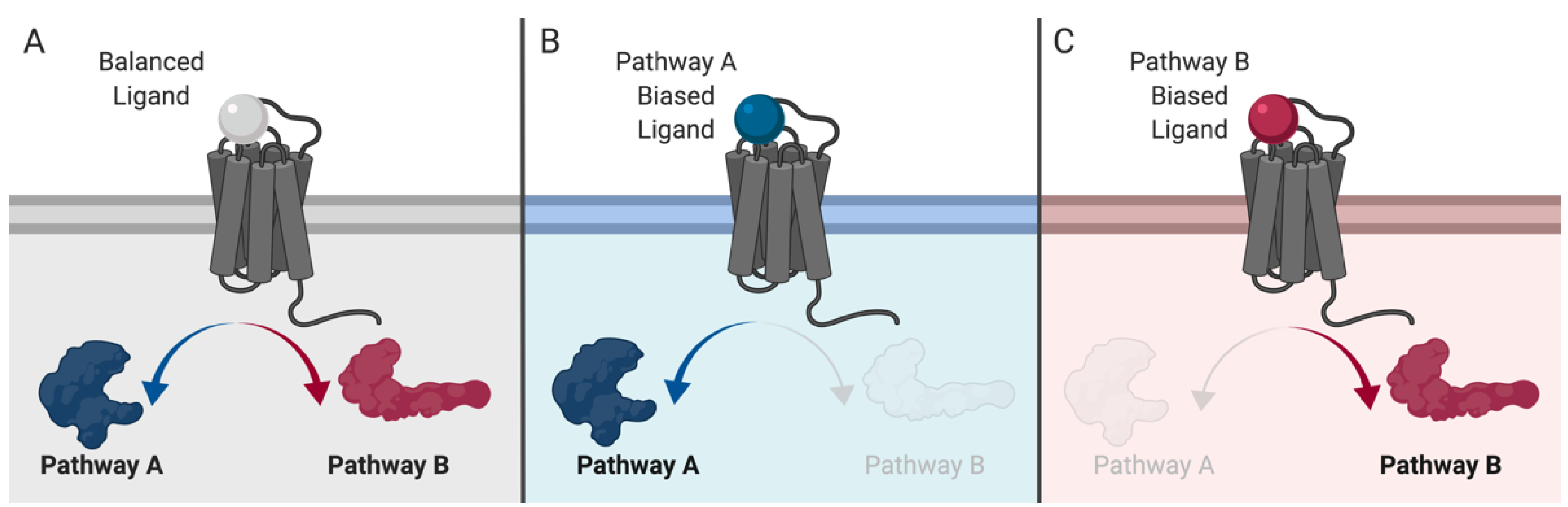
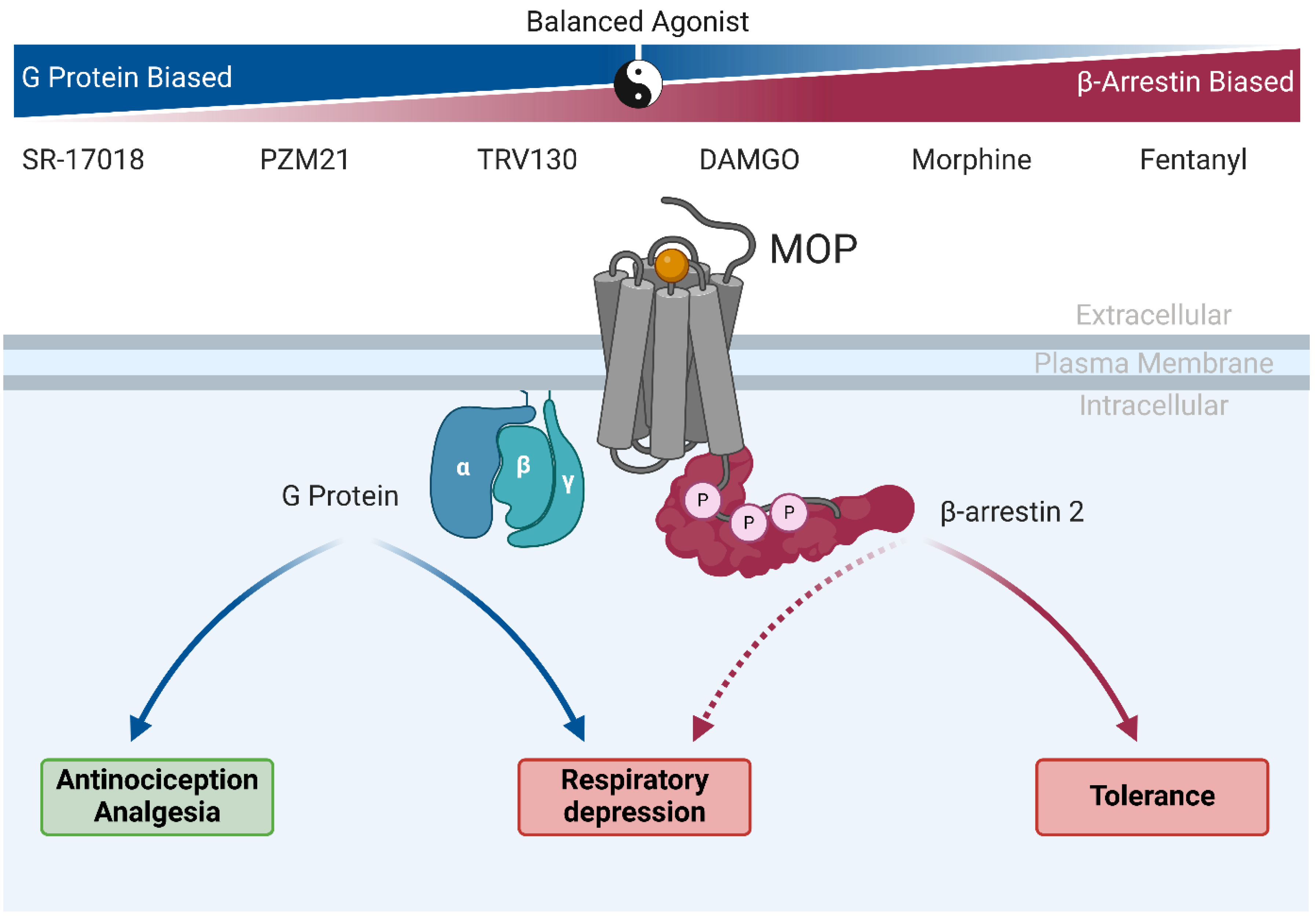
4. Old Drugs, New Tricks: MOP-Biased Agonists in Clinically Approved Opioids
5. Oliceridine, the Prototype MOP-Biased Agonist
6. MOP-Biased Ligands in Preclinical Development
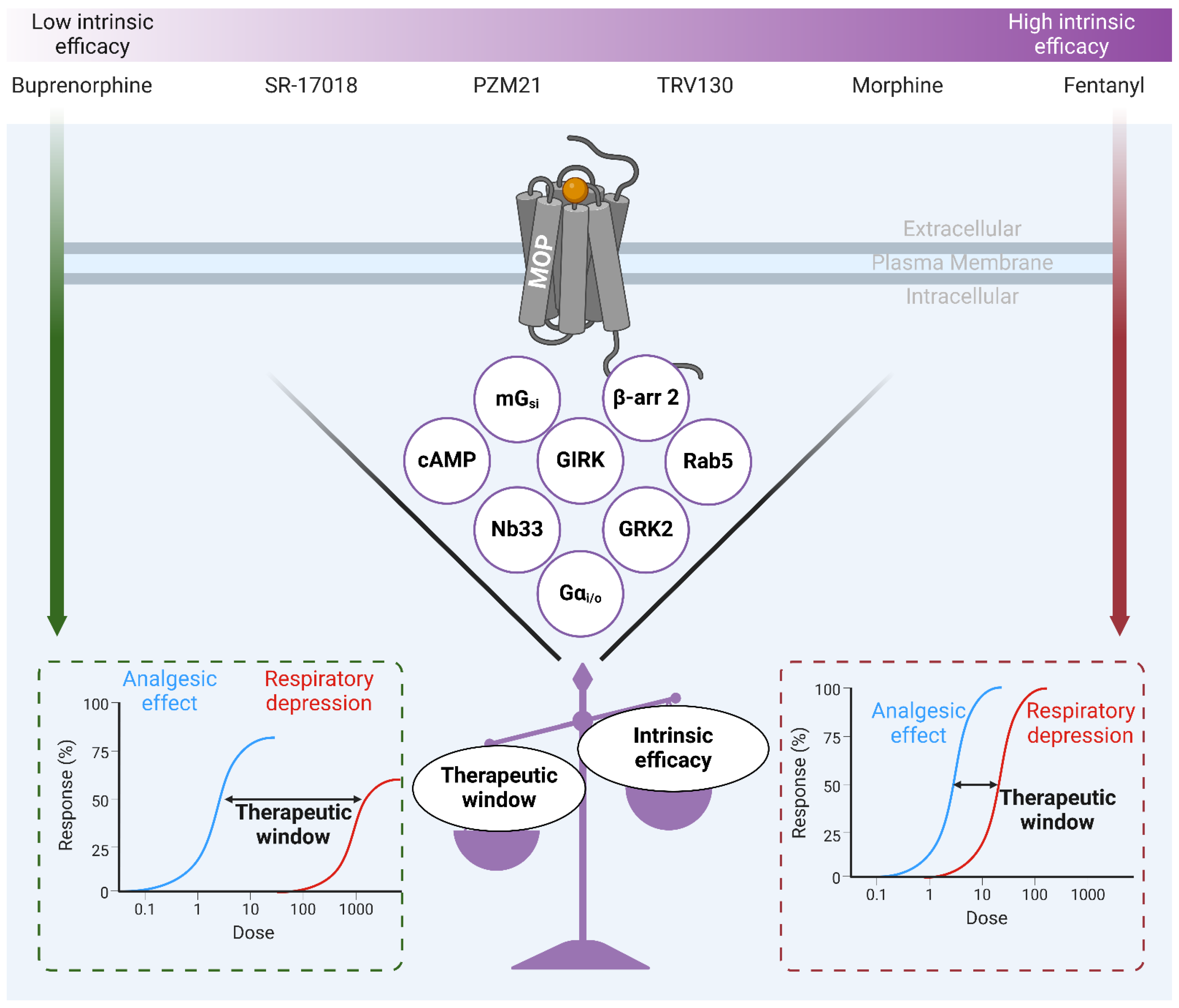
7. MOP-Biased Ligands Versus Low Intrinsic Efficacy Ligands at the µ-Opioid Receptor
8. Abuse Liability and Tolerance of Novel MOP Ligands in the Current Opioid Crisis
9. Alternatives to Biased Signaling at the µ-Opioid Receptor
10. The Still Unknown Component for an Ideal Analgesic
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Code Availability Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cohen SP, Vase L, Hooten WM. Chronic pain: an update on burden, best practices, and new advances. Lancet. 2021 May 29;397(10289):2082-97. [CrossRef]
- Volkow ND, Blanco C. The changing opioid crisis: development, challenges and opportunities. Mol Psychiatry. 2021 Jan;26(1):218-33. [CrossRef]
- Duarte DF. [Opium and opioids: a brief history.]. Rev Bras Anestesiol. 2005 Feb;55(1):135-46.
- Santoro D, Bellinghieri G, Savica V. Development of the concept of pain in history. J Nephrol. 2011 May-Jun;24 Suppl 17:S133-6. [CrossRef]
- Abdel Shaheed C, Maher CG, Williams KA, Day R, McLachlan AJ. Efficacy, Tolerability, and Dose-Dependent Effects of Opioid Analgesics for Low Back Pain: A Systematic Review and Meta-analysis. JAMA Intern Med. 2016 Jul 1;176(7):958-68.
- de Leon-Casasola OA. Opioids for chronic pain: new evidence, new strategies, safe prescribing. Am J Med. 2013 Mar;126(3 Suppl 1):S3-11.
- Kissin I. The development of new analgesics over the past 50 years: a lack of real breakthrough drugs. Anesth Analg. 2010 Mar 1;110(3):780-9. [CrossRef]
- WHO Expert Committee. The selection and use of essential medicines: report of the WHO Expert Committee, 2017 (including the 20th WHO Model List of Essential Medicines and the 6th WHO Model List of essential Medicines for Children). WHO technical report series ; no 1006. Geneva: World Health Organization; 2017. p. 604.
- Shipton EA, Shipton EE, Shipton AJ. A Review of the Opioid Epidemic: What Do We Do About It? Pain Ther. 2018 Jun;7(1):23-36.
- Manchikanti L, Helm S, 2nd, Fellows B, Janata JW, Pampati V, Grider JS, et al. Opioid epidemic in the United States. Pain Physician. 2012 Jul;15(3 Suppl):ES9-38.
- Webster L, Schmidt WK. Dilemma of Addiction and Respiratory Depression in the Treatment of Pain: A Prototypical Endomorphin as a New Approach. Pain Med. 2019 Jun 5. [CrossRef]
- Wang Y, Zhuang Y, DiBerto JF, Zhou XE, Schmitz GP, Yuan Q, et al. Structures of the entire human opioid receptor family. Cell. 2023 Jan 19;186(2):413-27 e17. [CrossRef]
- Matthes HW, Maldonado R, Simonin F, Valverde O, Slowe S, Kitchen I, et al. Loss of morphine-induced analgesia, reward effect and withdrawal symptoms in mice lacking the mu-opioid-receptor gene. Nature. 1996 Oct 31;383(6603):819-23.
- Hsia JA, Moss J, Hewlett EL, Vaughan M. ADP-ribosylation of adenylate cyclase by pertussis toxin. Effects on inhibitory agonist binding. J Biol Chem. 1984 Jan 25;259(2):1086-90. [CrossRef]
- Taussig R, Iniguez-Lluhi JA, Gilman AG. Inhibition of adenylyl cyclase by Gi alpha. Science. 1993 Jul 9;261(5118):218-21.
- Ippolito DL, Temkin PA, Rogalski SL, Chavkin C. N-terminal tyrosine residues within the potassium channel Kir3 modulate GTPase activity of Galphai. J Biol Chem. 2002 Sep 6;277(36):32692-6.
- Sadja R, Alagem N, Reuveny E. Gating of GIRK Channels. Neuron. 2003;39(1):9-12. [CrossRef]
- Díaz A, Flórez J, Pazos A, Hurlé MA. Opioid tolerance and supersensitivity induce regional changes in the autoradiographic density of dihydropyridine-sensitive calcium channels in the rat central nervous system. Pain. 2000;86(3):227-35. [CrossRef]
- Diaz A, Ruiz F, Florez J, Pazos A, Hurle MA. Regulation of dihydropyridine-sensitive Ca++ channels during opioid tolerance and supersensitivity in rats. J Pharmacol Exp Ther. 1995 Sep;274(3):1538-44. [CrossRef]
- Wickman K, Clapham DE. Ion channel regulation by G proteins. Physiol Rev. 1995 Oct;75(4):865-85. [CrossRef]
- Dang VC, Christie MJ. Mechanisms of rapid opioid receptor desensitization, resensitization and tolerance in brain neurons. Br J Pharmacol. 2012 Mar;165(6):1704-16. [CrossRef]
- Groer CE, Schmid CL, Jaeger AM, Bohn LM. Agonist-directed interactions with specific beta-arrestins determine mu-opioid receptor trafficking, ubiquitination, and dephosphorylation. J Biol Chem. 2011 Sep 9;286(36):31731-41.
- Melief EJ, Miyatake M, Bruchas MR, Chavkin C. Ligand-directed c-Jun N-terminal kinase activation disrupts opioid receptor signaling. Proc Natl Acad Sci U S A. 2010 Jun 22;107(25):11608-13. [CrossRef]
- Whistler JL, Chuang H-h, Chu P, Jan LY, von Zastrow M. Functional Dissociation of μ Opioid Receptor Signaling and Endocytosis. Neuron. 1999;23(4):737-46. [CrossRef]
- Gondin AB, Halls ML, Canals M, Briddon SJ. GRK Mediates mu-Opioid Receptor Plasma Membrane Reorganization. Front Mol Neurosci. 2019;12:104.
- Raffa RB, Martinez RP, Connelly CD. G-protein antisense oligodeoxyribonucleotides and μ-opioid supraspinal antinociception. European Journal of Pharmacology. 1994;258(1-2):R5-R7. [CrossRef]
- Bohn LM, Lefkowitz RJ, Gainetdinov RR, Peppel K, Caron MG, Lin FT. Enhanced morphine analgesia in mice lacking beta-arrestin 2. Science. 1999 Dec 24;286(5449):2495-8.
- Raehal KM, Walker JK, Bohn LM. Morphine Side Effects in β -Arrestin 2 Knockout Mice. J Pharmacol Exp Ther. 2005 Sep;314(3):1195-201. [CrossRef]
- Bohn LM, Gainetdinov RR, Lin FT, Lefkowitz RJ, Caron MG. Mu-opioid receptor desensitization by beta-arrestin-2 determines morphine tolerance but not dependence. Nature. 2000 Dec 7;408(6813):720-3.
- Gaveriaux-Ruff C, Kieffer BL. Opioid receptor genes inactivated in mice: the highlights. Neuropeptides. 2002 Apr-Jun;36(2-3):62-71. [CrossRef]
- Yang CH, Huang HW, Chen KH, Chen YS, Sheen-Chen SM, Lin CR. Antinociceptive potentiation and attenuation of tolerance by intrathecal beta-arrestin 2 small interfering RNA in rats. Br J Anaesth. 2011 Nov;107(5):774-81.
- Kliewer A, Schmiedel F, Sianati S, Bailey A, Bateman JT, Levitt ES, et al. Phosphorylation-deficient G-protein-biased mu-opioid receptors improve analgesia and diminish tolerance but worsen opioid side effects. Nat Commun. 2019 Jan 21;10(1):367.
- Bull FA, Baptista-Hon DT, Sneddon C, Wright L, Walwyn W, Hales TG. Src Kinase Inhibition Attenuates Morphine Tolerance without Affecting Reinforcement or Psychomotor Stimulation. Anesthesiology. 2017 Nov;127(5):878-89. [CrossRef]
- Kliewer A, Gillis A, Hill R, Schmiedel F, Bailey C, Kelly E, et al. Morphine-induced respiratory depression is independent of beta-arrestin2 signalling. Br J Pharmacol. 2020 Jul;177(13):2923-31.
- Chen YJ, Oldfield S, Butcher AJ, Tobin AB, Saxena K, Gurevich VV, et al. Identification of phosphorylation sites in the COOH-terminal tail of the mu-opioid receptor. J Neurochem. 2013 Jan;124(2):189-99.
- Miess E, Gondin AB, Yousuf A, Steinborn R, Mosslein N, Yang Y, et al. Multisite phosphorylation is required for sustained interaction with GRKs and arrestins during rapid mu-opioid receptor desensitization. Sci Signal. 2018 Jul 17;11(539).
- Bachmutsky I, Wei XP, Durand A, Yackle K. B-arrestin 2 germline knockout does not attenuate opioid respiratory depression. Elife. 2021 May 18;10.
- Gregory KJ. Commentary on "Morphine-induced respiratory depression is independent of beta-arrestin2 signalling". Br J Pharmacol. 2020 Jul;177(13):2904-5.
- Lam H, Maga M, Pradhan A, Evans CJ, Maidment NT, Hales TG, et al. Analgesic tone conferred by constitutively active mu opioid receptors in mice lacking beta-arrestin 2. Mol Pain. 2011 Apr 12;7:24.
- Stone LS, Molliver DC. In search of analgesia: emerging roles of GPCRs in pain. Mol Interv. 2009 Oct;9(5):234-51.
- Rajagopal S, Rajagopal K, Lefkowitz RJ. Teaching old receptors new tricks: biasing seven-transmembrane receptors. Nat Rev Drug Discov. 2010 May;9(5):373-86. [CrossRef]
- Audet M, Bouvier M. Restructuring G-protein- coupled receptor activation. Cell. 2012 Sep 28;151(1):14-23. [CrossRef]
- Kenakin T. Functional selectivity and biased receptor signaling. J Pharmacol Exp Ther. 2011 Feb;336(2):296-302. [CrossRef]
- Wootten D, Christopoulos A, Marti-Solano M, Babu MM, Sexton PM. Mechanisms of signalling and biased agonism in G protein-coupled receptors. Nat Rev Mol Cell Biol. 2018 Oct;19(10):638-53. [CrossRef]
- van der Westhuizen ET, Breton B, Christopoulos A, Bouvier M. Quantification of ligand bias for clinically relevant beta2-adrenergic receptor ligands: implications for drug taxonomy. Mol Pharmacol. 2014 Mar;85(3):492-509.
- Kenakin T, Christopoulos A. Signalling bias in new drug discovery: detection, quantification and therapeutic impact. Nat Rev Drug Discov. 2013 Mar;12(3):205-16. [CrossRef]
- Besserer-Offroy E, Sarret P. Sending out Biased Signals: an Appropriate Proposition for Pain? Douleur et Analgésie. 2019;32(2):108-10.
- Michel MC, Charlton SJ. Biased Agonism in Drug Discovery-Is It Too Soon to Choose a Path? Mol Pharmacol. 2018 Apr;93(4):259-65.
- Urs NM, Gee SM, Pack TF, McCorvy JD, Evron T, Snyder JC, et al. Distinct cortical and striatal actions of a beta-arrestin-biased dopamine D2 receptor ligand reveal unique antipsychotic-like properties. Proc Natl Acad Sci U S A. 2016 Dec 13;113(50):E8178-E86.
- Schattauer SS, Kuhar JR, Song A, Chavkin C. Nalfurafine is a G-protein biased agonist having significantly greater bias at the human than rodent form of the kappa opioid receptor. Cell Signal. 2017 Apr;32:59-65.
- Kenakin T. Bias translation: The final frontier? Br J Pharmacol. 2024 May;181(9):1345-60.
- Kenakin T. Know your molecule: pharmacological characterization of drug candidates to enhance efficacy and reduce late-stage attrition. Nat Rev Drug Discov. 2024 Aug;23(8):626-44. [CrossRef]
- Keith DE, Murray SR, Zaki PA, Chu PC, Lissin DV, Kang L, et al. Morphine activates opioid receptors without causing their rapid internalization. J Biol Chem. 1996 Aug 9;271(32):19021-4.
- Ehrlich AT, Kieffer BL, Darcq E. Current strategies toward safer mu opioid receptor drugs for pain management. Expert Opin Ther Targets. 2019 Feb 25:1-12. [CrossRef]
- McPherson J, Rivero G, Baptist M, Llorente J, Al-Sabah S, Krasel C, et al. mu-opioid receptors: correlation of agonist efficacy for signalling with ability to activate internalization. Mol Pharmacol. 2010 Oct;78(4):756-66.
- Schmid CL, Kennedy NM, Ross NC, Lovell KM, Yue Z, Morgenweck J, et al. Bias Factor and Therapeutic Window Correlate to Predict Safer Opioid Analgesics. Cell. 2017 Nov 16;171(5):1165-75 e13. [CrossRef]
- Butler S. Buprenorphine-Clinically useful but often misunderstood. Scand J Pain. 2013 Jul 1;4(3):148-52. [CrossRef]
- Lutfy K, Cowan A. Buprenorphine: a unique drug with complex pharmacology. Curr Neuropharmacol. 2004 Oct;2(4):395-402. [CrossRef]
- Davis MP, Pasternak G, Behm B. Treating Chronic Pain: An Overview of Clinical Studies Centered on the Buprenorphine Option. Drugs. 2018 Aug;78(12):1211-28. [CrossRef]
- Ehrlich AT, Darcq E. Recommending buprenorphine for pain management. Pain Manag. 2019 Jan 1;9(1):13-6. [CrossRef]
- Vlok R, An GH, Binks M, Melhuish T, White L. Sublingual buprenorphine versus intravenous or intramuscular morphine in acute pain: A systematic review and meta-analysis of randomized control trials. Am J Emerg Med. 2019 Mar;37(3):381-6.
- Gudin J, Fudin J, Nalamachu S. Levorphanol use: past, present and future. Postgrad Med. 2016 Jan;128(1):46-53. [CrossRef]
- Prommer E. Levorphanol: the forgotten opioid. Support Care Cancer. 2007 Mar;15(3):259-64. [CrossRef]
- Prommer E. Levorphanol: revisiting an underutilized analgesic. Palliat Care. 2014;8:7-10. [CrossRef]
- Le Rouzic V, Narayan A, Hunkle A, Marrone GF, Lu Z, Majumdar S, et al. Pharmacological Characterization of Levorphanol, a G-Protein Biased Opioid Analgesic. Anesth Analg. 2019 Feb;128(2):365-73. [CrossRef]
- Nair AS, Upputuri O, Mantha SSP, Rayani BK. Levorphanol: Rewinding an Old, Bygone Multimodal Opioid Analgesic! Indian J Palliat Care. 2019 Jul-Sep;25(3):483-4.
- Pham TC, Fudin J, Raffa RB. Is levorphanol a better option than methadone? Pain Med. 2015 Sep;16(9):1673-9.
- Chen X-T, Pitis P, Liu G, Yuan C, Gotchev D, Cowan CL, et al. Structure–Activity Relationships and Discovery of a G Protein Biased μ Opioid Receptor Ligand, [(3-Methoxythiophen-2-yl)methyl]({2-[(9R)-9-(pyridin-2-yl)-6-oxaspiro-[4.5]decan-9-yl]ethyl})amine (TRV130), for the Treatment of Acute Severe Pain. Journal of Medicinal Chemistry. 2013;56(20):8019-31.
- DeWire SM, Yamashita DS, Rominger DH, Liu G, Cowan CL, Graczyk TM, et al. A G protein-biased ligand at the mu-opioid receptor is potently analgesic with reduced gastrointestinal and respiratory dysfunction compared with morphine. J Pharmacol Exp Ther. 2013 Mar;344(3):708-17.
- Trevena Inc. Trevena Announces Submission of New Drug Application to U.S. FDA for Oliceridine Injection. 2017 [cited 2020-03-24]; Available from: https://www.trevena.com/investors/press-releases/detail/127/trevena-announces-submission-of-new-drug-application-to.
- Trevena Inc. Trevena Announces FDA Acceptance for Review of New Drug Application for Oliceridine Injection. 2018 [cited 2020-03-24]; Available from: https://www.trevena.com/investors/press-releases/detail/129/trevena-announces-fda-acceptance-for-review-of-new-drug.
- Trevena Inc. Trevena Announces Oliceridine FDA Advisory Committee Meeting Outcome. 2018 [cited 2020-03-24]; Available from: https://www.trevena.com/investors/press-releases/detail/154/trevena-announces-oliceridine-fda-advisory-committee.
- FDA AADPAC. Transcript for the October 11, 2018 Meeting of the Anesthetic and Analgesic Drug Products Advisory Committee (AADPAC). 2018 [cited 2020-03-24]; Available from: https://www.fda.gov/downloads/AdvisoryCommittees/CommitteesMeetingMaterials/Drugs/AnestheticAndAnalgesicDrugProductsAdvisoryCommittee/UCM629602.pdf.
- Trevena Inc. Trevena Announces Receipt of Type A Meeting Minutes and Provides Regulatory Update for Oliceridine. 2019 [cited 2020-03-14]; Available from: https://www.trevena.com/investors/press-releases/detail/160/trevena-announces-receipt-of-type-a-meeting-minutes-and.
- Trevena Inc. Trevena Announces Initiation of Healthy Volunteer Study for Oliceridine. 2019 [cited 2020-05-12]; Available from: https://www.trevena.com/investors/press-releases/detail/5/trevena-announces-initiation-of-healthy-volunteer-study-for.
- Nafziger AN, Arscott KA, Cochrane K, Skobieranda F, Burt DA, Fossler MJ. The Influence of Renal or Hepatic Impairment on the Pharmacokinetics, Safety, and Tolerability of Oliceridine. Clin Pharmacol Drug Dev. 2019 Nov 7. [CrossRef]
- Trevena Inc. Trevena Resubmits New Drug Application for Oliceridine. 2020 [cited 2020-06-16]; Available from: https://www.trevena.com/investors/press-releases/detail/224/trevena-resubmits-new-drug-application-for-oliceridine.
- Gan TJ, Wase L. Oliceridine, a G protein-selective ligand at the mu-opioid receptor, for the management of moderate to severe acute pain. Drugs Today (Barc). 2020 Apr;56(4):269-86. [CrossRef]
- Trevena Inc. Trevena Announces FDA Approval of OLINVYK™ (oliceridine) injection. 2020 [cited 2024-08-18]; Available from: https://www.trevena.com/investors/press-releases/detail/234/trevena-announces-fda-approval-of-olinvyk-oliceridine-injection.
- U.S. Food & Drug Administration. FDA Approves New Opioid for Intravenous Use in Hospitals, Other Controlled Clinical Settings. 2020 [cited; Available from: https://www.fda.gov/news-events/press-announcements/fda-approves-new-opioid-intravenous-use-hospitals-other-controlled-clinical-settings.
- Shah A, Shah R, Fahim G, Brust-Sisti LA. A Dive Into Oliceridine and Its Novel Mechanism of Action. Cureus. 2021 Oct;13(10):e19076. [CrossRef]
- Azzam AAH, Lambert DG. Preclinical discovery and development of oliceridine (Olinvyk(R)) for the treatment of post-operative pain. Expert Opin Drug Discov. 2022 Mar;17(3):215-23.
- Trevena Inc. Trevena, Inc. Announces DEA Scheduling of OLINVYK™ (oliceridine) injection. 2020 [cited; Available from: https://www.trevena.com/investors/press-releases/detail/244/trevena-inc-announces-dea-scheduling-of-olinvyk-oliceridine-injection.
- U.S. Food & Drug Administration. Second Cycle Review And Summary Basis for Approval - Oliceridine injection (NDA 210-730). 2020 2020-08-07 [cited 210730Orig1s000 ]; Available from: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2020/210730Orig1s000MultidisciplineR.pdf.
- Trevena Inc. Trevena Announces Approval of OLINVYK in China. 2023 [cited 2024-08-18]; Available from: https://www.trevena.com/investors/press-releases/detail/320/trevena-announces-approval-of-olinvyk-in-china.
- Ayad S, Demitrack MA, Burt DA, Michalsky C, Wase L, Fossler MJ, et al. Evaluating the Incidence of Opioid-Induced Respiratory Depression Associated with Oliceridine and Morphine as Measured by the Frequency and Average Cumulative Duration of Dosing Interruption in Patients Treated for Acute Postoperative Pain. Clin Drug Investig. 2020 Aug;40(8):755-64. [CrossRef]
- Brzezinski M, Hammer GB, Candiotti KA, Bergese SD, Pan PH, Bourne MH, et al. Low Incidence of Opioid-Induced Respiratory Depression Observed with Oliceridine Regardless of Age or Body Mass Index: Exploratory Analysis from a Phase 3 Open-Label Trial in Postsurgical Pain. Pain Ther. 2021 Jun;10(1):457-73. [CrossRef]
- Hammer GB, Khanna AK, Michalsky C, Wase L, Demitrack MA, Little R, et al. Oliceridine Exhibits Improved Tolerability Compared to Morphine at Equianalgesic Conditions: Exploratory Analysis from Two Phase 3 Randomized Placebo and Active Controlled Trials. Pain Ther. 2021 Dec;10(2):1343-53. [CrossRef]
- Beard TL, Michalsky C, Candiotti KA, Rider P, Wase L, Habib AS, et al. Oliceridine is Associated with Reduced Risk of Vomiting and Need for Rescue Antiemetics Compared to Morphine: Exploratory Analysis from Two Phase 3 Randomized Placebo and Active Controlled Trials. Pain Ther. 2021 Jun;10(1):401-13. [CrossRef]
- Hill DM, DeBoer E. State and Future Science of Opioids and Potential of Biased-ligand Technology in the Management of Acute Pain After Burn Injury. J Burn Care Res. 2023 May 2;44(3):524-34. [CrossRef]
- Manglik A, Lin H, Aryal DK, McCorvy JD, Dengler D, Corder G, et al. Structure-based discovery of opioid analgesics with reduced side effects. Nature. 2016 Sep 8;537(7619):185-90. [CrossRef]
- Emery MA, Eitan S. Members of the same pharmacological family are not alike: Different opioids, different consequences, hope for the opioid crisis? Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2019;92:428-49.
- Kudla L, Bugno R, Skupio U, Wiktorowska L, Solecki W, Wojtas A, et al. Functional characterization of a novel opioid, PZM21, and its influence on behavioural responses to morphine. Br J Pharmacol. 2019 Jul 26.
- Hill R, Disney A, Conibear A, Sutcliffe K, Dewey W, Husbands S, et al. The novel mu-opioid receptor agonist PZM21 depresses respiration and induces tolerance to antinociception. Br J Pharmacol. 2018 Jul;175(13):2653-61.
- Grim TW, Acevedo-Canabal A, Bohn LM. Toward Directing Opioid Receptor Signaling to Refine Opioid Therapeutics. Biol Psychiatry. 2020 Jan 1;87(1):15-21. [CrossRef]
- de Waal PW, Shi J, You E, Wang X, Melcher K, Jiang Y, et al. Molecular mechanisms of fentanyl mediated beta-arrestin biased signaling. PLoS Comput Biol. 2020 Apr;16(4):e1007394.
- Ramos-Gonzalez N, Groom S, Sutcliffe KJ, Bancroft S, Bailey CP, Sessions RB, et al. Carfentanil is a beta-arrestin-biased agonist at the mu opioid receptor. Br J Pharmacol. 2023 Sep;180(18):2341-60.
- Huang W, Manglik A, Venkatakrishnan AJ, Laeremans T, Feinberg EN, Sanborn AL, et al. Structural insights into micro-opioid receptor activation. Nature. 2015 Aug 20;524(7565):315-21.
- Zhuang Y, Wang Y, He B, He X, Zhou XE, Guo S, et al. Molecular recognition of morphine and fentanyl by the human mu-opioid receptor. Cell. 2022 Nov 10;185(23):4361-75 e19.
- Kelly E, Conibear A, Henderson G. Biased Agonism: Lessons from Studies of Opioid Receptor Agonists. Annu Rev Pharmacol Toxicol. 2023 Jan 20;63:491-515. [CrossRef]
- Kise R, Inoue A. GPCR signaling bias: an emerging framework for opioid drug development. J Biochem. 2024 Mar 25;175(4):367-76. [CrossRef]
- Gillis A, Gondin AB, Kliewer A, Sanchez J, Lim HD, Alamein C, et al. Low intrinsic efficacy for G protein activation can explain the improved side effect profiles of new opioid agonists. Sci Signal. 2020 Mar 31;13(625). [CrossRef]
- Stoeber M, Jullie D, Lobingier BT, Laeremans T, Steyaert J, Schiller PW, et al. A Genetically Encoded Biosensor Reveals Location Bias of Opioid Drug Action. Neuron. 2018 Jun 6;98(5):963-76 e5. [CrossRef]
- Wan Q, Okashah N, Inoue A, Nehme R, Carpenter B, Tate CG, et al. Mini G protein probes for active G protein-coupled receptors (GPCRs) in live cells. J Biol Chem. 2018 May 11;293(19):7466-73. [CrossRef]
- Stahl EL, Bohn LM. Low Intrinsic Efficacy Alone Cannot Explain the Improved Side Effect Profiles of New Opioid Agonists. Biochemistry. 2022 Sep 20;61(18):1923-35. [CrossRef]
- Clark DJ, Schumacher MA. America's Opioid Epidemic: Supply and Demand Considerations. Anesth Analg. 2017 Nov;125(5):1667-74. [CrossRef]
- Killeen N, Lakes R, Webster M, Killoran S, McNamara S, Kavanagh P, et al. The emergence of nitazenes on the Irish heroin market and national preparation for possible future outbreaks. Addiction. 2024 Sep;119(9):1657-8. [CrossRef]
- Kozell LB, Eshleman AJ, Wolfrum KM, Swanson TL, Bloom SH, Benware S, et al. Pharmacologic Characterization of Substituted Nitazenes at mu, kappa, and Delta Opioid Receptors Suggests High Potential for Toxicity. J Pharmacol Exp Ther. 2024 Apr 18;389(2):219-28.
- Bendjilali-Sabiani JJ, Eiden C, Lestienne M, Cherki S, Gautre D, Van den Broek T, et al. Isotonitazene, a synthetic opioid from an emerging family: The nitazenes. Therapie. 2024 May 28. [CrossRef]
- Amaducci A, Aldy K, Campleman SL, Li S, Meyn A, Abston S, et al. Naloxone Use in Novel Potent Opioid and Fentanyl Overdoses in Emergency Department Patients. JAMA Netw Open. 2023 Aug 1;6(8):e2331264. [CrossRef]
- Hauser W, Schug S, Furlan AD. The opioid epidemic and national guidelines for opioid therapy for chronic noncancer pain: a perspective from different continents. Pain Rep. 2017 May;2(3):e599. [CrossRef]
- Rudd RA, Seth P, David F, Scholl L. Increases in Drug and Opioid-Involved Overdose Deaths - United States, 2010-2015. MMWR Morb Mortal Wkly Rep. 2016 Dec 30;65(50-51):1445-52.
- Busse JW, Craigie S, Juurlink DN, Buckley DN, Wang L, Couban RJ, et al. Guideline for opioid therapy and chronic noncancer pain. CMAJ. 2017 May 8;189(18):E659-E66. [CrossRef]
- Bohn LM, Gainetdinov RR, Sotnikova TD, Medvedev IO, Lefkowitz RJ, Dykstra LA, et al. Enhanced Rewarding Properties of Morphine, but not Cocaine, in βarrestin-2 Knock-Out Mice. The Journal of Neuroscience. 2003;23(32):10265-73. [CrossRef]
- Zamarripa CA, Edwards SR, Qureshi HN, Yi JN, Blough BE, Freeman KB. The G-protein biased mu-opioid agonist, TRV130, produces reinforcing and antinociceptive effects that are comparable to oxycodone in rats. Drug Alcohol Depend. 2018 Nov 1;192:158-62.
- Altarifi AA, David B, Muchhala KH, Blough BE, Akbarali H, Negus SS. Effects of acute and repeated treatment with the biased mu opioid receptor agonist TRV130 (oliceridine) on measures of antinociception, gastrointestinal function, and abuse liability in rodents. J Psychopharmacol. 2017 Jun;31(6):730-9. [CrossRef]
- Liang DY, Li WW, Nwaneshiudu C, Irvine KA, Clark JD. Pharmacological Characters of Oliceridine, a mu-Opioid Receptor G-Protein[FIGURE DASH]Biased Ligand in Mice. Anesth Analg. 2018 Jul 21.
- Soergel DG, Subach RA, Burnham N, Lark MW, James IE, Sadler BM, et al. Biased agonism of the mu-opioid receptor by TRV130 increases analgesia and reduces on-target adverse effects versus morphine: A randomized, double-blind, placebo-controlled, crossover study in healthy volunteers. Pain. 2014 Sep;155(9):1829-35.
- Grim TW, Schmid CL, Stahl EL, Pantouli F, Ho JH, Acevedo-Canabal A, et al. A G protein signaling-biased agonist at the mu-opioid receptor reverses morphine tolerance while preventing morphine withdrawal. Neuropsychopharmacology. 2020 Jan;45(2):416-25.
- Quirion B, Bergeron F, Blais V, Gendron L. The Delta-Opioid Receptor; a Target for the Treatment of Pain. Front Mol Neurosci. 2020;13:52. [CrossRef]
- Blaine AT, van Rijn RM. Receptor expression and signaling properties in the brain, and structural ligand motifs that contribute to delta opioid receptor agonist-induced seizures. Neuropharmacology. 2023 Jul 1;232:109526.
- Conibear AE, Asghar J, Hill R, Henderson G, Borbely E, Tekus V, et al. A Novel G Protein-Biased Agonist at the delta Opioid Receptor with Analgesic Efficacy in Models of Chronic Pain. J Pharmacol Exp Ther. 2020 Feb;372(2):224-36.
- PharmNovo A. PharmNovo announces positive Phase I results for neuropathic pain management drug PN6047. 2024 [cited 2024-08-18]; Available from: https://www.mediconvillage.se/pharmnovo-announces-positive-phase-i-results-for-neuropathic-pain-management-drug-pn6047/.
- Cayir S, Zhornitsky S, Barzegary A, Sotomayor-Carreno E, Sarfo-Ansah W, Funaro MC, et al. A review of the kappa opioid receptor system in opioid use. Neurosci Biobehav Rev. 2024 Jul;162:105713. [CrossRef]
- Santino F, Gentilucci L. Design of kappa-Opioid Receptor Agonists for the Development of Potential Treatments of Pain with Reduced Side Effects. Molecules. 2023 Jan 1;28(1).
- Dalefield ML, Scouller B, Bibi R, Kivell BM. The Kappa Opioid Receptor: A Promising Therapeutic Target for Multiple Pathologies. Front Pharmacol. 2022;13:837671. [CrossRef]
- Mores KL, Cummins BR, Cassell RJ, van Rijn RM. A Review of the Therapeutic Potential of Recently Developed G Protein-Biased Kappa Agonists. Front Pharmacol. 2019;10:407. [CrossRef]
- Valentino RJ, Volkow ND. Untangling the complexity of opioid receptor function. Neuropsychopharmacology. 2018 Dec;43(13):2514-20. [CrossRef]
- Cahill CM, Lueptow L, Kim H, Shusharla R, Bishop A, Evans CJ. Kappa Opioid Signaling at the Crossroads of Chronic Pain and Opioid Addiction. Handb Exp Pharmacol. 2022;271:315-50.
- Arendt-Nielsen L, Olesen AE, Staahl C, Menzaghi F, Kell S, Wong GY, et al. Analgesic efficacy of peripheral kappa-opioid receptor agonist CR665 compared to oxycodone in a multi-modal, multi-tissue experimental human pain model: selective effect on visceral pain. Anesthesiology. 2009 Sep;111(3):616-24.
- Meyrath M, Szpakowska M, Zeiner J, Massotte L, Merz MP, Benkel T, et al. The atypical chemokine receptor ACKR3/CXCR7 is a broad-spectrum scavenger for opioid peptides. Nat Commun. 2020 Jun 19;11(1):3033.
- Gach-Janczak K, Biernat M, Kuczer M, Adamska-Bartlomiejczyk A, Kluczyk A. Analgesic Peptides: From Natural Diversity to Rational Design. Molecules. 2024 Mar 29;29(7). [CrossRef]
- Palmer CB, Meyrath M, Canals M, Kostenis E, Chevigne A, Szpakowska M. Atypical opioid receptors: unconventional biology and therapeutic opportunities. Pharmacol Ther. 2022 May;233:108014. [CrossRef]
- O'Neill A, Lirk P. Multimodal Analgesia. Anesthesiol Clin. 2022 Sep;40(3):455-68.
- Young A, Buvanendran A. Recent advances in multimodal analgesia. Anesthesiol Clin. 2012 Mar;30(1):91-100. [CrossRef]
- Giummarra MJ, Gibson SJ, Allen AR, Pichler AS, Arnold CA. Polypharmacy and chronic pain: harm exposure is not all about the opioids. Pain Med. 2015 Mar;16(3):472-9. [CrossRef]
- Arumugam S, Lau CS, Chamberlain RS. Use of preoperative gabapentin significantly reduces postoperative opioid consumption: a meta-analysis. J Pain Res. 2016;9:631-40. [CrossRef]
- Pergolizzi JV, Jr., van de Laar M, Langford R, Mellinghoff HU, Merchante IM, Nalamachu S, et al. Tramadol/paracetamol fixed-dose combination in the treatment of moderate to severe pain. J Pain Res. 2012;5:327-46. [CrossRef]
- Pergolizzi J, Varrassi G, LeQuang JAK, Breve F, Magnusson P. Fixed Dose Versus Loose Dose: Analgesic Combinations. Cureus. 2023 Jan;15(1):e33320. [CrossRef]
- Frantz S. The trouble with making combination drugs. Nat Rev Drug Discov. 2006 Nov;5(11):881-2.
- Gascon N, Almansa C, Merlos M, Miguel Vela J, Encina G, Morte A, et al. Co-crystal of tramadol-celecoxib: preclinical and clinical evaluation of a novel analgesic. Expert Opin Investig Drugs. 2019 May;28(5):399-409. [CrossRef]
- Caputi FF, Nicora M, Simeone R, Candeletti S, Romualdi P. Tapentadol: an analgesic that differs from classic opioids due to its noradrenergic mechanism of action. Minerva Med. 2019 Feb;110(1):62-78. [CrossRef]
- Barakat A. Revisiting Tramadol: A Multi-Modal Agent for Pain Management. CNS Drugs. 2019 May;33(5):481-501. [CrossRef]
- Cannella N, Lunerti V, Shen Q, Li H, Benvenuti F, Soverchia L, et al. Cebranopadol, a novel long-acting opioid agonist with low abuse liability, to treat opioid use disorder: Preclinical evidence of efficacy. Neuropharmacology. 2024 Oct 1;257:110048. [CrossRef]
- Kiguchi N, Ding H, Ko MC. Therapeutic potentials of NOP and MOP receptor coactivation for the treatment of pain and opioid abuse. J Neurosci Res. 2022 Jan;100(1):191-202. [CrossRef]
- Sanam M, Ashraf S, Saeed M, Khalid A, Abdalla AN, Qureshi U, et al. Cebranopadol: An Assessment for Its Biased Activation Potential at the Mu Opioid Receptor by DFT, Molecular Docking and Molecular Dynamic Simulation Studies. ChemistrySelect. 2023;8(37). [CrossRef]
- Kleczkowska P, Lipkowski AW, Tourwe D, Ballet S. Hybrid opioid/non-opioid ligands in pain research. Curr Pharm Des. 2013;19(42):7435-50. [CrossRef]
- Breault E, Desgagne M, Neve J, Cote J, Barlow TMA, Ballet S, et al. Multitarget ligands that comprise opioid/nonopioid pharmacophores for pain management: Current state of the science. Pharmacol Res. 2024 Nov;209:107408. [CrossRef]
- Smith MT, Kong D, Kuo A, Imam MZ, Williams CM. Multitargeted Opioid Ligand Discovery as a Strategy to Retain Analgesia and Reduce Opioid-Related Adverse Effects. J Med Chem. 2023 Mar 23;66(6):3746-84. [CrossRef]
- Kenakin T. Biased Receptor Signaling in Drug Discovery. Pharmacol Rev. 2019 Apr;71(2):267-315. [CrossRef]
- Luttrell LM, Maudsley S, Gesty-Palmer D. Translating in vitro ligand bias into in vivo efficacy. Cell Signal. 2018 Jan;41:46-55. [CrossRef]
- Walker CS, Sundrum T, Hay DL. PACAP receptor pharmacology and agonist bias: analysis in primary neurons and glia from the trigeminal ganglia and transfected cells. Br J Pharmacol. 2014 Mar;171(6):1521-33. [CrossRef]
- Ehrlich AT, Semache M, Gross F, Da Fonte DF, Runtz L, Colley C, et al. Biased Signaling of the Mu Opioid Receptor Revealed in Native Neurons. iScience. 2019 Apr 26;14:47-57. [CrossRef]
- Rivero G, Llorente J, McPherson J, Cooke A, Mundell SJ, McArdle CA, et al. Endomorphin-2: a biased agonist at the mu-opioid receptor. Mol Pharmacol. 2012 Aug;82(2):178-88.
- Kelly E. Efficacy and ligand bias at the mu-opioid receptor. Br J Pharmacol. 2013 Aug;169(7):1430-46.
- Pedersen MF, Wrobel TM, Marcher-Rorsted E, Pedersen DS, Moller TC, Gabriele F, et al. Biased agonism of clinically approved mu-opioid receptor agonists and TRV130 is not controlled by binding and signaling kinetics. Neuropharmacology. 2019 Jul 24:107718.

| Compound | Structure | Biased toward1 |
Contradictory literature |
| Morphine | 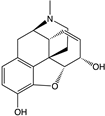 |
β-arrestins [55,68,150] |
No bias [101,151,152] |
| Fentanyl | 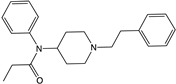 |
β-arrestins [55,95] |
No bias [96] |
| Buprenorphine | 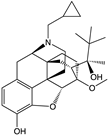 |
G protein [101,118,150,153] |
|
| Levorphanol |  |
G protein [65] |
|
| Oliceridine | 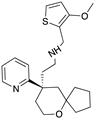 |
G protein [67,68] |
Tendency towards G protein [101] |
| PZM21 | 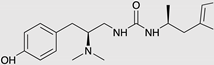 |
G protein [90,93] |
Tendency towards G protein [101] |
| SR-17018 | 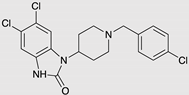 |
G protein [55] |
Tendency towards G protein [101] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



