Submitted:
07 April 2024
Posted:
23 April 2024
You are already at the latest version
Abstract
Melatonin is an important hormone with a methoxy-indole structure. It is secreted regularly by the pineal gland of mammals. It takes part in many chemical, biological, and physiological activities of the body. Melatonin, acts as a basic hormone, and is responsible for circadian rhythm regulation and sleep control. It has been shown to occur in the highest concentration in organisms during the night times. In living organisms, many activities that have physiological, biological, hormonal, and psychological characteristics follows a certain rhythm. Body temperature, hormone levels, sleep-wakefulness, and some other cognitive functions in humans also change with the circadian rhythm. Its synthesis and release begin with darkness and end with light. This review aims to explore the functions of melatonin such as its stimulating effects on antioxidant enzymes, reducing lipid peroxidation, and protecting brain tissue from oxidative damage. It also has properties that stop the proliferation of cancer cells and prevent tumor growth. Melatonin, which was previously known only as an animal hormone, has been found to also exist in plants. Research has shown that melatonin is present in almost all plant species analyzed to date and that it performs functions similar to those in animals. In plants, it plays a vital role in dealing with various abiotic and biotic stresses. Melatonin acts as an important antioxidant in post-harvest technology and extends the post-harvest life of fruits and vegetables. It also takes part in the regulation of stress-specific genes and the activation of pathogenesis-related proteins and antioxidant enzymes. The most prominent roles of melatonin in plants include regulating circadian rhythm, growth, and development, and increasing tolerance to a wide range of environmental stress factors. The review reveals the effects of melatonin as an extremely important secondary metabolite, on human, animal, and plant metabolism. The use of PRISMA guidelines was used for reporting, search, selection, and synthesis. Information up to 2024 was searched from search engines for the occurrence, nature and function of melatonin. Key words such as structural function, hormonal function, secretion and metabolism were used in the search of publications.
Keywords:
melatonin
; immunity
; cancer
; circadian rhythm
; growth and development
; aging
; plant
; seed
; seed viability
1. Introduction
The pineal gland was identified by Herophilus in the third century BC.450 years later, Galen revealed that this is a tiny endocrine gland in the middle of the brain tissue and has a similar function to the lymph nodes [1,2,3]. The pineal gland is a red-gray gland located in the central part of the brain, on the roof of the third ventricle of vertebrates, that produces the hormone melatonin, a derivative of serotonin that affects sleep-wake modulation patterns and seasonal functions.[4]. Galen, (129 – 216 AD) used the Latin word "conarium" to describe the human pineal gland, which he likened to the top of a pine tree (Emre, 1993). Although it is very small compared to body size, it is the second organ that receives the most blood after the kidneys. Although many peptide hormones are synthesized in the pineal gland, the main hormone is melatonin (2,5,6). Melatonin hormone was first discovered by dermatologist Lerner in 1958 and its function was revealed [7].
The formula of melatonin (N-acetyl-5 methoxytryptamine), an indoleamine tryptophan derivative shown in Figure 1, was reported in bovine pineal gland tissue in the late 1950s and became an important discovery for the scientific world [7]. In general, it is the main hormone secreted by the pineal gland. Secondary sources of this hormone are other structures such as the retina, intestine, skin, platelets, and bone marrow [8,9]. This methoxy indole compound is synthesized from serotonin. The melatonin has been reported to lightens the skin color, hence its name which is a combination of the Latin words "melas" meaning black, and "tosos" meaning wife [10,11]. It is found in all living organisms, especially vertebrates and mammals and detected in the blood [12] (Figure 1).
Melatonin was first found in algae and was later thought to be in the structures of different plants [13]. As a result of the studies, it has been determined that the roots, leaves, seeds and fruits of many plant species contain significant levels of melatonin [3,14].
Additionally, melatonin has been found to be synthesized by subphyla including bacteria[15,16,17]. Melatonin hormone, which is secreted from the pineal gland and known to increase in the dark, is a powerful antioxidant and is known to play a role in important functions such as cell renewal, strengthening the immune system, sleep rhythm and body temperature regulation. The circadian rhythm in melatonin release becomes irregular with age. It has been determined that the decrease in melatonin production is effective not only in physiological conditions but also in some clinical disorders such as mood disorders [18]. The metabolism of melatonin starts as tryptophan, an amino acid found in plants, animals, algae, and yeast. Tryptophan is first converted into serotonin which is then converted to melatonin in pinealocytes [10]. Tryptophan hydroxylase enzyme uses tetrahydrobiopterin (BH4) and O2 as cofactors, and vitamin B6 serves as a coenzyme in this reaction [19]. Although, melatonin is synthesized from the tryptophan precursor in different ways, the most commonly accepted way is; It is in the form of Tryptophan, Tryptamine, Serotonin, 5-methoxytryptamine, Melatonin and a wide variety of enzymes are involved during this synthesis [20].
Research as shown that melatonin hormone is secreted more during long nights in winter and less in summer. The functiom of the hormone Melatonin, is regulation of human biochemical metabolism and physiological rhythm while alleviating related disorders such as jet lag and insomnia, scavenging free radicals, and strengthening the immune system. Melatonin has been implecated as the cause of the phenomenon of jet lag. Studies have shown that people who work at night get sick more quickly and are more likely to get cancer. Additionally, this compound has anti-aging, anti-cancer effects as well as neuroprotective effects. It has also been shown that it has effects in the control of chronic diseases such as heart, diabetes and obesity [18,21].
There are many important factors that ensure the synthesis and release of melatonin, the most important of which is light. In other words, the light-dark, day-night rhythm provides control of the synthesis and release of the compound. This regulation mechanism is defined as “photo neuroendocrine control”. Light intensity is especially important in measuring the suppression level of melatonin, and as the brightness of the light stimulus increases, the suppression power increases [22,23]. Melatonin shows its effect through specific receptors located in target tissues.
In addition to the pineal gland, receptors have been reported to be present in most peripheral tissues such as the retina, brain, pituitary gland, spleen, retinal erythrocyte, leukocyte, thyroid gland, thymus, placenta, endometrium and gastrointestinal tract [11,12,24].
This review aims to reveal the effects of melatonin on human, animal, and plant metabolism, based on all its functional properties.
To source out the results in this review, a Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines were formulated using the online scholar databases such as google scholar, BMC, science direct and PubMed. Information from this article was based on development of keywords such as structural function, hormonal function, secretion and metabolism. An overview of scientific publications (research articles) until March 2024 and those focused on structural function in human, animals, chemistry, and in vitro systems published in English language were used. A total of 356 research articles were screened by the authors (ARN, AKY, N.Ç. and H.Ö.D) to assess their relevance on the objective of the proposed review paper. Therefore, all authors’ separately recorded ‘yes’ or ‘no’ as whether that article should be included. Later after the assessment, a total of 134 articles were included as of the data based on the motive of the study. In cases where the articles would meet a set criteria of relevance full paper article were downloaded and saved in the Menderly® referencing tool. Selection criteria of articles were constructed on the compound, melatonin. However, to a great extent, articles without adequate data, reproducible results and methods, and article written in other languages apart for English were excluded. In addition, in cases where results in different articles would show a contradicting fact, consultation with other colleagues whom are expects in this filed was done. Data extraction from the reviewed literature followed a standardized procedure. The extracted data included: materials and methods, results, interpretations and discussions. Data were extracted autonomously by the authors (ARN, AKY, XXXXX), and incongruities were resolved through constant discussions. Therefore, the use of PRISMA guidelines were used for reporting, search, selection, and synthesis.
2. The Importance of Melatonin in Humans and Animals
2.1. Melatonin Functions
Melatonin has exhibited anti-oxidative, metabolism regulator, immune/defense system booster, melanogenesis regulation, nervous system regulation, bioenergetic (energy) regulation, anti-inflammatory properties, sleep regulation, circadian (periodic behavior) rhythm regulation, onco-static (preventing tumor development) effects [24,25]. Melatonin is a powerful heart-healthy and exerts significant beneficial effects on cardiological disorders and many other health problems [26,27]. Additionally, melatonin has been shown to play a role in regulating human mood, sleep and wake cycles and reproduction. It is also known to reduce anxiety and stimulate immunity [28]. Melatonin is a lipophilic and hydrophilic substance and quickly passes into the blood and other body fluids before being stored in the body. Since it is soluble in water and lipid phases, it quickly passes into all intracellular components and has a potential antioxidant effect by effectively protecting the cell membrane, organelles and nucleus from free radical damage [29,30]. At least 70% of melatonin in the blood binds to albumin and is metabolized first in the liver and secondly in the kidneys to 6-hydroxy melatonin sulfate and 6-hydroxy melatonin glucuronide, and the resulting metabolites are excreted in the urine [5,19].
Melatonin has remarkable importance as a growth and development regulator in plants. It has important effects on plants such as increasing tolerance to environmental factors, protecting chlorophyll, and supporting root regeneration [31]. The effect of melatonin on its role in plant physiology has been demonstrated by many studies. It acts as an antistress agent against abiotic stress factors such as melatonin and biotic stress factors such as microbial infections. It also regulates plant growth and photosynthesis [28].
2.2. Factors Affecting Melatonin Synthesis and Release
Although many factors affect the synthesis and release of melatonin, the most important factor affecting this situation is light. This hormone, dark-light; controls the regulation of day-night rhythm. This mechanism is known as “photo neuroendocrine control”. Although light synchronizes the circadian rhythm in pineal melatonin synthesis, it can also severely suppress melatonin synthesis. Exposure of the eyes to artificial light during the night also causes a rapid decrease in melatonin synthesis and release. The intensity of light is an important factor in determining the level of suppression of melatonin, and as the brightness of the light increases, the level of suppression also increases [11,32,33,34].
2.3. Melatonin Biosynthesis and Sources
Melatonin is produced from tryptophan taken from the bloodstream and converted into serotonin. Serotonin is converted into melatonin in a two-step process as a result of the activity of two enzymes. Serotonin-N-acetyl transferase and hydroxyindole-O-methyl transferase, which are limiting enzymes for melatonin synthesis, are involved. mRNAs encoding these enzymes are synthesized in the pineal region depending on the day/night rhythm [35].
When it gets dark, photoreceptors in the retina are stimulated and this information is transmitted to the suprachiasmatic nucleus (SCN) in the hypothalamus via the monosynaptic retinohypothalamic pathway. This system works autonomously, centrally and circadianly. Incoming signals pass to the paraventricular nucleus and from there they combine in the medial forebrain stem and reticular formation and go to the intermediolateral nucleus of the spinal cord. The signals received by the preganglionic adrenergic nerves of the sympathetic nervous system are carried to the upper cervical ganglion, and the final sympathetic information from here reaches the pineal gland. Sympathetic nerves rhythmically secrete norepinephrine in the terminals of the parenchymal cells of the pineal gland, depending on daily light-dark changes. This substance is recognized by ß-adrenergic receptors in the pine-alocyte membrane and accordingly the cyclic nucleotide system is stimulated. [36,37,38]. In mammals, the pineal gland acts as a neuroendocrine transformer, converting electrical impulses into hormonal secretions. It is in high concentration during the night because melatonin synthesis and release are affected by light. In the dark, the stimulus from the superior sympathetic ganglion via β-adrenergic post-ganglionic sympathetic fibers increases its synthesis, while in bright conditions its synthesis is inhibited [33,39,40,41].
Melatonin shows its effect in animal cells mostly through membrane-bound G-protein coupled receptors [42]. In mammals, three nuclear receptors of melatonin, called MT1, MT2, and MT3, have been identified [43,44]. Melatonin receptors are found in many organs, especially in the brain, cardiovascular system, aorta, heart ventricular wall, cerebral and coronary arteries, gallbladder, liver, retina, parotid gland, appendix, cecum, colon, skin, pancreas, platelets, and immune system. [45,46]. The majority of melatonin receptors are typically found in the jejunal and colonic mucosa of the gastrointestinal tract [36]. Melatonin shows its effect with the participation of various molecular pathways. The most described pathways involve the activation of specific membrane receptors, namely high affinity (ML1) and low affinity (ML2) domains [47,48]. ML1 acts directly on target cells or through G-protein coupled receptors. There are three types of receptors in the synthesis of melatonin: MT1, MT2 and MT3: MT1 and MT2 show G-protein-mediated effects. While MT1 has high affinity, MT2 has low affinity [49]. The third type belonging to the quinone reductase family is the MT3 protein [50]. Depending on their chromosome location and molecular structure, MT1 and MT2 melatonin receptors are defined as different subtypes [43,44,51,52,53]. MT1 receptors are located in the pars tuberalis part of the pituitary, and MT2 receptors are located in the retina. Additionally, both receptors were found in the suprachiasmatic nucleus of the hypothalamus. The presence of the MT3 receptor has also been demonstrated in amphibians and birds. It is thought that circadian and reproductive effects occur through the MT1 receptor, and the MT2 receptor is associated with dopaminergic functions in the brain and retina.
As expressed in Figure 2, the amino acid tryptophan is the precursor of melatonin synthesis and is taken from the plasma by the pineal gland [54]. Tryptophan is hydroxylated to 5-hydroxytryptophan by tryptophan hydroxylase (Figure 2). Vitamin B6 serves as a coenzyme in this reaction. The resulting 5-hydroxytryptophan is a naturally occurring intermediate metabolite. 5-hydroxytryptophan can easily cross the blood-brain barrier and turns into serotonin by losing its carboxyl group through the L-amino acid decarboxylase enzyme. Serotonin cannot cross the blood-brain barrier. However, it is then acetylated by the N-acetyltransferase enzyme and converted into N acetylserotonin, which in turn is converted into melatonin by the hydroxyindole-O-methyl transferase enzyme (Figure 2) [5,11,23,28].
Melatonin has high lipid and water solubility, which facilitates passage through cell membranes. After being released into circulation, it passes into various fluids such as saliva, urine, cerebrospinal fluid, semen, amniotic fluid and milk [1].
In humans, melatonin release begins immediately after darkness falls (20:00-23:00), reaches peak levels in the middle of the night (02:00-04:00), and ends in the morning hours (07:00-09:00). Nocturnal melatonin production in young adults is typically around 10–80 g [55]. In a study conducted on 170 people (85 women; 26.2 ± 6.0 years), it was reported that the highest melatonin concentration ranged between 2 and 84 pg/mL [56]. It is stated that the concentrations of melatonin in the blood and cells at night are 3-10 times higher than during the day [11]. Melatonin level in serum varies according to age. In newborns, melatonin levels are quite low. After the third month, melatonin secretion becomes rhythmic. While the highest melatonin level in the life cycle is detected between the third and fifth years of life, melatonin secretion and level decrease with advancing age [57,58,59].
Although melatonin is found in many food sources, melatonin content varies greatly among foods. The richest sources of melatonin taken externally (exogenously) through the diet are products such as eggs, fish, mushrooms, barley, oats, sprouted seeds, nuts, and medicinal plants [60]. Studies are showing that the seeds, roots, leaves, and fruits of plants are good sources of melatonin. The change in melatonin level varies depending on the type of plant, climatic conditions, horticultural practices, processing and storage conditions [28]. Melatonin levels in plants not only vary from species to species. It may also differ between genotypes and cultivars within the same species or at different growth stages of individuals of the same genotype [61,62].
2.4. Melatonin Synthesis Sites
Melatonin is secreted from the pineal gland, ovaries, lens and bone marrow cells, and the bile and gastrointestinal systems of mammals [34]. The pineal gland synthesizes approximately 80% of the amount of melatonin in the bloodstream. In many species, the main source of melatonin other than the pineal gland is the gastrointestinal tract. The intestines contain 400 times more melatonin than the pineal gland. Although the release of melatonin, synthesized in enterochromaffin cells in the gastrointestinal tract, shows a diurnal rhythm in birds, it is dependent on the presence of nutritional factors in many other species [63]. Melatonin, synthesized in the retina, plays a role in regulating retinal pigment epithelial function and the response of the retina to day-night variation in photoreceptors. Melatonin, which is responsible for the change of pigment granules in the skin and the protection of deep tissues against harmful radiation of the sun, is synthesized in enterochromaffin cells in the gastrointestinal tract and released into the circulation postprandial. Melatonin, synthesized in bile, protects the mucosa and epithelium of the biliary tract against oxidative damage against oxidized cholesterol derivatives and bile acid. However, the contribution of this synthesized amount to the melatonin level in the bloodstream is very small [1,23,38].
2.5. Effects of Melatonin on Physiological Functions
2.5.1. Effect on Lymphoid Tissues
Melatonin increases the size of tissues and organs directly related to immune function, such as the thymus and spleen of mammals. It has been reported that the thymus gland atrophy in animals whose pineal gland was removed experimentally or in which drugs were used to suppress the pineal function, and when melatonin was given externally to these animals, hyperplasia was observed in the thymus gland [64].
2.5.2. Presence of Melatonin in the Digestive System
In general, the level of melatonin secreted from the gastrointestinal system is 10-100 times higher than the level of melatonin secreted from the pineal gland. In particular, there are different levels of melatonin in different parts of the digestive system, and the amount of melatonin in this region is completely related to the serotonin concentration [8].
2.5.3. Effect of Melatonin on the Cardiovascular System
The function of melatonin on the cardiovascular system occurs through two different mechanisms. These are mechanisms with and without receptors. Melatonin causes vasoconstriction in cerebral arteries and vasodilation in peripheral vascular beds. Studies have reported that melatonin levels are very low in coronary heart patients at risk of myocardial infarction and sudden death [52,65]. Additionally, melatonin levels are low in people with high LDL-cholesterol levels, as well as in the body of hypertensive patients, and melatonin has been found to lower blood pressure. The role of melatonin's vasodilator effect in inducing sleep through thermoregulation is also very important [66].
2.5.4. Daily and Annual Rhythm Regulation and Photoperiodic Response
Melatonin has been found to act as a photoperiodic regulator or circadian rhythm regulator in animals and humans, as the number of changes in blood helps tissues and cells sense the time of day or year and mimics the dark application of external melatonin applications [5]. Circadian rhythm or circadian clock, also known as the biological clock, regulates the physiological, behavioral, and metabolic functions of a wide variety of organisms, from bacteria and fungi to plants and animals, in approximately 24-hour cycles, depending on the time it takes for the Earth to rotate once around itself. Cells and organisms react to the time of day according to their internal clock, thus cell renewal and the secretion of hormones occur, and control is provided over events such as photosynthesis and flowering in plants. According to some researchers, melatonin; can regulate many biological processes in plants, including circadian rhythm, regulation of genes and metabolites, protein stability, daily and seasonal cycles, increases photosynthesis and growth rates, can increase seed yield in the product by affecting flowering, and can affect biotic and abiotic responses [67,68]. It has been reported that it functions as a photoperiodic regulator or 24-hour rhythm regulator in plants. In addition, research has shown that although melatonin varies depending on the species, its amount increases especially at night or in the dark, and some plants it peaks a second time during the day. In the first study, in which a change similar to that in animals was observed in plants and the change in melatonin within 24 hours was revealed, it was found that the amount of melatonin in Chenopodium rubrum L. plants was at a very low level in the light but reached the highest level towards the end of the dark period [69]. Although it has been reported that melatonin acts as a 24-hour rhythm regulator in plants and that the amount of synthesis generally increases in the dark, some researchers have reported that the amount of synthesis reaches the highest level just before sunset [70,71]. In some research, it is reported that melatonin is constantly present in plants, with level changes during the day [3]. For example, in a study conducted with water hyacinth, it was observed that the melatonin level in plants grown under natural conditions increased towards the end of the light period, unlike the rhythm detected in animals, and the reason for this was related to the processes of photosynthesis and protection from light. It has been stated that it may be possible [71]. In another study examining the melatonin change in apples during the day, it was found that the melatonin level in leaves peaked twice during the day, at 14:30 and 05:30 [72].
2.5.5. Effect of Melatonin on Aging
In animals, melatonin has been found to prevent aging-related disorders and extend the lifespan of some mammals to some extent, depending on genetic conditions [73]. In studies on the aging process in plants, researchers focus on the aging effect occurring in the leaves rather than the life of the organism, so a different process is experienced than the aging seen in animals. However, the common point seen in both is that as aging progresses, the damage caused by oxidative damage in the relevant cell increases. In both cases, it is a known fact that the antioxidant activity of melatonin contributes to delaying aging [74].
2.5.6. Effects of Melatonin on the Immune System and Cancer
Melatonin is a powerful free radical scavenger and has an antioxidant effect by blocking nitric oxide synthase enzyme activity [71,75]. It also prevents oxidative damage caused by neutrophil activation in tissues in inflammation cases such as burn sepsis, ischemia re-perfusion injury [75]. Another function of melatonin on the immune system is its anti-inflammatory effect. It has also been suggested that melatonin may have an effect on diseases that may involve viruses and be contagious, such as bacterial infections, cancer, and HIV[76].
Melatonin also provides daily and seasonal regulation of immune system cells [77]. In studies conducted at the in vivo level, it has been determined that high doses of melatonin provide general immune system stimulation. It has been determined that it increases T cell activation, lymphocyte growth, humoral response and inhibits thymus involution that occurs with age. (Simonneaux and Ribelayga 2003). In vitro studies have shown that melatonin increases helper T cell and NK cell activity, IL-2 and IFNγ production, and IL 1 mRNA expression in human monocytes [77]. In addition, considering the helplessness experienced during the treatment of viral infections, it has been reported that melatonin can act as a supporter in the treatment of many viral infections [78].
It has been demonstrated that MT1 melatonin receptors are constantly active in breast and prostate cancer cells and, as a result, inhibit the proliferation of cancer cells [79]. In recent years, the majority of research on the effects of melatonin on cancer has focused on breast cancer. The results of the studies have revealed that nighttime applications of melatonin provide more successful results in cancer [3,80]. The effect of seasons on cancer development is proportional to melatonin. During the winter months, when nights are long, melatonin production is high and tumor development slows down during this period [81].
Melatonin has a pure anticarcinogenic effect by reducing the receptors that allow linoleic acid, one of the substances that the cancer cell needs for growth, to enter the cancer cell, puts the cancer cell to sleep through the Ca-calmodulin complex, and slows down the growth of the tumor [82]. It has been determined that melatonin binds to Ca2+ activated calmodulin with high affinity. Thus, it has been suggested that melatonin prevents tumor growth by removing calmodulin from calcium, slowing down the cell cycle. Melatonin also has the feature of preventing the formation of apoptosis in healthy cells [3,76].
2.5.7. Free Radical Scavenging and Antioxidant Effect of Melatonin
Oxygen, which makes up 21% of the Earth's atmosphere, is necessary for aerobic organisms to survive. The majority of inhaled oxygen is used in the production of ATP, and a relatively significant portion of inhaled oxygen (about 5%) is converted into extremely toxic free radicals. Melatonin is a powerful hydroxyl and peroxyl radical scavenger and its radical scavenging effect is lower than vitamin E and therefore has less neutralizing effect against the lipoperoxyl radical [75,83].
This is due to the production of free radicals during oxidative phosphorylation in mitochondria and fatty acid oxidation in peroxisomes. These free radicals are toxic substances called hydroxyl radicals (OH), hydrogen peroxide radicals (H2O2), superoxide anion radicals or singlet oxygen radicals (O2) [84,85]. Melatonin is a powerful antioxidant that eliminates the hydroxyl radical, the most harmful radical, thus preventing lipid peroxidation [85]. Melatonin has also been shown to have indirect effects on free radicals. Melatonin also has an antioxidant effect by activating the GSH-Px enzyme that metabolizes hydroperoxides, increasing the SOD activity that catalyzes the O2- radical to H2O2, preventing the decrease in catalase activity during oxidative stress, and inhibiting the nitric oxide synthase (NOS) enzyme responsible for NO formation [1,86,87].
Oxidative stress caused by some toxins that cause oxidative tissue damage is prevented by melatonin. Since melatonin is soluble in both water and lipid phases, it easily reaches all intracellular components and effectively protects the cell membrane, organelles and nucleus from free radical damage. Melatonin binds to the out of the cell membrane surface, traps radicals before they enter the membrane, and protects the membrane by detoxifying them. It also reduces the production of radicals such as oxygen (O2), hydrogen peroxide (H2O2) and hydroxy (OH) that occur in mitochondrial respiration. Its ability to reach the nucleus protects DNA against oxidative damage [88].
2.5.8. Effect of Melatonin on Sleep
Melatonin ensures homeostasis as an endogenous regulator. It provides information to all cells with daily, monthly and annual secretion differences with the signals coming from the SCN. This hormone epigenetically controls clock genes in cells [18,82]. It is known that its effects on sleep are probably chronobiological effects. It is known that the effect of melatonin synthesis on sleep is related to the onset, quality and latent phase of sleep rather than the length of total sleep time and that this is achieved through the hypothermic effect and thermoregulation. Melatonin increases the nocturnal temperature drop and makes it easier to fall asleep [18]. While sleep is not necessary for melatonin production, a dark environment is sufficient [89].
Sleep is a very important time period for human health. The basic mechanism that ensures sleep is; It was the light-dark cycle. The dark environment stimulates the nerve network that starts from the retina and extends to the hypothalamic region of the brain. The suprachiasmatic nucleus in the hypothalamic region of the brain acts as a "biological clock" by regulating activities that affect the whole body by giving signals that control body temperature, hormones, sleep-wake status and other regions [90].
The pineal gland remains inactive until darkness falls, and the production of sleep-related melatonin and other similar hormones is restricted. When the environment darkens, the signals that suppress melatonin production created by the suprachiasmatic nucleus disappear and the pineal gland is stimulated to release melatonin. As the melatonin level increases, less stimulation is perceived in the brain and the likelihood of falling asleep increases. It has been found that melatonin level is related to the onset, latent phase and sleep quality rather than the length of the total sleep cycle. [91].
2.5.9. Melatonin's Relationship with the Reproductive System and Other Hormones
It has been determined in numerous studies that melatonin release affects the reproductive system [92,93]. Exogenous melatonin also regulates reproduction, although it varies depending on species, age, dose and time of administration [85]. It has been suggested that melatonin may have an inhibitory effect on the hypothalamic pituitary gonadal system and also it has antigonadotropic effects [94]. Reportedly as a result of the chronic melatonin administration in adult females, ovarian size decreases and the frequency of estrus decreases [95]. While high doses of melatonin decrease gonadotropin levels, it increases prolactin and growth hormone levels. Melatonin controls reproductive activity in seasonally breeding animals [96].
While Melataonin has a stimulating effect on the gonads in animals that reproduce in short days, such as goats, sheep and deer, it has an inhibitory effect in animals that reproduce in long days, such as hamsters, camels and horses [97]. Especially in seasonally breeding hamsters, prolonged darkness prevents reproduction by causing more melatonin secretion and causes testicles to shrink in male hamsters and anoestrus in females. Epidemiological studies show that although there is no seasonal reproduction in humans; It shows that pregnancy and birth rates have a seasonal distribution in different geographical regions. It has been revealed that in regions where winters are long, hypothalamo-gonadal system activation and pregnancy rate are lower compared to summer months [98].
It is a center that regulates endocrinological activity in the organism with the help of the pineal gland, thyroid, adrenal gland, gonads and pituitary [54]. It is accepted that melatonin and therefore the pineal gland have effects on the release of gonadotropic hormones as well as other hormones from the pituitary. Administration of melatonin increases prolactin secretion in women. Melatonin application in adult men increases basal growth hormone secretion and leads to an increase in the effect of growth hormone release hormone on growth hormone secretion. Melatonin probably exerts this effect by inhibiting somatostatin secretion from the hypothalamus [99,100] showed in their study that 1 mg melatonin given to people at 09.00 in the morning reduced PRL and LH levels, but did not affect GH, FSH and TSH levels.
2.5.10. Effect of Melatonin on Aging
In recent studies, it has been determined that the plasma melatonin level in humans decreases with aging. In parallel with this, the circadian rhythm of melatonin changes. In studies determined that the plasma melatonin level detected in elderly people was 40-50% lower than its normal value [54,84].
It has been determined that anatomical and functional degeneration in organs with aging is associated with the decrease in antioxidant capacity that occurs with the decrease in melatonin level and the damage caused by increased free radicals [101]. As it is known, melatonin has the effect of protecting brain tissue from oxidative damage, preventing lipid peroxidation and stimulating antioxidative enzymes. Research has revealed that neurodegenerative damage in the brain occurs as a result of an increase in free radicals due to a decrease in melatonin [102]. The decrease in melatonin, an important radical scavenger, over time causes brain tissue to become vulnerable to oxidative attacks [102]. With age, the number of β-adrenergic receptors in the pinealocyte membrane decreases and probably melatonin synthesis in the pineal gland decreases. β-adrenergic receptors mediate the secretion of norepinephrine from sympathetic neurons to the pineal gland at night and initiate a series of reactions that increase melatonin synthesis [84,101].
2.5.11. Effect of Melatonin on Jetlag
Studies have relaved that transmeridian journeys have an effect on sleep, circadian rhythm and daily activity [103]. Flight dysrhythmia, as it is known, manifests itself with symptoms such as inability to loss of appetite, wake up, daytime fatigue, jetlag, nighttime insomnia, poor psychomotor coordination, digestive problems, poor cognition, and depressive mood, [103]. To relieve jetlag symptoms, especially on flights from west to east; Studies have shown that the most effective treatment is melatonin application [103].
2.5.12. Effect of Melatonin on İnsulin Secretion
There is evidence that the circadian rhythm of melatonin affects the pancreas and ensures insulin secretion [104,105]. Many studies suggest that melatonin has a suppressive effect on the activity of the pineal gland, β-cell, as it reduces insulin levels in rats [106] and revealed that these effects are compatible with the decrease in glucose tolerance. Increased insulin rate has an inhibitory effect on the pineal gland and melatonin. This situation reveals the fact that there is a functional antagonism between insulin and melatonin [107,108].
3. Presence of Melatonin ın Plants
Studies investigating melatonin in plants have a shorter history than those in animals. Recently, significant progress has been made in research based on the effects of melatonin in plants. It has been determined that melatonin controls many physiological processes, especially during vegetative and reproductive periods, due to its ability to reduce oxidative stress in plants [109,110,111]. Melatonin regulates circadian rhythm and photoperiodic properties in plants.
It also has effects such as regulating growth, gene expression and development, protecting plants by preparing against endo- or exogenous biological stress factors, ensuring interaction with biotic substances (e.g. herbivores/pests) and slowing down aging. Additionally, as an indoleamine neurotransmitter, it functions primarily as an antioxidant agent by controlling reactive oxygen and nitrogen in plants. Melatonin, a signaling agent, increases growth, rooting, photosynthesis, carbon fixation, seed germination and defense against various biotic and abiotic stress factors in plants and induces a variety of specific physiological responses. It also functions as an important mediator in the metabolism of cytokinin, gibberellin, ethylene, indole-3-acetic acid, and oxygen carrier proteins, as well as gene expression related to plant hormones. Again, regulation of stress-specific genes and activation of pathogenesis-related protein and antioxidant enzyme genes under stress conditions have made it a versatile molecule. Melatonin is one of the main function regulators of plants due to its diversity of effects and its role in regulating plant growth, development, and behavior [109,110,111,112].
3.1. Melatonin Formation in Plants
Melatonin production in plants occurs differently than in animals. Melatonin biosynthesis in plants occurs in main regions such as chloroplast and mitochondria. These organelles contain enzyme groups that synthesize melatonin through different procedures. Melatonin production can occur in two ways. If melatonin production is prevented in mitochondria, synthesis takes place in chloroplasts. Production in plants occurs by a specific M3H enzyme [113]. Although the amount of melatonin in plants varies depending on plant tissues and light level, melatonin has been detected in almost all plant species. It has been determined that the melatonin level in leaves and aromatic plants is higher than in seeds [16,20]. In higher plants, melatonin was first identified in 1993 in the fruits of Convolvulaceae, namely ivy morning glory and tomato plants [114,115]. Especially plants belonging to the Rosaceae, Apiaceae, Brassicaceae, Poaceae and Vitaceae families contain more endogenous melatonin than other plant species. Over the last decade, many researchers have reported that melatonin concentration varies among varieties of the same species, depending on factors such as location, growth stage, part of the plant, and harvest time [116,117].
3.2. Edible Vegetable Sources of Melatonin
Melatonin is found especially in edible plants and various plant-based food products. Plants contain not only melatonin but also its precursor. Melatonin, a bioactive substance, is found in different compositions and concentrations in fruits, vegetables, and herbs such as tomatoes, strawberries, grapes, rice, walnuts, oranges, cherries, olive oil, and barley. Melatonin levels in plants are universal; However, concentrations in plant tissues can vary from picograms to micrograms. In the growth and germination of seeds in plants, in reducing oxidative stress, in increasing resistance to stress factors (drought, salinity, toxic chemicals, heavy metals, UV rays, high and low and high ambient temperature, and light), in stimulating the immune system, in regulating circadian rhythms, in leaves. It plays an important role in controlling stomatal closure. In addition, since melatonin contributes to the development of various properties such as antibacterial, antifungal and antiviral, it also contributes to coping with biotic stress in plants [117,118].
3.3. Melatonin Uptake in Plants
Plants containing melatonin can synthesize this indolamine; It is also possible for them to receive melatonin applied externally to the growing medium, that is, the soil or the plant itself (leaf) [75]. Studies have shown that melatonin is not only taken from the roots but also the seeds and leaves can absorb it. For example, it was determined that the amount of melatonin in barley leaves given different amounts of melatonin increased depending on the concentration [119].
3.4. Function of Melatonin in the Regulation of Plant Development and Physiology
While melatonin is known to improve the overall growth of plants, its primary function is as an antioxidant activity [118,120]. Melatonin plays a role in the control of growth and development, coping with stress, antioxidant defense mechanism and various physiological processes in plants. In plants grown in the garden, it can control seed germination, root development, shoot release and biomass increase. It also plays a role in ensuring vegetative reproduction and growth stages, flower formation and regulating leaf senescence [60,121]. A similar role of melatonin was also determined in etiolated Lupinus albus L.; here it was found to be responsible for promoting vegetative growth and regeneration of lateral and adventitious roots [122]. Again, in a study conducted on cucumbers, melatonin was applied to cucumber plants under conditions of intense salt stress, and as a result, it was found that there was an increase in ide growth, improvement in nutrient uptake and productivity, and an increase in nitrogen metabolism [123]. In another study, it was found that melatonin supplementation stimulated root growth and elongation, increased photosynthetic activity, and improved redox homeostasis in wheat, barley and rice [124]. In summary, melatonin can be regarded as an important biological plant growth regulator that increases the reproductive rate in plants.
3.4.1. Function of Melatonin on Seed Germination
Germination and seedling formation are two important steps in the formation of plant populations. Melatonin has an important contribution to this. In the natural cycle, melatonin level increases rapidly in the initial phase of seed germination and reaches its highest level at the end of the 14-hour period. However, as the germination period progresses, its concentration begins to decrease and reaches a steady state in the later stages of seedling formation. In particular, the increase in melatonin in the cytoplasm at the beginning of the plant development process may contribute to faster delivery of nutrients to the metabolic regions of germinating seeds [125].
3.4.2. Function of Melatonin on Photosynthesis
Photosynthesis in plants is sensitive to biotic and abiotic factors. Under plant stress conditions, it delays fermentation by closing stomata, reducing chlorophyll content and destroying excitons in chloroplasts. However, melatonin supplementation to the plant is useful in alleviating these negative effects of stressors. In particular, it ensures the photosynthetic efficiency of the plant at the cellular level by increasing stomatal conductance, reducing the rate of chlorophyll degradation and helping stomata to both open and close. Thus, the plant shows a higher rate of CO2 assimilation and photosynthetic efficiency, as well as an increase in antioxidative effect. Melatonin reduces the negative effects of abiotic stress factors as well as biotic stress factors. Thus, the chloroplast is protected, sugar assimilation is supported, and proline accumulation, which is necessary to maintain normal cellular functions under stress, is increased. [109,126]. Additionally, genes related to carbon assimilation, photosynthesis, and ascorbate biosynthesis are also stimulated by melatonin. In a study investigating the application of melatonin to cherry rootstocks under stress, it was determined that the production of photosynthetic pigments increased with the application of melatonin to the rootstock, resulting in higher carbohydrate accumulation, an increase in proline content and an increase in biomass.[127,128]
3.4.3. The Function of Melatonin in the Regulation of Reproduction, Development and Circadian Rhythm of Plants
After the existence and importance of melatonin was discovered in plants, it was thought to have a similar function to that in mammals. To this end, initial studies focused on factors related to circadian rhythm, such as vernalization, photoreceptors, flowering, and hormones, and on identifying the potential role of melatonin [20,123,129].
3.4.4. Function of Melatonin on the Antioxidant Defense System
The main function of melatonin in plants is to act as an antioxidant and to protect the plant organism against various environmental pollutants. In the study conducted on barley (Hordeum vulgare), barley was exposed to multiple stressors such as salt, hydrogen peroxide and some heavy metals, and as a result, it was determined that there was an increase in the endogenous melatonin level of the plant. Similar results were also detected in Lupinus albus (lupine) seedlings [130,131,132].
3.4.5. Effect of Melatonin on Supporting Defense Mechanisms against Various Stress Elements in Plants
As explained in the previous sections, melatonin has a protective role and defense mechanism for the plant against various stress factors such as drought, heat, cold, chemicals, heavy metals, ultraviolet rays and salinity [127].
3.4.6. Function of Melatonin in Biomass Increase in Plants
Apart from the main factors mentioned, melatonin also has an effect on increasing the biomass of plants. Especially in order to develop transgenic plants, different modifications have been made on the metabolic indoleamine enzyme of melatonin and it has been determined that this increases plant biomass [118]. In addition, melatonin increases plant development and productivity, improves the germination of seeds and seedlings by developing defense mechanisms against external and internal factors, and increases plant biomass [133,134].
4. Conclusions
Based on the available data and research in this review study, it is seen that melatonin has an undeniable importance in human, animal, plant and agricultural production. Melatonin is widely found in nature and is a biomolecule with very important functions produced by humans, animals, plants, fungi and single-celled organisms. In case of deficiencies in melatonin synthesis or impaired receptor in the organism, various abnormal conditions such as hypertension, diabetes, neurological and autoimmune disorders, mood changes, breast cancer, obesity and prostate cancer may occur. In particular, it has been revealed that the deficiency of the Melatonin molecule plays an important role in disorders such as sleep disorders and chronic insomnia. It is also vital in reducing oxidative damage. Therefore, exogenous melatonin is applied in the prevention and treatment of various diseases, as well as in many clinical applications. For this purpose, various melatonin agonists have been developed and produced to perform various treatments. Melatonin is also a hormonal molecule with very important functions in plants. Melatonin, an antioxidant, increases the resistance of plants to chemical pollutants, environmental factors, low and high temperatures, and other environmental factors. In addition, melatonin accelerates photosynthesis in plant metabolism by protecting chlorophyll against oxidative stress elements, and ensures root renewal by signaling the development of the root system. Due to all these important effects, it is of great importance to reveal and better understand the functions of the melatonin molecule in humans, animals, and plants.
Author Contributions
Conceptualization, A.R.N., A.K.Y., N.Ç. and H.Ö.D.; resources A.R.N., A.K.Y., N.Ç. and H.Ö.D.; data curation, A.R.N., A.K.Y., N.Ç. and H.Ö.D.; writing—original draft preparation, A.R.N., A.K.Y. and N.Ç.; writing—review and editing, A.R.N., A.K.Y., N.Ç. and H.Ö.D.; funding acquisition, A.R.N. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research was funded by the Department of Science and Innovation, Pretoria (South African Government), grant number DSI-IKS DSI/CON C2235/2021.
Data Availability Statement
Not applicable.
Acknowledgments
The authors (A.R.N., A.K.Y., N.Ç. and H.Ö.D) acknowledge the funding provided by the Department of Science and Innovation, Pretoria, South African Government.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Reiter, R.J. Melatonin: The Chemical Expression of Darkness. Molecular and Cellular Endocrinology 1991, 79, C153–C158. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.B.; Siopes, T.D. Melatonin Enhances Cellular and Humoral Immune Responses in the Japanese Quail (Coturnix Coturnix Japonica) via an Opiatergic Mechanism. General and Comparative Endocrinology 2003, 131, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Paredes, S.D.; Manchester, L.C.; Tan, D.-X. Reducing Oxidative/Nitrosative Stress: A Newly-Discovered Genre for Melatonin. Critical Reviews in Biochemistry and Molecular Biology 2009, 44, 175–200. [Google Scholar] [CrossRef] [PubMed]
- Çam, A.; Erdoğan, M.F. Melatonin. Ankara Üniversitesi Tıp Fakültesi Mecmuası 2003, 56. [Google Scholar] [CrossRef]
- Reiter, R.J. Pineal Melatonin: Cell Biology of Its Synthesis and of Its Physiological Interactions*. Endocrine Reviews 1991, 12, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Vico, A.; Calvo, J.R.; Abreu, P.; Lardone, P.J.; García-Mauriño, S.; Reiter, R.J.; Guerrero, J.M. Evidence of Melatonin Synthesis by Human Lymphocytes and Its Physiological Significance: Possible Role as Intracrine, Autocrine, and/or Paracrine Substance. The FASEB Journal 2004, 18, 537–539. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. ISOLATION OF MELATONIN, THE PINEAL GLAND FACTOR THAT LIGHTENS MELANOCYTES1. J. Am. Chem. Soc. 1958, 80, 2587–2587. [Google Scholar] [CrossRef]
- Bubenik, G.A. Localization, Physiological Significance and Possible Clinical Implication of Gastrointestinal Melatonin. Biological Signals and Receptors 2001, 10, 350–366. [Google Scholar] [CrossRef]
- Melatonin Effects on Bone: Experimental Facts and Clinical Perspectives - Cardinali - 2003 - Journal of Pineal Research - Wiley Online Library Available online:. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1034/j.1600-079X.2003.00028.x?sid=nlm%3Apubmed (accessed on 4 March 2024).
- Beyer, C.E.; Steketee, J.D.; Saphier, D. Antioxidant Properties of Melatonin—an Emerging Mystery. Biochemical Pharmacology 1998, 56, 1265–1272. [Google Scholar] [CrossRef]
- Atasoy, N. Melatonin ve Antioksidan Etkileri. Düzce Üniversitesi Sağlık Bilimleri Enstitüsü Dergisi 2019, 9, 196–201. [Google Scholar] [CrossRef]
- Juhnevica-Radenkova, K.; Moreno, D.A.; Ikase, L.; Drudze, I.; Radenkovs, V. Naturally Occurring Melatonin: Sources and Possible Ways of Its Biosynthesis. Comprehensive Reviews in Food Science and Food Safety 2020, 19, 4008–4030. [Google Scholar] [CrossRef] [PubMed]
- Poeggeler, B.; Hardeland, R. Detection and Quantification of Melatonin in a Dinoflagellate, Gonyaulax Polyedra: Solutions to the Problem of Methoxyindole Destruction in Non-Vertebrate Material. Journal of Pineal Research 1994, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- BAGHURST, R.; COGHILL, R. A STUDY OF MELATONIN IN PLANT TISSUES AND ITS DIETARY AND HEALTH IMPLICATIONS. In Proceedings of the BIOELECTROMAGNETICS Current Concepts; Ayrapetyan, S.N., Markov, M.S., Eds.; Springer Netherlands: Dordrecht, 2006; pp. 405–412. [Google Scholar]
- Hardeland, R. Melatonin and 5-methoxytryptamine in non-metazoans. Reprod. Nutr. Dev. 1999, 39, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B. Phytomelatonin: Discovery, Content, and Role in Plants. Advances in Botany 2014, 2014, 1–11. [Google Scholar] [CrossRef]
- Feng, X.; Wang, M.; Zhao, Y.; Han, P.; Dai, Y. Melatonin from Different Fruit Sources, Functional Roles, and Analytical Methods. Trends in Food Science & Technology 2014, 37, 21–31. [Google Scholar] [CrossRef]
- Claustrat, B.; Geoffriau, M.; Brun, J.; Chazot, G. Melatonin in Humans: A Biochemical Marker of the Circadian Clock and an Endogenous Synchronizer. Neurophysiologie Clinique/Clinical Neurophysiology 1995, 25, 351–359. [Google Scholar] [CrossRef]
- Brzezinski, A. Melatonin in Humans. New England Journal of Medicine 1997, 336, 186–195. [Google Scholar] [CrossRef]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front Endocrinol (Lausanne) 2019, 10, 249. [Google Scholar] [CrossRef]
- Garaulet, M.; Qian, J.; Florez, J.C.; Arendt, J.; Saxena, R.; Scheer, F.A.J.L. Melatonin Effects on Glucose Metabolism: Time To Unlock the Controversy. Trends in Endocrinology & Metabolism 2020, 31, 192–204. [Google Scholar] [CrossRef]
- Scheer, F.A.J.L.; Czeisler, C.A. Melatonin, Sleep, and Circadian Rhythms. Sleep Medicine Reviews 2005, 9, 5–9. [Google Scholar] [CrossRef]
- Özçelik, F.; Erdem, M.; Bolu, A.; Gülsün, M. Melatonin: Genel Özellikleri ve Psikiyatrik Bozukluklardaki Rolü. Psikiyatride Güncel Yaklaşımlar 2013, 5, 179–203. [Google Scholar] [CrossRef]
- Yılmaz, C.; Gökmen, V. Neuroactive Compounds in Foods: Occurrence, Mechanism and Potential Health Effects. Food Research International 2020, 128, 108744. [Google Scholar] [CrossRef] [PubMed]
- Kleszczyński, K.; Slominski, A.T.; Steinbrink, K.; Reiter, R.J. Clinical Trials for Use of Melatonin to Fight against COVID-19 Are Urgently Needed. Nutrients 2020, 12, 2561. [Google Scholar] [CrossRef] [PubMed]
- Favero, G.; Franceschetti, L.; Buffoli, B.; Moghadasian, M.H.; Reiter, R.J.; Rodella, L.F.; Rezzani, R. Melatonin: Protection against Age-Related Cardiac Pathology. Ageing Research Reviews 2017, 35, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Sharopov, F.; Fokou, P.V.T.; Kobylinska, A.; Jonge, L. de; Tadio, K.; Sharifi-Rad, J.; Posmyk, M.M.; Martorell, M.; Martins, N.; et al. Melatonin in Medicinal and Food Plants: Occurrence, Bioavailability, and Health Potential for Humans. Cells 2019, 8, 681. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.M.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Dehpour, A.R.; Shirooie, S.; Silva, A.S.; Baldi, A.; Khan, H.; Daglia, M. Anti-Inflammatory Effects of Melatonin: A Mechanistic Review. Critical Reviews in Food Science and Nutrition 2019, 59, S4–S16. [Google Scholar] [CrossRef]
- Karunanithi, D.; Radhakrishna, A.; Sivaraman, K.P.; Biju, V.M.N. Quantitative Determination of Melatonin in Milk by LC-MS/MS. J Food Sci Technol 2014, 51, 805–812. [Google Scholar] [CrossRef] [PubMed]
- ÖZTÜRK, G.; AKBULUT, K.; GÜNEY, Ş. Melatonin, Aging, and COVID-19: Could Melatonin Be Beneficial for COVID-19 Treatment in the Elderly? Turkish Journal of Medical Sciences 2020, 50, 1504–1512. [Google Scholar] [CrossRef]
- Yakupoğlu, G.; Köklü, Ş.; Korkmaz, A. Bitkilerde Melatonin ve Üstlendiği Görevler. Kahramanmaraş Sütçü İmam Üniversitesi Doğa Bilimleri Dergisi 2018. [Google Scholar] [CrossRef]
- Brainard, G.C.; Rollag, M.D.; Hanifin, J.P. Photic Regulation of Melatonin in Humans: Ocular and Neural Signal Transduction. J Biol Rhythms 1997, 12, 537–546. [Google Scholar] [CrossRef]
- Koçak, D.A.; Çolak, D.A. Melatonin ve Santral Sinir Sistemi. Turgut Özal Tıp Merk Derg 1996, 3. [Google Scholar]
- Erzincan Binali Yildirim Universitesi Tip Fakultesi, Fizyoloji Ana Bilim Dali, Erzincan, Turkiye; Ustundag, H.; Senturk, E.; Agri Ibrahim Cecen Universitesi Saglik Bilimleri Fakultesi, Hemsirelik Bolumu, Agri, Turkiye; Gul, M.; Ataturk Universitesi Tip Fakultesi, Fizyoloji Ana Bilim Dali, Ezurum, Turkiye Melatonin and Hyperthyroidism. Arch Basic Clin Res 2020, 2, 59–64. [CrossRef]
- Bernard, M.; Guerlotté, J.; Grève, P.; Gréchez-Cassiau, A.; Iuvone, M.P.; Zatz, M.; Chong, N.W.; Klein, D.C.; Voisin, P. Melatonin synthesis pathway: circadian regulation of the genes encoding the key enzymes in the chicken pineal gland and retina. Reprod. Nutr. Dev. 1999, 39, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Bolliet, V.; Ali, M.A.; Lapointe, F.-J.; Falcón, J. Rhythmic Melatonin Secretion in Different Teleost Species: An in Vitro Study. J Comp Physiol B 1996, 165, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Ekmekcioglu, C. Melatonin Receptors in Humans: Biological Role and Clinical Relevance. Biomedicine & Pharmacotherapy 2006, 60, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Şahin, D. In vitro koşullarda sirkadien melatonin etkisine maruz bırakılan embriyolarda SOD ve HMGB1 genlerinin ekspresyonları ile melatonin etkisinin takibi. masterThesis, İstanbul Bilim Üniversitesi, Sağlık Bilimleri Enstitüsü., 2014.
- Siu, A.W.; Reiter, R.J.; To, C.H. The Efficacy of Vitamin E and Melatonin as Antioxidants against Lipid Peroxidation in Rat Retinal Homogenates. Journal of Pineal Research 1998, 24, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J. The Pineal Gland and Melatonin in Relation to Aging: A Summary of the Theories and of the Data. Experimental Gerontology 1995, 30, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.P.N.; Shiu, S.Y.W.; Chow, P.H.; Pang, S.F. Regional and Diurnal Studies of Melatonin and Melatonin Binding Sites in the Duck Gastro-Lntestinal Tract. Neurosignals 1996, 4, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Jockers, R.; Delagrange, P.; Dubocovich, M.L.; Markus, R.P.; Renault, N.; Tosini, G.; Cecon, E.; Zlotos, D.P. Update on Melatonin Receptors: IUPHAR Review 20. British Journal of Pharmacology 2016, 173, 2702–2725. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Godson, C.; Mahle, C.D.; Weaver, D.R.; Slaugenhaupt, S.A.; Gusella, J.F. Molecular Characterization of a Second Melatonin Receptor Expressed in Human Retina and Brain: The Mel1b Melatonin Receptor. Proceedings of the National Academy of Sciences 1995, 92, 8734–8738. [Google Scholar] [CrossRef]
- Nosjean, O.; Ferro, M.; Cogé, F.; Beauverger, P.; Henlin, J.-M.; Lefoulon, F.; Fauchère, J.-L.; Delagrange, P.; Canet, E.; Boutin, J.A. Identification of the Melatonin-Binding SiteMT 3 as the Quinone Reductase 2 *. Journal of Biological Chemistry 2000, 275, 31311–31317. [Google Scholar] [CrossRef]
- Uz, T.; Arslan, A.D.; Kurtuncu, M.; Imbesi, M.; Akhisaroglu, M.; Dwivedi, Y.; Pandey, G.N.; Manev, H. The Regional and Cellular Expression Profile of the Melatonin Receptor MT1 in the Central Dopaminergic System. Molecular Brain Research 2005, 136, 45–53. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin in Aging and Disease —Multiple Consequences of Reduced Secretion, Options and Limits of Treatment. Aging Dis 2011, 3, 194–225. [Google Scholar]
- Dubocovich, M.L. Melatonin Receptors: Are There Multiple Subtypes? Trends in Pharmacological Sciences 1995, 16, 50–56. [Google Scholar] [CrossRef]
- Morgan, P.J.; Barrett, P.; Howell, H.E.; Helliwell, R. Melatonin Receptors: Localization, Molecular Pharmacology and Physiological Significance. Neurochemistry International 1994, 24, 101–146. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Y.; Smith, D.G.; Hardeland, R.; Yang, M.Y.; Xu, H.L.; Zhang, L.; Yin, H.D.; Zhu, Q. Melatonin Receptor Genes in Vertebrates. International Journal of Molecular Sciences 2013, 14, 11208–11223. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Golombek, D.A.; Rosenstein, R.E.; Cutrera, R.A.; Esquifino, A.I. Melatonin Site and Mechanism of Action: Single or Multiple? Journal of Pineal Research 1997, 23, 32–39. [Google Scholar] [CrossRef]
- Slaugenhaupt, S.A.; Roca, A.L.; Liebert, C.B.; Altherr, M.R.; Gusella, J.F.; Reppert, S.M. Mapping of the Gene for the Mel1a-Melatonin Receptor to Human Chromosome 4 (MTNR1A) and Mouse Chromosome 8 (Mtnr1a). Genomics 1995, 27, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Dubocovich, M.L.; Markowska, M. Functional MT1 and MT2 Melatonin Receptors in Mammals. Endocr 2005, 27, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Witt-Enderby, P.A.; Bennett, J.; Jarzynka, M.J.; Firestine, S.; Melan, M.A. Melatonin Receptors and Their Regulation: Biochemical and Structural Mechanisms. Life Sciences 2003, 72, 2183–2198. [Google Scholar] [CrossRef]
- Atasoy, Ö.B.; Erbaş, O. Melatonin hormonunun fizyolojik etkileri. İstanbul Bilim Üniversitesi Florence Nightingale Tıp Dergisi 2017, 3, 52–62. [Google Scholar] [CrossRef]
- Peuhkuri, K.; Sihvola, N.; Korpela, R. Diet Promotes Sleep Duration and Quality. Nutrition Research 2012, 32, 309–319. [Google Scholar] [CrossRef]
- Burgess, H.J.; Fogg, L.F. Individual Differences in the Amount and Timing of Salivary Melatonin Secretion. PLOS ONE 2008, 3, e3055. [Google Scholar] [CrossRef] [PubMed]
- WALDHAUSER, F.; WEISZENBACHER, G.; TATZER, E.; GISINGER, B.; WALDHAUSER, M.; SCHEMPER, M.; FRISCH, H. Alterations in Nocturnal Serum Melatonin Levels In Humans With Growth and Aging*. The Journal of Clinical Endocrinology & Metabolism 1988, 66, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Molina-Carballo, A.; Munoz-Hoyos, A.; Martin-Garcia, J. a.; Uberos-Feritindez, J.; Rodriguez-Cabezas, T.; Acuna-Castroviejo, D. 5-Methoxytryptophol and Melatonin in Children: Differences Due to Age and Sex. Journal of Pineal Research 1996, 21, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.P.; Vacas, M.I. Pineal Gland, Photoperiodic Responses, and Puberty. J Endocrinol Invest 1984, 7, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Kamran, M.; Ding, R.; Meng, X.; Wang, H.; Ahmad, I.; Fahad, S.; Han, Q. Exogenous Melatonin Confers Drought Stress by Promoting Plant Growth, Photosynthetic Capacity and Antioxidant Defense System of Maize Seedlings. PeerJ 2019, 7, e7793. [Google Scholar] [CrossRef]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of Melatonin in Plants and Its Effects on Plasma Melatonin Levels and Binding to Melatonin Receptors in Vertebrates. Biochem Mol Biol Int 1995, 35, 627–634. [Google Scholar] [PubMed]
- Posmyk, M.M.; Janas, K.M. Melatonin in Plants. Acta Physiologiae Plantarum 2009, 31. [Google Scholar] [CrossRef]
- Martin, M.T.; Azpiroz, F.; Malagelada, J.R. Melatonin and the Gastrointestinal Tract. Therapie 1998, 53, 453–458. [Google Scholar]
- Maestroni, G.J.M.; Conti, A.; Pierpaoli, W. The Pineal Gland and the Circadian, Opiatergic, Immunoregulatory Role of Melatonin. Annals of the New York Academy of Sciences 1987, 496, 67–77. [Google Scholar] [CrossRef]
- Sewerynek, E. Melatonin and the Cardiovascular System. Neuro Endocrinol Lett 2002, 23 Suppl 1, 79–83. [Google Scholar]
- Paulis, L.; Simko, F. Blood Pressure Modulation and Cardiovascular Protection by Melatonin: Potential Mechanisms Behind. Physiol Res 2007, 56, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Atkins, K.A.; Dodd, A.N. Circadian Regulation of Chloroplasts. Current Opinion in Plant Biology 2014, 21, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Buttar, Z.A.; Wu, S.N.; Arnao, M.B.; Wang, C.; Ullah, I.; Wang, C. Melatonin Suppressed the Heat Stress-Induced Damage in Wheat Seedlings by Modulating the Antioxidant Machinery. Plants 2020, 9, 809. [Google Scholar] [CrossRef] [PubMed]
- Kolář, J.; Macháčková, I.; Eder, J.; Prinsen, E.; van Dongen, W.; van Onckelen, H.; Illnerová, H. Melatonin: Occurrence and Daily Rhythm in Chenopodium Rubrum. Phytochemistry 1997, 44, 1407–1413. [Google Scholar] [CrossRef]
- Wolf, K.; Kolář, J. a. n.; Witters, E.; van Dongen, W.; van Onckelen, H.; Macháčková, I. Daily Profile of Melatonin Levels in Chenopodium Rubrum L. Depends on Photoperiod. Journal of Plant Physiology 2001, 158, 1491–1493. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.-X.; Manchester, L.C.; Simopoulos, A.P.; Maldonado, M.D.; Flores, L.J.; Terron, M.P. Melatonin in Edible Plants (Phytomelatonin): Identification, Concentrations, Bioavailability and Proposed Functions. World Rev Nutr Diet 2007, 97, 211–230. [Google Scholar] [CrossRef]
- Zuo, B.; Zheng, X.; He, P.; Wang, L.; Lei, Q.; Feng, C.; Zhou, J.; Li, Q.; Han, Z.; Kong, J. Overexpression of MzASMT Improves Melatonin Production and Enhances Drought Tolerance in Transgenic Arabidopsis Thaliana Plants. Journal of Pineal Research 2014, 57, 408–417. [Google Scholar] [CrossRef]
- Hardeland, R.; Poeggeler, B. Melatonin and Synthetic Melatonergic Agonists: Actions and Metabolism in the Central Nervous System. Central Nervous System Agents in Medicinal Chemistry 12, 189–216.
- Hardeland, R. Melatonin and the Theories of Aging: A Critical Appraisal of Melatonin’s Role in Antiaging Mechanisms. Journal of Pineal Research 2013, 55, 325–356. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.; Osuna, C.; Gitto, E. Actions of Melatonin in the Reduction of Oxidative Stress. J Biomed Sci 2000, 7, 444–458. [Google Scholar] [CrossRef]
- Maestroni, G.J.M. Therapeutic Potential of Melatonin in Immunodeficiency States, Viral Diseases, and Cancer. In Tryptophan, Serotonin, and Melatonin: Basic Aspects and Applications; Huether, G., Kochen, W., Simat, T.J., Steinhart, H., Eds.; Advances in Experimental Medicine and Biology; Springer US: Boston, MA, 1999; pp. 217–226. ISBN 978-1-4615-4709-9. [Google Scholar]
- Simonneaux, V.; Ribelayga, C. Generation of the Melatonin Endocrine Message in Mammals: A Review of the Complex Regulation of Melatonin Synthesis by Norepinephrine, Peptides, and Other Pineal Transmitters. Pharmacol Rev 2003, 55, 325–395. [Google Scholar] [CrossRef] [PubMed]
- Gümüşova, S.; Memi̇ş, Y.S. Bazı Viral Enfeksiyonlarda Melatoninin Etkileri. Atatürk Üniversitesi Veteriner Bilimleri Dergisi 2014, 9, 50–54. [Google Scholar] [CrossRef]
- Liu, J.; Clough, S.J.; Hutchinson, A.J.; Adamah-Biassi, E.B.; Popovska-Gorevski, M.; Dubocovich, M.L. MT1 and MT2 Melatonin Receptors: A Therapeutic Perspective. Annual Review of Pharmacology and Toxicology 2016, 56, 361–383. [Google Scholar] [CrossRef]
- Hotchkiss, A.K.; Nelson, R.J. Melatonin and Immune Function: Hype or Hypothesis? Crit Rev Immunol 2002, 22, 351–371. [Google Scholar]
- Lissoni, P.; Barni, S.; Tancini, G.; Mainini, E.; Piglia, F.; Maestroni, G.J.; Lewinski, A. Immunoendocrine Therapy with Low-Dose Subcutaneous Interleukin-2 plus Melatonin of Locally Advanced or Metastatic Endocrine Tumors. Oncology 1995, 52, 163–166. [Google Scholar] [CrossRef]
- Topal, T.; Öter, S.; Korkmaz, A. Melatonin ve Kanserle Ilişkisi: Journal of General Medicine / Genel Tıp Dergisi. Journal of General Medicine / Genel Tıp Dergisi 2009, 19, 137–143. [Google Scholar]
- Escames, G.; Guerrero, J.M.; Reiter, R.J.; Garcia, J.J.; Munoz-Hoyos, A.; Ortiz, G.G.; Oh, C.S. Melatonin and Vitamin E Limit Nitric Oxide-Induced Lipid Peroxidation in Rat Brain Homogenates. Neuroscience Letters 1997, 230, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Touitou, Y. Human Aging and Melatonin. Clinical Relevance. Experimental Gerontology 2001, 36, 1083–1100. [Google Scholar] [CrossRef] [PubMed]
- Salt, A.; Çenesiz, M.; Çenesiz, S. Melatonin, Etkileri ve Kullanım Alanları. Etlik Vet. Mik. Derg. 2017, 28, 7–12. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Osuna, C.; Gitto, E. Actions of Melatonin in the Reduction of Oxidative Stress. A Review. J Biomed Sci 2000, 7, 444–458. [Google Scholar] [CrossRef]
- Reiter, R.J.; Calvo, J.R.; Karbownik, M.; Qi, W.; Tan, D.X. Melatonin and Its Relation to the Immune System and Inflammation. Annals of the New York Academy of Sciences 2000, 917, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J. Interactions of the Pineal Hormone Melatonin with Oxygen-Centered Free Radicals: A Brief Review. Braz J Med Biol Res 1993, 26, 1141–1155. [Google Scholar] [PubMed]
- Garfinkel, D.; Laudon, M.; Nof, D.; Zisapel, N. Improvement of Sleep Quality in Elderly People by Controlled-Release Melatonin. Lancet 1995, 346, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Macchi, M.M.; Bruce, J.N. Human Pineal Physiology and Functional Significance of Melatonin. Frontiers in Neuroendocrinology 2004, 25, 177–195. [Google Scholar] [CrossRef]
- Arendt, J. Melatonin, Circadian Rhythms, and Sleep. New England Journal of Medicine 2000, 343, 1114–1116. [Google Scholar] [CrossRef] [PubMed]
- Oztekin, E.; Mogulkoc, R.; Baltaci, A.K.; Tiftik, A.M. The Influence of Estradiol and Progesterone and Melatonin Supplementation on TNF-Alpha Levels in Ovariectomized and Pinealectomized Rats. Acta Biol Hung 2006, 57, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Oztürk, A.; Baltaci, A.K.; Bediz, C.S.; Mogulkoc, R.; Güngör, S. Effects of Zinc and Melatonin Deficiency on Testicular Tissue of Rats. Biol Trace Elem Res 2003, 96, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Baltaci, A.K. Melatonin, immün sistem ve çinko. Selcuk Med J 2001, 17, 267–272. [Google Scholar]
- Wurtman, R.J.; Axelrod, J.; Chu, E.W. Melatonin, a Pineal Substance: Effect on the Rat Ovary. Science 1963, 141, 277–278. [Google Scholar] [CrossRef]
- Nagy, P.; Guillaume, D.; Daels, P. Seasonality in Mares. Animal Reproduction Science 2000, 60–61, 245–262. [Google Scholar] [CrossRef]
- Uyar, A.; Alan, M. Koyunlarda Erken Anöstrüs Döneminde Melatonin Uygulamalarının Ovulasyon Ve Gebelik Üzerine Etkisi. YYU Vet Fak Derg 2008, 19, 47–54. [Google Scholar]
- Ölmez, E.; Şahna, E.; Ağkadi̇r, M.; Acet, A. MELATONİN: EMEKLİLİK YAŞI 80 OLUR MU? Turgut Özal Tıp Merk Derg 2000, 7. [Google Scholar]
- Ninomiya, T.; Iwatani, N.; Tomoda, A.; Miike, T. Effects of Exogenous Melatonin on Pituitary Hormones in Humans. Clinical Physiology 2001, 21, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Turgut, M.; Uysal, A.; Yurtseven, M. Epifiz Bezinin Morfolojik Özellikleri, Embriyolojik Gelişimi ve Deneysel Greftleme İşlemleri. aktd 2003, 12. [Google Scholar]
- Mollaoğlu, H.; Özgüner, M. fehmi Yaşlanma Sürecinde Melatoninin Rolü. SDÜ Tıp Fak Derg 2009, 12, 52–56. [Google Scholar] [CrossRef]
- Kerman, M.; Cirak, B.; Ozguner, M.F.; Dagtekin, A.; Sutcu, R.; Altuntas, I.; Delibas, N. Does Melatonin Protect or Treat Brain Damage from Traumatic Oxidative Stress? Exp Brain Res 2005, 163, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Spence, D.W.; Pandi-Perumal, S.R.; Trakht, I.; Cardinali, D.P. Jet Lag: Therapeutic Use of Melatonin and Possible Application of Melatonin Analogs. Travel Medicine and Infectious Disease 2008, 6, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Diabetic Goto Kakizaki Rats as Well as Type 2 Diabetic Patients Show a Decreased Diurnal Serum Melatonin Level and an Increased Pancreatic Melatonin-receptor Status - Peschke - 2006 - Journal of Pineal Research - Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1600-079X.2005.00287.x (accessed on 29 March 2024).
- Peschke, E.; Stumpf, I.; Bazwinsky, I.; Litvak, L.; Dralle, H.; Mühlbauer, E. Melatonin and Type 2 Diabetes - a Possible Link? J Pineal Res 2007, 42, 350–358. [Google Scholar] [CrossRef]
- Wolden-Hanson, T.; Mitton, D.R.; McCants, R.L.; Yellon, S.M.; Wilkinson, C.W.; Matsumoto, A.M.; Rasmussen, D.D. Daily Melatonin Administration to Middle-Aged Male Rats Suppresses Body Weight, Intraabdominal Adiposity, and Plasma Leptin and Insulin Independent of Food Intake and Total Body Fat. Endocrinology 2000, 141, 487–497. [Google Scholar] [CrossRef]
- Cagnacci, A.; Arangino, S.; Renzi, A.; Paoletti, A.M.; Melis, G.B.; Cagnacci, P.; Volpe, A. Influence of Melatonin Administration on Glucose Tolerance and Insulin Sensitivity of Postmenopausal Women. Clin Endocrinol (Oxf) 2001, 54, 339–346. [Google Scholar] [CrossRef]
- Champney, T.H.; Steger, R.W.; Christie, D.S.; Reiter, R.J. Alterations in Components of the Pineal Melatonin Synthetic Pathway by Acute Insulin Stress in the Rat and Syrian Hamster. Brain Res 1985, 338, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Plant Growth Regulator and/or Biostimulator during Stress? Trends in Plant Science 2014, 19, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Xie, Y.; Zhang, Z.; Chen, L. Melatonin: A Multifunctional Factor in Plants. Int J Mol Sci 2018, 19, 1528. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S. Novel Perspectives on the Molecular Crosstalk Mechanisms of Serotonin and Melatonin in Plants. Plant Physiology and Biochemistry 2018, 132, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lv, Y.; Shi, Y.; Li, T.; Chen, Y.; Zhao, D.; Zhao, Z. The Role of Phyto-Melatonin and Related Metabolites in Response to Stress. Molecules 2018, 23, 1887. [Google Scholar] [CrossRef]
- Acharya, D.; Satapathy, S.; Somu, P.; Parida, U.K.; Mishra, G. Apoptotic Effect and Anticancer Activity of Biosynthesized Silver Nanoparticles from Marine Algae Chaetomorpha Linum Extract Against Human Colon Cancer Cell HCT-116. Biol Trace Elem Res 2021, 199, 1812–1822. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Xie, C.; Zhang, H.; Arnao, M.B.; Ali, M.; Ali, Q.; Muhammad, I.; Shalmani, A.; Nawaz, M.A.; Chen, P.; et al. Melatonin and Its Effects on Plant Systems. Molecules 2018, 23, 2352. [Google Scholar] [CrossRef] [PubMed]
- Van Tassel, D.L.; O’Neill, S.D. Putative Regulatory Molecules in Plants: Evaluating Melatonin. Journal of Pineal Research 2001, 31, 1–7. [Google Scholar] [CrossRef]
- Burkhardt, S.; Tan, D.X.; Manchester, L.C.; Hardeland, R.; Reiter, R.J. Detection and Quantification of the Antioxidant Melatonin in Montmorency and Balaton Tart Cherries (Prunus Cerasus). J Agric Food Chem 2001, 49, 4898–4902. [Google Scholar] [CrossRef]
- Wei, Y.; Liu, G.; Chang, Y.; Lin, D.; Reiter, R.J.; He, C.; Shi, H. Melatonin Biosynthesis Enzymes Recruit WRKY Transcription Factors to Regulate Melatonin Accumulation and Transcriptional Activity on W-Box in Cassava. J Pineal Res 2018, 65, e12487. [Google Scholar] [CrossRef]
- Tan, D.-X.; Manchester, L.C.; Liu, X.; Rosales-Corral, S.A.; Acuna-Castroviejo, D.; Reiter, R.J. Mitochondria and Chloroplasts as the Original Sites of Melatonin Synthesis: A Hypothesis Related to Melatonin’s Primary Function and Evolution in Eukaryotes. Journal of Pineal Research 2013, 54, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Protective Effect of Melatonin against Chlorophyll Degradation during the Senescence of Barley Leaves. Journal of Pineal Research 2009, 46, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Campbell, S.S.B.; Saxena, P.K. The Role of Serotonin and Melatonin in Plant Morphogenesis: Regulation of Auxin-Induced Root Organogenesis in in Vitro-Cultured Explants of St. John’s Wort (Hypericum Perforatum L.). In Vitro Cell.Dev.Biol.-Plant 2001, 37, 786–793. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. The Physiological Function of Melatonin in Plants. Plant Signal Behav 2006, 1, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin Promotes Adventitious- and Lateral Root Regeneration in Etiolated Hypocotyls of Lupinus Albus L. J Pineal Res 2007, 42, 147–152. [Google Scholar] [CrossRef]
- Zhang, R.; Sun, Y.; Liu, Z.; Jin, W.; Sun, Y. Effects of Melatonin on Seedling Growth, Mineral Nutrition, and Nitrogen Metabolism in Cucumber under Nitrate Stress. J Pineal Res 2017, 62. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wei, J.-P.; Scott, E.R.; Liu, J.-W.; Guo, S.; Li, Y.; Zhang, L.; Han, W.-Y. Exogenous Melatonin Alleviates Cold Stress by Promoting Antioxidant Defense and Redox Homeostasis in Camellia Sinensis L. Molecules 2018, 23, 165. [Google Scholar] [CrossRef]
- Aguilera, Y.; Herrera, T.; Liébana, R.; Rebollo-Hernanz, M.; Sanchez-Puelles, C.; Martín-Cabrejas, M.A. Impact of Melatonin Enrichment during Germination of Legumes on Bioactive Compounds and Antioxidant Activity. J Agric Food Chem 2015, 63, 7967–7974. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of Melatonin in Plants: A Review. J Pineal Res 2015, 59, 133–150. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Synthesis from Tryptophan and Its Role in Higher Plant. Amino acids in higher plants 2015, 390–435. [Google Scholar] [CrossRef]
- Sarropoulou, V.; Dimassi-Theriou, K.; Therios, I.; Koukourikou-Petridou, M. Melatonin Enhances Root Regeneration, Photosynthetic Pigments, Biomass, Total Carbohydrates and Proline Content in the Cherry Rootstock PHL-C (Prunus Avium × Prunus Cerasus). Plant Physiology and Biochemistry 2012, 61, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Assessment of Different Sample Processing Procedures Applied to the Determination of Melatonin in Plants. Phytochem Anal 2009, 20, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin and Reactive Oxygen and Nitrogen Species: A Model for the Plant Redox Network. Melatonin Research 2019, 2, 152–168. [Google Scholar] [CrossRef]
- Manchester, L.C.; Tan, D.-X.; Reiter, R.J.; Park, W.; Monis, K.; Qi, W. High Levels of Melatonin in the Seeds of Edible Plants: Possible Function in Germ Tissue Protection. Life Sciences 2000, 67, 3023–3029. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Protective Effect of Melatonin against Chlorophyll Degradation during the Senescence of Barley Leaves. J Pineal Res 2009, 46, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Posmyk, M.M.; Kuran, H.; Marciniak, K.; Janas, K.M. Presowing Seed Treatment with Melatonin Protects Red Cabbage Seedlings against Toxic Copper Ion Concentrations. J Pineal Res 2008, 45, 24–31. [Google Scholar] [CrossRef]
- Kolář, J.; Johnson, C.H.; Macháčková, I. Exogenously Applied Melatonin (N-Acetyl-5-Methoxytryptamine) Affects Flowering of the Short-Day Plant Chenopodium Rubrum. Physiologia Plantarum 2003, 118, 605–612. [Google Scholar] [CrossRef]
Figure 1.
N-acetyl-5-methoxy tryptamine (Melatonin).
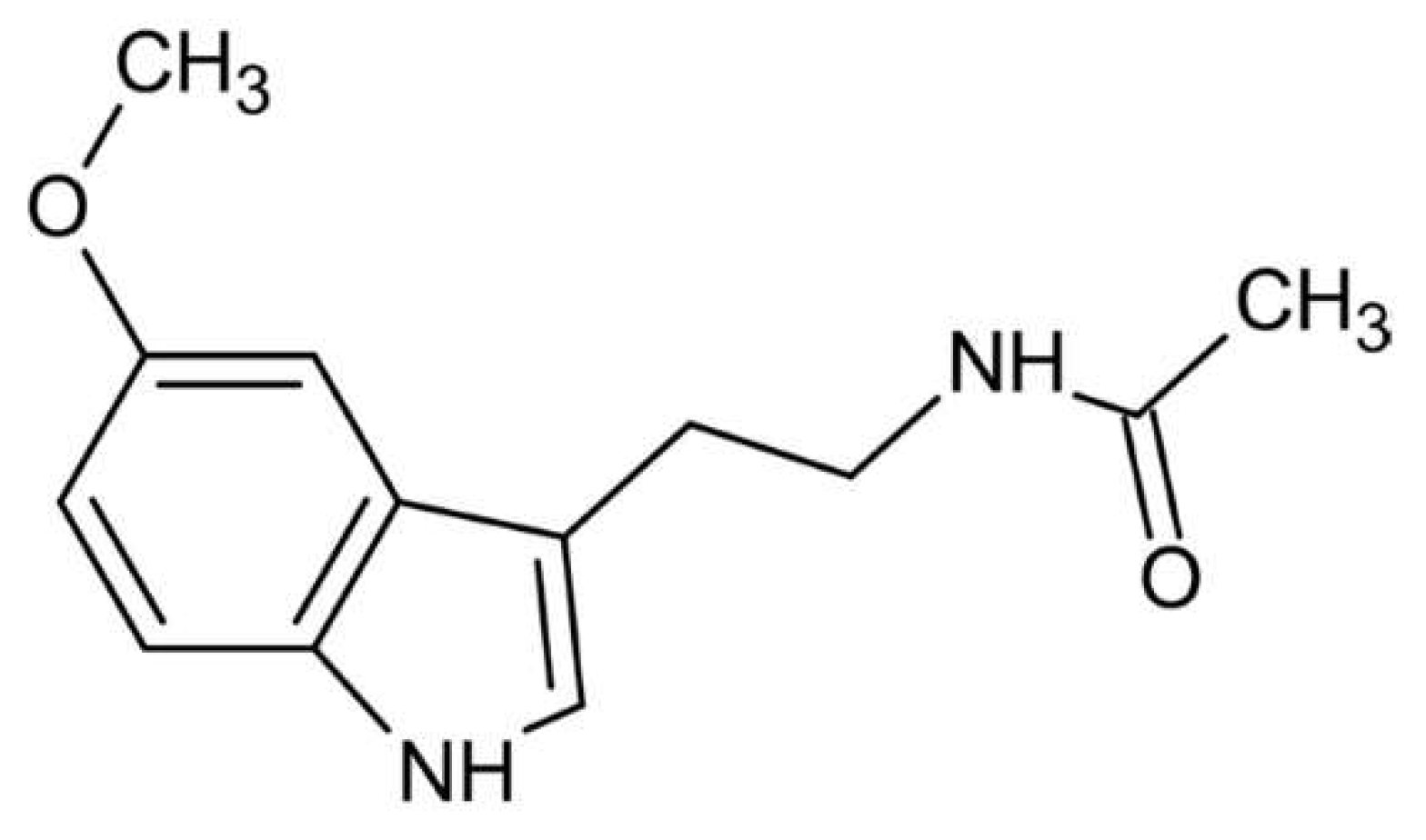
Figure 2.
Melatonin synthesis [23].
Figure 2.
Melatonin synthesis [23].
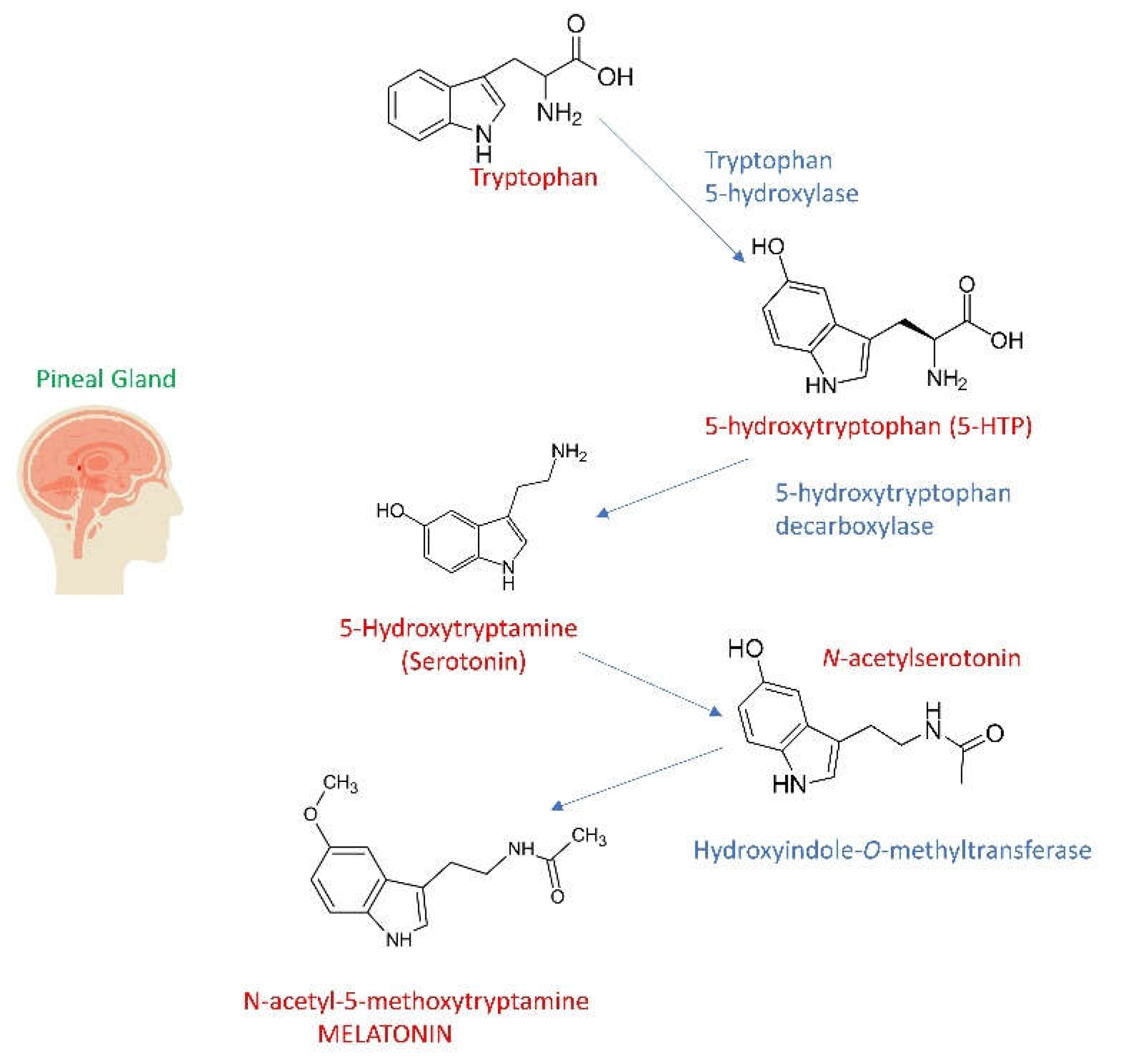
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



