
Submitted:
30 August 2024
Posted:
31 August 2024
You are already at the latest version
Abstract
Obesity is one of the most prevalent and damaging metabolic conditions and is a risk factor for the development of other disorders that define the metabolic syndrome. Recent studies have identified a significant role of AGE accumulation and RAGE signaling pathways in the underlying mechanisms that cause the chronic inflammatory response present in obesity which, ultimately, leads to adipose tissue dysfunction. Given the current interest and increasing evidence on possible therapeutic opportunities in the AGE-RAGE axis, the purpose of this work is to review the key findings on new small molecules and bioactive compounds that act as AGE or RAGE inhibitors and their effects on metabolic parameters of people with obesity. The search queries were defined according to the following PICO question: “What are the effects of AGE-RAGE antagonism on adipose tissue metabolism and other metabolic parameters of people with obesity? “. Article search was performed in three online databases: PubMed, Scopus, and Web of Science. In the end, 20 studies were selected for this review. Overall, the reviewed molecules showed beneficial effects on body weight, glucose tolerance, insulin resistance, inflammatory profile, and adipose tissue metabolism in mice models with obesity. There are still very few studies on human application of these molecules, but the ones that were reviewed revealed controversial data when compared to animal studies. Therefore, further human clinical trials should be performed to retrieve more conclusive data.
Keywords:
obesity
; advanced glycation end products
; receptor for advanced glycation end products
; pyridoxamine
; pentoxifylline
; phytochemicals
; aminoguanidine
; carnosine
; adipose tissue dysfunction
; metabolic inflammation
; insulin resistance
; glucose intolerance
1. Introduction
Obesity and its related metabolic disorders are the most prevalent and damaging metabolic conditions [1]. According to the World Obesity Federation, in 2020, over 2.6 billion people were overweight or obese, corresponding to 38% of the population over 5 years old [2]. Obesity impacts every country, and none have seen a decrease in obesity rates among their population. Obesity, as defined by the World Health Organization, is characterized as an abnormal or excessive accumulation of adipose tissue [3]. This condition is associated with a chronic inflammatory response that emerges, but is not exclusively derived, from the abnormal hypertrophy and hyperplasia of adipose tissue, which disrupt the normal formation of new vessels, resulting in adipocyte necrosis [1,4]. The adipose tissue is also an important secretory organ, so when the excess nutrients accumulate in adipocytes, they lead to mitochondrial disfunction and, thus, oxidative stress. This oxidative stress and inflammation are not limited to the adipose tissue and can impair various other organs functions, ultimately leading to altered adipokine and cytokine secretion and adipose tissue dysfunction.
Advanced glycation end products (AGEs) are generated by nonenzymatic irreversible modifications of macromolecules through a reaction called Maillard reaction, between aldehydes group of, for example, reducing sugars and the free amino groups of proteins, lipids, or nucleic acids [5]. The receptor for AGEs (RAGE) is a multi-ligand member of the immunoglobulin superfamily. It is expressed in many cell surfaces and plays an important role in various physiological processes, such as cell signaling, generation of reactive oxygen species (ROS), inflammatory response, and cellular events [6]. AGEs and RAGE have been implicated in numerous ageing-related and inflammatory diseases (Alzheimer’s disease, diabetes, atherosclerosis and several cancers) [5,7]. AGEs and its receptor have been implicated in the pathogenesis of obesity and associated complications. Several studies have shown higher levels of AGE accumulation and higher RAGE expression in serum, adipose tissue, and other organs such as the liver and kidneys, of mice and people with obesity [5,8,9]. Furthermore, these increased endogenous levels of AGE and RAGE and high-AGE diets have been linked to the aggravation of many metabolic parameters (glucose tolerance, insulin resistance, lipid profile, adipokine secretion) and the progression of obesity-associated comorbidities (cardiovascular complications and renal dysfunction) [10,11,12]. In physiological conditions, the formation of AGEs is slow. But, there are several endogenous and exogenous factors that contribute to the acceleration of this process. AGEs can occur during food processing, especially when foods are high in sugar, proteins, and fat and when it is cooked at high temperatures [13]. Dietary exogenous AGEs are also rapidly absorbed in the body [13]. At the same time, endogenous AGEs production is augmented in several conditions (like hyperglycemia, oxidative stress, obesity, renal insufficiency and other inflammatory diseases), that in turn are related to consuming high-AGE diets.
Giving the increasing evidence of the importance of the AGE-RAGE axis in the pathogenesis of obesity, there has been a growing interest in exploring the therapeutic opportunities in this signaling pathway. Recently, numerous studies on AGE and RAGE inhibitors as therapeutic strategies in cell-lines, animal models and humans with obesity and other metabolic disorders have been conducted. This work aims to collect and review knowledge on small molecules and bioactive compounds that act as AGE or RAGE inhibitors and their effects on metabolic parameters of people with obesity, and propose new research directions on the subject.
2. Methods
2.1. Search Query and Inclusion Criteria
This narrative review was conducted and reported according to the Scale for the Assessment of Narrative Review Articles (SANRA). The participants, intervention comparators, outcomes, and study design (PICO) criteria were adapted from the following question “What are the effects of AGE-RAGE antagonism on adipose tissue metabolism and other metabolic parameters of people with obesity?“ and are: “P: obese population; I: AGE or RAGE inhibitors; C: no treatment obese population; O: effects of the antagonism of AGE-RAGE axis in adipose tissue metabolism”. Original articles were searched using the following electronic databases: MEDLINE (PubMed, www.pubmed.com), Web of Science (www.webofscience.com/), and Scopus (www.scopus.com). The search query included the keywords “obese”, “obesity”, “advanced glycation end products”, “Receptor for Advanced Glycation End Products”, “RAGE”, “sRAGE”, “inhibitor”, “inhibition”, “antagonist”, “small molecule”, “liraglutide”, “pyridoxamine”, “azeliragon”, “S100-derived peptide”, “HMGB1-derived Peptide”, “alagebrium”, “carnosine”, “FPS-ZM1”, “GM-1111”, “semaglutide”, “GLP1 analog”, “adipose tissue metabolism”, “adipose tissue dysfunction”, “metabolic inflammation” and “insulin resistance”. Search strategies in all databases are described in Table 1. There were no data or language restrictions applied. The last search was conducted on May 23rd, 2024. A reverse manual search was also performed to identify relevant articles cited in the selected studies.
The inclusion criteria are studies that assess the effects of molecules with AGE or RAGE antagonizing properties in metabolic parameters (such as weight, adipose tissue metabolism, insulin resistance, glucose tolerance impairment, metabolic inflammation, lipid metabolism) of human or animal models of obesity, with or without other metabolic disorders or comorbidities.
2.2. Study Selection
Study selection was divided in three phases. First, included studies were screened through reading of titles and abstracts. After, full-texts of eligible articles were retrieved and read thoroughly before deciding to include or exclude. Authors were contacted to retrieve full-text articles.
3. Results
3.1. Search Results
The search performed retrieved 937 records. After resolving duplicates, 888 articles were left for the first screening. After title and abstract reading, 50 studies were selected for full-text screening. In the end, 20 articles met the inclusion criteria and were eligible to be included in this review. Study selection is further described on a flow diagram in Figure 1. The characteristics and the key findings of each study are summarized in Table 2 and described in the following subsections, with data aggregated by molecule. 26 articles were excluded because they did not evaluate the anthropomorphic and metabolic parameters mentioned above, or effects on the AGE-RAGE axis. 2 were excluded because their study population did not include animals or humans with obesity.
3.2. Pyridoxamine
Pyridoxamine (PM) is the 4-aminomethyl form of vitamin B6, that inhibits the formation of AGEs by trapping its intermediates and blocking oxidative degradation[35]. Most of the existing studies with this substance analyzed in this review are animal studies, with obesity and/or insulin resistant rat models, focusing on various parameters including weight gain, adipocyte hypertrophy, lipid metabolism, inflammation, insulin resistance, and glucose tolerance.
Alderson N. et al. described the effects of PM treatment in Zucker rats with obesity. After 30 weeks of treatment, they found no significant difference in body weight between the fatty control group and the treated group. Likewise, PM showed no effect on glycemia, demonstrated by the mean values of fasting blood glucose (in mmol/L: 6.6±0.4 lean control; 6.9±0.2 fatty control vs 6.8±0.5 fatty PM-treated) and total glycated hemoglobin (in %: 6.6±0.4 lean group vs 6.9±0.2 fatty control vs 6.8±0.5 fatty PM-treated) [14]. In accordance, they also found that PM had no significant effect on plasma fasting insulin levels between the two obese groups (7.1±0.6 µg/L fatty control vs 7.3±0.6 µg/L in fatty PM-treated at 30 weeks). However, there was a difference in the lipid profile of the two fatty groups. PM treatment significantly reduced triglycerides and cholesterol levels, with mean values in the treated group being 50% (P< 0.0001) and 40%(P<0.0001), respectively, lower than those of the fatty control [14].
Muellenbach E. et al. [15] performed a study to determine the effects of individual and combined treatments with PM and R-(+)-α-lipoic acid (R-ALA) on whole-body and skeletal muscle insulin resistance in Zucker rats with obesity. After a 6-week treatment, they found that individual treatment with PM did not affect average final body weight and rather increased total abdominal fat, as a percentage of total final body weight in comparison with the obese control group. There was also no significant change in fasting plasma values for glucose, insulin, and fatty-free acids (FFA). PM treatment did significantly reduce oxidative stress biomarkers (by 35%) and positively impacted glucose tolerance and insulin sensitivity (reduction of 14% compared to the obesity control) [15]. The combination of the PM and R-ALA treatments resulted in substantial reductions in the parameters above, eliciting the only significant amelioration of body weight gain (23%, P < .05), causing the greatest reductions in fasting plasma glucose (in mg/dL: 98 ± 3 in PM+R-ALA vs 127 ± 5 in obesity control), insulin (in μU/mL: 130 ± 8 PM+R-ALA vs 154 ± 10 obesity control), and FFA (in mmol/L: 1.23 ± 0.04 vs 1.62 ± 0.07) and resulted in the greatest improvement in glucose tolerance and whole-body insulin sensitivity during the glucose tolerance test (HOMA-IR units: 31.2 ± 1.2 vs 48.3 ± 3.9; reduction of 35%) [15]. At 22 weeks of treatment, PM elicited significant enhancements of metabolic regulation, inducing a significant diminution of fasting plasma glucose and fasting insulin resistance (reflected by HOMA-IR) and improved glucose tolerance [16]. Moreover, treatment with PM alone significantly reduced muscle tissue oxidative damage, and muscle triglyceride levels were similarly decreased. These findings are in line with those of Unoki-Kubota et al. [18]. They also found no significant difference in blood glucose levels of KK-Ay mice in the PM-treated group compared with control and the slight reduction in body weight was only significant with the higher dose of PM at 240 mg/l (34.5±1.0 g in PM group vs 39.3±2.6g in control, P=0.015). Moreover, they found a dose-dependent effect of PM in decreasing serum levels of AGE and insulin (which were positively correlated with each other, R2=0.7568, P<0.001), as well as in the improvement of insulin sensitivity during an insulin tolerance test (ITT). Three more studies analyzed in this review showed that obesity-associated metabolic dysfunction and complications in mice with obesity were ameliorated by an intervention with PM [17,23,28]. Increases in body weight, hyperglycemia, hypercholesterolemia, and levels of leptin and insulin were all reduced by PM treatment, as well as it generated an increase in glucose tolerance and insulin sensitivity [17,23,28].
Besides the effects in these metabolic parameters, Maessen et al. also found that PM demonstrated inhibition of expansion of visceral adipose tissue and reduced hypertrophy of adipocytes in mice with obesity [23]. This inhibition of adipogenesis by PM in high-fat diet (HFD) mice did not lead to fatty liver disease. On the contrary, PM treatment resulted in less fat content in HFD livers. Hagiwara et al. and Seyeon O. et al. also demonstrated a PM-induced reduction in white adipose tissues [17,28]. PM was also associated with a normalization of the inflammatory phenotype of adipose tissues through reduction of the accumulation of macrophages, expression of pro-inflammatory cytokines (like TNFα, MCP-1, CD11c, and MHC-II) [23,28], increase of the expression of the anti-inflammatory cytokine adiponectin, increased antioxidative enzymes in white adipose tissues[17]. Moreover, PM led to an increase in glyoxalase-1 (GLO 1) activity [23,28], an important enzyme in the detoxification of methylglyoxal (MGO), one of the most important AGE precursors. However, the anti-inflammatory and anti-adipogenic effects seem to be dependent on the adipocyte location, as Seyeon O. et al. found that pyridoxamine only demonstrated these changes significantly on visceral and perivascular fat tissue, but not in subcutaneous fat tissue of HFD mice [28].
Still, in a recent study, Van den Eynde M. et al. tested the effects of 8-week treatment with PM in a randomized double-blind placebo-controlled trial on individuals with abdominal obesity. They described a significant reduction in plasma MGO levels (reduction of plasma MGO of 22 nmol/L with supplementation of 200mg/day compared to placebo, 95% CI [- 39, - 4]; P = 0.017) and total plasma AGEs (Z-score: - 0.38; 95% CI [-0.77, 0.02]; P = 0.06) [32]. Still, PM treatment did not show significant changes in weight, BMI, fat percentage, and waist circumference. Likewise, there was also no significant difference in fasting plasma glucose, insulin, and C-peptide concentrations nor in insulin sensitivity. The oral glucose tolerance test values were also not affected by PM supplementation. Furthermore, metabolic inflammation was also evaluated and they found no significant changes in plasma levels of IL-6, IL-8, IL-10, TNF-α, C-reactive protein, and serum amyloid A [32]. Despite some beneficial effect of PM administration on animal models, there’s still a lack of human information.
3.3. Carnosine
Carnosine (β-alanyl-l-histidine) is a dipeptide naturally present in the heart, skeletal muscle, and brain. It’s refered as having anti-inflammatory, antioxidant, anti-glycation, anti-ischaemic, and chelating roles[36]. Carnosine (CAR) inhibits AGEs and advanced lipoxidation end-products (ALEs) formation produced by both a non-enzymatic glycation and a direct reaction with reactive carbonyl species (RCS) generated by lipid and sugar oxidation [22]. Animal studies found that CAR and its analog (carnosinol) reduced obesity-related diseases in rats with obesity, significantly restraining the development of dyslipidemia, hypertension, and even renal injury. Aldini G. et al. [20] reported that treatment with L-CAR or D-CAR did not significantly affect the plasma glucose levels. However, plasma insulin concentration increase in Zucker rats was significantly restrained by L-CAR and D-CAR, which also improved insulin resistance. They also found a significant decrease in body weight (35.8% by L-CAR and 40% by D-CAR) and plasma and kidney cholesterol (in (µg/mg: L-CAR 6.31 ± 0.5115 and D-CAR 6.668 ± 0.5087 vs 9.757 ± 0.895 in untreated control; P<0.05) and triglycerides (in µg/mg: L-CAR 12.01 ± 0.9528 and D-CAR 10.66 ± 0.3616 vs 17.92 ± 2.135 untreated control; P<0.05) levels with a 24-week treatment of obese Zucker rats with L-CAR and D-CAR [20]. Anderson E. et al. found that carnosinol at the high dose of 45 mg/kg/day had no effect on body weights and adiposity of high-fat/high-sucrose (HFHS) diet-induced obesity mice, but fasting serum triglyceride and cholesterol levels were decreased in treated mice. Carnosinol treatment also led to enhanced glucose disposal following oral glucose challenge, despite not significantly improving insulin sensitivity [26]. On one hand, a systematic review of randomized controlled trials on humans with obesity or diabetes undergoing long term (2 weeks) supplementation with carnosine [31] found no significant difference between the carnosine group and the control group in terms of triglycerides (−14.46 mg/dl, 95 % CI: −29.11, 0.19, I2:94 %), cholesterol (−2.79 mg/dl, 95 % CI: −14.59, 9.02,I2:89 %), HDL (6.38 mg/dl, 95 % CI: −6.46, 19.22, I2:98 %), and HOMA-IR levels (−0.41 mg/dl, 95 % CI: −1.00,0.18, I2:66 %) [31]. On the other hand, a recent review [22] showed that supplementation with 2g/day of carnosine for 12 weeks reduced fasting glucose and insulin, insulin resistance, as well as insulin secretion in nondiabetic overweight and obese individuals. It also showed a decrease in fat mass and an increase in fat-free mass with a trend for improvement of insulin secretion, although these last two effects were seen in a carnosine, cinnamon, and chromium supplement combination trial, therefore it is not clear if these effects are due to any individual compound or a combination of them.
3.4. Aminoguanidine
Xiong D. et al. studied the effect of aminoguanidine treatment for 6 weeks in Sprague Dawley (SD) rats with high-fat diet-induced obesity. They found that treatment with 100 mg/kg/day of aminoguanidine, in comparison with the obesity control group, significantly reduced body weight (675.32g±39.73 fatty control vs 569.71g±31.25 treated group; P<0.05), serum AGE levels (P<0.05) and total cholesterol (1.83±0.66 vs 1.51±0.31 P<0.05), triglycerides (2.65±0.75 vs 1.77±0.39, P<0.05) and LDL-C levels (1.17±0.36 vs 0.92±3.23, P<0.05), while increasing HDL-C levels (0.51±0.08 vs 0.78±0.09, P<0.05) [24]. Additionally, aminoguanidine treatment effectively suppressed the expression of TNF-α and IL-6 mRNA in liver tissues and decreased serum levels of these cytokines, in comparison with the fatty untreated group.
Aminoguanidine has been extensively studied in animal models and it showed promising effects as a novel therapeutic intervention, mainly in diabetic complications such as diabetic nephropathy [37]. At least two randomized, double-blinded, placebo-controlled human trials were performed to evaluate the efficacy and safety of this molecule in attenuating the progression of nephropathy in patients with type 1 and type 2 diabetes[38,39]. The study in patients with type 1 diabetes did not show a statistically significant benefit in the use of aminoguanidine on the progression of overt nephropathy [39] and the study on patients with type 2 diabetes was terminated due to safety concerns and lack of efficacy. In the latter study, patients reported several side-effects, such as gastrointestinal symptoms, flu-like symptoms, abnormalities in liver function tests, and, rarely, an anti-neutrophil cytoplasmic antibody (ANCA)–associated vasculitis [37,38].
3.5. Nateglinide and Telmisartan
Miura K. et al. [19] investigated the effect of a 6-week treatment with combination therapy of NAT and TEL on insulin resistance in the Zucker fatty animal model of obesity and discovered if said effect was brought through the suppression of the AGE-RAGE axis. In a previous paper with the same study design, they reported that NAT + TEL treatment, but not monotherapies, significantly decreased fasting plasma insulin, triglycerides, and FFA levels, and also improved ITT values [40]. Later on, they demonstrated that the same combination therapy inhibited serine phosphorylation (mostly regarded as a negative regulator of IRS-1 function, inhibiting insulin signaling [41]) and restored tyrosine phosphorylation ( which results in intracellular transduction of the insulin signal [41]) levels of IRS-1 in the liver of Zucker fatty rats [19]. Furthermore, treatment with NAT+TEL significantly diminished serum levels of AGEs, hepatic RAGE expression levels, and hepatic AGE-RAGE index [19]. These results suggest that combination therapy with NAT and TEL could improve metabolic parameters by decreasing AGE burden and by suppressing the AGE-RAGE axis in the liver. These findings cast a light on a possible novel application for these familiar and well known drugs, as a treatment of obesity, but further human data is needed.
3.6. Pentoxifylline
Inacio M. et al. performed an experimental study on C57BL-6J mice, fed with a high-fat diet to induce obesity and treated with 50 mg/kg pentoxifylline (PTX) for 7 weeks. They evaluated the effect of PTX on glycoxidative stress in mice with obesity, in particular, changes in the renal levels of AGE metabolism components. They found PTX treatment promoted a significant reduction in body weight and weight of adipose tissues, with the treated group mice showing medium values of body weight 21% lower than those of high-fat diet control group [30], although they attributed these effects, at least partially, to reductions in food and energy intake in the PTX group. In accordance, serum leptin levels were also decreased in the treated mice by 29% in comparison with the fatty control (in ng/mL: 2.97 ± 0.19 vs. 4.17 ± 0.41, P<0.05) [30]. In terms of glucose and insulin impairments, treatment with pentoxifylline led to a reduction in the plasma levels of glucose (in mg/dL: 190.0 ± 6.3 in high fat control vs. 152.8 ± 4.9 PTX group, P<0.05) and insulin (in ng/mL: 1.00 ± 0.13 fatty control vs. 0.67 ± 0.09 PTX group, P<0.05), attenuated glucose intolerance and improved insulin sensitivity, as shown by the OGTT and ITT and increased levels of insulin-stimulated protein kinase B (AKT) phosphorylation in eWAT, sWAT and iBAT (32%, 37%, 108%). These effects were accompanied by a reduction in the plasma levels of fluorescent AGEs (in AU/ mg protein: 83.5 ± 4.1 in fatty control vs 59.9 ± 6.9 PTX group, P<0.05, and 56.6 ± 5.2 in lean control) [30], with values of the PTX group returning to similar levels of the lean control group mice. Conversely, PTX treatment did not decrease the serum levels of TNF-α (in pg/mL: 9.61 ± 0.45 fatty control vs 9.89 ± 1.04 PTX) or IL-6 (in pg/mL: 10.28 ± 0.82 fatty control vs 11.84 ± 1.53 PTX), but still reduced serum levels of MCP-1 (in pg/mL: 37.74 ± 5.11 fatty control vs 19.53 ± 5.53, P<0.05) [30].
3.7. RAGE229
N(4-(7-cyano-4-(morpholin-4ylmethyl)quinolin-2-yl)phenyl)acetamide, RAGE229, is a chemical probe that antagonizes RAGE signaling and reduces inflammation and diabetic complications in mice. Wilson R. et al. [33] tested the hypothesis that pharmacological antagonism of RAGE signaling with this small molecule improves metabolic health in lean mice and mice with obesity undergoing diet-induced weight loss. They found in HFD-induced obese mice undergoing diet induced weight loss, administration of a rage229 in 150ppm dose diet improved thermogenesis as shown by a lesser decline in body temperature compared to control group. RAGE229 also inflicted an increase in lost body mass and significantly lower white adipose tissue (WAT) mass (in epididymal WAT and inguinal WAT), with no significant differences in lean mass. It also showed important metabolic effects, as mice on the RAGE229 diet showed better insulin tolerance, higher AKT phosphorylation in liver and muscle tissues, and lower fasting insulin levels and HOMA-IR index [33]. They also reported significantly improved glucose clearance and better insulin signaling post- IP. As for lipid metabolism, plasma cholesterol concentrations were significantly lower and, although plasma triglyceride concentrations did not differ, hepatic triglyceride concentrations were significantly reduced. Treatment with RAGE229 diet also showed adipocyte remodeling capacities, since treated mice had smaller adipocytes in epididymal WAT (which corresponds to visceral adipose tissue), a higher proportion of smaller adipocytes compared to larger ones and significantly lower leptin/fat mass, and significantly higher adiponectin/fat mass and adiponectin/leptin concentration ratio. Moreover, the RAGE inhibitor showed significantly lower TNF-α mRNA expression in epididymal WAT. All of these effects appear to be dose-dependent, since a lower dose (50 ppm) of RAGE229 did not significantly affect body mass, adiposity, glucose tolerance, or insulin sensitivity in mice with obesity [33].
3.8. Pyrogallol-Phloroglucinol-6,6-Bieckol
Pyrogallol-Phloroglucinol-6,6-Bieckol(PPB) is an antioxidant phlorotannin isolated from the Ecklonia cava seaweed[42]. Choi J. et al. investigated the effects of oral administration of PPB (2 mg/kg/day for 4 weeks) in a diet- induced obesity mouse model (C57BL/6N mice fed a 45% high-fat diet). They found that PPB treatment led to a reduction in body weight and the size of adipocytes of visceral adipose tissues. Additionally, PPB also lowered serum levels of triglycerides and total cholesterol [27], although the authors attribute this effect partially to the decrease in fat-mass as well. In terms of adipose tissue inflammation, PPB showed modulation of macrophage markers, significantly reducing the intensity of CD86 signal after staining visceral adipose tissue samples of mice with obesity and, on the other hand, increasing the intensity of CD163, when compared with the fatty untreated control. The relative mRNA expression levels of these macrophage markers showed equivalent changes. In concordance, PPB supplementation led to a decrease in secretion of inflammatory cytokines TNF-α and IL-1β in visceral adipose tissue, exhibited by the decrease in relative TNF-α and IL-1β mRNA expression levels in the visceral fat of the PPB group. Moreover, and accordingly with the effects on the metabolic parameters, PPB showed a reduction in the levels of AGEs, HMGB1, and S100β in visceral fat and significantly reduced RAGE expression, shown by the reduced RAGE signal intensity in immunofluorescence staining of visceral adipose tissue of PPB group and by the diminished AGEs, HMGB1 and S100β- RAGE binding ratios [27].
3.9. Genistein
Dietary genistein is an abundant isoflavone in soybean and other plant-derived foods, that can prevent the formation of AGEs by trapping MGO and regulating MGO metabolism [43]. Zhao Y. et al. [29] demonstrated that genistein treatment in mice with obesity significantly ameliorated very high fat (VHF) diet –induced and high fat diet- exogenous MGO (HFM) induced indicators of metabolic syndrome by significantly inhibiting the increases in body weight (in g: 23.03 ± 2.94 VHF vs 25.58 ± 1.47 HFM vs 6.3 ± 2.48 in HFM+ 0.067% genistein treatment), fasting blood glucose concentration, organ weight, plasma cholesterol, ALT, and AST concentrations, liver TG concentration, and fatty liver. Furthermore, genistein dramatically lowered plasma MGO and AGE concentrations in comparison with VHF diet mice, and wih the healthy control (low-fat (LF) diet mice). The expressions of GLO 1 and GLO 2, important MGO detoxification systems, were upregulated in the VHF diet plus genistein group to either a similar level or a significantly higher level than those in the LF group. In the liver, dietary genistein inhibited the formation of AGEs, but also blocked the activation of the RAGE pathway to prevent liver damage induced by VHF diet[29].
3.10. Carnosic Acid
Carnosic acid (CA) is an active rosemary extract (RE) component. Zhao Y. et al. [21] studied the effects of CA on HF -diet-induced obesity and metabolic syndrome in mice. They found that RE supplementation significantly reduced body weight gain (), total fat mass, and percent body fat in HF diet-fed mice. This effect was dose-dependent, with RE with higher CA content showing greater efficacy. They also noted improved markers of metabolic syndrome including fasting blood glucose levels, insulin resistance (measured by HOMA-IR index), and lipid profiles in the liver (triglycerides and free fatty acids), with the added effect of reducing liver weight and ALT and AST levels. Oxidative stress and inflammation were also decreased with RE supplementation, with diminished MDA and TNF-α levels. All of these changes were accompanied by an inhibition of AGE accumulation in plasma and liver and a reduced expression of RAGE.
3.11. Indian Gooseberry Extract
Indian gooseberry (Phyllanthus emblica L.) is a traditional medicinal used against common cold, fever, diabetes, dyspepsia, peptic ulcer, inflammation, and skin diseases. It contains abundant bioactive components, such as vitamin C, gallic acid (GA), ellagic acid (EA). Several pharmacological studies have elucidated that the extract and its active compounds display antioxidative, anti-inflammatory, hepatoprotective, hypolipidemic, and hypoglycemic abilities[44]. A recent study revealed that water extract of Indian gooseberry fruit (WEIG) and its phenolic compounds (GA and EA) prevent MGO glycation-induced leptin resistance [45]. Chen S- et al. investigated WEIG supplementation and its bioactive compound GA in preventing leptin resistance in peripheral tissues by regulating MGO, inflammation, and oxidative stress in HFD-induced obesity rats [34]. They found that administration of WEIG and GA significantly reduced the body weight in rats subjected to HFD. WEIG and GA treatment also effectively lowered MGO levels in peripheral tissues, increased Glo-1 activity in liver and perinephric tissues and significantly reduced AGE activity in the liver, kidney and perinephric fat tissue. Furthermore, WEIG and GA supplementation significantly inhibited leptin expression in the serum and liver of HFD-induced rats and promote the increase in adiponectin content in the liver and perirenal fat. However, serum and renal adiponectin levels were not significantly changed. Supplementation with WEIG reduced oxidative stress and inflammation, as demonstrated by a reduced MDA expression and ameliorated activity of SOD and GPx, and reduced secretion of IL-6, IL-1b, TNF-a in liver and kidney tissues.
3.12. Epigallocatechin-3-gallate (EGCG)
The most abundant bioactive catechins present in green tea is epigallocatechin-3-gallate (EGCG), which has shown positive results for its anti-carcinogenic, antioxidant, radical scavenging and anti-inflammatory properties. Clinical studies have reported that consumption of green tea extracts (rich in EGCG) reduces blood glucose levels in obese and diabetic subjects. So, Sampath C. et al [25] hypothesized that EGCG would mitigate AGEs formation via activating the nuclear factor erythroid-2-related-factor-2 (Nrf2) pathway in high fat diet-induced obese mice. They showed that administration of EGCG at 25 mg/kg had positive effects on body weight (in g; 675.32±39.73 obesity control vs 569.71±31.25 EGCG; P<0,05) and size of organs (Liver wet weight: 14.89±0.82 obesity control vs 11.46±0.66; P<0,05), as it significantly slowed the weight gain observed in mice on HFD. Moreover, a higher dose of EGCG (75 mg/kg) showed almost 100% less weight gain. The apparent dose-dependent effects translated to other metabolic effects, as the higher dose of EGCG exhibited significant reduction in fasting blood glucose levels, but administration of EGCG at 25 mg/kg showed only a slight blood glucose lowering effect. EGCG at 75 mg/kg brought insulin levels to nearly identical levels as the control group (mice on low-fat diet) and EGCG decreased the HOMA-IR index of mice on HFD by up to 89.2%. The green tea bioactive compound also ameliorated oxidative stress and inflammation, as it affected HFD-induced dicarbonyl stress, AGEs accumulation and the expression of RAGE, Nrf2, and HO-1. Supplementation with EGCG at 75 mg/kg significantly increased the GSH levels (exhibits an essential role in detoxification in the tissues exposed to dicarbonyl stress) in the liver and adipose tissue indicating a beneficial effect on oxidative stress. It also significantly inhibited AGEs accumulation in plasma and adipose tissue, inhibiting AGEs by up to 91% in the latter. Finally, the data show that RAGE expression significantly reduced or inhibited by high-dose EGCG, which reduced the RAGE expression by about 3 folds from the levels observed in HFD control.
4. Discussion
The rising worldwide incidence of obesity significantly increases both its healthcare and financial impact [2]. Current treatments for obesity consist of lifestyle changes, primarily in diet and nutrition, and anti-obesogenic drugs, but the latter can result in several side effects with long term administration [46]. Therefore, it is important to continue research to find better alternatives, that are equally or more effective and with less side effects, thus improving and innovating obesity care. This review summarizes the state of the art on potential beneficial effects of supplementation with molecules that interfere in the AGE-RAGE axis, which has been implicated in central mechanisms underlying the pathogenesis of adipose tissue dysfunction and other metabolic disorders associated with obesity [4,8,47].
Overall, reviewed substances showed promising therapeutic effects in treating obesity and obesity- associated metabolic disorders. In rodents models of obesity, these molecules exhibited a significant role in reducing body weight, improving the lipid profile, lowering the levels of triglycerides, cholesterol and FFAs in the serum and liver. They also improved glucose impairment, insulin sensitivity and even ameliorated the inflammatory profile of adipose tissue, reducing the expression of Il-6, IL-1b, TNF-a and leptin and increasing the secretion of adiponectin. All of these effects are correlated with the inhibition of AGE accumulation and RAGE expression in adipose tissue and systemically, which is in line with a previous study that shows the same improvement in metabolic markers through a reduction in dietary AGEs [10]. However, the few human trials included in this review showed controversial outcomes, in comparison with the animal studies, with only one of three studies showing significant improvements in fasting glucose and insulin, insulin resistance and secretion, and decreasing fat mass in people with obesity. These differences between animal and human study outcomes might be because the sample sizes in the human trials are small and do not have the same characteristics as the rodent models. While rodent obesity models have concomitant metabolic disorders, like dyslipidemia, insulin resistance, glucose intolerance and hypertension, or diseases, most commonly type 2 diabetes, Van den Eynde M. et al human study only included non-diabetic individuals and excluded those with metabolic or vascular diseases [32]. Furthermore, the dosage and duration of supplementation used in humans is lower than that used in animal models, which could affect the efficacy of these molecules, since the reviewed animal studies showed a dose-dependent effect for most substances. Additionally, all reviewed studies focused on evaluating disease-oriented parameters of obesity. In future research, it would be interesting to understand if these beneficial impacts on clinical parameters also translate into an improvement in patient-oriented outcomes.
In conclusion, this review demonstrates that these AGE-RAGE inhibitors may serve as possible new treatments for obesity and obesity-related metabolic disorders. They showed a significant role in improving adipose tissue inflammation and dysfunction, not only through the reduction of oxidative stress (by directly interfering with AGE and RAGE expression), but also by regulating cytokine secretion and aiding in weight loss [48]. Furthermore, a lot of these molecules are phytochemicals, which have had an increasing interest in various research fields due to their safety and fewer side effects. There is a need to perform more clinical trials with larger samples to further establish their effects on humans, and to define ideal doses for treatment.
Acknowledgments
An immense deal of gratitude goes to Baltazar Oliveira for aiding in designing the search queries and proofreading this review.
Author Contributions
Nádia Oliveira was involved and contributed to all steps of this review and was the main author. Manuel Nuno Alçada contributed as a supervisor, helping with the study design and providing corrections to the paper.
Competing Interests
The authors declare there are no competing interests. This is an independent review.
List of Abbreviations
AGE – advanced glycation end products
AKT – protein kinase B
ALE- advanced lipoxidation end products
CA – carnosic acid
D-CAR – D-carnosine
DIO – diet-induced obesity
EGCG – epigallocatechin-3-gallate
eWAT – epididimal white adipose tissue
FFA – free fatty acid
GA - gallic acid
GHS – glutathione
GLO 1/2 – glyoxalase 1/2
GPx- glutathione peroxidase
HFD – high-fat diet
HFHS – high-fat/high-sucrose diet
HNE – 4-Hydroxynonenal
HOMA – IR – Homeostatic Model Assessment for Insulin Resistance
HWEIG – High - water extract of Indian gooseberry fruit
iBAT- interscapular brown adipose tissue
IRS- 1 – insulin receptor substrate 1
ITT- Insulin tolerance test
L-CAR - L-Carnosine
LWEIG – low- water extract of Indian gooseberry fruit
MCP-1 – monocyte chemoattractant protein-1
MDA – malondialdehyde
MGO – methylglyoxal
NAT - nateglinide
OGTT – oral glucose tolerance test
PM- pyridoxamine
PPB – Pyrogallol-Phloroglucinol-6,6-Bieckol
PTX – pentoxifylline
R- ALA – α-lipoic acid
RAGE- receptor for advanced glycation end products
RCS- reactive carbonyl species
RE – rosemary extract
ROS- reactive oxygen species
sICAM-1 – soluble intercellular adhesion molecule-1
SOD – superoxide dismutase
sVCAM-1 – soluble vascular cell adhesion molecule 1
sWAT- subcutaneous white adipose tissue
TEL - telmisartan
WEIG – water extract of Indian gooseberry fruit
References
- Hotamisligil KEWaGS. Obesity-induced inflammatory changes in adipose tissue. J Clin Invest. 2003;112(12):4. [CrossRef]
- Federation WO. World Obesity Atlas 2023. https://data.worldobesity.org/publications/?cat=19: World Obesity Federation; 2023 march.
- Organization WH. Obesity https://www.who.int/health-topics/obesity#tab=tab_12024 [.
- Gutowska K, Czajkowski K, Kurylowicz A. Receptor for the Advanced Glycation End Products (RAGE) Pathway in Adipose Tissue Metabolism. Int J Mol Sci. 2023;24(13). [CrossRef]
- Shen CY, Lu CH, Wu CH, Li KJ, Kuo YM, Hsieh SC, et al. The Development of Maillard Reaction, and Advanced Glycation End Product (AGE)-Receptor for AGE (RAGE) Signaling Inhibitors as Novel Therapeutic Strategies for Patients with AGE-Related Diseases. Molecules. 2020;25(23). [CrossRef]
- Jangde N, Ray R, Rai V. RAGE and its ligands: from pathogenesis to therapeutics. Crit Rev Biochem Mol Biol. 2020;55(6):555-75. [CrossRef]
- Ahmad S, Khan H, Siddiqui Z, Khan MY, Rehman S, Shahab U, et al. AGEs, RAGEs and s-RAGE; friend or foe for cancer. Semin Cancer Biol. 2018;49:44-55. [CrossRef]
- C S-B, NA B, CG F, M K, CK N, D P, et al. Advanced glycation end-products regulate extracellular matrix-adipocyte metabolic crosstalk in diabetes. England2019 2019-12-24. 19748 p.
- Z F, L Z, J W. RAGE signalling in obesity and diabetes: focus on the adipose tissue macrophage. Adipocyte. 2020;9(1):563-6. [CrossRef]
- PVM R, JF T, MAC C, JB M, RCG A. Effect of reducing dietary advanced glycation end products on obesity-associated complications: a systematic review. Nutrition reviews. 2019;77(10):725-34. [CrossRef]
- SK M, N V, SK M. Advanced glycation end products induce lipogenesis: regulation by natural xanthone through inhibition of ERK and NF-κB. Journal of cellular physiology. 2014;229(12):1972-80. [CrossRef]
- Villegas-Rodriguez ME, Uribarri J, Solorio-Meza SE, Fajardo-Araujo ME, Cai W, Torres-Graciano S, et al. The AGE-RAGE Axis and Its Relationship to Markers of Cardiovascular Disease in Newly Diagnosed Diabetic Patients. PLoS One. 2016;11(7):e0159175. [CrossRef]
- Vijaykrishnaraj M, Wang K. Dietary natural products as a potential inhibitor towards advanced glycation end products and hyperglycemic complications: A phytotherapy approaches. Biomedicine and Pharmacotherapy. 2021;144. [CrossRef]
- Alderson N, Chachich M, Youssef N, Beattie R, Nachtigal M, Thorpe S, et al. The AGE inhibitor pyridoxamine inhibits lipemia and development of renal and vascular disease in Zucker obese rats. KIDNEY INTERNATIONAL. 2003;63(6):2123-33. [CrossRef]
- EA M, CJ D, MK T, KA L, TL A, NB H, et al. Interactions of the advanced glycation end product inhibitor pyridoxamine and the antioxidant alpha-lipoic acid on insulin resistance in the obese Zucker rat. Metabolism: clinical and experimental. 2008;57(10):1465-72. [CrossRef]
- EM M, CJ D, MK T, KA L, O H, M M, et al. Metabolic interactions of AGE inhibitor pyridoxamine and antioxidant alpha-lipoic acid following 22 weeks of treatment in obese Zucker rats. Life sciences. 2009;84(15):563-8. [CrossRef]
- S H, T G, M T, T I, M M, I O, et al. Effects of pyridoxamine (K-163) on glucose intolerance and obesity in high-fat diet C57BL/6J mice. Metabolism: clinical and experimental. 2009;58(7):934-45. [CrossRef]
- H U-K, S Y, M T, H B, Y S. Pyridoxamine, an inhibitor of advanced glycation end product (AGE) formation ameliorates insulin resistance in obese, type 2 diabetic mice. Protein and peptide letters. 2010;17(9):1177-81. [CrossRef]
- Miura K, Kitahara Y, Kajioka T, Takeuchi M, Yamagishi S. Combination therapy with nateglinide and telmisartan ameliorates insulin resistance in zucker Fatty rats by suppressing advanced glycation end product receptor axis. Horm Metab Res. 2011;43(3):226-8. [CrossRef]
- G A, M O, G R, F S, P B, G V, et al. The carbonyl scavenger carnosine ameliorates dyslipidaemia and renal function in Zucker obese rats. Journal of cellular and molecular medicine. 2011;15(6):1339-54. [CrossRef]
- Y Z, R S, P W, H C, Y Z, S S. Carnosic acid as a major bioactive component in rosemary extract ameliorates high-fat-diet-induced obesity and metabolic syndrome in mice. Journal of agricultural and food chemistry. 2015;63(19):4843-52. [CrossRef]
- Baye E, Ukropcova B, Ukropec J, Hipkiss A, Aldini G, de Courten B. Physiological and therapeutic effects of carnosine on cardiometabolic risk and disease. AMINO ACIDS. 2016;48(5):1131-49. [CrossRef]
- Maessen DE, Brouwers O, Gaens KH, Wouters K, Cleutjens JP, Janssen BJ, et al. Delayed Intervention With Pyridoxamine Improves Metabolic Function and Prevents Adipose Tissue Inflammation and Insulin Resistance in High-Fat Diet-Induced Obese Mice. DIABETES. 2016;65(4):956-66. [CrossRef]
- DD X, M Z, N L, JF G, L M, M L. Mediation of inflammation, obesity and fatty liver disease by advanced glycation endoproducts. European review for medical and pharmacological sciences. 2017;21(22):5172-8. [CrossRef]
- Sampath C, Rashid MR, Sang S, Ahmedna M. Green tea epigallocatechin 3-gallate alleviates hyperglycemia and reduces advanced glycation end products via nrf2 pathway in mice with high fat diet-induced obesity. Biomed Pharmacother. 2017;87:73-81. [CrossRef]
- Anderson EJ, Vistoli G, Katunga LA, Funai K, Regazzoni L, Blake Monroe T, et al. A carnosine analog mitigates metabolic disorders of obesity by reducing carbonyl stress. Journal of Clinical Investigation. 2018;128(12):5280-93. [CrossRef]
- J C, S O, M S, K B. Pyrogallol-Phloroglucinol-6,6-Bieckol Alleviates Obesity and Systemic Inflammation in a Mouse Model by Reducing Expression of RAGE and RAGE Ligands. Marine drugs. 2019;17(11). [CrossRef]
- S O, H A, H P, JI L, KY P, D H, et al. The attenuating effects of pyridoxamine on adipocyte hypertrophy and inflammation differ by adipocyte location. The Journal of nutritional biochemistry. 2019;72:108173. [CrossRef]
- Y Z, P W, S S. Dietary Genistein Inhibits Methylglyoxal-Induced Advanced Glycation End Product Formation in Mice Fed a High-Fat Diet. The Journal of nutrition. 2019;149(5):776-87. [CrossRef]
- Inacio MD, Costa MC, Oliveira Lima TF, Figueiredo ID, Motta BP, Spolidorio LC, et al. Pentoxifylline mitigates renal glycoxidative stress in obese mice by inhibiting AGE/RAGE signaling and increasing glyoxalase levels. LIFE SCIENCES. 2020;258. [CrossRef]
- W P, P M, L L, K C, Y Z, W X, et al. Effect of carnosine supplementation on lipid profile, fasting blood glucose, HbA1C and insulin resistance: A systematic review and meta-analysis of long-term randomized controlled trials. Complementary therapies in medicine. 2020;48:102241. [CrossRef]
- MDG VdE, AJHM H, JLJM S, AMA L, PM N, N S, et al. Pyridoxamine reduces methylglyoxal and markers of glycation and endothelial dysfunction, but does not improve insulin sensitivity or vascular function in abdominally obese individuals: A randomized double-blind placebo-controlled trial. Diabetes, obesity & metabolism. 2023;25(5):1280-91. [CrossRef]
- RA W, L A, HH R, B Z, K Q, MB M, et al. Pharmacological antagonism of receptor for advanced glycation end products signaling promotes thermogenesis, healthful body mass and composition, and metabolism in mice. Obesity (Silver Spring, Md). 2023;31(7):1825-43. [CrossRef]
- Chen S-Y, Huang Y-N, Lin J-A, Yen G-C. Effect of Indian gooseberry extract on improving methylglyoxal-associated leptin resistance in peripheral tissues of high-fat diet-fed rats. JOURNAL OF FOOD AND DRUG ANALYSIS. 2024;32(1). [CrossRef]
- Metz TO, Alderson NL, Chachich ME, Thorpe SR, Baynes JW. Pyridoxamine Traps Intermediates in Lipid Peroxidation Reactions in Vivo: EVIDENCE ON THE ROLE OF LIPIDS IN CHEMICAL MODIFICATION OF PROTEIN AND DEVELOPMENT OF DIABETIC COMPLICATIONS*. Journal of Biological Chemistry. 2003;278(43):42012-9. [CrossRef]
- Sabliov CM, Astete CE. 12 - Encapsulation and controlled release of antioxidants and vitamins. In: Garti N, editor. Delivery and Controlled Release of Bioactives in Foods and Nutraceuticals: Woodhead Publishing; 2008. p. 297-330. [CrossRef]
- Thornalley PJ. Use of aminoguanidine (Pimagedine) to prevent the formation of advanced glycation endproducts. Archives of Biochemistry and Biophysics. 2003;419(1):31-40. [CrossRef]
- Freedman BI, Wuerth J-P, Cartwright K, Bain RP, Dippe S, Hershon K, et al. Design and Baseline Characteristics for the Aminoguanidine Clinical Trial in Overt Type 2 Diabetic Nephropathy (ACTION II). Controlled Clinical Trials. 1999;20(5):493-510. [CrossRef]
- Bolton WK, Cattran DC, Williams ME, Adler SG, Appel GB, Cartwright K, et al. Randomized trial of an inhibitor of formation of advanced glycation end products in diabetic nephropathy. Am J Nephrol. 2004;24(1):32-40. [CrossRef]
- Kajioka T, Miura K, Kitahara Y, Yamagishi S. Potential utility of combination therapy with nateglinide and telmisartan for metabolic derangements in Zucker Fatty rats. Horm Metab Res. 2007;39(12):889-93. [CrossRef]
- Langlais P, Yi Z, Finlayson J, Luo M, Mapes R, De Filippis E, et al. Global IRS-1 phosphorylation analysis in insulin resistance. Diabetologia. 2011;54(11):2878-89. [CrossRef]
- Lee JH, Ko JY, Oh JY, Kim CY, Lee HJ, Kim J, et al. Preparative isolation and purification of phlorotannins from Ecklonia cava using centrifugal partition chromatography by one-step. Food Chem. 2014;158:433-7. [CrossRef]
- Lo CY, Li S, Tan D, Pan MH, Sang S, Ho CT. Trapping reactions of reactive carbonyl species with tea polyphenols in simulated physiological conditions. Mol Nutr Food Res. 2006;50(12):1118-28. [CrossRef]
- Saini R, Sharma N, Oladeji OS, Sourirajan A, Dev K, Zengin G, et al. Traditional uses, bioactive composition, pharmacology, and toxicology of Phyllanthus emblica fruits: A comprehensive review. J Ethnopharmacol. 2022;282:114570. [CrossRef]
- Huang Y-N, Chen S-Y, Lin J-A, Chiang IC, Yen G-C. Phyllanthus emblica L. extract alleviates leptin resistance and lipid accumulation by inhibiting methylglyoxal production. Food Bioscience. 2023;53:102619. [CrossRef]
- Aruwa CE, Sabiu S. Adipose tissue inflammation linked to obesity: A review of current understanding, therapies and relevance of phyto-therapeutics. Heliyon. 2024;10(1):e23114. [CrossRef]
- Wu Y, Zong M, Wu H, He D, Li L, Zhang X, et al. Dietary Advanced Glycation End-Products Affects the Progression of Early Diabetes by Intervening in Carbohydrate and Lipid Metabolism. Mol Nutr Food Res. 2022;66(11):e2200046. [CrossRef]
- Goran MI, Alderete TL. Targeting Adipose Tissue Inflammation to Treat the Underlying Basis of the Metabolic Complications of Obesity. In: Drewnowski A, Rolls BJ, editors. Obesity Treatment and Prevention: New Directions: 73rd Nestlé Nutrition Institute Workshop, Carlsbad, Calif, September 2011. 73: S.Karger AG; 2012. p. 0.
Figure 1.
PRISMA Flowchart for study selection.
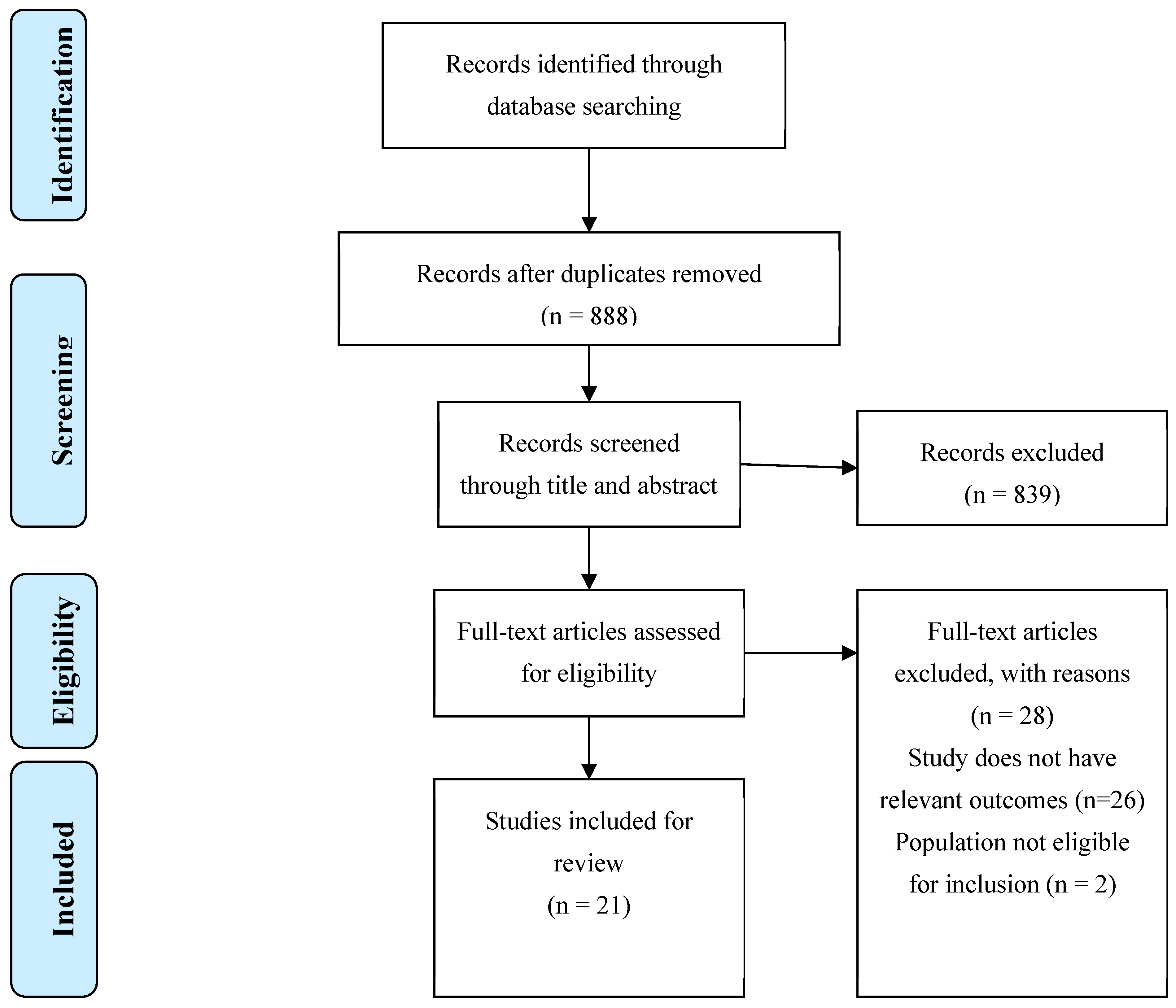

Table 1.
Search strategies used in electronic databases.
| Electronic Database | Search Date | Search Strategy | Nº of results |
| MEDLINE(PubMed) | 23/05/2024 | ((obese OR obesity[MeSH Terms])) AND ((advanced glycation end products[MeSH Terms]) OR (Receptor for Advanced Glycation End Products[MeSH Terms]) OR (RAGE) OR (sRAGE)) AND ((inhibitor) OR (inhibition) OR (antagonist) OR (small molecule) OR (liraglutide) OR (pyridoxamine) OR (azeliragon) OR (S100-derived peptide) OR (HMGB1-derived Peptide) OR (alagebrium) OR (carnosine) OR (FPS-ZM1) OR (GM-1111) OR (semaglutide) OR (GLP1 analog)) AND ((adipose tissue metabolism) OR (adipose tissue dysfunction) OR (metabolic inflammation) OR (insulin resistance)) | 245 |
| SCOPUS | 23/05/2024 | ALL ( obese OR obesity ) AND ALL ( "advanced glycation end products" OR "Receptor for Advanced Glycation End Products" OR "RAGE" OR "sRAGE" ) AND ALL ( inhibitor OR inhibition OR antagonist OR "small molecule" OR liraglutide OR pyridoxamine OR azeliragon OR "S100-derived peptide" OR "HMGB1-derived Peptide" OR "alagebrium" OR "carnosine" OR "FPS-ZM1" OR "GM-1111" OR "semaglutide" OR glp1 AND analog ) AND TITLE-ABS-KEY ( "adipose tissue metabolism" OR "adipose tissue dysfunction" OR "metabolic inflammation" OR "insulin resistance") | 588 |
| Web of Science | 23/05/2024 | ALL=(Obese OR Obesity) AND ALL=((advanced glycation end products) OR (Receptor for Advanced Glycation End Products) OR (RAGE) OR (sRAGE)) AND ALL=((inhibitor) OR (inhibition) OR (antagonist) OR (small molecule) OR (liraglutide) OR (pyridoxamine) OR (azeliragon) OR (S100-derived peptide) OR (HMGB1-derived Peptide) OR (alagebrium) OR (carnosine) OR (FPS-ZM1) OR (GM-1111) OR (semaglutide) OR (GLP1 analog)) AND ALL=((adipose tissue metabolism) OR (adipose tissue dysfunction) OR (metabolic inflammation) OR (insulin resistance)) | 104 |
Table 2.
Characteristics of included studies.
| Authors | Year | Study type | Methodology | Key findings |
| Alderson N. et al.[14] | 2003 | Animal study | Three groups of Zucker rats were studied: lean, untreated fatty and fa/fa treated with PM (2 g/Ldrinking water) for 32 weeks. | PM inhibited the increases in AGE/ALEs in collagen, and significantly decreased the rise in plasma triglycerides, cholesterol, and creatinine, corrected hypertension and thickening of the vascular wall in Zucker fa/fa rats. |
| Muellenbach E. et al. [15] |
2008 | Animal study | Obese Zucker rats were assigned to either a control group or to a treatment group receiving daily injections of the R-ALA (92 mg/kg) or PM (60 mg/kg), individually or in combination, for 6 weeks. | Individual and combined treatments with R-ALA and PM significantly (P < .05) reduced markers of oxidative stress. Combination treatment resulted in the largest reductions of fasting plasma glucose (23%), insulin (16%), and free fatty acids (24%) and of muscle triglycerides (45%). It also elicited the greatest enhancement of whole-body insulin sensitivity. |
| Muellenbach E. et al.[16] |
2009 | Animal study | Female Obese Zucker rats received vehicle (OV), PM (OP, 60 mg/kg body wt), racemic ALA (rac-ALA; OM, 92 mg/kg), the R- (+)-enantiomer of ALA (R-ALA; OR, 92 mg/kg), or combined treatments with PM and rac-ALA (OPM) or PM and R-ALA (OPR), daily for 22 weeks. | Individual and combined treatments with PM, rac-LA, and R-LA significantly inhibited markers of oxidative damage, and triglyceride levels, and plasma free fatty acids, with the greatest decrease (26%) elicited in OPR. Insulin resistance was lowered (20%) only in combined treatment. |
| Hagiwara S. et al. [17] | 2009 | Animal study | C57BL/6J mice were divided into 3 groups: low-fat diet, high-fat diet, and high-fat diet with pyridoxamine treatment, for 12 weeks | Body and adipose tissue weights of PM treatment group were diminished. PM also diminished serum AGE; increased antioxidant enzyme expression; and improved dysregulation of adipocytokines in adipose tissues. PM treatment improved blood glucose levels and fasting hyperinsulinemia. |
| Unoki-Kubota H. et al. [18] |
2010 | Animal study | Of 37 KK-Ay/Ta Jcl mice, 21 aged at 5 weeks were followed up to 15 weeks of age, and body weight and food consumption were monitored during the study periods. The rest of 5-week-old KK- Ay mice were divided randomly into four groups; untreated control group, 24, 120, 240 mg/l PM treatment, and followed for 5 more weeks. | PM treatment dose-dependently decreased fasting insulin levels and improved insulin sensitivity in KK-Ay mice of 10 weeks old, but it did not affect fasting blood glucose levels. |
| Miura K. et al. [19] | 2011 | Animal study | Male Zucker fatty rats were divided into 4 groups (n = 6 each): vehicle (VEH), 50 mg / kg of NAT, 5 mg / kg of TEL, or both (NAT / TEL). Treatment was administered for 6 weeks. |
Combination therapy with NAT and TEL, but not each monotherapy, inhibited IRS-1 serine phosphorylations at 307 and 636 / 639 residues and restored the decrease in IRS-1 tyrosine phosphorylation in the liver. It also reduced levels of AGEs, hepatic RAGE expression and hepatic AGE-RAGE index. |
| Aldini G. et al[20]. | 2011 | Animal study | Zucker fa/fa rats with obesity were randomly divided into three groups of 12 each: untreated control with obesity, L-CAR treated and D-CAR- treated. D-CAR and L-CAR were administered to fa/fa rats in the drinking water (24 hr drug dose of 30 mg/kg) for 24 weeks. | L-CAR and D-CAR restrained the development of dyslipidaemia, hypertension and renal injury. Body weight was reduced. L-CAR and D- CAR-fed rats, after 24 weeks, had plasma cholesterol and triglycerides levels reduced, but plasma glucose levels weren’t affeced. L- CAR and D-CAR restrained the development of hyperinsulinemia and improved insulin resistance. |
| Zhao Y. et al.[21] | 2015 | Animal study | Male C57BL/6J mice were given a low-fat diet, a high-fat diet or a high-fat diet supplemented with either 0.14% CA- enriched rosemary extract or 0.28% CA-enriched RE or 0.5% of commercial RE, for a period of 16 weeks. | RE supplementation significantly reduced body weight gain, percent of fat, plasma ALT, AST, glucose, insulin levels, liver weight, liver triglyceride, and free fatty acid levels It also decreased the levels of plasma and liver AGEs, and the liver expression of RAGE. |
| Baye E. et al. [22] | 2016 | Narrative review | Not disclosed | Carnosine supplementation reduced fasting insulin, decreased insulin resistance and reduced insulin secretion in healthy humans with obesity or overweight. |
| Maessen D. et al. [23] | 2016 | Animal study | Male C57BL/6J 12-week-old mice were divided into three groups. The low- fat diet group, and 2 high fat diet groups. After 6 weeks of HFD, one group started to receive PM (2 g/L) in the drinking water (HFD + PM) for 18 weeks. Male db/db mice were included in the study at an age of 6 weeks. They were also treated with PM in their drinking water for 18 weeks. | Delayed intervention with PM protected against HFD-induced body weight gain, hyperglycemia and hypercholesterolemia. PM also inhibited impaired glucose and insulin resistance in HFD-induced and db/db obese mice. PM prevented expansion of adipose tissue, adipocyte hypertrophy and attenuated expression of proinflammatory genes in visceral adipose tissue. |
| Xiong D-D. et al. [24] | 2017 | Animal study | Sprague Dawley (SD) rats were randomly divided into control, model and AGEs inhibitor groups. Obesity fatty liver model was prepared by the application of high-fat diet and subcutaneous injection of CCl4 at 0.2 mg/100 g for six weeks. Aminoguanidine (100 mg/kg·d) was given by subcutaneous injection for 6 weeks. | Administration of aminoguanidine significantly improved liver functions, improved the metabolism of fatty acids and lowered TNF-α or IL-6 levels. |
| Sampath C. et al. [25] | 2017 | Animal study | Dietary EGCG was tested in C57BL/6 mice placed on a high-fat diet with or without ECGC for 17 weeks, compared to a control group placed on low-fat diet for the same period. | Dietary EGCG significantly reduced weight gain, plasma glucose, insulin level, liver and kidney weight. EGCG also decreased levels of AGEs in plasma and liver, and inhibited RAGE expression. |
| Anderson E. et al. [26] | 2018 | Animal study | C57BL6/J female mice were crossed with male GPx4+/– mice. At 8 to 12 weeks of age, WT and GPx4+/– male age-matched littermates were randomly assigned to groups. Mice were fed either a control or a HFHS diet for 25 weeks. After 8 weeks of the HFHS diet, half of the mice in the HFHS diet were administered carnosinol (45 mg/kg/day) until study termination at 20 weeks. | In models of diet-induced obesity and metabolic syndrome, carnosinol dose-dependently reduced HNE adduct formation in liver and skeletal muscle, and mitigated inflammation, dyslipidemia, insulin resistance, and steatohepatitis. |
| Choi J. et al. [27] | 2019 | Animal study | C57BL/6N male mice were fed a 45% high-fat diet for 8 weeks. Mice were separated into three groups: control, DIO/saline, DIO/PPB. Isolated PPB was dissolved in 0.9% saline and each group was orally administrated saline and PPB (2.5 mg/kg) daily for 4 weeks. | In visceral fat, PPB significantly inhibited RAGE ligands, reduced the RAGE expression, and reduced binding ratio between RAGE and RAGE ligands. PPB reduced differentiation of macrophages in visceral fat into M1-type and related pro-inflammatory cytokines. |
| Oh S. et al. [28] | 2019 | Animal study | Sprague-Dawley rats were divided into three groups and fed a 45% high-fat diet or a normal diet for 8 weeks. Rats in the pyridoxamine treated group were fed a HFD for 4 weeks and then pyridoxamine (2 mg/day dissolved in 1 ml of saline) for another 4 weeks with feeding a 45% high-fat diet. | Pyridoxamine reduced HFD-induced weight gain, adipocyte size, RAGE ligand accumulations, AGE-RAGE ligands binding, decreased macrophage M1 polarization and increased M2 polarization in visceral fat tissues, but not in subcutaneous tissues. PM induced Glo-1 expression in visceral fat in the HFD group. |
| Zhao Y. et al[29]. | 2019 | Animal study | Male C57BL/6J mice (n = 15) were fed a LF diet or a VHF diet alone or including 0.25% genistein for 16 weeks in study 1. In study 2, 75 similar mice were fed the LF diet or the HF diet or in combination with up to 0.2% MGO in water (HFM) and 0.067% or 0.2% dietary genistein for 18 weeks. | Body weight gain, fat deposits, dyslipidemia, hyperglycemia, and fatty liver were ameliorated by dietary genistein. Plasma MGO and plasma, liver and kidney AGEs concentration were significantly lower with genistein. Genistein upregulated the expressions of GLO- 1 and 2 |
| Inacio M. et al. [30] | 2020 | Animal study | C57BL-6J mice were fed a high-fat diet for 14 weeks and treated with 50 mg/kg pentoxifylline during the last 7 weeks. | Pentoxifylline reduced body weight gain, improved insulin sensitivity and glucose tolerance and downregulated biomarkers of glycoxidative stress. |
| Peng W. et al. [31] | 2020 | Systematic review and meta-analysis | PubMed, Scopus and Web of sciences were investigated to identify relevant articles up to June 2019. Inclusion criteria: (1) RCTs, (2) Carnosine use versus any control, (3) intervention for at least 2 weeks. Exclusion criteria: (1) animal studies; (2) studies that surveyed the effect of carnosine along with other components. | Carnosine use versus control for at least 2 weeks showed reduced HbA1C levels in intervention vs control groups. triglyceride levels were not significant reduced. No significant change in HOMA-IR, Cholesterol, fasting blood sugar, or HDL-C. |
| Van den Eynde M. et al.[32] | 2023 | Randomized controlled trial | Individuals with abdominal obesity were randomized to an 8- week intervention with either placebo (n = 36), 25 mg PM (n = 36) or 200 mg PM (n = 36). | PM reduces MGO, AGEs, sVCAM-1 and sICAM-1. No treatment effects on insulin sensitivity, vascular function or other functional outcome measurements. |
| Wilson R. et al. [33] | 2023 | Animal study | RAGE229 (150 parts per million [ppm], approximately 30 mg/kg/d or 50 ppm, approximately 10 mg/kg/d;) was administered to lean mice and mice with obesity undergoing diet-induced weight loss. | RAGE229 reduced body mass and adiposity and improved glucose, insulin, and lipid metabolism in male mice with obesity undergoing weight loss. |
| Chen S.-Y. et al. [34] | 2024 | Animal study | Sprague-Dawley rats were randomly allocated to six groups. The experimental groups included Control, HFD, LWEIG (WEIG, 250 mg kg/b.w./daily), HWEIG (WEIG, 500 mg kg/b.w./daily), ALA (1 mg kg/b.w./daily), and GA (GA, 100 mg kg/b.w./daily), which were administered for 112 days. |
WEIG and GA prevented leptin resistance and MGO, and AGEs accumulation in the liver, kidney, and perinephric fat. WEIG and GA supplementation increased adiponectin, glutathione peroxidase, superoxide dismutase and catalase, and decreased IL-6, IL-1b, TNF-a in the peripheral tissues. |
|
Abbreviations: AGE – advanced glycation end products; RAGE – receptor for advanced glycation end products; ROS- reactive oxygen species; PM- pyridoxamine; ALE- advanced lipoxidation end products; R- ALA – α-lipoic acid; L-CAR – L-carnosine; D-CAR – D- carnosine ; CA – carnosic acid; RE – rosemary extract; HFD – high-fat diet; EGCG – epigallocatechin-3-gallate; HFHS – high-fat/high-sucrose diet; HNE – 4-Hydroxynonenal; DIO – diet-induced obesity; PPB – Pyrogallol-Phloroglucinol-6,6-Bieckol; Glo 1/2 – glyoxalase 1 /2; HOMA – IR – Homeostatic Model Assessment for Insulin Resistance; MGO – methylglyoxal; sVCAM-1 – soluble vascular cell adhesion molecule 1; sICAM-1 – soluble intercellular adhesion molecule-1; LWEIG – low- water extract of Indian gooseberry fruit; WEIG – water extract of Indian gooseberry fruit; HWEIG – High - water extract of Indian gooseberry fruit; GA - gallic acid | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



