
Submitted:
16 May 2024
Posted:
17 May 2024
You are already at the latest version
Abstract
The growing population in Yucatan has led to the expansion of construction in the Mayan jungle for tourist spaces, residential areas, and agriculture. Recently, rabies cases in cats (Felis catus) have increased. This study aimed to perform the antigenic and genetic characterization of the RABV and to try to identify factors contributing to the increased transmission of rabies virus from wildlife to cats in the State of Yucatan. The ArcGIS software was employed to generate maps depicting the geographic locations of rabies outbreaks in cats. A mixed-effects Poisson regression model was applied to investigate the association between cat rabies cases and sociodemographic and ecological variables. A total of nine feline rabies cases occurred, three antigenic variants were detected: RVV1 (n=1); (RVV3) (n=1), and the canine-originated atypical variant (n=7). Rabies cases were spatially distributed at the urban-jungle interface, which may favor contact between cats and wildlife. No statistically significant associations with sociodemographic/ecological variables were observed. Urban areas with low (0-2%) to moderate (2-4%) changes in land cover showed a substantial increase in the probability of feline rabies cases. This study highlights the concerning resurgence of rabies infections in cats, emphasizing the looming threat of its reintroduction in dogs should vaccination rates diminish. The genetic affinity between the atypical variant and the canine virus underscores the urgent need for vigilance in maintaining high vaccination coverage across all susceptible species.
Keywords:
Rabies
; Felis catus
; urbanization
; spillover
1. Introduction
Rabies is disease caused by neurotropic viruses of the genus Lyssavirus in the family Rhabdoviridae, it is transmissible to all mammals and it is almost uniformly fatal [1].
Cross-species transmission (CST) occurs when a rabies virus variant infects non-reservoir species, leading to disease. The frequency of CST events results from a complex interplay of eco-epidemiological, genetic, and evolutionary factors conditioning the specific virus-host relationship [2,3,4,5].
CST frequency is primarily driven by the rate of physical contact between species rather than their relatedness [6]. Changes in wildlife migration patterns and the resulting abundance in new regions correspondingly alter interspecific interactions [7,8].
While all rabies reservoirs are also vectors for the virus, not all vectors function as reservoirs. Cats (Felis catus), whether domestic, free roaming, or feral, can effectively transmit the rabies virus (RABV). However, there is no perpetuation of cat-to-cat rabies transmission. In comparison, infected dogs typically act as the predominant reservoir. Furthermore, no unique feline rabies virus variant has been documented [9].
Although human rabies transmitted by cats is infrequent [10], the potential for CST from wild-living mammals to domestic animal populations increases the risk of transmission to humans. This is due to the multidimensional connections with domestic animals within our societies [2,11,12,13].
According to the SIRVERA (Regional Information System for Epidemiological Surveillance of Rabies) database, there were 42 cases of human rabies in the Americas region up until 2020. Among these, cats transmitted eight: three in Cuba, two in Colombia (one with rabies virus variant (RVV) 4), one in Brazil (RVV3), one in Argentina (RVV3), and one in Mexico [14].
Meanwhile, cats stand out as the most commonly reported rabid domestic animal in the United States [15]. The virus can be found in the saliva of rabid cats, leading to transmission to humans through bites. It has been documented that cats may shed the virus up to 10 days before the onset of clinical signs. Since 1990, reported cases of domestic cats have consistently surpassed those of dogs in the United States [16].
In Mexico, three rabies cases transmitted by cats have been recorded to date: one in the state of Quintana Roo in 2004 (RVV 3), one in Nayarit in 2022, and the most recent one in Quintana Roo in 2024 (RVV5 related to the common vampire bat Desmodus rotundus was detected).
Based on the last census by the National Institute of Statistics and Geography (INEGI) in 2020, Mexico’s pet population totaled 80 million, distributed as follows: 55% dogs, 20% cats, and 30% other species. This translates to four dogs and one cat for every ten inhabitants [17].
The Ministry of Health carries out rabies vaccination campaigns, administering around 18 million doses yearly, with 80% allocated for canines and 20% for cats (Supplementary Material) [18].
According to the INEGI, the state of Yucatan has seen an annual growth rate of 1.8%, resulting in a current total population of 2,320,898 inhabitants. Its pet population ranks third at the national level, with 2,074,423 pets. Of these, 44% are dogs, 23% are cats, and 33% belong to other species. Each year, about 143,000 rabies vaccines are administered in the state [17].
The population increase has resulted in the expansion of human activity into the Mayan forest to meet the needs of the growing population. In Mexico, the Mayan forest experiences an approximate loss of 80,000 hectares per year due to human modification of natural landscapes. Over the last two decades, the state of Yucatan has witnessed a loss of 266,613 hectares, with an average annual deforestation rate of 12,696 hectares [19]. Moreover, deforestation stemming from the conversion of forestland to human settlements from 2001 to 2021 has averaged 362 hectares per year [20].
In the Yucatan Peninsula, where the state of Yucatan is located, landscape fragmentation primarily stems from anthropogenic activities. These include rainfed agriculture, irrigated agriculture, cultivated grasslands, and nomadic agriculture often associated with remnants of secondary vegetation resulting from forestry practices, both industrial and domestic landscapes [21].
These practices, along with conventional tourism, have led to substantial modifications of coastal landscapes and urbanization in certain areas. Additionally, mining activities such as salt and limestone extraction, as well as petroleum and petroleum products production, have further contributed to the impact on the region’s landscapes [21].
Landscape fragmentation in Yucatan currently plays a significant role in ecological restoration and preservation of biodiversity, affecting processes such as regeneration, reintroduction, and species flows [21].
The translocation and introduction of animals to new geographic regions, driven by increased human global travel and commerce, serve as underlying factors for the emergence of infectious diseases. Additionally, introducing potential hosts into new geographic areas without the co-introduction of pathogens can also lead to disease emergence [22].
Thus, addressing the impacts of landscape fragmentation and human activities on both human and animal health necessitates the implementation of a One Health approach. This integrated methodology aims to sustainably balance and optimize the health of people, animals, and the environment by recognizing the interconnectedness and interdependence among human health, domestic and wild animals, plants, and the environment [23].
In this study, we conduct the antigenic and genetic characterization of RABV isolated from cats in the state of Yucatan. Additionally, we investigate potential factors, from a One Health perspective, that may be linked to the transmission of RABV from wildlife to cats.
2. Materials and Methods
2.1. Samples
The total number of registered cases (n=9) in cats (Felis catus) from the state of Yucatan was obtained from the positive sample bank of the InDRE Rabies Laboratory. Each sample contained the brain and brain stem of each individual.
2.2. Rabies Diagnosis
Rabies positivity was determined by the fluorescent antibody test [24], which uses a fluorescently labeled anti-rabies monoclonal globulin (Fujirebio, Diagnostics Inc.), in brain and brain stem tissue samples from all nine cases. Further confirmatory tests including antigenic characterization, RT-PCR, and sequencing were performed.
2.3. Antigenic Characterization
The antigenic characterization was performed using an indirect fluorescent antibody technique with a reduced panel of eight monoclonal antibodies (MAbs) (C1, C4, C9, C10, C12, C15, C18, C19) as previously described [25,26] This reduced panel can identify 11 reactivity patterns associated with different animals involved with rabies virus maintenance and transmission in Mexico and South America [27]. Antigenic characterization was applied directly to the brain smear (Table 1).
2.4. Viral RNA Extraction
Fifty milligrams (mg) of brain tissue were macerated in lysis buffer: Tris 1M, NaCl 5M, MgCl2 0.5M, and NP40 (Sigma). The buffer aids in the homogenization of brain tissues and the hypotonic lysis of the cells to free cytoplasmic RNA. The samples were subsequently centrifuged and the supernatant collected [28]. Finally, the QIAmp® Viral RNA mini kit was used to collect viral RNA following the manufacturer´s instructions.
2.5. RT-PCR and Sequencing
Retrotranscription coupled to PCR was performed using the SuperScript Platinum kit. The primers that were used: 550FW (5’-ATGTGYGCTAAYTGGAGYAC-3’) position: 647-666 of the genome of the Challenge Virus Strain (CVS) [29] and 304 (5´-CGCTCTAGATTGACGAAGATCTTGCTCAT-3´); (position 1514-1533) [30] generated an 886-bp amplicon overlapping on 200 nucleotides, obtaining nearly the complete coding sequence of the nucleoprotein gene. PCR products were visualized by electrophoresis on the Agilent Bioanalyzer.
The DNA sequencing was performed with the BigDye Terminator v3.1 Cycle Sequencing kit® employing the ABI PRISM® 3130xl Genetic Analyzer (Applied Biosystems, Foster City, CA) according to the manufacturer’s recommendations. Sequences obtained in both senses were edited in BioEdit Sequence Alignment Editor 7.2.5 [31] and BLAST (Basic Local Alignment Search Tool) analysis (https://blast.ncbi.nlm.nih.gov/Blast.cgi) [32] from NCBI was performed. Edited files were converted to FASTA format to be used in the phylogenetic analysis.
2.6. Phylogenetic Analysis
The phylogenetic analysis was based on 41 sequences involving reference sequences, and domestic and wild-living mammals from Yucatan. Multiple alignments of all sequences were performed with MEGA X [33] using the ClustalW Multiple Alignment application. The evolutionary model was Kimura 2P+G using the Neighbor-Joining method.
2.7. Geographic Data
Advanced Geographic Information Systems (GIS) tools in ArcGIS and the National Institute of Statistics and Geography (INEGI) were used to obtain detailed geographic data at the municipal level for the state of Yucatan. This process included the overlaying of case data and ecological variables (land cover change, type of land use, and vegetation).
Data on land cover change for the state of Yucatan between 2016 and 2020 were obtained from the National Forest Monitoring System-National Forestry Commission (SNMF-CONAFOR) website: https://snmf.cnf.gob.mx/cobertura-del-suelo/ [19,20].
2.8. Regression Analysis
A mixed-effects Poisson regression model was used to explore the association between feline rabies cases and sociodemographic (population size, social deprivation index) and ecological (land cover change) variables. A mixed effects model was employed to account for variability at the level of municipalities (random effects) when estimating the association of all other variables (fixed effects).
The population size for each municipality was based on the INEGI census of 2020. The 2020 social deprivation index was obtained from the National Council for the Evaluation of Social Development Policy (CONEVAL) [34]. For the land cover change variable, the percentage of the total area in each municipality that underwent land cover change was derived and subsequently used in the regression model. These analyses were performed using the glmer () function in R.
3. Results
3.1. Rabies Diagnoses
Between 2003 and 2022, all nine samples from cats in the state of Yucatan tested positive for RABV via the FAT method. Notably, between 2016 and 2022, an increase in sample positivity was detected (Figure 1).
3.2. Antigenic Characterization
Antigenic characterization revealed three virus variants. The oldest sample, dating back to 2003, corresponded to rabies virus variant (RVV) 1 (n=1; 11%), the canine variant that circulated in dogs. Among the positive samples obtained between 2017 and 2021, seven (77%) corresponded to atypical RVV. Additionally, the most recent sample from 2021 corresponded to RVV3 (n=1; 11%), which is related to the common vampire bat from the species Desmodus rotundus (Table 1).
3.3. Genetic Characterization and Phylogenetic Analysis
For the phylogenetic analysis, a dataset of 41 sequences, each approximately 800 bp in length and representative of rabies cases in the Yucatan Peninsula was included. Two main clades were identified that clustered the sequences of two rabies cycles, one related to terrestrial mammals and one related to bats (Figure 2). Phylogenetic reconstruction confirmed that the seven sequences corresponding to atypical RVV (751MxcatYuc17, 752MxcatYuc17, 753MxcatYuc17, 754MxcatYuc17, 1832MxcatYuc18, 613MxcatYuc19, and 399MxcatYuc21) clustered with a terrestrial cycle variant. This variant originated from the canine variant that was previously prevalent among dogs in Yucatan and became extinct due to mass vaccination programs.
Currently, the atypical variant is believed to have its reservoir in certain wild-living species, most likely skunks living within the Mayan jungle. Additionally, it has been detected in several mammals, such as the Lowland paca (Cuniculus paca), the white-tailed deer (Odocoileus virginianus), and the white-nosed coati (Nasua narica).
The virus sequences from cases 3419MxcatYuc03 and 400MxcatYuc2021 were grouped according to the antigenic characterization that was previously carried out on the samples. The virus from the 2003 case, (sequence 3419MxcatYuc03), clustered with viruses isolated from terrestrial mammals, but this one was grouped with the dog rabies cases with RVV1.
In this clade, the most recent case in dogs was detected in 2009, congruent with the history of elimination of this RVV1 in Yucatan. Since then, the atypical variant has been isolated most commonly from other terrestrial mammals (domestic and wild-living).
The virus sequence from the most recent case detected in 2024 (sequence 400MxcatYuc2021), clustered with the rabies cycle related to bats. In this clade, the sequences are related to the common vampire bat Desmodus rotundus, the main reservoir of rabies in Yucatan in the bat- related cycle. So far, this is the only bat species that is submitted for rabies diagnosis and the only one detected in other species submitted for rabies diagnosis in the state of Yucatan.
3.4. Geographical Data
3.4.1 Spatial Distribution of Cat Rabies Cases
To gain insights into the spatial distribution of cat rabies cases these were plotted at the municipality level. Out of the nine cases, four occurred in Muna, four in Merida, and one in the municipality of Cuncunul. Both Muna and Merida are predominantly urban areas, with populations of approximately 14,000 and 100,000 individuals, respectively. In contrast, Cuncunul is mostly rural, with a population of around 2000 individuals (Figure 3).
Soil degradation is caused by human activities such as agriculture, livestock, deforestation, urban and industrial development. Between 2002 and 2022 the state of Yucatan presents soil degradation (62.2%), in the case of physical degradation was 19.2% and chemical degradation was 55.1% being the highest at the national level, resulting in changes in soil structure and loss of vegetation cover. This shows that urbanization and industrial development have grown significantly in Yucatan [35,36,37]. As the areas of urbanization increase, the possibility of keeping domestic animals such as dogs and cats in these zones increases [22,38].
Changes in land cover varied across the state, with a higher percentage towards the central part and extending southeastward. The largest proportion of land cover change (8.2%) was observed in Dzan, followed by the rural municipalities of Chumayel (4.7%) and Kantunil (4.2%). While Dzan is a primarily urban municipality with approximately 6000 inhabitants, Chumayel and Kantunil are rural municipalities, with 3200 and 5500 inhabitants, respectively.
The municipalities of Merida and Muna where feline rabies cases were reported, are all predominantly urban and exhibited low (0-2%) to moderate (2-4%) changes in land cover.
3.5. Risk Factors for Feline Rabies Cases
Using a mixed-effects Poisson regression model, we investigated the association between feline rabies cases, and the sociodemographic factors population size and social deprivation index. Additionally, to assess the impact of environmental changes, we included the percentage change in land cover in the model. The model without social deprivation index had a lower AIC and is thus presented on Table 2. No statistically significant associations were observed (Table 2).
4. Discussion
Cats are explorative mammals [39] and efficient and abundant predators. Free-ranging cats can significantly impact ecosystems as generalist predators, exploiting a wide range of prey [40].
Since prey selection is correlated with prey availability [41,42], cats pose a threat to certain groups of animals such as bats, birds, and rodents, both in human-modified and natural landscapes [40,43,44]. Consequently, cats are recognized as some of the most harmful to biodiversity among invasive or commensal species [45,46,47].
In urban areas, domestic cats are the most abundant carnivores [48,49] and thus can prey on a large amount of wildlife every year [50]. Wildlife predation by cats originates from the heterogeneous conditions of free-ranging felines (owned or feral).
A study conducted in Italy revealed that predation on bats mainly occurred in rural areas or areas characterized by single buildings interspersed with large vegetation patches where cats are often allowed to roam outdoors in rural or sparsely populated urban areas [46]. Conversely, owned cats in more densely populated urban areas were frequently kept indoors, potentially explaining the lower numbers of bats caught in such environments, despite the possibility of a higher absolute density of cats [51].
In this study, infected cats were found in proximity to roads and were rescued by residents, indicating that they can be considered free-roaming cats, except for one domestic cat. According to reports from the residents, this particular domestic cat had kittens and established its shelter within the premises of a school, where it also sought refuge alongside other cats.
In all cases, cats displayed symptoms of aggression, inability to ingest water or food, and difficulty walking. Additionally, in three instances, there were reports of attacks on people and a pet (canine).
Although the cases have occurred in an urban context, it is not known with certainty where they were infected, in some cases the point of CST may have been in a wilder environment. Since the cats were rescued, the case report was registered in an urbanized locality.
In our study, an important factor that influences the fact that cats are becoming potential vectors of the rabies virus is that none of the cases were previously vaccinated, probably due to their young age (between one and three months old).
Consequently, it became necessary to apply post-exposure anti-rabies vaccination schedules to 34 persons, 18 of whom were bitten or scratched, along with a canine that was directly or indirectly exposed to RABV-infected cats.
The phylogenetic analyses conducted suggest the presence of two epidemiological cycles of the rabies virus in the state of Yucatan, both associated with the wild-living cycle of rabies. One of these cycles corresponds to the bat-related cycle, in which evidence thus far indicates that only hematophagous bats of the species Desmodus rotundus are responsible for enzootically sustaining the rabies virus in this state.
D. rotundus can transmit RABV to non-livestock species, including both wildlife and domestic animals in captive and free-range settings [52,53,54], in other non-hematophagous bat species, and wild-living terrestrial mammals known to play a role in both sylvatic and urban cycles of the disease [45,55,56,57].
The other cycle corresponds to the terrestrial cycle, which involves some species of skunk that appear to be responsible for maintaining and transmitting the rabies virus to other wild-living species and domestic animals such as dogs and cats [55,56]. Evidence suggests that the atypical antigenic variant originated from the dog variant that circulated in Yucatan but became extinct due to vaccination.
It is now sustained and transmitted by some species of wild mammals, most likely skunks. Its canine origin poses a serious risk of reintroducing the virus into canine populations if vaccination coverage is not maintained [58,59].
It is widely recognized that human modification of natural landscapes continues to rapidly transform ecosystems [60,61,62]. Anthropogenic disturbances such as land-use change, exploitation of natural resources, climate anomalies, and agricultural expansion are persistently altering biodiversity assemblages [63,64,65,66].
In Latin America, this phenomenon remains disproportionately significant, given the abundant ecosystem services provided by this region [67,68].
INEGI data revealed a steady growth in Yucatan’s urban and rural population between 2000 and 2020. During this period, the urban population increased by an average of 1.416% while the rural population increased by 14.39%. This growth translated into an overall increase in the state’s total population, which increased from 1,658,210 in 2000 to 2,320,892 in 2020.
This increase in population has been accompanied by a significant loss of forestland to human settlement. The current rate of forest loss is 362 hectares per year. If this rate continues, it is estimated that the total loss of forest hectares for the period 2000 to 2024 will be approximately 8,688 hectares.
According to the data obtained from INEGI, urbanization areas have been increasing in the state of Yucatan over the years. However, over the past 20 years, there has not been a significant increase in the chemical degradation of the territory [17].
The urbanization of natural areas can be considered a factor in promoting contact between wildlife and domestic animals. Importantly, wildlife can serve as reservoir of diseases, like rabies, which can affect domestic animals and potentially turn them into vectors of zoonotic diseases [22,38].
Relating this data to the cases of cat rabies, three cases were located in areas with extreme physical soil degradation, while the other six cases were in areas with slight chemical degradation. These results suggest a possible association between soil degradation and the occurrence of rabies cases in cats, highlighting the importance of considering the environmental setting when addressing the health and safety of domestic animals.
However, there has been a very significant increase in physical degradation, and when observing the maps of the location of rabies cases in cats, four of them occurred in the years 2003, 2018, 2019, and 2021 in the area of extreme degradation and the remaining five cases occurred in areas of light degradation.
The process of deforestation and urban expansion can have important implications for public health, especially with regard to the incidence of zoonotic diseases such as rabies. The increased proximity between human settlements and wildlife areas due to urban sprawl may increase the interaction between humans, domestic animals, and wildlife, thereby increasing the risk of transmission of rabies.
Sociodemographic factors, such as poverty and population density, have been associated with the transmission of canine and human rabies in certain contexts [69]. Additionally, ecological factors and attributes of the landscape, such as land cover, have been implicated in rabies transmission by wildlife species, like skunks and raccoons [70].
Changes in land cover can exacerbate landscape fragmentation, potentially facilitating pathogen spillover by increasing interactions between wild and domestic animals [71].
In our regression model, however, we did not detect any statistically significant associations between feline rabies cases and population size, social deprivation index, and land cover change. The absence of significance may stem from differences in how variables are defined between our study and others. The social deprivation index, for example, summarizes four aspects of social deprivation according to CONEVAL’s poverty measurement: educational backwardness, access to health services, access to basic services in housing, and quality and spaces in housing. Consequently, it is not a multidimensional measure of poverty [34].
Some studies have shown that the incidence and transmission of rabies correlate negatively with a country’s economic development, as this has an impact on access to basic health services, the availability of post-exposure treatments and case reporting [73,74,75,76]. However, further investigations would be necessary to consider whether, at least in the case of feline rabies in the state of Yucatan, economic development does not negatively impact rabies transmission. This could signify an epidemiological transition of rabies in Yucatan, especially as canine rabies is brought under control.
Alternatively, our findings may suggest that other unexamined factors may be relevant in understanding the complex dynamics of wildlife-domestic cat interactions and pathogen transmission. For instance, cat-hunting behavior has been suggested as a contributing factor to the elevated rate of feline rabies in Brazil [72]. Moreover, the limited number of reported rabies cases may have contributed to the lack of significant factors identified in our study. Further investigations are warranted to better understand the underlying factors driving these dynamics.
Understanding the mechanisms that underpin the cross-species transmission and host adaptation of RABV remains an important part of the ongoing goal to reduce and eliminate rabies [4].
Changes in predator-prey dynamics and disruptions in the trophic structure are unavoidable and expected consequences of changes to biodiversity [77]. Geographic ranges, prolonged exposure to prey during feeding, co-species roosting, and social behavior have numerous implications for disease transmission [78].
CST of rabies among terrestrial animals is a complex interaction that likely depends on animal susceptibility to the virus, animal population densities, animal behavior, niche overlap, landscape characteristics, human population distribution, environmental conditions, and other factors. These multivariate associations can be difficult to evaluate over large geographic areas, even within a single virus variant region [2].
Dogs and cats may have frequent opportunities for encounters with rabid animals and remain a critical barrier to human rabies exposure. The finding that CST rates increased in cats is a reminder of the importance of maintaining current vaccination status to protect animal health and prevent human exposure [2].
Understanding the mechanisms of host adaptation and interspecies transmission of RABV remains an important part of the ongoing goal to reduce and eliminate rabies [4]. A One Health approach to animal rabies prevention is crucial. This needs to be an interdisciplinary approach that considers host, pathogen, and environmental factors [79].
5. Conclusions
Our study sheds light on the emerging role of cats as a source of rabies virus transmission to humans in the state of Yucatan. We underscore the intricate relationship between land degradation, land cover change, and the incidence of cat rabies cases, potentially intensifying interactions between wildlife and domestic animals. This heightened proximity between human settlements and wild areas raises concerns about the heightened risk of zoonotic disease transmission, including rabies.
The One Health approach is invaluable for understanding the complexities of cross-species transmission, enabling the identification of crucial variables that demand attention for the implementation of effective prevention and control strategies.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
AMSG, JIOL performed experiments; NAC, AAS, conceived and designed the study; NAC, AAS, ILM, acquired the funding; AMSG, IM, JIOL performed analyses; NAC Wrote the paper. All co-authors revised and edited the paper.
Funding
This research received no external funding.
Data Availability Statement
All sequences generated in this study are available at Genbank with the following accession numbers: PP105582, PP105583, PP105584, PP105585, PP105586, PP105587, PP105588, PP105589 and PP105590.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Franka, R., Smith, T.G.; Dyer, J.L.; Wu, X.; Niezgoda M.; Rupprecht C.E. Current and future tools for global canine rabies elimination Antiviral Res. 2013. [CrossRef]
- Wallace, R.M.; Gilbert, A.; Slate, D.; Chipman, R; Singh, A.; Cassie, W., et al. Right place, wrong species: a 20-year review of rabies virus cross species transmission among terrestrial mammals in the United States. PLoS One. 2014, 9(10):e107539. [CrossRef]
- Marston D.A, Banyard A.C., McElhinney, L.M., Freuling, C.M., Finke, S., de Lamballerie X, et al. The lyssavirus host-specificity conundrum-rabies virus-the exception not the rule. Curr. Opin. Virol. 2017, 28:68–73.
- Bonnaud, E.M.; Troupin, C.; Dacheux, L.; Holmes, E.; Monchatre-Leroy, E.; Tanguy, M.; et al. Comparison of intra- and inter-host genetic diversity in rabies virus during experimental cross-species transmission. PLoS Pathog. 2019, 15(6):e1007799. [CrossRef]
- Mollentze, N.; Streicker, D.G.; Murcia, P.R.; Hampson, K.; Biek, R. Virulence Mismatches in Index Hosts Shape the Outcomes of Cross-Species Transmission. Proc. Natl. Acad. Sci. USA 2020, 117, 28859–28866. [Google Scholar] [CrossRef] [PubMed]
- Jacquot, M., Wallace, M.A, Streicker, D.G., Biek, R. Geographic Range Overlap Rather than Phylogenetic Distance Explains Rabies Virus Transmission among Closely Related Bat Species. Viruses. 2022, 14(11):2399. [CrossRef]
- Fleming, P.A. and Bateman, P.W. Novel predation opportunities in anthropogenic landscapes. Anim. Behav. 2018, 138: 145–55. [CrossRef]
- Guiden, P.W., Bartel, S.L., Byer, N.W., Shipley, A.A., Orrock, J.L. Predator-Prey Interactions in the Anthropocene: Reconciling Multiple Aspects of Novelty. Trends Ecol. Evol. 2019, 34(7):616-627. [CrossRef]
- Rupprecht C.E. 2023. Rabies in animals in: MSD Veterinary Manual. Merck & Co., Inc., Rahway, NJ, USA. https://www.msdvetmanual.com/nervous-system/rabies/rabies-in-animals?query=Rabies%20in%20animals accessed January 24 ,2024.
- Grobbelaar, A.A.; Blumberg, L.H.; Dermaux-Msimang, V.; Le Roux, C.A.; Moolla, N.; Paweska, J.T., et al. Human rabies associated with domestic cat exposures in South Africa, 1983-2018. J. S. Afr. Vet. Assoc. 2020, 91:e1–4. [CrossRef]
- Ribeiro, J.; Staudacher, C.; Martins, C.M.; Ullmann, L.S.; Ferreira, F.; Araujo, J.P.J, et al. Bat rabies surveillance and risk factors for rabies spillover in an urban area of Southern Brazil. BMC Vet. Res. 2018, 14:173. [CrossRef]
- Brunt, S.; Solomon, H.; Brown, K.; Davis, A. Feline and Canine Rabies in New York State, USA. Viruses 2021, 13, 450. [Google Scholar] [CrossRef] [PubMed]
- Herrera, D.J.; Cove, M.V.; McShea, W.J.; Decker, S.; Flockhart, D.T.T.; Moore, S.M. and Gallo, T. Spatial and temporal overlap of domestic cats (Felis catus) and native urban wildlife. Front. Ecol. Evol. 2022, 10:1048585. [CrossRef]
- SIRVERA Regional Information System for Epidemiological Surveillance of Rabies 2024. https://sirvera.panaftosa.org.br/ accessed January 24, 2024.
- Ellis, R. and Ellis C. Dog and cat bites. Am. Fam. Physician. 2014, 90(4): 239-243.
- Rupprecht C.E. 2022. Rabies in cats. MSD Veterinary Manual. Merck & Co., Inc., Rahway, NJ, USA. https://www.msdvetmanual.com/cat-owners/brain,-spinal-cord,-and-nerve-disorders-of-cats/rabies-in-cats. Accessed: January 24, 2024.
- INEGI https://www.inegi.org.mx/ Accesed January 24, 2024.
- Secretaría de Salud, 285. México alcanza 15 años sin registro de casos de rabia humana transmitida por perro. gob.mx. https://www.gob.mx/salud/prensa/285-mexico-alcanza-15-anos-sin-registro-de-casos-de-rabia-humana-transmitida-por-perro Accesed on January 24, 2024.
- SNMF, National Forest Monitoring System DEFORESTACIÓN. 2022. Accesed: January 20, 2024, de https://snmf.cnf.gob.mx/deforestacion/.
- CONAFOR, National Forestry Commission Deforestación bruta a nivel estatal en el periodo 2001 al 2021. (2022). https://tableros_snmf.cnf.gob.mx/deforestacion_estatal Accesed: January 20, 2024.
- Chiappy C, Gama L. Modifications and fragmentation of tropical geocomplexes of the Yucatan Peninsula. Universidad y Ciencia Num Esp I. 2004, 17–25.
- Daszak, P., Cunningham, A.A., Hyatt, A.D. Emerging infectious diseases of wildlife--threats to biodiversity and human health. Science. 2000, 21;287(5452):443-9. Erratum in: Science 2000, 10;287(5459):1756. [CrossRef]
- OHHLEP, One Health High-Level Expert Panel Adisasmito WB, Almuhairi S, Behravesh CB, Bilivogui P, Bukachi SA, et al. One Health: A new definition for a sustainable and healthy future. PLoS Pathog. 2022, 18(6): e1010537. [CrossRef]
- Dean, D.J.; Abelseth, M.K.; Atanasiu, W. The fluorescent antibody test. In Laboratory Techniques in Rabies; Meslin, F.-X., Kaplan, M.M., Koprowski, H., Eds.;World Health Organization: Geneva, Switzerland, 1996; pp. 88–95.
- Smith, J.S. Rabies virus epitopic variation: use in ecologic studies. Adv. Virus Res. 1989, 36, 215–253. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.S.; Orciari, L.A.; Yager, P.A.; Seidel, H.D.; Warner, C.K. Epidemiologic and historical relationships among 87 rabies virus isolates as determined by limited sequence analysis. J. Infect. Dis. 1992, 166, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.M.; Papo, S.; Rodriguez, A.; Smith, J.S. Antigenic analysis of rabies-virus isolates from Latin America and the Caribbean. J. Vet. Intern. Med., Series B. 1994, 41(1-10), 153-160. [CrossRef]
- McQuiston, J.H.; Yager, P.A.; Smith, J.S.; Rupprecht, C.E. Epidemiologic characteristics of rabies virus variants in dogs and cats in the United States. J. Am. Vet. Med. Assoc. 2001, 218, 1939–1942. [Google Scholar] [CrossRef] [PubMed]
- Markotter, W.; Kuzmin, I.; Rupprecht, C.E.; Randles, J.; Sabeta, C.T.; Wandeler, A.I.; Nel, L.H. Isolation of Lagos bat virus from water mongoose. Emerg. Infect. Dis. 2006, 12, 1913–1918. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.R.; Baron, E.J.; Pfaller, M.A.; Tenover, F.C.; Yolken, R.H.; Morgan, D.R. Manual of Clinical Microbiology (6th ed). Trends Microbiol. 1995, 3, 449. [Google Scholar]
- Hall, T.A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. 1999, Ser. 41:95-98.
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D. J. Basic local alignment search tool. J. Mol. Biol. 1990, 215(3), 403–410. [Google Scholar] [CrossRef] [PubMed]
- Kumar. S., Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35:1547-1549. [CrossRef]
- CONEVAL 2020 .National Council for the Evaluation of Social Development Policy. https://www.coneval.org.mx/Medicion/IRS/Paginas/Indice_Rezago_Social_2020. aspx Accesed on April 29, 2024.
- SEMARNAT Ministry of Environment and Natural Resources 2024 https://apps1.semarnat.gob.mx:8443/dgeia/informe18/tema/cap3.html Accesed on April 23, 2024.
- Bautista, F., Frausto, O., Ihl, T., Aguilar Y. An update soil map of The Yucatan State, Mexico: Geomorphopedological approach and WRB. Ecosist. y Rec.Agro. 2015. 2(6):303-315.
- CONABIO 2024. National Commission for the Knowledge and Use of Biodiversity. www.conabio.gob.mx Accesed on May 14, 2024.
- Cunningham, A.A., Daszak, P., Wood, J.L.N. One Health, emerging infectious diseases and wildlife: two decades of progress? Phil. Trans. R. Soc. B 2017, 372:20160167. [CrossRef]
- Machado, J.C.; Genaro, G. Comportamento exploratòrio em gatos domésticos (Felis silvestris catus Linnaeus, 1758): uma revisão. Archit. Vet. Sci. 2010, 15, 107–117. [Google Scholar] [CrossRef]
- Loyd, K. A. T.; Hernandez, S. M.; Carroll, J. P.; Abernathy, K.J.; Marshall, G. J. Quantifying free-roaming domestic cat predation using animal-borne video cameras. Biol. Conserv. 2013, 160, 183–189. [Google Scholar] [CrossRef]
- Liberg, O. Food Habits and Prey Impact by Feral and House-Based Domestic Cats in a Rural Area in Southern Sweden. J. Mammal.1984, 65(3):424-432.
- Molsher, R.; Newsome, A.; Dickman, C.R. Feeding ecology and population dynamics of the feral cat in relation to the availability of prey in central-eastern New SouthWales. Wildl.Res. 1999, 26, 593–607. [Google Scholar] [CrossRef]
- Ancillotto, L.; Serangeli, M.T.; Russo, D. Curiosity killed the bat: Domestic cats as bat predators. Mamm. Biol. 2013, 78, 369–373. [Google Scholar] [CrossRef]
- Welch, J.N. and Leppanen C. The threat of invasive species to bats: a review. Mamm. Rev. 2017, 47 (4): 277-290. [CrossRef]
- Lowe, S., Browne, M., Boudjelas, S., De Poorter, M. 2004. 100 of the World’s worst invasive alien species. A selection from the Global Invasive Species Database. The Invasive Species Specialist Group (ISSG) a specialist group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN).
- Loss, Scott R. Will, Tom, Marra, Peter P. The impact of free-ranging domestic cats on wildlife of the United States. Nat. Comm. 2013, 4(1). [CrossRef]
- Doherty, T.S., Glen, A.S., Nimmo, D.G., Ritchie, E.G., Dickman, C.R. Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. U S A. 2016, 113(40):11261-11265. [CrossRef]
- Coleman, J. S., & Temple, S. A. Rural Residents’ Free-Ranging Domestic Cats: A Survey. Wildl. Soc. B. 1993, 21(4), 381–390. http://www.jstor.org/stable/3783408.
- Lepczyk, C.A.; Mertig, A.G.; Liu, J. Landowners and cat predation across rural to- urban landscapes. Biol. Conserv. 2003, 115, 191–201. [Google Scholar] [CrossRef]
- Woods, M., Mcdonald, R.A, and Harris, S. Predation of wildlife by domestic cats Felis catus in Great Britain. Mammal. Review 2003, 33(2)0305-1838. [CrossRef]
- Natoli, E. Spacing pattern in a colony of urban stray cats (Felis catus L.) in the historic center of Rome. Appl. Anim. Behav. Sci. 1985, 14, 289–304. [Google Scholar] [CrossRef]
- Gonçalves F, Galetti M, Streicker DG. Management of vampire bats and rabies: a precaution for rewilding projects in the neotropics. Perspec.t Ecol. Conserv. 2021;19:37–42. [CrossRef]
- Schneider MC, Aron J, Santos-Burgoa C, Uieda W, Ruiz-Velazco S. Common vampire bat attacks on humans in a village of the Amazon region of Brazil. Cad. Saude Publ. 2001, 17:1531–1536. [CrossRef]
- Schneider MC, Romijn PC, Uieda W, Tamayo H, da Silva DF, Belotto A, da Silva JB, Leanes LF. Rabies transmitted by vampire bats to humans: an emerging zoonotic disease in Latin America? Rev Panam Salud Publ. 2009, 25:260–269. [CrossRef]
- Delpietro HA, Lord RD, Russo RG, Gury-Dhomen F. Observations of sylvatic rabies in northern Argentina during outbreaks of paralytic cattle rabies transmitted by vampire bats (Desmodus rotundus) J. Wildl. Dis. 2009, 45:1169–1173. [CrossRef]
- Favoretto SR, Carrieri ML, Cunha EMS, Aguiar EAC, Silva LHQ, Sodré MM, Souza MCAM, Kotait I. Antigenic typing of Brazilian rabies virus samples isolated from animals and humans, 1989–2000. Rev. Inst. Med. Tro.p Sao Paulo. 2002, 44:91–95. [CrossRef]
- Kobayashi Y, Sato G, Shoji Y, Sato T, Itou T, Cunha EMS, Samara SI, Carvalho AAB, Nociti DP, Ito FH, Sakai T. Molecular epidemiological analysis of bat rabies viruses in Brazil. J. Vet Med Sci. 2005, 67:647–652. [CrossRef]
- Garcés-Ayala, F.; Aréchiga-Ceballos, N.; Ortiz-Alcántara, J.M.; González-Durán, E.; Pérez-Agüeros, S.I.; Méndez-Tenorio, A.; Torres-Longoria, B.; López-Martínez, I.; Hernández-Rivas, L.; Díaz-Quiñonez, J.A.; Ramírez-González, J.E. Molecular characterization of atypical antigenic variants of canine rabies virus reveals its reintroduction by wildlife vectors in southeastern Mexico. Arch. Virol. 2017, 162(12):3629-3637. [CrossRef]
- Puebla-Rodríguez, P.; Almazán-Marín, C.; Garcés-Ayala, F.; Rendón-Franco, E.; Chávez-López., S.; Gómez-Sierra, M.; Sandoval-Borja, A.; Martínez-Solís, D.; Escamilla-Ríos, B.; Sauri-González, I.; Alonzo-Góngora, A.; López-Martínez, I.; Aréchiga-Ceballos, N. Rabies virus in white-nosed coatis (Nasua narica) in Mexico: What do we know so far?.Front. Vet. Sci. 2023, 10:1090222. [CrossRef]
- Barlow, J.; Lennox, G.D.; Ferreira, J;, Berenguer, E.; Lees, A.C.; Nally, R.M.; Thomson, J.R.; de Ferraz, S.F.B.; Louzada, J.; Oliveira, V.H.; Parry, L.; de Castro, R.; Solar, R.; Vieira, I.C.G.; Aragão, L.E.O.C.; Begotti, R.A.; Braga, R.F.; Cardoso, T.M.; de Oliveira, R.C, Souza, C.M. Jr.; Moura, N.G.; Nunes, S.S.; Siqueira, J.V.; Pardini, R.; Silveira, J.M.; Vaz-de-Mello, F.Z.; Veiga, .RC.S.; Venturieri, A.; Gardner, T.A. Anthropogenic disturbance in tropical forests can double, biodiversity loss from deforestation. Nature. 2016, 535:144–147. [CrossRef]
- Ellis, E.C.; Klein, G. K, Siebert, S.; Lightman, D.; Ramankutty, N. Anthropogenic transformation of the biomes, 1700 to 2000. Glob. Ecol. Biogeogr. 2010, 19:589–606. [CrossRef]
- Grantham, H.S.; Duncan, A.; Evans, T.D.; Jones, K.R.; Beyer, H.L.; Schuster, R.; Walston, J.; Ray, J.C.; Robinson, J.G.; Callow, M.; Clements, T.; Costa, H.M.; DeGemmis, A.; Elsen, P.R.; Ervin, J.; Franco, P;, Goldman, E., Goetz, S., Hansen, A., Hofsvang, E., Jantz, P., Jupiter, S., Kang, A., Langhamme,r P., Laurance, W.F., Lieberman, S., Linkie, M., Malhi, Y., Maxwell, S., Mendez, M., Mittermeier, R., Murray, N.J., Possingham, H., Radachowsky, J., Saatchi, S., Samper, C., Silverman, J., Shapiro, A., Strassburg, B., Stevens, T., Stokes, E., Taylor, R., Tear, T., Tizard, R., Venter, O., Visconti, P., Wang, S., Watson, J.E.M. Anthropogenic modification of forests means only 40% of remaining forests have high ecosystem integrity. Nat. Commun. 2020, 11:5978. [CrossRef]
- Bengtsson, J.; Ahnström, J.; Weibull, A.C. The effects of organicagriculture on biodiversity and abundance: a meta-analysis. J .Appl. Ecol. 2005, 42:261–269. [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15:365–377. [CrossRef]
- Chaudhary, A., Mooers, A.O. Terrestrial vertebrate biodiversity loss under future global land use change scenarios. Sustainability 2018, 10:2764. [CrossRef]
- Hooper, D.U., Adair, E.C., Cardinale, B.J., Byrnes, J.E., Hungate, B.A., Matulich, K.L., Gonzalez, A., Duffy, J.E., Gamfeldt, L., O’Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature. 2012, 486(7401):105-8. [CrossRef]
- Nobre, C.A.; Sampaio, G.; Borma, L.S.; Castilla-Rubio, J.C.; Silva, J.S.; Cardoso, M. Land-use and climate change risks in the Amazon and the need of a novel sustainable development paradigm. Proc. Natl .Acad. Sci. USA. 2016, 113:10759–10768. [CrossRef]
- Salazar, A.; Baldi, G.; Hirota, M., Syktus, J.; McAlpine, C. Land use and land cover change impacts on the regional climate of non-Amazonian South America: a review. Glob. Planet. Change. 2015, 128:103–119. [CrossRef]
- Mshelbwala, P.P.J., Soares Magalhães, R., Weese, J.S., Ahmed, N.O., Rupprecht, C.E., Clark, N.J. Modelling modifiable factors associated with the probability of human rabies deaths among self-reported victims of dog bites in Abuja, Nigeria. PLoS Negl. Trop. Dis. 2023 21;17(2):e0011147. [CrossRef]
- Smith, D.L., Waller, L.A., Russell, C.A., Childs, J.E., Real, L.A. Assessing the role of long-distance translocation and spatial heterogeneity in the raccoon rabies epidemic in Connecticut. Prev. Vet. Med. 2005, 71(3-4):225-40. [CrossRef]
- Borremans, B., Faust, C., Manlove, K.R., Sokolow, S.H., Lloyd-Smith, J.O. Cross-species pathogen spillover across ecosystem boundaries: mechanisms and theory. Philos. Trans. R. Soc. Lond. B. Biol Sci. 2019, 374(1782):20180344. [CrossRef]
- de Lima, J.S., Mori, E., Kmetiuk, L.B., Biondo, L.M., Brandão, P.E., Biondo, A.W., Maiorka, P.C. Cat rabies in Brazil: a growing One Health concern. Front. Public. Health. 2023, 11:1210203. [CrossRef]
- Yu, J.; Xiao, H.; Yang, W.; Dellicour, S.; Kraemer, M.U.G.; Liu, Y.; Cai, J.; Huang, Z.X.Y.; Zhang, Y.; Feng, Y.; et al. The Impact of Anthropogenic and Environmental Factors on Human Rabies Cases in China. Transbound. Emerg. Dis. 2020, 67, 2544–2553. [Google Scholar] [CrossRef] [PubMed]
- Arias-Orozco, P.; Bá Stida-Gonzá Lez, F.; Cruz, L.; Villatoro, J.; Espinoza, E.; Zárate-Segura, P.B.; Recuenco, S.; Díaz, S.S.; Esquina, M.; De, P.; et al. Spatiotemporal Analysis of Canine Rabies in El Salvador: Violence and Poverty as Social Factors of Canine Rabies. PLoS ONE 2018, 13, e0201305. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, R.K.; Hanlon, C.A.; Goodin, D.G.; Davis, R.; Moore, M.; Moore, S.; Anderson, G.A. Bayesian Spatiotemporal Pattern and Eco-Climatological Drivers of Striped Skunk Rabies in the North Central Plains. PLoS Negl. Trop. Dis. 2016, 10, e0004632. [Google Scholar] [CrossRef] [PubMed]
- Subedi, D.; Chandran, D.; Subedi, S.; Acharya, K.P. Ecological and Socioeconomic Factors in the Occurrence of Rabies: A Forgotten Scenario. Infect. Dis. Rep. 2022, 14, 979–986. [Google Scholar] [CrossRef]
- Guiden, P.W., Bartel, S.L., Byer, N.W., Shipley, A.A., Orrock, J.L. Predator-Prey Interactions in the Anthropocene: Reconciling Multiple Aspects of Novelty. Trends Ecol. Evol. 2019, 34(7):616-627. [CrossRef]
- Brown, N.; Escobar, L.E. A review of the diet of the common vampire bat (Desmodus rotundus) in the context of anthropogenic change. Mamm. Bio.l 2023, 103, 433–453. [Google Scholar] [CrossRef]
- Nadal, D., Beeching, S.; Cleaveland, S.; Cronin, K.; Hampson, K.; Steenson, R.; Abela-Ridder, B. Rabies and the pandemic: Lessons for One Health. Trans. R. Soc. Trop. Med. Hyg. 2022, 116:197–200. [CrossRef]
Figure 1.
Time series of laboratory-confirmed rabies cases in cats in the state of Yucatan, 2003 to 2022.
Figure 1.
Time series of laboratory-confirmed rabies cases in cats in the state of Yucatan, 2003 to 2022.
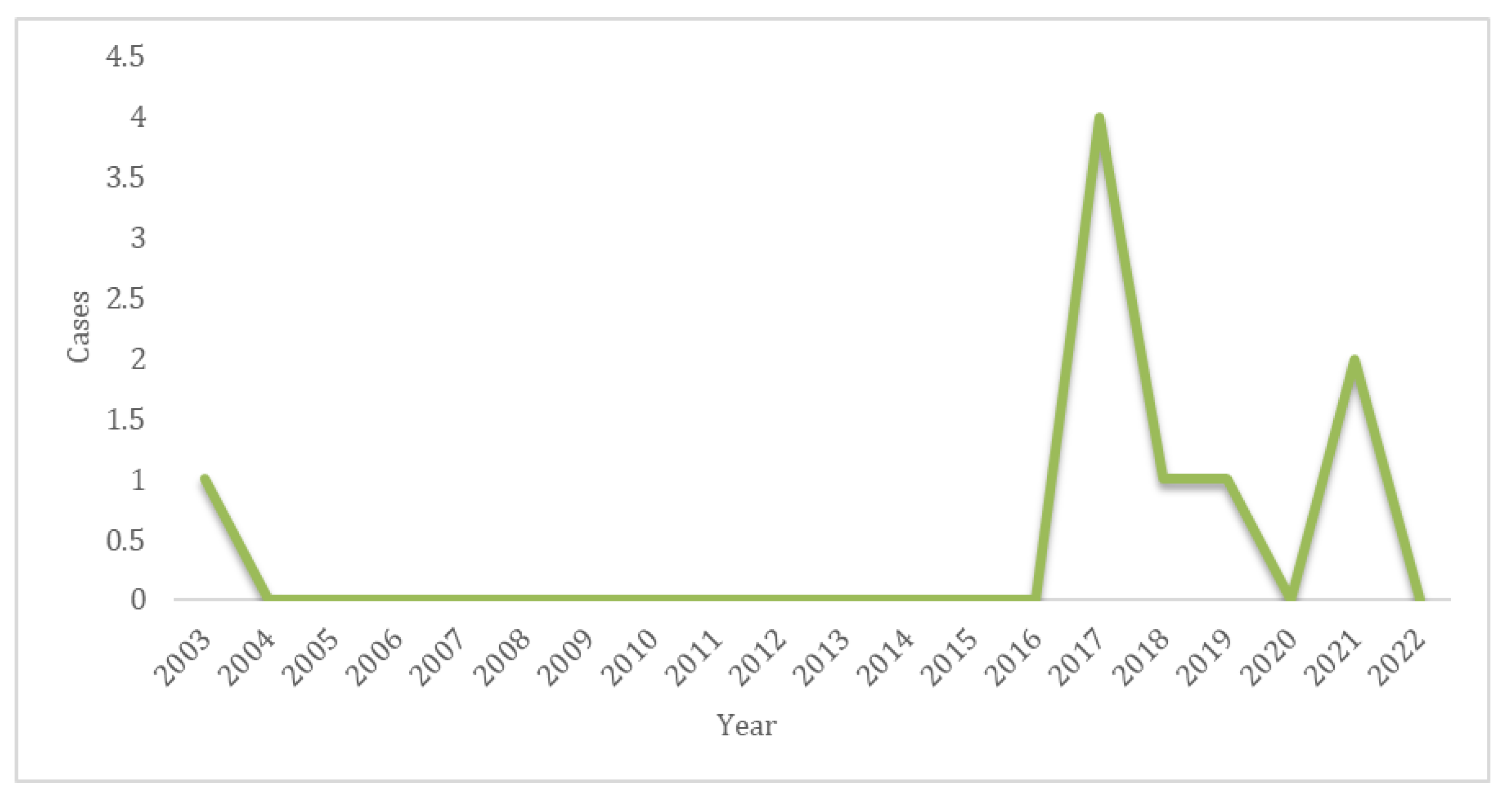
Figure 2.
The phylogenetic reconstruction of the nine positive rabies cases in cats (F. catus) in Yucatan, was based on a dataset of 41 RABV partial nucleoprotein sequences belonging to RVV of the Yucatan Peninsula. The Neighborg joining analysis was carried out using the evolutionary model Kimura 2P+G.
Figure 2.
The phylogenetic reconstruction of the nine positive rabies cases in cats (F. catus) in Yucatan, was based on a dataset of 41 RABV partial nucleoprotein sequences belonging to RVV of the Yucatan Peninsula. The Neighborg joining analysis was carried out using the evolutionary model Kimura 2P+G.
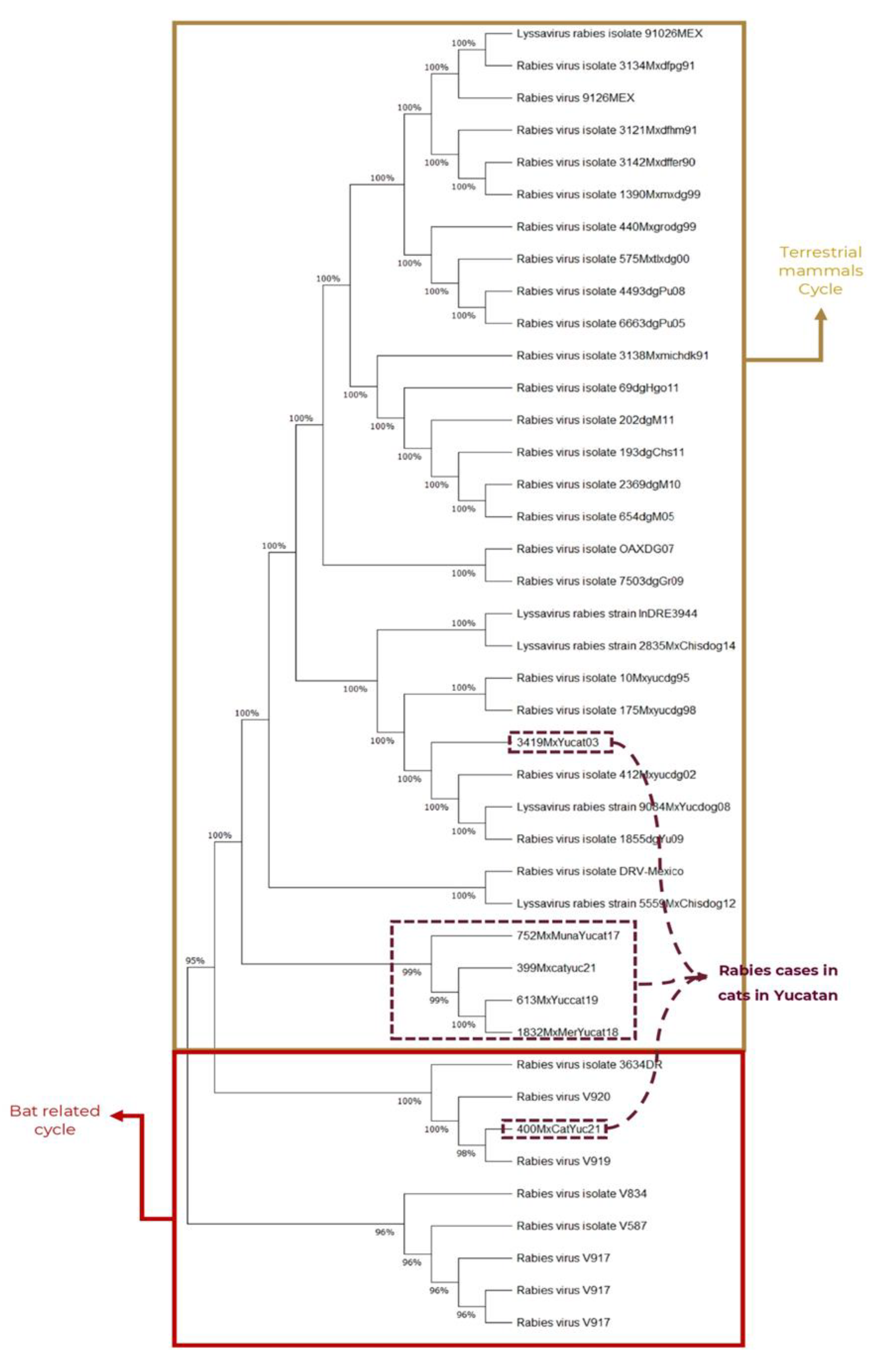
Figure 3.
Geographic distribution of cat rabies cases (n=9) in Yucatan at the municipality level. Panel A. Types of vegetation in the state and the rabies virus antigenic variants detected. Panel B. Population size and the rabies cases in cats (green triangles).
Figure 3.
Geographic distribution of cat rabies cases (n=9) in Yucatan at the municipality level. Panel A. Types of vegetation in the state and the rabies virus antigenic variants detected. Panel B. Population size and the rabies cases in cats (green triangles).
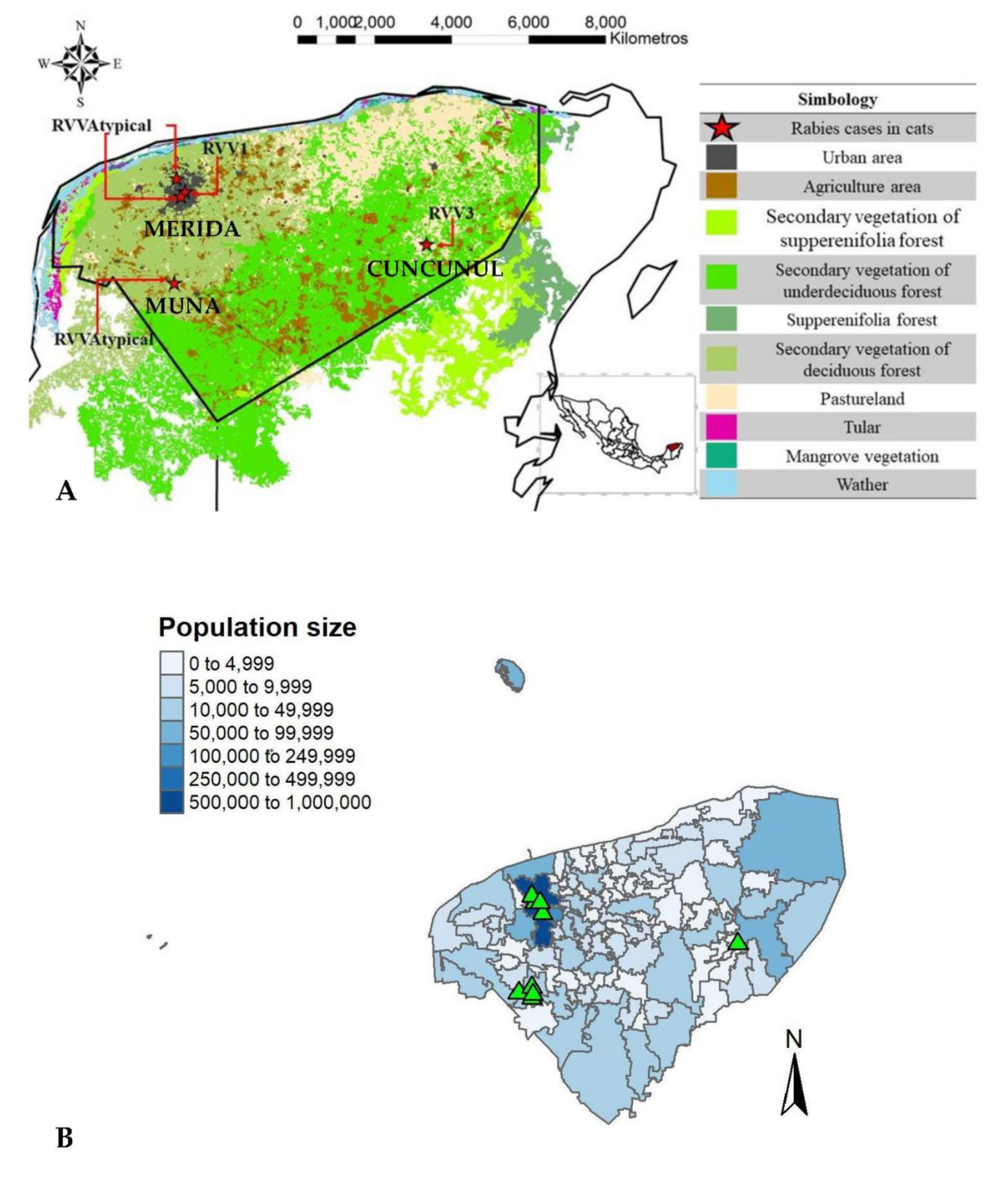

Table 1.
Summary of rabies cases in cats in the state of Yucatan.
| # | Case | Year | Municipality | Antigenic RVV |
Sequence Name | GenBank Accesion Number |
|---|---|---|---|---|---|---|
| 1 | 3419 | 2003 | Merida | RVV1 | 3419MxcatYuc03 | PP105582 |
| 2 | 751 | 2017 | Muna | Atypical | 751MxcatYuc17 | PP105583 |
| 3 | 752 | 2017 | Muna | Atypical | 752MxcatYuc17 | PP105584 |
| 4 | 753 | 2017 | Muna | Atypical | 753MxcatYuc17 | PP105585 |
| 5 | 754 | 2017 | Muna | Atypical | 754MxcatYuc17 | PP105586 |
| 6 | 1832 | 2018 | Merida | Atypical | 1832MxcatYuc18 | PP105587 |
| 7 | 613 | 2019 | Merida | Atypical | 613MxcatYuc19 | PP105588 |
| 8 | 399 | 2021 | Merida | Atypical | 399MxcatYuc21 | PP105589 |
| 9 | 400 | 2021 | Cuncunul | RVV3 | 400MxCatYuc21 | PP105590 |
Table 2.
Mixed-effects Poisson regression model with random-effects for municipality.
| Factor | Estimate | IRR | p- value |
|---|---|---|---|
| Population size | 0. 974 | 2.649 | 0.159 |
| % Land cover change | -0.304 | 0.738 | 0.863 |
IRR= incidence rate ratio.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



