Submitted:
13 April 2024
Posted:
16 April 2024
You are already at the latest version
Abstract
Infection with the malaria parasite Plasmodium, remains a major cause of illness and death world-wide. The development of evasion strategies by the parasite from host immunity and its adaptability to antimalarial drugs has raised the urgency for developing new strategies to combat this disease. One promising avenue is targeting telomeres – sequences found at chromosome termini– and telomerase – the enzyme that catalysis synthesis of telomeres using an intrinsic RNA template. As telomeres shorten critically, cells undergo replicative senescence or apoptosis. Plasmodium, lacking telomerase activity in its human stages, exploits alternative mechanisms, accelerating telomere shortening, aging, and immune evasion. Chemical telomerase inhibitors show promise as antimalarials. In this review, we examine the potential of targeting this system in malaria therapy, with telomerase inhibitors offering a novel approach. A systematic search on PubMed using Boolean techniques identified 246 relevant articles from 1985 to March 31, 2024. After filtering, 43 articles met inclusion criteria, supplemented by snowball sampling. The telomerase inhibitor drug strategy is widely applicable against cancer. However, given the drugability of telomerase and the absolute requirement for active telomere maintenance in the vast majority of parasites including plasmodium, telomerase and telomere maintenance could well represent the Achilles heel of the parasite
Keywords:
Malaria
; Telomeres Maintenance
; Plasmodium
; Therapeutic Target
Introduction
Malaria infection poses a significant public health burden, affecting over half of the population worldwide. The disease, transmitted by female Anopheles mosquitoes and caused by the Plasmodium parasite, persists as a formidable threat. Plasmodium falciparum and Plasmodium vivax, among the five identified parasites causing human malaria, pose a substantial public health threat and economic burden [1]. In 2022, a total of 249 million malaria cases and 608,000 deaths were reported globally across 85 countries. The Sub-Saharan African area experiences the greatest impact of malaria, accounting for 94% of instances (equivalent to 233 million cases) and 95% of fatalities (amounting to 580,000 deaths), with roughly 80% of these deaths occurring in children under the age of 5. [2,168]. The years coinciding with the peak of COVID-19 pandemic, added an extra burden to malaria control efforts, with a distressing increase in new cases and deaths during this period [3,4]. The African region, particularly countries like Nigeria, Congo, Tanzania, and Mozambique remains the hardest hit, accounting for over half of the global cases [1,2,5]. According to GBD 2016 Healthcare Access and Quality Collaborators, 2018, these challenges are partly due to limited resources and inadequate healthcare systems.
Over the years, numerous malaria control measures have been implemented to lower disease incidence and mortality rates, including the development of better healthcare systems, increased funding, use of vector-repellent bed nets, Indoor residual sprays (IRS), and artemisinin combination treatments (ACTs), the challenges of malaria control measures keep increasing in sub-Saharan Africa [7,8].These statistics emphasize the urgent need for sustained efforts to combat malaria, especially in heavily affected regions. One of the underexplored areas that provide new avenues for malaria therapy is the mechanistic pathways of genomic stability of both plasmodium and humans during host-parasite interaction [9,10]. The telomere-telomerase system which composed of cellular elements that play essential role in maintaining genomic integrity and regulating cellular aging, offers innovative possibilities in targeting malaria [11,12,13,14]. Telomeres are specialized repetitive DNA sequences at linear chromosome ends, act as protective chromosomes ends from degradation [15,16]. Together with its associated proteins, telomeres prevent the loss of genetic information during cell division and protect chromosome ends from degradation and fusion. The DNA sequences of telomeres typically consist of tandem repeats of short nucleotide sequences, such as TTAGGG in humans, extending for hundreds to thousands of base pairs (Figure 1).
- hTERT: Human Telomerase Reverse Transcriptase; telomere elongation, maintaining chromosome stability. hTERC: Human Telomerase RNA Component; template for telomere synthesis, enabling telomerase action.
- POT1: Protection of Telomeres 1, TPP1: Adrenocortical Dysplasia Homolog (ACD) Protein or TINT1; telomerase recruitment to telomeres. TERF1: Telomeric Repeat-binding Factor 1; telomere protection. TERF2IP: Telomeric Repeat-binding Factor 2 Interacting Protein; stabilizes telomeres against chromosome end fusions. TIN2: TRF1-Interacting Nuclear Factor 2; Mediates interactions between telomere proteins, maintaining telomere integrity. DKC1: Dyskerin Pseudouridine Synthase 1; Modifies telomerase RNA, facilitating telomerase complex assembly. NHP2: Nucleolar Protein 2; Aids in telomerase complex assembly, crucial for telomere elongation. NOP10: Nucleolar Protein 10; Essential for telomerase stability and function in telomere maintenance. GAR1: Guide to the Function of NHP2-like Protein 1; Maintains telomerase RNA stability, ensuring proper telomerase function. TCAB1: Telomerase Cajal Body Protein 1; Guides telomerase to nuclear compartments.
This repetitive nature allows telomeres to cap the chromosome ends, shielding them from DNA breaks repair enzymes and preventing degradation or fusion with neighbouring chromosomes [18,19]. Telomeres undergo shortening with each round of cell division. This is as a result of a phenomenon of the end-replication problem, where the lagging strand cannot replicate to full length [20,21,22]. As a result, telomeres gradually erode over time, contributing to cellular aging and senescence [23]. Ultimately, as telomeres reach a critical threshold, cells enter a state of replicative senescence or apoptosis, constraining their ability to proliferate and contributing to tissue aging and overall decline in organismal health [24,25,26].
To counteract telomere shortening, some cells express an enzyme called telomerase [27,28]. Telomerases are a specialized ribonucleoprotein complexes that add repetitive DNA sequences at the ends of telomeres, thereby maintaining or even lengthening them. Telomerase consists of a catalytic protein subunit, which carries out the synthesis of telomeric DNA, and an RNA component that serves as a template for telomere extension [29,30]. The activity of the enzyme telomerase is typically high in embryonic cells, germline, and certain stem cell populations, but is typically repressed in most somatic cells [31]. However, aberrant activation of telomerase or alterations in telomere maintenance mechanisms can contribute to various diseases, including cancer, where unchecked telomere elongation enables unlimited cell proliferation [32,33]. Understanding the dynamics of telomeres and telomerase is crucial for elucidating the mechanisms underlying cellular aging, disease development, and potential therapeutic interventions aimed at modulating telomere dynamics to promote health and longevity [24,28,34,35].
While telomere shortening is linked to aging and diseases like cancer, Plasmodium parasites possess active telomerase, allowing them to evade cell death [36,37,38]. Targeting telomeres and telomerase emerges as a promising strategy for malaria therapy, with studies indicating that inhibiting telomerase activity can kill Plasmodium parasites [14,39,40]. Telomerase activity is crucial for maintaining telomere length and enabling unlimited cellular proliferation, a characteristic shared by both cancer cells and many single-cell eukaryotic pathogens [32,41,42]. While telomerase's role in cancer has been extensively studied, its significance in human parasites remained largely unexplored. These parasites, like cancer cells, depend on a continuous enzyme activity of telomerase to sustain their proliferative ability within their hosts [42,43,44]. Studying telomerase dyanamics in human parasites could reveal insights into their biology and identify new therapeutic targets for treating parasitic diseases. [45].
Telomeres and Telomerase implication in Health and Disease
Telomeres are structures that function as “caps” on the tips of linear chromosomes to maintain genomic stability. They prevent chromosome degradation and fusion by safeguarding the internal regions of the chromosomes [16]. These complexes of nucleoproteins are made up of repetitive DNA sequences and accompanying proteins, that shield and safeguard chromosome ends, preventing them from being identified as DNA damage [46]. Telomerase is a ribonucleoprotein that act as an enzyme for catalysing the replenishing of these sequences which are gradually lost each time a cell undergoes division [47]. This enzyme activity is typically elevated in germ line, embryonic cells, and certain stem cells that are constantly dividing [48]. Nevertheless, the enzyme's function is suppressed in most somatic cells [31]. Dysregulation of mechanisms responsible for maintaining telomeres, including shifts in telomerase enzyme activity or alterations in the normal length of telomeres, plays a significant role in the development of several diseases [24,27].
In various host-parasite interactions, specific telomere processes are involved in gene regulation. For the highly proliferative human pathogen P. falciparum, telomerase plays a dual role. It not only supports telomeres maintenance but also involve in repairing of damaged chromosome ends [37,49]. Telomere length (TL) is influenced by both genetic inheritance and environmental factors, such as reactive oxygen species (ROS) [49,50,51] and inflammation [52,53,54] through different mechanisms as highlighted in Figure 2. Other lifestyle factors contribute to telomere length alterations including nutrition [55,56,57,58].
Numerous pieces of evidence, including observational data indicate that individuals who have been exposed to infectious diseases tend to exhibit shorter telomeres [59]. As such, the implication infectious diseases exert on telomere/telomerase dynamics is increasingly recognized as a critical aspect of host-pathogen interactions and disease progression [60]. Evidence from human observational studies indicates that chronic viral infections and inflammatory liver diseases correlate with telomere attrition [61,62] in specific cells potentially compromising immunocompetence and longevity. Moreover, individuals with telomerase deficiency due to rare genetic disorders exhibit heightened vulnerability to infectious diseases, and other complications [63,64]. This supports the notion that telomerase plays a crucial role in immune function and disease resistance. In vitro experiments reveal that acute antigen exposure initially boosts telomerase activity in T lymphocytes [64,65], but repeated encounters lead to its decline, suggesting a link between immune activation and telomere dynamics [66]. Studies with telomerase-deficient mutant mice further supports the significance of telomerase in immune cell proliferation and function, as well as organ regeneration following injury [63]. Decreased telomere length has been implicated in viral infections including HIV [67], Hepatitis [62], and HPV [61].
Other research works agree on the concept that exposure to infectious agents could expedite telomere shortening, contributing to immunosenescence and cellular aging process. Notably, research in avian species highlights that encounters with pathogenic infections lead to telomere shortening in blood cells and multiple tissues [12,68]. Other studies provided evidence that long-term exposure to infection can reduce telomere length which in turn modulate the host's response to inflammatory signals throughout life [53,60,69].
Telomere Length and Telomerase dynamics in Malaria
Malaria mostly affects children, especially those below the ages of five (WHO, 2023). However, elderly people experience higher rates of morbidity and mortality from infectious diseases than young people, at least in part because of age-related declines in immune function that make them more vulnerable to infections [70,71]. Changes in both genes and environment contribute to the aging process. Many age-related diseases may originate in cellular aging [56], which is defined as a decline in cellular function, growth, and division. Infections have been linked to premature aging [72,73,74]. Recent progress in aging research indicates a reciprocal relationship between biological age and susceptibility to disease, suggesting that exposure to infectious diseases can expedite cellular aging [68,75]. Moreover, research has illustrated that telomeres undergo gradual erosion with cell division, eventually leading to cellular senescence or programmed cell death (Figure 4), unless they are sufficiently restored by telomerase [24,52].
Cellular stress orchestrated by parasite infection and immune response can lead to DNA damage and TL shortening. Telomerase helps restores telomeres which are vital for cell cycle, apoptosis, and cellular maintenance. Impaired telomerase enzymatic activity can precipitate aging, cellular senescence and immune function impairment [49]. The telomeres of Plasmodium parasites have attracted great interest from aging biologists. Telomere regulation has been implicated in a number of host-parasite interactions [14], including the promotion of telomere maintenance and chromosome end repair processes [37]. P. falciparum has been observed to accomplish the process of linear chromosome replication, thereby elucidating the parasites' ability to persist despite their high replication rate [76]. Additionally, disease severity depends on parasite load in the host species [77,78]. In P. falciparum, the telomere-associated sequence (TAS), which is 15–30 kb in length, precedes each of the 14 linear chromosomes of the parasite [79,80].
The telomere-associated sequence (TAS) on each chromosome consists of six distinct blocks of telomere-associated repeat elements (TAREs) [37,81]. Within the TAS region, the genes encoding the virulent var and rifin genes in P. falciparum, as well as the vir gene in P. vivax, are located [82,83,84].This region of the Plasmodium genome also contains telomere-associated silent information regulator 2 (Sir2), which plays a role in chromatin silencing. Sir2 is thought to bind to upstream promoter region of var genes, effectively silencing them. This results in regulation of expression of antigenic genes [85,86]. However, the aberrant lengthening of telomeres caused by Sir2A deficiency and its relationship with telomerase-mediated telomere length homeostasis and the regulation of virulence genes in P. falciparum are yet to be understood [44]. The telomerase of Plasmodium parasites could be targeted to prevent their multiplication and cell survival, opening novel malaria prevention strategies.
Interestingly, recent studies have reported a potential link between telomere length, telomerase activity, and malaria infection [72,73,74,87,88]. Other studies have reported an elevated telomerase activity in many cancers, while in malaria, telomerase may play a role in parasite development and virulence (Asghar et al. 2016b; Mohanty, Gupta, and Bhatnagar 2015; Religa et al. 2014). Recent research (past decade) demonstrates eukaryotic telomere transcription resulting in TERRA, a G-rich RNA molecule [91]. Plasmodium species, during their complex life cycle, infect both human and mosquito hosts, leading to chronic infections and repeated invasion of red blood cells (Figueiredo and Scherf 2005). This repetitive infection cycle places substantial stress on the telomeres of both parasite and host cells, necessitating the maintenance of telomere length for survival and propagation [92].
In recent years, research has revealed that eukaryotic telomeres undergo transcription, generating a G-rich RNA known as TERRA. Initially, it was thought that TERRA solely served a structural role and did not contain any protein-coding sequences [91]. However, the discovery of two novel signaling proteins encoded by telomeres may provide a new biomarker for biological age, and a simple blood test for these proteins may provide an early warning of cancer, inflammation, and malaria infection [93,94]. Further research on telomerase in malaria is imperative due to its pivotal role in controlling telomere length, which is linked to the advancement of these diseases. Understanding telomerase's function and activity may facilitate the creation of improved diagnostic methods and targeted treatments for malaria and cancer, potentially enhancing patient outcomes and survival rates [93,95].
Diagnostic and Prognostic Potential of Telomeres and Telomerase in Malaria
Recent research has explored the diagnostic potential of telomere and telomerase dynamics in malaria [73,88]. Studies highlight that TL in lymphocytes of infected patients is quite shorter as compared to uninfected cells [96,97], and that telomerase enzyme activity can be measured in serum of malaria patients [93]. It has been demonstrated that variations of TL in malaria parasites correlates with disease severity and treatment outcomes [72]. Other reports have revealed correlations between diminished immune function and reduced telomere length in blood samples [69,89]. However, changes in TL may be considered as a biomarker of disease and not the underlining cause [98]. Additionally, quantification of telomerase activity has already shown promise as a biomarker for detecting malaria infection and monitoring treatment response [74].
Research has firmly established that reduced telomere lengths in leukocytes serve as a predictor of poor prognostic outcomes, suggesting compromised immune functions and heightened susceptibility to diseases and infections, including malaria [98,99]. Additionally, TL attrition has been linked with risk of overall mortality [100]. Moreover, advanced molecular techniques like qPCR and LAMP has been shown to enhance the detection and measurement of telomeres and telomerase enzyme activity in clinical samples [38,77,101,102].
Telomere Length Regulation in Malaria Parasite
Telomerase systems in Plasmodium and humans differ significantly. Table 1 summarizes the key differences between the two systems [37]. In Plasmodium parasites, telomerase exhibit an essential role in the preservation of, T.L. This enzyme comprises two integral components: telomerase reverse transcriptase (TERT) and the telomerase RNA component (TERC) [29,90,103]. The TERT protein possess the catalytic functions of the enzyme, while TERC RNA is a template to be used the synthesis of new telomeric sequences [37]. The TERT protein in P. falciparum has an N-terminal extension that is absent in human TERT, and TERC RNA lacks the conserved template region found in human TERC [104,105]. This contrast extends to telomerase activity regulation, as Plasmodium maintains stable telomere length through tight regulation, whereas human telomerase is generally repressed, leading to gradual telomere shortening [88,106].
Interestingly, Plasmodium telomeres exhibit rapid elongation and recombination-based exchange, which distinguishes them from the dynamics observed in human telomeres. Although telomerase is not normally expressed in most somatic cells, it has been shown to be overexpressed in some adult stem cells, embryonic stem cells, and cancer cells [107].
It has been reported that inhibition of telomerase enzyme activity in Plasmodium can result to gradual loss of telomere function and, ultimately, cell death [44]. Human telomerase, hTERT is typically expressed only in few stem cells and cancer cells, whereas in Plasmodium species, telomerase is constitutively expressed throughout the parasite life cycle [108,109]. The telomeres of Plasmodium parasites are generally much shorter than those of human cells. For instance, while human somatic cells typically have telomeres ranging from 5-15 kb, those in P. falciparum are notably shorter, measuring only approximately 1-2 kb on average [36,88]. One other key difference that characterize the human telomerase is that it is primarily responsible for maintaining telomere length, while in Plasmodium parasites, telomere maintenance involves a combination of telomerase-mediated extension and recombination-based mechanisms [13,44]. Although telomerase activity is not essential for replication in most human cells, it has been shown to be very crucial for Plasmodium replication [44,110].
In addition, Plasmodium telomerase activity is regulated by both cell cycle molecules and telomere-associated proteins, while hTERT is modulated by a complex network of factors, including telomere-binding proteins, epigenetic modifications, and signalling pathways [35,111] . The telomerase system in Plasmodium species has evolved independently from that in humans, and appears to have undergone extensive diversification and adaptation to the unique requirements for the parasite's life cycle [112,113]. Telomerase enzyme of Plasmodium species may have roles in evading host immune responses. For example, it has been suggested that telomerase-mediated telomere maintenance in Plasmodium may help the parasite avoid detection and destruction by the host immune system [88,114].
Furthermore, the differences between the Plasmodium and human telomerase systems have made telomerase an attractive target for the development of new antimalarial drugs [36,40,45,115]. Several telomerase inhibitors have been identified that specifically target Plasmodium telomerase have shown promise as potential therapeutic agents. Linear DNA replication presents a challenge due to incomplete copying at the 5' ends, resulting in 3' overhangs rich in guanine (G-overhangs) on both strands. strands [116]. Consequently, these overhangs necessitate protection from cellular enzymes to prevent the loss of genetic information [117,118]. Cells have evolved two primary mechanisms to safeguard their chromosomal termini and prevent DNA loss [119,120].
One of these mechanisms involves telomeres. These repetitive DNA sequences were initially considered nonessential for gene expression, telomeres gradually shorten with each cell division [23] as depicted in Figure 5. When telomeres shortens, the cell gradually loses its capacity to divide, ultimately leading to cell death [121,122].
The second crucial component of this protective system is telomerase. This protein enzyme is used for maintaining TL [123]. This is achieved by adding specific DNA repeat sequences to the chromosome termini. Notably, telomerase exhibits activity in specific cell types, such as stem cells and germ cells, while remaining inactive in most other cell types [124].
In humans, telomere shortening has been associated with cellular aging and contributes to the development of various diseases. This phenomenon leads to a state known as cell senescence, characterized by irreversible growth arrest. In contrast, cancer cells often reactivate telomerase, allowing them to evade cell senescence and continue unchecked proliferation [125]. The prospect of targeting telomeres and telomerase holds promise as a strategy for developing novel treatments for cancer and other diseases. However, it is essential to emphasize that telomerase also plays a crucial role in the normal functioning of certain cells, such as stem cells. Consequently, any efforts to target telomeres and telomerase must be designed with precision to ensure their safety and effectiveness in preserving the functionality of these critical cell types [40,126,127].
Top of Form
In humans and other mammals, telomere sequences gradually shorten with each replication cycle until they reach a critical point called "Hayflick limit," after which cells enter into replicative senescence, see Figure 6 [107]. This process of telomere shortening serves as a protective mechanism against uncontrolled cell proliferation, including tumour cells [107]. However, what was originally conceived as a protective mechanism soon became problematic, as it can lead to various aberrations in cellular processes, such as the loss of telomere G-overhang, resection of the C strand, increased recombination frequency at chromosome ends, chromosome fusion, genome instability, altered gene expression patterns, growth arrest, and cell death [121,128].
Telomerase activity is subject to tight regulation but is expressed in highly proliferative cells to preserve telomere length and prevent cell death and growth arrest [35,124]. Researchers have reported traditional methods of protecting human cells against disease through antioxidant protection using specific diet [129], treatment of Hemolytic and inflammatory diseases [130,131]. However, the efforts have not been enough to protect cellular senescence and death in malaria. The mechanisms governing telomere length maintenance are currently under active area of research, as aberrations in these mechanisms have been associated with variety of human ailment, including cancer [132,133], cardiovascular and nervous disorders [125], and malaria [72,73,89]. Notably, the Plasmodium parasite employs telomerase to maintain the length of its telomeres during replication [13,37]. This presents an opportunity to target the telomerase enzyme, disrupting the parasite's ability to replicate and cause disease [134].
Telomeres and Telomerase-Targeting Drugs
The persistent global challenge of malaria, primarily attributed to the Plasmodium species necessitates the continuous exploration of innovative therapeutic targets [135,136].Telomeres and the enzyme telomerase have attracted significant attention of scientists in the field of oncology due to their roles in cellular senescence and immortality [137,138]. The exploration of telomeres and telomerase-targeting strategies extends beyond cancer research. They are also implicated in the pathogenesis of infectious diseases, notably malaria [59,60,89]. Recent insights suggest that these components also play essential role in the survival and proliferation of Plasmodium parasites, presenting a unique target for antimalarial intervention [37,42,134]. Telomerase, particularly its TERT and TERC, is essential for telomere elongation, facilitating the parasite's replication and immune evasion within the host [88,103,114,139].
Malaria parasites exploit this telomere-telomerase system to replicate within the host cells [92,140]. This essential system plays a pivotal role in preserving the stability of the parasite's chromosomes during replication, facilitating continued replication, and inducing damage to the host's red blood cells [134,141]. Targeting this system emerges as an attractive focus for both diagnostic and therapeutic applications against Plasmodium parasites, offering the potential to disrupt parasite replication and control its spread [39,106,120,120]. In the quest for innovative antimalarial interventions, strategizing for targeting telomeres and telomerase could present a promising avenue for tackling this disease. While telomerase remains a widely explored target for disease treatment, there hasn't been a successful development of a telomerase-targeted inhibitor that has advanced to late-stage human clinical trials [142]. Given the pivotal role of telomerase in maintaining telomere length and genomic stability in Plasmodium parasites, efforts are underway to exploit this vulnerability for therapeutic purposes [34,120]. Efforts to target telomerase in Plasmodium parasites aim to selectively target telomerase in parasites and disrupt their ability to maintain telomere length, leading to genomic instability and cell death while sparing host cells [143].
There are several approaches that are currently explored, one of them include the design of inhibitors that specifically target telomerase activity in Plasmodium parasites [36,40,144]. Additionally, gene editing technologies such as CRISPR-Cas9 may offer a means to disrupt telomerase function in these parasites, potentially leading to new avenues for anti-malarial drug development [145,146,147]. A spectrum of telomere-telomerase targeting approaches which have been demonstrated in cancer, is illustrated in Figure 8.
GRN163(L), DN-TERT, and BIBR1532
GRN163(L), DN-TERT, and BIBR1532 are three distinct compounds that have demonstrated the ability to directly inhibit telomerase activity. These compounds exert their effects by targeting key components of the telomerase enzyme, such as the telomerase reverse transcriptase (TERT) or the telomerase RNA component (TERC). By of telomerase, these compounds interfere with the enzymatic function by disrupting its ability to extend telomeres, As a result, cells, including cancer cells, are unable to maintain their telomeres' length, leading to instability of the genome and ultimately cellular senescence and death [148,149,150].
BRACO19, RHPS4, and Telomestatin
BRACO19, RHPS4, and Telomestatin represent a class of compounds that have garnered significant attention for their ability to induce the formation of G-quadruplex structures at telomere termini (Figure 7). These structures, composed of guanine-rich sequences, are stable secondary DNA structures that can interfere with telomerase activity and impede telomere elongation. By promoting the formation of G-quadruplexes, BRACO19, RHPS4, and Telomestatin disrupt the normal function of telomerase, ultimately leading to telomere shortening and cellular senescence or apoptosis [151]. This approach has shown promise as a potential anticancer strategy, as cancer cells, which often exhibit high telomerase activity, are particularly vulnerable to telomere disruption. The development and optimization of compounds that target G-quadruplex formation at telomeres offer new avenues for the design of innovative cancer therapies aimed at selectively targeting telomere maintenance mechanisms.These compounds have been shown to induce G-quadruplex structural formation at telomere termini [152,153,154].
T-Oligo
T-oligo, a synthetic oligonucleotide, acts as a mimic for dysfunctional telomeres, thereby initiating cellular responses that culminate in the arrest of cell cycle process and senecence. This compound exerts its effects by activating ATM/ATR-mediated pathways, key regulators of DNA damage responses [155]. Upon exposure to T-oligo, cells perceive the presence of dysfunctional telomere-like structures, triggering a cascade of signaling events modulated by ataxia telangiectasia mutated(ATM) and ataxia telangiectasia and Rad3-related(ATR) protein kinases. These kinases orchestrate the activation of downstream effectors involved in DNA damage repair, cell cycle checkpoint control, and cellular senescence induction (Figure 8). Consequently, T-oligo-induced activation of ATM/ATR pathways halts cell cycle progression and promotes the establishment of a senescent phenotype, rendering cells incapable of further proliferation [148,156,157,158].
Immunization Involving Peptides Derived from TERT or the Introduction of TERT mRNA into Dendritic Cells
Immunization strategies utilizing peptides derived from TERT or the inoculation of TERT genes into dendritic cells offer innovative approaches to harnessing the immune system's potential in combating cancer [159]. Peptides derived from TERT, when administered as part of an immunization protocol, serve as antigens capable of activating both T and B cells. Upon exposure to these TERT-derived peptides, dendritic cells process and present them to T cells via major histocompatibility complex (MHC) molecules (Figure 8). This presentation triggers the activation and proliferation of T cells, including cytotoxic T lymphocytes (CTLs) and helper T cells, which play crucial roles in antitumor immunity [160]. Furthermore, B cells can recognize TERT-derived peptides as antigens, leading to the production of TERT-specific antibodies that contribute to the immune response against cancer cells expressing TERT. Alternatively, the introduction of TERT mRNA into dendritic cells serves as a means to directly activate the immune system. Dendritic cells transfected with TERT mRNA can express TERT protein, which is then presented to T cells, eliciting an immune response against cancer cells expressing TERT. This activation of T and/or B cells enables the recognition and elimination of cells expressing high TERT levels, thereby bolstering the immune system's ability to target and eradicate parasitic cells [45,66,157].
Figure 8.
Drugs Targeting Telomeres and Telomerase. Figure by T. N. WAKAI, designed in BioRender.com, adopted from [157].
Figure 8.
Drugs Targeting Telomeres and Telomerase. Figure by T. N. WAKAI, designed in BioRender.com, adopted from [157].
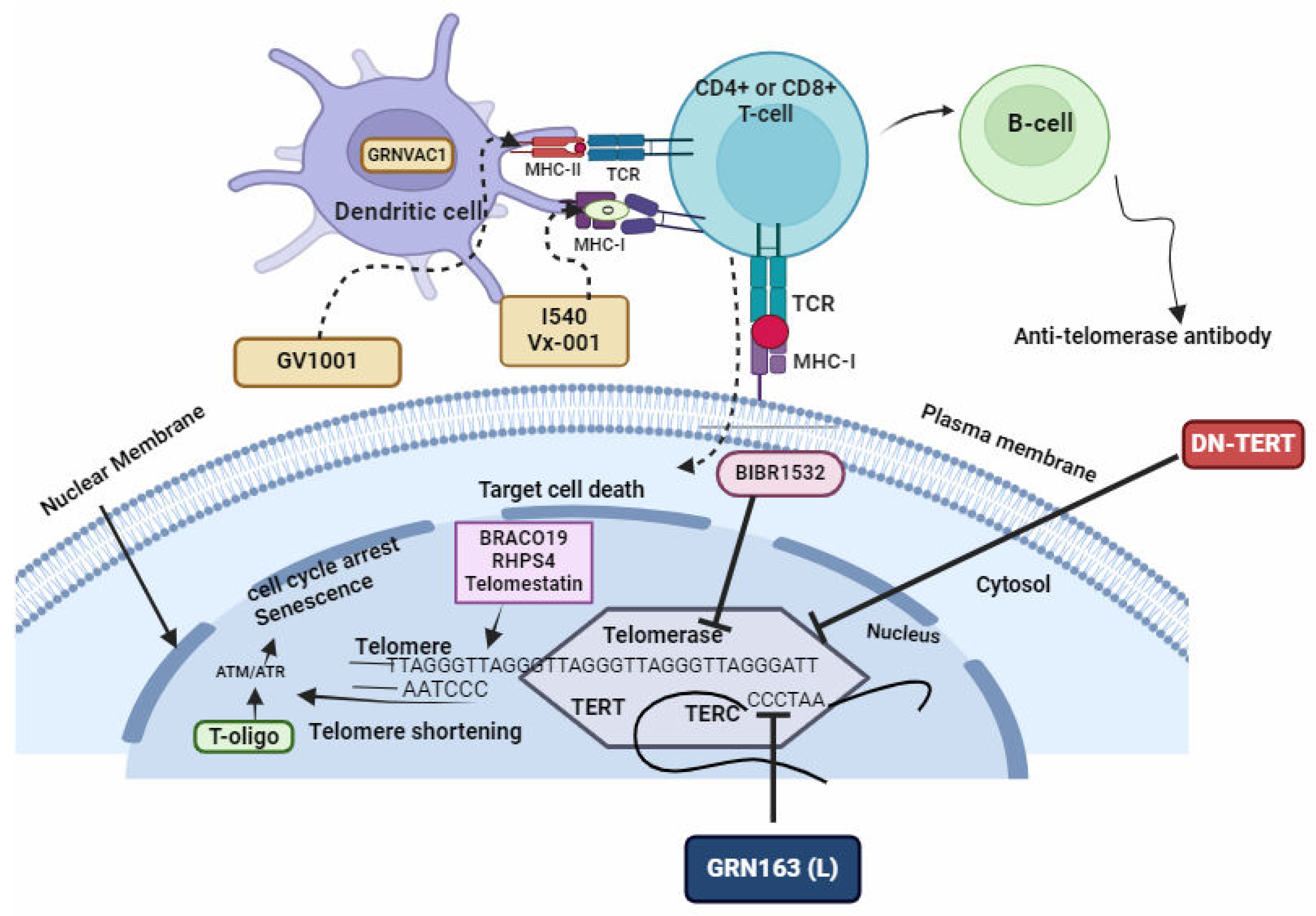
Several compounds have shown potential as telomere-targeting agents based on their ability to interact with telomeric DNA or disrupt telomere function [19,161]. However, drugs specifically designed to target telomeres and telomerase in malarial parasites have shown promising results [106]. However, they are still in the early stages of research, with no clinical drugs currently available [142,144,162,163].
Scientists have developed approaches aimed at developing drug targets against telomeres in malaria parasites (Table 2). These approaches can be classified into three: Firstly, the use of telomestatin analogs and G-quadruplex ligands: G-quadruplex ligands and analogs bind to G-quadruplex structures within the parasite's telomeres, stabilizing them and inhibiting telomerase function [14,115,151] (Figure 7). Secondly, the disruption of telomere maintenance system by synthesising compounds that interefere with telomere-associated proteins causing genomic instability and parasite cell death [92]. and thirdly, by synthesising compounds that induce telomere dysfunction through oxidative stress mechanisms [50,164]
Conclusion and Future Perspectives
Over the years, malaria control measures have aimed to reduce the global disease burden, but challenges persist, especially in sub-Saharan Africa, where effectiveness is hindered by drug-resistant parasites and insecticide-resistant mosquitoes. Identifying new drug targets and vaccines is crucial to enhance control efforts. Telomerase has garnered interest as a potential drug target for various diseases, including malaria. Recent studies have explored telomeres and telomerase in Plasmodium, revealing the parasite's unique mechanism for chromosome end maintenance. Plasmodium parasites exhibit high telomerase activity, making them vulnerable to inhibition, which disrupts telomere maintenance, leading to cell death. Despite possessing the genetic machinery for telomere synthesis, Plasmodium cannot elongate telomeres due to absent telomerase in asexually replicating stages in the human host. Malaria infection has been demonstrated to accelerates telomere shortening in humans, avian and rodent population. This shortening correlate with disease severity and treatment outcomes, including cellular senescence and accelerated cellular aging in malaria patients. Telomere length measurement and telomerase enzyme activity quantification may serve as an infection diagnostic and prognostic biomarker. Understanding how malaria induces telomere shortening and cellular senescence could offer insights for effective intervention. While telomerase-based therapies are in development, Plasmodium's unique telomerase system presents a promising avenue for future antimalarial drug discovery.
Author Contributions
TNW: Conceptualization, Scholarship acquisition, Visualization, Writing – original draft, Writing – review and editing. DOA: Writing – original draft, Writing – review and editing. ISA: Conceptualization, Supervision, Writing – review and editing.
Funding
The authors would like to acknowledge Covenant University Centre for Research Innovation and Development (CUCRID) for funding the Article Processing of this publication. This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Andrade, M.V.; Noronha, K.; Diniz, B.P.C.; et al. The economic burden of malaria: a systematic review. Malaria Journal 2022, 21, 283. [Google Scholar] [CrossRef] [PubMed]
- Gibson, L. World Malaria Report 2023: Key findings from the report. Target Malaria. Published , 2023. Accessed December 16, 2023. https://targetmalaria.org/latest/news/world-malaria-report-2023-key-findings-from-the-report/. 4 December.
- Gilbert, M.; Pullano, G.; Pinotti, F.; et al. Preparedness and vulnerability of African countries against importations of COVID-19: a modelling study. Lancet 2020, 395, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Nkengasong, J.N.; Mankoula, W. Looming threat of COVID-19 infection in Africa: act collectively, and fast. Lancet 2020, 395, 841–842. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.J.; Bertozzi-Villa, A.; Rumisha, S.F.; et al. Indirect effects of the COVID-19 pandemic on malaria intervention coverage, morbidity, and mortality in Africa: a geospatial modelling analysis. Lancet Infect Dis 2021, 21, 59–69. [Google Scholar] [CrossRef] [PubMed]
- GBD 2016 Healthcare Access and Quality Collaborators. Measuring performance on the Healthcare Access and Quality Index for 195 countries and territories and selected subnational locations: a systematic analysis from the Global Burden of Disease Study 2016. Lancet 2018, 391, 2236–2271. [Google Scholar] [CrossRef] [PubMed]
- Cotter, C.; Sturrock, H.J.W.; Hsiang, M.S.; et al. The changing epidemiology of malaria elimination: new strategies for new challenges. Lancet 2013, 382, 900–911. [Google Scholar] [CrossRef]
- Oladipo, H.J.; Tajudeen, Y.A.; Oladunjoye, I.O.; et al. Increasing challenges of malaria control in sub-Saharan Africa: Priorities for public health research and policymakers. Annals of Medicine and Surgery, 1043. [Google Scholar] [CrossRef]
- Siao, M.C.; Borner, J.; Perkins, S.L.; Deitsch, K.W.; Kirkman, L.A. Evolution of Host Specificity by Malaria Parasites through Altered Mechanisms Controlling Genome Maintenance. mBio 2020, 11, e03272–19. [Google Scholar] [CrossRef] [PubMed]
- Su, X.Z.; Lane, K.D.; Xia, L.; Sá, J.M.; Wellems, T.E. Plasmodium Genomics and Genetics: New Insights into Malaria Pathogenesis, Drug Resistance, Epidemiology, and Evolution. Clin Microbiol Rev 2019, 32, e00019–19. [Google Scholar] [CrossRef] [PubMed]
- Asghar, M.; Hasselquist, D.; Hansson, B.; Zehtindjiev, P.; Westerdahl, H.; Bensch, S. Hidden costs of infection: Chronic malaria accelerates telomere degradation and senescence in wild birds. Science 2015, 347, 436–438. [Google Scholar] [CrossRef] [PubMed]
- Asghar, M.; Palinauskas, V.; Zaghdoudi-Allan, N.; et al. Parallel telomere shortening in multiple body tissues owing to malaria infection. Proceedings of the Royal Society B: Biological Sciences 2016, 283, 20161184. [Google Scholar] [CrossRef]
- Calhoun, S.F.; Reed, J.; Alexander, N.; Mason, C.E.; Deitsch, K.W.; Kirkman, L.A. Chromosome End Repair and Genome Stability in Plasmodium falciparum. mBio 2017, 8, 10.1128/mbio.00547–17. [Google Scholar] [CrossRef]
- De Cian, A.; Grellier, P.; Mouray, E.; et al. Plasmodium telomeric sequences: structure, stability and quadruplex targeting by small compounds. Chembiochem 2008, 9, 2730–2739. [Google Scholar] [CrossRef]
- Abraham, R.S. Tell me more – telomere biology and understanding telomere defects. Pathology. [CrossRef]
- Lu, W.; Zhang, Y.; Liu, D.; Songyang, Z.; Wan, M. Telomeres - Structure, Function, and Regulation. Experimental cell research 2013, 319, 133. [Google Scholar] [CrossRef] [PubMed]
- Button, L.; Rogers, B.; Thomas, E.; et al. Telomere and Telomerase-Associated Proteins in Endometrial Carcinogenesis and Cancer-Associated Survival. International Journal of Molecular Sciences 2022, 23, 626. [Google Scholar] [CrossRef] [PubMed]
- Greider, C.W.; Blackburn, E.H. The telomere terminal transferase of Tetrahymena is a ribonucleoprotein enzyme with two kinds of primer specificity. Cell 1987, 51, 887–898. [Google Scholar] [CrossRef]
- Li, B. Telomere components as potential therapeutic targets for treating microbial pathogen infections. Frontiers in Oncology, /: Accessed , 2023. https, 2 March 2023. [Google Scholar]
- Bonnell, E.; Pasquier, E.; Wellinger, R.J. Telomere Replication: Solving Multiple End Replication Problems. Front Cell Dev Biol, 6681. [Google Scholar] [CrossRef]
- Levy, M.Z.; Allsopp, R.C.; Futcher, A.B.; Greider, C.W.; Harley, C.B. Telomere end-replication problem and cell aging. Journal of Molecular Biology 1992, 225, 951–960. [Google Scholar] [CrossRef]
- Maestroni, L.; Matmati, S.; Coulon, S. Solving the Telomere Replication Problem. Genes 2017, 8, 55. [Google Scholar] [CrossRef]
- Chakravarti, D.; LaBella, K.A.; DePinho, R.A. Telomeres: history, health, and hallmarks of aging. Cell 2021, 184, 306–322. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S. Telomeres in health and disease. J Oral Maxillofac Pathol 2017, 21, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Effros, R.B. Impact of the Hayflick Limit on T cell responses to infection: lessons from aging and HIV disease. Mech Ageing Dev 2004, 125, 103–106. [Google Scholar] [CrossRef]
- Lulkiewicz, M.; Bajsert, J.; Kopczynski, P.; Barczak, W.; Rubis, B. Telomere length: how the length makes a difference. Mol Biol Rep 2020, 47, 7181–7188. [Google Scholar] [CrossRef] [PubMed]
- Armanios, M. The Role of Telomeres in Human Disease. Annu Rev Genomics Hum Genet. [CrossRef]
- GOMEZ DE, ARMANDO RG, FARINA HG, et al. Telomere structure and telomerase in health and disease. Int J Oncol 2012, 41, 1561–1569. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, G.E.; Fountain, A.J.; van Roon, A.M.M.; et al. Structure of human telomerase holoenzyme with bound telomeric DNA. Nature 2021, 593, 449–453. [Google Scholar] [CrossRef]
- Lee, J.; Pellegrini, M.V. Biochemistry, Telomere And Telomerase. In: StatPearls. StatPearls Publishing; 2023. Accessed , 2023. http://www.ncbi.nlm.nih. 25 May 5764. [Google Scholar]
- Panasiak, L.; Kuciński, M.; Hliwa, P.; Pomianowski, K.; Ocalewicz, K. Telomerase Activity in Somatic Tissues and Ovaries of Diploid and Triploid Rainbow Trout (Oncorhynchus mykiss) Females. Cells (2073-4409) 2023, 12, 1772. [Google Scholar] [CrossRef] [PubMed]
- Akincilar, S.C.; Unal, B.; Tergaonkar, V. Reactivation of telomerase in cancer. Cell Mol Life Sci 2016, 73, 1659–1670. [Google Scholar] [CrossRef] [PubMed]
- Guterres, A.N.; Villanueva, J. Targeting telomerase for cancer therapy. Oncogene 2020, 39, 5811–5824. [Google Scholar] [CrossRef] [PubMed]
- Li, B. Telomere components as potential therapeutic targets for treating microbial pathogen infections. Frontiers in Oncology, /: Accessed , 2023. https, 2 March 2023. [Google Scholar]
- Roake, C.M.; Artandi, S.E. Regulation of human telomerase in homeostasis and disease. Nat Rev Mol Cell Biol 2020, 21, 384–397. [Google Scholar] [CrossRef] [PubMed]
- Bottius, E.; Bakhsis, N.; Scherf, A. Plasmodium falciparum Telomerase: De Novo Telomere Addition to Telomeric and Nontelomeric Sequences and Role in Chromosome Healing. Mol Cell Biol 1998, 18, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, L.; Scherf, A. Plasmodium telomeres and telomerase: the usual actors in an unusual scenario. Chromosome Res 2005, 13, 517–524. [Google Scholar] [CrossRef]
- Rubiano, C.C.; Wasserman, M. Short communication - detection of telomerase activity in Plasmodium falciparum using a nonradioactive method. Published online , 2003. Accessed February 22, 2023. https://tspace.library.utoronto. 31 December 1807. [Google Scholar]
- Sekhri, K. Telomeres and telomerase: understanding basic structure and potential new therapeutic strategies targeting it in the treatment of cancer. J Postgrad Med 2014, 60, 303–308. [Google Scholar] [CrossRef]
- Мoрoзoва ЛФ, Турбабина НА, Степанoва ЕВ, et al. Method of treating malaria using a therapeutic combination of telomerase inhibitors (iimatinib mesilate) and artemether. Published online , 2020. Accessed March 17, 2024. https://patents.google. 5 June 2722.
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; et al. Specific Association of Human Telomerase Activity with Immortal Cells and Cancer. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Religa, A.A.; Ramesar, J.; Janse, C.J.; Scherf, A.; Waters, A.P. P. berghei telomerase subunit TERT is essential for parasite survival. PLoS One 2014, 9, e108930. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.A.; Chakrabarti, K. Telomerase ribonucleoprotein and genome integrity—An emerging connection in protozoan parasites. WIREs RNA 2022, 13, e1710. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.; Chakrabarti, K. Current Perspectives of Telomerase Structure and Function in Eukaryotes with Emerging Views on Telomerase in Human Parasites. IJMS 2018, 19, 333. [Google Scholar] [CrossRef] [PubMed]
- Calvo, E.P.; Wasserman, M. G-Quadruplex ligands: Potent inhibitors of telomerase activity and cell proliferation in Plasmodium falciparum. Mol Biochem Parasitol 2016, 207, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Kurzhals, R.L.; Fanti, L.; Ebsen, A.C.G.; Rong, Y.S.; Pimpinelli, S.; Golic, K.G. Chromosome Healing Is Promoted by the Telomere Cap Component Hiphop in Drosophila. Genetics 2017, 207, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H. Telomeres and telomerase: their mechanisms of action and the effects of altering their functions. FEBS Lett 2005, 579, 859–862. [Google Scholar] [CrossRef] [PubMed]
- Benites-Zapata, V.A.; Ulloque-Badaracco, J.R.; Alarcón-Braga, E.A.; Fernández-Alonso, A.M.; López-Baena, M.T.; Pérez-López, F.R. Telomerase activity and telomere length in women with breast cancer or without malignancy: A systematic review and meta-analysis. Maturitas, 1078. [Google Scholar] [CrossRef]
- Lin, J.; Epel, E. Stress and telomere shortening: Insights from cellular mechanisms. Ageing Res Rev, 1015. [Google Scholar] [CrossRef]
- Barnes, R.P.; Fouquerel, E.; Opresko, P.L. The impact of oxidative DNA damage and stress on telomere homeostasis. Mech Ageing Dev. [CrossRef]
- Armstrong, E.; Boonekamp, J. Does oxidative stress shorten telomeres in vivo? A meta-analysis. Ageing Res Rev, 1018. [Google Scholar] [CrossRef]
- Niveta, J.P.S.; Kumar, M.A.; Parvathi, V.D. Telomere attrition and inflammation: the chicken and the egg story. Egyptian Journal of Medical Human Genetics 2022, 23, 131. [Google Scholar] [CrossRef]
- Colella, M.P.; Santana, B.A.; Conran, N.; et al. Telomere length correlates with disease severity and inflammation in sickle cell disease. Rev Bras Hematol Hemoter 2017, 39, 140–145. [Google Scholar] [CrossRef]
- Ahmed, W.; Lingner, J. Impact of oxidative stress on telomere biology. Differentiation. [CrossRef]
- Andreu-Sánchez, S.; Aubert, G.; Ripoll-Cladellas, A.; et al. Genetic, parental and lifestyle factors influence telomere length. Commun Biol 2022, 5, 565. [Google Scholar] [CrossRef]
- Lin, J.; Blackburn, E. Telomeres and lifestyle factors: Roles in cellular aging. Mutation research. [CrossRef]
- Chen, W.; Shi, S.; Jiang, Y.; et al. Association Between Riboflavin Intake and Telomere Length: A Cross-Sectional Study From National Health and Nutrition Examination Survey 1999–2002. Frontiers in Nutrition, /: Accessed , 2023. https, 3 December 2023. [Google Scholar]
- Tarry-Adkins, J.L.; Fernandez-Twinn, D.S.; Chen, J.H.; et al. Poor maternal nutrition and accelerated postnatal growth induces an accelerated aging phenotype and oxidative stress in skeletal muscle of male rats. Dis Model Mech 2016, 9, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Ilmonen, P.; Kotrschal, A.; Penn, D.J. Telomere Attrition Due to Infection. PLoS One 2008, 3, e2143. [Google Scholar] [CrossRef] [PubMed]
- Giraudeau, M.; Heidinger, B.; Bonneaud, C.; Sepp, T. Telomere shortening as a mechanism of long-term cost of infectious diseases in natural animal populations. Biol Lett 2019, 15, 20190190. [Google Scholar] [CrossRef] [PubMed]
- Albosale, A.H.; Mashkina, E.V. Association of Relative Telomere Length and Risk of High Human Papillomavirus Load in Cervical Epithelial Cells. Balkan J Med Genet 2021, 24, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.S.; Nguyen, M.T.N.; Pham, T.X.; et al. Hepatitis C Virus Nonstructural 5A Protein Interacts with Telomere Length Regulation Protein: Implications for Telomere Shortening in Patients Infected with HCV. Mol Cells 2022, 45, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E.; Samper, E.; Martín-Caballero, J.; Flores, J.M.; Lee, H.; Blasco, M.A. Disease states associated with telomerase deficiency appear earlier in mice with short telomeres. The EMBO Journal 1999, 18, 2950–2960. [Google Scholar] [CrossRef] [PubMed]
- Matthe, D.M.; Thoma, O.M.; Sperka, T.; Neurath, M.F.; Waldner, M.J. Telomerase deficiency reflects age-associated changes in CD4+ T cells. Immunity & Ageing 2022, 19, 16. [Google Scholar] [CrossRef]
- Patrick, M.; Weng, N.P. Expression and regulation of telomerase in human T cell differentiation, activation, aging and diseases. Cell Immunol, 1039. [Google Scholar] [CrossRef]
- Valenzuela, H.F.; Effros, R.B. Divergent telomerase and CD28 expression patterns in human CD4 and CD8 T cells following repeated encounters with the same antigenic stimulus. Clin Immunol 2002, 105, 117–125. [Google Scholar] [CrossRef]
- Dalzini, A.; Ballin, G.; Dominguez-Rodriguez, S.; et al. Size of HIV-1 reservoir is associated with telomere shortening and immunosenescence in early-treated European children with perinatally acquired HIV-1. J Int AIDS Soc 2021, 24, e25847. [Google Scholar] [CrossRef]
- Ilmonen, P.; Kotrschal, A.; Penn, D.J. Telomere Attrition Due to Infection. PLOS ONE 2008, 3, e2143. [Google Scholar] [CrossRef]
- Akbar, A.N.; Beverley, P.C.L.; Salmon, M. Will telomere erosion lead to a loss of T-cell memory? Nat Rev Immunol 2004, 4, 737–743. [Google Scholar] [CrossRef]
- Pawelec, G.; Akbar, A.; Caruso, C.; Solana, R.; Grubeck-Loebenstein, B.; Wikby, A. Human immunosenescence: is it infectious? Immunological Reviews 2005, 205, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Gray-Miceli, D.; Gray, K.; Sorenson, M.R.; Holtzclaw, B.J. Immunosenescence and Infectious Disease Risk Among Aging Adults. Advances in Family Practice Nursing 2023, 5, 27–40. [Google Scholar] [CrossRef]
- Asghar, M.; Yman, V.; Homann, M.V.; et al. Cellular aging dynamics after acute malaria infection: A 12-month longitudinal study. Aging Cell 2018, 17, e12702. [Google Scholar] [CrossRef]
- Miglar, A. The Effect of Plasmodium Infection on Cellular Aging in Humans. Inst för medicin, Solna / Dept of Medicine, Solna; 2023. Accessed , 2023. http://openarchive.ki. 10 April 1061. [Google Scholar]
- Miglar, A.; Reuling, I.J.; Yap, X.Z.; Färnert, A.; Sauerwein, R.W.; Asghar, M. Biomarkers of cellular aging during a controlled human malaria infection. Sci Rep 2021, 11, 18733. [Google Scholar] [CrossRef] [PubMed]
- Durso, D.F.; Silveira-Nunes, G.; Coelho, M.M.; et al. Living in endemic area for infectious diseases accelerates epigenetic age. Mech Ageing Dev, 1117. [Google Scholar] [CrossRef]
- Stanojcic, S.; Kuk, N.; Ullah, I.; Sterkers, Y.; Merrick, C.J. Single-molecule analysis reveals that DNA replication dynamics vary across the course of schizogony in the malaria parasite Plasmodium falciparum. Sci Rep 2017, 7, 4003. [Google Scholar] [CrossRef]
- Raj, D.K.; Das, B.R.; Dash, A.P.; Supakar, P.C. Identification of telomerase activity in gametocytes of Plasmodium falciparum. Biochemical and Biophysical Research Communications 2003, 309, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Kołodziej-Sobocińska, M. Factors affecting the spread of parasites in populations of wild European terrestrial mammals. Mamm Res 2019, 64, 301–318. [Google Scholar] [CrossRef]
- Edwards-Smallbone, J.; Jensen, A.L.; Roberts, L.E.; Totañes, F.I.G.; Hart, S.R.; Merrick, C.J. Plasmodium falciparum GBP2 Is a Telomere-Associated Protein That Binds to G-Quadruplex DNA and RNA. Front Cell Infect Microbiol, 2022; 12, 782537. [Google Scholar] [CrossRef]
- Gardner, M.J.; Hall, N.; Fung, E.; et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 2002, 419, 498–511. [Google Scholar] [CrossRef] [PubMed]
- Kwapisz, M.; Morillon, A. Subtelomeric Transcription and its Regulation. Journal of Molecular Biology 2020, 432, 4199–4219. [Google Scholar] [CrossRef]
- Scherf, A.; Figueiredo, L.M.; Freitas-Junior, L.H. Plasmodium telomeres: a pathogen’s perspective. Curr Opin Microbiol 2001, 4, 409–414. [Google Scholar] [CrossRef]
- del Portillo, H.A.; Fernandez-Becerra, C.; Bowman, S.; et al. A superfamily of variant genes encoded in the subtelomeric region of Plasmodium vivax. Nature 2001, 410, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Araujo, R.B.D.; Silva, T.M.; Kaiser, C.S.; et al. Independent regulation of Plasmodium falciparum rif gene promoters. Scientific Reports 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Duraisingh, M.T.; Voss, T.S.; Marty, A.J.; et al. Heterochromatin Silencing and Locus Repositioning Linked to Regulation of Virulence Genes in Plasmodium falciparum. Cell 2005, 121, 13–24. [Google Scholar] [CrossRef]
- Wang, W.F.; Zhang, Y.L. PfSWIB, a potential chromatin regulator for var gene regulation and parasite development in Plasmodium falciparum. Parasites & Vectors 2020, 13, 48. [Google Scholar] [CrossRef]
- Asghar, M.; Palinauskas, V.; Zaghdoudi-Allan, N.; et al. Parallel telomere shortening in multiple body tissues owing to malaria infection. Proc R Soc B 2016, 283, 20161184. [Google Scholar] [CrossRef]
- Reed, J.; Kirkman, L.A.; Kafsack, B.F.; Mason, C.E.; Deitsch, K.W. Telomere length dynamics in response to DNA damage in malaria parasites. iScience 2021, 24, 102082. [Google Scholar] [CrossRef] [PubMed]
- Asghar, M.; Palinauskas, V.; Zaghdoudi-Allan, N.; et al. Parallel telomere shortening in multiple body tissues owing to malaria infection. Proceedings of the Royal Society B: Biological Sciences 2016, 283, 20161184. [Google Scholar] [CrossRef]
- Mohanty, P.; Gupta, A.; Bhatnagar, S. Modeling of Plasmodium falciparum Telomerase Reverse Transcriptase Ternary Complex: Repurposing of Nucleoside Analog Inhibitors. Assay Drug Dev Technol 2015, 13, 628–637. [Google Scholar] [CrossRef]
- Wang, Y.; Feigon, J. Structural biology of telomerase and its interaction at telomeres. Curr Opin Struct Biol. [CrossRef]
- Edwards-Smallbone J, Jensen AL, Roberts LE, Totañes FIG, Hart SR, Merrick CJ. Plasmodium falciparum GBP2 Is a Telomere-Associated Protein That Binds to G-Quadruplex DNA and RNA. Frontiers in Cellular and Infection Microbiology. 2022;12. Accessed May 19, 2023. https://www.frontiersin.org/articles/10.3389/fcimb.2022.782537.
- Al-Turki, T.M.; Griffith, J.D. Mammalian telomeric RNA (TERRA) can be translated to produce valine–arginine and glycine–leucine dipeptide repeat proteins. Proceedings of the National Academy of Sciences 2023, 120, e2221529120. [Google Scholar] [CrossRef]
- Vaiserman, A.; Krasnienkov, D. Telomere Length as a Marker of Biological Age: State-of-the-Art, Open Issues, and Future Perspectives. Front Genet, 6301. [Google Scholar] [CrossRef]
- Shay, J.W. Role of Telomeres and Telomerase in Aging and Cancer. Cancer Discov 2016, 6, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Svenson, U.; Nordfjäll, K.; Baird, D.; et al. Blood Cell Telomere Length Is a Dynamic Feature. PLOS ONE 2011, 6, e21485. [Google Scholar] [CrossRef] [PubMed]
- Minami, R.; Takahama, S.; Yamamoto, M. Correlates of telomere length shortening in peripheral leukocytes of HIV-infected individuals and association with leukoaraiosis. PLoS One 2019, 14, e0218996. [Google Scholar] [CrossRef] [PubMed]
- Hamad, R.; Walter, S.; Rehkopf, D.H. Telomere length and health outcomes: A two-sample genetic instrumental variables analysis. Experimental Gerontology. [CrossRef]
- Helby, J.; Nordestgaard, B.G.; Benfield, T.; Bojesen, S.E. Shorter leukocyte telomere length is associated with higher risk of infections: a prospective study of 75,309 individuals from the general population. Haematologica 2017, 102, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.V.; Schneider, K.M.; Teumer, A.; et al. Association of Telomere Length With Risk of Disease and Mortality. JAMA Internal Medicine 2022, 182, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Tambo, M.; Mwinga, M.; Mumbengegwi, D.R. Loop-mediated isothermal amplification (LAMP) and Polymerase Chain Reaction (PCR) as quality assurance tools for Rapid Diagnostic Test (RDT) malaria diagnosis in Northern Namibia. PLoS One 2018, 13, e0206848. [Google Scholar] [CrossRef] [PubMed]
- Fali, T.; K’Ros, C.; Appay, V.; Sauce, D. Assessing T Lymphocyte Aging Using Telomere Length and Telomerase Activity Measurements in Low Cell Numbers. Methods Mol Biol. [CrossRef]
- Udroiu, I.; Marinaccio, J.; Sgura, A. Many Functions of Telomerase Components: Certainties, Doubts, and Inconsistencies. International Journal of Molecular Sciences 2022, 23, 15189. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.; Collins, K. Human Telomerase Activation Requires Two Independent Interactions between Telomerase RNA and Telomerase Reverse Transcriptase. Molecular cell. [CrossRef]
- Wyatt, H.D.M.; West, S.C.; Beattie, T.L. InTERTpreting telomerase structure and function. Nucleic Acids Res 2010, 38, 5609–5622. [Google Scholar] [CrossRef] [PubMed]
- Li, B. Keeping Balance Between Genetic Stability and Plasticity at the Telomere and Subtelomere of Trypanosoma brucei. Front Cell Dev Biol, 6996. [Google Scholar] [CrossRef]
- Shay, J.W.; Wright, W.E. Telomeres and telomerase: three decades of progress. Nat Rev Genet 2019, 20, 299–309. [Google Scholar] [CrossRef]
- Cong, Y.S.; Wright, W.E.; Shay, J.W. Human Telomerase and Its Regulation. Microbiol Mol Biol Rev 2002, 66, 407–425. [Google Scholar] [CrossRef]
- Simantov, K.; Goyal, M.; Dzikowski, R. Emerging biology of noncoding RNAs in malaria parasites. PLOS Pathogens 2022, 18, e1010600. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, L.; Scherf, A. Plasmodium telomeres and telomerase: the usual actors in an unusual scenario. Chromosome Res 2005, 13, 517–524. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, B.C.D.; Shiburah, M.E.; Paiva, S.C.; et al. Possible Involvement of Hsp90 in the Regulation of Telomere Length and Telomerase Activity During the Leishmania amazonensis Developmental Cycle and Population Proliferation. Front Cell Dev Biol, 2021; 9, 713415. [Google Scholar] [CrossRef]
- Calado, R.; Young, N. Telomeres, Telomerase, and Human Disease. The Hematologist, 2010. [Google Scholar] [CrossRef]
- Calado, R.T.; Yewdell, W.T.; Wilkerson, K.L.; et al. Sex hormones, acting on the TERT gene, increase telomerase activity in human primary hematopoietic cells. Blood 2009, 114, 2236–2243. [Google Scholar] [CrossRef] [PubMed]
- Belachew, E.B. Immune Response and Evasion Mechanisms of Plasmodium falciparum Parasites. Journal of Immunology Research, 2018; 2018, e6529681. [Google Scholar] [CrossRef]
- Belmonte-Reche, E.; Martínez-García, M.; Guédin, A.; et al. G-Quadruplex Identification in the Genome of Protozoan Parasites Points to Naphthalene Diimide Ligands as New Antiparasitic Agents. J Med Chem 2018, 61, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Lujan, S.A.; Williams, J.S.; Kunkel, T.A. DNA polymerases divide the labor of genome replication. Trends Cell Biol 2016, 26, 640–654. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A.A.; Watson, C.M.; Noble, J.R.; Pickett, H.A.; Tam, P.P.L.; Reddel, R.R. Alternative lengthening of telomeres in normal mammalian somatic cells. Genes Dev 2013, 27, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Cooper, G.M. DNA Replication. In: The Cell: A Molecular Approach. 2nd Edition. Sinauer Associates; 2000. Accessed , 2024. https://www.ncbi.nlm.nih. 28 March 9940. [Google Scholar]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA damage, repair and mutagenesis. Environ Mol Mutagen 2017, 58, 235–263. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Pickett, H.A. Targeting telomeres: advances in telomere maintenance mechanism-specific cancer therapies. Nat Rev Cancer 2022, 22, 515–532. [Google Scholar] [CrossRef]
- Muraki, K.; Nyhan, K.; Han, L.; Murnane, J.P. Mechanisms of telomere loss and their consequences for chromosome instability. Front Oncol. [CrossRef]
- Piñeiro-Hermida, S.; Bosso, G.; Sánchez-Vázquez, R.; Martínez, P.; Blasco, M.A. Telomerase deficiency and dysfunctional telomeres in the lung tumor microenvironment impair tumor progression in NSCLC mouse models and patient-derived xenografts. Cell Death Differ, 1: online , 2023, 21 April 2023. [Google Scholar] [CrossRef]
- Gavia-García, G.; Rosado-Pérez, J.; Arista-Ugalde, T.L.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. Telomere Length and Oxidative Stress and Its Relation with Metabolic Syndrome Components in the Aging. Biology (Basel) 2021, 10, 253. [Google Scholar] [CrossRef]
- Lupatov AYu, Yarygin, K. N. Telomeres and Telomerase in the Control of Stem Cells. Biomedicines 2022, 10, 2335. [Google Scholar] [CrossRef]
- Rossiello, F.; Jurk, D.; Passos, J.F.; d’Adda di Fagagna, F. Telomere dysfunction in ageing and age-related diseases. Nat Cell Biol 2022, 24, 135–147. [Google Scholar] [CrossRef]
- Ali, J.H.; Walter, M. Combining old and new concepts in targeting telomerase for cancer therapy: transient, immediate, complete and combinatory attack (TICCA). Cancer Cell International 2023, 23, 197. [Google Scholar] [CrossRef]
- Phatak, P.; Burger, A.M. Telomerase and its potential for therapeutic intervention. Br J Pharmacol 2007, 152, 1003–1011. [Google Scholar] [CrossRef]
- Verdun, R.E.; Karlseder, J. Replication and protection of telomeres. Nature 2007, 447, 924–931. [Google Scholar] [CrossRef]
- Olawole, T.D.; Okundigie, M.I.; Rotimi, S.O.; Okwumabua, O.; Afolabi, I.S. Preadministration of Fermented Sorghum Diet Provides Protection against Hyperglycemia-Induced Oxidative Stress and Suppressed Glucose Utilization in Alloxan-Induced Diabetic Rats. Front Nutr 2018, 5. [Google Scholar] [CrossRef]
- Osikoya, I.O.; Afolabi, I.S.; Rotimi, S.O.; Okafor, A.M.J. Anti-hemolytic and anti-inflammatory activities of the methanolic extract of Solenostemon Monostachyus (P.Beauv.) Briq. leaves in 2-butoxyethanol-hemolytic induced rats 2018, 1954:040015. [CrossRef]
- Afolabi, I.S.; Olawole, T.D.; Adams, K.A.; Shopeju, O.A.; Ezeaku, M.C. Anti-inflammatory effects and the molecular pattern of the therapeutic effects of dietary seeds of Adenanthera Pavonina in albino rats. AIP Conference Proceedings 2018, 1954, 040016. [Google Scholar] [CrossRef]
- Alessandrini, I.; Percio, S.; Naghshineh, E.; et al. Telomere as a Therapeutic Target in Dedifferentiated Liposarcoma. Cancers (Basel) 2022, 14, 2624. [Google Scholar] [CrossRef]
- De Vitis, M.; Berardinelli, F.; Sgura, A. Telomere Length Maintenance in Cancer: At the Crossroad between Telomerase and Alternative Lengthening of Telomeres (ALT). Int J Mol Sci 2018, 19, 606. [Google Scholar] [CrossRef]
- Dey, A.; Chakrabarti, K. Current Perspectives of Telomerase Structure and Function in Eukaryotes with Emerging Views on Telomerase in Human Parasites. Int J Mol Sci 2018, 19, 333. [Google Scholar] [CrossRef]
- Sinha, S.; Medhi, B.; Sehgal, R. Challenges of drug-resistant malaria. Parasite. [CrossRef]
- Tripathi, H.; Bhalerao, P.; Singh, S.; et al. Malaria therapeutics: are we close enough? Parasites & Vectors 2023, 16, 130. [Google Scholar] [CrossRef]
- Robinson, N.J.; Schiemann, W.P. Telomerase in Cancer: Function, Regulation, and Clinical Translation. Cancers (Basel) 2022, 14, 808. [Google Scholar] [CrossRef]
- Fathi, E.; Charoudeh, H.N.; Sanaat, Z.; Farahzadi, R. Telomere shortening as a hallmark of stem cell senescence. Stem Cell Investigation. [CrossRef]
- Zaug, A.J.; Crary, S.M.; Jesse Fioravanti, M.; Campbell, K.; Cech, T.R. Many disease-associated variants of hTERT retain high telomerase enzymatic activity. Nucleic Acids Res 2013, 41, 8969–8978. [Google Scholar] [CrossRef]
- Raj, D.K.; Das, B.R.; Dash, A.P.; Supakar, P.C. Identification of telomerase activity in gametocytes of Plasmodium falciparum. Biochem Biophys Res Commun 2003, 309, 685–688. [Google Scholar] [CrossRef]
- Bertschi, N.L. Novel insights into telomere biology and virulence gene expression in plasmodium falciparum. Published online 2015. [CrossRef]
- Mender, I.; Gryaznov, S.; Shay, J.W. A novel telomerase substrate precursor rapidly induces telomere dysfunction in telomerase positive cancer cells but not telomerase silent normal cells. Oncoscience 2015, 2, 693–695. [Google Scholar] [CrossRef]
- Bär, C.; Blasco, M.A. Telomeres and telomerase as therapeutic targets to prevent and treat age-related diseases. F1000Res, 2016. [Google Scholar] [CrossRef]
- Morozova, L.; Kondrashin, A.V.; Stepanova, E.V.; et al. In vivo effectiveness of the inhibitors of telomerase against malaria parasites. Infekcionnye bolezni, 2020. [Google Scholar] [CrossRef]
- Lee, M.C.S.; Lindner, S.E.; Lopez-Rubio, J.J.; Llinás, M. Cutting back malaria: CRISPR/Cas9 genome editing of Plasmodium. Brief Funct Genomics 2019, 18, 281–289. [Google Scholar] [CrossRef]
- Kudyba, H.M.; Cobb, D.W.; Florentin, A.; Krakowiak, M.; Muralidharan, V. CRISPR/Cas9 Gene Editing to Make Conditional Mutants of Human Malaria Parasite, P. falciparum. J Vis Exp, 5774. [Google Scholar] [CrossRef]
- Bryant, J.M.; Baumgarten, S.; Glover, L.; Hutchinson, S.; Rachidi, N. CRISPR in Parasitology: Not Exactly Cut and Dried! Trends in Parasitology 2019, 35, 409–422. [Google Scholar] [CrossRef]
- Lee, H.W. ; Roh J il, Sung, Y.H. Clinical implications of antitelomeric drugs with respect to the nontelomeric functions of telomerase in cancer. OTT, 1: online 13, 20 August 1161. [Google Scholar] [CrossRef]
- Lavanya, C.; Venkataswamy, M.M.; Sibin, M.K.; Srinivas Bharath, M.M.; Chetan, G.K. Down regulation of human telomerase reverse transcriptase (hTERT) expression by BIBR1532 in human glioblastoma LN18 cells. Cytotechnology 2018, 70, 1143–1154. [Google Scholar] [CrossRef]
- Altamura, G.; Degli Uberti, B.; Galiero, G.; et al. The Small Molecule BIBR1532 Exerts Potential Anti-cancer Activities in Preclinical Models of Feline Oral Squamous Cell Carcinoma Through Inhibition of Telomerase Activity and Down-Regulation of TERT. Front Vet Sci, 6207. [Google Scholar] [CrossRef]
- Sullivan, H.J.; Readmond, C.; Radicella, C.; Persad, V.; Fasano, T.J.; Wu, C. Binding of Telomestatin, TMPyP4, BSU6037, and BRACO19 to a Telomeric G-Quadruplex–Duplex Hybrid Probed by All-Atom Molecular Dynamics Simulations with Explicit Solvent. ACS Omega 2018, 3, 14788–14806. [Google Scholar] [CrossRef]
- Lagah, S.; Tan, I.L.; Radhakrishnan, P.; et al. RHPS4 G-Quadruplex Ligand Induces Anti-Proliferative Effects in Brain Tumor Cells. PLoS One 2014, 9, e86187. [Google Scholar] [CrossRef]
- Neidle, S. 5 - Telomeric Quadruplex Ligands II: Polycyclic and Non-fused Ring Compounds. In: Neidle, S.; ed. Therapeutic Applications of Quadruplex Nucleic Acids, 9: Press; 2012, 2012. [Google Scholar] [CrossRef]
- Machireddy, B.; Sullivan, H.J.; Wu, C. Binding of BRACO19 to a Telomeric G-Quadruplex DNA Probed by All-Atom Molecular Dynamics Simulations with Explicit Solvent. Molecules 2019, 24, 1010. [Google Scholar] [CrossRef]
- Maréchal, A.; Zou, L. DNA Damage Sensing by the ATM and ATR Kinases. Cold Spring Harb Perspect Biol 2013, 5, a012716. [Google Scholar] [CrossRef]
- Davé, A.; Pai, C.C.; Durley, S.C.; et al. Homologous recombination repair intermediates promote efficient de novo telomere addition at DNA double-strand breaks. Nucleic Acids Res 2020, 48, 1271–1284. [Google Scholar] [CrossRef]
- 157. Roh J il, Sung, Y.H.; Lee, H.W. Clinical implications of antitelomeric drugs with respect to the nontelomeric functions of telomerase in cancer. Onco Targets Ther. [CrossRef]
- Huang, R.X.; Zhou, P.K. DNA damage response signaling pathways and targets for radiotherapy sensitization in cancer. Sig Transduct Target Ther 2020, 5, 1–27. [Google Scholar] [CrossRef]
- Negrini, S.; De Palma, R.; Filaci, G. Anti-cancer Immunotherapies Targeting Telomerase. Cancers (Basel) 2020, 12, 2260. [Google Scholar] [CrossRef]
- Qiu, J.; Li, G.W.; Sui, Y.F.; et al. Immunization with truncated sequence of Telomerase Reverse Transcriptase induces a specific antitumor response in vivo. Acta Oncologica 2007, 46, 961–968. [Google Scholar] [CrossRef]
- Carvalho, V.S.; Gomes, W.R.; Calado, R.T. Recent advances in understanding telomere diseases. Fac Rev. [CrossRef]
- Nogueira, B.M.D.; da Costa Pantoja, L.; da Silva, E.L.; et al. Telomerase (hTERT) Overexpression Reveals a Promising Prognostic Biomarker and Therapeutical Target in Different Clinical Subtypes of Pediatric Acute Lymphoblastic Leukaemia. Genes (Basel) 2021, 12, 1632. [Google Scholar] [CrossRef]
- Anas, M.; Sharma, R.; Dhamodharan, V.; et al. Investigating Pharmacological Targeting of G-Quadruplexes in the Human Malaria Parasite. Biochemistry 2017, 56, 6691–6699. [Google Scholar] [CrossRef]
- Fouquerel, E.; Lormand, J.; Bose, A.; et al. Oxidative guanine base damage regulates human telomerase activity. Nat Struct Mol Biol 2016, 23, 1092–1100. [Google Scholar] [CrossRef]
- Guillon, J.; Cohen, A.; Gueddouda, N.M.; et al. Design, synthesis and antimalarial activity of novel bis{N-[(pyrrolo[1,2-a]quinoxalin-4-yl)benzyl]-3-aminopropyl}amine derivatives. J Enzyme Inhib Med Chem 2017, 32, 547–563. [Google Scholar] [CrossRef]
- Chien, H.D.; Pantaleo, A.; Kesely, K.R.; et al. Imatinib augments standard malaria combination therapy without added toxicity. J Exp Med 2021, 218, e20210724. [Google Scholar] [CrossRef] [PubMed]
- Sriwilaijareon, N.; Petmitr, S.; Mutirangura, A.; Ponglikitmongkol, M.; Wilairat, P. Stage specificity of Plasmodium falciparum telomerase and its inhibition by berberine. Parasitol Int 2002, 51, 99–103. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Malaria. Geneva: World Health Organization; 2023. Available from: https://www.who.int/health-topics/malaria#tab=tab_1. Accessed April 10, 2024. [Google Scholar]
Figure 1.
Telomere primary structure scheme. Figure by T. N. WAKAI, design was done in BioRender.com, and ideas adopted from [17].
Figure 1.
Telomere primary structure scheme. Figure by T. N. WAKAI, design was done in BioRender.com, and ideas adopted from [17].
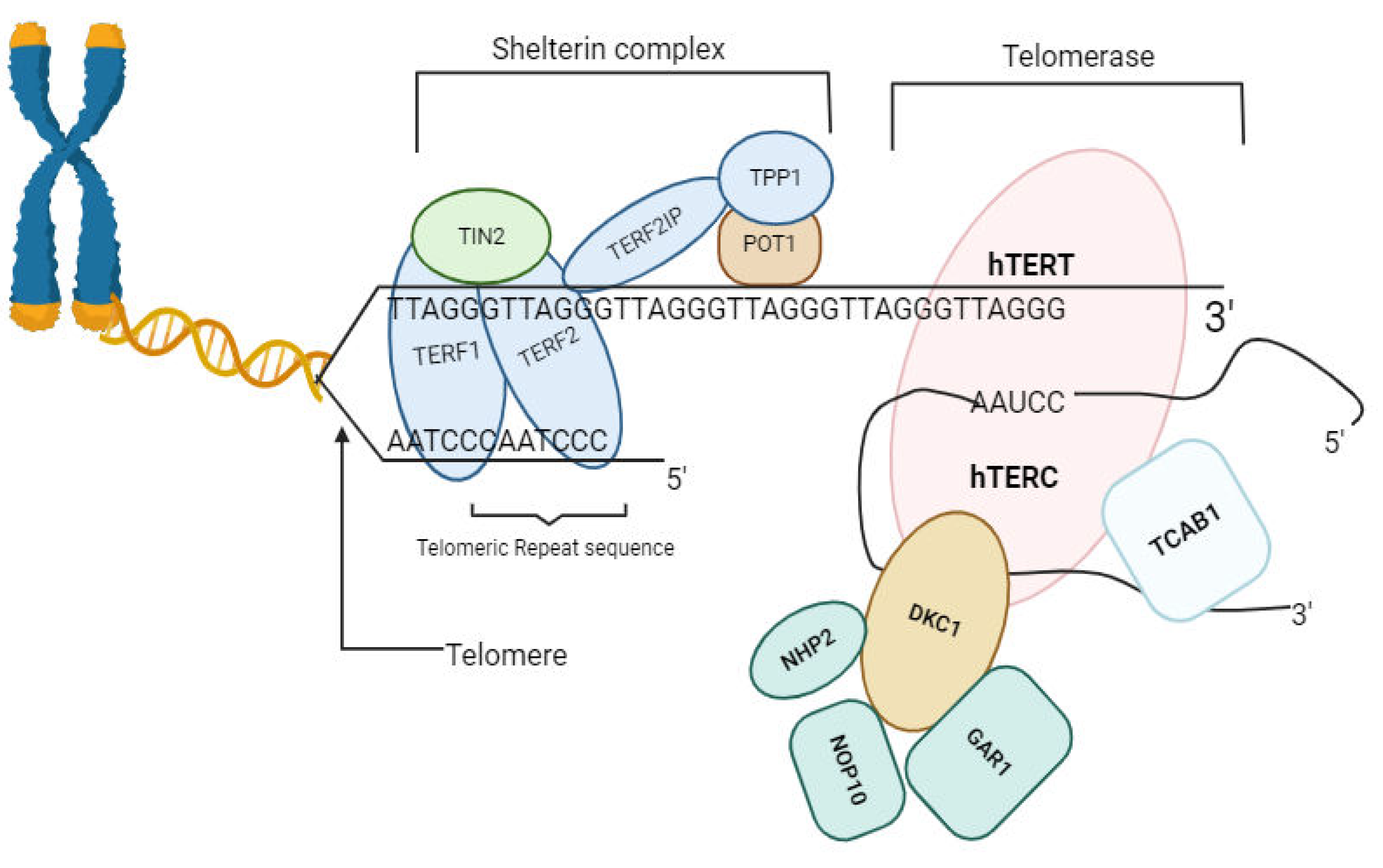
Figure 2.
The Role play by psychological Stress in Telomere Shortening and Dysfunction. Figure by T. N. WAKAI, designed was made in BioRender.com, and ideas adopted from [49].
Figure 2.
The Role play by psychological Stress in Telomere Shortening and Dysfunction. Figure by T. N. WAKAI, designed was made in BioRender.com, and ideas adopted from [49].
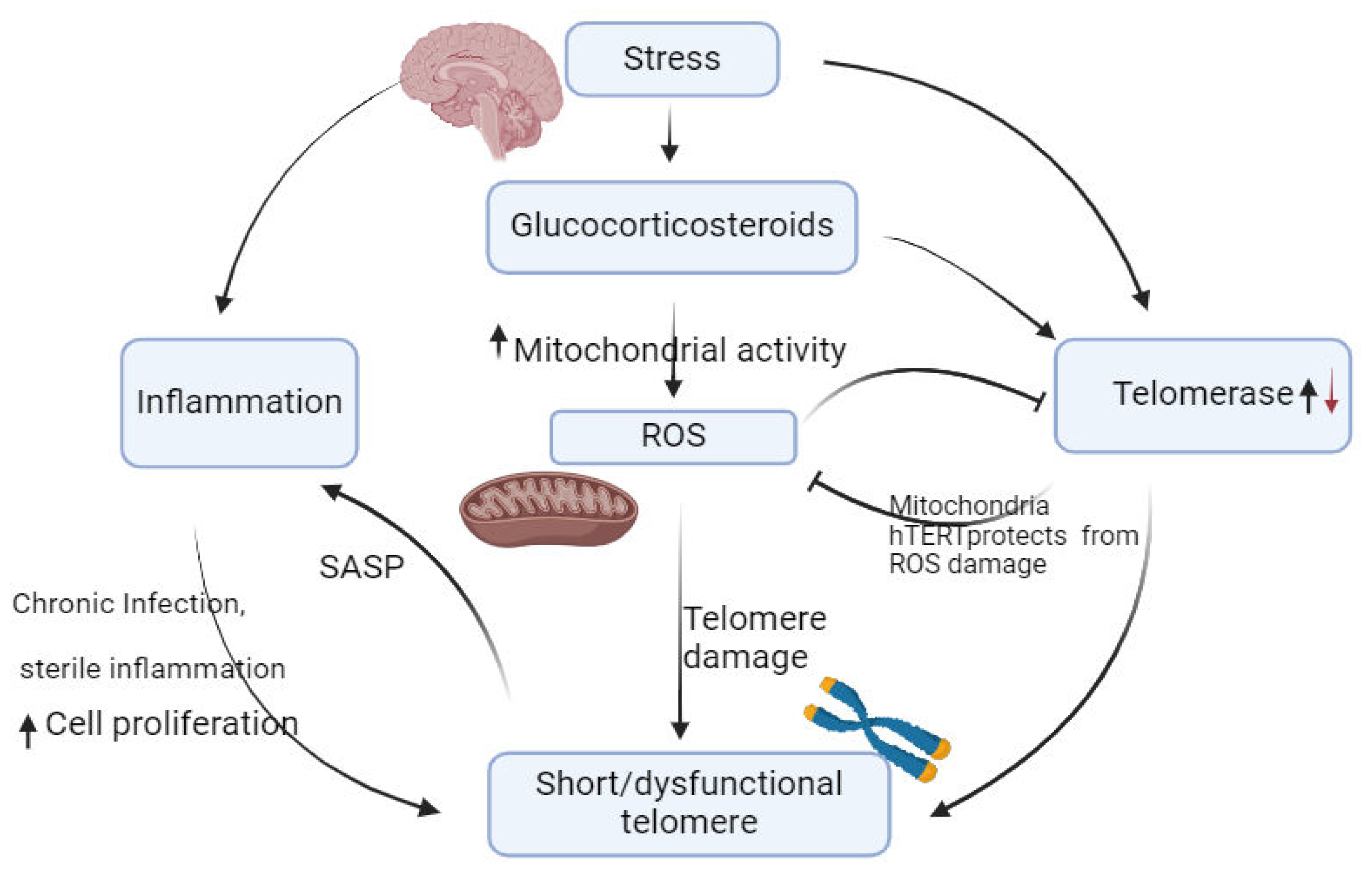
Figure 4.
Schematic representation of possible Telomerase-mediated stress response during malaria. Figure by T. N. WAKAI, designed in BioRender.com.
Figure 4.
Schematic representation of possible Telomerase-mediated stress response during malaria. Figure by T. N. WAKAI, designed in BioRender.com.
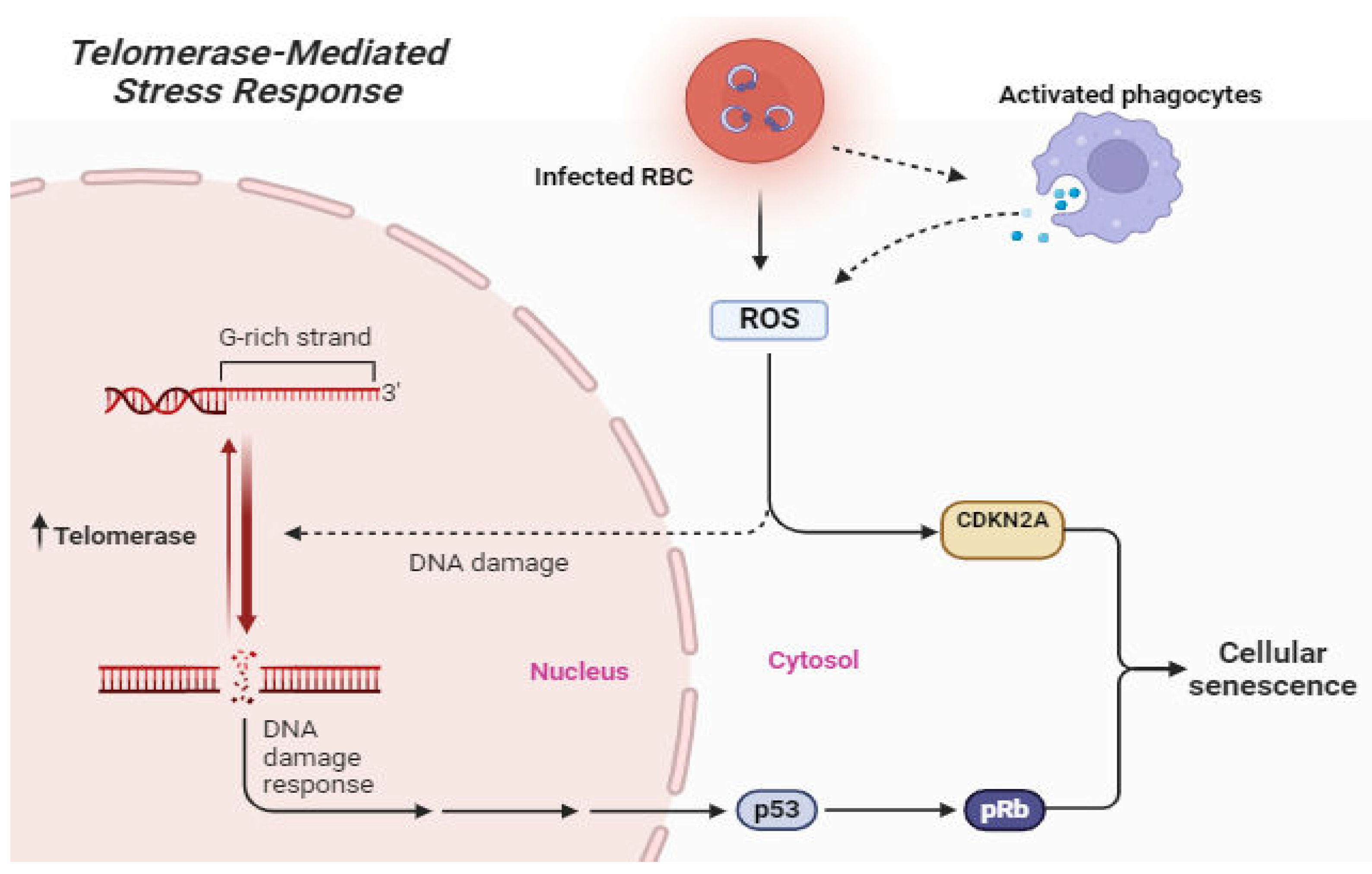
Figure 5.
Depletion of Repetitive DNA at Chromosome Ends Due to Telomere Shortening. Figure by T. N. WAKAI, designed in BioRender.com with ideas adopted from [35].
Figure 5.
Depletion of Repetitive DNA at Chromosome Ends Due to Telomere Shortening. Figure by T. N. WAKAI, designed in BioRender.com with ideas adopted from [35].
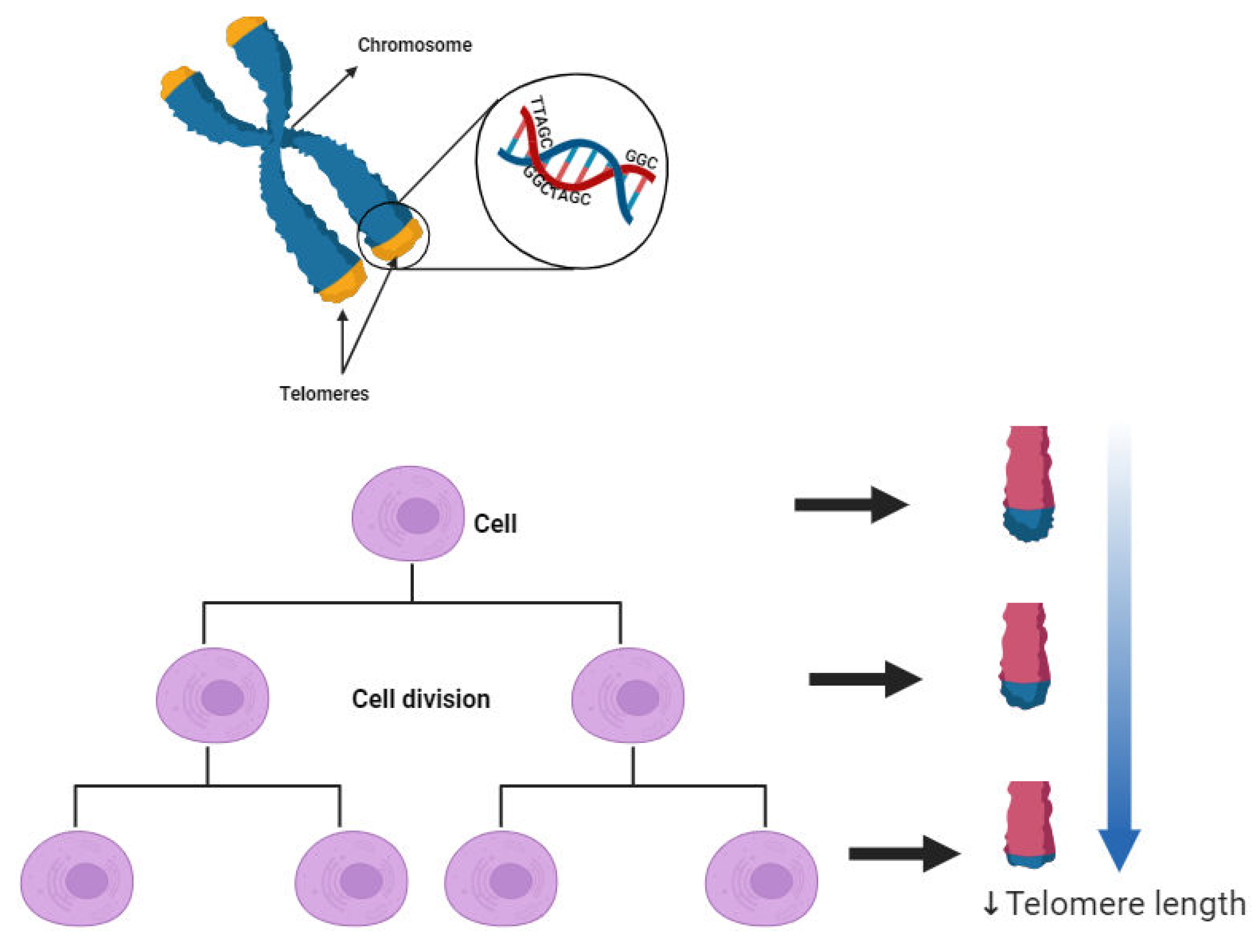
Figure 6.
Telomere Attrition and Telomerase-Mediated Extension [94].
Figure 6.
Telomere Attrition and Telomerase-Mediated Extension [94].
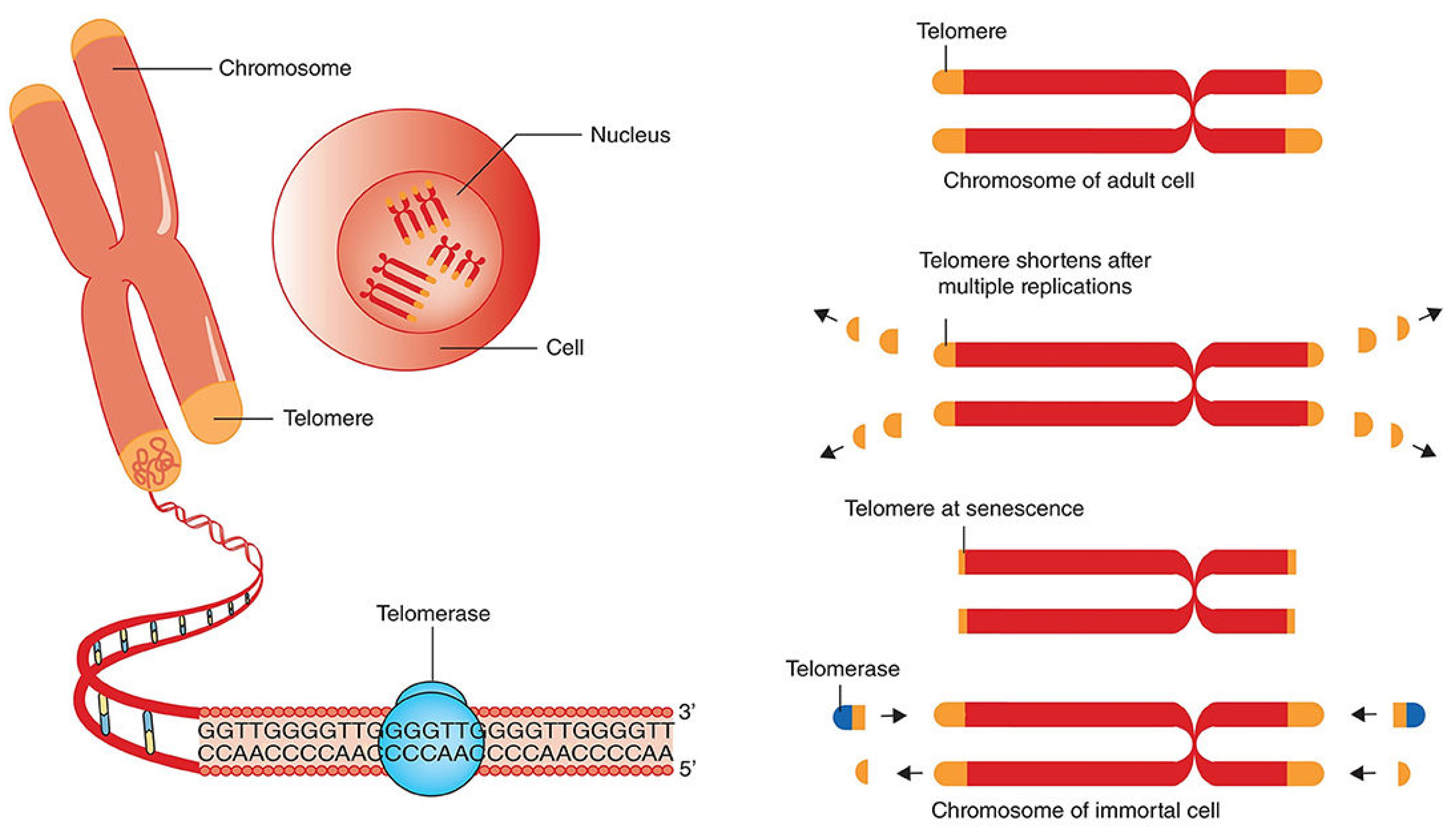
Figure 8.
Telomere shortening in Healthy (A) and Diseased cells(B and C). Figure by T. N. Wakai, designed in BioRender.com.
Figure 8.
Telomere shortening in Healthy (A) and Diseased cells(B and C). Figure by T. N. Wakai, designed in BioRender.com.
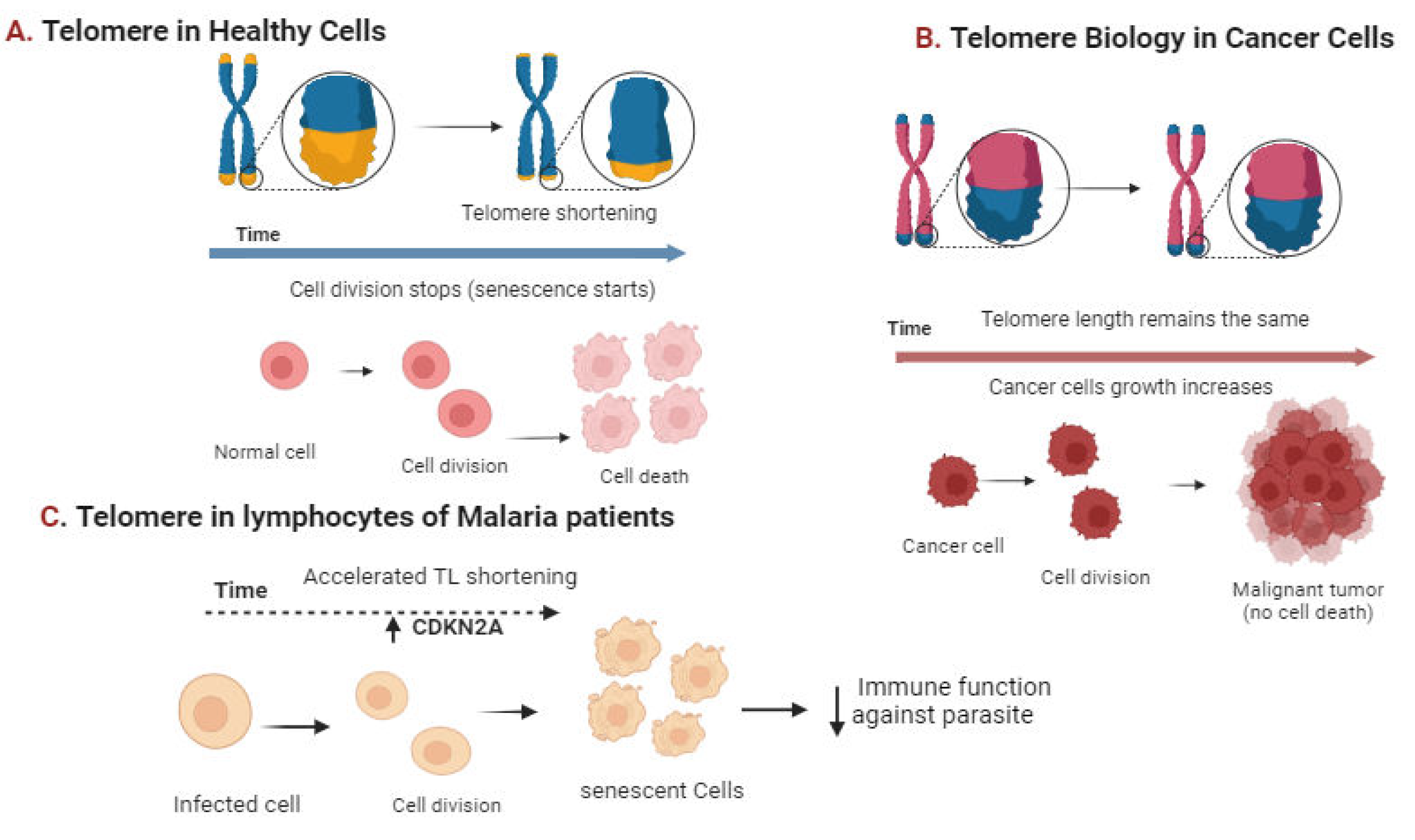
Figure 7.
Inhibition of telomerase by G-quadruplex ligands Figure by T. N. WAKAI, designed in BioRender.com, adopted from [45].
Figure 7.
Inhibition of telomerase by G-quadruplex ligands Figure by T. N. WAKAI, designed in BioRender.com, adopted from [45].
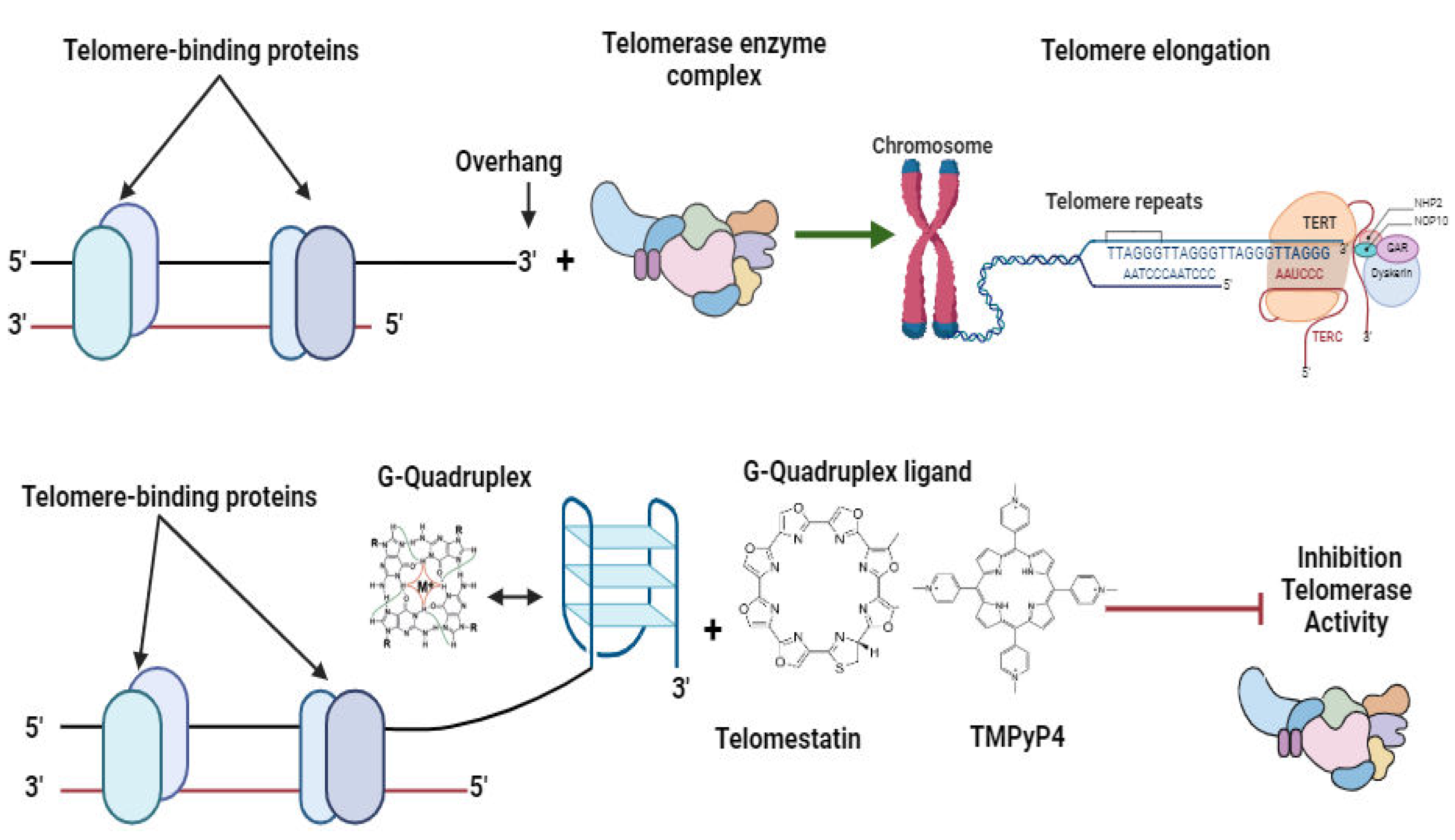

Table 1.
Key variations in Plasmodium and human telomerase systems.
|
Feature |
Plasmodium telomerase system |
Human telomerase system |
|---|---|---|
|
Telomere repeat sequence |
TTAGGG |
TTAGGG |
|
Telomerase RNA template |
TER1 |
TER1 |
|
Telomerase reverse transcriptase |
TERT |
TERT |
|
Accessory proteins |
TRF2, TPP1 |
TIN2, Pot1 |
|
Regulation |
Unknown |
Complex, regulated by multiple factors, including cell type, differentiation state, and stress |
Table 2.
Selected Compounds Targeting Telomeres and Telomerase in Malarial Parasites.
| Compound | Mechanism of action | Reference | |
|---|---|---|---|
| Bis{N-[(pyrrolo[1,2-a]quinoxalin-4-yl)benzyl]-3-aminopropyl}amine derivatives | Disrupts telomere maintenance by Stabilizing Plasmodium | Guillon et al., 2017 | |
| TMPyP4 (5,10,15,20-Tetrakis-(N-methyl-4-pyridyl)porphine) | Inhibition telomerase and parasite growth | [45] | |
| Dideoxy GPT |
Inhibition of telomerase activity and promotes p cell senescence in vitro |
[36] | |
| 17-AAG (Radicicol) | Depletion of Pfsir2 protein,deacetylation of Histone, shields telomerase access to telomeres | [42] | |
| Imatinib |
Inhibition of Plasmodium kinase PfPK5 and dyregulation of parasite cell cycle progression | [144,166] | |
| Berberine |
Inhibition of Telomerase activity in P. falciparum in stage specific manner |
[167] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



