Submitted:
21 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
The glucocorticoid receptor alpha (GRα), a pivotal component of the ancient glucocorticoid (GC) signaling system that emerged nearly 450 million years ago, has been instrumental in directing vertebrate evolution and is essential for survival, regulating fertility and fetal development, the function of every organ and circulating cells, metabolism, immune responses, and adaptation to stress. While its anti-inflammatory properties have been recognized since the mid-20th century, GRα’s broader role as the master regulator of homeostatic corrections, particularly in health and critical illness, has only recently been fully appreciated.GRα orchestrates three key phases of homeostatic correction: the Priming Phase, where it rapidly activates early immune responses, mobilizes energy reserves, and prepares the body to mount an immediate defense against various physiological stressors, including infection and injury. During this phase, GRα enhances glucose metabolism, supports mitochondrial function, and facilitates the release of key immune cells to the site of damage, ensuring the body is equipped to handle the initial assault. The Modulatory Phase follows, in which GRα fine-tunes the inflammatory response to prevent excessive tissue damage. By balancing pro- and anti-inflammatory signals, GRα helps contain the immune response, preventing the harmful effects of prolonged inflammation while maintaining immune surveillance. This phase also involves the regulation of oxidative stress, vascular tone, and the preservation of cellular integrity to protect vital organs from injury. Finally, in the Restorative Phase, GRα plays a critical role in resolving inflammation and initiating tissue repair. During this phase, GRα supports cellular regeneration, promotes the clearance of apoptotic cells and debris, and restores anatomical and physiological balance. It aids in rebuilding tissue architecture and returning the neuroendocrine-immune system to its baseline state, ensuring long-term recovery and stability.Throughout these phases, GRα manages complex interactions, including recently discovered co-regulation with pro-inflammatory transcription factors, while also regulating mitochondrial function to ensure bioenergetic and metabolic stability during stress. In critically ill patients, depletion of bioenergetic and micronutrient reserves severely impairs GRα’s capacity to regulate immune responses, heightening the risk of morbidity and mortality.This review, in light of this new understanding, underscores the need to re-evaluate current glucocorticoid (GC) treatment strategies, emphasizing the incorporation of micronutrient support to optimize GRα function across all phases of homeostatic correction. This integrated therapeutic approach has the potential to enhance both immediate immune responses and long-term recovery in critically ill patients, offering a more comprehensive strategy for managing the complex pathobiology of critical illness. Furthermore, this integrative approach could decrease glucocorticoid dose and duration requirements and potential side effects.
Keywords:
acute phase response
; acute respiratory distress syndrome
; allostasis
; chromatin
; chronic
; co-regulation
; critical illness
; dysregulated systemic inflammation
; endothelium
; evolution
; general adaptation syndrome
; glucocorticoid
; glucocorticoid receptor
; homeostasis
; homeostatic corrections
; long-term
; micronutrients
; mitochondria
; nuclear factor kappa B
; reactive oxygen species
; regulation
; resolution
; sepsis
; stress response
; systemic inflammation
; treatment
; vertebrates
For nearly 450 million years, the glucocorticoid (GC) signaling system, mediated by the glucocorticoid receptor (GR), has been integral to vertebrate evolution and survival, enabling precise adaptations to environmental stressors, metabolic regulation, and immune homeostasis. Although this system has ancient origins, its full significance became evident only in the early 20th century, with the discovery of cortisol's therapeutic value in treating inflammatory diseases [1]. This breakthrough marked a turning point in medicine. However, the predominant focus on glucocorticoids as anti-inflammatory agents has often overshadowed their broader biological roles. This review explores the evolutionary history of the GC-GR system and its central role in the development of vertebrates and humans. It examines its role in fertility, fetal development, and the regulation of all organs and circulating cells throughout life, its complex involvement in homeostatic corrections, and its vital contributions to disease resolution, particularly in critical illness.
The evolutionary history of the GC-GR system. The evolutionary development of homeostatic corrections, particularly involving the GC-GR network and its interaction with molecular pathways, is driven by organisms' need to survive and adapt to environmental challenges [2]. Steroid signaling, including that mediated by glucocorticoids, emerged 450 million years ago—about 250 million years before the development of the hypothalamic-pituitary-adrenal (HPA) axis in vertebrates. Interestingly, GR, Nuclear Factor- kappa B (NF-kB), and the mechanisms of hemostasis and evolved concurrently during the Cambrian explosion, highlighting their interconnected roles in regulating physiological responses. This early signaling system allowed vertebrates to regulate metabolism and respond to environmental stressors, which was essential for survival. (Table 1) [3]. The co-evolution of the GR with the immune system further enabled the regulation of inflammation and hemostasis, balancing immune defense with tissue protection. This regulatory balance is exemplified by the interaction between GR and transcription factors (TFs) such as NF-κB and Activator Protein-1 (AP-1) [4]. Over time, the GR system evolved to respond to a wide range of stressors—including infections, injuries, psychological stress, and metabolic demands—positioning GR as a master regulator of homeostasis by integrating signals from multiple pathways.
The co-evolution of the GR with mitochondrial function further underscores its critical role in energy production and metabolism during stress. Mitochondria, which originally formed a symbiotic relationship with early eukaryotic cells, became integral to GR-mediated stress responses, representing a significant advancement in maintaining both cellular and systemic homeostasis [5,6]. Mitochondria are under GC-GRα control due to the presence of GRs within them, and glucocorticoid response elements (GREs) have been identified in the mitochondrial genome [6,7].
Understanding the evolutionary development of these systems provides a critical framework for gaining greater insight into the complex pathobiology of critical illness, highlighting the deep-rooted interplay between stress response, inflammation, hemostasis, immune regulation, bioenergetics, and metabolic adaptation. Despite this long evolutionary history, it wasn’t until the 20th century that the body’s stress response was systematically studied, yielding groundbreaking insights into systemic inflammation and its progression to organ failure [8].
Brief History of the Discovery of Glucocorticoids and Advancements in Knowledge. The discovery of cortisone in the late 1940s, particularly for treating rheumatoid arthritis, revolutionized medicine due to its powerful anti-inflammatory effects [9]. Notably, the dangers of using glucocorticoid treatment without proper tapering were recognized early on, yet this critical aspect continues to be neglected in many randomized trials [10]. Since then, GCs have played a central role in inflammation management. However, their broader biological significance extends well beyond these properties. In the late 1960s and early 1970s, the identification of the GRα receptor significantly advanced understanding of GC mechanisms. By the 1980s and 1990s, it became clear that the GC-GRα complex was crucial for transcriptional regulation, highlighting GCs’ wider impact on cellular functions [11].
GC-GR System: Master Regulator of Homeostatic Corrections. Over the last two decades, the GC-GRα system has emerged as the master regulator of homeostasis, finely tuning physiological adjustments during disease progression and resolution [12,13]. This evolving perspective challenges the traditional view of GCs as merely anti-inflammatory agents, emphasizing their broader roles in metabolism, stress adaptation, and immune regulation—especially in critical illness, where their extensive actions are often underappreciated.
The regulatory functions of GRα have evolved to optimize survival and reproduction. The GRα system has been indispensable for vertebrate evolution, performing essential functions in both male and female physiology, from fertility [14] to establishing the cellular environment necessary for maintaining normal uterine biology and regulating placental development through its uterine-specific actions. GRα is also critical for the structural and functional maturation of all fetal organs [15,16] by influencing the expression of nearly 4,000 genes [17].
The profound importance of the GRα system in reproductive health and developmental processes ensures survival and proper function throughout the lifespan, with its investment in creating life continuing throughout life as GRα maintains physiological homeostasis, supports organ function, and ensures the body’s capacity to respond to stress and sustain health.
The GRα regulatory framework plays a critical role in maintaining vascular and neural integrity, regulating the function of every organ (Table 2) and circulating cells (Table 3), from immune to non-immune cells, ensuring coordinated and precise responses to stress, and is essential for preventing or resolving multi-organ failure in critical illness [13,18,19]. GC-GRα signaling pathway influence spans virtually every physiological system, impacting metabolism by promoting gluconeogenesis, mobilizing fatty acids, and modulating protein catabolism, all essential for maintaining energy balance during stress. Glucocorticoids regulate cardiovascular and vascular function by controlling vascular tone and reducing inflammation, ensuring proper blood flow and vascular integrity. They also play a key role in electrolyte balance by influencing sodium retention and potassium excretion in the kidneys, which is crucial for fluid balance and blood pressure regulation. Additionally, glucocorticoids are central to neuroendocrine signaling, particularly through their feedback within the HPA axis, which modulates the body’s overall stress response. Their effects extend to the nervous system, where they regulate neurotransmitter release, influence cognitive processes such as memory and learning, and modulate mood. In muscle tissue, glucocorticoids balance protein synthesis and degradation, but prolonged exposure can lead to muscle wasting. Furthermore, they are vital for tissue repair, transitioning from an early pro-inflammatory response to activating anti-inflammatory pathways, ensuring proper wound healing and recovery. Glucocorticoids are central agents in preserving homeostatic balance, ensuring that the body’s responses to stress are precisely coordinated, regulated, sustained, and ultimately resolved.
The interaction between GRα and pro-inflammatory TFs such as NF-κB and AP-1 is central to the stress response. For decades, pro- and anti-inflammatory actions were viewed as opposing forces, akin to a “tug of war.” However, emerging evidence reveals a more nuanced relationship, characterized by complementary co-regulation [20] (dualism) between GRα and the NF-κB and AP-1 pathways throughout every phase of the disease process. GRα and these pro-inflammatory TFs can function in a co-regulatory manner, dynamically adjusting gene expression based on the specific cellular context and inflammatory status.
Building on the intricate crosstalk between GRα and pro-inflammatory transcription factors, four seminal studies [21,22,23,24] have revealed a novel dual-phase modulation of chromatin accessibility by GRα and pro-inflammatory transcription factors. In the initial phase of the host response (see below), GRα actively interacts with NF-κB or AP-1 to drive the transcription of pro-inflammatory genes [21,22,23], ensuring a robust immune response to external threats. As homeostatic corrections progress, these interactions enable GRα to pivot, facilitating tissue recovery by promoting the transcription of anti-inflammatory genes [24]. During this phase, GRα modulates chromatin structure, shifting from promoting inflammation to restoring balance by activating anti-inflammatory pathways. This evolutionarily refined strategy demonstrates how GRα, in coordination with transcription factors such as NF-κB and AP-1, ensures that the immune system's response to stress is both swift and tightly regulated, allowing for an immediate inflammatory reaction to threats while simultaneously preparing for anti-inflammatory and tissue-repair processes, thereby ensuring efficient immune regulation and recovery during stress.
Temporal Phases of GRα Function and Micronutrient’ Impact on Its Regulation. The regulation of GRα involves six sequential phases [18] (Table 4) that are critical to its activity in stress response, immune modulation, and homeostatic corrections. These phases are influenced by various factors, including hormone availability, chromatin accessibility, post-translational modifications, oxidative stress, and cellular signaling interactions [12]. Each phase is supported by specific nutrients that facilitate GRα’s function and protect it from damage, ensuring proper cellular responses during stress and recovery.
1. The Initial Activation Phase The availability of cortisol, the primary ligand for GRα, initiates the Initial Activation Phase. Cortisol production is regulated by the adrenal glands, in response to stress and circadian rhythms. Corticosteroid-binding globulin (CBG) and cortisol biosynthetic enzymes facilitate cortisol’s availability, enabling it to bind to GRα and activate the receptor’s cellular functions. Essential nutrients supporting this phase include Vitamin B6, which plays a key role in cortisol synthesis and regulation, and Vitamin C, [6] which enhances adrenal gland function and cortisol production. Sodium is also crucial for adrenal health. Hormonal fluctuations, particularly in cortisol levels, significantly influence GRα activation, with peak cortisol levels occurring in the morning [25,26]. Additionally, hormonal fluctuations, particularly in cortisol levels, significantly influence GRα activation. Peak cortisol levels occur in the morning, with synthesis triggered by stress signals [25]. Vitamins B6 and C help modulate cortisol synthesis, while Vitamin C reduces oxidative stress during cortisol production [26].
2. Once GRα is activated, the Chromatin Remodeling and Gene Accessibility phase follows. During this phase, chromatin remodeling, including histone acetylation, enables GRα to access specific genes within the DNA to regulate gene expression. Key binding partners include chromatin remodeling complexes and histone acetyltransferases (HATs). Nutrients like Vitamin B9, which provides methyl groups for DNA and histone modifications, Vitamin C, which supports chromatin remodeling enzymes, and zinc, necessary for chromatin-remodeling enzymes, play integral roles in facilitating GRα’s regulatory functions during transcription [27].
3. The Immediate Post-translational Modifications phase involves hanges such as phosphorylation, acetylation, and ubiquitination, which regulate GRα's stability, activity, and protein interactions. These modifications fine-tune GRα’s function, with kinases (e.g., MAPK) and phosphatases acting as key binding partners. Nutrients like Vitamins B6, C, and E support enzymes that perform these modifications, while magnesium and zinc serve as cofactors for the modifying enzymes, ensuring GRα’s proper activity [25,28].
4. In the Early Signaling and Interaction Phase, GRα interacts with other receptors, including mineralocorticoid receptors (MR) and estrogen receptors (ER), to modulate cellular responses to stress, metabolism, and inflammation [28]. Inflammatory cytokines such as TNF-α and IL-6 activate pathways that interact with GRα, influencing its role in immune regulation. Binding partners like NF-κB and AP-1 further modulate GRα’s activity. Magnesium is important for receptor cross-talk, while antioxidant nutrients like Vitamins C and E help reduce inflammation by mitigating oxidative stress [25,26]. Selenium and zinc support immune function and the enzymes that regulate inflammation, while selenium and zinc support immune function and the enzymes that regulate inflammation.
5. As signaling continues, the Sustained Signaling Phase focuses on protecting GRα from oxidative damage caused by stress. Oxidative stress can impair GRα’s function and damage cellular components, making antioxidant defense mechanisms critical. Vitamins C and E provide protection against oxidative stress, while selenium, zinc, magnesium, and iron support mitochondrial function and oxidative stress mitigation [26].
6. In the Long-term Regulation and Maintenance Phase, epigenetic modifications, interactions with commensal microbiota, and prolonged cortisol exposure influence GRα’s long-term regulation. Binding partners like DNA methyltransferases, Toll-like receptors (TLRs), and hepatic enzymes (e.g., CYP3A4, 11β-HSD1, 11β-HSD2) are crucial in regulating these long-term effects. Vitamins B9, B12, D, and E support epigenetic stability, while zinc, magnesium, and iron contribute to prolonged GRα regulation, modulating immune responses and metabolic processes during chronic stress.
The General Adaptation Syndrome. While the discovery of glucocorticoids laid the groundwork for understanding their role in stress and inflammation, it was Hans Selye in the 1930s and 1940s who revolutionized the field by systematically studying the body's response to stress. Through his pioneering work, Selye formalized the concept of the General Adaptation Syndrome (GAS), demonstrating that organisms respond to various stressors—whether physical, emotional, or environmental—in a predictable stereotypical manner. Selye proposed that the body progresses through three distinct phases in response to stress: the Alarm Phase, where the body’s immediate defense mechanisms are activated; the Resistance Phase, where the body attempts to adapt and cope with the ongoing stressor; and the Exhaustion Phase, where the body’s resources become depleted. His research revealed that animals subjected to prolonged stress exhibited significant physical changes, most notably enlargement of the adrenal cortex, a sign of the body's adaptive efforts to increase glucocorticoid production—particularly cortisol in humans. This increase in cortisol is essential for mobilizing energy reserves and modulating immune and inflammatory responses during periods of intense physiological demand [8].
Selye later established that failure of the HPA-axis—the critical neuroendocrine system responsible for the stress response—was pivotal in the body's inability to cope with chronic or overwhelming stress. In the Exhaustion phase, the adrenal cortex’s capacity to produce cortisol is compromised, leading to a collapse of homeostatic control, multi-system dysfunction, and immune dysregulation. This failure marks a shift from adaptive to maladaptive responses, characterized by dysregulated systemic inflammation and the progression to organ failure [8,18]. In response to this understanding, therapeutic interventions such as intramuscular ACTH were explored to improve outcomes in patients with sepsis by stimulating cortisol production and mitigating the effects of immune dysregulation.
The General Adaptation Syndrome in Critical Illness. Critical illness represents the most extreme manifestation of the General Adaptation Syndrome, highlighting the body’s intense systemic response to overwhelming stress. The GC-GRα axis plays a pivotal role in orchestrating homeostatic corrections, with growing evidence revealing its central function in regulating these corrections throughout all phases of the syndrome, from modulating innate immunity to restoring organ integrity and function.
In critical illness, homeostatic corrections are essential for survival, demanding substantial bioenergetic and metabolic resources. By the time patients reach the stage of life support - such as need for vasopressors or mechanical ventilation - they are often nearing or have reached complete exhaustion of neuroendocrine compensatory mechanisms, cellular bioenergetic and metabolic reserves, and vital micronutrient stores [6]. This profound depletion critically impairs GRα's ability to effectively execute the Priming Phase (analogous to Selye’s Alarm Phase), undermining its anti-inflammatory capacity, ability to sustain cellular repair, regulate immune responses, and support vital organ functions. The resulting severe allostatic load drastically compromises the body’s recovery potential, significantly heightening the risk of both acute and long-term morbidity and mortality [18].
Understanding the evolutionary development of these homeostatic corrections provides a crucial framework for deciphering the complex pathobiology underlying critical illness (Table 1). The activated GC-GRα signaling system functions as a master regulator across all three major phases of homeostatic corrections, orchestrating precise, timely tissue-specific adjustments while dynamically integrating signals from other hormonal and molecular pathways [10,12].
This adaptive capability ensures that the body's responses are meticulously tailored to the specific context and target cells involved, allowing for finely tuned adjustments that meet the distinct demands of various tissues under both physiological and pathological conditions. Central to recovery, this system serves as the cornerstone of immunity, vascular and neural integrity, energy regulation, stress response, metabolism, electrolyte balance, and tissue repair, orchestrating these essential physiological processes with remarkable precision and adaptability. By continuously modulating these processes in response to evolving physiological demands and pathological stressors, this system prevents systemic failure and supports the restoration of homeostasis [10].
Vitamins and micronutrients are essential for supporting homeostatic corrections by serving as key cofactors in metabolic processes, antioxidant defense, and cellular repair, all of which are critical for maintaining physiological balance during stress and recovery. These nutrients are rapidly depleted in individuals with severe illness due to increased metabolic demands and oxidative stress. Over the course of evolution, organisms have adapted to utilize these nutrients to enhance mitochondrial energy production, regulate gene expression, and modulate immune responses, but in severe disease, replenishment is crucial to support resilience and recovery.
Table 5 provides an overview of the critical roles that thiamine, vitamin D, and vitamin C play in the three phases of homeostatic corrections during critical illness. Each micronutrient supports functions like immune regulation, energy metabolism, vascular integrity, and inflammation repression, contributing to the restoration and maintenance of cellular and tissue homeostasis. Thiamine is vital for ATP production, vitamin D modulates immune and vascular functions, and vitamin C acts as a potent antioxidant, supporting immune defense. For more details on how each micronutrient functions in these phases, refer to reference [6].
In addition to vitamins, essential micronutrients such as magnesium, zinc, selenium, copper, iron, and calcium are critical for the three phases of homeostatic regulation during critical illness. Magnesium supports ATP production and reduces oxidative stress, particularly in the early and inflammatory phases [29]. Zinc modulates immune function, aids in cytokine production, and supports tissue repair [30]. Selenium acts as an antioxidant, reducing oxidative stress and promoting inflammation resolution [31]. Copper and iron are essential for mitochondrial function, vascular integrity, and immune responses [32,33]. Calcium assists in intracellular signaling and endothelial repair [34]. Together, these micronutrients complement GRα in maintaining homeostasis and preventing multi-organ failure. A comprehensive review of their detailed interactions is outside the scope of this summary.
Meduri et al. first described the three major phases of homeostatic corrections [35]; this work presents a significant update to that model (Figure 1). In an adaptive response, these phases are successfully completed, leading to disease resolution and restoration of homeostasis. Conversely, in a maladaptive response, failure to complete these phases impedes recovery, perpetuates dysfunction, and can ultimately result in chronic disease or mortality [18].
1. Priming Phase: Activating innate immunity, bioenergetic supply, and cardiovascular response. The GC-GRα signaling system plays a central role in amplifying the body's initial immune response by ensuring energy availability and maintaining vascular integrity. This prepares the body to mount an immediate defense against various stress signals, including pathogenic and inflammatory stimuli. The GC-GRα complex is critical for the development and release of key immune cells—T cells, B cells, NK cells, dendritic cells, monocytes/macrophages, and neutrophils—regulating both innate and adaptive immune responses. Additionally, the GC-GRα complex enhances glucose metabolism, boosts mitochondrial biogenesis [36,37], and facilitates energy production via glycolysis and fatty acid breakdown [38], ensuring the energy reserves necessary for heightened immune activity. Throughout all phases of homeostatic corrections, the GC-GRα complex supports mitochondrial function, not only ensuring bioenergetic stability but also modulating oxidative stress responses and maintaining mitochondrial integrity. This sustained support is crucial for balancing the metabolic demands of immune activation, tissue repair, and restoration of systemic homeostasis, as mitochondria play a key role in energy production, cell survival, and adaptive signaling [6].
Priming innate immunity through key mechanisms. The priming of innate immunity occurs through several mechanisms. First, the GC-GRα complex triggers and modulates the systemic and hepatic acute-phase protein (APP) response via IL-6 and nitric oxide, with GRα co-regulating alongside IL-6 [23,39]. This response results in the production of proteins such as C-reactive protein (CRP) and serum amyloid A (SAA), both essential for inflammation and innate immunity. Second, the complex induces the expression of critical immune receptors, including Toll-like receptors 2 and 4 (TLR2 and TLR4), p38 MAPK, the NLRP3 inflammasome [40,41], and the purinergic receptor P2Y2R [42]. These receptors are vital for recognizing pathogen-associated molecular patterns (PAMPs) and danger-associated molecular patterns (DAMPs), enabling the innate immune system to respond swiftly to infections and cellular stressors. Third, the GC-GRα complex cooperates with pro-inflammatory transcription factors leading to the synergistic induction of pro-inflammatory genes (see below).
Amplifying early immune activation through cooperation with pro-inflammatory transcription factors. Although the GC-GRα complex typically restrains inflammation, during the early stages of homeostatic corrections, it cooperates with NF-κB, AP-1, and TNF-α to modify chromatin and regulate previously inaccessible genes, leading to the induction of pro-inflammatory genes [21,22,23] that enhance immune processes such as leukocyte trafficking, chemokine production, and complement component activation [23,43]. Additionally, GC-GRα mediates the induction of NLRP3, sensitizing cells to extracellular ATP and promoting the release of pro-inflammatory molecules like TNF-α, IL-1β, and IL-6 [40,41]. The GC-GRα signaling system collaborates with other hormonal systems, including the HPA-axis and thyroid hormones, to orchestrate a comprehensive adaptive response.
Regulation of phagocytic activity and bacterial killing. The GC-GRα complex plays a pivotal role in preventing excessive activation of phagocytic cells while simultaneously enhancing intracellular bacterial killing [44]. During the early phase of innate immunity, the upregulation of GILZ (Glucocorticoid-Induced Leucine Zipper) boosts interferon-gamma (IFN-γ) expression, which promotes macrophage activation, bacterial clearance, and the induction of apoptosis in infected cells. This dual action balances the immune response by inhibiting pro-inflammatory transcription factors like NF-κB and AP-1, while promoting anti-inflammatory mediators via GILZ [45,46,47]. This coordination ensures that pathogens are cleared while uncontrolled inflammation is prevented.
Glucocorticoid regulation in immune control. Glucocorticoid regulation is crucial in immune control. In the absence of adrenal glucocorticoid (GC) production, pathogen clearance may occur more rapidly, but this leads to higher mortality due to unchecked T-cell responses [48]. Pro-inflammatory cytokines further increase GC bioavailability by enhancing cortisol production and activating 11β-HSD1 [7]. This cytokine-GC synergy strengthens early immune responses while preventing overactivation of phagocytes. In the Priming Phase, the GC-GRα complex also suppresses acquired immunity [4,43,48], and limits non-essential functions such as digestion, reproduction, and growth, conserving energy for the immediate needs of the innate immune system.
Cardiovascular response and endothelial protection. The GC-GRα complex also plays a vital role in enhancing cardiovascular responses and protecting endothelial cells. It increases the sensitivity of adrenergic receptors and upregulates beta-adrenergic receptor expression in cardiac and vascular smooth muscle cells [49,50]. This leads to enhanced catecholamine synthesis and release from the adrenal medulla, ensuring adequate hormone supply during stress. The complex also boosts nitric oxide (NO) production by upregulating endothelial nitric oxide synthase (eNOS) [51,52,53], which maintains vascular tone, reduces resistance, and prevents endothelial dysfunction.
Antioxidant defense and vascular health. In addition to its cardiovascular effects, GR activation upregulates antioxidant enzymes and protective factors within endothelial cells, mitigating oxidative stress [54,55]. These antioxidant actions provide sustained protection to vascular tissues throughout the resolution phase of immune responses, ensuring long-term vascular health and functionality.
2. Modulatory Phase: Repressing inflammation, mitigating oxidative stress, and restoring vascular integrity. After the initial immune response is reinforced and stabilized, the body enters the Modulatory Phase, where the primary goal is to prevent further damage by downregulating the inflammatory response and reducing oxidative stress. This shift, driven by the GC-GRα signaling system, is critical because unchecked inflammation and oxidative stress can compromise the body’s recovery by damaging tissues, impairing GRα function, and leading to cellular dysfunction. In this phase, restoring vascular integrity is equally important, as maintaining proper blood flow and organ perfusion is essential for tissue restoration and preventing further deterioration.
Balancing inflammation and repair. The GC-GRα system plays a crucial role in balancing this process. By fine-tuning the inflammatory response, it not only reduces the immediate risk of tissue damage but also lowers the risk of chronic inflammation, which could hinder recovery. At the same time, the system initiates the restoration of vascular integrity, ensuring that nutrient delivery and blood flow to affected tissues are maintained or improved. This phase requires a delicate balance between dampening harmful inflammatory and oxidative processes and promoting early tissue repair and recovery, laying the groundwork for the subsequent healing phase.
Anti-inflammatory mechanisms of the GC-GRα complex. During this phase, the GC-GRα complex exerts its anti-inflammatory effects by inhibiting key inflammatory pathways, including NF-κB, AP-1, and MAPK [56]. Simultaneously, it increases the transcription of anti-inflammatory genes and the NF-κB inhibitory protein IκBα [19,57], which further strengthens the anti-inflammatory response. The complex interaction between glucocorticoids and NF-κB depends on the specific stimulus, leading to a flexible and adaptive inflammatory control mechanism [24].
Collaboration with pro-inflammatory transcription factors. One notable feature of this phase is the cooperative interaction between GRα and pro-inflammatory transcription factors (TFs), which paradoxically enhances the overall anti-inflammatory response. This collaboration between GRα and TFs contributes to the resolution of inflammation and the stabilization of vascular function [24]. Chromatin accessibility plays a key role here, allowing GRα to work in tandem with pro-inflammatory TFs, amplifying the anti-inflammatory effects of glucocorticoids. For example, AP-1 primes chromatin, enabling GRα to bind more effectively and suppress inflammation, while enhanced chromatin accessibility enables GRα to fine-tune NF-κB activity [22].
Preventing nosocomial infections in critical illness. In conditions like acute respiratory distress syndrome (ARDS) and sepsis, dysregulated inflammation impairs neutrophil function, increasing the risk of secondary infections [58,59,60,61]. By effectively controlling inflammation with a robust anti-inflammatory response, the risk of nosocomial infections is significantly reduced [62,63,64,65]. In ARDS survivors, lower levels of inflammatory cytokines suppress bacterial growth, while in non-survivors, higher cytokine levels promote pathogen proliferation [66]. Prolonged glucocorticoid treatment reduces systemic and pulmonary inflammation, lowering the risk of infections by shortening the duration of mechanical ventilation, creating a less favorable environment for pathogen growth [44,66,67,68,69], and enhancing neutrophil function [59,60].
Restoring vascular integrity through endothelial regulation. In addition to controlling inflammation, the GC-GRα complex plays a crucial role in restoring vascular integrity by upregulating mediators that support endothelial cell homeostasis. It upregulates Sphingosine Kinase 1 (SphK1) [70], which produces sphingosine-1-phosphate (S1P), a molecule essential for maintaining endothelial barrier function. Additionally, the complex enhances Angiopoietin-1 (Angpt-1) expression, stabilizing endothelial cell junctions and reducing vascular permeability. These actions are essential for maintaining vascular stability during the recovery process.
Supporting vascular stability and endothelial health. The GC-GRα complex also induces MAPK Phosphatase-1 (MKP-1) [71], which mitigates inflammation by deactivating MAPKs, and upregulates Serum Glucocorticoid Kinase-1 (SGK-1) [72], which promotes cell survival and supports endothelial stability. GILZ, another key mediator upregulated by the complex [73], inhibits pro-inflammatory transcription factors such as NF-κB, further protecting vascular integrity. The GC-GRα complex also boosts nitric oxide (NO) production via Endothelial Nitric Oxide Synthase (eNOS) [51,52,53], which maintains vascular tone, reduces vascular resistance, and prevents endothelial dysfunction. Together, these actions preserve the vascular system as the body transitions to the resolution phase of homeostatic corrections.
Experimental evidence in sepsis. In experimental sepsis, low-dose glucocorticoid treatments such as hydrocortisone or dexamethasone have been shown to preserve the endothelial glycocalyx, sustain the vascular barrier, reduce interstitial edema [74,75], and improve mesenteric blood flow, all contributing to the resolution of organ injury [76]. These findings underscore the critical role of the GC-GRα signaling system in facilitating the transition from inflammation to tissue repair and recovery, highlighting the need for ongoing modulation to promote long-term recovery in critically ill patients.
3. Restorative Phase: Resolving inflammation, facilitating tissue repair, and restoring normal structure. The sequential phases of inflammation reflect an adaptive response when systemic inflammation is well-regulated. In contrast, maladaptive responses occur when the GC-GRα complex fails to properly downregulate inflammation and support tissue repair, resulting in incomplete recovery [18].
Timely downregulation of inflammation. Resolving disease requires the timely downregulation of systemic inflammation, beginning in the Modulatory Phase. In the early stages of critical illness, dysregulated inflammation can lead to prolonged organ support, depletion of neuroendocrine compensation, cellular bioenergetic reserves, and essential micronutrients—all critical for an adaptive response. NF-κB drives persistent inflammation, oxidative stress, and micronutrient depletion. If GRα downregulation is insufficient, patients either improve slowly (partial failure) or not at all (complete failure), leading to ongoing tissue damage and increasing the risk of in-hospital death or chronic morbidity among survivors [77].
From pro-inflammatory to pro-resolving mediators. In the Restorative Phase, the resolution of inflammation is an active process involving the transition from pro-inflammatory to pro-resolving mediators [12,78]. As inflammation subsides, the activated GC-GRα complex engages various pro-resolution mechanisms, altering the phenotypes of immune cells such as granulocytes and macrophages. Through genomic mechanisms, GRα increases the expression of key pro-resolving proteins, including AnxA1, its receptor ALXR, and GILZ. These molecules are crucial for controlling the resolution phase of inflammation, particularly in macrophages and neutrophils [79].
Clearance of cellular debris and preparation for tissue repair. Once inflammation begins, damaged tissues release debris and intracellular molecules due to cell damage or death, which signals immune cells to initiate the healing process [80]. Effective tissue repair requires this debris to be cleared [80]. During this phase, the GC-GRα system promotes the removal of damaged cells through phagocytosis and breaks down fibrin and clots, creating a clean foundation for tissue repair.
Supporting cellular regeneration and structural restoration. The GC-GRα system is essential for driving cellular regeneration and structural restoration. It stimulates the production of growth factors that promote cell proliferation and tissue regeneration, while also synthesizing extracellular matrix components necessary for rebuilding normal tissue architecture [18]. To support effective repair, the local tissue environment must return to a state of homeostasis, with normalized pH, oxygen levels, and other biochemical conditions.
Restoration of normal anatomical structure. A key objective of this phase is the restoration of normal anatomical structure and function. The GC-GRα system guides tissue remodeling, ensuring that affected organs and systems regain their proper function. This involves both the physical reconstruction of tissues and the normalization of cellular functions, ultimately leading to the full restoration of health and homeostasis.
Granulocyte and macrophage transitions in inflammation resolution. In neutrophils, GILZ expression increases in a significant number of GC-treated ARDS patients [81], promoting their apoptosis and reducing inflammatory potential. Macrophages recognize and remove apoptotic neutrophils through efferocytosis, transitioning from a pro-inflammatory (M1) to an anti-inflammatory (M2) phenotype [82,83]. M2 macrophages release anti-inflammatory mediators and facilitate tissue regeneration through enhanced chemokinesis and lymphatic clearance. For an expanded review see reference [18].
Role of AnxA1 and macrophages in tissue repair. The GC-mediated AnxA1 peptide (Ac2–26), acting via the ALXR receptor, plays a key role in removing apoptotic cells. During later stages, macrophages undergo a phenotypic shift to a resolution-promoting phenotype (Mres), which helps limit tissue damage and fibrosis. These macrophages produce antifibrotic and antioxidant molecules, further aiding in tissue repair. GCs also regulate genes in monocytes, promoting anti-inflammatory pathways.
GC-GRα and TNF-α in tissue remodeling. The GC-GRα complex, in conjunction with TNF-α, co-regulates the expression of SERPINE1 (plasminogen activator inhibitor-1), a key factor in tissue remodeling. SERPINE1 influences extracellular matrix dynamics, angiogenesis, and fibrosis. TNF-α upregulates SERPINE1 during inflammation, promoting ECM accumulation. In the resolution phase, glucocorticoids modulate this effect, balancing pro-fibrotic and pro-resolving pathways, enhancing angiogenesis, and promoting fibrosis resolution [23,40,41].
Preserving muscle mass during prolonged illness. In cases of prolonged illness, where patients often experience immobility and systemic catabolic stress, glucocorticoids help preserve muscle mass and function. By modulating protein metabolism and reducing muscle catabolism, glucocorticoids prevent severe muscle wasting, which is crucial for recovery and rehabilitation [84].
Conclusion. Over 450 million years, the GC-GRα regulatory network has evolved as a cornerstone of biological survival, meticulously maintaining and restoring homeostasis under a variety of stresses and challenges. This finely tuned system is not a passive element of health management but a dynamic mechanism governing all phases of homeostatic correction—priming, modulation, and restoration—ultimately leading to disease resolution. Despite its pivotal role, research efforts have yet to fully harness the immense potential of this system. GRα’s ability to maintain homeostasis is intricately tied to essential micronutrients, which are rapidly depleted during critical illness. Future research must prioritize understanding how these micronutrients, alongside GCs, can further enhance GR function, providing critical support in both acute and chronic disease management.
This evolving understanding of GRα function demands a comprehensive re-evaluation of current GC treatment strategies. Until now, randomized controlled trials (RCTs) investigating GC treatment in critically ill patients have relied on outdated knowledge from forty years ago, neglecting significant advancements in GR research. Future RCTs must focus on optimizing GC administration modalities, while ensuring essential micronutrient support to allow GRα to function optimally across all adaptive phases.
This shift in focus is not merely an advancement in research but a necessary evolution in clinical practice. By embracing an integrative therapeutic approach—combining glucocorticoids (GCs) with vital micronutrients that support glucocorticoid receptor (GR) function and homeostatic corrections—future research holds the potential to significantly improve both immediate therapeutic outcomes and long-term recovery in critically ill patients. This comprehensive strategy not only addresses the complex pathobiology of critical illness but also has the potential to reduce the required glucocorticoid doses and treatment durations, thereby minimizing associated side effects. Such a transformation could improve the standard of care for critically ill patients, help alleviate the burden of chronic disease and provide potential cost savings to healthcare systems over time.
References
- Wu C, Hou D, Du C, et al. Corticosteroid therapy for coronavirus disease 2019-related acute respiratory distress syndrome: a cohort study with propensity score analysis. Crit Care. Nov 10 2020;24(1):643. [CrossRef]
- McEwen BS, Wingfield JC. The concept of allostasis in biology and biomedicine. Horm Behav. Jan 2003;43(1):2-15.
- Thornton J. Evolution of vertebrate steroid receptors from an ancestral estrogen receptor by ligand exploitation and serial genome expansions. Proceedings of the National Academy of Sciences. 2001;98 (10):5671-5676.
- Cain DW, Cidlowski JA. Immune regulation by glucocorticoids. Nature reviews Immunology. Apr 2017;17(4):233-247. [CrossRef]
- Psarra AM, Sekeris CE. Glucocorticoid receptors and other nuclear transcription factors in mitochondria and possible functions. Biochim Biophys Acta. May 2009;1787(5):431-6. [CrossRef]
- Meduri GU, Psarra A-M, Amrein K. General Adaptation in Critical Illness 2: The Glucocorticoid Signaling System as a Master Rheostat of Homeostatic Corrections in Concerted Action with Mitochondrial and Essential Micronutrient Support.
- In: Fink G, ed. Handbook of Stress: Stress, Immunology and Inflammation San Diego, Elsevier.; 2024:263-287:chap 23. vol. 5.
- Dejager L, Vandevyver S, Petta I, Libert C. Dominance of the strongest: inflammatory cytokines versus glucocorticoids. Cytokine Growth Factor Rev. Feb 2014;25(1):21-33. [CrossRef]
- Selye H. The general adaptation syndrome and the diseases of adaptation. J Clin Endocrinol. 1946;6:117-230.
- Hench PS, Kendall EC, Slocumb CH, Polley HF. Arch Internal Med. 1950;85:545-666.
- Meduri GU, Confalonieri M, Chaudhuri D, Rochwerg B, Meibohm B. Prolonged glucocorticoid treatment in ARDS: pathobiological rationale and pharmacological principles. . In: Fink G, ed. Handbook of Stress: Stress, Immunology and Inflammation Academic Press; 2024:289-323:chap 24. vol. Encyclopedia of Stress.
- Munck A, Guyre PM, Holbrook NJ. Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocr Rev. 1984;5(1):25-44.
- Busillo JM, Cidlowski JA. The five Rs of glucocorticoid action during inflammation: ready, reinforce, repress, resolve, and restore. Trends in endocrinology and metabolism: TEM. Mar 2013;24(3):109-19. [CrossRef]
- Meduri GU, Muthiah MP, Carratu P, Eltorky M, Chrousos GP. Nuclear factor-kappaB- and glucocorticoid receptor alpha- mediated mechanisms in the regulation of systemic and pulmonary inflammation during sepsis and acute respiratory distress syndrome. Evidence for inflammation-induced target tissue resistance to glucocorticoids. Neuroimmunomodulation. 2005;12(6):321-38. [CrossRef]
- Whirledge SD, Oakley RH, Myers PH, Lydon JP, DeMayo F, Cidlowski JA. Uterine glucocorticoid receptors are critical for fertility in mice through control of embryo implantation and decidualization. Proceedings of the National Academy of Sciences. 2015;112(49):15166-15171.
- Rog-Zielinska EA, Thomson A, Kenyon CJ, et al. Glucocorticoid receptor is required for foetal heart maturation. Hum Mol Genet. 2013;22(16):3269-3282.
- Bird AD, McDougall AR, Seow B, Hooper SB, Cole TJ. Minireview: glucocorticoid regulation of lung development: lessons learned from conditional GR knockout mice. Mol Endocrinol. 2015;29(2):158-171.
- Oakley RH, Ramamoorthy S, Foley JF, Busada JT, Lu NZ, Cidlowski JA. Glucocorticoid receptor isoform–specific regulation of development, circadian rhythm, and inflammation in mice. The FASEB Journal. 2018;32(10):5258-5271.
- Meduri GU, Chrousos GA. General Adaptation in Critical Illness: The Glucocorticoid Signaling System as Master Rheostat of Homeostatic Corrections in Concerted Action with Nuclear Factor-kB.
- In: Fink G, ed. Handboo of Stress, Immunology and Inflammation Academic Press; 2024:231-261:chap 22. Handbook of Stress; vol. .
- Meduri GU, Tolley EA, Chrousos GP, Stentz F. Prolonged methylprednisolone treatment suppresses systemic inflammation in patients with unresolving acute respiratory distress syndrome. Evidence for inadequate endogenous glucocorticoid secretion and inflammation-induced immune cell resistance to glucocorticoids. Am J Respir Crit Care Med. Apr 1 2002;165(7):983-991. [CrossRef]
- Ratman D, Berghe WV, Dejager L, et al. How glucocorticoid receptors modulate the activity of other transcription factors: a scope beyond tethering. Mol Cell Endocrinol. 2013;380(1-2):41-54.
- Rao NA, McCalman MT, Moulos P, et al. Coactivation of GR and NFKB alters the repertoire of their binding sites and target genes. Genome Res. 2011;21(9):1404-1416.
- Biddie SC, John S, Sabo PJ, et al. Transcription factor AP1 potentiates chromatin accessibility and glucocorticoid receptor binding. Mol Cell. 2011;43(1):145-155.
- Lannan EA, Galliher-Beckley AJ, Scoltock AB, Cidlowski JA. Proinflammatory actions of glucocorticoids: glucocorticoids and TNFα coregulate gene expression in vitro and in vivo. Endocrinology. 2012;153(8):3701-3712.
- Kadiyala V, Sasse SK, Altonsy MO, et al. Cistrome-based cooperation between airway epithelial glucocorticoid receptor and NF-κB orchestrates anti-inflammatory effects. J Biol Chem. 2016;291(24):12673-12687.
- Scheschowitsch K, Leite JA, Assreuy J. New insights in glucocorticoid receptor signaling—more than just a ligand-binding receptor. Front Endocrinol (Lausanne). 2017;8:16.
- Carr AC, Maggini S. Vitamin C and immune function. Nutrients. 2017;9(11):1211.
- Hayashi R, Wada H, Ito K, Adcock IM. Effects of glucocorticoids on gene transcription. Eur J Pharmacol. 2004;500(1-3):51-62.
- Reichardt HM, Tuckermann JP, Göttlicher M, et al. Repression of inflammatory responses in the absence of DNA binding by the glucocorticoid receptor. The EMBO journal. 2001;
- Barbagallo M, Dominguez L. Magnesium and aging. Curr Pharm Des. 2010;16(7):832-839.
- Ibs K-H, Rink L. Zinc-altered immune function. The Journal of nutrition. 2003;133(5):1452S-1456S.
- MP R. Selenium and human health. Lancet. 2012;379:1256-1268.
- Lutsenko S. Human copper homeostasis: a network of interconnected pathways. Curr Opin Chem Biol. 2010;14(2):211-217.
- Ganz T. Systemic iron homeostasis. Physiol Rev. 2013;93(4):1721-1741.
- Berridge MJ. The inositol trisphosphate/calcium signaling pathway in health and disease. Physiol Rev. 2016;96(4):1261-1296.
- Meduri GU, Chrousos GP. General Adaptation in Critical Illness: Glucocorticoid Receptor-alpha Master Regulator of Homeostatic Corrections. Review. Front Endocrinol (Lausanne). 2020-April-22 2020;11(161):161. [CrossRef]
- Psarra AM, Sekeris CE. Glucocorticoids induce mitochondrial gene transcription in HepG2 cells: role of the mitochondrial glucocorticoid receptor. Biochim Biophys Acta. Oct 2011;1813(10):1814-21. [CrossRef]
- Karra AG, Sioutopoulou A, Gorgogietas V, Samiotaki M, Panayotou G, Psarra A-MG. Proteomic analysis of the mitochondrial glucocorticoid receptor interacting proteins reveals pyruvate dehydrogenase and mitochondrial 60 kDa heat shock protein as potent binding partners. J Proteomics. 2022;257:104509.
- Picard M, McManus MJ, Gray JD, et al. Mitochondrial functions modulate neuroendocrine, metabolic, inflammatory, and transcriptional responses to acute psychological stress. Proc Natl Acad Sci U S A. Dec 1 2015;112(48):E6614-23. [CrossRef]
- Gruys E, Toussaint MJ, Niewold TA, Koopmans SJ. Acute phase reaction and acute phase proteins. J Zhejiang Univ Sci B. Nov 2005;6(11):1045-56. [CrossRef]
- Busillo JM, Azzam KM, Cidlowski JA. Glucocorticoids Sensitize the Innate Immune System through Regulation of the NLRP3 Inflammasome. J Biol Chem. Nov 4 2011;286(44):38703-38713. [CrossRef]
- Amrani Y, Panettieri RA, Ramos-Ramirez P, Schaafsma D, Kaczmarek K, Tliba O. Important lessons learned from studies on the pharmacology of glucocorticoids in human airway smooth muscle cells: too much of a good thing may be a problem. Pharmacol Ther. 2020;213:107589.
- Ding Y, Gao ZG, Jacobson KA, Suffredini AF. Dexamethasone enhances ATP-induced inflammatory responses in endothelial cells. J Pharmacol Exp Ther. Dec 2010;335(3):693-702. [CrossRef]
- Galon J, Franchimont D, Hiroi N, et al. Gene profiling reveals unknown enhancing and suppressive actions of glucocorticoids on immune cells. Faseb J. Jan 2002;16(1):61-71. [CrossRef]
- Meduri GU, Kanangat S, Bronze M, et al. Effects of methylprednisolone on intracellular bacterial growth. Clin Diagn Lab Immunol. Nov 2001;8(6):1156-63. [CrossRef]
- Ayroldi E, Riccardi C. Glucocorticoid-induced leucine zipper (GILZ): a new important mediator of glucocorticoid action. Faseb J. 2009;23. [CrossRef]
- Ellouze M, Vigouroux L, Tcherakian C, et al. Overexpression of GILZ in macrophages limits systemic inflammation while increasing bacterial clearance in sepsis in mice. European journal of immunology. 2020;50(4):589-602.
- Ballegeer M, Vandewalle J, Eggermont M, et al. Overexpression of Gilz Protects Mice Against Lethal Septic Peritonitis. Shock. Aug 2019;52(2):208-214. [CrossRef]
- Taves MD, Ashwell JD. Glucocorticoids in T cell development, differentiation and function. Nature Reviews Immunology. 2021;21(4):233-243.
- Oakley RH, Cidlowski JA. The biology of the glucocorticoid receptor: new signaling mechanisms in health and disease. J Allergy Clin Immunol. Nov 2013;132(5):1033-44. [CrossRef]
- Sapolsky RM, Romero LM, Munck AU. How Do Glucocorticoids Influence Stress Responses? Integrating Permissive, Suppressive, Stimulatory, and Preparative Actions. Endocrine Reviews. 2000;21(1):55-89. doi:. [CrossRef]
- Bouazza Y, Sennoun N, Strub C, et al. Comparative effects of recombinant human activated protein C and dexamethasone in experimental septic shock. Comparative Study.
- Research Support, Non-U.S. Gov't. Intensive care medicine. Nov 2011;37(11):1857-64. [CrossRef]
- Limbourg FP, Huang Z, Plumier JC, et al. Rapid nontranscriptional activation of endothelial nitric oxide synthase mediates increased cerebral blood flow and stroke protection by corticosteroids. J Clin Invest. Dec 2002;110(11):1729-38. [CrossRef]
- Hafezi-Moghadam A, Simoncini T, Yang Z, et al. Acute cardiovascular protective effects of corticosteroids are mediated by non-transcriptional activation of endothelial nitric oxide synthase. Nat Med. May 2002;8(5):473-9. [CrossRef]
- Ehrchen J, Steinmuller L, Barczyk K, et al. Glucocorticoids induce differentiation of a specifically activated, anti-inflammatory subtype of human monocytes. Blood. Feb 1 2007;109(3):1265-74.
- Tanaka S, Couret D, Tran-Dinh A, et al. High-density lipoproteins during sepsis: from bench to bedside. Crit Care. Apr 7 2020;24(1):134. [CrossRef]
- Kim EK, Choi EJ. Pathological roles of MAPK signaling pathways in human diseases. Biochim Biophys Acta. Apr 2010;1802(4):396-405. [CrossRef]
- Vandevyver S, Dejager L, Libert C. Comprehensive overview of the structure and regulation of the glucocorticoid receptor. Endocr Rev. Aug 2014;35(4):671-93. [CrossRef]
- Mascellino MT, Delogu G, Pelaia MR, Ponzo R, Parrinello R, Giardina A. Reduced bactericidal activity against Staphylococcus aureus and Pseudomonas aeruginosa of blood neutrophils from patients with early adult respiratory distress syndrome. Journal of medical microbiology. Jan 2001;50(1):49-54.
- Kaufmann I, Briegel J, Schliephake F, et al. Stress doses of hydrocortisone in septic shock: beneficial effects on opsonization-dependent neutrophil functions. Intensive Care Med. Feb 2008;34(2):344-9.
- Keh D BT, Weber-Cartens S, Schulz C, Ahlers O, Bercker S, Volk HD, Doecke WD, Falke KJ, Gerlach H. Immunologic and hemodynamic effects of "low-dose" hydrocortisone in septic shock: a double-blind, randomized, placebo-controlled, crossover study. Am J Respir Crit Care Med. 2003;167(4):512-20.
- Conway Morris A, Kefala K, Wilkinson TS, et al. C5a mediates peripheral blood neutrophil dysfunction in critically ill patients. Multicenter Study.
- Research Support, Non-U.S. Gov't. Am J Respir Crit Care Med. Jul 1 2009;180(1):19-28. [CrossRef]
- Headley AS, Tolley E, Meduri GU. Infections and the inflammatory response in acute respiratory distress syndrome. Chest. May 1997;111(5):1306-21.
- Steinberg KP, Hudson LD, Goodman RB, et al. Efficacy and safety of corticosteroids for persistent acute respiratory distress syndrome. N Engl J Med. Apr 20 2006;354(16):1671-84. [CrossRef]
- Meduri GU, Golden E , Freire AX, et al. Methylprednisolone infusion in early severe ARDS: results of a randomized controlled trial. Original Research. Chest. April 2007 2007;131(4):954-963. [CrossRef]
- Roquilly A, Mahe PJ, Seguin P, et al. Hydrocortisone therapy for patients with multiple trauma: the randomized controlled HYPOLYTE study. Multicenter Study.
- Randomized Controlled Trial.
- Research Support, Non-U.S. Gov't. JAMA : the journal of the American Medical Association. Mar 23 2011;305(12):1201-9. [CrossRef]
- Meduri GU. Clinical review: a paradigm shift: the bidirectional effect of inflammation on bacterial growth. Clinical implications for patients with acute respiratory distress syndrome. Crit Care. Feb 2002;6(1):24-9.
- Kanangat S, Meduri GU, Tolley EA, et al. Effects of cytokines and endotoxin on the intracellular growth of bacteria. Infect Immun. Jun 1999;67(6):2834-40.
- Kanangat S, Bronze MS, Meduri GU, et al. Enhanced extracellular growth of Staphylococcus aureus in the presence of selected linear peptide fragments of human interleukin (IL)-1beta and IL-1 receptor antagonist. J Infect Dis. Jan 1 2001;183(1):65-69. [CrossRef]
- Sibila O, Luna C, Agustí C, et al. Effects of corticosteroids an animal model of ventilator-associated pneumonia. . abstract. Proceeding of the American Thoracic Society. 2006;3:A 21.
- Vettorazzi S, Bode C, Dejager L, et al. Glucocorticoids limit acute lung inflammation in concert with inflammatory stimuli by induction of SphK1. Nat Commun. 2015;6:7796. [CrossRef]
- Fürst R, Schroeder T, Eilken HM, et al. MAPK phosphatase-1 represents a novel anti-inflammatory target of glucocorticoids in the human endothelium. The FASEB journal. 2007;21(1):74-80.
- Kim H, Lee JM, Park JS, et al. Dexamethasone coordinately regulates angiopoietin-1 and VEGF: a mechanism of glucocorticoid-induced stabilization of blood-brain barrier. Biochem Biophys Res Commun. Jul 18 2008;372(1):243-8. [CrossRef]
- Hahn RT, Hoppstadter J, Hirschfelder K, et al. Downregulation of the glucocorticoid-induced leucine zipper (GILZ) promotes vascular inflammation. Atherosclerosis. Jun 2014;234(2):391-400. [CrossRef]
- Chappell D, Jacob M, Hofmann-Kiefer K, et al. Hydrocortisone preserves the vascular barrier by protecting the endothelial glycocalyx. In Vitro.
- Research Support, Non-U.S. Gov't. Anesthesiology. Nov 2007;107(5):776-84. [CrossRef]
- Chappell D, Hofmann-Kiefer K, Jacob M, et al. TNF-alpha induced shedding of the endothelial glycocalyx is prevented by hydrocortisone and antithrombin. Research Support, Non-U.S. Gov't. Basic research in cardiology. Jan 2009;104(1):78-89. [CrossRef]
- Aytac HO, Iskit AB, Sayek I. Dexamethasone effects on vascular flow and organ injury in septic mice. J Surg Res. May 15 2014;188(2):496-502. [CrossRef]
- Voiriot G, Oualha M, Pierre A, et al. Chronic critical illness and post-intensive care syndrome: from pathophysiology to clinical challenges. Ann Intensive Care. 2022;12(1):1-14.
- Basil MC, Levy BD. Specialized pro-resolving mediators: endogenous regulators of infection and inflammation. Nature Reviews Immunology. 2016/01/01 2016;16(1):51-67. [CrossRef]
- Vago JP, Tavares LP, Garcia CC, et al. The role and effects of glucocorticoid-induced leucine zipper in the context of inflammation resolution. J Immunol. May 15 2015;194(10):4940-50. [CrossRef]
- Morioka S, Maueröder C, Ravichandran KS. Living on the edge: efferocytosis at the interface of homeostasis and pathology. Immunity. 2019;50(5):1149-1162.
- Espinasse M-A, Hajage D, Montravers P, et al. Neutrophil expression of glucocorticoid-induced leucine zipper (GILZ) anti-inflammatory protein is associated with acute respiratory distress syndrome severity. journal article. Ann Intensive Care. 2016;6(1):105. [CrossRef]
- Desgeorges T, Caratti G, Mounier R, Tuckermann J, Chazaud B. Glucocorticoids shape macrophage phenotype for tissue repair. Front Immunol. 2019;10:1591.
- Schif-Zuck S, Gross N, Assi S, Rostoker R, Serhan CN, Ariel A. Saturated-efferocytosis generates pro-resolving CD11blow macrophages: Modulation by resolvins and glucocorticoids. European journal of immunology. 2011;41(2):366-379.
- Meduri GU, Schwingshackl A, Hermans G. Prolonged Glucocorticoid Treatment in ARDS: Impact on Intensive Care Unit-Acquired Weakness. Mini Review. Front Pediatr. 2016-August-2 2016;4:69. [CrossRef]
- Wang T-T, Nestel FP, Bourdeau V, et al. Cutting edge: 1, 25-dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. The Journal of Immunology. 2004;173(5):2909-2912.
- Dimitrov V, Barbier C, Ismailova A, et al. Vitamin D-regulated Gene Expression Profiles: Species-specificity and Cell-specific Effects on Metabolism and Immunity. Endocrinology. Feb 1 2021;162(2):bqaa218. [CrossRef]
- Greulich T, Regner W, Branscheidt M, et al. Altered blood levels of vitamin D, cathelicidin and parathyroid hormone in patients with sepsis-a pilot study. Anaesth Intensive Care. Jan 2017;45(1):36-45.
- Ang A, Pullar JM, Currie MJ, Vissers MCM. Vitamin C and immune cell function in inflammation and cancer. Biochem Soc Trans. Oct 19 2018;46(5):1147-1159. [CrossRef]
- Sorice A, Guerriero E, Capone F, Colonna G, Castello G, Costantini SJMrimc. Ascorbic acid: its role in immune system and chronic inflammation diseases. 2014;14(5):444-452.
- Sae-Khow K, Tachaboon S, Wright HL, et al. Defective neutrophil function in patients with sepsis is mostly restored by ex vivo ascorbate incubation. Journal of Inflammation Research. 2020;13:263.
- Kim Y, Kim H, Bae S, et al. Vitamin C is an essential factor on the anti-viral immune responses through the production of interferon-α/β at the initial stage of influenza A virus (H3N2) infection. Immune Netw. 2013;13(2):70-74.
- Combs Jr GF, McClung JP. The vitamins: fundamental aspects in nutrition and health. Edition 5. Chapter 11 Thiamin. Academic press; 2016:298-314.
- Collie JTB, Greaves RF, Jones OAH, Lam Q, Eastwood GM, Bellomo R. Vitamin B1 in critically ill patients: needs and challenges. Clin Chem Lab Med. Oct 26 2017;55(11):1652-1668. [CrossRef]
- Ryan ZC, Craig TA, Folmes CD, et al. 1α, 25-dihydroxyvitamin D3 regulates mitochondrial oxygen consumption and dynamics in human skeletal muscle cells. 2016;291(3):1514-1528.
- Luo G, Xie ZZ, Liu FY, Zhang GB. Effects of vitamin C on myocardial mitochondrial function and ATP content in hypoxic rats. Zhongguo Yao Li Xue Bao. Jul 1998;19(4):351-5.
- Charoenngam N, Holick MF. Immunologic effects of vitamin D on human health and disease. Nutrients. 2020;12(7):2097.
- Dancer RC, Parekh D, Lax S, et al. Vitamin D deficiency contributes directly to the acute respiratory distress syndrome (ARDS). Thorax. Jul 2015;70(7):617-24. [CrossRef]
- Tyml K. Vitamin C and microvascular dysfunction in systemic inflammation. Antioxidants. 2017;6(3):49.
- Oudemans-van Straaten HM, Elbers PW, Spoelstra-de Man AM. How to give vitamin C a cautious but fair chance in severe sepsis. Editorial. Chest. 2017;151(6):1199-1200.
- Marik PE. Glucocorticosteroids as Adjunctive Therapy for Acute Respiratory Distress Syndrome and Sepsis? Yes, But Not as Monotherapy. Critical care medicine. 2017;45(5):910-911.
- Carr AC, Shaw GM, Natarajan R. Ascorbate-dependent vasopressor synthesis: a rationale for vitamin C administration in severe sepsis and septic shock? Critical Care. 2015;19(1):1-8.
- Mohammed BM, Fisher BJ, Kraskauskas D, et al. Vitamin C: a novel regulator of neutrophil extracellular trap formation. Nutrients. Aug 9 2013;5(8):3131-51. [CrossRef]
- Yadav UC, Kalariya NM, Srivastava SK, Ramana KV. Protective role of benfotiamine, a fat-soluble vitamin B1 analogue, in lipopolysaccharide-induced cytotoxic signals in murine macrophages. Free Radic Biol Med. May 15 2010;48(10):1423-34. [CrossRef]
- Bozic I, Savic D, Laketa D, et al. Benfotiamine attenuates inflammatory response in LPS stimulated BV-2 microglia. PLoS One. 2015;10(2):e0118372.
- Menezes RR, Godin AM, Rodrigues FF, et al. Thiamine and riboflavin inhibit production of cytokines and increase the anti-inflammatory activity of a corticosteroid in a chronic model of inflammation induced by complete Freund's adjuvant. Pharmacol Rep. Oct 2017;69(5):1036-1043. [CrossRef]
- Bagnoud M, Hoepner R, Pistor M, et al. Vitamin D augments glucocorticosteroid efficacy via inhibition of mTORc1. 2018:33-34.
- Zhang Y, Leung DY, Goleva E. Vitamin D enhances glucocorticoid action in human monocytes: involvement of granulocyte-macrophage colony-stimulating factor and mediator complex subunit 14. J Biol Chem. May 17 2013;288(20):14544-53. [CrossRef]
- Kassel O, Sancono A, Krätzschmar J, Kreft B, Stassen M, Cato AC. Glucocorticoids inhibit MAP kinase via increased expression and decreased degradation of MKP-1. The EMBO journal. Dec 17 2001;20(24):7108-7116. [CrossRef]
- Zhang Y, Leung DY, Richers BN, et al. Vitamin D inhibits monocyte/macrophage proinflammatory cytokine production by targeting MAPK phosphatase-1. J Immunol. Mar 1 2012;188(5):2127-35. [CrossRef]
- Stio M, Martinesi M, Bruni S, et al. The Vitamin D analogue TX 527 blocks NF-kappaB activation in peripheral blood mononuclear cells of patients with Crohn's disease. J Steroid Biochem Mol Biol. Jan 2007;103(1):51-60. [CrossRef]
- Ojaimi S, Skinner NA, Strauss BJ, Sundararajan V, Woolley I, Visvanathan K. Vitamin D deficiency impacts on expression of toll-like receptor-2 and cytokine profile: a pilot study. J Transl Med. Jul 22 2013;11(1):176. [CrossRef]
- Patak P, Willenberg HS, Bornstein SR. Vitamin C is an important cofactor for both adrenal cortex and adrenal medulla. Endocr Res. 2004;30(4):871-875. [CrossRef]
- Moskowitz A, Andersen LW, Huang DT, et al. Ascorbic acid, corticosteroids, and thiamine in sepsis: a review of the biologic rationale and the present state of clinical evaluation. Crit Care. Oct 29 2018;22(1):283. [CrossRef]
- Okamoto K, Tanaka H, Makino Y, Makino I. Restoration of the glucocorticoid receptor function by the phosphodiester compound of vitamins C and E, EPC-K1 (L-ascorbic acid 2-[3,4-dihydro-2,5,7,8-tetramethyl-2-(4,8,12-trimethyltridecyl)-2H-1-benzopyran-6 -yl hydrogen phosphate] potassium salt), via a redox-dependent mechanism. Biochem Pharmacol. Jul 1 1998;56(1):79-86.
- Chen Y, Luo G, Yuan J, et al. Vitamin C mitigates oxidative stress and tumor necrosis factor-alpha in severe community-acquired pneumonia and LPS-induced macrophages. Mediators Inflamm. 2014:426740. [CrossRef]
- Bowie AG, O’Neill LA. Vitamin C inhibits NF-κB activation by TNF via the activation of p38 mitogen-activated protein kinase. The Journal of Immunology. 2000;165(12):7180-7188.
- Donnino MW, Andersen LW, Chase M, et al. Randomized, Double-Blind, Placebo-Controlled Trial of Thiamine as a Metabolic Resuscitator in Septic Shock: A Pilot Study. Critical Care Medicine. Feb 2016;44(2):360-367. [CrossRef]
- Schreiber SN. The transcriptional coactivator PGC-1 [alpha] as a modulator of ERR [alpha] and GR signaling: function in mitochondrial biogenesis. University_of_Basel; 2004.
- Latham CM, Brightwell CR, Keeble AR, et al. Vitamin D Promotes Skeletal Muscle Regeneration and Mitochondrial Health. Front Physiol. 2021;12:463.
- Jain SK, Micinski D. Vitamin D upregulates glutamate cysteine ligase and glutathione reductase, and GSH formation, and decreases ROS and MCP-1 and IL-8 secretion in high-glucose exposed U937 monocytes. Biochem Biophys Res Commun. Jul 19 2013;437(1):7-11. [CrossRef]
- Mohammed BM, Fisher BJ, Kraskauskas D, et al. Vitamin C promotes wound healing through novel pleiotropic mechanisms. Int Wound J. 2016;13(4):572-584.
- Vissers M, Hampton M. The role of oxidants and vitamin C on neutrophil apoptosis and clearance. Biochem Soc Trans. 2004;32(3):499-501.
Figure 1.
Glucocorticoid receptor-α regulation of the three major phases of homeostatic corrections. Legend: The glucocorticoid receptor alpha (GRα) serves as a primary regulator of essential physiological processes, playing a central role in sustaining life by orchestrating the body’s adaptive responses to stress and maintaining intricate homeostatic balance. GRα ensures two fundamental functions critical to survival: (i) the provision of an adequate bioenergetic supply by mobilizing glucose, fatty acids, and other substrates required to meet the energy demands of cells and tissues under stress; and (ii) the precise regulation of the neuroendocrine-immune axis, harmonizing the interactions between hormones, neurotransmitters, and immune signals. This fine-tuned coordination allows GRα to modulate the body's response across the three distinct phases of adaptive homeostatic corrections: In the Priming Phase (1a. and 1b.), the GRα activates early cellular and molecular pathways that prepare the body for an immediate defense, enhancing energy availability and initiating immune surveillance. During the Modulatory Phase (2), the GRα primary goal is to prevent further damage by downregulating the inflammatory response and reducing oxidative stress. In the Restorative Phase (3a.3b.), the GRα supports the resolution of inflammation and recovery of systemic balance, ensuring long-term stabilization by promoting tissue repair, replenishing energy stores, and returning the neuroendocrine-immune system to its baseline state. By tightly regulating these processes, GRα enables the body to adapt dynamically to environmental and internal challenges, safeguarding health and ensuring survival. It's essential to recognize that disease processes can vary widely depending on the specific condition and individual factors. There may be varying degrees of overlap, and these phases may not always occur in a linear fashion Modified with permission from reference [18]. Abbreviations: Angpt-1, angiopoietin-1; AnxA1, annexinA1; ALXR, AnxA1 receptor; AP-1, activator protein 1; APR, acute phase response; APPs, acute phase proteins; BBB, blood brain barrier; GILZ, glucocorticoid-induced leucine zipper; NOD-like receptor pyrin containing 3; eNOS, endothelial nitric oxide synthase; IFN-g, interferon-g; iNOS, inducible nitric oxide synthase; IL, interleukin; MKP-1, mitogen-activated protein kinase phosphatase 1; Na+, sodium; NF-kB, nuclear factor-kB; NLRP3, NOD-like receptor family and the pyrin domain containing 3 gene; PI3K, phosphoinositide 3-kinase; purinergic receptor P2Y2R; SGK-1, serum glucocorticoid kinase-1; sICAM-1, soluble intercellular adhesion molecule-1; SphK1, sphingosine kinase 1; TF, transcription factor; TGFb, transforming growth factor beta; TIMPs, tissue inhibitors of metalloproteinases; TLR2, toll-like receptor 2; TLR4, toll-like receptor 4. VEGF, vascular endothelial growth factor.
Figure 1.
Glucocorticoid receptor-α regulation of the three major phases of homeostatic corrections. Legend: The glucocorticoid receptor alpha (GRα) serves as a primary regulator of essential physiological processes, playing a central role in sustaining life by orchestrating the body’s adaptive responses to stress and maintaining intricate homeostatic balance. GRα ensures two fundamental functions critical to survival: (i) the provision of an adequate bioenergetic supply by mobilizing glucose, fatty acids, and other substrates required to meet the energy demands of cells and tissues under stress; and (ii) the precise regulation of the neuroendocrine-immune axis, harmonizing the interactions between hormones, neurotransmitters, and immune signals. This fine-tuned coordination allows GRα to modulate the body's response across the three distinct phases of adaptive homeostatic corrections: In the Priming Phase (1a. and 1b.), the GRα activates early cellular and molecular pathways that prepare the body for an immediate defense, enhancing energy availability and initiating immune surveillance. During the Modulatory Phase (2), the GRα primary goal is to prevent further damage by downregulating the inflammatory response and reducing oxidative stress. In the Restorative Phase (3a.3b.), the GRα supports the resolution of inflammation and recovery of systemic balance, ensuring long-term stabilization by promoting tissue repair, replenishing energy stores, and returning the neuroendocrine-immune system to its baseline state. By tightly regulating these processes, GRα enables the body to adapt dynamically to environmental and internal challenges, safeguarding health and ensuring survival. It's essential to recognize that disease processes can vary widely depending on the specific condition and individual factors. There may be varying degrees of overlap, and these phases may not always occur in a linear fashion Modified with permission from reference [18]. Abbreviations: Angpt-1, angiopoietin-1; AnxA1, annexinA1; ALXR, AnxA1 receptor; AP-1, activator protein 1; APR, acute phase response; APPs, acute phase proteins; BBB, blood brain barrier; GILZ, glucocorticoid-induced leucine zipper; NOD-like receptor pyrin containing 3; eNOS, endothelial nitric oxide synthase; IFN-g, interferon-g; iNOS, inducible nitric oxide synthase; IL, interleukin; MKP-1, mitogen-activated protein kinase phosphatase 1; Na+, sodium; NF-kB, nuclear factor-kB; NLRP3, NOD-like receptor family and the pyrin domain containing 3 gene; PI3K, phosphoinositide 3-kinase; purinergic receptor P2Y2R; SGK-1, serum glucocorticoid kinase-1; sICAM-1, soluble intercellular adhesion molecule-1; SphK1, sphingosine kinase 1; TF, transcription factor; TGFb, transforming growth factor beta; TIMPs, tissue inhibitors of metalloproteinases; TLR2, toll-like receptor 2; TLR4, toll-like receptor 4. VEGF, vascular endothelial growth factor.
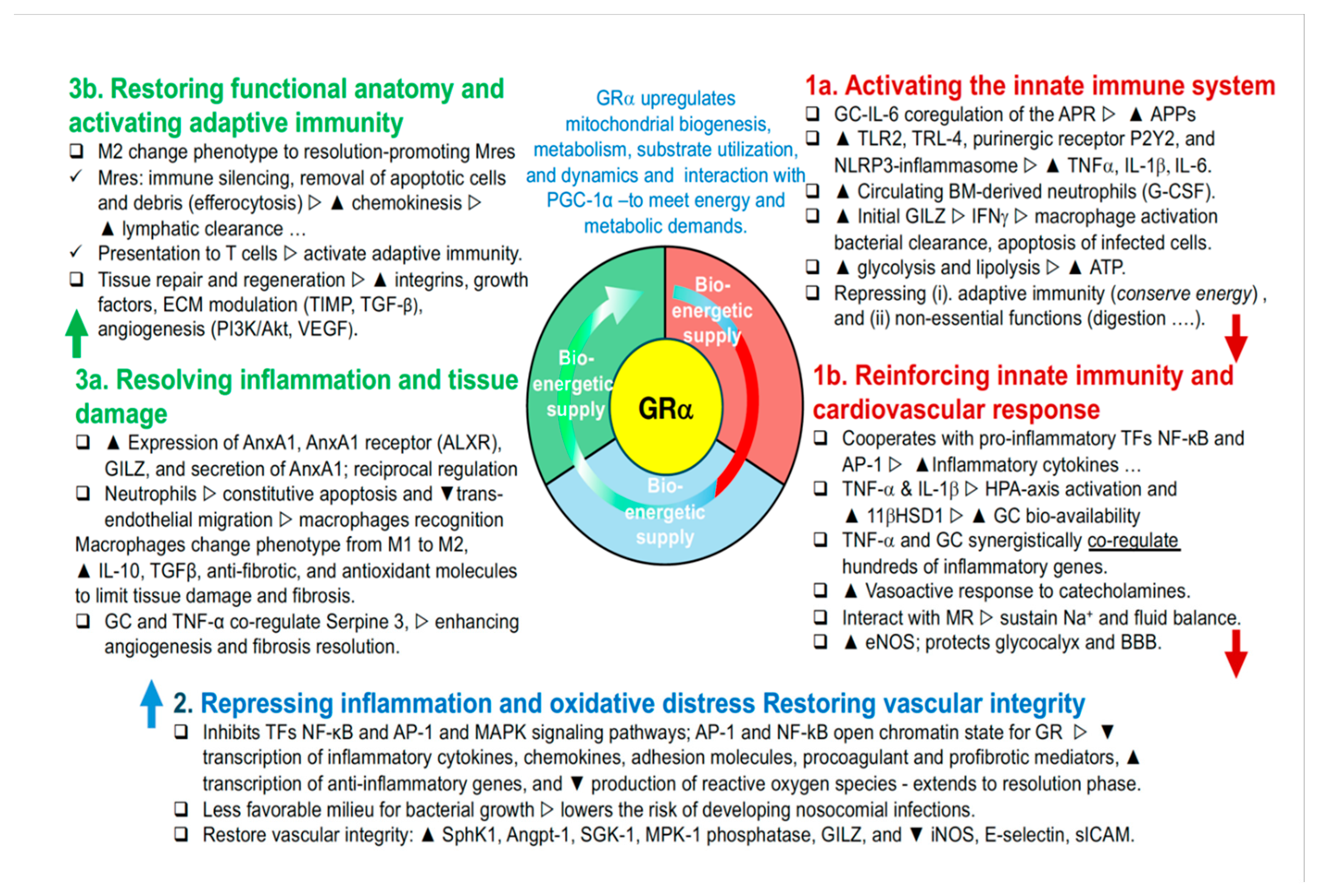

Table 1.
The Evolutionary Development of Homeostatic Corrections.
| Stage | Description | Key Points | Approximate Timeline |
|---|---|---|---|
| 1. Ancient Origins of Steroid Signaling | Evolution of steroid signaling pathways to regulate metabolism and stress responses in early vertebrates. These pathways allowed organisms to manage energy resources and respond to environmental changes. [3] | - Primitive mechanisms for managing energy resources and respond to environmental changes.- Crucial for survival. | ~450-500 million years ago |
| 2. Co-evolution with Immune, Inflammatory, and hemostatic Responses | GR co-evolved with the immune system to regulate inflammation and prevent tissue damage. [4] Hemostasis and inflammatory mechanisms evolved alongside, underscoring their interconnected roles. | - Interaction between GR, NF-kB, AP-1, and hemostasis.- Coordinated response to infection, wounds, and tissue protection. | ~400-450 million years ago |
| 3. Adaptation to Diverse Stressors | GR system evolved to manage a wide range of stressors, including infections, injuries, psychological, and metabolic stress. [50] | - GR as a master regulator.- Integrates signals from various pathways to maintain homeostasis. [50] | ~300-350 million years ago |
| 4. Integration with Mitochondrial Function | GR co-evolved with mitochondrial function, reflecting the role of energy production in stress response. Mitochondria contain glucocorticoid response elements (GREs) in mitochondrial DNA (mtDNA) [5] | - Mitochondria originated from symbiosis with proteobacteria.- GR-mediated stress response integrated with energy metabolism. MtGRE directly influence mitochondrial gene expression and energy production. | ~1.5–2 billion years ago (mitochondria origin), integration with GR: ~400 million years ago |
| 5. Essential Micronutrients and Antioxidant Systems | GR-mediated corrections rely on micronutrients and antioxidants incorporated into stress responses as organisms evolved more complex diets and metabolic systems. [57] | - Micronutrients provided a survival advantage in environments where oxidative stress and energy demands were high. | ~400 million years ago |
Table 2.
Comprehensive Roles of GRα in the Regulation of Organ and System Functions.
| Organ/System | GRα Regulation |
|---|---|
| Immune System | GRα plays a crucial role in modulating both innate and adaptive immunity by ensuring the immune response is proportionate and controlled. It regulates innate immune cells like macrophages, neutrophils, and dendritic cells, guiding their response to pathogens and injury. In adaptive immunity, GRα regulates T and B cell proliferation, differentiation, and cytokine production, helping maintain immune homeostasis and preventing autoimmunity. GRα upregulates GILZ, which attenuates MAPK/ERK signaling, and Annexin 1, which inhibits neutrophil migration, promotes macrophage-mediated clearance of apoptotic cells, and modulates T and B cell activity. As the immune response progresses, GRα shifts towards repressing pro-inflammatory mediators, promoting the resolution of inflammation and preventing chronic immune activation, thus ensuring a balanced and effective immune response. |
| Lymphatic system | The glucocorticoid receptor (GR) plays a significant role in modulating the function of lymphatic endothelial cells, which are crucial for the integrity and operation of lymphatic vessels. By influencing the permeability and contractility of these vessels, GR affects the flow of lymph, which is essential for the transport of immune cells and antigens throughout the body. Additionally, GR regulates the expression of various transporters and receptors within the lymphatic system, thereby enhancing the efficiency of lymphatic clearance and ensuring effective immune surveillance and response |
| Central Nervous System (CNS) | GRα plays a crucial role in regulating the hypothalamic-pituitary-adrenal (HPA) axis, controlling the body's response to stress and helping to restore homeostasis once a threat has passed. In the CNS, GRα modulates neurotransmitter systems, including serotonergic and dopaminergic pathways, influencing mood, cognition, and behavior. It also impacts synaptic transmission by regulating the release and uptake of neurotransmitters, which affects neuronal excitability and synaptic plasticity, further influencing cognitive functions such as memory, attention, and executive function. Additionally, GRα is involved in mood regulation by modulating the activity of brain regions like the amygdala and prefrontal cortex, and it supports brain energy metabolism by regulating glucose availability and utilization. |
|
Peripheral Nervous System (PNS) |
GRα regulates the function of the peripheral nervous system by modulating the stress response at the level of peripheral nerves. It influences the sensitivity of peripheral sensory neurons to pain and inflammation, helping to modulate pain perception and inflammatory responses. GRα is also involved in nerve regeneration and repair, influencing the healing process following peripheral nerve injuries by managing inflammation and tissue repair. Additionally, GRα affects the autonomic nervous system, contributing to the regulation of heart rate, blood pressure, and gastrointestinal motility under stress. |
|
Endocrine System |
GRα regulates various endocrine functions by modulating the HPA axis and influencing the production of key hormones such as cortisol, which impacts metabolism, immune response, and stress adaptation. GRα also interacts with other hormones, like insulin, thyroid hormones, and reproductive hormones, ensuring coordinated endocrine responses to stress and maintaining overall hormonal balance. |
| Reproductive System | GRα influences reproductive function by regulating the expression of genes involved in hormone production, ovulation, and pregnancy. It modulates the effects of stress on reproductive health, ensuring that stress responses do not interfere with normal reproductive processes. GRα also plays a role in fetal development by regulating placental function and fetal growth. |
|
Cardiovascular System |
GRα plays a vital role in regulating blood pressure, vascular tone, and overall heart function. It modulates the expression of genes involved in the production of vasodilators, such as endothelial nitric oxide synthase (eNOS), essential for maintaining vascular health. GRα also influences glucose uptake, utilization, and storage in the heart, ensuring that cardiac cells have sufficient energy during stress. Additionally, GRα affects cardiac electrophysiology by modulating ion channel function and action potential duration, which is crucial for maintaining normal heart rhythms. |
| Endothelium | GR plays a key role in endothelial homeostasis by regulating the expression of adhesion molecules and cytokines involved in the inflammatory response, thereby reducing inflammation. It also maintains the integrity of the endothelial barrier and glycocalyx, regulates vascular tone and blood pressure through nitric oxide production, protects against oxidative stress, and promotes angiogenesis and vascular repair. These functions are essential for preserving vascular integrity and preventing diseases like atherosclerosis and hypertension. |
| Lungs | GRα plays a vital role in maintaining lung function by modulating the immune response to inhaled pathogens and allergens, reducing airway inflammation, and preventing excessive immune responses that can lead to tissue damage. It supports the repair of lung tissue following injury or infection, ensuring proper respiratory function. Additionally, GRα exerts bronchodilatory effects by relaxing airway smooth muscle and modulates the expression of genes involved in maintaining smooth muscle tone and producing surfactant, a substance crucial for keeping the airways open and facilitating efficient gas exchange. |
| Kidneys | In the kidneys, GRα is involved in the regulation of electrolyte balance and fluid homeostasis. It influences the expression of sodium channels and transporters in the renal tubules, which helps control the reabsorption of sodium and water. This regulation is essential for maintaining blood pressure and overall fluid balance in the body. |
| Liver | GRα plays a crucial role in maintaining liver health and overall immune homeostasis by regulating the expression of key genes involved in metabolism and immune function. In the liver, GRα modulates glucose metabolism by promoting gluconeogenesis and influencing glycogen storage, ensuring that the body has sufficient energy during periods of stress or inflammation. Additionally, GRα is integral to lipid metabolism, detoxification processes, and bile acid metabolism, all essential for processing lipids, eliminating toxins, and supporting digestive functions. GRα enhances protein synthesis, crucial for producing acute phase proteins and enzymes necessary for immune responses and detoxification. Furthermore, GRα works in concert with IL-6 to coactivate the acute phase response (APR), boosting the liver’s production of proteins that manage inflammation and bolster immune defenses during injury or stress. |
| Gastrointestinal Tract | GRα regulates gastrointestinal function by modulating the immune response within the gut. It helps maintain the integrity of the gut lining by controlling inflammation and promoting the repair of damaged tissues. Additionally, GRα plays a role in regulating the gut microbiome by modulating the local immune environment and inflammatory responses, and influences gut motility and secretion, contributing to the proper digestion and absorption of nutrients. |
| Pancreas | GRα helps regulate insulin production and glucose homeostasis in the pancreas. It modulates the function of pancreatic beta cells, which are responsible for insulin secretion. By balancing the production and release of insulin, GRα helps maintain normal blood glucose levels, particularly during stress or fasting. The GR is expressed in various cell types within the pancreatic islets, including beta cells, alpha cells, and delta cells. Activation of the GR in these cells can affect their function and hormone secretion |
| Adipose Tissue | GRα is involved in lipid metabolism within adipose tissue. It promotes lipolysis, the breakdown of stored triglycerides into free fatty acids and glycerol, which can then be used as energy sources. GRα also regulates the balance between lipid storage and mobilization, ensuring energy availability during periods of stress or fasting. |
| Muscle | GRα is important for preserving muscle function, especially during stress. It regulates protein breakdown to provide energy through gluconeogenesis while also modulating inflammation to support muscle repair after injury. GRα helps maintain a balance between muscle breakdown and building, ensuring muscle strength and resilience during stress or recovery. |
| Bone | GRα plays a role in maintaining bone health by regulating the balance between bone formation and resorption. It influences the activity of osteoblasts (bone-forming cells) and osteoclasts (bone-resorbing cells), ensuring proper bone remodeling and mineral homeostasis, which is crucial for maintaining bone density and structural integrity. |
| Skin | GRα helps maintain skin homeostasis by regulating the skin's inflammatory response. It controls the production of cytokines and other inflammatory mediators, ensuring that the skin's immune responses are appropriate and do not result in excessive inflammation. This is important for protecting the skin from infections and environmental stressors. |
Legend: This table outlines the essential functions of GRα in various organs and systems under normal conditions. GRα plays a critical role in maintaining homeostasis, modulating stress responses, and ensuring the proper functioning of vital physiological processes. This table was generated with the assistance of AI using GPT-4. I would like to acknowledge the assistance of AI-powered tools in the research, structuring, and refinement of this table.
Table 3.
Roles of GRα in the Regulation of Circulating Cells.
| Cell Type | Brief Description of GRa Role |
|---|---|
| 1. Immune Cells | |
| - T cells | GRa modulates T cell function by inhibiting the production of pro-inflammatory cytokines (e.g., IL-2, IFN-γ), promoting the differentiation of regulatory T cells (Tregs), and reducing the proliferation of effector T cells. This results in the suppression of excessive immune responses and maintenance of immune tolerance. |
| - B cells | GRa plays a role in suppressing B cell activation and differentiation into plasma cells, thereby reducing antibody production. It also impacts B cell survival and modulates the production of regulatory cytokines like IL-10, which further influences immune responses. |
|
- Monocytes/ Macrophages |
GRa regulates the transition of monocytes into macrophages and affects their polarization into either pro-inflammatory (M1) or anti-inflammatory (M2) phenotypes. It suppresses the production of inflammatory mediators (e.g., TNF-α, IL-6) and enhances phagocytic activity and tissue repair functions. |
| - Dendritic Cells | GRa modulates the maturation and function of dendritic cells (DCs), reducing their ability to present antigens and activate T cells. This leads to a decrease in adaptive immune responses and helps to maintain immune homeostasis, especially during chronic inflammation. |
| - Natural Killer (NK) Cells | GRa influences the cytotoxic activity of NK cells, reducing their ability to target and destroy virus-infected or tumor cells. It also modulates the production of cytokines like IFN-γ, which plays a role in shaping the overall immune response. |
| Eosinophils | GRa suppresses eosinophil activation, reducing the production of pro-inflammatory cytokines and chemokines involved in allergic responses and asthma. It also decreases eosinophil survival and migration, helping to control inflammation and tissue damage in allergic conditions |
| 2. Non-Immune Cells | |
| - Erythrocytes (Red Blood Cells) | GRa regulates erythropoiesis by influencing the production of erythropoietin and other factors involved in red blood cell maturation. It also impacts hemoglobin synthesis and the capacity of erythrocytes to transport oxygen, particularly under stress conditions. |
| - Platelets | GRa affects platelet function by modulating the expression of surface receptors involved in platelet activation and aggregation. This regulation is crucial in balancing hemostasis and preventing excessive clot formation during inflammation. |
| - Endothelial Progenitor Cells | GRa plays a role in the mobilization and differentiation of endothelial progenitor cells (EPCs), which are essential for vascular repair and regeneration. It also modulates the expression of factors that influence endothelial function and vascular integrity, particularly in response to injury or stress. |
Legend: This table provides an overview of the regulatory roles of GRα in various circulating cells within the body, divided into two main categories: immune cells and non-immune cells. Each cell type is listed along with a brief description of how GRa influences its function: Immune Cells: These are cells primarily involved in the body's immune response, including T cells, B cells, monocytes/macrophages, dendritic cells, and natural killer (NK) cells. GRα modulates various aspects of their activity, such as cytokine production, cell proliferation, and differentiation, to maintain immune homeostasis and prevent excessive inflammation. Non-Immune Cells: These are circulating cells not directly involved in the immune response but play critical roles in maintaining physiological balance. This category includes erythrocytes (red blood cells), platelets, endothelial progenitor cells, mesenchymal stem cells (MSCs), and eosinophils. GRα influences processes like oxygen transport, hemostasis, vascular repair, tissue regeneration, and the modulation of allergic responses and inflammation in these cells. I would like to acknowledge the assistance of AI-powered tools in the research, structuring, and refinement of this table.
Table 4.
Phases of Glucocorticoid Receptor Alpha (GRα) Regulation: Key Factors, Binding Partners, Vitamins, and Micronutrients Supporting Each Phase.
Table 4.
Phases of Glucocorticoid Receptor Alpha (GRα) Regulation: Key Factors, Binding Partners, Vitamins, and Micronutrients Supporting Each Phase.
| Temporal Phase |
Factors Affecting GRα Function |
Binding Partners |
Vitamins and Micronutrients Supporting GRα Function |
|---|---|---|---|
| Initial Activation Phase | Ligand availability (e.g., cortisol levels), adrenal gland function | Corticosteroid-binding globulin (CBG), Cortisol biosynthetic enzymes | Vitamins: B6, C; Minerals: Sodium |
| Chromatin Remodeling and Gene Accessibility | Chromatin accessibility, histone modifications | Chromatin remodeling complexes, Histone acetyltransferases (HATs) | Vitamins: B9, C; Minerals: Zinc |
| Immediate Post-translational Modifications | Phosphorylation, acetylation, ubiquitination | Kinases (e.g., MAPK), Phosphatases | Vitamins: B6, C, E; Minerals: Magnesium, Zinc |
| Early Signaling and Interaction Phase | Receptor interactions (MR, ER), cytokine signaling (e.g., TNF-α, IL-6) | Estrogen receptors (ER), Mineralocorticoid receptors (MR), NF-κB, AP-1 | Vitamins: C, E; Minerals: Selenium, Zinc; Regulatory processes: Immune signaling |
| Sustained Signaling Phase | Oxidative stress, mitochondrial function | Antioxidant enzymes (e.g., glutathione peroxidase), Mitochondrial transcription factors (e.g., TFAM) | Vitamins: C, E; Minerals: Selenium, Zinc, Magnesium, Iron |
| Long-term Regulation and Maintenance | Epigenetic modifications, gut-brain axis signaling, prolonged cortisol exposure | DNA methyltransferases, Toll-like receptors (TLRs), Hepatic enzymes (e.g., CYP3A4, 11β-HSD1, 11β-HSD2) | Vitamins: B9, B12, D, E; Minerals: Zinc, Magnesium, Iron |
Legend: Refer to the manuscript text for an explanation of the temporal organization of GRα function. Regulatory Processes: Refers to the biological mechanisms and pathways that influence GRα function during each phase, including hormone regulation, chromatin remodeling, and stress response. Binding Partners: These are molecules or proteins that interact with GRα to regulate its activity, such as enzymes, receptors, and transcription factors, crucial for cellular responses and gene regulation. I would like to acknowledge the assistance of AI-powered tools in the research, structuring, and refinement of this table.
Table 5.
Role of Thiamin, Vitamin D, and Vitamin C in the three phases of homeostatic correction.
| Homeostatic Phase |
Thiamin | Vitamin D | Vitamin C |
|---|---|---|---|
| Reinforce innate immunity | - |
Supports innate and adaptive immune system. ⇑ TLR coreceptor CD14. ⇑ antimicrobial peptides cathelicidin and LL-37. [85] ⇑ neutrophil recruitment, activation, and function. [86] ⇑ antibacterial activity. [87] |
Supports neutrophil anti-bacterial function at hypoxic inflammatory sites. [88] ⇑ neutrophil and macrophage chemotaxis, phagocytic capacity, lysozyme activity for cell elimination, and bacterial killing. [88,89,90] Supports lymphocyte proliferation and differentiation. [88] ⇑ production of type I interferons (IFNs) for anti-viral immune responses against influenza virus infection. [91] |
| Bioenergetic supply | Essential for energy metabolism and carbohydrate breakdown/ATP production [92] Thiamin pyrophosphate is a cofactor for PDH, a-KGDC. [93] |
⇑ mitochondrial number, morphology, physiology, and expression of key mitochondrial proteins, resulting in increased ATP synthesis. [94] | ⇑ ATP synthesis. [95] |
| Vascular integrity | - | Modulate endothelial function (non-genomic up-regulation of eNOS gene expression) and vascular permeability (prevents the formation of intracellular endothelial gaps) via multiple genomic and extra-genomic pathways. [96] Protective effect on the alveolar capillary membrane. [97] |
Improves endothelial permeability, microvascular and macrovascular function. [98] Preserves endothelial barrier integrity [99] in synergy with GC. [100] Cofactor for dopamine and vasopressin. [101] Down regulator of NET formation in sepsis. [90,102] |
| Repress inflammation | Exerts significant anti-inflammatory effects: (i) ⇓ activation of p38-MAPK, (ii) ⇓ degradation of Ik-Ba, and (iii) ⇑ activation and nuclear translocation of NF-kB, ⇓ expression of cytokines and chemokines, iNOS and COX-2. [103] ⇓ nuclear NF-kB/p65 protein level, ⇑ IL-10 synthesis – ⇓ synthesis of iNOS, COX-2, Hsp70, TNF-a, and IL-6. [104] Sinergy with glucocorticoids in inhibiting IL-6 transcription. [105] |
⇑ GR concentration [106] and GC function. [107] ⇓ synthesis of TNF-a and IL-1b. [87] ⇑ GC-mediated MKP-1 ⇒ ⇓ p38 MAPK-mediated inflammatory genes. [107,108,109] ⇑ IkBa expression ⇒ ⇓ NF-kB. [110,111] GR represses Vitamin D inactivator CYP24A1. |
Cofactor for GC synthesis. [112] Improves GR function. [113] Reverses oxidation of the GR. [114] GC facilitate Vit C cellular uptake ⇓ synthesis of TNF-α and IL-6. [115] ⇓ Ik-Ba degradation ⇒ ⇓ NF-kB activation and nuclear translocation. [116] |
| Repress oxidative stress | ⇑ Transketolase a key enzyme for the pentose phosphate pathway and for the synthesis of NADPH with glutathione cycling, an important antioxidant pathway. [117] |
VDR is a GR target for PGC-1a induction a. [118] Protective against ROS production [119] ⇑ Glutathione and glutamate formation ⇒ ⇓ ROS formation. [120] |
General role: electron donation as one of the most potent antioxidants Suppress NADPH oxidase (NOX) pathway. [113] Prevents the depletion of other circulatory antioxidants, such as lipid-soluble vitamin E and glutathione. [99] |
| Resolve and restore anatomical function | - | Restore alveolar epithelial barrier, promoting the proliferation of type 2 epithelial cells, and inhibiting fibroproliferation | ⇑ expression of pro-resolution and wound healing biomarkers, better matrix organization, and collagen deposition consistent with adaptive repair. [121] ⇑ neutrophils apoptosis and clearance from inflammatory sites. [122] ⇑ Collagen synthesis, recycles other antioxidants, improves wound healing |
Legend: ⇑ = increase; ⇓ = decrease. Reproduced with permission form reference [6].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



