Submitted:
08 August 2024
Posted:
12 August 2024
You are already at the latest version
Abstract
Chronic pancreatitis is a progressive fibroinflammatory disorder with no currently satisfactory treatment. Emerging evidence suggests an association between gut microbial dysbiosis and chronic pancreatitis. Although direct causative evidence is lacking, it is hypothesized that the gut microbiota may play a pivotal role in modulating pancreatic function via the gut-pancreas axis. Thus, modulating the gut microbiota through the administration of probiotics or prebiotics may alleviate pancreatic disorders. In this review, we first propose the potential mechanisms by which specific probiotics or prebiotics may ameliorate chronic pancreatitis, including the alleviation of small intestinal bacterial overgrowth (SIBO), facilitation of short-chain fatty acids (SCFAs) production, activation of the glucagon-like peptide 1 receptors (GLP-1Rs) in the pancreas. Since there are currently no probiotics or prebiotics used for the treatment of chronic pancreatitis, we have discussed research in other disease models that use probiotics or prebiotics to modulate pancreatic endocrine and exocrine functions and prevent pancreatic fibrosis. This provides indirect evidence for their potential application in the treatment of chronic pancreatitis. We anticipate that this research will stimulate further investigation into the gut-pancreas axis and the potential therapeutic value of probiotics and prebiotics in chronic pancreatitis.
Keywords:
chronic pancreatitis
; probiotics
; prebiotics
; gut microbiota
1. Introduction
Chronic pancreatitis (CP) is a progressive fibroinflammatory syndrome with an annual incidence of 5 to 8 and a prevalence of 42-73 cases per 100,000 adults in the United States [1,2,3]. With repetitive episodes of inflammation, the pancreas is irreversibly replaced by fibrotic tissues, resulting in chronic abdominal pain, endocrine and exocrine insufficiency, reduced quality of life, and a shorter life expectancy [4]. Current therapeutic approaches primarily focus on symptom alleviation and supportive care, rather than targeting the underlying pathophysiological mechanisms [5].
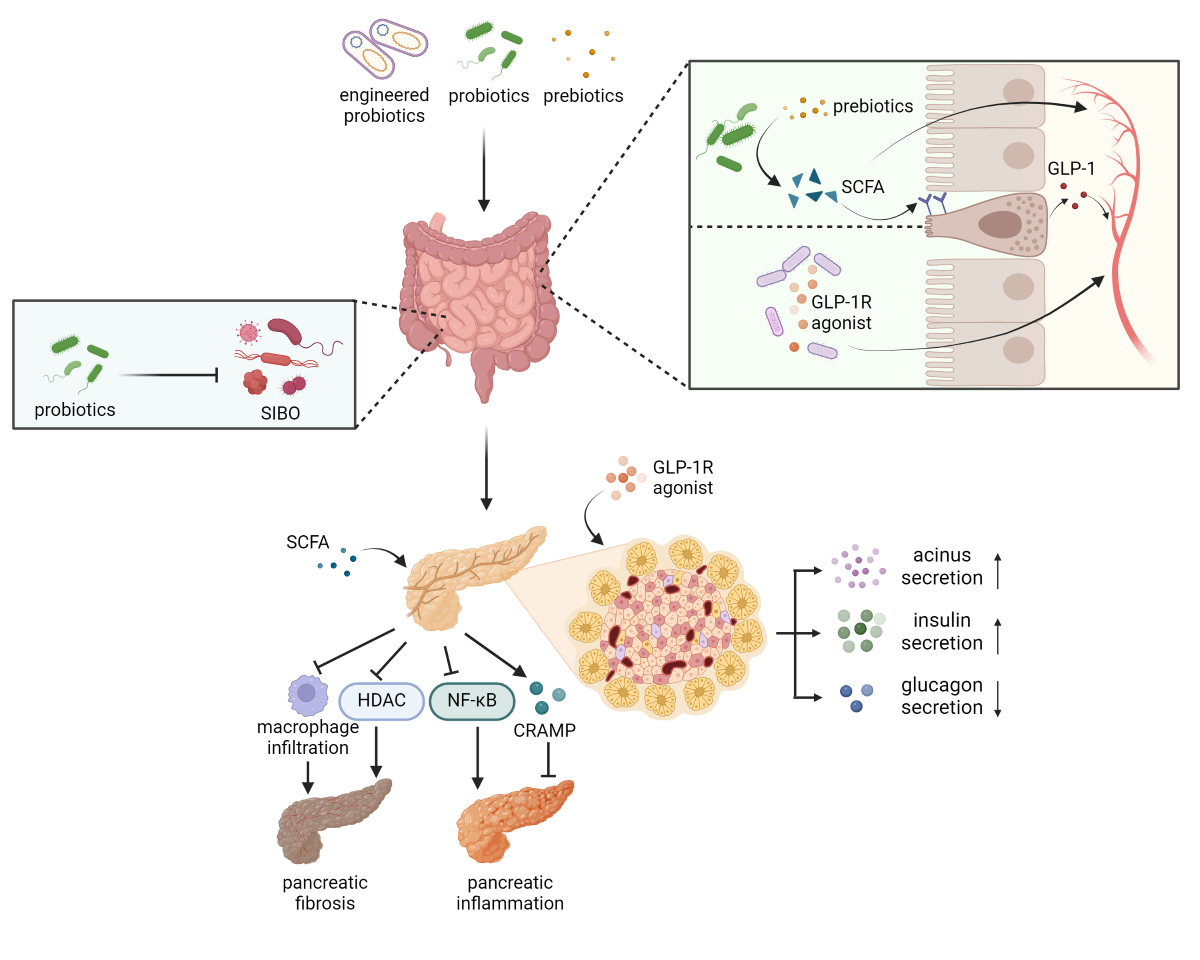
In recent years, accumulating evidence has highlighted the crucial role of the pancreas in regulating gut microbiota and the reciprocal influence of gut microbiota on pancreatic function, which indicates the presence of a bidirectional relationship referred to as the "gut-pancreas axis". Gut microbiota plays a pivotal role in this axis through its involvement in metabolism and nutrition, protection against pathogens, and immune system regulation [6]. Bidirectional alteration of the gut-pancreas axis has been observed in many pancreatic diseases, including CP (Figure 1) [7]. Regarding its role in its role in the homeostasis of the gut-pancreas axis, microbiota-based treatments, such as probiotics and prebiotics, may offer effective therapeutic options for CP.
Probiotics, defined as live microorganisms that confer health benefits to the host, and prebiotics, non-digestible food components that selectively stimulate beneficial gut bacteria, have shown potential in managing various gastrointestinal and systemic disorders [8]. In the context of CP, several mechanisms by which probiotics and prebiotics might exert therapeutic effects have been proposed.
Although probiotics or prebiotics have been proposed as potential treatments for chronic pancreatitis [6,7], their efficacy has not yet been validated in animal models or clinical trials. Additionally, the possible mechanisms of their action have not been thoroughly explored. This review aims to explore the potential of probiotics and prebiotics as therapeutic agents for CP by examining their effects on the gut-pancreas axis. We will discuss possible mechanisms that ameliorate CP, including endocrine and exocrine function improvement, inflammation reduction, and pancreatic fibrosis alleviation. The effects of probiotics and prebiotics on these targets and their feasibility as intervention methods are reviewed. We seek to provide deeper insights into the potential of probiotics and prebiotics in the treatment of CP.
2. Search Strategy
We conducted a comprehensive search in PubMed on July 2024. The search strategy involved the following main queries: (1) "(probiotics OR prebiotics OR synbiotics) AND (pancreatitis)". Literature and reference screening were conducted to select potentially relevant articles. This approach provided a general overview of the current research landscape and potential therapeutic mechanisms of probiotics and prebiotics in CP. After identifying the potential mechanisms of action, the following search queries were involved: (2) (small intestine bacterial overgrowth) AND (chronic pancreatitis); (3) (small intestine bacterial overgrowth) AND (probiotics OR prebiotics OR synbiotics); (4) (short-chain fatty acid) AND (pancreas*); (5) (short-chain fatty acid) AND (probiotics OR prebiotics OR synbiotics); (6) (GLP-1) AND (pancreas*); (7) (GLP-1) AND (probiotics OR prebiotics OR synbiotics).
3. Alleviation of Small Intestinal Bacterial Overgrowth
In a healthy small intestine, several defective mechanisms maintain a relatively sterile environment: gastric acid secretion, an intact ileocecal valve, intestinal motility, immunoglobulins in intestinal secretions, and the bacteriostatic properties of pancreatic and biliary secretions [9,10]. When these protective mechanisms are disrupted, small intestinal bacterial overgrowth syndrome (SIBO) can occur. SIBO is characterized by an excessive number of bacteria in the small bowel, leading to gastrointestinal symptoms such as bloating, abdominal distension, diarrhea, and nutrient deficiencies [11,12]. A systematic review found that SIBO is present in 38% of patients with CP [13]. Current evidence links SIBO in CP to diabetes mellitus, pancreatic exocrine insufficiency, and the severity of CP, with treatment often resulting in symptomatic improvement [13,14,15,16].
The standard treatment for SIBO involves antibiotics aimed at eradicating bacteria in the small intestine [17]. However, with a combined normalization rate of 51% for antibiotics, about half of the patients may remain symptomatic despite treatment [18]. This necessitates refined treatment strategies. Probiotics and prebiotics are believed to benefit SIBO by preventing the growth of pathogenic flora through direct competition and the production of bacteriocins [12]. Several randomized controlled trials have shown that adding probiotics to antibiotic therapy results in higher clinical remission rates [19,20,21]. In a randomized prospective pilot study of patients with SIBO and chronic abdominal distension, the group receiving a combination of probiotics (Lactobacillus casei, Lactobacillus plantarum, Streptococcus faecalis, Bifidobacterium brevis) showed significantly better clinical improvement compared to the sole metronidazole group [22]. A systematic review concluded that while probiotics are unavailable to prevent SIBO, they can effectively decontaminate SIBO and relieve abdominal pain [23]. Probiotics also aid in repairing and reconstructing intestinal mucosa. In rats treated with probiotic formulations containing coconut oil and traces of peppermint-lemon-patchouli essential oil, researchers observed mitotic figures and regression of the inflammatory response in villus epithelium and crypts previously damaged by SIBO-induced gut dysbiosis [24].
Probiotic supplementation to reduce SIBO has been attempted in various diseases, including irritable bowel syndrome [25,26,27,28,29], hypothyroidism during pregnancy [30,31,32], systemic sclerosis [33], liver diseases [34,35,36], and gastric and colorectal cancer [37]. However, there is a lack of research evidence on the application of probiotics for SIBO in chronic pancreatitis. Further investigation is needed to explore the potential benefits of probiotics in alleviating SIBO in CP.
4. Facilitation of Short-Chain Fatty Acids Production
Short-chain fatty acids (SCFAs), primarily acetate, propionate and butyrate, are produced via fermentation of dietary fibers by gut microbiota. They have significant effects on various tissues, including the pancreas. Sodium butyrate is capable of inhibiting histone deacetylases (HDACs), which are crucial in inflammation and fibrogenesis. Post-treatment with sodium butyrate significantly reduces the expression of α-smooth muscle actin, interleukin-1β, inducible nitric oxide synthase, and 3-nitrotyrosine, thereby alleviating L-arginine-induced pancreatic damage and fibrosis in rats [38]. SCFAs modulate pancreatic fibrosis by inhibiting macrophage infiltration and M2 phenotype switching [39]. SCFAs have also been confirmed to play an immunoregulatory and anti-inflammatory role. Cathelicidin-related antimicrobial peptide (CRAMP) is an immunoregulatory antimicrobial peptide and can be produced by acinar cells. It modulates the phenotypic switch of intrapancreatic macrophages and changes the production of transforming growth factor-β, thereby defending against inflammation. Research has revealed that the production of CRAMP is regulated by SCFAs produced by gut microbiota [40]. Additionally, SCFAs, especially butyrate, exhibit anti-inflammatory effects by inhibiting the activation of NF-κB and HDACs [41,42,43,44]. SCFAs also act directly on acinar cells to stimulate secretion, similar to incretins, through increasing cellular calcium concentration [45,46,47,48].
Extensive studies have investigated SCFAs' effects on insulin secretion, acting as ligands to G-protein-coupled receptors (GPCRs), specifically free fatty acid receptor-2 (FFA2, previously termed GPR43) and FFA3 (previously termed GPR41). These receptors are found in various human tissues, including gut enteroendocrine cells and pancreatic islets [49,50]. FFA2 and FFA3 receptors on enteroendocrine cells trigger GLP-1 secretion [49], which has multiple positive effects and will be discussed in the next part. Enhanced secretion of insulin after SCFA treatment has been reported in a number of studies and is thought to be associated with FFA2 and FFA3 receptors on β-cells, but contradicting evidence also exists in several researches [50,51]. Therefore, no clear consensus has been achieved on the effect of SCFAs on FFA2 and FFA3 receptors in pancreatic islets.
Patients with CP exhibit a reduced abundance of SCFA producers, such as Faecalibacterium and Fusicatenibacter [52]. There is a noticeable reduction in Faecalibacterium prausnitzii from healthy controls to CP non-diabetics to CP diabetics [53]. Depletion of SCFA-producing Gram-positive bacteria worse CP independently of TLR4, but supplementing exogenous SCFAs ameliorates the condition [39]. These studies implicated the role of SCFAs in protecting pancreatic function from damage of CP. Therefore, supplementing probiotics or prebiotics that contribute to SCFA production may offer a novel intervention for managing CP.
Both in vivo and in vitro studies confirm that probiotics can increase SCFA levels. Probiotics capable of producing SCFAs are summarized in Table 1. In an in vitro human gut model, an aqueous probiotic suspension, containing L. plantarum, L. rhamnosus, L. acidophilus and Enterococcus faecium, exerted anti-inflammatory effects through increased SCFA production, especially butyrate [54].
Prebiotics also show potential as clinical targets by promoting the growth and activity of probiotics. Prebiotics, typically complex carbohydrates such as starch, pectin, xylan, and arabinogalactan, serve as substrates for bacterial fermentation, resulting in the production of SCFAs [62]. The metabolism of different polysaccharides is associated with the production of different SCFAs. For example, pectin metabolism leads to a proportional increase in acetate concentration, while starch fermentation significantly boosts butyrate production over other SCFAs [63,64]. Overall, the microbial hydrolysis of insoluble substrates can promote the biosynthesis of high concentrations of SCFAs, with about 60% present as acetate, while butyrate and propionate each account for approximately 20% of gastrointestinal SCFAs [65]. Colonic SCFAs increase after consuming inulin or arabinoxylan oligosaccharides-enriched food in healthy humans [66,67]. Inulin supplementation elevates the abundance of butyrate-producing microbiota, including Bifidobacterium, Clostridium cluster IV, and Akkermansia muciniphila [68]. When supplemented with oligofructose or inulin as the sole energy source, cross-feeding interactions between bifidobacteria and butyrate-producing bacteria like Faecalibacterium prausnitzii are observed. These interactions may enhance the colon ecosystem and contribute to combined bifidogenic and butyrogenic effects [69,70].
In summary, the use of probiotics and prebiotics to produce SCFAs shows promise as a management for CP. This approach could help modulate inflammation, fibrosis, and pancreatic function, offering a potential therapeutic avenue worth further exploration.
5. Activation of Glucagon-like Peptide 1 Receptors in the Pancreas
Glucagon-like peptide 1 (GLP-1) is released from gut enteroendocrine cells at low levels during fasting and increases significantly within minutes of food digestion. GLP-1 is a multifaceted hormone with broad pharmacological potential, including incretin-like activity, stimulation of glucose-dependent insulin secretion, and inhibition of glucagon secretion, food intake, and gastric emptying [71,72]. These properties have led to the development of GLP-1 receptor (GLP-1R) agonists for treating T2DM, and subsequently, obesity [73]. The multifunctional role of GLP-1 in the pancreas suggests additional potential for clinical management.
The physiological importance of GLP-1R on β-cells has been well-established in animal studies. GLP-1 normalizes glucose tolerance and enhances glucose-dependent insulin secretion via GLP-1R on pancreatic β-cells [74]. The mechanisms by which GLP-1 restores glucose sensitivity in β-cells involve crosstalk between membrane ion channels, cyclic AMP (cAMP)-dependent signaling, and intracellular glucose metabolism. Additionally, GLP-1 inhibits glucagon secretion, although the expression levels of GLP-1R on α-cells are debated. Some studies report GLP-1R on a subset of α-cells [75], suggesting direct inhibition of glucagon secretion, while others show very low or undetectable levels [76,77,78]. Moreover, GLP-1 acts on GLP-1R on pancreatic δ-cells, stimulating somatostatin secretion, which inhibits glucagon secretion from α-cells via the somatostatin-2 receptor (SSTR2) [78,79].
Beyond regulating blood glucose through modulating levels of insulin and glucagon, GLP-1 inhibits β-cell apoptosis, induces β-cell proliferation, and increases β-cell mass [80]. In diabetic mouse models, GLP-1R activation alleviates ER stress in β-cells via cAMP-dependent enhancement of ATF4 translation, promoting β-cell survival [81]. Although GLP-1R agonists can increase β-cell mass in diabetic rodent models, this effect is modest and short-lived, with older rodents showing reduced response [82,83,84]. Nevertheless, these drugs are believed to help prevent further loss of β-cell mass and function, especially if treatment begins early in disease progression. In baboons subjected to partial pancreatectomy and treated with the GLP-1R agonist exenatide, immunofluorescent staining revealed ductal cells co-expressing insulin, suggesting exenatide might promote the differentiation of ductal cells into β-like cells [85].
While most GLP-1 research focuses on α and β cells in the endocrine pancreas, GLP-1 also affects the exocrine pancreas. GLP-1R is expressed in a significant proportion of pancreatic acinar cells, though at lower levels than in β cells [86,87,88]. In caerulein-induced experimental pancreatitis, GLP-1R agonists increased pancreas weight and induced anti-inflammatory protein expression while reducing proinflammatory markers [89]. Preclinical studies show that GLP-1R activation increases acinar cell mass and protein content via S6 phosphorylation, independent of DNA content or cell proliferation changes [90]. GLP-1 induces amylase secretion in pancreatic acini through stimulated cAMP production and increased protein kinase A-mediated phosphorylation [86]. Moreover, elevation of plasma enzyme levels in human subjects treated with GLP-1R agonists is dose-independent and reversible [91].
The effects of GLP-1 and its analogs on the exocrine pancreas have raised concerns about the risk of pancreatitis. Most of the evaluations were done in patients with type 2 diabetes mellitus (T2DM). A population-based cohort study found that incretin users had a 1.5-fold increased risk of any pancreatitis and a 2.0-fold increased risk of acute pancreatitis, although no increased risk was found for chronic pancreatitis [92]. A review of liraglutide clinical trials reported a higher incidence of pancreatitis, but conclusions were inconclusive due to confounding variables [93]. While the risk of GLP-1 causing pancreatitis remains uncertain, further research is needed to understand its effects in CP compared to T2DM, as the existing evidence is all based on the T2DM population.
Pancreatic stellate cells (PSCs) are activated in the CP microenvironment, contributing to pancreatic fibrosis progression. With the activation of PSC, GLP-1R on it is markedly increased. Studies suggest that GLP-1R agonist liraglutide does not induce inflammatory gene expression in activated PSCs but does induce proliferation [94]. Other studies found chronic GLP-1R agonist treatment can lead to PSC activation, causing the expression of fibrosis markers and chronic inflammation [95]. In the context of CP, further research is needed to determine whether GLP-1R agonists exacerbate pancreatic fibrosis and to understand the underlying mechanisms. It remains to be explored whether there are ways to modify GLP-1R agonists to enhance their positive effects on pancreatic endocrine and exocrine functions while minimizing their impact on PSCs.
In many animal models of other diseases, certain probiotics have been found to induce GLP-1 secretion (Table 2). In addition, using probiotics as oral vectors for recombinant GLP-1R agonists delivery has been explored to replace costly chemical synthesis and inconvenient injections. Probiotics can efficiently target the pancreas, offering high bioavailability. Lactobacillus paracasei L14 transformed with a plasmid encoding the exendin-4 gene has shown efficient secretion and facilitated transport of exendin-4, enhancing insulin secretion and maintaining β cells [96]. Engineered probiotic yeast Saccharomyces boulardii administered orally has also produced bioactive GLP-1R agonists [97]. Apart from delivering GLP-1R agonists, protease-resistant modified GLP-1 (mGLP-1) has been constructed with added arginine to ensure the structural integrity of mGLP-1 released in vivo [98]. In addition to producing bioactive GLP-1R agonists, engineered probiotics as carriers also exert their inherent function of regulating the microbiota. Engineered Clostridium butyricum significantly improved gut microbiota dysbiosis in rats via downregulating the relative abundance of Porphyromonadaceae at the family level and upregulating Lactobacillus at the genus level [99]. Similarly, engineered Escherichia coli Nissle 1917 expressing GLP-1 regulated intestinal flora and increased probiotic diversity in mice [100].
An increase in GLP-1 secretion levels has also been observed following the addition of prebiotics, including dietary resistant starch [101,102], resistant maltodextrin [103], fructooligosaccharides [103,104], chondroitin sulfate [105], and Dendrobium officinale polysaccharide (DOP) [106]. The stimulative effect of prebiotics on GLP-1 secretion may be through stimulating SCFA production [101,102].

Table 2.
Summary of probiotics that can promote GLP-1 expression in various disease models.
| Genus | Species | Disease models | References |
|---|---|---|---|
| Lactobacillus | L. casei CCFM419 | T2DM | [107] |
| L. plantarum MTCC5690 | [108] | ||
| L. fermentum MTCC5689 | [108] | ||
| Lactobacillus CGMCC No. 21661 | [109] | ||
| L. rhamnosus NCDC 17 | [110] | ||
| L. paracasei JY062 | Glycolipid metabolic disorders | [111] | |
| L. reuteri | Glucose metabolism disorder induced by acrylamide; glucose-tolerant humans | [112,113] | |
| L. paracasei subsp. paracasei L. casei W8 | isolated pig intestine | [114] | |
| Lacticaseibacillus | L. paracasei L-21 | STC-1 cell line | [115] |
| Bifidobacterium | selenium-enriched B. longum DD98 | T2DM | [116] |
| B. animalis subsp. lactis MN-Gup | [117] | ||
| B. animalis subsp. lactis NJ241 | Parkinson’s disease | [118] | |
| B. animalis subsp. lactis GCL2505 | Metabolic syndrome | [119] | |
| B. longum subsp. longum B-53 | STC-1 cell line | [115] | |
| Akkermansia | Pasteurized A. muciniphila | T2DM | [120] |
| Bacteroides | B. thetaiotaomicron | alcoholic fatty liver disease | [121] |
| Limosilactobacillus | L. fermentum MG4295 | T2DM | [122] |
| Clostridium | C. butyricum | chronic unpredictable mild stress; T2DM | [123,124] |
In conclusion, the potential of engineered probiotics to express GLP-1 analogs offers a promising avenue for treating CP. Unfortunately, there is currently a lack of experimental evidence regarding the use of engineered probiotics in CP. However, considering their mechanisms of action and the positive effects observed in other disease models, their application in the treatment of CP holds great promise. This approach could improve pancreatic function and manage symptoms more effectively, although further research is needed to fully understand the implications and optimize treatment strategies.
6. Conclusion
Current treatments for CP lack innovation, underscoring the need for novel therapeutic approaches. The gut microbiota can influence pancreatic function through its metabolic activities in the gut, via the gut-pancreas axis. Probiotics and prebiotics may hold the potential for treating CP via this axis.
The three possible intervention mechanisms discussed in this review—alleviating small intestine bacterial overgrowth, facilitating SCFAs production, and activating GLP-1R in the pancreas—are largely based on theoretical extrapolations from existing research, much of which is derived from other pancreatic disease models. Although there is a scarcity of experimental evidence specifically targeting CP, these mechanisms show strong potential for its treatment, including improvement of pancreatic endocrine and exocrine functions, and maintaining cellular and structural integrity. Therefore, there is an urgent need for experimental validation in the field of chronic pancreatitis. This exploration forms the core focus of this review, highlighting the promising potential of these interventions to address the pressing need for improved chronic pancreatitis therapies.
Author Contributions
Conceptualization, D.W. and Y.P.; methodology, Y.P.; software, Y.P.; validation, Y.P., J.L. and Z.F.; formal analysis, Y.C.; investigation, Y.C.; resources, Z.F.; data curation, Z.F.; writing—original draft preparation, Y.P. and J.L.; writing—review and editing, D.W.; visualization, X.H.; supervision, D.W.; project administration, D.W.; funding acquisition, D.W. and J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (grant number 32170788), Beijing Natural Science Foundation (grant number 7232123, 7244390), the National High Level Hospital Clinical Research Funding (grant number 2022- PUMCH-B-023), and the National Key Clinical Specialty Construction Project (grant number ZK108000).
Data Availability Statement
The data underlying this article are available in the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- YADAV D, TIMMONS L, BENSON J T, et al. Incidence, prevalence, and survival of chronic pancreatitis: a population-based study [J]. Am J Gastroenterol, 2011, 106(12): 2192-9. [CrossRef]
- YADAV D, MUDDANA V, O'CONNELL M. Hospitalizations for chronic pancreatitis in Allegheny County, Pennsylvania, USA [J]. Pancreatology, 2011, 11(6): 546-52.
- MACHICADO J D, DUDEKULA A, TANG G, et al. Period prevalence of chronic pancreatitis diagnosis from 2001-2013 in the commercially insured population of the United States [J]. Pancreatology, 2019, 19(6): 813-8.
- PETROV M S, OLESEN S S. Metabolic Sequelae: The Pancreatitis Zeitgeist of the 21st Century [J]. Gastroenterology, 2023, 165(5): 1122-35.
- SINGH V K, YADAV D, GARG P K. Diagnosis and Management of Chronic Pancreatitis: A Review [J]. JAMA, 2019, 322(24): 2422-34.
- THOMAS R M, JOBIN C. Microbiota in pancreatic health and disease: the next frontier in microbiome research [J]. Nat Rev Gastroenterol Hepatol, 2020, 17(1): 53-64.
- LUPU V V, BRATU R M, TRANDAFIR L M, et al. Exploring the Microbial Landscape: Gut Dysbiosis and Therapeutic Strategies in Pancreatitis-A Narrative Review [J]. Biomedicines, 2024, 12(3).
- SANDERS M E, MERENSTEIN D J, REID G, et al. Author Correction: Probiotics and prebiotics in intestinal health and disease: from biology to the clinic [J]. Nat Rev Gastroenterol Hepatol, 2019, 16(10): 642.
- BURES J, CYRANY J, KOHOUTOVA D, et al. Small intestinal bacterial overgrowth syndrome [J]. World J Gastroenterol, 2010, 16(24): 2978-90.
- GRACE E, SHAW C, WHELAN K, et al. Review article: small intestinal bacterial overgrowth--prevalence, clinical features, current and developing diagnostic tests, and treatment [J]. Aliment Pharmacol Ther, 2013, 38(7): 674-88.
- PIMENTEL M, SAAD R J, LONG M D, et al. ACG Clinical Guideline: Small Intestinal Bacterial Overgrowth [J]. Am J Gastroenterol, 2020, 115(2): 165-78.
- ZAFAR H, JIMENEZ B, SCHNEIDER A. Small intestinal bacterial overgrowth: current update [J]. Curr Opin Gastroenterol, 2023, 39(6): 522-8.
- EL KURDI B, BABAR S, EL ISKANDARANI M, et al. Factors That Affect Prevalence of Small Intestinal Bacterial Overgrowth in Chronic Pancreatitis: A Systematic Review, Meta-Analysis, and Meta-Regression [J]. Clin Transl Gastroenterol, 2019, 10(9): e00072. [CrossRef]
- LEE A A, BAKER J R, WAMSTEKER E J, et al. Small Intestinal Bacterial Overgrowth Is Common in Chronic Pancreatitis and Associates With Diabetes, Chronic Pancreatitis Severity, Low Zinc Levels, and Opiate Use [J]. Am J Gastroenterol, 2019, 114(7): 1163-71.
- Ní CHONCHUBHAIR H M, BASHIR Y, DOBSON M, et al. The prevalence of small intestinal bacterial overgrowth in non-surgical patients with chronic pancreatitis and pancreatic exocrine insufficiency (PEI) [J]. Pancreatology, 2018, 18(4): 379-85.
- CAPURSO G, SIGNORETTI M, ARCHIBUGI L, et al. Systematic review and meta-analysis: Small intestinal bacterial overgrowth in chronic pancreatitis [J]. United European Gastroenterol J, 2016, 4(5): 697-705.
- RAO S S C, BHAGATWALA J. Small Intestinal Bacterial Overgrowth: Clinical Features and Therapeutic Management [J]. Clin Transl Gastroenterol, 2019, 10(10): e00078.
- SHAH S C, DAY L W, SOMSOUK M, et al. Meta-analysis: antibiotic therapy for small intestinal bacterial overgrowth [J]. Aliment Pharmacol Ther, 2013, 38(8): 925-34.
- REDONDO-CUEVAS L, BELLOCH L, MARTíN-CARBONELL V, et al. Do Herbal Supplements and Probiotics Complement Antibiotics and Diet in the Management of SIBO? A Randomized Clinical Trial [J]. Nutrients, 2024, 16(7).
- ROSANIA R, GIORGIO F, PRINCIPI M, et al. Effect of probiotic or prebiotic supplementation on antibiotic therapy in the small intestinal bacterial overgrowth: a comparative evaluation [J]. Curr Clin Pharmacol, 2013, 8(2): 169-72.
- KHALIGHI A R, KHALIGHI M R, BEHDANI R, et al. Evaluating the efficacy of probiotic on treatment in patients with small intestinal bacterial overgrowth (SIBO)--a pilot study [J]. Indian J Med Res, 2014, 140(5): 604-8.
- SOIFER L O, PERALTA D, DIMA G, et al. [Comparative clinical efficacy of a probiotic vs. an antibiotic in the treatment of patients with intestinal bacterial overgrowth and chronic abdominal functional distension: a pilot study] [J]. Acta Gastroenterol Latinoam, 2010, 40(4): 323-7.
- ZHONG C, QU C, WANG B, et al. Probiotics for Preventing and Treating Small Intestinal Bacterial Overgrowth: A Meta-Analysis and Systematic Review of Current Evidence [J]. J Clin Gastroenterol, 2017, 51(4): 300-11.
- ASLAN I, TARHAN CELEBI L, KAYHAN H, et al. Probiotic Formulations Containing Fixed and Essential Oils Ameliorates SIBO-Induced Gut Dysbiosis in Rats [J]. Pharmaceuticals (Basel), 2023, 16(7). [CrossRef]
- BARRETT J S, CANALE K E K, GEARRY R B, et al. Probiotic effects on intestinal fermentation patterns in patients with irritable bowel syndrome [J]. World J Gastroenterol, 2008, 14(32): 5020-4.
- YAO C K, BARRETT J S, PHILPOTT H, et al. Poor predictive value of breath hydrogen response for probiotic effects in IBS [J]. J Gastroenterol Hepatol, 2015, 30(12): 1731-9.
- LEE S-H, JOO N-S, KIM K-M, et al. The Therapeutic Effect of a Multistrain Probiotic on Diarrhea-Predominant Irritable Bowel Syndrome: A Pilot Study [J]. Gastroenterol Res Pract, 2018, 2018: 8791916.
- LUO M, LIU Q, XIAO L, et al. Golden bifid might improve diarrhea-predominant irritable bowel syndrome via microbiota modulation [J]. J Health Popul Nutr, 2022, 41(1): 21.
- BUSTOS FERNáNDEZ L M, MAN F, LASA J S. Impact of Saccharomyces boulardii CNCM I-745 on Bacterial Overgrowth and Composition of Intestinal Microbiota in Diarrhea-Predominant Irritable Bowel Syndrome Patients: Results of a Randomized Pilot Study [J]. Dig Dis, 2023, 41(5): 798-809.
- HAO Y, XU Y, BAN Y, et al. Efficacy evaluation of probiotics combined with prebiotics in patients with clinical hypothyroidism complicated with small intestinal bacterial overgrowth during the second trimester of pregnancy [J]. Front Cell Infect Microbiol, 2022, 12: 983027.
- ZHANG M, XU Y, SUN Z, et al. Evaluation of probiotics in the treatment of hypothyroidism in early pregnancy combined with small intestinal bacterial overgrowth [J]. Food Sci Nutr, 2024, 12(4): 2671-8.
- OUYANG Q, XU Y, BAN Y, et al. Probiotics and Prebiotics in Subclinical Hypothyroidism of Pregnancy with Small Intestinal Bacterial Overgrowth [J]. Probiotics Antimicrob Proteins, 2024, 16(2): 579-88.
- GARCíA-COLLINOT G, MADRIGAL-SANTILLáN E O, MARTíNEZ-BENCOMO M A, et al. Effectiveness of Saccharomyces boulardii and Metronidazole for Small Intestinal Bacterial Overgrowth in Systemic Sclerosis [J]. Dig Dis Sci, 2020, 65(4): 1134-43.
- EFREMOVA I, MASLENNIKOV R, ZHARKOVA M, et al. Efficacy and Safety of a Probiotic Containing Saccharomyces boulardii CNCM I-745 in the Treatment of Small Intestinal Bacterial Overgrowth in Decompensated Cirrhosis: Randomized, Placebo-Controlled Study [J]. J Clin Med, 2024, 13(3).
- FEROLLA S M, COUTO C A, COSTA-SILVA L, et al. Beneficial Effect of Synbiotic Supplementation on Hepatic Steatosis and Anthropometric Parameters, But Not on Gut Permeability in a Population with Nonalcoholic Steatohepatitis [J]. Nutrients, 2016, 8(7).
- KWAK D S, JUN D W, SEO J G, et al. Short-term probiotic therapy alleviates small intestinal bacterial overgrowth, but does not improve intestinal permeability in chronic liver disease [J]. Eur J Gastroenterol Hepatol, 2014, 26(12): 1353-9.
- LIANG S, XU L, ZHANG D, et al. Effect of probiotics on small intestinal bacterial overgrowth in patients with gastric and colorectal cancer [J]. Turk J Gastroenterol, 2016, 27(3): 227-32.
- KANIKA G, KHAN S, JENA G. Sodium Butyrate Ameliorates L-Arginine-Induced Pancreatitis and Associated Fibrosis in Wistar Rat: Role of Inflammation and Nitrosative Stress [J]. J Biochem Mol Toxicol, 2015, 29(8): 349-59. [CrossRef]
- PAN L-L, REN Z-N, YANG J, et al. Gut microbiota controls the development of chronic pancreatitis: A critical role of short-chain fatty acids-producing Gram-positive bacteria [J]. Acta Pharm Sin B, 2023, 13(10): 4202-16.
- SUN J, FURIO L, MECHERI R, et al. Pancreatic β-Cells Limit Autoimmune Diabetes via an Immunoregulatory Antimicrobial Peptide Expressed under the Influence of the Gut Microbiota [J]. Immunity, 2015, 43(2): 304-17.
- VINOLO M A R, RODRIGUES H G, HATANAKA E, et al. Suppressive effect of short-chain fatty acids on production of proinflammatory mediators by neutrophils [J]. J Nutr Biochem, 2011, 22(9): 849-55.
- PARK J-S, LEE E-J, LEE J-C, et al. Anti-inflammatory effects of short chain fatty acids in IFN-gamma-stimulated RAW 264.7 murine macrophage cells: involvement of NF-kappaB and ERK signaling pathways [J]. Int Immunopharmacol, 2007, 7(1): 70-7.
- USAMI M, KISHIMOTO K, OHATA A, et al. Butyrate and trichostatin A attenuate nuclear factor kappaB activation and tumor necrosis factor alpha secretion and increase prostaglandin E2 secretion in human peripheral blood mononuclear cells [J]. Nutr Res, 2008, 28(5): 321-8.
- PARK G Y, JOO M, PEDCHENKO T, et al. Regulation of macrophage cyclooxygenase-2 gene expression by modifications of histone H3 [J]. Am J Physiol Lung Cell Mol Physiol, 2004, 286(5): L956-L62.
- HARADA E, KATO S. Effect of short-chain fatty acids on the secretory response of the ovine exocrine pancreas [J]. Am J Physiol, 1983, 244(3): G284-G90.
- KATOH K, TSUDA T. Effects of secretagogues on membrane potential and input resistance of pancreatic acinar cells of sheep [J]. Res Vet Sci, 1985, 38(2): 250-1.
- KATOH K, TSUDA T. Effects of acetylcholine and short-chain fatty acids on acinar cells of the exocrine pancreas in sheep [J]. J Physiol, 1984, 356: 479-89.
- KATOH K, TSUDA T. Effects of intravenous injection of butyrate on the exocrine pancreatic secretion in guinea pigs [J]. Comp Biochem Physiol A Comp Physiol, 1987, 87(3): 569-72.
- LU V B, GRIBBLE F M, REIMANN F. Free Fatty Acid Receptors in Enteroendocrine Cells [J]. Endocrinology, 2018, 159(7): 2826-35.
- TEYANI R, MONIRI N H. Gut feelings in the islets: The role of the gut microbiome and the FFA2 and FFA3 receptors for short chain fatty acids on β-cell function and metabolic regulation [J]. Br J Pharmacol, 2023, 180(24): 3113-29.
- ROSLI N S A, ABD GANI S, KHAYAT M E, et al. Short-chain fatty acids: possible regulators of insulin secretion [J]. Mol Cell Biochem, 2023, 478(3): 517-30.
- FROST F, WEISS F U, SENDLER M, et al. The Gut Microbiome in Patients With Chronic Pancreatitis Is Characterized by Significant Dysbiosis and Overgrowth by Opportunistic Pathogens [J]. Clin Transl Gastroenterol, 2020, 11(9): e00232. [CrossRef]
- JANDHYALA S M, MADHULIKA A, DEEPIKA G, et al. Altered intestinal microbiota in patients with chronic pancreatitis: implications in diabetes and metabolic abnormalities [J]. Sci Rep, 2017, 7: 43640.
- MOENS F, VAN DEN ABBEELE P, BASIT A W, et al. A four-strain probiotic exerts positive immunomodulatory effects by enhancing colonic butyrate production in vitro [J]. Int J Pharm, 2019, 555.
- KOH A, DE VADDER F, KOVATCHEVA-DATCHARY P, et al. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites [J]. Cell, 2016, 165(6): 1332-45.
- LEBLANC J G, CHAIN F, MARTíN R, et al. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria [J]. Microb Cell Fact, 2017, 16(1): 79.
- MEIMANDIPOUR A, HAIR-BEJO M, SHUHAIMI M, et al. Gastrointestinal tract morphological alteration by unpleasant physical treatment and modulating role of Lactobacillus in broilers [J]. Br Poult Sci, 2010, 51(1): 52-9.
- SIVIERI K, MORALES M L V, ADORNO M A T, et al. Lactobacillus acidophilus CRL 1014 improved "gut health" in the SHIME reactor [J]. BMC Gastroenterol, 2013, 13: 100.
- AMARETTI A, BERNARDI T, TAMBURINI E, et al. Kinetics and metabolism of Bifidobacterium adolescentis MB 239 growing on glucose, galactose, lactose, and galactooligosaccharides [J]. Appl Environ Microbiol, 2007, 73(11): 3637-44.
- ABDIN A A, SAEID E M. An experimental study on ulcerative colitis as a potential target for probiotic therapy by Lactobacillus acidophilus with or without "olsalazine" [J]. J Crohns Colitis, 2008, 2(4): 296-303.
- SALAZAR N, BINETTI A, GUEIMONDE M, et al. Safety and intestinal microbiota modulation by the exopolysaccharide-producing strains Bifidobacterium animalis IPLA R1 and Bifidobacterium longum IPLA E44 orally administered to Wistar rats [J]. Int J Food Microbiol, 2011, 144(3): 342-51.
- BYRNE C S, CHAMBERS E S, MORRISON D J, et al. The role of short chain fatty acids in appetite regulation and energy homeostasis [J]. Int J Obes (Lond), 2015, 39(9): 1331-8.
- MORRISON D J, PRESTON T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism [J]. Gut Microbes, 2016, 7(3): 189-200.
- PASCALE N, GU F, LARSEN N, et al. The Potential of Pectins to Modulate the Human Gut Microbiota Evaluated by In Vitro Fermentation: A Systematic Review [J]. Nutrients, 2022, 14(17).
- MACFARLANE S, MACFARLANE G T. Regulation of short-chain fatty acid production [J]. Proc Nutr Soc, 2003, 62(1): 67-72.
- BOETS E, DEROOVER L, HOUBEN E, et al. Quantification of in Vivo Colonic Short Chain Fatty Acid Production from Inulin [J]. Nutrients, 2015, 7(11): 8916-29.
- FRANçOIS I E J A, LESCROART O, VERAVERBEKE W S, et al. Effects of a wheat bran extract containing arabinoxylan oligosaccharides on gastrointestinal health parameters in healthy adult human volunteers: a double-blind, randomised, placebo-controlled, cross-over trial [J]. Br J Nutr, 2012, 108(12): 2229-42.
- GUIMARãES J B, RODRIGUES V F, PEREIRA Í S, et al. Inulin prebiotic ameliorates type 1 diabetes dictating regulatory T cell homing via CCR4 to pancreatic islets and butyrogenic gut microbiota in murine model [J]. J Leukoc Biol, 2024, 115(3): 483-96.
- FALONY G, VLACHOU A, VERBRUGGHE K, et al. Cross-feeding between Bifidobacterium longum BB536 and acetate-converting, butyrate-producing colon bacteria during growth on oligofructose [J]. Appl Environ Microbiol, 2006, 72(12): 7835-41. [CrossRef]
- MOENS F, WECKX S, DE VUYST L. Bifidobacterial inulin-type fructan degradation capacity determines cross-feeding interactions between bifidobacteria and Faecalibacterium prausnitzii [J]. Int J Food Microbiol, 2016, 231: 76-85.
- DRUCKER D, J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1 [J]. Cell Metab, 2018, 27(4): 740-56.
- MüLLER T D, FINAN B, BLOOM S R, et al. Glucagon-like peptide 1 (GLP-1) [J]. Mol Metab, 2019, 30.
- NOGUEIRAS R, NAUCK M A, TSCHöP M H. Gut hormone co-agonists for the treatment of obesity: from bench to bedside [J]. Nat Metab, 2023, 5(6): 933-44.
- DRUCKER D J, HABENER J F, HOLST J J. Discovery, characterization, and clinical development of the glucagon-like peptides [J]. J Clin Invest, 2017, 127(12): 4217-27.
- RICHARDS P, PARKER H E, ADRIAENSSENS A E, et al. Identification and characterization of GLP-1 receptor-expressing cells using a new transgenic mouse model [J]. Diabetes, 2014, 63(4): 1224-33.
- MURARO M J, DHARMADHIKARI G, GRüN D, et al. A Single-Cell Transcriptome Atlas of the Human Pancreas [J]. Cell Syst, 2016, 3(4).
- SEGERSTOLPE Å, PALASANTZA A, ELIASSON P, et al. Single-Cell Transcriptome Profiling of Human Pancreatic Islets in Health and Type 2 Diabetes [J]. Cell Metab, 2016, 24(4): 593-607.
- WASER B, BLANK A, KARAMITOPOULOU E, et al. Glucagon-like-peptide-1 receptor expression in normal and diseased human thyroid and pancreas [J]. Mod Pathol, 2015, 28(3): 391-402.
- DE HEER J, RASMUSSEN C, COY D H, et al. Glucagon-like peptide-1, but not glucose-dependent insulinotropic peptide, inhibits glucagon secretion via somatostatin (receptor subtype 2) in the perfused rat pancreas [J]. Diabetologia, 2008, 51(12): 2263-70.
- CAMPBELL J E, DRUCKER D J. Pharmacology, physiology, and mechanisms of incretin hormone action [J]. Cell Metab, 2013, 17(6): 819-37.
- YUSTA B, BAGGIO L L, ESTALL J L, et al. GLP-1 receptor activation improves beta cell function and survival following induction of endoplasmic reticulum stress [J]. Cell Metab, 2006, 4(5): 391-406.
- LAMONT B J, ANDRIKOPOULOS S. Hope and fear for new classes of type 2 diabetes drugs: is there preclinical evidence that incretin-based therapies alter pancreatic morphology? [J]. J Endocrinol, 2014, 221(1): T43-T61.
- RANKIN M M, KUSHNER J A. Adaptive beta-cell proliferation is severely restricted with advanced age [J]. Diabetes, 2009, 58(6): 1365-72.
- TSCHEN S-I, GEORGIA S, DHAWAN S, et al. Skp2 is required for incretin hormone-mediated β-cell proliferation [J]. Mol Endocrinol, 2011, 25(12): 2134-43.
- FIORENTINO T V, OWSTON M, ABRAHAMIAN G, et al. Chronic continuous exenatide infusion does not cause pancreatic inflammation and ductal hyperplasia in non-human primates [J]. Am J Pathol, 2015, 185(1): 139-50.
- HOU Y, ERNST S A, HEIDENREICH K, et al. Glucagon-like peptide-1 receptor is present in pancreatic acinar cells and regulates amylase secretion through cAMP [J]. Am J Physiol Gastrointest Liver Physiol, 2016, 310(1): G26-G33.
- PYKE C, HELLER R S, KIRK R K, et al. GLP-1 receptor localization in monkey and human tissue: novel distribution revealed with extensively validated monoclonal antibody [J]. Endocrinology, 2014, 155(4): 1280-90.
- KIRK R K, PYKE C, VON HERRATH M G, et al. Immunohistochemical assessment of glucagon-like peptide 1 receptor (GLP-1R) expression in the pancreas of patients with type 2 diabetes [J]. Diabetes Obes Metab, 2017, 19(5): 705-12.
- KOEHLER J A, BAGGIO L L, LAMONT B J, et al. Glucagon-like peptide-1 receptor activation modulates pancreatitis-associated gene expression but does not modify the susceptibility to experimental pancreatitis in mice [J]. Diabetes, 2009, 58(9): 2148-61.
- KOEHLER J A, BAGGIO L L, CAO X, et al. Glucagon-like peptide-1 receptor agonists increase pancreatic mass by induction of protein synthesis [J]. Diabetes, 2015, 64(3): 1046-56.
- STEINBERG W M, ROSENSTOCK J, WADDEN T A, et al. Erratum. Impact of Liraglutide on Amylase, Lipase, and Acute Pancreatitis in Participants With Overweight/Obesity and Normoglycemia, Prediabetes, or Type 2 Diabetes: Secondary Analyses of Pooled Data From the SCALE Clinical Development Program. Diabetes Care 2017;40:839-848 [J]. Diabetes Care, 2018, 41(7): 1538.
- KNAPEN L M, DE JONG R G P J, DRIESSEN J H M, et al. Use of incretin agents and risk of acute and chronic pancreatitis: A population-based cohort study [J]. Diabetes Obes Metab, 2017, 19(3): 401-11.
- JENSEN T M, SAHA K, STEINBERG W M. Erratum. Is There a Link Between Liraglutide and Pancreatitis? A Post Hoc Review of Pooled and Patient-Level Data From Completed Liraglutide Type 2 Diabetes Clinical Trials. Diabetes Care 2015;38:1058-1066 [J]. Diabetes Care, 2015, 38(8): 1622.
- NAKAMURA T, ITO T, UCHIDA M, et al. PSCs and GLP-1R: occurrence in normal pancreas, acute/chronic pancreatitis and effect of their activation by a GLP-1R agonist [J]. Lab Invest, 2014, 94(1): 63-78.
- YANG Y, YU X, HUANG L, et al. GLP-1R agonist may activate pancreatic stellate cells to induce rat pancreatic tissue lesion [J]. Pancreatology, 2013, 13(5): 498-501.
- ZENG Z, YU R, ZUO F, et al. Heterologous Expression and Delivery of Biologically Active Exendin-4 by Lactobacillus paracasei L14 [J]. PLoS One, 2016, 11(10): e0165130.
- HEDIN K A, ZHANG H, KRUSE V, et al. Cold Exposure and Oral Delivery of GLP-1R Agonists by an Engineered Probiotic Yeast Strain Have Antiobesity Effects in Mice [J]. ACS Synth Biol, 2023, 12(11): 3433-42.
- WANG Q, GUO H, MAO W, et al. The Oral Delivery System of Modified GLP-1 by Probiotics for T2DM [J]. Pharmaceutics, 2023, 15(4).
- WANG X-L, CHEN W-J, JIN R, et al. Engineered probiotics Clostridium butyricum-pMTL007-GLP-1 improves blood pressure via producing GLP-1 and modulating gut microbiota in spontaneous hypertension rat models [J]. Microb Biotechnol, 2023, 16(4): 799-812. [CrossRef]
- WANG Y, SHI Y, PENG X, et al. Biochemotaxis-Oriented Engineering Bacteria Expressing GLP-1 Enhance Diabetes Therapy by Regulating the Balance of Immune [J]. Adv Healthc Mater, 2024, 13(11): e2303958.
- ZHOU J, MARTIN R J, TULLEY R T, et al. Dietary resistant starch upregulates total GLP-1 and PYY in a sustained day-long manner through fermentation in rodents [J]. Am J Physiol Endocrinol Metab, 2008, 295(5): E1160-E6.
- SHEN L, KEENAN M J, RAGGIO A, et al. Dietary-resistant starch improves maternal glycemic control in Goto-Kakizaki rat [J]. Mol Nutr Food Res, 2011, 55(10): 1499-508.
- HIRA T, SUTO R, KISHIMOTO Y, et al. Resistant maltodextrin or fructooligosaccharides promotes GLP-1 production in male rats fed a high-fat and high-sucrose diet, and partially reduces energy intake and adiposity [J]. Eur J Nutr, 2018, 57(3): 965-79.
- WONGKRASANT P, PONGKORPSAKOL P, CHITWATTANANONT S, et al. Fructo-oligosaccharides alleviate inflammation-associated apoptosis of GLP-1 secreting L cells via inhibition of iNOS and cleaved caspase-3 expression [J]. J Pharmacol Sci, 2020, 143(2): 65-73.
- PICHETTE J, FYNN-SACKEY N, GAGNON J. Hydrogen Sulfide and Sulfate Prebiotic Stimulates the Secretion of GLP-1 and Improves Glycemia in Male Mice [J]. Endocrinology, 2017, 158(10): 3416-25.
- LIU H, XING Y, WANG Y, et al. Dendrobium officinale Polysaccharide Prevents Diabetes via the Regulation of Gut Microbiota in Prediabetic Mice [J]. Foods, 2023, 12(12).
- WANG G, LI X, ZHAO J, et al. Lactobacillus casei CCFM419 attenuates type 2 diabetes via a gut microbiota dependent mechanism [J]. Food Funct, 2017, 8(9): 3155-64.
- BALAKUMAR M, PRABHU D, SATHISHKUMAR C, et al. Improvement in glucose tolerance and insulin sensitivity by probiotic strains of Indian gut origin in high-fat diet-fed C57BL/6J mice [J]. Eur J Nutr, 2018, 57(1): 279-95.
- WANG Y, WANG X, XIAO X, et al. A Single Strain of Lactobacillus (CGMCC 21661) Exhibits Stable Glucose- and Lipid-Lowering Effects by Regulating Gut Microbiota [J]. Nutrients, 2023, 15(3).
- SINGH S, SHARMA R K, MALHOTRA S, et al. Lactobacillus rhamnosus NCDC17 ameliorates type-2 diabetes by improving gut function, oxidative stress and inflammation in high-fat-diet fed and streptozotocintreated rats [J]. Benef Microbes, 2017, 8(2): 243-55.
- SU Y, REN J, ZHANG J, et al. Lactobacillus paracasei JY062 Alleviates Glucolipid Metabolism Disorders via the Adipoinsular Axis and Gut Microbiota [J]. Nutrients, 2024, 16(2).
- YUE Z, ZHAO F, GUO Y, et al. Lactobacillus reuteri JCM 1112 ameliorates chronic acrylamide-induced glucose metabolism disorder via the bile acid-TGR5-GLP-1 axis and modulates intestinal oxidative stress in mice [J]. Food Funct, 2024, 15(12): 6450-8.
- SIMON M-C, STRASSBURGER K, NOWOTNY B, et al. Intake of Lactobacillus reuteri improves incretin and insulin secretion in glucose-tolerant humans: a proof of concept [J]. Diabetes Care, 2015, 38(10): 1827-34.
- BJERG A T, KRISTENSEN M, RITZ C, et al. Lactobacillus paracasei subsp paracasei L. casei W8 suppresses energy intake acutely [J]. Appetite, 2014, 82: 111-8.
- CHENG Z, CHEN J, ZHANG Y, et al. In Vitro Hypoglycemic Activities of Lactobacilli and Bifidobacterium Strains from Healthy Children's Sources and Their Effect on Stimulating GLP-1 Secretion in STC-1 Cells [J]. Foods, 2024, 13(4). [CrossRef]
- ZHAO D, ZHU H, GAO F, et al. Antidiabetic effects of selenium-enriched Bifidobacterium longum DD98 in type 2 diabetes model of mice [J]. Food Funct, 2020, 11(7): 6528-41.
- ZHANG C, FANG B, ZHANG N, et al. The Effect of Bifidobacterium animalis subsp. lactis MN-Gup on Glucose Metabolism, Gut Microbiota, and Their Metabolites in Type 2 Diabetic Mice [J]. Nutrients, 2024, 16(11).
- DONG Y, QI Y, CHEN J, et al. Neuroprotective Effects of Bifidobacterium animalis subsp. lactis NJ241 in a Mouse Model of Parkinson's Disease: Implications for Gut Microbiota and PGC-1α [J]. Mol Neurobiol, 2024.
- AOKI R, KAMIKADO K, SUDA W, et al. A proliferative probiotic Bifidobacterium strain in the gut ameliorates progression of metabolic disorders via microbiota modulation and acetate elevation [J]. Sci Rep, 2017, 7: 43522.
- NIU H, ZHOU M, JI A, et al. Molecular Mechanism of Pasteurized Akkermansia muciniphila in Alleviating Type 2 Diabetes Symptoms [J]. J Agric Food Chem, 2024, 72(23): 13083-98.
- SANGINETO M, GRANDER C, GRABHERR F, et al. Recovery of Bacteroides thetaiotaomicron ameliorates hepatic steatosis in experimental alcohol-related liver disease [J]. Gut Microbes, 2022, 14(1): 2089006.
- KIM J E, LEE J Y, KANG C-H. Limosilactobacillus fermentum MG4295 Improves Hyperglycemia in High-Fat Diet-Induced Mice [J]. Foods, 2022, 11(2).
- SUN J, WANG F, HU X, et al. Clostridium butyricum Attenuates Chronic Unpredictable Mild Stress-Induced Depressive-Like Behavior in Mice via the Gut-Brain Axis [J]. J Agric Food Chem, 2018, 66(31): 8415-21.
- JIA L, LI D, FENG N, et al. Anti-diabetic Effects of Clostridium butyricum CGMCC0313.1 through Promoting the Growth of Gut Butyrate-producing Bacteria in Type 2 Diabetic Mice [J]. Sci Rep, 2017, 7(1): 7046. [CrossRef]
Figure 1.
Bidirectional gut-pancreas interactions in the context of CP. SCFA, short-chain fatty acid; GLP-1, glucagon-like peptide 1; SIBO, small intestinal bacterial overgrowth; AMP, antimicrobial peptide. This figure was created with BioRender.com.
Figure 1.
Bidirectional gut-pancreas interactions in the context of CP. SCFA, short-chain fatty acid; GLP-1, glucagon-like peptide 1; SIBO, small intestinal bacterial overgrowth; AMP, antimicrobial peptide. This figure was created with BioRender.com.
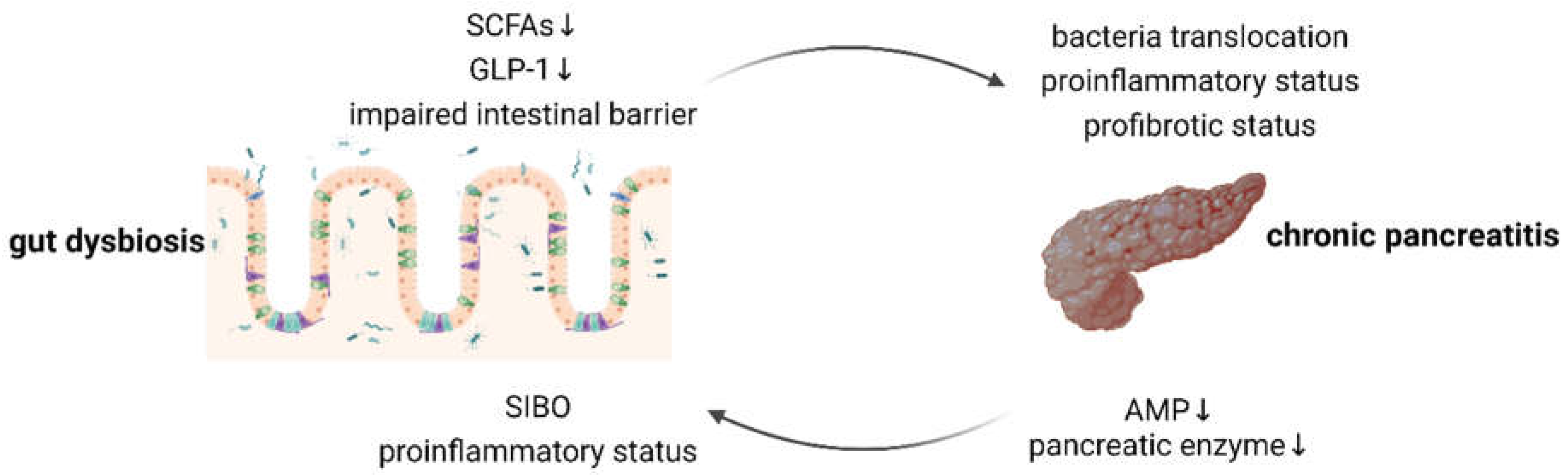
Table 1.
Main probiotics producing short-chain fatty acids.
| Probiotics | Products | References |
|---|---|---|
| Bifidobacterium spp. | acetate, butyrate | [55] |
| Lactobacillus rhamnosus GG | propionate | [56] |
| Lactobacillus gasseri PA 16/8 | ||
| Bifidobacterium longum SP 07/3 | acetate, propionate | |
| Bifidobacterium bifidum MF 20/5 | ||
| Lactobacillus salivarius spp salcinius JCM 1230 | propionate, butyrate | [57] |
| Lactobacillus agilis JCM 1048 | ||
| Lactobacillus acidophilus CRL 1014 | acetate, propionate, butyrate | [58,59,60,61] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



