
Submitted:
29 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
Oxidative stress and chronic inflammation, both at the systemic and central level, are critical early events in atherosclerosis and Alzheimer’s disease (AD). Purpose. To investigate the oxidative stress, inflammatory, and Tau-phosphorylation lowering effects of pomegranate polyphenols (PP) (punicalagin, ellagic acid, peels, and arils extracts). Methods. We used flow cytometry to quantify protein expression of proinflammatory cytokines (IL-1β) and anti-inflammatory mediators (IL-10) in THP-1 macrophages, as well as M1/M2 cell-specific markers (CD86 and CD163) expression in human microglia HMC3 cells. IL-10 protein expression was also quantified in U373-MG human astrocytes. The effect of PP on human amyloid beta 1-42 (Aβ1-42)-induced oxidative stress was assessed in microglia by measuring ROS generation and lipid peroxidation, using respectively two ′,7′-dichlorofluorescein diacetate (DCFH-DA) and thiobarbituric acid reactive substances (TBARS) tests. Neuronal viability and cell apoptotic response to Aβ1-42 toxicity were assayed using the MTT (3-(4, 5-dimethyl thiazolyl-2)-2, 5-diphenyltetrazolium bromide) assay and the annexinV-FITC apoptosis detection kit; respectively. Finally, flow cytometry analysis was also performed to evaluate the ability of PP to modulate Aβ1-42-induced Tau-181 phosphorylation (pTau-181). Results. Our data indicate that PP was significantly (p<0.05) effective in countering Aβ1-42-induced inflammation through increasing the anti-inflammatory cytokines (IL-10) (in U373-MG astrocytes and THP1 macrophages) and decreasing proinflammatory markers (IL-1β) expression in THP1 macrophages. PP was also significantly (p<0.05) effective in inducing the phenotypic transition of THP-1 macrophages and microglial cells from M1 to M2 by decreasing CD86 and increasing CD163 surface receptor expression. Moreover, our treatments have a significant (p<0.05) beneficial impact on oxidative stress, illustrated in the reduction of TBARS and ROS generation. Our treatments have significant (p<0.05) cell viability improvement capacities and anti-apoptotic effects on human H4 neurons. Furthermore, our results suggest that Aβ1-42 significantly (p<0.05) increases pTau-181. This effect was significantly (p<0.05) attenuated by arils, peels, and punicalagin and drastically reduced by ellagic acid treatments. Conclusion. Our results are attributed to PP's anti-inflammatory, antioxidant, anti-apoptotic, and anti-Tau pathology potential. Future studies should aim to extend our knowledge of the potential role of PP on A1-42-induced neurodegeneration, mainly its association with the tauopathy involved in AD.
Keywords:
Neuroinflammation
; Alzheimer’s disease
; pomegranate (Punica granatum L)
; oxidative stress
; microglia
; amyloid-beta
; phospho-Tau-181
; ellagic acid
; punicalagin
1. Introduction
It was necessary to wait until 1988 so that Rogers and coworkers could highlight the involvement of immune response in the aetiology of Alzheimer’s disease (AD) [1]. Neuroinflammation and neuroimmune response in AD involve astrocytes and microglia activation. Astrocytes are multifunctional “housekeeping” cells and the most abundant type of glial cells in the central nervous system (CNS). These dynamic regulators ensure vital functions such as brain water homeostasis [2], blood flow regulation [3], dynamic synaptic structure control, extracellular pH regulation [5], and detoxification of brain free radical species [6]. Microglia cells are dynamic immunocompetent cells and “brain resident macrophages” that phagocyte neuronal debris and clean aggregated amyloid-beta (Aβ) fragments [7]. They maintain brain homeostasis by ensuring critical CNS activities, including brain development [8], neuronal survival [9], neurogenesis, and synaptic remodelling [10].
In injured or diseased conditions, these cells become chronically activated and adopt the M1 phenotype to express several cell surface receptors, such as CD80 and CD86, as well as a variety of reactive oxygen species and proinflammatory cytokines (interleukin-1 beta (IL-1β), interleukin-6 (IL-6), interferon-alpha (INF-α), and tumour necrosis factor-alpha (TNF-α)), and chemokines. In most cases, they can accelerate the healing process and strengthen recovery. However, for various reasons, including chronic exposure to Aβ [11] due to its overexpression or due to the failure of its clearance by microglia cells, the M1 phenotype may persist, leading to brain injury, chronic neuroinflammation, oxidative stress, reduced neuronal viability, and brain atrophy. Mechanistically, soluble Aβ can bind numerous microglia molecular pattern recognition and G protein-coupled receptors, such as toll-like and formyl peptide receptors, to induce MAPK/Erk and NFκB signalling pathways directly [12,13]. This leads to the expression of many pro-inflammatory genes, such as TNFα, IL-1β, and IL6 [14].
Consequently, Aβ-peptide targets neuroinflammation [15], oxidative stress [16], blood-brain barrier transporter disruption [17], neuronal apoptosis [18], and Tau-phosphorylation [19], as well as enhances astrocyte reactivity [20]. All these events appear to be causative alterations in AD. On the other hand, the implication of neuroinflammation in AD progression is also supported by numerous epidemiological studies, which indicated that the intake of non-steroidal anti-inflammatory drugs (NSAIDs) can reduce the risk of developing AD [21,22,23]. In addition, the anti-inflammatory therapies were reported to be effective in altering Aβ processing and deposition [24], providing additional proof and pointing to the aberrant involvement of neuroinflammation in AD-aetiology.
Systemic inflammation is an immune process that localises, opsonises, and eliminates toxic compounds, pathological aggressors, and the source of insult. Failure of this healing process results in chronic inflammation, which, over time, may lead to numerous chronic inflammatory diseases such as cancer, neurodegenerative diseases, and atherosclerosis. Atherosclerosis is a multi-stage process of arterial plaque formation and an immunoinflammatory disease that affects large and medium-sized arteries, causing fatal complications such as heart attack, vascular dementia, and stroke. Macrophage plasticity and their M1/M2 phenotypes contribute to atherosclerosis. M1-macrophages phenotype seems to be adopted in response to exogenous/endogenous stressors, while M2-phenotype is generated to help tissue repair, modulate inflammatory phases, and accelerate the recovery process.
It has been shown that peripheral inflammation can induce neuroinflammation [25,26]. Systemic proinflammatory mediators can modulate the neuronal microenvironment, activating microglial cells [25,26]. This immunity decline results in the organism's inability to adequately manage its inflammatory state, leading to various pathological consequences, such as reduced brain-blood barrier selectivity [27] and death of glial and neuronal cells [28,29]. This gradual deterioration of brain homeostasis can result in progressive cognitive and behavioural deficits and point to a strong link between peripheral and central inflammation.
Ellagitannins-rich foods and diet-containing polyphenols have received considerable attention due to their correlation with a lower risk of developing several chronic inflammatory and oxidative stress-related diseases [30,31,32]. As our team previously published [33], pomegranate (Sefri variety) contains various active phytochemicals such as anthocyanins, flavonoids, gallic acid, ellagic acid, α and β-punicalagin. Also, we demonstrated that pomegranate peels and arils phenolic extracts inhibit reactive oxygen species (ROS) and conjugated diene formation and improve paraoxonase 1 (PON1) expression and activity [33]. However, their neuroprotective, central anti-inflammatory impact and possible preventive effects against AD-related neuroinflammation have not been sufficiently studied. To the best of our knowledge, only a few published papers [34,35,36,37,38,39] have focused their research interest on investigating PP effects on neuroinflammation, and only one study has evaluated the impact of pomegranate urolithins, an ellagic acid derivative, on p-Tau [40]. These studies did not fully cover the subject, and many critical aspects of this topic still need to be investigated. This includes the effects of punicalagin and ellagic acid, a potent pomegranate-derived active molecules with remarkable antioxidant and anti-inflammatory properties, on Aβ-induced Tau-phosphorylation, as well as their impact on the phenotypic transition of microglia from M1 to M2-state. Therefore, a considerable need exists to cover this gap and explore pomegranate bioeffects on molecular and physiological processes leading to CNS degeneration.
2. Materials and Methods
2.1. Chemicals and Reagents
Tert butyl hydroperoxide (TBHP), 2-thiobarbituric acid (TBA), hydrochloric acid (HCL), butylated hydroxytoluene (BHT), radioimmunoprecipitation assay buffer (RIPA), dimethyl sulfoxide (DMSO) and 3-(4, 5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide (MTT), were purchased from Sigma Aldrich (ST Louis, MO, USA), except for lipopolysaccharide (LPS) and Elacridar which were from Sigma Aldrich (Canada, LTD). Trichloroacetic acid (TCA) and butanol were from Fisher Scientific (Loughborough, UK). Punicalagin and ellagic acid were purchased from Cayman Chemical (Michigan, USA). Human beta-amyloid-(1-42) was supplied by Abcam (Canada). Dichlorodihydrofluorescein diacetate (H2-DCFH-DA) was purchased from Invitrogen (Waltham, Massachusetts, USA). The Annexin V-FITC apoptosis detection assay kit was purchased from BioLegend. Pierce BCA protein assay kit was from Thermo Fisher Scientific (Waltham, MA USA). Ethanol, phorbol myristate acetate (PMA), and bovine serum albumin were from Wisent Inc (Saint-Jean-Baptiste, QC-Canada).
2.2. Plant Material
Moroccan pomegranate (Punica granatum L.) fruits of the Sefri variety were grown and harvested from a local farm in the Beni Mellal-Khenifra region-Oulad Abdallah area (Central Morocco; Latitude: 23°50′05″ E; Longitude: 6°48′98″ N). Professor Abbas Younes, a taxonomist and professor at Sultan Moulay Slimane University, confirmed the authenticity of our variety.
2.2.1. Pomegranate Polyphenols Extraction and Chemical Characterization
Peels and arils polyphenol extracts were prepared as previously described by Benchagra et al. [33]. Briefly, aril molasses and peel powder were subjected to extraction (methanol/water, 70: 30) for 48 h at 4°C. The hydroalcoholic extracts were centrifuged, filtered, lyophilised, and stored at -20°C until use. We have previously determined the chemical content (of total flavonoids, total anthocyanins, and total polyphenols) and specific polyphenols by HPLC analysis of arils and peels phenolic extracts [33].
2.3. Human Aβ1-42 Preparation
Human Aβ1-42 peptide was supplied by Abcam company (MW 4514.08; AB120301), and it was dissolved in DMSO (0.1%) and diluted in PBS to a final concentration of 1 mg/ml. After that, it was immediately sonicated for 15 min at room temperature and centrifuged at 15,000 × g at 4 ◦C for 20 min. As described, the supernatant was stored at −80◦C in 50 µL aliquots until use [41].
2.4. Cell Culture
Human H4 neuroglioma (ATCC HTB-148), HMC3 immortalised human microglial cell line (ATCC CRL-3304), and Human THP-1 monocytes (ATCC TIB-202) were purchased from the American Type Culture Collection (Manassas, VA, USA) via Cederlane® company (Burlington, ON, Canada). THP1 cells were differentiated into macrophages using 100 nM of phorbol myristate acetate (PMA) for 96 h. H4, HMC3, and THP-1 cell lines were cultured in Dulbecco’s modified Eagle’s medium (DMEM), Eagle's Minimum Essential Medium (EMEM), and Roswell Park Memorial Institute (RPMI) medium, respectively. The U373-MG human astrocytes cell line was kindly provided by Professor Denis Gris, University of Sherbrooke-Canada, and was grown in DMEM. All mediums were supplemented with 10% (v/v) heat-inactivated fetal bovine serum, 10 U/ml penicillin, and 10 μg/ml of streptomycin. The cells were maintained in a 5% CO2 humidified incubator at 37 °C.
2.5. Cell Viability Measurement
The effects of human Aβ1-42 and PP on cell growth were studied using the MTT test. Briefly, H4 neurons were seeded in 96-well plates (104 cells/well) and incubated for 24 h with 20 µg/ml of Aβ1-42, in the presence or not of PP. The medium was removed, and cells were incubated for two hours with MTT (0,5 mg/ml) (Sigma Aldrich-USA-CAS-No: 298-93-1). After washing, DMSO-dissolved formazan crystals formed, and the absorbance was recorded at 570 nm using a microplate reader (PerkinElmer, Canada) [42].
2.6. Cell Apoptosis Assay
Apoptotic cells were detected using the FITC annexin-V apoptosis detection kit (Biolegend; Cat: 640914) according to the manufacturer’s instructions. In brief, H4 neurons were incubated with or without 15 µg/ml of Aβ1-42 in the presence or absence of PP for 24h. All cells were collected, washed with PBS, and stained with 5 µl of FITC-conjugated annexin-V (20 °C, in the dark for 15 min). Then, cells received 400 µl of annexin-v binding buffer. Data was acquired using flow cytometry (Beckman Coulter, Brea, California, USA) and analysed by FlowJo 10.2 software (Tree Star Inc., Ashland, OR, USA).
2.7. Measurement of Intracellular ROS
Oxidative stress imbalance is one of the leading causes of various disorders and illnesses. In this assay, the intracellular ROS was measured using the fluorescent probe dichlorodihydrofluorescein diacetate (H2DCF-DA) (Invitrogen-USA; C400) as previously described [43]. Briefly, 2 x 105 cells (HMC3 microglial) were seeded in a 24-well plate in a complete EMEM medium for 24h to stabilise at 37 °C. The cells were then pre-treated with PP for two hours before their treatment with human Aβ1-42 for an additional four hours. The cells were washed and incubated with ten µM of DCFH-DA solution at 37°C in the dark for 30 min. After removing DCFH-DA and washing, protein analysis was done using BCA assay according to the manufacturer’s recommendations (Thermo Fisher Scientific). The DFC fluorescence intensity of the cells was measured at an excitation and emission wavelength of 485 nm and 530 nm, respectively, using a VICTOR Multilabel Plate Reader (PerkinElmer, Canada). Results were expressed as fluorescence emission intensity/mg of protein for each condition.
2.8. Lipid Peroxidation Assay
Lipids are the primary biological components targeted by ROS, leading to cellular damage and injury. In this assay, we tested the ability of our treatments to prevent lipid peroxidation. Briefly, HMC3 cells were seeded in 24 well-plates and were subjected to PP treatments at different concentrations for 24h before incubation with TBHP (Cat: 458139; Sigma Aldrich, USA) (200 µM) for one h. Cell-free supernatants were added to 300 µl of the following mixture (0,5% TBA, 30 % TCA, 0,33 M HCL, and 0,005 % of butylated hydroxytoluene) and boiled for 60 min. The samples were cooled, and 300 µl of the butanol was added to each vial for MDA-TBA extraction. After centrifugation, 100 µl of the butanol fraction was transferred to 96 healthy plates designed for fluorescence-based assays (PerkinElmer, Canada) to quantify the fluorescence intensity at excitation and emission wavelengths of 530 and 590 nm, respectively [44]. The obtained data were normalised using BCA assay (Thermo scientific-USA; Cat: 2322).
2.9. M1/M2 Polarization of Microglia Cells and Systemic Macrophages
We attempted to evaluate the ability of our treatments to push M1/M2 shifting in HMC3 and THP1 cells. We investigated the expression of CD86 and CD163 cell surface receptors as M1 and M2-specific markers, respectively. Briefly, cells were stimulated by LPS (1 µg/ml) (Sigma Aldrich-Canada; CAS-No: 93572-42-0) and cotreated simultaneously with pomegranate extracts (peels or arils) or with pomegranate-purified compounds (punicalagin or ellagic acid) at increasing concentrations for 24h. Then, cells were harvested and washed twice with cold PBS (1x) and centrifuged (350xg, at four °C, for 6 min). Then, they were stained for CD86 and CD163 cell surface receptor expression using FITC-mouse anti-human CD86 (1:200; BD-Biosciences-Cat: 555657) and BV711-mouse anti-human CD163 monoclonal antibodies (1:200; BD-Biosciences-Cat: 563889), for 50 min at four °C in darkness. Following this period, the samples were rewashed, centrifuged, and resuspended in cold PBS for cytometry analysis. Data were collected using a CytoFLEX instrument (Beckman Coulter, Brea, California, USA) and analysed by FlowJo 10.2 software (Tree Star Inc., Ashland, OR, USA).
2.10. The Assessment of Interleukin 1-Beta (IL-1β) and Interleukin-10 (IL-10) Proteins Expression
The U373-MG human astrocytes and THP-1 macrophages were subjected to intracellular staining to quantify the protein expression of IL-10 and IL-1β according to the manufacturer’s instructions (BD-Biosciences-Canada). Briefly, cells were stimulated by LPS (1 µg/ml) and cotreated simultaneously with increasing concentrations of PP. Cells were treated during the last four h of culture with protein transport inhibitor A (Brefeldin A: 1:1000; BD-Biosciences-Cat: ab51-2301KZ) to allow the intracytoplasmic accumulation of cytokines. Then, cells were fixed/permeabilised (1h at 4 °C, in darkness) using Fix/perm buffer (BD-Biosciences; Cat: 51-9008314). Cells were then stained with monoclonal APC-rat anti-human IL-10 (1:400; BD-Biosciences; Cat: 554707)) and PE-mouse anti-human IL-1β (1:200; BD-Biosciences; Cat: 340516) for 50 min at four °C, in darkness. Data was collected using a CytoFLEX instrument (Beckman Coulter, Brea, California, USA) and analysed by FlowJo 10.2 software (Tree Star Inc., Ashland, OR, USA).
2.11. Tau-Phosphorylation at Threonine 181
Human H4 neuroglioma cells were cultured in a 6-well plate at a concentration of 1 x 106 and grew to about 80% of confluence. All cells, except the control group, were stimulated with ten µg/ml of human Aβ1-42 and simultaneously cotreated or not with PP for 24h. Then, cells were fixed/permeabilised (1h at four °C, in darkness) using Fix/perm buffer (BD-Biosciences; Cat: 51-9008314) and stained with primary rabbit monoclonal anti-phospho-Tau181 antibody (1:500; Abcam-Canada; Cat: ab254409) for an additional one h. After washing, the unconjugated anti-phospho-Tau181 antibody was conjugated with Alexa-Fluor 647 anti-rabbit secondary antibody (1:200; Invitrogen; Cat: A-21244) in darkness for 45 min at four °C. Following this period, the samples were washed twice, centrifuged, and resuspended in cold PBS for cytometry analysis. Data were collected using a CytoFLEX instrument (Beckman Coulter, Brea, California, USA) and analysed by FlowJo 10.2 software (Tree Star Inc., Ashland, OR, USA).
2.12. Statistical Analysis
The statistical analysis was performed using version 10.2.3 of GraphPad Prism software (GraphPad Software®, Inc., La Jolla, CA, USA), and the obtained results were expressed as mean ± SEM. Differences between groups were analysed using a t-test (two groups) or one-way ANOVA with Dunnett's multiple comparisons test. The significance levels were: *p < 0.05, **p < 0.01, ***p < 0.001.
3. Results and Discussion
3.1. Phytochemical Analysis
Our previously published results have demonstrated that peels are richer than arils extracts in total phenolic and flavonoid compounds, where no significant difference was observed in anthocyanin content. Moreover, HPLC analysis showed that both extracts are especially rich in α-punicalagin and β-punicalagin but also contain ellagic and gallic acids, with a higher proportion in peels than arils extracts [33].
3.2. Cell Viability Measurement
We first investigated the cytotoxic effect of human Aβ1-42 by measuring the metabolic activity of H4 neurons and, subsequently, the cytoprotective effects of PP. The used concentration was fixed upon results obtained from a dose-dependent experiment that we carried out with increased concentrations of Aβ1-42 (5 to 20 µg/ml, supplementary data, figure 1S). The obtained data show that Aβ1-42 treatments resulted in a significant (p<0.0001) decrease in cell viability in all the used concentrations (5 to 20 µg/ml), with considerable cytotoxicity at 20 µg/ml. Therefore, we tested the capability of our treatments to improve neuronal viability in the presence of this concentration (20 µg/ml). Our results in Figure 1A show that human Aβ1-42 induce a significant decrease in neuronal viability (p<0.01). However, the treatment with PP significantly attenuates this cytotoxic effect (Figure 1B). As illustrated in Figure 1B, ellagic acid and pomegranate peels polyphenols, but not punicalagin and pomegranate arils treatments, have attenuated the cytotoxic effects of human Aβ1-42 on H4 neurons. Peel treatment appears to be more potent in protecting cells against Aβ1-42.
On the other hand, no cytotoxic effect was observed under PP treatments when used alone (data not shown). Previously published reports agree with these findings on PP's ability to improve cell viability. Data from Dasilva et al., [39], showed that pomegranate urolithins, a derivative compound of ellagic acid, can improve SH-SY5Y viability and attenuate LPS-induced toxicity. Moreover, pomegranate peel extract increased cell viability and showed neuroprotective effects against cisplatin-induced neurotoxicity in a dose-depending manner [34,45]. Furthermore, punicalagin was shown to suppress LPS-mediated cytotoxicity in BV2 microglial cells [45]. All these findings support the improvement of PP's safety and cellular viability.
Arils extract and punicalagin treatments, which have no significant effect (p>0.05) on Aβ1-42-mediated neuronal death (Figure 1B), were combined with Elacridar, a third generation of P-glycoprotein (P-gp) inhibitor and potent bioenhancer that targets multiple drug resistance, to improve outcomes potentially. It has been shown that P-gp pomp plays a vital role in drug resistance [46], and its pharmacological inhibition has emerged as a promising therapeutical approach to improve intracellular drug deposition [47]. Many studies have attributed Elacridar's significant capacities in modulating drug resistance involving efflux transporters and substantial effects in enhancing the intracellular bio-disponibility of many anticancer agents [48,49]. We hypothesised that the effects of PP, particularly arils extract and punicalagin, which do not significantly impact Aβ1-42-mediated neuronal death, could be potentiated by their combination with Elacridar. Indeed, this intervention significantly (p<0.01) improved neuronal viability (Figure 2), which was reflected by the enhancement of cellular respiration and metabolic activity. Arils + Elacridar treatments significantly attenuated the cytotoxic effect Aβ1-42, suggesting a decisive synergistic action between pomegranate arils extract polyphenols and Elacridar. Similarly, combining punicalagin with Elacridar significantly enhanced the protective effect of punicalagin against Aβ1-42-induced neuronal death (Figure 2). Therefore, co-therapy using natural molecules and Elacridar could be envisaged to potentiate their biological effects.
To confirm our findings regarding the viability improvement effect of our treatments and to validate the neurotoxic impact of Aβ1-42 against H4 neurons, we performed an annexin-V assay to measure neuronal apoptosis.
3.3. Human Aβ1-42 Induces Apoptosis of H4 Neurons
Cell apoptosis is an evolutionarily conserved process and a cellular response to harmful stimuli. Our results indicate that Aβ1-42 significantly (p<0.05) increases neuronal apoptosis. These results align with those published by Brouillette et al. [50] and Cizas et al. [51], who found that the repeated neuronal exposition to Aβ1-42 induces neuronal loss. Additionally, Aβ1-42 has been suggested to trigger neurodegeneration via age-dependent autophagic-lysosomal damage [52] and was correlated with brain atrophy in many scientific reports [53,54,55]. This neuronal toxicity was significantly (p<0.001) attenuated by PP (peels, arils, punicalagin and ellagic acid) (Figure 3), suggesting an anti-apoptotic effect of PP against cellular degeneration caused by Aβ1-42. Ellagic acid was more effective in reducing apoptosis than the peels and punicalagin, while no significant difference was observed between ellagic acid and arils (p>0.05). Peels and punicalagin showed comparable effects (p>0.05). Interestingly, this confirms and supports the reported MTT results in Figure 1. From a mechanistic perspective, ellagic acid can trigger and alter intrinsic apoptotic pathways by inhibiting caspase 3 and 9 , serine-aspartyl proteases essential in triggering programmed cellular death, to increase neuronal viability. Similarly, it has been shown [45] that pomegranate peels phenolic-rich extract reduce caspase-3 and caspase nine mRNA transcription, and other potent pomegranate flavonoids such as quercetin and pomegranate non-flavonoids including resveratrol, also exert neuroprotective effects against neuronal apoptosis [57]. The anti-apoptotic effect of punicalagin was reported by El-Missiry et al., [58] who indicated that punicalagin reduces Bax/Bcl2 and caspase 3 and 9. We can suggest pomegranate as an effective strategy and a complementary medicine to limit neuronal degeneration.
3.4. Measurement of intracellular ROS
Excessive generation of reactive oxygen species (ROS) induces pathological changes in susceptible biomolecules such as lipids, proteins, and DNA. Oxidative modifications in the structure/function of these vital molecules may cause a series of redox-related diseases, including neurological disorders [59]. Therefore, maintaining the balance between pro/antioxidants is fundamental for cellular homeostasis and disease prevention. We attempted to investigate the capability of the human Aβ1-42 to induce oxidative stress in microglial cells. The results showed that peels at 400 µg/ml can significantly (p<0.05) reduce the Aβ1-42-induced oxidative stress in HMC3 microglia (Figure 4) in a dose-depending manner (Figure 4A.S). The peel is well known for its antioxidant properties, and the reported effect may result from synergistic effects between the potent antioxidant polyphenols in the peel extract, such as hydrolysable tannins and flavonoids. Mechanistically, antioxidant molecules can use their free hydroxyl groups attached to aromatic rings to reduce oxidative species through hydrogen atom transfer reactions. They can also alleviate oxidative damage by acting as chelating agents [60] or as electron-donating compounds [61]. The active phytochemicals of pomegranate, especially the polyphenols extracted from the peels, have the potential for multi-targeted effects, limiting proteins, DNA, and lipid damage. Aril treatments showed a significant trend (p<0.05) to attenuate the induced oxidative stress (Figure 4B). This tendency varies significantly (p<0.05) in a concentration-dependent way (Figure 4B.S). Punicalagin polyphenols were able to reduce ROS generation only at 100 µg/ml (Figure 4C) and do not follow a dose-response model (p>0.05) (Figure 4C.S). Under the present conditions, ellagic acid polyphenols were not significantly (p>0.05) effective in reducing the oxidative stress induced by Aβ1-42.
3.5. Lipid Peroxidation Assay
Brain lipids contribute to cellular energy capital and essential intermediate effectors in cell signalling pathways [62]. Lipids are also a structural component of cell membranes and contribute to cell architecture. They are a particular target of ROS, and the oxidative modifications of lipids can induce numerous dyshomeostasis in the CNS [63]. In this experiment, we pretreated the HMC3 microglial cells with pomegranate treatments for 24h, followed by TBHP stimulation for one h. As illustrated in Figure 5, pomegranate polyphenols were able to significantly (p<0.05) reduce lipid peroxidation in a concentration-depending manner (Figure 5.S). Peels polyphenols, at 100, 200 and 400 µg/ml, attenuate significantly (p<0.001) the pro-oxidative effect of TBHP in a dose-depending manner (Figure 5A.S). Arils and punicalagin polyphenols showed a similar trend and were significantly (p<0.05 and p<0.01; respectively) effective in decreasing the provoked oxidative stress. The linear regression analysis indicates that this reduction follows significantly (p<0.05) a dose-response trend (Figure 5B.S,C.S).
Ellagic acid was only significantly (p<0.05) effective at higher concentrations (100 µM). The peels contain a mixture of powerful antioxidant polyphenolic compounds such as punicalagin-β and punicalagin-α [60], cyanidin 3-glucoside, pelargonidin 3-glucoside, cyanidin 3,5-diglucoside [64], quercetin [65], which are known for their antioxidant activities. In vivo evidence, achieved by Morzelle et al., [66] demonstrated that pomegranate peel extract can reduce lipid peroxidation in male C57BI/6 mice. Similarly, Kim et al. [34] reported that four weeks of administration of 1.5 mg/kg/day of punicalagin polyphenol to male ICR mice decreased the hydrogen peroxide and MDA levels and increased the GSH/GSSG ratio. These results are comparable to those obtained by Yaidikar et al. [67], who reported that pomegranate downregulates the malondialdehyde level. The preventive role of pomegranate polyphenols against neurodegeneration could be due to its corrective effect of redox imbalance regarding the richness of pomegranate in antioxidant molecules.
3.6. M1/M2 Polarization of Microglia and Systemic Macrophages
To exert their protective effects both at the peripheric and central level, pomegranate polyphenols should reach the bloodstream and cross the brain-blood barrier (BBB). Previous scientific investigations showed the capability of pomegranate polyphenols, especially punicalagin and ellagic acid, to get human blood. According to Cerdá et al., [68] punicalagin compounds have been detected in the plasma of Sprague-Dawley rats at concentrations (C-max) around 30 µg/ml. Similarly, ellagic acid was detected at a concentration (C max) of 31.9 ng/ml in human plasma after consuming 180 ml of pomegranate juice [69]. Furthermore, Serdar et al. [70] have demonstrated that punicalagin and ellagic acid can cross an in vitro BBB model. Similarly, in vivo evaluation of BBB penetrability of ellagic acid indicated its capacity to achieve the neuronal compartment 0.5–4 h after a dose of 50 mg/kg orally [71].
Neuroinflammation mediated by the M1 phenotype of microglia is crucial in the neurodegenerative process. Therefore, we attempted to evaluate our treatments' capability to drive the move of macrophages from an M1 pro-inflammatory/destructive phenotype to an M2 anti-inflammatory/protective state. Chronic M1 phenotype is associated with increased release of pro-inflammatory cytokines and can induce neuronal damage and neurotoxicity. The M2 phenotype is more likely to be associated with increased expression of anti-inflammatory markers and is thought to support neuronal survival and cellular homeostasis. We reported that LPS significantly (p<0.05) upregulate CD86 expression in HMC3 microglia cells (Figure 6.1). This finding agrees with results obtained by Lu et al., [72] who reported around a 29% increase in CD86 expression in HMC3. In our experiment, pomegranate treatments showed a trend to mitigate M1-related neurotoxicity by decreasing the expression of the cell surface receptor CD86 and increasing CD163 protein expression (Figure 6). Peels rich-phenolic extract dose-dependently reduced the CD86 expression (p<0.001) (Figure 6.1A and Figure 6.1A.S). The peel treatment was also dose-dependently (p<0.0001) (Figure 6.2A.S) effective in improving M2-phenotype through CD163 up-regulation (Figure 6.2A). Peel extract contains a mixture of polyphenolic constituents recognised to drive anti-inflammatory pharmacological activities, such as ellagitannins bioactive compounds [73]. We have demonstrated that arils phenolic extract also modulated LPS-induced neuroinflammation (Figure 6 and Figure 6.S). However, arils appear less potent than peels in modulating microglia's M1/M2 phenotypic transition. The previous publication by our team [74] could explain this difference between peels and arils extracts. Our team previously demonstrated that peels are more prosperous in antioxidant and anti-inflammatory compounds such as punicalagin polyphenols than aril extract [74].
Similarly, the incubation of cells with punicalagin resulted in a significant (p<0.0001) shift of microglial HMC3 cells from M1 to M2-phenotype via the downregulation of CD86 (Figure 6.1C) and upregulation of CD163 (Figure 6.2C) cell surface receptors expression. The linear regression analysis suggests the mentioned effect of punicalagin polyphenols follows a concentration-depending model (Figure 6.S). Ellagic acid is a natural antioxidant polyphenol and a potent punicalagin metabolite that gives rise to the anti-inflammatory urolithin constituents [75], and it was reported to drive various neuroprotective activities [76]. In our hand, we demonstrated that ellagic acid can modulate the pro-inflammatory state of microglia by decreasing (in a non-dose-response manner, (p>0.05); Figure 6.1.S) the expression of CD 86 (Figure 6.1D) and increasing in concentration-depending manner (Figure 6.2.S) the one of CD163 (Figure 6.2D).
Only one previously published study [77] attributed comparable effects to cyanidin-3-O-glucoside, an anthocyanin compound found in many vegetables and fruits, including pomegranate. According to the authors, cyanidin-3-O-glucoside could shift the M1-phenotype of HMC3 microglial cells to the M2-protective phenotype, as it can decrease the expression CD86 and CD80, inflammatory cytokines (IL-1β, IL-6, TNF-α), and oxidative stress, as well as increase M2-specific markers, particularly the CD206 and CD163. These results were confirmed by the same authors under in vivo conditions, using APPswe/PS1ΔE9 mice. However, there are no previous reports on the modulatory effect of punicalagin, ellagic acid, pomegranate peels and arils polyphenols on microglia (M1/M2) polarisation.
Systemic chronic inflammation is involved in the aetiology of many incurable diseases [78,79], including atherosclerosis. In the present paper, we first investigated the response of human systemic THP-1 macrophages to LPS stimulation. Secondly, we attempted to modulate the resulting M1-phenotype by using increasing concentrations of the used PP. Our data suggest that LPS significantly (p<0.05) enhance the expression of CD86 in THP-1 macrophages and, therefore, corroborate those previously published by Hennen et al. [80]. Results showed in Figure 7.1A indicates that peels significantly (p<0.001) and dose-dependently (p<0.0001) (Figure 7.1A.S) downregulated CD86 receptor expression. This agrees with the data from Xin-Yu Lu and coworkers , who reported that pomegranate peel extract can reduce inflammation by reducing the percentages and absolute numbers of CD80+ and CD86+ cells. Pomegranate peels are rich in hydrolysable tannins and various gallic acid esters, widely recognised for mediating anti-inflammatory activities [82,83,84,85]. The arils-rich phenolic extract was also significantly effective in reducing CD86 expression, notably at a concentration of 200 µg/ml (p<0.001) (Figure 7.1B). This effect follows a dose-response relationship (p<0.0001) (Figure 7.1B.S). Arils contain various active substances, including flavonoids and anthocyanins [86]. These active phytochemicals are endowed with anti-inflammatory properties [87,88]. In addition, punicalagin significantly (p<0.001) reversed the M2 to M1 phenotypic transition of THP-1 macrophages induced by LPS stimulation. Punicalagin reduced CD86 expression (Figure 7.1C) in a dose-depending manner (p<0.01) (Figure 7.1C.S). Punicalagin treatments were reported to promote M2-like macrophage polarisation via up-regulation of HO-1 in murine macrophages [89] . They could be a preventive strategy against inflammatory disorders. Furthermore, ellagic acid significantly (p<0.01) attenuated LPS-induced M1 phenotype in THP1 macrophages and downregulated in a concentration-depending manner (p<0.01) (Figure 7.1D.S) CD86 protein expression (Figure 7.1D). Recent in vivo research [90] has revealed similar findings, suggesting that ellagic acid at 50 and 100 µM attenuated LPS-induced neuroinflammation. These effects may be exerted directly by ellagic acid or indirectly by its metabolites, including urolithin A, which can achieve neuronal microenvironment [91].
The effect of our treatments on CD163 expression is illustrated in Figure 7.2. The reported findings indicate a potent modulation of the THP-1 macrophage state. Incubation of cells with LPS resulted in a significant (p<0.05) decrease in CD163 cell surface receptors (Figure 7.2). This downregulation was corrected by peels polyphenols, which significantly (p<0.001) increased CD163 receptors (Figure 7.2A) in a concentration-depending manner (Figure 7.2A.S; p<0.0001). Arils treatments, especially at 400 µg/ml, remarkably and positively shifted the expression of CD163 in a concentration-depending way (p<0.01) (Figure 7.2B,B.S). A similar trend was observed with punicalagin polyphenols, which upregulated in a dose-depending manner (Figure 7.2C.S; p<0.0001) CD163 protein expression, and the most potent effect was achieved at 100 µg/ml (Figure 7.2C). On the other hand, it appears that ellagic acid at 25 and 50 µM was not able to significantly improve CD163 M2 markers (p>0.05), while at higher concentrations, its effect was significant (100 µM; p<0.05) (Figure 7.2D). Comparable findings were obtained also by Aharoni et al.,[92] using a J774-A1 macrophage-like cell line. The authors of this study reported that punicalagin and pomegranate juice polyphenols could promote macrophage switch to an anti-inflammatory M2 response [92]. Furthermore, pomegranate juice can limit mice's age-associated switch from M2 to M1 [92].
The reported findings and the discussed literature point to a significant protective transition from M1 to M2 polarisation mediated by PP at central and peripheric levels. These add additional support to the central role of polyphenols from natural resources in contracting significant inflammatory alterations that occur in atherosclerosis and AD.
3.7. The Assessment of Interleukin 1-beta (IL-1β) and Interleukin-10 (IL-10) Cytokines Expression
Inflammation is a crucial mechanism of innate immunity that aims to guide, enhance, and accelerate the healing process. However, chronic inflammation is harmful to neurons, and it can lead to irreversible degeneration, causing a progressive decline in brain vital functions. Various scans of the AD brain showed a high level of inflammatory markers, and numerous genome-wide association studies suggested that several immune-related loci can increase the susceptibility to AD [93,94]. On the other hand, many nonsteroidal anti-inflammatory agents (NSAIDs) can reduce the risk of developing AD [95,96]. This evidence, among others, points to the potential involvement of the immune system in the aetiology of AD. The paper evaluated the possible inflammatory lowering effect of pomegranate peels, arils, punicalagin, and ellagic acid in U373-MG human astrocytes and THP1-monocytes differentiated into systemic macrophages. Flow cytometry quantified the expression of proinflammatory cytokines (Il-1β) and anti-inflammatory markers (IL-10).
In U373-MG astrocytes, LPS (1 µg/ml) significantly (p<0.001) reduced the expression of IL-10 (Figure 8). These proinflammatory effects were attenuated in a dose-depending manner by peels (100-200 and 400 µg/ml, p<0.01) (Figure 8A and Figure 8A.S) and by arils treatments (Figure 8B and Figure 8B.S). Similarly, punicalagin (Figure 8C) significantly (25-50 and 100 µg/ml, p<0.01) reduced in a dose-depending manner (Figure 8C.S; p<0.05) the cytotoxicity of LPS-induced neuroinflammation and significantly increased the expression of IL-10 cytokines in human astrocytes cells. Furthermore, ellagic acid treatment upregulated (Figure 8 D) dose-dependently (Figure 8D.S; p<0.0001) IL-10 expression. Previous in vivo studies reported PP as a potent glial inflammatory process regulator. Findings obtained by [66] and by [38] suggested that peel-rich-phenolic extract can reduce TNF-α and Il-1β expression both in C57BI/6 and in APPsw/Tg2576 mice. Similarly, punicalagin suppresses Il-1β, IL-6, and TNF-α in male ICR mice [34]. Moreover, a short-term intervention (14 days) using urolithins A, an ellagic acid derivative, significantly downregulated gene expression of IL-1β, IL-6, and TNF-α in a transgenic female model that expressed APP/PS1 mutation [97]. From a mechanistic point of view, PP can act through IKK and Ikb inhibition [34,35], decrease NF-kB DNA binding activity, and reduce p50 and p65 subunit translocation [34,35].
We also found that pomegranate treatments can alleviate LPS-induced systemic inflammation in human THP1-monocytes differentiated into macrophages. Indeed, LPS stimulation (1µg/ml) resulted in a significant (p<0.001) decrease in IL-10 cytokine expression (Figure 9.1). The induced inflammation was significantly (p<0.001) ameliorated by peels and arils (Figure 9.1A and B; respectively) as well as drastically suppressed (p<0.001) in a concentration-depending manner by punicalagin (p<0.0001) (Figure 9.1C.S) and ellagic acid (p<0.05) (Figure 9.1D and figure 9.1D.S). In the same sense, the incubation of THP-1 macrophages with LPS resulted in a significant (p<0.05) increase of IL-1β proinflammatory cytokines in comparison to the untreated group (Figure 9.2). Polyphenols from pomegranate approximatively suppressed the proinflammatory effects of LPS. Peels, arils and punicalagin approximatively returned (p<0.05) (Figure 9.2A–C; respectively) in a dose-depending manner (p<0.05) THP-1 macrophages to the basal state (Figure 9.2A.S–C.S; respectively), while ellagic acid reduced in a very remarkable way the expression of IL-1β especially at 100 µM (Figure 9.2D).
Our data joins the previously published scientific reports [98,99] on the anti-inflammatory effects mediated by pomegranate polyphenols. Indeed, it has been reported that punicalagin from a pomegranate can inhibit NF-κB and MAPK activation in response to LPS stimulation in RAW264.7 macrophages, as well as enhanced LC3II and p62 protein expression [98]. Furthermore, a study published by Du et al. [99] attributed peel extract suppressive activities against the TLR4/NF-κB pathway. Preventive and corrective effects of pomegranate against other neurodegenerative diseases are debated as Tapias et al. [100] reported essential and unexpected results in a rotenone model of Parkinson's disease. The authors showed adverse effects of PP, such as dopaminergic neuronal death, aggravation of the inflammatory response, and caspase activity enhancement. This may be due to the high concentration used, as polyphenols are recognised to promote proinflammatory pathways at high doses [101]. However, the reported unexpected findings have not been confirmed in a recent study published by Kujawska et al. [102], who suggested opposite findings. To our knowledge, no published data exists regarding the effect of ellagic acid and arils on the investigated cytokines. Also, human findings related to the modulatory effect of PP on neuroinflammation are not available. This shows the deep gap and the vast need to examine the reported in vitro and the discussed in vivo beneficial effects on human beings.
3.8. Tau-Phosphorylation at Threonine 181
Various scientific investigations have associated the abnormal phosphorylation of Tau-protein with AD development [103,104,105]. From a mechanical point of view, Tau-phosphorylation can be involved in neurodegeneration via the alteration of Tau-related physiological functions such as microtubule-binding activities [106] and neurite outgrowth [107,108]. In addition, much clinical evidence has found an elevated hyperphosphorylated Tau in the brain of patients suffering from AD [109,110] , and this was correlated with cognitive decline in several published papers [111,112]. On our hand, we performed the flow cytometry analysis first to examine whether human Aβ1-42 can induce the phosphorylation of Tau-protein at threonine 181 and secondary to evaluate the modulatory impact of our treatments against Aβ1-42-induced Tau-phosphorylation. We report that incubating H4 neurons with human Aβ1-42 resulted in a significant (p<0.001) increase of pTau-181. Previous analysis reported that Aβ1-42 stimulation significantly increased the phosphorylation of Tau (by 1.8-fold) [113]. Our intervention using PP significantly suppressed (p<0.0001) the Aβ1-42-stimulating effect on Tau-phosphorylation (Figure 10). Pomegranate peels and ellagic acid appear to be more potent than the other treatments (p<0.05), followed by punicalagin and arils (p<0.05) polyphenols. It has been shown that ellagic acid can ameliorate learning and memory impairments by inhibiting Aβ and Tau-phosphorylation [114]. Our findings could explain this effect, and we can suspect the indirect impact of ellagic acid on p-Tau via the reduction of Aβ-production. Also, possible direct interactions between PP, in particularly ellagic acid and Tau-kinases/phosphatases, including glycogen-synthase kinase-3β (GSK3β), cyclin-dependent kinase 5 (CdK5), and cAMP-dependent protein kinase (PKA) [115,116], could explain this dynamic regulation of Tau. In all, natural substances and diets rich in polyphenols can positively impact limiting AD-associated Tau-pathology.
4. Conclusion and perspectives
The current study provides direct evidence to support the beneficial effects of PP in Tau pathology against oxidative stress and neuroinflammation-mediated neurodegeneration. The pharmacotherapeutic potential of pomegranate should be deeply evaluated in future investigations, particularly in AD-relevant models and through well-designed randomised placebo-controlled trials. Future work should also determine the most effective and safest dose and the potent bioactive compound regarding the richness of pomegranate in several other active ingredients. In this context, we suggest, as a perspective, to investigate the gut-based (microbiome) polyphenol metabolites of ellagic acid, especially the urolithins constituents, as they showed higher blood-brain barrier penetrability and a diverse array of neuroprotective actions with multi-targeted physiological effects.
Author Contributions
Conceptualization, H.B. and M.A; methodology, H.B., K.B., M.A., Z.E.; software, H.B., M.A. and K.B; validation, H.B., T.F. and A.K.; formal analysis, H.B., S.B., C.R. and M.A.; investigation, H.B., T.F and M.A.; resources, H.B., S.B., T.F, T.B; writing—original draft preparation, M.A., H.B., A.K. and S.B.; visualization.; B.L., J.M.W., T.F.; A.K., N.Z. and H.B. supervision, project administration and funding acquisition, H.B., T.F. and A.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Agence Nationale des Plantes Médicinales et Aromatiques), (Centre National de Recherche Scientifique et Technique) and (Université Sultan Moulay Slimane), Morocco. 2020–2023 (H.B.) and by Canadian Institutes of Health Research (grant Number # PJT-162366) (A.K.).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and the protocol was approved by the Ethics Committee of the Sherbrooke University Institute of Geriatrics (# 2009/19). Informed consent was obtained from all subjects involved in the study.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are contained within the article.
Acknowledgments
work partially supported by an unrestricted fund provided by Izumi Bioscience, the Research Centre on Aging and the Alzheimer Foundation.
Conflicts of Interest
The authors declare no conflict of interest. The company (Izumi Bioscience) had no role in the design of the study; in the collection, analysis, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Rogers, J.; Luber-Narod, J.; Styren, S.D.; Civin, W.H. Expression of Immune System-Associated Antigens by Cells of the Human Central Nervous System: Relationship to the Pathology of Alzheimer’s Disease. Neurobiol. Aging 1988, 9, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Fiacco, T.A.; Carthy, K.E.N.D.M.C. Astrocyte Calcium Elevations : Properties , Propagation , and Effects on Brain Signaling. 2006, 690, 676–690. [CrossRef]
- Floyd, R.A.; Hensley, K. Nitrone Inhibition of Age-Associated. 222–237.
- Ventura, R.; Harris, K.M. Three-Dimensional Relationships between Hippocampal Synapses and Astrocytes. 1999, 19, 6897–6906.
- Boyarsky, G.; Schlue, B.R.W.; Davis, M.B.E.; Boron, W.F. Intracellular PH Regulation in Single Cultured Astrocytes From Rat Forebrain. 1993, 248.
- Drukarch, B.; Schepens, E.; Stoof, J.C.; Langeveld, C.H.; Van Muiswinkel, F.L. Astrocyte-Enhanced Neuronal Survival Is Mediated by Scavenging of Extracellular Reactive Oxygen Species. Free Radic. Biol. Med. 1998, 25, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Rostami, J.; Mothes, T.; Kolahdouzan, M.; Eriksson, O.; Moslem, M.; Bergström, J.; Ingelsson, M.; O’Callaghan, P.; Healy, L.M.; Falk, A.; et al. Crosstalk between Astrocytes and Microglia Results in Increased Degradation of α-Synuclein and Amyloid-β Aggregates. J. Neuroinflammation 2021, 18, 1–20. [Google Scholar] [CrossRef]
- Bilimoria, P.M.; Stevens, B. Microglia Function during Brain Development: New Insights from Animal Models. Brain Res. 2015, 1617, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.H.; Lee, D.Y.; Park, K.W.; Kim, S.U.; Yang, M.S.; Joe, E.H.; Jin, B.K. Microglia Expressing Interleukin-13 Undergo Cell Death and Contribute to Neuronal Survival in Vivo. Glia 2004, 46, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Aparicio, I.; Paris, I.; Sierra-Torre, V.; Plaza-Zabala, A.; Rodríguez-Iglesias, N.; Márquez-Ropero, M.; Beccari, S.; Huguet, P.; Abiega, O.; Alberdi, E.; et al. Microglia Actively Remodel Adult Hippocampal Neurogenesis through the Phagocytosis Secretome. J. Neurosci. 2020, 40, 1453–1482. [Google Scholar] [CrossRef] [PubMed]
- Meda, L.; Cassatella, M.A.; Szendrei, G.I.; Otvos, L.; Baron, P.; Villalba, M.; Ferrari, D.; Rossi, F. Activation of Microglial Cells by β-Amyloid Protein and Interferon-γ. Nature 1995, 374, 647–650. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Ding, M.; Xue, J.; Leng, W. The Role of TLR2/JNK/NF-ΚB Pathway in Amyloid β Peptide-Induced Inflammatory Response in Mouse NG108-15 Neural Cells. Int. Immunopharmacol. 2013, 17, 880–884. [Google Scholar] [CrossRef]
- Trojan, E.; Tylek, K.; Schröder, N.; Kahl, I.; Brandenburg, L.O.; Mastromarino, M.; Leopoldo, M.; Basta-Kaim, A.; Lacivita, E. The N-Formyl Peptide Receptor 2 (FPR2) Agonist MR-39 Improves Ex Vivo and In Vivo Amyloid Beta (1–42)-Induced Neuroinflammation in Mouse Models of Alzheimer’s Disease. Mol. Neurobiol. 2021, 58, 6203–6221. [Google Scholar] [CrossRef]
- Choi, J.Y.; Yeo, I.J.; Kim, K.C.; Choi, W.R.; Jung, J.K.; Han, S.B.; Hong, J.T. K284-6111 Prevents the Amyloid Beta-Induced Neuroinflammation and Impairment of Recognition Memory through Inhibition of NF-ΚB-Mediated CHI3L1 Expression. J. Neuroinflammation 2018, 15, 1–13. [Google Scholar] [CrossRef]
- Ismail, R.; Parbo, P.; Madsen, L.S.; Hansen, A.K.; Hansen, K. V.; Schaldemose, J.L.; Kjeldsen, P.L.; Stokholm, M.G.; Gottrup, H.; Eskildsen, S.F.; et al. The Relationships between Neuroinflammation, Beta-Amyloid and Tau Deposition in Alzheimer’s Disease: A Longitudinal PET Study. J. Neuroinflammation 2020, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cheignon, C.; Tomas, M.; Bonnefont-Rousselot, D.; Faller, P.; Hureau, C.; Collin, F. Oxidative Stress and the Amyloid Beta Peptide in Alzheimer’s Disease. Redox Biol. 2018, 14, 450–464. [Google Scholar] [CrossRef] [PubMed]
- Farkas, I.G.; Czigner, A.; Farkas, E.; Dobó, E.; Soós, K.; Penke, B.; Endrész, V.; Mihály, A. Beta-Amyloid Peptide-Induced Blood-Brain Barrier Disruption Facilitates T-Cell Entry into the Rat Brain. Acta Histochem. 2003, 105, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Selznick, L.A.; Zheng, T.S.; Flavell, R.A.; Rakic, P.; Roth, K.A. Amyloid Beta-Induced Neuronal Death Is Bax-Dependent but Caspase- Independent. J. Neuropathol. Exp. Neurol. 2000, 59, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Shepardson, N.; Yang, T.; Chen, G.; Walsh, D.; Selkoe, D.J. Soluble Amyloid β-Protein Dimers Isolated from Alzheimer Cortex Directly Induce Tau Hyperphosphorylation and Neuritic Degeneration. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 5819–5824. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Akama, K.T.; Krafft, G.A.; Chromy, B.A.; Van Eldik, L.J. Amyloid-β Peptide Activates Cultured Astrocytes: Morphological Alterations, Cytokine Induction and Nitric Oxide Release. Brain Res. 1998, 785, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Hanji, M.B. Meta-Analysis of Observational Studies. Meta-Analysis Psychiatry Res. 2017, 327, 111–121. [Google Scholar] [CrossRef]
- Szekely, C.A.; Zandi, P.P. Non-Steroidal Anti-Inflammatory Drugs and Alzheimers Disease: The Epidemiological Evidence. CNS Neurol. Disord. - Drug Targets 2012, 9, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Szekely, C.A.; Thorne, J.E.; Zandi, P.P.; Ek, M.; Messias, E.; Breitner, J.C.S.; Goodman, S.N. Nonsteroidal Anti-Inflammatory Drugs for the Prevention of Alzheimer’s Disease: A Systematic Review. Neuroepidemiology 2004, 23, 159–169. [Google Scholar] [CrossRef]
- Yan, Q.; Zhang, J.; Liu, H.; Babu-khan, S.; Vassar, R.; Biere, A.L.; Citron, M.; Landreth, G. Anti-Inflammatory Drug Therapy Alters  -Amyloid Processing and Deposition in an Animal Model of Alzheimer ’ s Disease. 2003, 23, 7504–7509.
- Sun, Y.; Koyama, Y.; Shimada, S. Inflammation From Peripheral Organs to the Brain : How Does Systemic Inflammation Cause Neuroinflammation ? 2022, 14, 1–10. [CrossRef]
- Walker, K.A.; Page, L.M. Le; Terrando, N.; Duggan, M.R.; Heneka, M.T.; Bettcher, B.M. The Role of Peripheral Inflammatory Insults in Alzheimer ’ s Disease : A Review and Research Roadmap. Mol. Neurodegener. 2023, 1–19. [Google Scholar] [CrossRef]
- Huang, X.; Hussain, B.; Chang, J. Peripheral Inflammation and Blood–Brain Barrier Disruption: Effects and Mechanisms. CNS Neurosci. Ther. 2021, 27, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Tansey, M.G.; Goldberg, M.S. Neuroinflammation in Parkinson’s Disease: Its Role in Neuronal Death and Implications for Therapeutic Intervention. Neurobiol. Dis. 2010, 37, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Mander, P.; Brown, G.C. Activation of Microglial NADPH Oxidase Is Synergistic with Glial INOS Expression in Inducing Neuronal Death: A Dual-Key Mechanism of Inflammatory Neurodegeneration. J. Neuroinflammation 2005, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Danesi, F.; Ferguson, L.R. Could Pomegranate Juice Help in the Control of Inflammatory Diseases? Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Rosillo, M.A.; Sánchez-Hidalgo, M.; Cárdeno, A.; Aparicio-Soto, M.; Sánchez-Fidalgo, S.; Villegas, I.; De La Lastra, C.A. Dietary Supplementation of an Ellagic Acid-Enriched Pomegranate Extract Attenuates Chronic Colonic Inflammation in Rats. Pharmacol. Res. 2012, 66, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Doostkam, A.; Bassiri-Jahromi, S.; Iravani, K. Punica Granatum with Multiple Effects in Chronic Diseases. Int. J. Fruit Sci. 2020, 20, 471–494. [Google Scholar] [CrossRef]
- Benchagra, L.; Berrougui, H.; Islam, M.O.; Ramchoun, M.; Boulbaroud, S.; Hajjaji, A.; Fulop, T.; Ferretti, G.; Khalil, A. Antioxidant Effect of Moroccan Pomegranate (Punica Granatum L. Sefri Variety) Extracts Rich in Punicalagin against the Oxidative Stress Process. Foods 2021, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.E.; Hwang, C.J.; Lee, H.P.; Kim, C.S.; Son, D.J.; Ham, Y.W.; Hellström, M.; Han, S.B.; Kim, H.S.; Park, E.K.; et al. Inhibitory Effect of Punicalagin on Lipopolysaccharide-Induced Neuroinflammation, Oxidative Stress and Memory Impairment via Inhibition of Nuclear Factor-KappaB. Neuropharmacology 2017, 117, 21–32. [Google Scholar] [CrossRef]
- Velagapudi, R.; Baco, G.; Khela, S.; Okorji, U.; Olajide, O. Pomegranate Inhibits Neuroinflammation and Amyloidogenesis in IL-1β-Stimulated SK-N-SH Cells. Eur. J. Nutr. 2016, 55, 1653–1660. [Google Scholar] [CrossRef]
- Rojanathammanee, L.; Puig, K.L.; Combs, C.K. Pomegranate Polyphenols and Extract Inhibit Nuclear Factor of Activated T-Cell Activity and Microglial Activation Invitroand Inatransgenic Mouse Model of Alzheimer Disease. J. Nutr. 2013, 143, 597–605. [Google Scholar] [CrossRef]
- Essa, M.M.; Subash, S.; Akbar, M.; Al-Adawi, S.; Guillemin, G.J. Long-Term Dietary Supplementation of Pomegranates, Figs and Dates Alleviate Neuroinflammation in a Transgenic Mouse Model of Alzheimer’s Disease. PLoS One 2015, 10, 1–17. [Google Scholar] [CrossRef]
- Braidy, N.; Essa, M.M.; Poljak, A.; Selvaraju, S.; Al-Adawi, S.; Manivasagm, T.; Thenmozhi, A.J.; Ooi, L.; Sachdev, P.; Guillemin, G.J. Consumption of Pomegranates Improves Synaptic Function in a Transgenic Mice Model of Alzheimer’s Disease. Oncotarget 2016, 7, 64589–64604. [Google Scholar] [CrossRef]
- DaSilva, N.A.; Nahar, P.P.; Ma, H.; Eid, A.; Wei, Z.; Meschwitz, S.; Zawia, N.H.; Slitt, A.L.; Seeram, N.P. Pomegranate Ellagitannin-Gut Microbial-Derived Metabolites, Urolithins, Inhibit Neuroinflammation in Vitro. Nutr. Neurosci. 2019, 22, 185–195. [Google Scholar] [CrossRef]
- Tu, H.J.; Su, C.J.; Peng, C.S.; Lin, T.E.; Fu, W.C.H.; Hsu, K.C.; Hwang, T.L.; Pan, S.L. Urolithin A Exhibits a Neuroprotective Effect against Alzheimer’s Disease by Inhibiting DYRK1A Activity. J. Food Drug Anal. 2023, 31, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Olajide, O.J.; La Rue, C.; Bergdahl, A.; Chapman, C.A. Inhibiting Amyloid Beta (1–42) Peptide-Induced Mitochondrial Dysfunction Prevents the Degradation of Synaptic Proteins in the Entorhinal Cortex. Front. Aging Neurosci. 2022, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Denizot, F.; Lang, R. Rapid Colorimetric Assay for Cell Growth and Survival. Modifications to the Tetrazolium Dye Procedure Giving Improved Sensitivity and Reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Hyeoncheol, K.; Xiang, X. Detection of Total Reactive Oxygen Species in Adherent Cells by 2’,7’-Dichlorodihydrofluorescein Diacetate Staining. J. Vis. Exp. 2020, 160. [Google Scholar]
- Venkatesan, T.; Choi, Y.W.; Kim, Y.K. Effect of an Extraction Solvent on the Antioxidant Quality of Pinus Densiflora Needle Extract. J. Pharm. Anal. 2019, 9, 193–200. [Google Scholar] [CrossRef]
- ÇINAR, İ.; YAYLA, M.; DEMİRBAĞ, Ç.; BİNNETOĞLU, D. Pomegranate Peel Extract Reduces Cisplatin-Induced Toxicity and Oxidative Stress in Primary Neuron Culture. Clin. Exp. Heal. Sci. 2021, 11, 59–64. [Google Scholar] [CrossRef]
- Karthika, C.; Sureshkumar, R.; Zehravi, M.; Akter, R.; Ali, F.; Ramproshad, S.; Mondal, B.; Tagde, P.; Ahmed, Z.; Khan, F.S.; et al. Multidrug Resistance of Cancer Cells and the Vital Role of P-Glycoprotein. Life 2022, 12, 1–20. [Google Scholar] [CrossRef]
- Binkhathlan, Z.; Lavasanifar, A. P-Glycoprotein Inhibition as a Therapeutic Approach for Overcoming Multidrug Resistance in Cancer: Current Status and Future Perspectives. Curr. Cancer Drug Targets 2013, 13, 326–346. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.; Gonçalves, A.C.; Jorge, J.; Almeida, A.M.; Sarmento-ribeiro, A.B. Combination of Elacridar with Imatinib Modulates Resistance Associated with Drug Efflux Transporters in Chronic Myeloid Leukemia. Biomedicines 2022, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Siddig, S.; Uzu, M.; Suzuki, S.; Nomura, Y.; Kashiba, T.; Gushimiyagi, K.; Sekine, Y.; Uehara, T.; Arano, Y.; et al. Elacridar Enhances the Cytotoxic Effects of Sunitinib and Prevents Multidrug Resistance in Renal Carcinoma Cells. Eur. J. Pharmacol. 2015, 746, 258–266. [Google Scholar] [CrossRef]
- Brouillette, J.; Caillierez, R.; Zommer, N.; Alves-Pires, C.; Benilova, I.; Blum, D.; de Strooper, B.; Buée, L. Neurotoxicity and Memory Deficits Induced by Soluble Low-Molecular-Weight Amyloid-β 1-42 Oligomers Are Revealed in Vivo by Using a Novel Animal Model. J. Neurosci. 2012, 32, 7852–7861. [Google Scholar] [CrossRef] [PubMed]
- Cizas, P.; Budvytyte, R.; Morkuniene, R.; Moldovan, R.; Broccio, M.; Lösche, M.; Niaura, G.; Valincius, G.; Borutaite, V. Size-Dependent Neurotoxicity of β-Amyloid Oligomers. Arch. Biochem. Biophys. 2010, 496, 84–92. [Google Scholar] [CrossRef]
- Ling, D.; Song, H.J.; Garza, D.; Neufeld, T.P.; Salvaterra, P.M. Abeta42-Induced Neurodegeneration via an Age-Dependent Autophagic-Lysosomal Injury in Drosophila. PLoS One 2009, 4. [Google Scholar] [CrossRef]
- Tosun, D.; Schuff, N.; Mathis, C.A.; Jagust, W.; Weiner, M.W. Spatial Patterns of Brain Amyloid-β Burden and Atrophy Rate Associations in Mild Cognitive Impairment. Brain 2011, 134, 1077–1088. [Google Scholar] [CrossRef]
- Kemppainen, N.; Johansson, J.; Teuho, J.; Helin, S.; Liu, Y.; Helisalmi, S.; Soininen, H.; Parkkola, R.; Ngandu, T.; Kivipelto, X.M.; et al. Brain -Amyloid and Atrophy in Individuals at Increased Risk of Cognitive Decline. Am. J. Neuroradiol. 2019, 40, 80–85. [Google Scholar] [CrossRef]
- Chételat, G.; Villemagne, V.L.; Bourgeat, P.; Pike, K.E.; Jones, G.; Ames, D.; Ellis, K.A.; Szoeke, C.; Martins, R.N.; O’Keefe, G.J.; et al. Relationship between Atrophy and β-Amyloid Deposition in Alzheimer Disease. Ann. Neurol. 2010, 67, 317–324. [Google Scholar] [CrossRef]
- Iflazoglu Mutlu, S.; Seven, I.; Arkali, G.; Birben, N.; Sur Arslan, A.; Aksakal, M.; Tatli Seven, P. Ellagic Acid Plays an Important Role in Enhancing Productive Performance and Alleviating Oxidative Stress, Apoptosis in Laying Quail Exposed to Lead Toxicity. Ecotoxicol. Environ. Saf. 2021, 208, 111608. [Google Scholar] [CrossRef]
- Bureau, G.; Longpré, F.; Martinoli, M.G. Resveratrol and Quercetin, Two Natural Polyphenols, Reduce Apoptotic Neuronal Cell Death Induced by Neuroinflammation. J. Neurosci. Res. 2008, 86, 403–410. [Google Scholar] [CrossRef] [PubMed]
- El-Missiry, M.A.; ElKomy, M.A.; Othman, A.I.; AbouEl-ezz, A.M. Punicalagin Ameliorates the Elevation of Plasma Homocysteine, Amyloid-β, TNF-α and Apoptosis by Advocating Antioxidants and Modulating Apoptotic Mediator Proteins in Brain. Biomed. Pharmacother. 2018, 102, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Bolaños, J.P.; Moro, M.A.; Lizasoain, I.; Almeida, A. Mitochondria and Reactive Oxygen and Nitrogen Species in Neurological Disorders and Stroke: Therapeutic Implications. Adv. Drug Deliv. Rev. 2009, 61, 1299–1315. [Google Scholar] [CrossRef]
- Sabraoui, T.; Khider, T.; Nasser, B.; Eddoha, R.; Moujahid, A.; Benbachir, M.; Essamadi, A. Determination of Punicalagins Content, Metal Chelating, and Antioxidant Properties of Edible Pomegranate (Punica Granatum L) Peels and Seeds Grown in Morocco. Int. J. Food Sci. 2020, 2020, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Robert Cronin Yung Peng, Rose Khavari, N.D. 乳鼠心肌提取 HHS Public Access. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Yoshida, Y.; Shichiri, M. Oxidative Stress Biomaker and Its Application to Health Maintainance The Role of Lipid Peroxidation in Neurological Disorders. J. Clin. Biochem. Nutr 2014, 54, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Smaoui, S.; Hlima, H. Ben; Mtibaa, A.C.; Fourati, M.; Sellem, I.; Elhadef, K.; Ennouri, K.; Mellouli, L. Pomegranate Peel as Phenolic Compounds Source: Advanced Analytical Strategies and Practical Use in Meat Products. Meat Sci. 2019, 158, 107914. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, S.; Khan, J.A.; Aslam, B.; Faisal, M.N. Pomegranate Peel Extract and Quercetin Possess Antioxidant and Hepatoprotective Activity against Concanavalin A-Induced Liver Injury in Mice. Pak. Vet. J. 2021, 41, 197–202. [Google Scholar] [CrossRef]
- Morzelle, M.C.; Salgado, J.M.; Telles, M.; Mourelle, D.; Bachiega, P.; Buck, H.S.; Viel, T.A. Neuroprotective Effects of Pomegranate Peel Extract after Chronic Infusion with Amyloid-β Peptide in Mice. PLoS One 2016, 11, 1–20. [Google Scholar] [CrossRef]
- Yaidikar, L.; Byna, B.; Thakur, S.R. Neuroprotective Effect of Punicalagin against Cerebral Ischemia Reperfusion-Induced Oxidative Brain Injury in Rats. J. Stroke Cerebrovasc. Dis. 2014, 23, 2869–2878. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Llorach, R.; Cerón, J.J.; Espín, J.C.; Tomás-Barberán, F.A. Evaluation of the Bioavailability and Metabolism in the Rat of Punicalagin, an Antioxidant Polyphenol from Pomegranate Juice. Eur. J. Nutr. 2003, 42, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Lee, R.; Heber, D. Bioavailability of Ellagic Acid in Human Plasma after Consumption of Ellagitannins from Pomegranate (Punica Granatum L.) Juice. Clin. Chim. Acta 2004, 348, 63–68. [Google Scholar] [CrossRef]
- Serdar, B.S.; Erkmen, T.; Koçtürk, S. Combinations of Polyphenols Disaggregate Aβ1-42 by Passing through in Vitro Blood-Brain Barrier Developed by Endothelium, Astrocyte, and Differentiated SH-SY5Y Cells. Acta Neurobiol. Exp. (Wars). 2021, 335–349. [Google Scholar] [CrossRef]
- Wang, W.; Yang, L.; Liu, T.; Wang, J.; Wen, A.; Ding, Y. Aging-12-103270. 2020, 12, 10457–10472.
- Lu, Z.; Liu, S.; Lopes-Virella, M.F.; Wang, Z. LPS and Palmitic Acid Co-Upregulate Microglia Activation and Neuroinflammatory Response. Compr. Psychoneuroendocrinology 2021, 6, 100048. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Ma, J.; Gao, W.; Zhang, L.; Li, J.; Li, J.; Zang, J. Pomegranate Peel as a Source of Bioactive Compounds: A Mini Review on Their Physiological Functions. Front. Nutr. 2022, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Benchagra, L.; Berrougui, H.; Islam, M.O.; Ramchoun, M. Antioxidant Effect of Moroccan Pomegranate ( Punica Granatum L. Foods 2021, 10, 2219. [Google Scholar] [CrossRef]
- Iglesias-Aguirre, C.E.; García-Villalba, R.; Beltrán, D.; Frutos-Lisón, M.D.; Espín, J.C.; Tomás-Barberán, F.A.; Selma, M. V. Gut Bacteria Involved in Ellagic Acid Metabolism To Yield Human Urolithin Metabotypes Revealed. J. Agric. Food Chem. 2023, 71, 4029–4035. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, A.K.; Kumar, R.; Jamieson, S.; Pandey, A.K.; Bishayee, A. Neuroprotective Potential of Ellagic Acid: A Critical Review. Adv. Nutr. 2021, 12, 1211–1238. [Google Scholar] [CrossRef]
- Sanjay; Shin, J.H.; Park, M.; Lee, H.J. Cyanidin-3-O-Glucoside Regulates the M1/M2 Polarization of Microglia via PPARγ and Aβ42 Phagocytosis Through TREM2 in an Alzheimer’s Disease Model. Mol. Neurobiol. 2022, 59, 5135–5148. [Google Scholar] [CrossRef]
- Straub, R.H.; Schradin, C. Chronic Inflammatory Systemic Diseases – an Evolutionary Trade-off between Acutely Beneficial but Chronically Harmful Programs. Evol. Med. Public Heal. 2016, eow001. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Across the Life Span. Nat. Med. 2019, 25. [Google Scholar] [CrossRef]
- Hennen, J.; Aeby, P.; Goebel, C.; Schettgen, T.; Oberli, A.; Kalmes, M.; Blömeke, B. Cross Talk between Keratinocytes and Dendritic Cells: Impact on the Prediction of Sensitization. Toxicol. Sci. 2011, 123, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.Y.; Han, B.; Deng, X.; Deng, S.Y.; Zhang, Y.Y.; Shen, P.X.; Hui, T.; Chen, R.H.; Li, X.; Zhang, Y. Pomegranate Peel Extract Ameliorates the Severity of Experimental Autoimmune Encephalomyelitis via Modulation of Gut Microbiota. Gut Microbes 2020, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Romier-Crouzet, B.; Van De Walle, J.; During, A.; Joly, A.; Rousseau, C.; Henry, O.; Larondelle, Y.; Schneider, Y.J. Inhibition of Inflammatory Mediators by Polyphenolic Plant Extracts in Human Intestinal Caco-2 Cells. Food Chem. Toxicol. 2009, 47, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Romier, B.; Schneider, Y.J.; Larondelle, Y.; During, A. Dietary Polyphenols Can Modulate the Intestinal Inflammatory Response. Nutr. Rev. 2009, 67, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Yaidikar, L.; Thakur, S. Punicalagin Attenuated Cerebral Ischemia–Reperfusion Insult via Inhibition of Proinflammatory Cytokines, up-Regulation of Bcl-2, down-Regulation of Bax, and Caspase-3. Mol. Cell. Biochem. 2015, 402, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Vezza, T.; Rodríguez-Nogales, A.; Algieri, F.; Utrilla, M.P.; Rodriguez-Cabezas, M.E.; Galvez, J. Flavonoids in Inflammatory Bowel Disease: A Review. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Viuda-Martos, M.; Fernández-Lóaez, J.; Pérez-álvarez, J.A. Pomegranate and Its Many Functional Components as Related to Human Health: A Review. Compr. Rev. Food Sci. Food Saf. 2010, 9, 635–654. [Google Scholar] [CrossRef]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B. V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Li, S.; Wu, B.; Fu, W.; Reddivari, L. The Anti-Inflammatory Effects of Dietary Anthocyanins against Ulcerative Colitis. Int. J. Mol. Sci. 2019, 20, 1–18. [Google Scholar] [CrossRef]
- Xu, X.; Guo, Y.; Zhao, J.; He, S.; Wang, Y.; Lin, Y.; Wang, N.; Liu, Q. Punicalagin, a PTP1B Inhibitor, Induces M2c Phenotype Polarization via up-Regulation of HO-1 in Murine Macrophages. Free Radic. Biol. Med. 2017, 110, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Huang, H.J.; Sheu, S.Y.; Liu, Y.C.; Lee, I.J.; Chiang, S.C.; Lin, A.M.Y. Oral Ellagic Acid Attenuated LPS-Induced Neuroinflammation in Rat Brain: MEK1 Interaction and M2 Microglial Polarization. Exp. Biol. Med. 2023, 248, 656–664. [Google Scholar] [CrossRef]
- Seeram, N.P.; Aronson, W.J.; Zhang, Y.; Henning, S.M.; Moro, A.; Lee, R.P.; Sartippour, M.; Harris, D.M.; Rettig, M.; Suchard, M.A.; et al. Pomegranate Ellagitannin-Derived Metabolites Inhibit Prostate Cancer Growth and Localize to the Mouse Prostate Gland. J. Agric. Food Chem. 2007, 55, 7732–7737. [Google Scholar] [CrossRef]
- Aharoni, S.; Lati, Y.; Aviram, M.; Fuhrman, B. Pomegranate Juice Polyphenols Induce a Phenotypic Switch in Macrophage Polarization Favoring a M2 Anti-Inflammatory State. BioFactors 2015, 41, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Reitz, C. Genetic Loci Associated with Alzheimer’s Disease. Future Neurol. 2014, 9, 119–122. [Google Scholar] [CrossRef]
- Waring, S.C.; Rosenberg, R.N. Genome-Wide Association Studies in Alzheimer Disease. Arch. Neurol. 2008, 65, 329–334. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, Y.; Wang, D.; Zhang, J.; Zhang, F. NSAID Exposure and Risk of Alzheimer’s Disease: An Updated Meta-Analysis from Cohort Studies. Front. Aging Neurosci. 2018, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, L.; Ongini, E.; Wenk, G. Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) in Alzheimer’s Disease: Old and New Mechanisms of Action. J. Neurochem. 2004, 91, 521–536. [Google Scholar] [CrossRef]
- Gong, Z.; Huang, J.; Xu, B.; Ou, Z.; Zhang, L.; Lin, X.; Ye, X.; Kong, X.; Long, D.; Sun, X.; et al. Urolithin A Attenuates Memory Impairment and Neuroinflammation in APP/PS1 Mice. J. Neuroinflammation 2019, 16, 1–13. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, J.; Ren, G.; Zhang, Y.; Tan, X.; Yang, L. Punicalagin Prevents Inflammation in Lps-Induced Raw264.7 Macrophages by Inhibiting Foxo3a/Autophagy Signaling Pathway. Nutrients 2019, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Li, J.; Zhang, X.; Wang, L.; Zhang, W.; Yang, M.; Hou, C. Pomegranate Peel Polyphenols Inhibits Inflammation in LPS-Induced RAW264.7 Macrophages via the Suppression of TLR4/NF-ΚB Pathway Activation. Food Nutr. Res. 2019, 63, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tapias, V.; Cannon, J.R.; Greenamyre, J.T. Pomegranate Juice Exacerbates Oxidative Stress and Nigrostriatal Degeneration in Parkinson’s Disease. Neurobiol. Aging 2014, 35, 1162–1176. [Google Scholar] [CrossRef] [PubMed]
- Chachay, V.S.; Macdonald, G.A.; Martin, J.H.; Whitehead, J.P.; O’Moore-Sullivan, T.M.; Lee, P.; Franklin, M.; Klein, K.; Taylor, P.J.; Ferguson, M.; et al. Resveratrol Does Not Benefit Patients With Nonalcoholic Fatty Liver Disease. Clin. Gastroenterol. Hepatol. 2014, 12, 2092–2103. [Google Scholar] [CrossRef] [PubMed]
- Kujawska, M.; Jourdes, M.; Kurpik, M.; Szulc, M.; Szaefer, H.; Chmielarz, P.; Kreiner, G.; Krajka-ku, V. Neuroprotective Effects of Pomegranate Juice against Parkinson ’ s Disease and Presence of Ellagitannins- Derived Metabolite — Urolithin A — In the Brain. 2020.
- Fath, T.; Eidenmüller, J.; Brandt, R. Tau-Mediated Cytotoxicity in a Pseudohyperphosphorylation Model of Alzheimer’s Disease. J. Neurosci. 2002, 22, 9733–9741. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Wood, J.G. Functional Studies of Alzheimer’s Disease Tau Protein. J. Neurosci. 1993, 13, 508–515. [Google Scholar] [CrossRef]
- Mi, K.; Johnson, G. The Role of Tau Phosphorylation in the Pathogenesis of Alzheimers Disease. Curr. Alzheimer Res. 2006, 3, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martín, T.; Cuchillo-Ibáñez, I.; Noble, W.; Nyenya, F.; Anderton, B.H.; Hanger, D.P. Tau Phosphorylation Affects Its Axonal Transport and Degradation. Neurobiol. Aging 2013, 34, 2146–2157. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.A.; Lee, G.; Jay, D.G. Tau Is Required for Neurite Outgrowth and Growth Cone Motility of Chick Sensory Neurons. Cell Motil. Cytoskeleton 1999, 43, 232–242. [Google Scholar] [CrossRef]
- Johnson, G.V.W.; Stoothoff, W.H. Tau Phosphorylation in Neuronal Cell Function and Dysfunction. 2004. [Google Scholar] [CrossRef]
- Hamilton, R.L.; Kofler, J.K.; Klunk, W.E.; Oscar, L. With Psychosis. 2015, 39, 759–773. [CrossRef]
- Perotti, A.; Oliveira, M.H.; Portugal. Ministério da Educação Apologia Do Intercultural. Educ. Intercult. 1997, 7, 88. [Google Scholar]
- Murray, M.E.; Lowe, V.J.; Graff-Radford, N.R.; Liesinger, A.M.; Cannon, A.; Przybelski, S.A.; Rawal, B.; Parisi, J.E.; Petersen, R.C.; Kantarci, K.; et al. Clinicopathologic and 11C-Pittsburgh Compound B Implications of Thal Amyloid Phase across the Alzheimer’s Disease Spectrum. Brain 2015, 138, 1370–1381. [Google Scholar] [CrossRef] [PubMed]
- Tai, X.Y.; Koepp, M.; Duncan, J.S.; Fox, N.; Thompson, P.; Baxendale, S.; Liu, J.Y.W.; Reeves, C.; Michalak, Z.; Thom, M. Hyperphosphorylated Tau in Patients with Refractory Epilepsy Correlates with Cognitive Decline: A Study of Temporal Lobe Resections. Brain 2016, 139, 2441–2455. [Google Scholar] [CrossRef] [PubMed]
- Manassero, G.; Guglielmotto, M.; Zamfir, R.; Borghi, R.; Colombo, L.; Salmona, M.; Perry, G.; Odetti, P.; Arancio, O.; Tamagno, E.; et al. Beta-Amyloid 1-42 Monomers, but Not Oligomers, Produce PHF-like Conformation of Tau Protein. Aging Cell 2016, 15, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Liu, H.; Zhang, W.; Liu, X.; Jiang, B.; Fei, H.; Sun, Z. Ellagic Acid Ameliorates Learning and Memory Impairment in APP/PS1 Transgenic Mice via Inhibition of β-Amyloid Production and Tau Hyperphosphorylation. Exp. Ther. Med. 2018, 16, 4951–4958. [Google Scholar] [CrossRef]
- Ferrer, I.; Gomez-Isla, T.; Puig, B.; Freixes, M.; Ribe, E.; Dalfo, E.; Avila, J. Current Advances on Different Kinases Involved in Tau Phosphorylation, and Implications in Alzheimers Disease and Tauopathies. Curr. Alzheimer Res. 2005, 2, 3–18. [Google Scholar] [CrossRef]
- Sanbe, A.; Osinska, H.; Villa, C.; Gulick, J.; Klevitsky, R.; Glabe, C.G.; Kayed, R.; Robbins, J. Reversal of Amyloid-Induced Heart Disease in Desmin-Related Cardiomyopathy. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 13592–13597. [Google Scholar] [CrossRef]
Figure 1.
The modulatory effect of peels, arils, punicalagin and ellagic acid on human Aβ1-42-induced neuronal cytotoxicity. Neuronal H4 cells were exposed or not to 20 µg/ml of Aβ1-42 in the presence or absence of the above treatments for 24h. (A): represents the effect of Aβ1-42 on neuronal cell viability. (B): illustrates the effects of PP (peels: 200 µg/ml; arils: 200 µg/ml; punicalagin: 50 µg/ml and ellagic acid: 50 µM) on the Aβ1-42-related cytotoxicity. Data are expressed as means ± SEM. (*) vs Aβ1-42: * p<0.05, ** p<0.01.
Figure 1.
The modulatory effect of peels, arils, punicalagin and ellagic acid on human Aβ1-42-induced neuronal cytotoxicity. Neuronal H4 cells were exposed or not to 20 µg/ml of Aβ1-42 in the presence or absence of the above treatments for 24h. (A): represents the effect of Aβ1-42 on neuronal cell viability. (B): illustrates the effects of PP (peels: 200 µg/ml; arils: 200 µg/ml; punicalagin: 50 µg/ml and ellagic acid: 50 µM) on the Aβ1-42-related cytotoxicity. Data are expressed as means ± SEM. (*) vs Aβ1-42: * p<0.05, ** p<0.01.
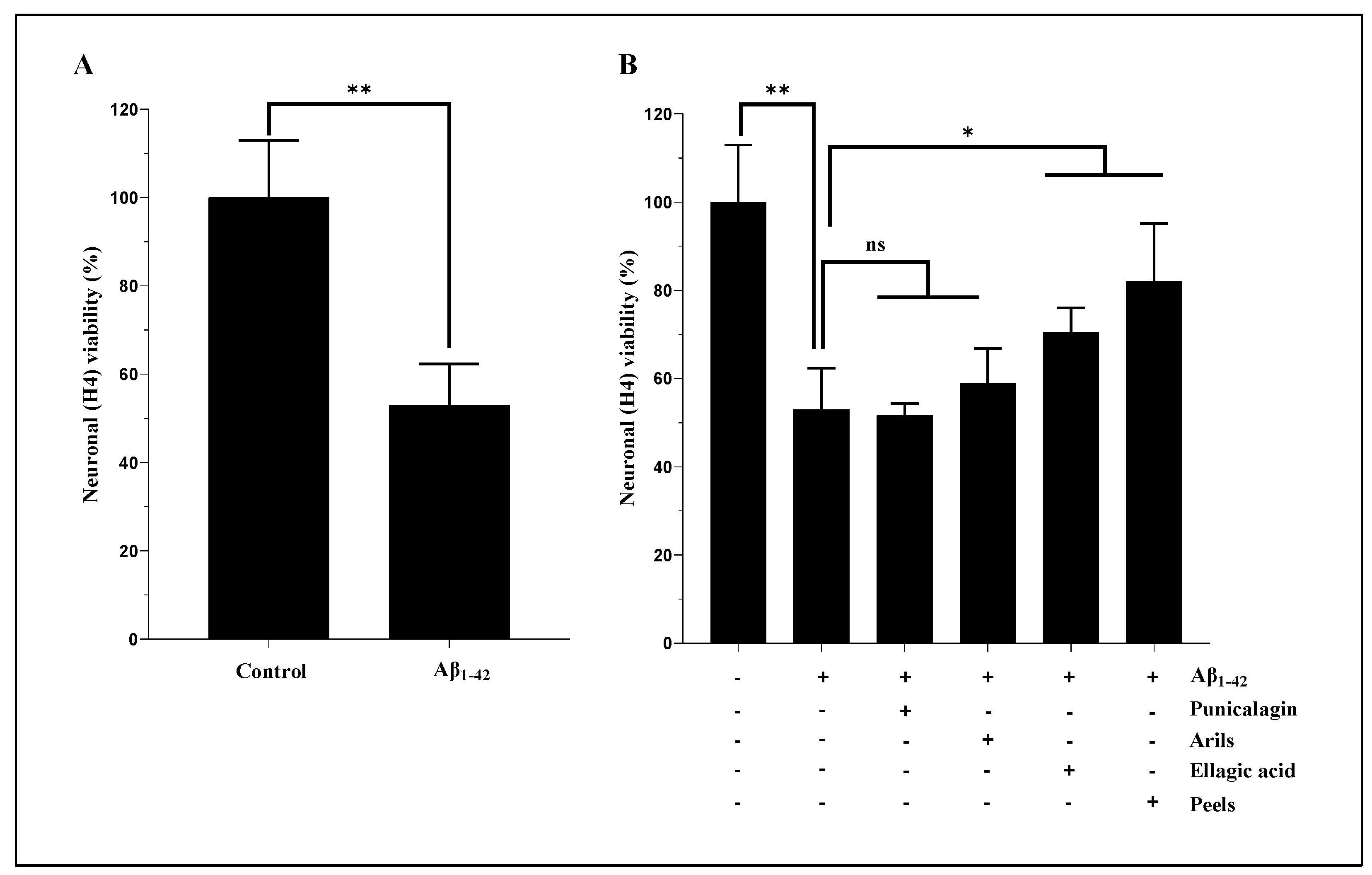
Figure 2.
Elacridar improves the effect of pomegranate arils-rich phenolic extract and punicalagin polyphenols against human Aβ1-42-induced neuronal death. H4 cells were stimulated or not with Aβ1-42 (20 µg/ml) for 24h and simultaneously treated with pomegranate arils extract (200 µg/ml) or punicalagin (50 µg/ml) in the presence of Elacridar (500 ng/ml). Data are expressed as means ± SEM. (*) vs Aβ1-42: * p<0.05, ** p<0.01, ***p<0.001.
Figure 2.
Elacridar improves the effect of pomegranate arils-rich phenolic extract and punicalagin polyphenols against human Aβ1-42-induced neuronal death. H4 cells were stimulated or not with Aβ1-42 (20 µg/ml) for 24h and simultaneously treated with pomegranate arils extract (200 µg/ml) or punicalagin (50 µg/ml) in the presence of Elacridar (500 ng/ml). Data are expressed as means ± SEM. (*) vs Aβ1-42: * p<0.05, ** p<0.01, ***p<0.001.
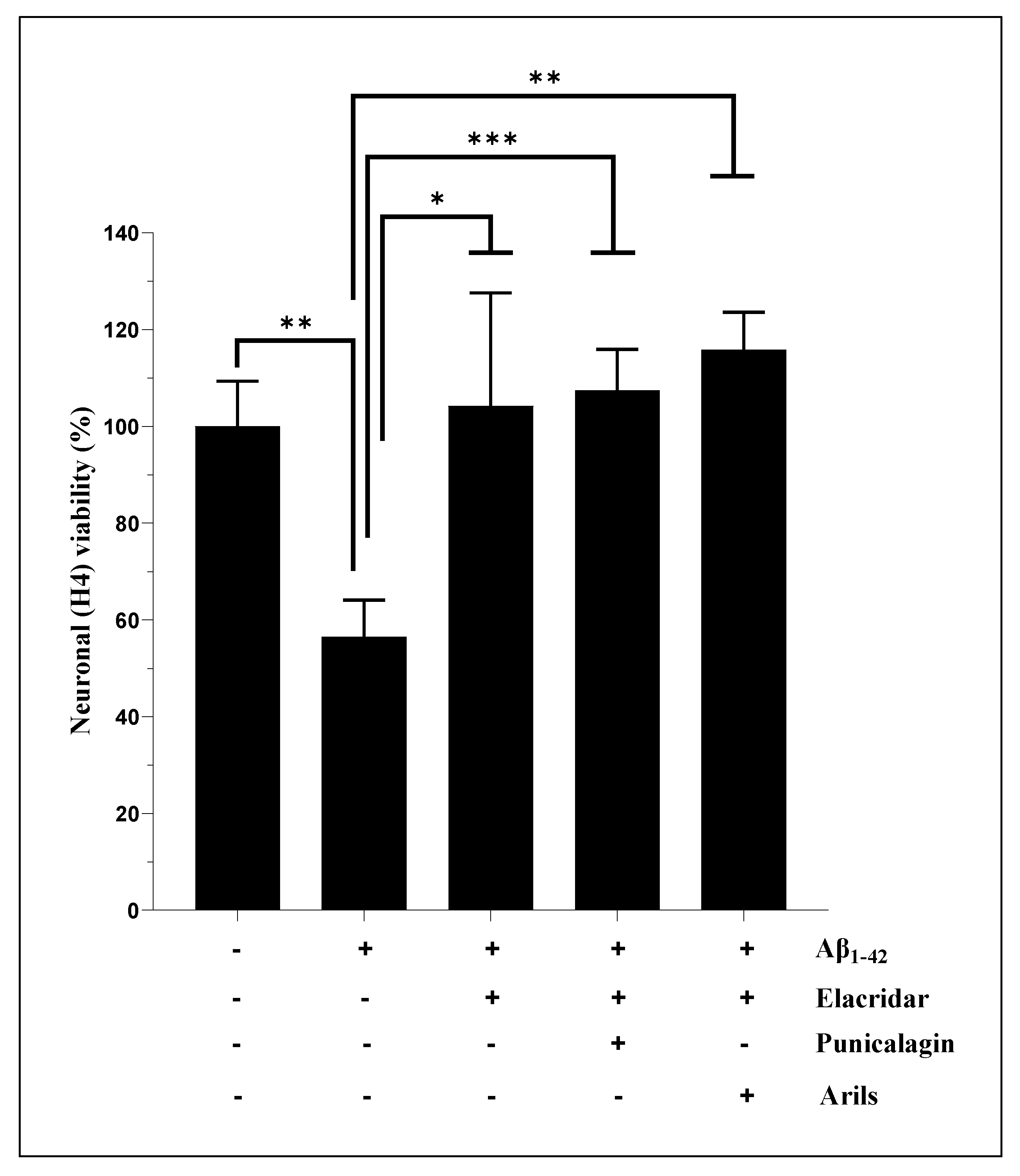
Figure 3.
The neuronal surviving enhancement effect of pomegranate peels (200 µg/ml), arils (200 µg/ml), punicalagin (50 µg/ml), and ellagic acid (50 µM). The H4 neurons were incubated with or without 15 µg/ml of human Aβ1-42 for 24h in the presence or absence of the above treatments. The obtained results are expressed as mean ± SEM. (*) vs Aβ1-42: *p< 0.05, ***p< 0.001.
Figure 3.
The neuronal surviving enhancement effect of pomegranate peels (200 µg/ml), arils (200 µg/ml), punicalagin (50 µg/ml), and ellagic acid (50 µM). The H4 neurons were incubated with or without 15 µg/ml of human Aβ1-42 for 24h in the presence or absence of the above treatments. The obtained results are expressed as mean ± SEM. (*) vs Aβ1-42: *p< 0.05, ***p< 0.001.
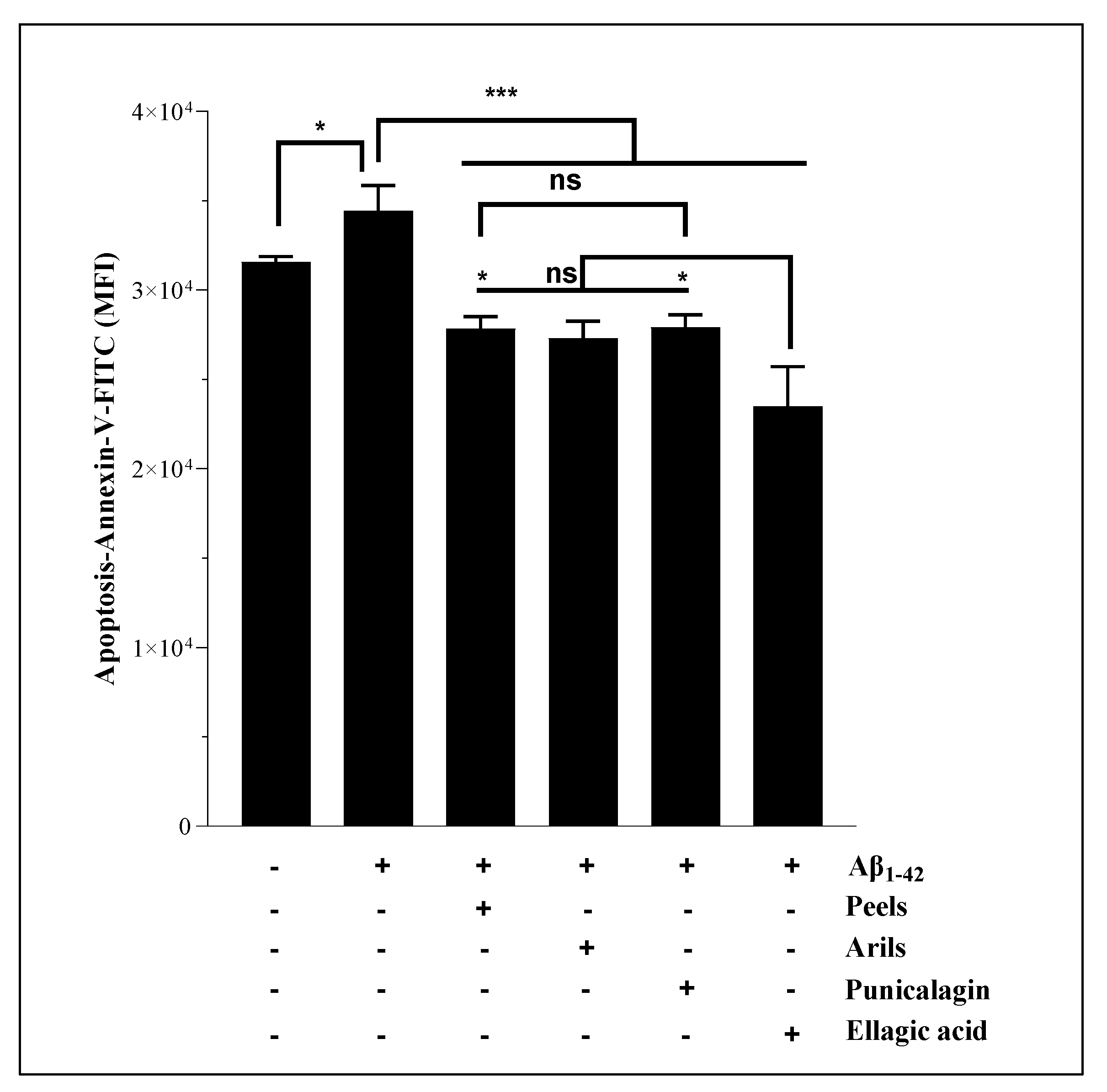
Figure 4.
Pomegranate polyphenols reduce ROS generation in human microglia HMC3 cells. Cells were pretreated with human Aβ1-42 (5 µg/ml) in the presence or not of different concentrations of pomegranate peels (A), arils (B), punicalagin (C) and ellagic acid (D). The obtained data are presented as mean ± SEM. (*) vs Aβ1-42: * p<0.05.
Figure 4.
Pomegranate polyphenols reduce ROS generation in human microglia HMC3 cells. Cells were pretreated with human Aβ1-42 (5 µg/ml) in the presence or not of different concentrations of pomegranate peels (A), arils (B), punicalagin (C) and ellagic acid (D). The obtained data are presented as mean ± SEM. (*) vs Aβ1-42: * p<0.05.
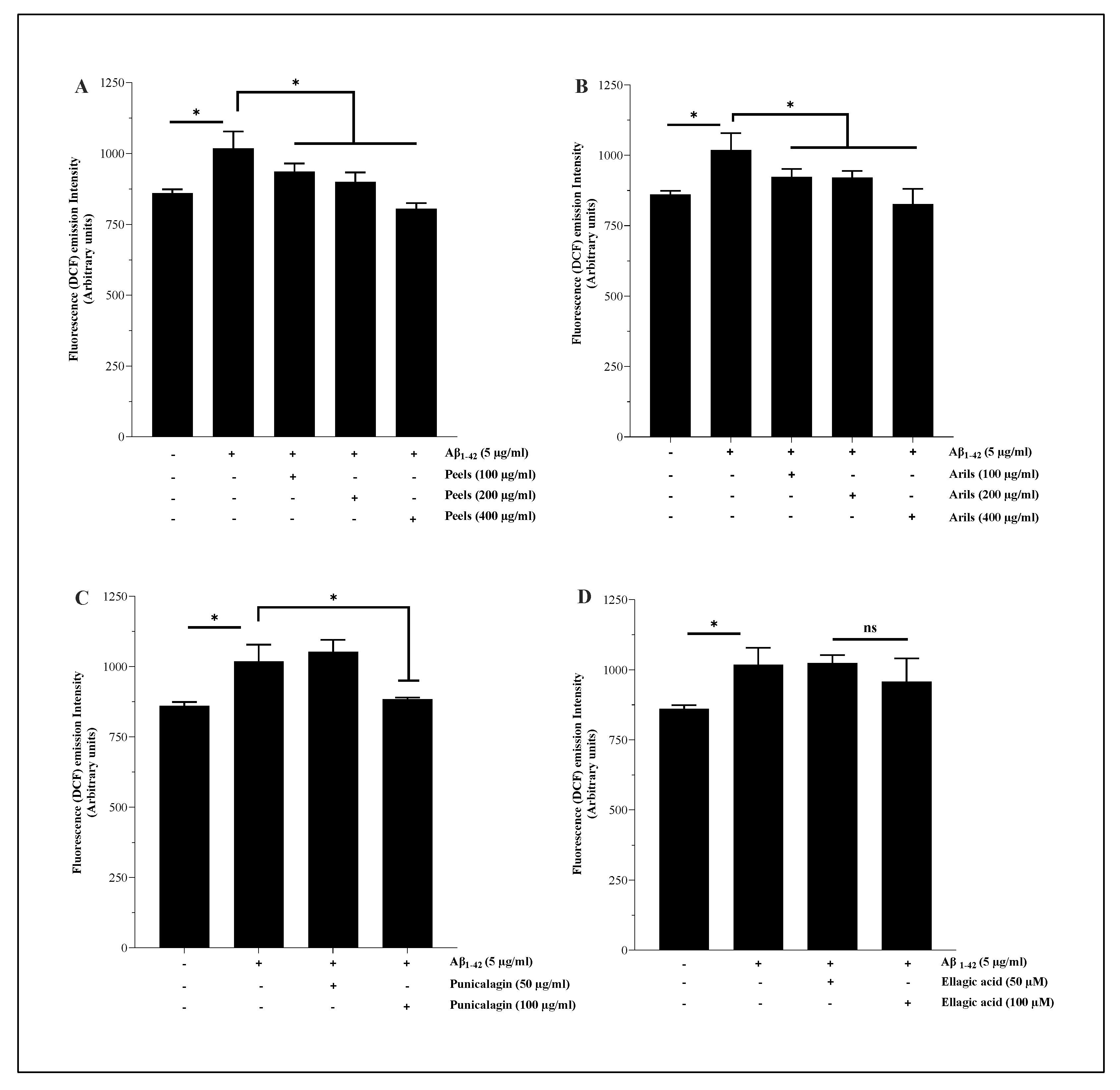
Figure 5.
The protective effect of pomegranate polyphenols on lipid peroxidation. HMC3 microglia cells were pretreated (24h) with peels (A) and arils (B) extracts, punicalagin (C) and ellagic acid (D) compounds before their stimulation with TBHP (200 µM) for one h. The data are presented as mean ± SEM of at least three independent experiments. *p< 0.05; ** p< 0.01; *** p< 0.001.
Figure 5.
The protective effect of pomegranate polyphenols on lipid peroxidation. HMC3 microglia cells were pretreated (24h) with peels (A) and arils (B) extracts, punicalagin (C) and ellagic acid (D) compounds before their stimulation with TBHP (200 µM) for one h. The data are presented as mean ± SEM of at least three independent experiments. *p< 0.05; ** p< 0.01; *** p< 0.001.
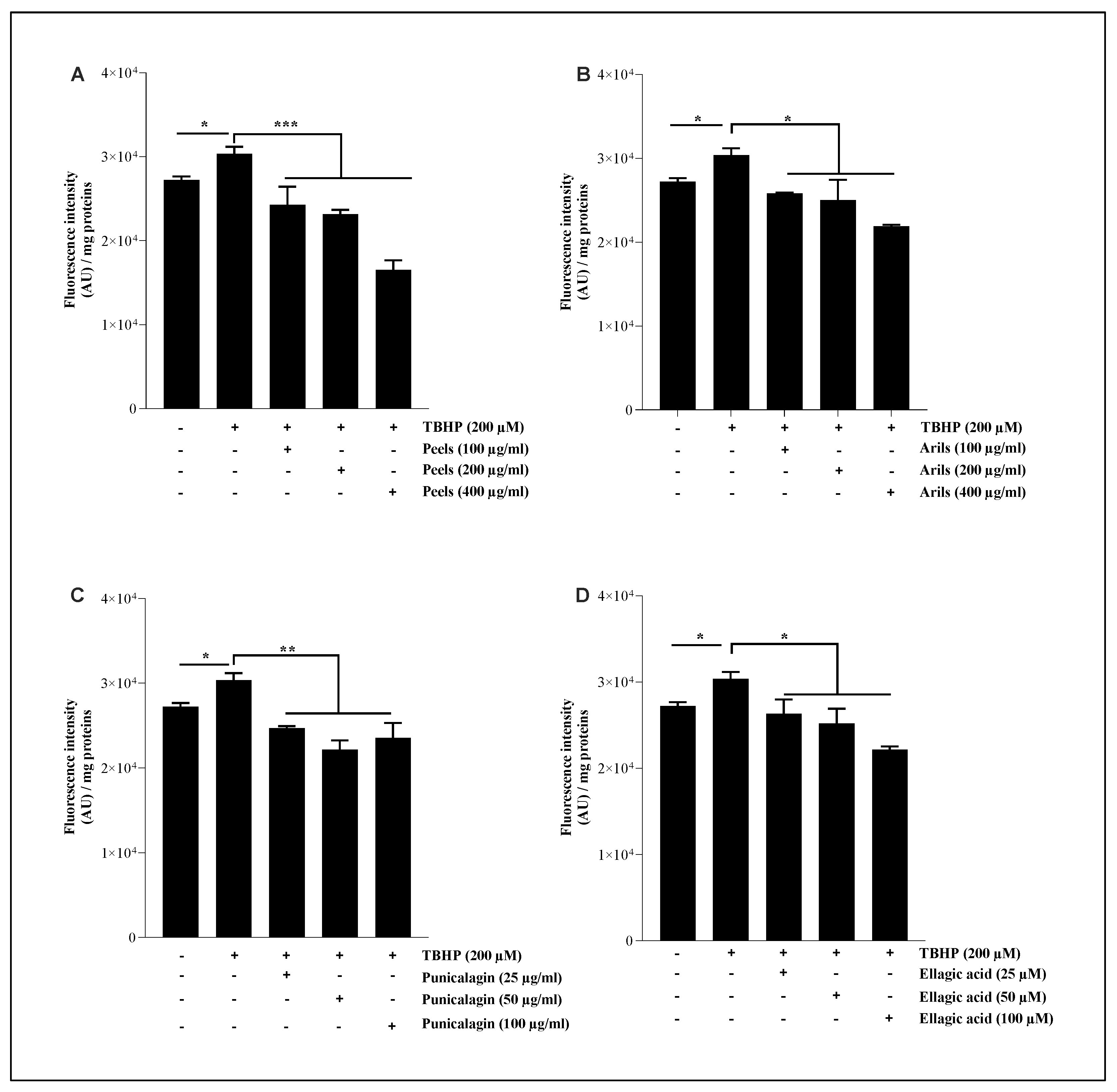
Figure 6.
The modulatory effect of peels, arils, punicalagin, and ellagic acid on M1/M2 polarisation of HMC3 microglia cells. HMC3 cells were stimulated by one µg/ml of LPS and cotreated simultaneously with pomegranate peels, arils, punicalagin, and ellagic acid for 24h. Figure 6.1 represents the effect of PP on CD86 expression. Figure 6.2 illustrates the impact of PP on CD163 receptor expression. (*) vs LPS: *p< 0.05; **p< 0.01; ***p< 0.001; ****p< 0.0001.
Figure 6.
The modulatory effect of peels, arils, punicalagin, and ellagic acid on M1/M2 polarisation of HMC3 microglia cells. HMC3 cells were stimulated by one µg/ml of LPS and cotreated simultaneously with pomegranate peels, arils, punicalagin, and ellagic acid for 24h. Figure 6.1 represents the effect of PP on CD86 expression. Figure 6.2 illustrates the impact of PP on CD163 receptor expression. (*) vs LPS: *p< 0.05; **p< 0.01; ***p< 0.001; ****p< 0.0001.
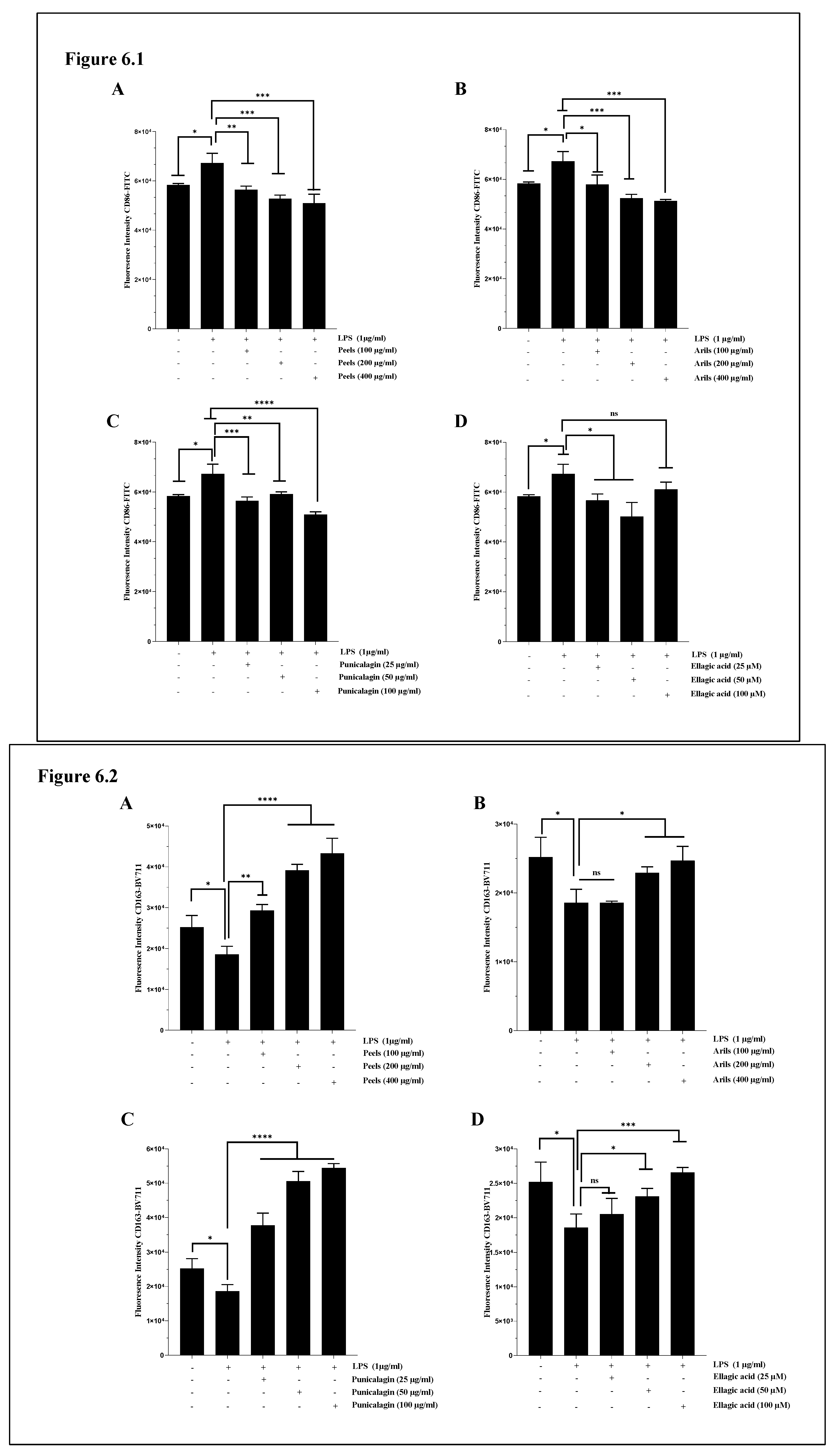
Figure 7.
The effect of pomegranate polyphenols on CD86 (Figure 7.1) and CD163 (Figure 7.2) protein expression. Human THP-1 derived macrophages were stimulated by LPS (1µg/ml) and cotreated with pomegranate peels (A), arils rich-phenolic extracts (B), punicalagin (C), and ellagic acid (D) for 24h. (*) vs LPS: *p< 0.05; ** p< 0.01; *** p< 0.001.
Figure 7.
The effect of pomegranate polyphenols on CD86 (Figure 7.1) and CD163 (Figure 7.2) protein expression. Human THP-1 derived macrophages were stimulated by LPS (1µg/ml) and cotreated with pomegranate peels (A), arils rich-phenolic extracts (B), punicalagin (C), and ellagic acid (D) for 24h. (*) vs LPS: *p< 0.05; ** p< 0.01; *** p< 0.001.
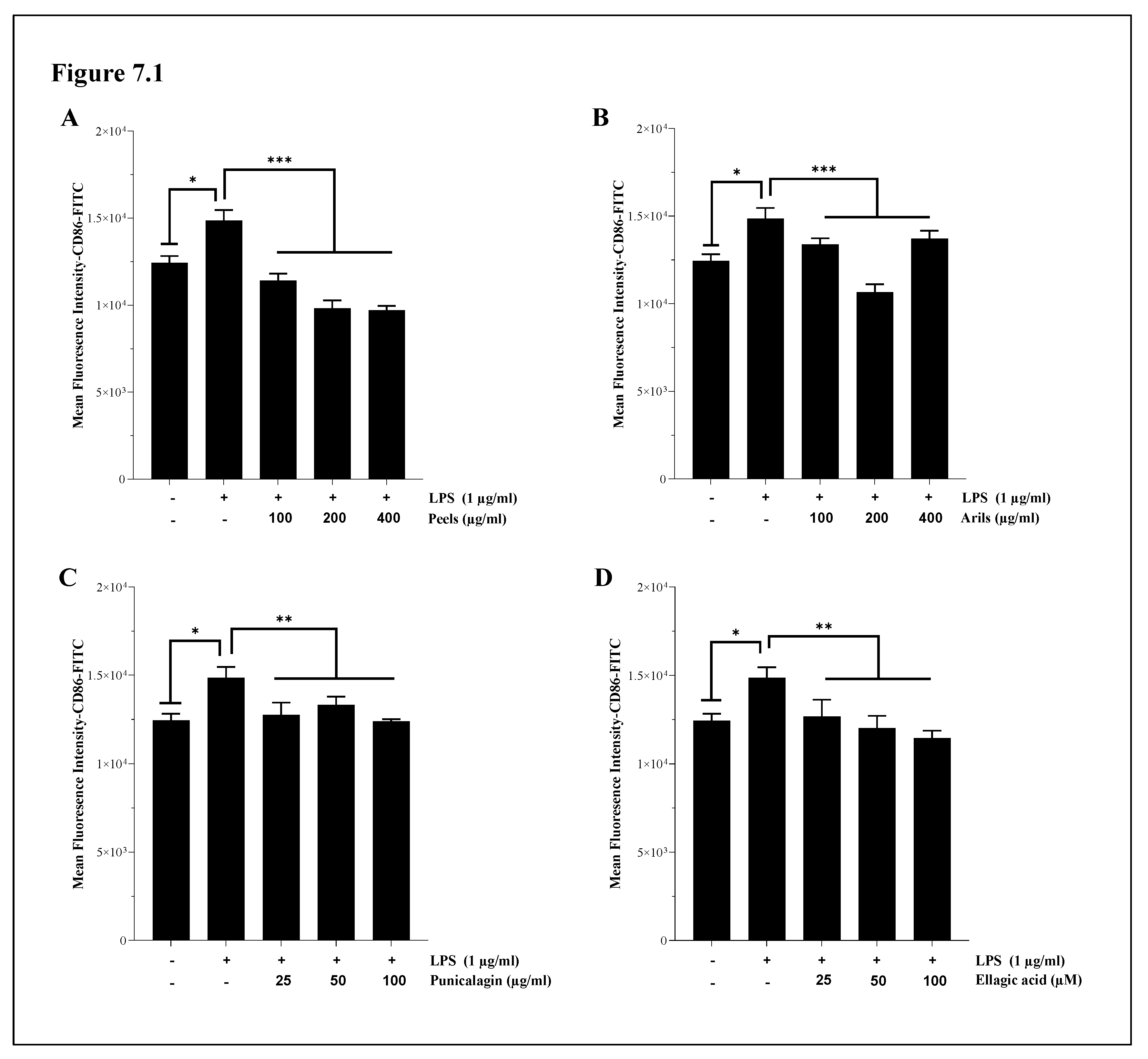
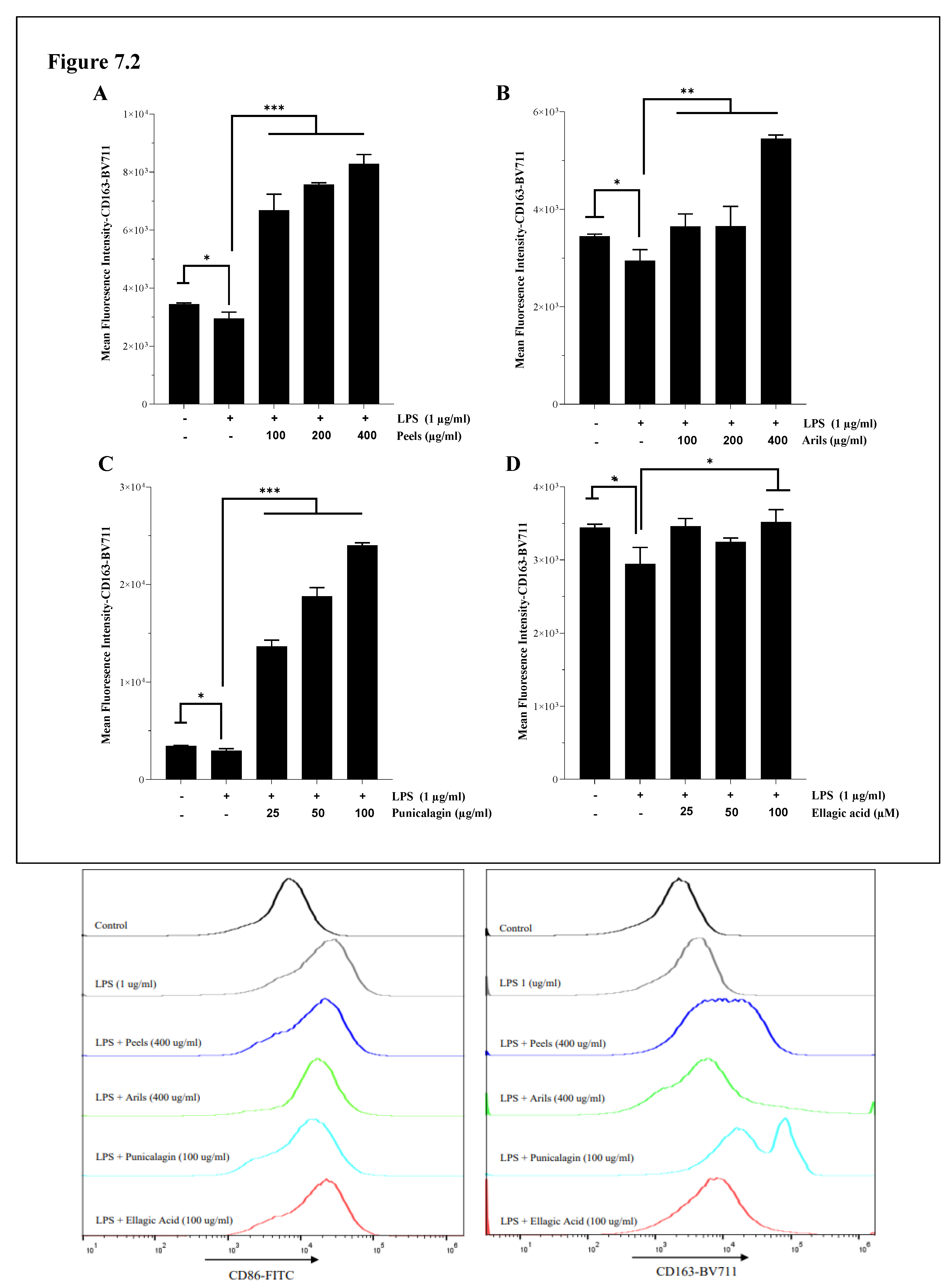
Figure 8.
The bioeffects of pomegranate polyphenols on gene expression of IL-10. The U373-MG human astrocytes were stimulated by one µg/ml of LPS and cotreated simultaneously overnight with pomegranate peels, arils, punicalagin, and ellagic acid at different concentrations. (*) vs LPS: **p< 0.01; *** p< 0.001.
Figure 8.
The bioeffects of pomegranate polyphenols on gene expression of IL-10. The U373-MG human astrocytes were stimulated by one µg/ml of LPS and cotreated simultaneously overnight with pomegranate peels, arils, punicalagin, and ellagic acid at different concentrations. (*) vs LPS: **p< 0.01; *** p< 0.001.
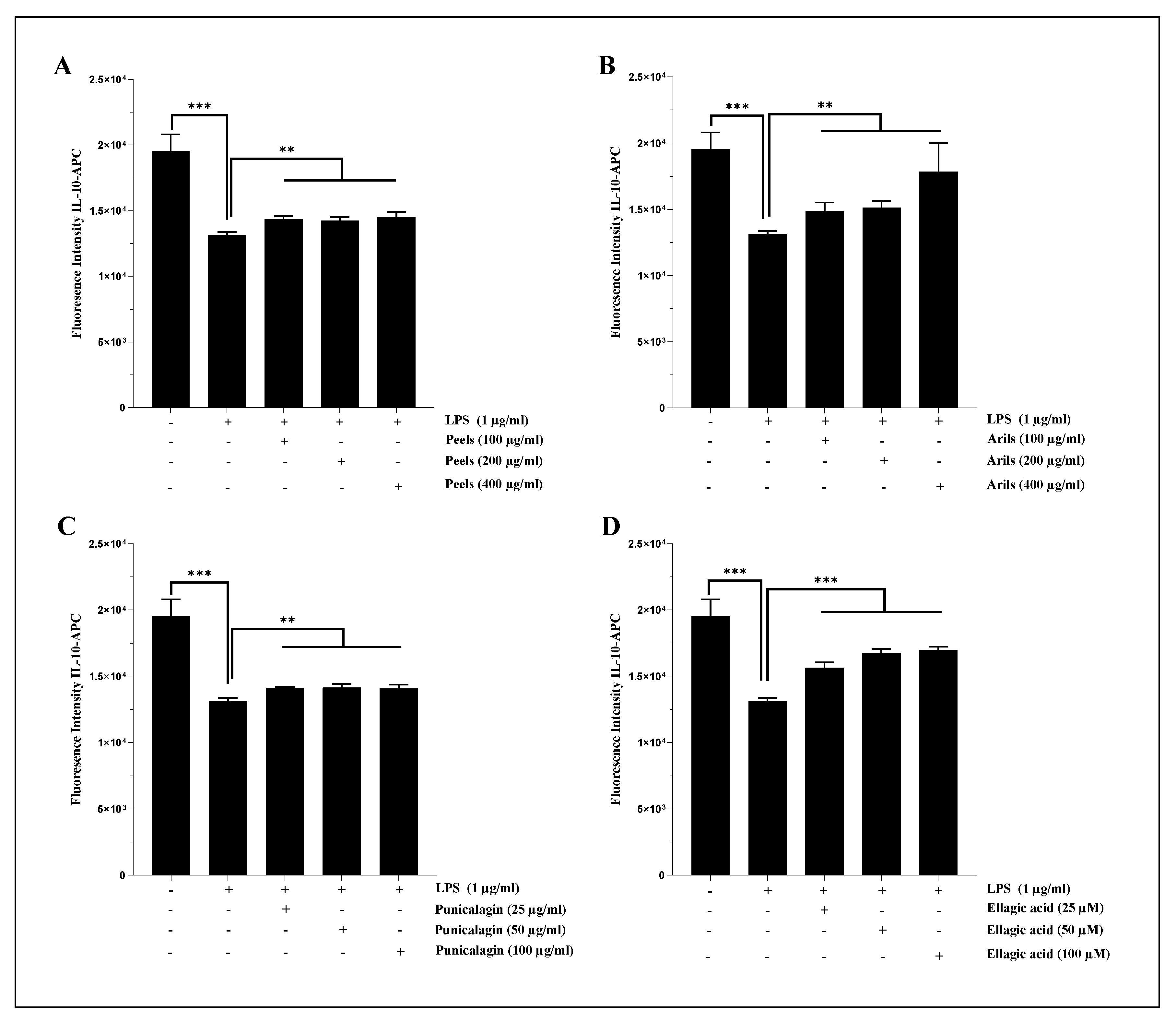
Figure 9.
The modulatory effect of pomegranate peels, arils, punicalagin, and ellagic acid on IL-10 (Figure 9.1) and IL-1β (Figure 9.2) protein expression in THP-1 derived macrophages. Cells were stimulated by one µg/ml of LPS and co-treated simultaneously with pomegranate polyphenols for 24h. (*) vs LPS: *p<0.05; ** p<0.01; *** p<0.001.
Figure 9.
The modulatory effect of pomegranate peels, arils, punicalagin, and ellagic acid on IL-10 (Figure 9.1) and IL-1β (Figure 9.2) protein expression in THP-1 derived macrophages. Cells were stimulated by one µg/ml of LPS and co-treated simultaneously with pomegranate polyphenols for 24h. (*) vs LPS: *p<0.05; ** p<0.01; *** p<0.001.
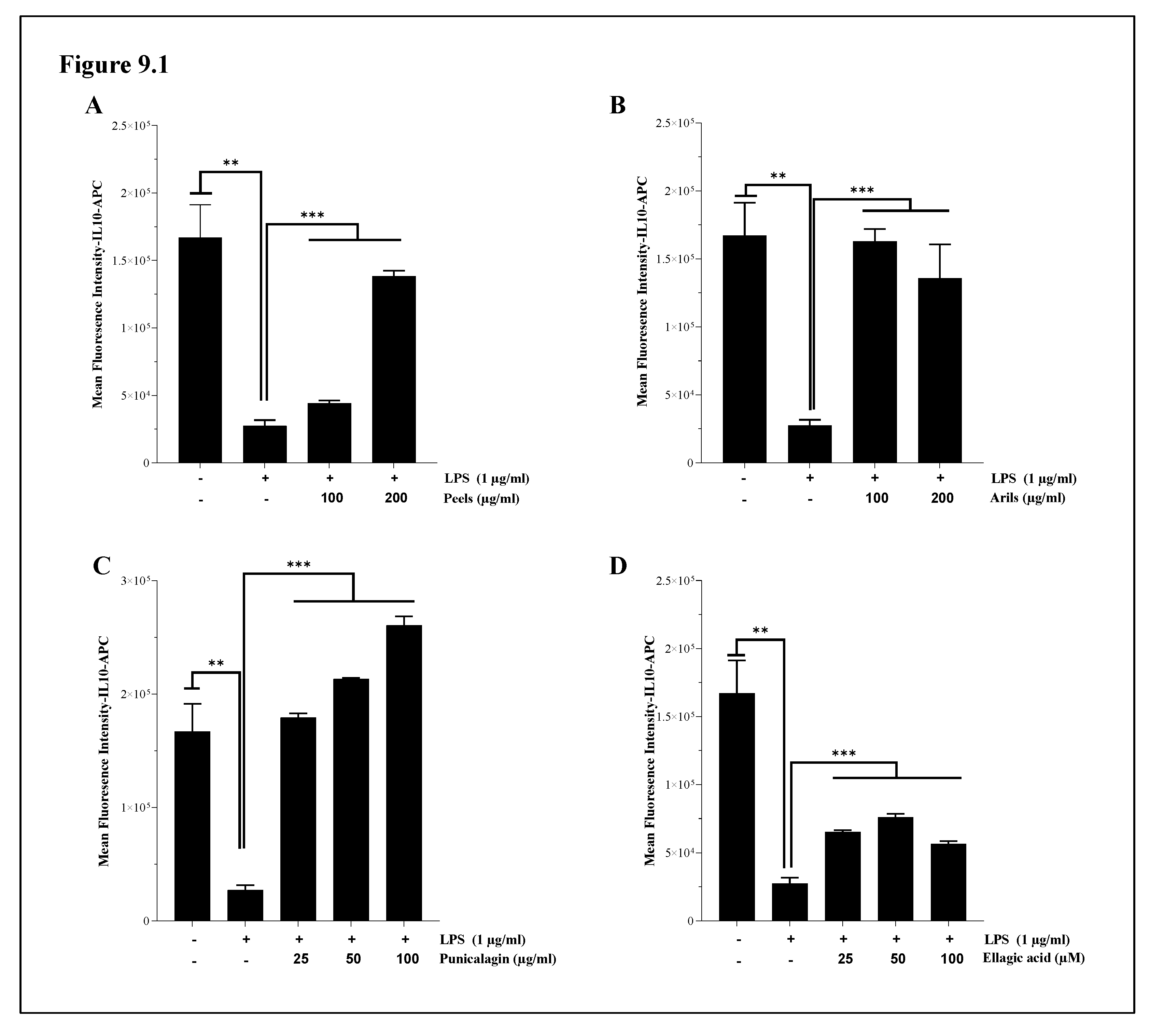
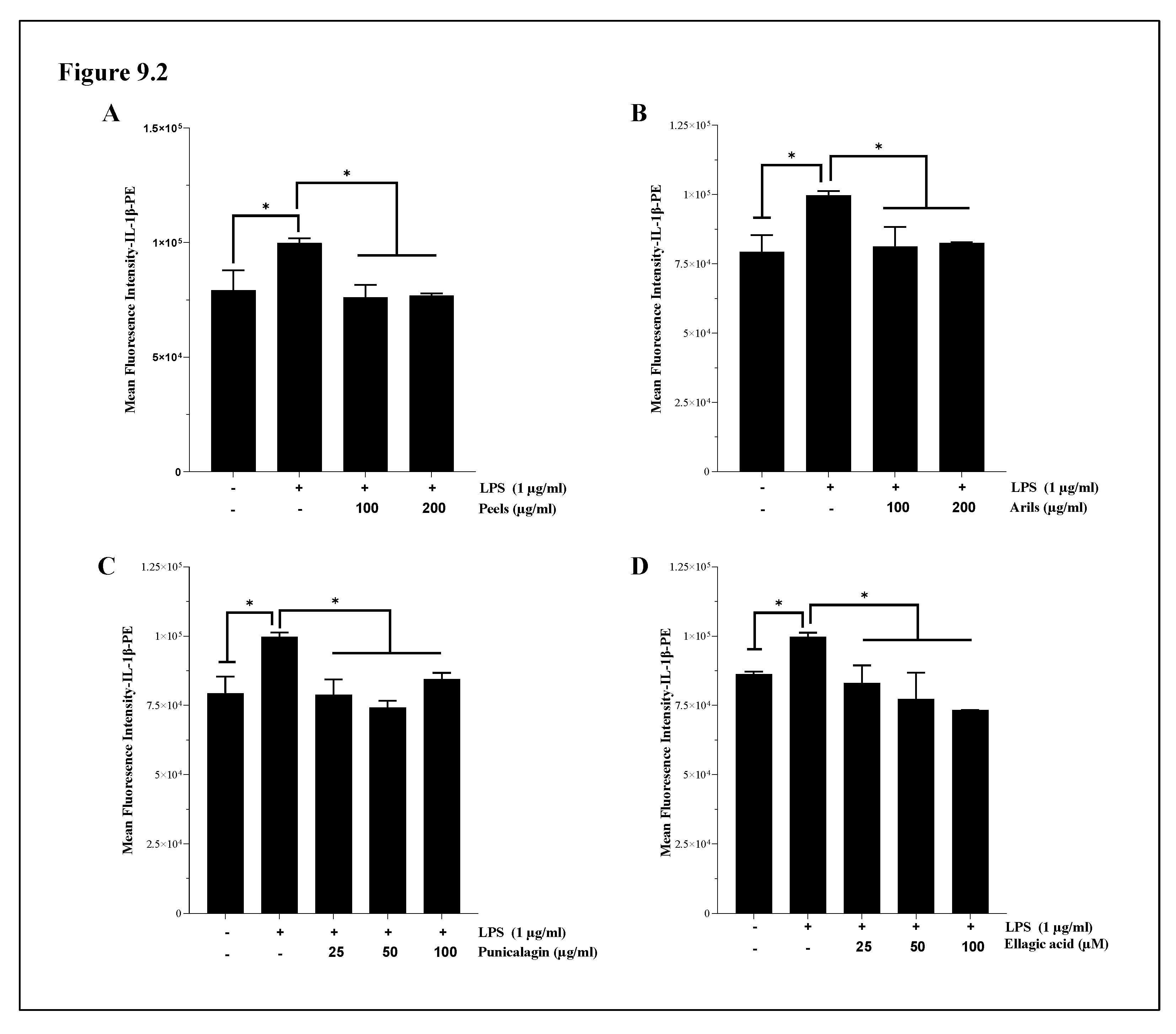
Figure 10.
The modulatory effect of pomegranate polyphenols on human Aβ1-42-induced Tau-phosphorylation at threonine 181 in H4 human neuroglioma cells. Cells were either unstimulated or stimulated with human Aβ1-42 (10 µg/ml) in the presence or not of pomegranate polyphenols simultaneously for 24h. Peels: 200 µg/ml; arils: 200 µg/ml; punicalagin: 50 µg/ml; ellagic acid: 50 µM. (*) vs Aβ1-42: *p<0.05; ** p<0.01; *** p<0.001.
Figure 10.
The modulatory effect of pomegranate polyphenols on human Aβ1-42-induced Tau-phosphorylation at threonine 181 in H4 human neuroglioma cells. Cells were either unstimulated or stimulated with human Aβ1-42 (10 µg/ml) in the presence or not of pomegranate polyphenols simultaneously for 24h. Peels: 200 µg/ml; arils: 200 µg/ml; punicalagin: 50 µg/ml; ellagic acid: 50 µM. (*) vs Aβ1-42: *p<0.05; ** p<0.01; *** p<0.001.
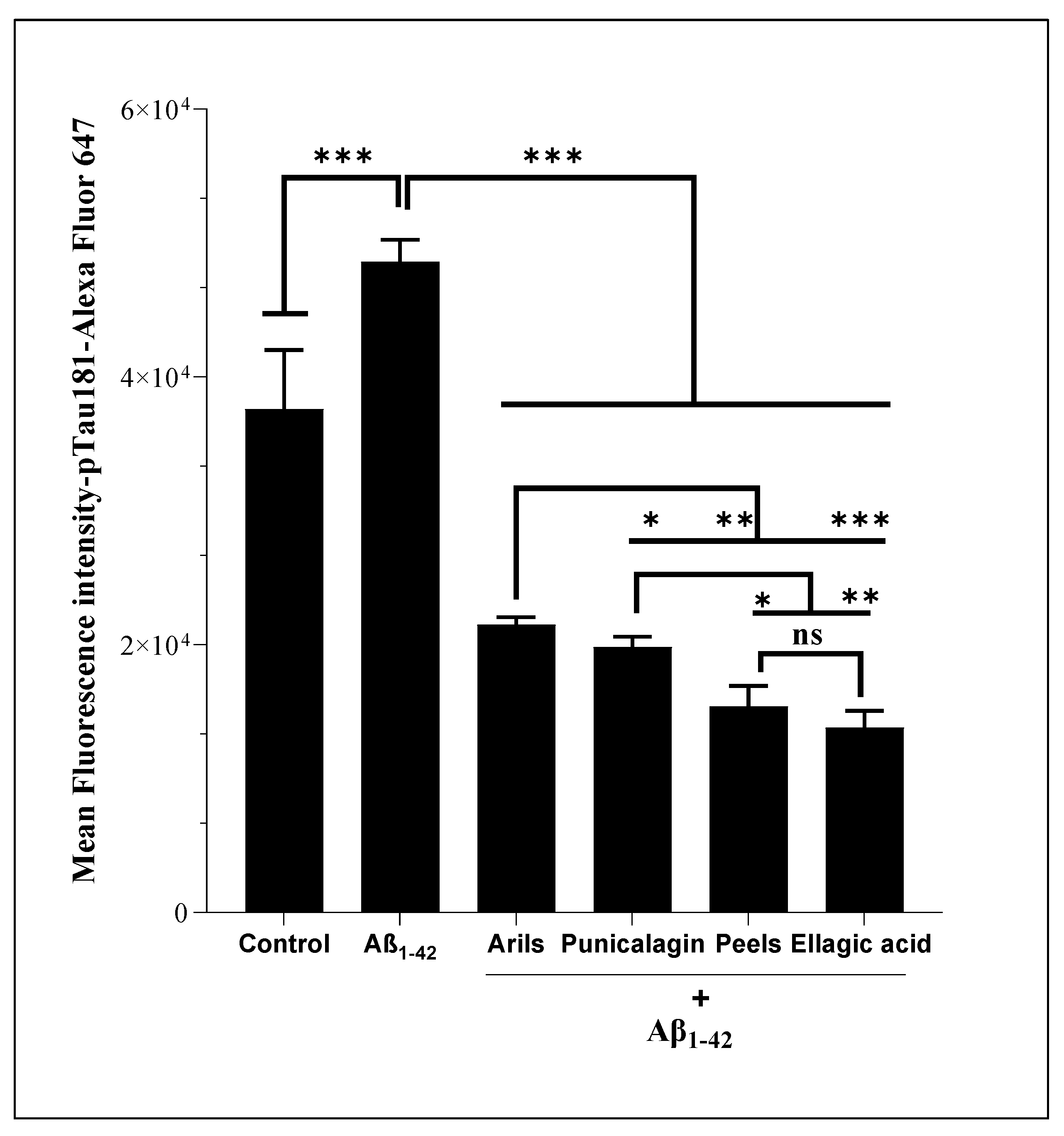
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



