
Submitted:
09 October 2024
Posted:
10 October 2024
You are already at the latest version
Abstract
One of the most common nutritional problems in athletes is iron deficiency. Although a lot of research has been done, the problem is still not solved. It is believed that iron deficiency is basically a lack of iron intake due to low dietary intake. Therefore, athletes who are found to be iron deficient are required to increase their iron intake (supplementation or intravenous administration). However, there are many supplements available on the market today, and if the problem is simply a lack of iron intake, it is a nutritional problem that should have been solved. In other words, there are reasons why the problem cannot be solved by simply taking iron, and it is important to conduct further research on the relationship between iron status and physical activity in the future.
In this paper, we summarize our recent research, focusing on the studies we have done so far.
Keywords:
athlete
; aerobic exercise
; resistance exercise
; iron deficiency
; ACD
【Iron Distribution in the Body】
Sixty to seventy percent of the iron in the body is present as part of the heme in hemoglobin (Figure 1). Heme iron, other than hemoglobin, accounts for approximately 5% in body and acts as an oxygen transporter in tissues. Enzymes that are not heme iron include enzymes that carry out oxidation reactions. In mitochondria, these non-heme iron enzyme groups are more abundant than in cytochromes. Iron stores, which account for approximately 25% of the body iron, include ferritin and hemosiderin. When iron loss is lower than the iron intake, apoferritin synthesis increases with an increasing iron intake, and iron adsorption onto ferritin within free polyribosomes proceeds, increasing ferritin stores in the liver. At the same time, hepatic hemosiderin stores are increased, apoferritin synthesis in the reticuloendothelial system and endoplasmic reticulum of hepatocytes is enhanced, and serum ferritin is also increased. Conversely, when iron loss is greater than the iron uptake, iron is released from ferritin in the liver and the serum iron concentration increases. In addition, iron stored in the reticuloendothelial system and hepatocytes is released into peripheral blood at a rate of 20-25 mg/day, and serum iron is taken up by erythroblasts in the bone marrow at the same rate [1,2,3]. Iron incorporated into erythroblasts is used for hemoglobin synthesis, and erythroblasts differentiate into erythrocytes and are released into the peripheral blood. Erythrocytes released into the peripheral blood eventually age and are phagocytosed by the reticuloendothelial macrophages. Approximately 85% of iron in erythrocytes phagocytosed by reticuloendothelial macrophages is released back into the body in the form of transferrin and ferritin, and the remainder is stored in the form of ferritin and hemosiderin. Thus, the uptake of iron into erythroblasts is complemented and maintained by iron supply from macrophages [1,2,4].
Dietary iron is absorbed from the upper gastrointestinal tract and enters the bloodstream, where it is bound to transferrin (Tf) and transported throughout the body. Some of the iron is stored in the liver and used by cells throughout the body, including muscle cells. The erythrocytes produced circulate throughout the body to carry oxygen, and at the end of their 120-day life span, the waste erythrocytes are destroyed in the endoreticular system, and the iron removed from hemoglobin is returned to the blood and reused. The body has no mechanism to actively excrete iron, and only a small amount of iron is lost through the shedding of epithelial cells in the gastrointestinal tract. On the other hand, the amount of iron absorbed from the gastrointestinal tract is small (about 1 mg/day in adults), and most of the iron is provided by recycled iron, creating a semi-closed loop.
A key player in iron homeostasis is hepcidin [5,6]. Hepcidin is a 25 amino acid peptide hormone produced from an 84 amino acid prepropeptide. Hepatocytes mainly secrete hepcidin, which binds to α2-macroglobulin and circulates in the plasma [7].
Dietary iron, both heme and non-heme, is absorbed by intestinal epithelial cells [8]. Non-heme iron, which is predominantly trivalent, is reduced to divalent iron by duodenal cytochrome b (Dcyt b) at the intestinal epithelial cell membrane and taken into the cells by divalent metal transporter 1 (DMT1) [9,10]. After its uptake by intestinal epithelial cells, heme iron is transported to the cytoplasm by heme carrier protein 1 (HCP1), which is degraded by heme oxygenase and released as iron ions [11,12]. Intracytoplasmic iron is exported into circulation by ferroportins on the basement membrane of the intestinal epithelium [13,14,15]. Iron released through the basement membrane by ferroportins is oxidized to trivalent iron by membrane-bound hephaestin or soluble ceruloplasmin, and then bound to transferrin (TF) in the blood for safe transport and distribution throughout the body [10,16,17,18] (Figure 2).
Transferrin receptor 1 (TfR1), which binds specifically to transferrin with high affinity, is highly expressed on the membrane surface of erythroblasts in the bone marrow, allowing transferrin-bound iron to be taken up by the cell. When transferrin binds to TfR1 on the cell membrane surface, it enters the cell by endocytosis, and iron is translocated from endosomes to the cytoplasm via DMT1 and is eventually used for hemoglobin production with the involvement of mitochondria [19]. When waste erythrocytes are trapped and destroyed by macrophages in the reticulospinal system, the resulting iron is released into the blood as divalent iron via FPN, which is then oxidized to trivalent iron by the oxidative action of ceruloplasmin (CP) and rebounds to transferrin for circulation and reuse in the body. Although TfR1 and its homologue transferrin receptor 2 (TfR2) have been implicated in the uptake of iron by hepatocytes [20], a pathway independent of these receptors has also been postulated [21]. Thus, iron metabolism in vivo is regulated by several related molecules, forming a semi-closed loop. Hepcidin is a small peptide consisting of 25 amino acids in its active form, which is mainly produced in the liver and circulates in blood. Hepcidin decreases the membrane surface expression of FPN in intestinal cells and the endoplasmic reticulum [5] and has been shown to be a general negative regulator of iron metabolism in vivo, inhibiting iron absorption in the gastrointestinal tract and release from macrophages [22,23].
Hepcidin is synthesized and secreted by the liver. Synthesized hepcidin is distributed throughout the body by the bloodstream and acts on cells expressing ferroportin (FPN), including small intestine, liver, and spleen macrophages. When hepcidin binds to FPN, it promotes FPN internalization and degradation, thereby inhibiting iron export from cells. In the small intestine, particularly in the duodenum, non-heme iron (Fe3+) from the diet is reduced to ferrous iron (Fe2+) by duodenal cytochrome b (Dcyt b), which is located on the luminal membrane of duodenal epithelial cells. The reduced iron is taken into the cell by the divalent metal transporter 1 (DMT1) and then exported to the bloodstream through the FPN on the basolateral side. Fe2+ exported from the FPN is oxidized to Fe3+ by membrane-bound hephaestin (HEPH) or soluble ceruloplasmin (CP). The oxidized iron rapidly binds to transferrin (Tf) in the blood and circulates as an Fe-Tf complex. Macrophages in the spleen play a critical role in phagocytosing senescent red blood cells and recovering iron from hemoglobin. FPN, also present on the macrophage surface, is responsible for releasing the recovered iron into the bloodstream, but hepcidin also controls this process. In inflammatory conditions, hepcidin production increases, resulting in systemic suppression of iron mobilization via FPN. This leads to decreased serum iron, sequestration of iron in reticuloendothelial macrophages, and restriction of iron available for hematopoiesis, resulting in anemia of chronic disease (ACD) [19].
【Iron Deficiency】
Iron deficiency is one of the most common nutritional issues worldwide. One in 49 people worldwide is anemic [24]. An iron imbalance is one of the causes of iron deficiency. In iron deficiency, daily iron loss and utilization exceed the daily iron intake [25]. In addition, disorders of iron metabolism, namely iron recycling from the spleen and macrophages, and iron absorption in the duodenum via hepcidin, are also associated with iron deficiency [25].
Iron is involved in many metabolic processes including mitochondrial electron transfer, neurotransmitter synthesis, protein synthesis, and organ formation. In addition, iron deficiency significantly reduces mitochondrial iron-sulfur content, cytochrome content, and total oxidative capacity [26]. Furthermore, iron deficiency limits hematopoiesis in hematopoietic tissues, leading to a decrease in hemoglobin concentration and resulting in iron deficiency anemia. Therefore, many biological functions, including the immune function, sympathetic nerve function, endocrine metabolism, and thermoregulation are reduced when iron is reduced in the body [27,28,29,30].
【Iron Deficiency in Athletes】
Iron deficiency (ID) is a common nutritional disorder among endurance athletes [31]. Iron is responsible for transporting oxygen to muscles during exercise and plays an important role in energy production during exercise [32]. Therefore, iron deficiency leads to impaired athletic performance during endurance events [33,34].
Iron deficiency is common in endurance athletes because of prolonged training and repetitive ground contact [31]. Several physiological mechanisms have been proposed to explain the reduced iron status, including gastrointestinal bleeding [35], hemolysis from the impact on the plantar surface and other areas [36], iron deficiency in the daily diet [37], and loss due to heavy sweating [38]. Iron storage deficiency occurs rapidly or very slowly, depending on the balance of the iron intake, storage, and iron requirements. In addition, the rate at which true iron deficiency occurs in individual tissues and intracellular organelles depends on the intracellular mechanisms of iron recycling and the metabolic turnover rate of iron-containing proteins [25].
As mentioned above, hepcidin regulates iron homeostasis by binding to ferroportin, the only known intracellular iron exporter, and inhibits its function through occlusion or degradation [39,40]. It has recently been reported to be associated with the onset of symptoms of apathy and negative mood disorders [41,42]. Consequently, hepcidin interferes with iron absorption and recycling. However, the detailed mechanisms underlying exercise-induced iron deficiency in athletes remain unclear.
【Exercise and Iron】
Hepcidin is a peptide hormone synthesized in the liver. The expression of hepcidin is upregulated by inflammation [43], increased iron stores [44], and exercise [45].
Banzet et al. observed that in rats subjected to treadmill exercise, liver hepcidin mRNA levels were elevated above resting levels immediately after exercise and returned to resting levels 6 h after exercise [45] Newlin et al. observed that plasma hepcidin levels increased significantly 3 h after exercise and returned to baseline levels 24 h after exercise [46]. Peeling et al. reported a significant increase in hepcidin mRNA levels in rats after 5 weeks of treadmill exercise. It has been suggested that hepcidin levels are influenced by exercise intensity and duration [47].
Intense exercise also increases the levels of the inflammatory cytokine interleukin-6 (IL-6), which stimulates hepcidin production. Serum hepcidin levels have also been found to increase approximately 3-6 h ours after a single bout of exercise [48,49]. Prolonged exercise also causes a marked increase in circulating IL-6 levels [50,51]. This has been suggested to be the mechanism responsible for the increase in hepcidin levels after exercise [51], and the mechanism of hepcidin expression from elevated IL-6 has been reported [52].
In recent years, mild to moderate anemia lasting more than a few months has been observed in patients with infectious, inflammatory, or neoplastic diseases, and is called anemia of inflammation or anemia of chronic disorder (ACD) [53]. This condition does not include chronic anemia due to chronic exanthema, hemolysis, renal failure, liver disease, endocrine disorders, and so on. Although anorexia-cachexia syndrome (ACD) is a widely used term, its meaning is somewhat ambiguous and the use of the term inflammatory anemia is recommended. Recently, hepcidin, a newly discovered endogenous antimicrobial substance, was shown to be closely involved in the development of this condition and has attracted renewed attention [54].
【Aerobic Exercise and Iron】
Because of the high prevalence of iron deficiency in persons with exercise habits, not only in the athletic population [31], and because many of these are aerobic exercise disciplines, much of the research on exercise and its effects on the body’s iron nutritional status has been conducted on aerobic exercise.
The proposed mechanisms by which iron status is affected by vigorous exercise include gastrointestinal bleeding after running and increased hematuria as a result of the destruction of red blood cells in the sole of the foot during running [55,56,57]. The possibility of increased erythrocyte metabolic turnover in athletes was also supported by iron kinetic measurements performed by Ehn et al., who showed that iron loss was 20% faster in female athletes than in non-athletes in experiments using radioactive iron and that iron loss was faster in adult males [58]. He also reported that female athletes had normal hemoglobin and plasma iron levels but a latent iron deficiency in the bone marrow [58]. Animal studies have reported that animals that exercise in a low-iron state have faster metabolic turnover of erythrocyte iron than those that do not exercise [59].
In contrast, animal studies have reported that aerobic exercise reduces or ameliorates the decline in in vivo iron indices caused by iron deficiency [60,61]. Perkkio et al. found that aerobic exercise (treadmill running) in iron-deficient rats significantly increased hemoglobin concentration, endurance capacity, and VO2max [62]. In contrast, in iron-sufficient rats, there was no decrease in hemoglobin concentration in the exercise group relative to the non-exercise group [62]. Willis et al. also reported that aerobic exercise (treadmill running) results in significantly higher hemoglobin concentrations in iron-deficient rats than in resting rats [63]. However, Gagine et al. observed a significant decrease in bone marrow iron in the exercise group relative to the resting group when rats were subjected to aerobic exercise (swimming exercise), suggesting that the decrease in bone marrow iron was due to increased iron metabolic turnover, including iron release from cells and the hemoglobin synthesis rate [64]. Qian et al. reported that in animals that underwent intense exercise (swimming exercise), the uptake of iron by bone marrow erythrocytes and hemoglobin synthesis increased, but tissue iron levels in the liver, spleen, kidneys, and heart decreased [65]. They also suggested that the decrease in tissue iron levels may be due to increased the uptake of iron into bone marrow cells for hemoglobin synthesis and that exercise may lead to a shift of iron from storage sites to bone marrow cells to promote hemoglobin synthesis [66]. Exercise-induced decreases in tissue iron content have been observed in other studies. Strause et al. also reported that exercised rats had lower liver and spleen iron than resting rats [66], and Ruckman et al. observed an increase in hemoglobin in exercised rats relative to resting rats [67]. However, they observed a trend toward iron deficiency in the liver and spleen, suggesting that decreased organ iron levels are associated with increased hemoglobin levels [67].
Nachitgall et al. investigated the blood status of 45 long-distance runners and reported that serum ferritin levels were < 35 μg/L in 51% of athletes and < 20 μg/L in 16% [68]. In addition, Simon et al. studied the blood status of professional soccer players and elite rowers and found that some players had normal hemoglobin levels, but serum ferritin levels below 12 μg/L (cutoff) [69]. In animal studies, iron deficiency in the absence of anemia has been shown to reduce endurance and exercise capacity. Hinton et al. also reported improved endurance when iron supplementation was administered to women with normal hemoglobin concentrations. Hinton et al. also reported improved exercise capacity when iron supplements were prescribed to women with normal hemoglobin and low serum ferritin levels [70]. In other words, the fact that hemoglobin concentrations were within the normal range does not mean that the iron intake was adequate. These reports suggest that assessing anemia by measuring the hemoglobin concentration alone may miss truly anemic individuals.
In recent years, serum and plasma ferritin concentrations have been commonly used to determine iron deficiency. The ferritin concentration in the blood reflects the amount of stored iron. The World Health Organization (WHO) recommends 15 ng/mL as the cutoff value of the serum ferritin concentration for iron deficiency in healthy individuals (age: 10-59 years) [71]. Many studies have also used a cutoff value of serum ferritin > 30 ng/mL, although studies related to iron deficiency and iron metabolism have used broader cutoff values, such as 12-40 ng/mL. However, a study investigating the association between serum ferritin, hepcidin, and iron absorption in young women recommended a ferritin cutoff value of 50 ng/mL for the early detection of iron deficiency [72]. Mielgo-Ayuso et al. also reported that 30–99 ng/mL was considered functional iron deficiency and that a serum ferritin level of ≥ 100 ng/mL was considered necessary [73]; therefore, the optimal cutoff value is still controversial.
【Resistance Exercise and Iron】
Compared to aerobic exercise, there are fewer studies on the effects of resistance exercise on iron metabolism. Resistance exercise is characterized by increased body protein synthesis [74] and muscle protein synthesis with faster post-exercise recovery [75,76,77,78].
Mild resistance exercise has been reported to improve subclinical iron deficiency in young women without iron supplementation [79]. In this study, 12 weeks of resistance exercise (dumbbell exercise) in women with subclinical iron deficiency resulted in significant increases in serum ferritin levels, hemoglobin concentration, red blood cell count, and total iron-binding capacity, suggesting that daily mild resistance exercise improves anemia and iron deficiency anemia.
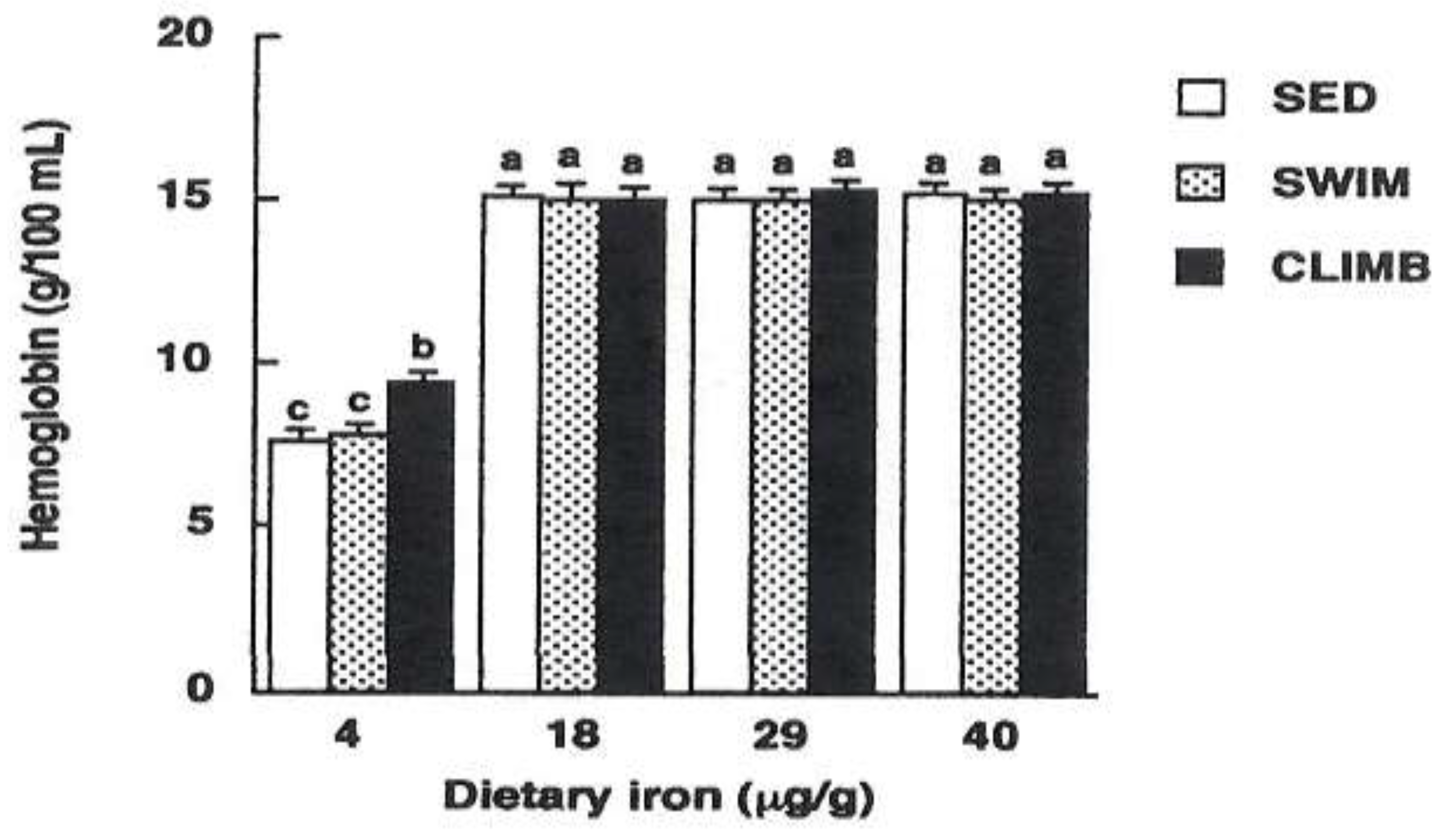
For example, Matsuo et al. reported that iron deficiency anemia was reduced in a study in which rats were subjected to a climbing exercise in which they voluntarily ascended and descended a 2-meter wire mesh tower cage for drinking water [78,80,81]. They suggested that this effect was related to an increase in ALAD activity, an enzyme involved in heme biosynthesis, due to resistance exercise [78,80,81]. Bone marrow ALAD activity increases after exercise in rats subjected to habitual climbing exercises. Matsuo et al. compared iron-deficient rats that performed habitual climbing or swimming exercise for 8 weeks and reported that they observed an increase in ALAD activity during climbing exercise, although swimming exercise did not alter bone marrow ALAD activity [78]. Climbing exercise has been shown to increase bone marrow ALAD activity, whereas aerobic exercise does not increase bone marrow ALAD activity. These findings suggest that resistance exercise, as opposed to aerobic exercise, may increase hemoglobin levels [81,82] (Figure 2). On the other hand, it has been suggested that even if climbing exercise increases heme biosynthetic capacity, blood hemoglobin concentration may not recover if the supply of iron, the source of hemoglobin, from diet or stored iron is insufficient [78,82,83].
It was observed that the blood hemoglobin concentration was improved and maintained in resistance-trained rats without a decrease in tissue iron concentration relative to resting rats [84]. In addition, there were no significant differences in blood iron indices or total body iron levels between groups of rats performing resistance exercise and those at rest. Furthermore, we observed that resistance exercise in rats increased iron excretion relative to resting rats [85]. These results suggest that the effect of resistance exercise on improving the iron nutritional status may not be due to increased iron absorption but rather to increased iron recycling. We speculate that this result is due to the fact that resistance exercise promotes iron recycling [82,83].
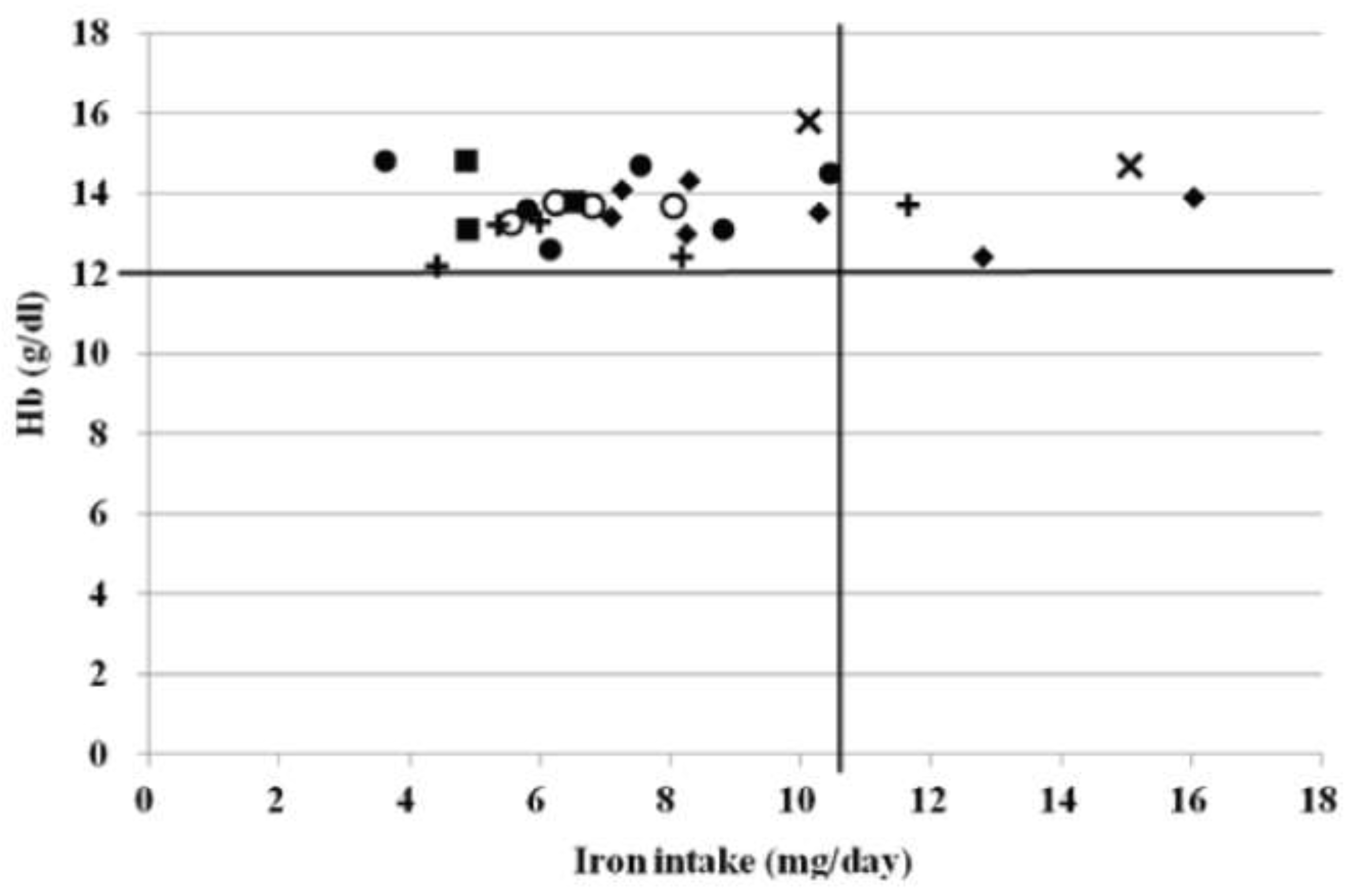
Therefore, because different types of exercise may have different effects on the iron nutritional status, we investigated the effects of different types of exercise on the iron nutritional status in athletes, focusing on track and field teams that have different exercise modalities, such as long-distance athletes who perform aerobic exercise and short-distance athletes and throwers who often perform strength training, which is a resistance exercise. This study examined the effects of different exercise modalities on the nutritional status of athletes. None of the male or female athletes in this study had anemia. It is believed that most men were not anemic because they consumed more than the recommended amount of iron. However, none of the females were anemic, although many did not consume the estimated average requirement (average iron intake: 8.1 ± 2.8 mg/day) (Figure 3). Short distance running, throwing, jumping, judo, and American football require strength and endurance, and strength training (resistance exercise) is normally incorporated into regular practice. For these reasons, resistance exercises are used in the training of athletes. These results suggest that resistance exercise increases iron recycling and may not induce anemia, even when the iron intake is low.
This result suggests that the iron intake varies with exercise type.
ALAS and ALAD, key enzymes in hemoglobin synthesis, were measured as indicators of heme biosynthetic capacity. When rats were subjected to 8 weeks of resistance exercise (climbing), bone marrow heme biosynthetic capacity increased, and the effect was greater than that of aerobic exercise. Previous experiments observing the effects of exercise on heme biosynthesis have inconsistently reported that habitual resistance exercise (climbing) increases heme biosynthesis, whereas no consistent conclusions have been drawn about the effects of habitual aerobic exercise.
【Nutrition and Exercise】
(1) Diet and Iron
The effects of various athletic activities on the iron status of college athletes have been studied [84,85]. Many male athletes were not anemic because they consumed more than the recommended amount of iron. Although many female athletes in the aforementioned study did not consume the estimated average requirement, none of them became anemic. Heme iron absorption was more efficient than non-heme iron absorption. There is also a protein (meat factor) that enhances iron absorption; meat factor increases the absorption of non-heme iron by almost a factor of 2. Lyle et al. reported that consumption of one meat-containing meal per day is effective in maintaining serum ferritin levels for long periods, even with moderate exercise [85]. Tetens et al. reported that serum ferritin levels decreased in women of childbearing age who consumed a plant-based diet, whereas serum ferritin was maintained in women who consumed a meat-based diet [86]. The subjects consumed more than 1.0 g of protein per kg of body weight. In addition, the animal protein intake was approximately 50% in both men and women. Based on the above, the subjects were considered to have an adequate meat intake, which may have increased iron absorption and prevented anemia [87].
Reinke et al. also suggested that the key physical factors of performance in rowing (i.e., power and endurance) may reduce the decline in stored iron relative to soccer, which requires speed and agility [88]. This may be one of the factors to consider for different iron intakes for different types of exercise.
(2) Diet and Nutrients
The first step in optimizing iron supplementation in athletes is to correct existing iron deficiency. Clenin et al. suggested that iron deficiency in athletes can be treated by dietary modification, oral supplementation, or intravenous/intramuscular supplementation [89]. Macrophages efficiently recycle old red blood cells, and under normal conditions, only 10% of the lost iron can be replenished by the diet [90,91]. It is important for athletes to consume meat, fish, whole grains, and green vegetables in their diet. In addition, foods rich in vitamin C increase iron absorption, whereas polyphenols found in coffee, tea, and certain plants inhibit iron absorption [91]. When iron deficiency occurs, the first step is usually to improve diet, aiming for an iron intake of 14 mg/day [89]. This number is approximately twice that of the general population in Japan. Furthermore, considering that the recommended iron intake for female endurance runners is 18 mg/day [92], this is a considerable burden for athletes in Japan, who are unable to achieve the iron recommendations of the general population.
Recently, the effects of the carbohydrate and energy intake on the hepcidin response in athletes have been investigated [93,94]. Decreased muscle glycogen content has been shown to increase exercise-induced IL-6 [93,95,96,97,98]; and McKay et al. found that both low energy availability (LEA) and a low carbohydrate intake increased the hepcidin response and that acute and long-term carbohydrate restriction had different effects on the hepcidin response and iron status [93]. They also concluded that while acute effects may influence the hepcidin response, the long-term approach is highly dependent on the athlete’s initial iron intake and not on their long-term carbohydrate intake. Although the results of this study may be considered a recent finding, it has been reported that the carbohydrate intake is also important for muscle protein synthesis [98]. To the best of our knowledge, it was reported as early as 1986 by Henderson et al., who reported increased utilization of glucose as an energy source in iron-deficient rats [99]. However, more research is required on this topic [100].
In recent years, in addition to iron deficiency anemia, relative energy deficit in sport (RED-S) has become a concern [101]. RED-S occurs in both males and females, resulting in decreased endurance, increased risk of injury, and decreased glycogen stores. Indeed, LEA is commonly observed during daily training in endurance athletes [102] and may increase the risk of iron deficiency [103]. It has also been suggested that severe energy deprivation may worsen hepcidin levels, even in the absence of inflammation [104]. Therefore, it can be said that nutritional deficiencies may indirectly affect iron status via increased hepcidin activity.
Low energy availability (LEA) caused by a decreased energy intake (EI) and/or increased exercise energy expenditure (EEE) has been reported to cause undesirable conditions such as decreased resting energy expenditure and disruption of various hormonal, metabolic, and functional properties [105]. However, the effects of LEA on iron metabolism, particularly hepcidin levels, during endurance exercise remain unknown.
A number of studies have examined the total energy intake in athletes and associated LEA with an increased hepcidin response [106]. Hennigar et al. found that athletes experiencing energy deprivation had an increased hepcidin response after exercise relative to athletes with a normal energy intake and found that iron absorption was reduced [106]. In addition, Barney et al. observed that in runners with low iron stores, prolonged running increased hepcidin levels and decreased dietary iron absorption compared with resting [107]. This suggests that maintaining an adequate energy balance may prevent exercise-induced increase in hepcidin response and reduce dietary iron absorption [108].
These results suggest that when appropriate for iron-deficient athletes, providing adequate energy and carbohydrates to meet training demands may be an important factor in optimizing an athlete’s iron status through diet [108].
【Influence of Exercise and Diet Timing on Iron Status in the Body】
The authors investigated the effect of meal timing on the effect of resistance exercise in improving iron nutritional status in iron-deficient rats [109]. Rats were subjected to resistance exercise and maintained for 3 weeks in two groups: one group received a meal immediately after exercise, and the other group received a meal 4 h after exercise. The results showed that plasma iron increased immediately after exercise in our study, and no increase in plasma iron due to the food intake was observed. However, both plasma iron levels and bone marrow ALAD activity increase after exercise. It has been reported that when anemic rats are subjected to resistance exercise, hemoglobin concentration increases significantly in comparison to resting rats [110]. It has also been reported that plasma iron is used for hemoglobin synthesis [111]. This suggests that the simultaneous increase in bone marrow ALAD activity and plasma iron levels indicates that resistance exercise promotes hemoglobin synthesis and increases hemoglobin concentration [77].
McCormick et al. found that despite the increase in hepcidin concentrations after exercise, more iron is absorbed when a meal is eaten after morning exercise than when fasting at rest or from an evening meal [109]. Although the physiological mechanisms that promote iron absorption after exercise remain to be elucidated, the present study suggests that, in addition to transient physiological changes after exercise, overall iron absorption is influenced by cumulative responses to inflammation and hepcidin. Therefore, they advocate that exercisers should be advised to take or supplement iron immediately after morning exercise to enhance iron absorption. This focuses on the fact that the increase in hepcidin occurs 3 h after exercise, regardless of whether exercise is performed in the morning or afternoon [110].
【Treatment and Iron】
In recent years, the use of iron supplements has become common as a preventive measure against anemia, not only for athletes, but also for people who do not exercise. Many types of supplements and other products to improve iron balance are available in the market. Brigham et al. reported that a supplement containing ferrous sulfate (39 mg/day of iron) prevented a decrease in serum ferritin in female swimmers [112]. Hinton et al. also reported that the administration of ferrous sulfate (36.8 mg/day iron) to late iron-deficient women significantly increased serum ferritin levels and improved endurance exercise capacity [112]. In addition, Kang et al. reported that supplementation of young female soccer players with iron (40 mg/day) significantly increased serum ferritin levels and prevented hemoglobin decline [113]. The doses of iron used in these studies were all low, suggesting that iron supplements and preparations can improve iron stores even at low doses. However, supplements can lead to overdosing because a certain amount is consumed regardless of the individual’s iron status. However, the side effects of oral iron supplementation may include gastrointestinal disturbances such as nausea, abdominal pain, and constipation [114]. These side effects may interfere with the daily performance and require careful consideration [115].
It is also recommended that athletes monitor their iron status regularly and treat deficiencies as soon as they occur [116,117,118], Clarke et al. recommend hematological testing every 6 months for women and annually for men unless there are other clinical indications [116]. However, no specific ferritin level is recommended for athletes to determine improvements in iron deficiency [119]. Clearer guidelines for iron supplementation and the associated hepcidin response to improve exercise performance are needed [120]. It has been reported that intravenous iron supplementation has no particular advantage over oral supplementation with respect to iron status in athletes [121,122,123]. It is well known that
【Our Latest Developments】
Hepcidin plays a central role in the pathophysiology of ACD by regulating the systemic iron homeostasis [124,125,126]. Hepcidin binds to ferroportin, promotes its internalization and degradation, and reduces cellular iron efflux [127]. In ACD, persistent inflammation increases hepcidin production, which in turn reduces the iron available for erythropoiesis and may lead to the development of anemia [128].
The regulation of the hepcidin expression is complex and involves multiple signaling pathways, including the BMP/SMAD, IL-6/STAT3, and HFE-TFR2 pathways [125]. Hemojuvelin (HJV), also known as repulsion-inducing molecule C (RGMc) or hemochromatosis type 2 protein (HFE2), plays an important role in regulating these pathways and, ultimately the expression of hepcidin [129,130]. HJV is a co-receptor for BMP and acts as a co-receptor for BMPs, enhancing BMP signaling and promoting hepcidin transcription [129,130,131].
HJV in hepcidin regulation is not a new area, but is often overlooked in sports nutrition. In fact, much of the research has focused on the association of hepcidin with IL-6 and nutrients, and we believe that new methods of preventing anemia around HJV would be beneficial not only to athletes but to non-athletes.
Therefore, we conducted a comprehensive study on the involvement of the RGM family in hepcidin regulation in ACD.
Figure 4.
The relationship between the daily iron intake and hemoglobin concentration in females. ○: sprinting, ●: throwing, x: jumping, +: middle and long distance running, ♦: judo, ∎: gymnastics. The black transverse line indicates the cutoff value for the hemoglobin concentration. The black vertical line indicates the recommended dietary allowance (RDA) for females.
Figure 4.
The relationship between the daily iron intake and hemoglobin concentration in females. ○: sprinting, ●: throwing, x: jumping, +: middle and long distance running, ♦: judo, ∎: gymnastics. The black transverse line indicates the cutoff value for the hemoglobin concentration. The black vertical line indicates the recommended dietary allowance (RDA) for females.
Date Availability Statement
All information and findings presented in this review are based on previously published literature and are cited throughout the manuscript. No new datasets were generated or analyzed in this study.
Author Contributions
Conceptualization, TF; Review of bibliographies, TF, KH and KO; Creation of figures, TF; Writing—review and editing, RT, TF and KH. All authors have read and agreed to the published version of the manuscript.
Funding
The authors did not receive support from any organization for the submitted work.
References
- Muñoz Gómez, M.; Campos Garríguez, A.; García Erce, J.A.; Ramírez Ramírez, G. Fisiopatología del metabolismo del hierro: implicaciones diagnósticas y terapéuticas [Fisiopathology of iron metabolism: diagnostic and therapeutic implications]. Nefrologia : publicacion oficial de la Sociedad Espanola Nefrologia 2005, 25, 9–19. [Google Scholar] [PubMed]
- Andrews, N. C. Disorders of iron metabolism. The New England journal of medicine 1986, 341, 1986–1995. [Google Scholar] [CrossRef] [PubMed]
- Siah, C.W.; Ombiga, J.; Adams, L.A.; Trinder, D.; Olynyk, J.K. Normal iron metabolism and the pathophysiology of iron overload disorders. The Clinical biochemist. Reviews 2006, 27, 5–16. [Google Scholar]
- Fleming, R.E.; Bacon, B.R. Orchestration of iron homeostasis. The New England journal of medicine 2005, 352, 1741–1744. [Google Scholar] [CrossRef]
- Ganz, T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood 2003, 102, 783–788. [Google Scholar] [CrossRef]
- Lee, P.L.; Beutler, E. Regulation of hepcidin and iron-overload disease. Annu. Rev. Pathol 2009, 4, 489–515. [Google Scholar] [CrossRef]
- Peslova, G.; Petrak, J.; Kuzelova, K.; Hrdy, I.; Halada, P.; Kuchel, P.W.; Soe-Lin, S.; Ponka, P.; Sutak, R.; Becker, E.; et al. Hepcidin, the hormone of iron metabolism, is bound specifically to alpha-2-macroglobulin in blood. Blood 2009, 113, 6225–6236. [Google Scholar] [CrossRef]
- Nicolas, G.; Bennoun, M.; Devaux, I.; Beaumont, C.; Grandchamp, B.; Kahn, A.; Vaulont, S. Lack of hepcidin gene expression and severe tissue iron overload in upstream stimulatory factor 2 (USF2) knockout mice. Proc. Natl. Acad. Sci. USA 2001, 98, 8780–8785. [Google Scholar] [CrossRef]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef]
- Hentze, M.W.; Muckenthaler, M.U.; Galy, B.; Camaschella, C. Two to tango: Regulation of Mammalian iron metabolism. Cell 2010, 142, 24–38. [Google Scholar] [CrossRef]
- Shayeghi, M.; Latunde-Dada, G.O.; Oakhill, J.S.; Laftah, A.H.; Takeuchi, K.; Halliday, N.; Khan, Y.; Warley, A.; McCann, F.E.; Hider, R.C.; et al. Identification of an intestinal heme transporter. Cell 2005, 122, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Raffin, S.B.; Woo, C.H.; Roost, K.T.; Price, D.C.; Schmid, R. Intestinal absorption of hemoglobin iron-heme cleavage by mucosal heme oxygenase. J. Clin. Investig. 1974, 54, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Abboud, S.; Haile, D.J. A novel mammalian iron-regulated protein involved in intracellular iron metabolism. J. Biol. Chem. 2000, 275, 19906–19912. [Google Scholar] [CrossRef] [PubMed]
- Donovan, A.; Brownlie, A.; Zhou, Y.; Shepard, J.; Pratt, S.J.; Moynihan, J.; Paw, B.H.; Drejer, A.; Barut, B.; Zapata, A.; et al. Positional cloning of zebrafish ferroportin1 identifies a conserved vertebrate iron exporter. Nature 2000, 403, 776–781. [Google Scholar] [CrossRef]
- McKie, A.T.; Marciani, P.; Rolfs, A.; Brennan, K.; Wehr, K.; Barrow, D.; Miret, S.; Bomford, A.; Peters, T.J.; Farzaneh, F.; et al. A novel duodenal iron-regulated transporter, IREG1, implicated in the basolateral transfer of iron to the circulation. Mol. Cell 2000, 5, 299–309. [Google Scholar]
- Vulpe, C.D.; Kuo, Y.M.; Murphy, T.L.; Cowley, L.; Askwith, C.; Libina, N.; Gitschier, J.; Anderson, G.J. Hephaestin, a ceruloplasmin homologue implicated in intestinal iron transport, is defective in the sla mouse. Nat. Genet. 1999, 21, 195–199. [Google Scholar] [CrossRef]
- Harris, Z.L.; Durley, A.P.; Man, T.K.; Gitlin, J.D. Targeted gene disruption reveals an essential role for ceruloplasmin in cellular iron efflux. Proc. Natl. Acad. Sci. USA 1999, 96, 10812–10817. [Google Scholar] [CrossRef]
- Aisen PTransferrin receptor 1. The international journal of biochemistry & cell biology 2004, 36, 2137–2143.
- Fujii T, Kobayashi K, Kaneko M, Osana S, Tsai CT, Ito S, Hata. RGM Family Involved in the Regulation of Hepcidin Expression in Anemia of Chronic Disease. Immuno 2024, 4, 266–285.
- Kawabata, H.; Yang, R.; Hirama, T.; et al. Molecular cloning of transferrin receptor 2. A new member of the transferrin receptor-like family. J Biol Chem 1999, 274, 20826–20832. [Google Scholar] [CrossRef]
- Ikuta, K.; Zak, O.; Aisen, P. Recycling, degradation and sensitivity to the synergistic anion of transferrin in the receptor-independent route of iron uptake by human hepatoma (HuH-7) cells. The international journal of biochemistry & cell biology 2004, 36, 340–352. [Google Scholar]
- Andrews, N. C. Forging a field: the golden age of iron biology. Blood 2008, 112, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Fleming, R.E.; Sly, W.S. Hepcidin: a putative iron-regulatory hormone relevant to hereditary hemochromatosis and the anemia of chronic disease. Proceedings of the National Academy of Sciences of the United States of America 2001, 98, 8160–8162. [Google Scholar] [CrossRef]
- WHO 2019. Prevalence of anaemia in women of reproductive age (aged 15–49) (%). Retrieved from: https://www.who.int/data/gho/data/indicators/indicator-details/GHO/prevalence-of-anaemia-in-women-of-reproductive-age-(-).
- Cappellini, M.D.; Musallam, K.M.; Taher, A.T. Iron deficiency anaemia revisited. Journal of internal medicine 2020, 287, 153–170. [Google Scholar] [CrossRef]
- Paul, B.T.; Manz, D.H.; Torti, F.M.; Torti, S.V. Mitochondria and Iron: current questions. Expert review of hematology 2017, 10, 65–79. [Google Scholar] [CrossRef]
- Dallman, P. R. Manifestations of iron deficiency. Seminars in hematology 1982, 19, 19–30. [Google Scholar]
- Dallman, P. R. Biochemical basis for the manifestations of iron deficiency. Annual review of nutrition 1986, 6, 13–40. [Google Scholar] [CrossRef] [PubMed]
- Scrimshaw, N. S. Functional consequences of iron deficiency in human populations. Journal of nutritional science and vitaminology 1984, 30, 47–63. [Google Scholar] [CrossRef]
- Cook, J.D.; Lynch, S.R. The liabilities of iron deficiency. Blood 1986, 68, 803–809. [Google Scholar] [CrossRef]
- Beard, J.; Tobin, B. Iron status and exercise. The American journal of clinical nutrition 2000, 72(2 Suppl), 594S–7S. [Google Scholar] [CrossRef]
- Lukaski, H.C. Vitamin and mineral status: effects on physical performance. Nutrition (Burbank, Los Angeles County, Calif.) 2004, 20, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Celsing, F.; Ekblom, B. Anemia causes a relative decrease in blood lactate concentration during exercise. European journal of applied physiology and occupational physiology 1986, 55, 74–78. [Google Scholar] [CrossRef]
- Haas, J.D.; Brownlie, T. 4th Iron deficiency and reduced work capacity: a critical review of the research to determine a causal relationship. The Journal of nutrition 2001, 131, 676S–690S. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.G.; Ahlquist, D.A.; McGill, D.B.; Ilstrup, D.M.; Schwartz, S.; Owen, R.A. Gastrointestinal blood loss and anemia in runners. Annals of internal medicine 1984, 100, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.J.; Pate, R.R.; Burgess, W. Foot impact force and intravascular hemolysis during distance running. International journal of sports medicine 1988, 9, 56–60. [Google Scholar] [CrossRef] [PubMed]
- King, N.; Fridlund, K.E.; Askew, E.W. Nutrition issues of military women. Journal of the American College of Nutrition 1993, 12, 344–348. [Google Scholar] [CrossRef]
- Brune, M.; Magnusson, B.; Persson, H.; Hallberg, L. Iron losses in sweat. Am. J. Clin. Nutr. 1986, 43, 438–443. [Google Scholar] [CrossRef]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science (New York, N.Y.) 2004, 306, 2090–2093. [Google Scholar] [CrossRef]
- Nemeth, E.; Ganz, T. Hepcidin-Ferroportin Interaction Controls Systemic Iron Homeostasis. International journal of molecular sciences 2021, 22, 6493. [Google Scholar] [CrossRef]
- Sawada, T.; Konomi, A.; Yokoi, K. Iron deficiency without anemia is associated with anger and fatigue in young Japanese women. Biological trace element research 2014, 159, 22–31. [Google Scholar] [CrossRef]
- Sim, M.; Garvican-Lewis, L.A.; Cox, G.R.; Govus, A.; McKay, A.K.A.; Stellingwerff, T.; Peeling, P. Iron considerations for the athlete: a narrative review. European journal of applied physiology 2019, 119, 1463–1478. [Google Scholar] [CrossRef] [PubMed]
- Recalcati, S.; Minotti, G.; Cairo, G. Iron regulatory proteins: from molecular mechanisms to drug development. Antioxidants & redox signaling 2010, 13, 1593–1616. [Google Scholar]
- Nemeth, E.; Valore, E.V.; Territo, M.; Schiller, G.; Lichtenstein, A.; Ganz, T. Hepcidin, a putative mediator of anemia of inflammation, is a type II acute-phase protein. Blood 2003, 101, 2461–2463. [Google Scholar] [CrossRef]
- Banzet, S.; Sanchez, H.; Chapot, R.; Bigard, X.; Vaulont, S.; Koulmann, N. Interleukin-6 contributes to hepcidin mRNA increase in response to exercise. Cytokine 2012, 58, 158–161. [Google Scholar] [CrossRef]
- Weiss, G.; Goodnough, L.T. Anemia of chronic disease. The New England journal of medicine 2005, 352, 1011–1023. [Google Scholar] [CrossRef]
- Peeling, P.; Dawson, B.; Goodman, C.; Landers, G.; Wiegerinck, E.T.; Swinkels, D.W.; Trinder, D. Cumulative effects of consecutive running sessions on hemolysis, inflammation and hepcidin activity. European journal of applied physiology 2009, 106, 51–59. [Google Scholar] [CrossRef]
- Peeling, P.; Dawson, B.; Goodman, C.; Landers, G.; Wiegerinck, E.T.; Swinkels, D.W.; Trinder, D. Training surface and intensity: inflammation, hemolysis, and hepcidin expression. Medicine and science in sports and exercise 2009, 41, 1138–1145. [Google Scholar] [CrossRef]
- Newlin, M.K.; Williams, S.; McNamara, T.; Tjalsma, H.; Swinkels, D.W.; Haymes, E.M. The effects of acute exercise bouts on hepcidin in women. International journal of sport nutrition and exercise metabolism 2012, 22, 79–88. [Google Scholar] [CrossRef]
- Ronsen, O.; Lea, T.; Bahr, R.; Pedersen, B.K. Enhanced plasma IL-6 and IL-1ra responses to repeated vs. single bouts of prolonged cycling in elite athletes. Journal of applied physiology (ethesda, Md. : 1985) 2002, 92, 2547–2553. [Google Scholar] [CrossRef]
- Banzet, S.; Sanchez, H.; Chapot, R.; Bigard, X.; Vaulont, S.; Koulmann, N. Interleukin-6 contributes to hepcidin mRNA increase in response to exercise. Cytokine 2012, 58, 158–161. [Google Scholar] [CrossRef]
- Means R: The anemia of chronic disorders. Wintrobe’s Clinical Hematology. Lee R et al, eds Williams & Willkins 1999, 1011-1021.
- Andrews, N. C. Anemia of inflammation: the cytokine-hepcidin link. The Journal of clinical investigation 2004, 113, 1251–1253. [Google Scholar] [CrossRef] [PubMed]
- Siegel, A.J.; Hennekens, C.H.; Solomon, H.S.; Van Boeckel, B. Exercise-related hematuria. Findings in a group of marathon runners. JAMA 1979, 241, 391–392. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.G.; Ahlquist, D.A.; McGill, D.B.; Ilstrup, D.M.; Schwartz, S.; Owen, R.A. Gastrointestinal blood loss and anemia in runners. Annals of internal medicine 1984, 100, 843–845. [Google Scholar] [CrossRef]
- Brune, M.; Magnusson, B.; Persson, H.; Hallberg, L. Iron losses in sweat. The American journal of clinical nutrition 1986, 43, 438–443. [Google Scholar] [CrossRef]
- Ehn, L.; Carlmark, B.; Höglund, S. Iron status in athletes involved in intense physical activity. Medicine and science in sports and exercise 1980, 12, 61–64. [Google Scholar] [CrossRef]
- Tobin, B.W.; Beard, J.L. Interactions of iron deficiency and exercise training in male Sprague-Dawley rats: ferrokinetics and hematology. The Journal of nutrition 1989, 119, 1340–1347. [Google Scholar] [CrossRef]
- Radomski, M.W.; Sabiston, B.H.; Isoard, P. Development of „sports anemia” in physically fit men after daily sustained submaximal exercise. Aviation, space, and environmental medicine 1980, 51, 41–45. [Google Scholar]
- Nicolas, G.; Chauvet, C.; Viatte, L.; Danan, J.L.; Bigard, X.; Devaux, I.; Beaumont, C.; Kahn, A.; Vaulont, S. The gene encoding the iron regulatory peptide hepcidin is regulated by anemia, hypoxia, and inflammation. The Journal of clinical investigation 2002, 110, 1037–1044. [Google Scholar] [CrossRef]
- Perkkiö, M.V.; Jansson, L.T.; Henderson, S.; Refino, C.; Brooks, G.A.; Dallman, P.R. Work performance in the iron-deficient rat: improved endurance with exercise training. The American journal of physiology 1985, 249(3 Pt 1), E306–E311. [Google Scholar] [CrossRef]
- Willis, W.T.; Brooks, G.A.; Henderson, S.A.; Dallman, P.R. Effects of iron deficiency and training on mitochondrial enzymes in skeletal muscle. Journal of applied physiology (Bethesda, Md. : 1985) 1987, 62, 2442–2446. [Google Scholar] [CrossRef]
- Gagne, C.M.; Walberg-Rankin, J.L.; Ritchey, S.J. Effects of exercise on iron status in mature female rats. Nutr, Res 2014, 14, 211–219. [Google Scholar] [CrossRef]
- Ming Qian, Z.; Sheng Xiao, D.; Kui Liao, Q.; Ping Ho, K. (2002). Effect of different durations of exercise on transferrin-bound iron uptake by rat erythroblast. The Journal of nutritional biochemistry 2002, 13, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Strause, L.; Hegenauer, J.; Saltman, P. Effects of exercise on iron metabolism in rats. Nutr Res 1983, 79–89. [Google Scholar] [CrossRef]
- Ruckman, K.S.; Sherman, A.R. Effects of exercise on iron and copper metabolism in rats. The Journal of nutrition 1981, 111, 1593–1601. [Google Scholar] [CrossRef]
- Nachtigall, D.; Nielsen, P.; Fischer, R.; Engelhardt, R.; Gabbe, E.E. Iron deficiency in distance runners. A reinvestigation using Fe-labelling and non-invasive liver iron quantification. International journal of sports medicine 1996, 17, 473–479. [Google Scholar] [CrossRef]
- Reinke, S.; Taylor, W.R.; Duda, G.N.; von Haehling, S.; Reinke, P.; Volk, H.D.; Anker, S.D.; Doehner, W. Absolute and functional iron deficiency in professional athletes during training and recovery. International journal of cardiology 2012, 156, 186–191. [Google Scholar] [CrossRef]
- Hinton, P. S. Iron and the endurance athlete. Applied physiology, nutrition, and metabolism = Physiologie appliquee, nutrition et metabolisme 2014, 39, 1012–1018. [Google Scholar] [CrossRef]
- Kuwabara, A.M.; Tenforde, A.S.; Finnoff, J.T.; Fredericson, M. Iron deficiency in athletes: A narrative review. PM & R : the journal of injury, function, and rehabilitation 2022, 14, 620–642. [Google Scholar]
- Galetti, V.; Stoffel, N.U.; Sieber, C.; Zeder, C.; Moretti, D.; Zimmermann, M.B. Threshold ferritin and hepcidin concentrations indicating early iron deficiency in young women based on upregulation of iron absorption. EClinicalMedicine 2021, 39, 101052. [Google Scholar] [CrossRef]
- Mielgo-Ayuso, J.; Zourdos, M.C.; Calleja-González, J.; Córdova, A.; Fernandez-Lázaro, D.; Caballero-García, A. Eleven Weeks of Iron Supplementation Does Not Maintain Iron Status for an Entire Competitive Season in Elite Female Volleyball Players: A Follow-Up Study. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Yang, R.C.; Mack, G.W.; Wolfe, R.R.; Nadel, E.R. Albumin synthesis after intense intermittent exercise in human subjects. Journal of applied physiology (Bethesda, Md. : 1985) 1998, 84, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Doi, T.; Hamada, K.; Sakurai, M.; Matsumoto, K.; Imaizumi, K.; Yoshioka, Y.; Shimizu, S.; Suzuki, M. Effect of amino acid and glucose administration during postexercise recovery on protein kinetics in dogs. The American journal of physiology 1997, 272(6 Pt 1), E1023–E1030. [Google Scholar] [CrossRef]
- Esmarck, B.; Andersen, J.L.; Olsen, S.; Richter, E.A.; Mizuno, M.; Kjaer, M. Timing of postexercise protein intake is important for muscle hypertrophy with resistance training in elderly humans. The Journal of physiology 2001, 535 (Pt 1), 301–311. [Google Scholar] [CrossRef]
- Levenhagen, D.K.; Gresham, J.D.; Carlson, M.G.; Maron, D.J.; Borel, M.J.; Flakoll, P.J. Postexercise nutrient intake timing in humans is critical to recovery of leg glucose and protein homeostasis. American journal of physiology. Endocrinology and metabolism 2001, 280, E982–E993. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Suzuki, H.; Suzuki, M. Resistance Exercise Increases the Capacity of Heme Biosynthesis More Than Aerobic Exercise in Rats. J Clin Biochem Nutr 2000, 29, 19–27. [Google Scholar] [CrossRef]
- Matsuo, T.; Suzuki, H.; Suzuki, M. Dubbell exercise improves non-anemic iron deficiency in young women without iron supplementation. Health Sci 2000, 16, 236–243. [Google Scholar]
- Matsuo, T.; Kang, H.S.; Suzuki, H.; Suzuki, M. Voluntary resistance exercise improves blood hemoglobin concentration in severely iron-deficient rats. Journal of nutritional science and vitaminology 2002, 48, 161–164. [Google Scholar] [CrossRef]
- Matsuo, T. Effects of resistance exercise on iron metabolism in iron-adequate or iron-deficient rats. The Korean Joumal of Exercise Nutrition 2004, 8, 1–15. [Google Scholar]
- Fujii, T.; Matsuo, T.; Okamura, K. Effects of resistance exercise on iron absorption and balance in iron-deficient rats. Biological trace element research 2014, 161, 101–106. [Google Scholar] [CrossRef]
- Fujii, T.; Asai, T.; Matsuo, T.; Okamura, K. Effect of resistance exercise on iron status in moderately iron-deficient rats. Biological trace element research 2011, 144, 983–991. [Google Scholar] [CrossRef]
- Fujii, T.; Okumura, Y.; Maeshima, E.; Okamura, K. Dietary Iron Intake and Hemoglobin Concentration in College Athletes in Different Sports. Int J Sports Exerc Med 2015, 1, 5. [Google Scholar] [CrossRef]
- Taguchi, M.; Ishikawa, T.K.; Tatsuta, W.; Katsuragi, C.; et al. Resting energy expenditure can be assessed by fat-free mass in female athletes regardless of body size. Journal of Nutritional Science and Vitaminology 2011. [Google Scholar] [CrossRef] [PubMed]
- Lyle, R.M.; Weaver, C.M.; Sedlock, D.A.; Rajaram, S.; Martin, B.; Melby, C.L. Iron status in exercising women: the effect of oral iron therapy vs increased consumption of muscle foods. The American journal of clinical nutrition 1992, 56. [Google Scholar] [CrossRef] [PubMed]
- Tetens, I.; Bendtsen, K.M.; Henriksen, M.; Ersbøll, A.K.; Milman, N. The impact of a meat- versus a vegetable-based diet on iron status in women of childbearing age with small iron stores. European journal of nutrition 2007, 46, 439–445. [Google Scholar] [CrossRef]
- Reinke, S.; Taylor, W.R.; Duda, G.N.; von Haehling, S.; Reinke, P.; Volk, H.D.; Anker, S.D.; Doehner, W. Absolute and functional iron deficiency in professional athletes during training and recovery. International journal of cardiology 2012, 156, 186–191. [Google Scholar] [CrossRef]
- Clenin, G.; Cordes, M.; Huber, A.; Schumacher, Y.O.; Noack, P.; Scales, J.; Kriemler, S. Iron deficiency in sports—Definition, influence on performance and therapy. Swiss Med. Wkly. 2015, 145, w14196. [Google Scholar] [CrossRef]
- Dominguez, R.; Sanchez-Oliver, A.J.; Mata-Ordonez, F.; Feria-Madueno, A.; Grimaldi-Puyana, M.; Lopez-Samanes, A.; Perez-Lopez, A. Effects of an Acute Exercise Bout on Serum Hepcidin Levels. Nutrients 2018, 10, 209. [Google Scholar] [CrossRef]
- Ems, T.; St Lucia, K.; Huecker, M.R. Biochemistry, Iron Absorption. In StatPearls; Ineligible Companies: Treasure Island, FL, USA, 2023. [Google Scholar]
- Deldicque, L.; Francaux, M. Recommendations for Healthy Nutrition in Female Endurance Runners: An Update. Frontiers in nutrition 2015, 2, 17. [Google Scholar] [CrossRef]
- McKay, A.K.A.; Pyne, D.B.; Burke, L.M.; Peeling, P. Iron Metabolism: Interactions with Energy and Carbohydrate Availability. Nutrients 2020, 12, 3692. [Google Scholar] [CrossRef]
- Hayashi, N.; Ishibashi, A.; Iwata, A.; Yatsutani, H.; Badenhorst, C.; Goto, K. Influence of an energy deficient and low carbohydrate acute dietary manipulation on iron regulation in young females. Physiol. Rep. 2022, 10, e15351. [Google Scholar] [CrossRef]
- Burke, L. M. Ketogenic low-CHO, high-fat diet: the future of elite endurance sport? The Journal of physiology 2021, 599. [Google Scholar] [CrossRef] [PubMed]
- Steensberg, A.; Febbraio, M.A.; Osada, T.; Schjerling, P.; van Hall, G.; Saltin, B.; Pedersen, B.K. Interleukin-6 production in contracting human skeletal muscle is influenced by pre-exercise muscle glycogen content. J. Physiol 2001, 537, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Badenhorst, C.E.; Black, K.E.; O’Brien, W.J. Hepcidin as a Prospective Individualized Biomarker for Individuals at Risk of Low Energy Availability. International journal of sport nutrition and exercise metabolism 2019, 29, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Badenhorst, C.E.; Dawson, B.; Cox, G.R.; Laarakkers, C.M.; Swinkels, D.W.; Peeling, P. Acute dietary carbohydrate manipulation and the subsequent inflammatory and hepcidin responses to exercise. Eur. J. Appl. Physiol. 2015, 115, 2521–2530. [Google Scholar] [CrossRef]
- Henderson, S.A.; Dallman, P.R.; Brooks, G.A. Glucose turnover and oxidation are increased in the iron-deficient anemic rat. The American journal of physiology 1986, 250(4 Pt 1), E414–E421. [Google Scholar] [CrossRef]
- Ishibashi, A.; Kojima, C.; Tanabe, Y.; Iwayama, K.; Hiroyama, T.; Tsuji, T.; Kamei, A.; Goto, K.; Takahashi, H. Effect of low energy availability during three consecutive days of endurance training on iron metabolism in male long distance runners. Physiol. Rep. 2020, 8, e14494. [Google Scholar] [CrossRef]
- Statuta, S.M.; Asif, I.M.; Drezner, J.A. Relative energy deficiency in sport (RED-S). British journal of sports medicine 2017, 51, 1570–1571. [Google Scholar] [CrossRef]
- Loucks, A. B. Low energy availability in the marathon and other endurance sports. Sports medicine (Auckland, N.Z.) 2007, 37, 348–352. [Google Scholar] [CrossRef]
- Petkus, D.L.; Murray-Kolb, L.E.; De Souza, M.J. The Unexplored Crossroads of the Female Athlete Triad and Iron Deficiency: A Narrative Review. Sports medicine (Auckland, N.Z.) 2017, 47, 1721–1737. [Google Scholar] [CrossRef]
- Papillard-Marechal, S.; Sznajder, M.; Hurtado-Nedelec, M.; Alibay, Y.; Martin-Schmitt, C.; Dehoux, M.; Westerman, M.; Beaumont, C.; Chevallier, B.; Puy, H.; et al. Iron metabolism in patients with anorexia nervosa: elevated serum hepcidin concentrations in the absence of inflammation. The American journal of clinical nutrition 2012, 95, 548–554. [Google Scholar] [CrossRef]
- Mountjoy, M.L.; Burke, L.M.; Stellingwerff, T.; Sundgot-Borgen, J. Relative Energy Deficiency in Sport: The Tip of an Iceberg. International journal of sport nutrition and exercise metabolism 2018, 28, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Hennigar, S.R.; McClung, J.P.; Hatch-McChesney, A.; Allen, J.T.; Wilson, M.A.; Carrigan, C.T.; Murphy, N.E.; Teien, H.K.; Martini, S.; Gwin, J.A.; et al. Energy deficit increases hepcidin and exacerbates declines in dietary iron absorption following strenuous physical activity: a randomized-controlled cross-over trial. The American journal of clinical nutrition 2021, 113, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Barney, D.E.; Ippolito, J.R.; Berryman, C.E.; Hennigar, S.R. A Prolonged Bout of Running Increases Hepcidin and Decreases Dietary Iron Absorption in Trained Female and Male Runners. The Journal of nutrition 2022, 152, 2039–2047. [Google Scholar] [CrossRef]
- Clarke, A.C.; Anson, J.M.; Dziedzic, C.E.; Mcdonald, W.A.; Pyne, D.B. Iron monitoring of male and female rugby seven players over an international season. The Journal of sports medicine and physical fitness 2018, 58, 1490–1496. [Google Scholar] [CrossRef]
- Fujii, T.; Matsuo, T.; Okamura, K. The effects of resistance exercise and post-exercise meal timing on the iron status in iron-deficient rats. Biological trace element research 2012, 147, 200–205. [Google Scholar] [CrossRef]
- McCormick, R.; Moretti, D.; McKay, A.K.A.; Laarakkers, C.M.; Vanswelm, R.; Trinder, D.; Cox, G.R.; Zimmerman, M.B.; Sim, M.; Goodman, C.; et al. The Impact of Morning versus Afternoon Exercise on Iron Absorption in Athletes. Medicine and science in sports and exercise 2019, 51, 2147–2155. [Google Scholar] [CrossRef]
- Brigham, D.E.; Beard, J.L.; Krimmel, R.S.; Kenney, W.L. Changes in iron status during competitive season in female collegiate swimmers. Nutrition 1993, 9, 418–422. [Google Scholar]
- Hinton, P. S. Iron and the endurance athlete. Applied physiology, nutrition, and metabolism = Physiologie appliquee, nutrition et metabolisme 2014, 39, 1012–1018. [Google Scholar] [CrossRef]
- Kang, H.S.; Matsuo, T. Effects of 4 weeks iron supplementation on haematological and immunological status in elite female soccer players. Asia Pacific journal of clinical nutrition 2004, 13, 353–358. [Google Scholar]
- Grzywacz, A.; Lubas, A.; Fiedor, P.; Fiedor, M.; Niemczyk, S. AFETY AND EFFICACY OF INTRAVENOUS ADMINISTRATION OF IRON PREPARATIONS. Acta poloniae pharmaceutica 2017, 74, 13–24. [Google Scholar]
- Low, M.S.; Speedy, J.; Styles, C.E.; De-Regil, L.M.; Pasricha, S.R. Daily iron supplementation for improving anaemia, iron status and health in menstruating women. The Cochrane database of systematic reviews 2016, 4, CD009747. [Google Scholar] [CrossRef] [PubMed]
- Fallon, K.E. Utility of hematological and iron-related screening in elite athletes. Clin. J. Sport Med. 2004, 14, 145–152. [Google Scholar] [CrossRef]
- Sim, M.; Garvican-Lewis, L.A.; Cox, G.R.; Govus, A.; McKay, A.K.A.; Stellingwerff, T.; Peeling, P. Iron considerations for the athlete: A narrative review. Eur. J. Appl. Physiol. 2019, 119, 1463–1478. [Google Scholar] [CrossRef]
- Rodenberg, R.E.; Gustafson, S. Iron as an ergogenic aid: Ironclad evidence? Curr. Sports Med. Rep. 2007, 6, 258–264. [Google Scholar] [CrossRef]
- Solberg, A.; Reikvam, H. Iron Status and Physical Performance in Athletes. Life (Basel, Switzerland) 2023, 13, 2007. [Google Scholar] [CrossRef]
- Garvican-Lewis, L.A.; Vuong, V.L.; Govus, A.D.; Peeling, P.; Jung, G.; Nemeth, E.; Hughes, D.; Lovell, G.; Eichner, D.; Gore, C.J. Intravenous Iron Does Not Augment the Hemoglobin Mass Response to Simulated Hypoxia. Med. Sci. Sports Exerc 2018, 50, 1669–1678. [Google Scholar] [CrossRef]
- Friedrisch, J.R.; Cancado, R.D. Intravenous ferric carboxymaltose for the treatment of iron deficiency anemia. Rev. Bras. Hematol. Hemoter 2015, 37, 400–405. [Google Scholar] [CrossRef]
- Girelli, D.; Ugolini, S.; Busti, F.; Marchi, G.; Castagna, A. Modern iron replacement therapy: Clinical and pathophysiological insights. Int. J. Hematol 2018, 107, 16–307. [Google Scholar] [CrossRef]
- Fraenkel, P.G. Understanding anemia of chronic disease. Hematol. Am. Soc. Hematol. Educ. Progra. 2015, 2015, 14–18. [Google Scholar] [CrossRef]
- Wang, C.Y.; Babitt, J.L. Hepcidin regulation in the anemia of inflammation. Curr. Opin. Hematol 2016, 23, 189–197. [Google Scholar] [CrossRef]
- Pagani, A.; Nai, A.; Silvestri, L.; Camaschella, C. Hepcidin and Anemia: A Tight Relationship. Front. Physiol 2019, 10, 1294. [Google Scholar] [CrossRef] [PubMed]
- Babitt, J.L.; Huang, F.W.; Wrighting, D.M.; Xia, Y.; Sidis, Y.; Samad, T.A.; Campagna, J.A.; Chung, R.T.; Schneyer, A.L.; Woolf, C.J.; et al. Bone morphogenetic protein signaling by hemojuvelin regulates hepcidin expression. Nat. Genet. 2006, 38, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Corradini, E.; Babitt, J.L.; Lin, H.Y. The RGM/DRAGON family of BMP co-receptors. Cytokine Growth Factor. Rev. 2009, 20, 389–398. [Google Scholar] [CrossRef]
- Andriopoulos, B., Jr.; Corradini, E.; Xia, Y.; Faasse, S.A.; Chen, S.; Grgurevic, L.; Knutson, M.D.; Pietrangelo, A.; Vukicevic, S.; Lin, H.Y.; et al. BMP6 is a key endogenous regulator of hepcidin expression and iron metabolism. Nat. Genet. 2009, 41, 482–487. [Google Scholar] [CrossRef]
- Meynard, D.; Kautz, L.; Darnaud, V.; Canonne-Hergaux, F.; Coppin, H.; Roth, M.P. Lack of the bone morphogenetic protein BMP6 induces massive iron overload. Nat. Genet. 2009, 41, 478–481. [Google Scholar] [CrossRef]
- Monnier, P.P.; Sierra, A.; Macchi, P.; Deitinghoff, L.; Andersen, J.S.; Mann, M.; Flad, M.; Hornberger, M.R.; Stahl, B.; Bonhoeffer, F.; et al. RGM is a repulsive guidance molecule for retinal axons. Nature 2002, 419, 392–395. [Google Scholar] [CrossRef]
- Hata, K.; Fujitani, M.; Yasuda, Y.; Doya, H.; Saito, T.; Yamagishi, S.; Mueller, B.K.; Yamashita, T. RGMa inhibition promotes axonal growth and recovery after spinal cord injury. J. Cell Biol. 2006, 173, 47–58. [Google Scholar] [CrossRef]
- Samad, T.A.; Srinivasan, A.; Karchewski, L.A.; Jeong, S.J.; Campagna, J.A.; Ji, R.R.; Fabrizio, D.A.; Zhang, Y.; Lin, H.Y.; Bell, E.; et al. DRAGON: A member of the repulsive guidance molecule-related family of neuronal- and muscle-expressed membrane proteins is regulated by DRG11 and has neuronal adhesive properties. J. Neurosci. 2004, 24, 2027–2036. [Google Scholar] [CrossRef]
Figure 1.
Kinetics of Iron Absorption in the Body.
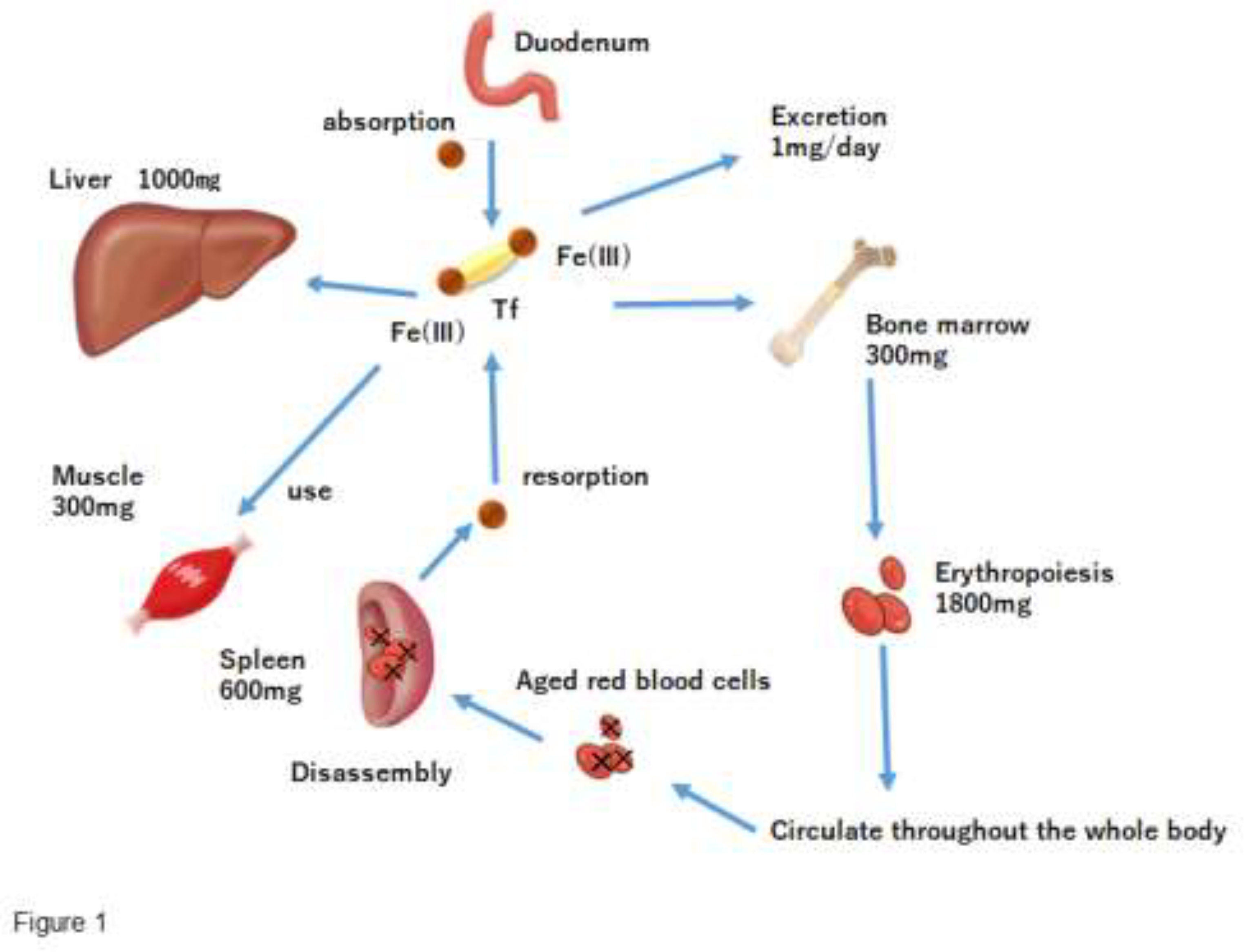
Figure 2.
The comprehensive mechanism of iron metabolism regulation centered on hepcidin.
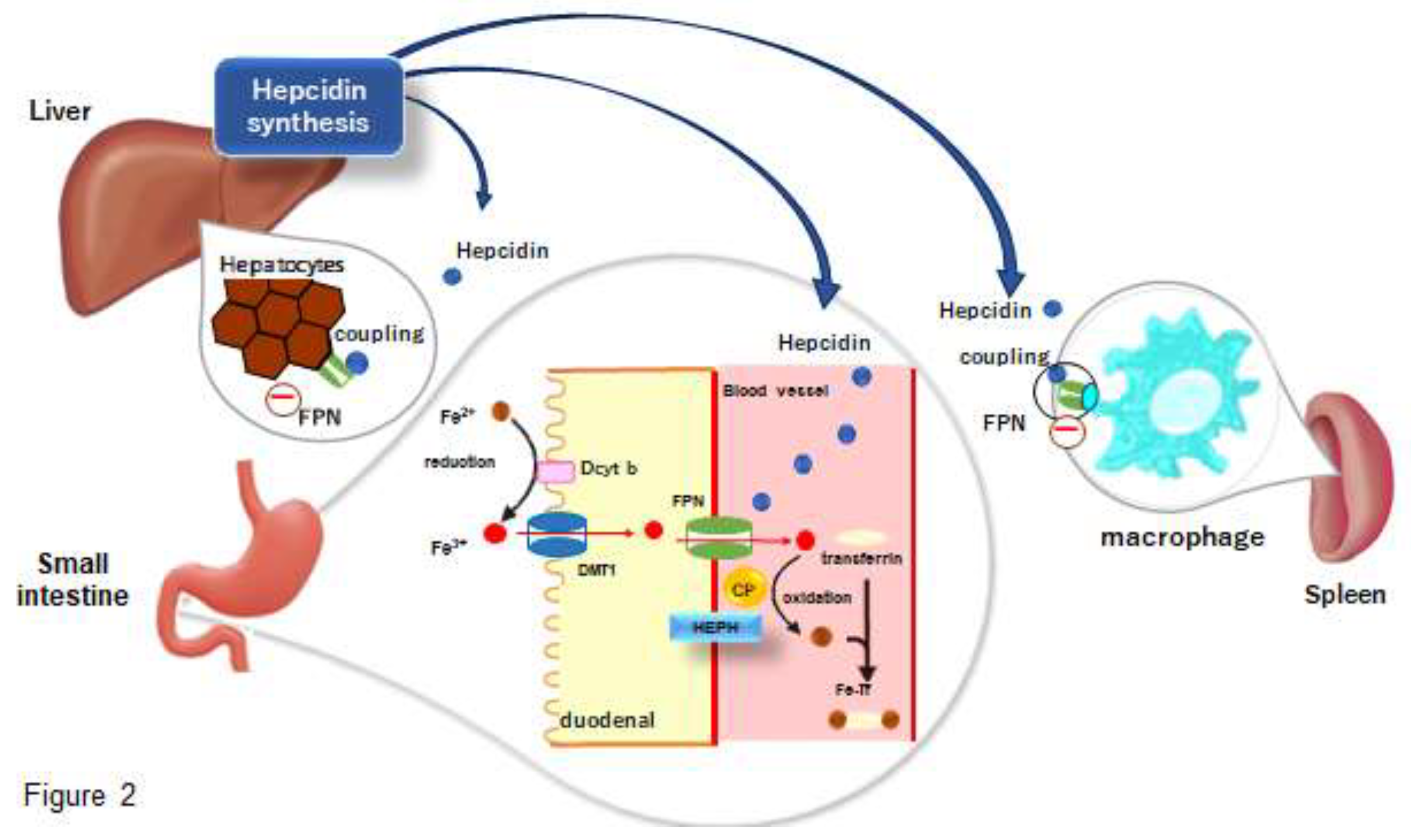
Figure 3.
Effects of swimming or climbing exercise on hemoglobin concentration in rats receiving 4, 18, 29, and 40 mg Fe/kg diet for 8 wk. Values are means and SE for 6 rats. Means with different superscripts are significantly different at p<0.05, determined by ANOVA and Fisher’s PLSD test. SED, sedentary; SWIM, swimming exercise; CLIMB, climbing exercise.
Figure 3.
Effects of swimming or climbing exercise on hemoglobin concentration in rats receiving 4, 18, 29, and 40 mg Fe/kg diet for 8 wk. Values are means and SE for 6 rats. Means with different superscripts are significantly different at p<0.05, determined by ANOVA and Fisher’s PLSD test. SED, sedentary; SWIM, swimming exercise; CLIMB, climbing exercise.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



