Submitted:
16 February 2024
Posted:
19 February 2024
You are already at the latest version
Abstract
Cannabis sativa is one of the oldest plants utilized by humans for both economic and medical purposes. Although the use of cannabis started millennia ago in the Eastern part of the hemisphere, its use has moved and flourished in the Western Nations in more recent centuries. C. sativa is the source of psychoactive cannabinoids that are consumed as recreational drugs worldwide. The C21 aromatic hydrocarbons are restricted in their natural occurrence to cannabis (with a few exceptions). Delta-9-tetrahydrocannabinol (Δ9-THC) is the main psychoactive component in cannabis with many pharmacological effects and various approved medical applications. However, a wide range of side effects are associated with the use of Δ9-THC, limiting its medical use. In 1966, another psychoactive cannabinoid, Delta-8-tetrahydrocannabinol (Δ8-THC) was isolated from marijuana grown in Maryland but in very low yield. Δ8-THC is gaining increased popularity due to its better stability and easier synthetic manufacturing procedures compared to Δ9-THC. Passing the U.S. Farm Bill in 2018 led to an increase in the sale of Δ8-THC in the United States. The marketed products contain Δ8-THC from synthetic sources. In this review, methods of extraction, purification, and structure elucidation of Δ8-THC will be presented. The issue of whether Δ8-THC is a natural compound or an artifact will be discussed, and the different strategies for its chemical synthesis will be presented. Δ8-THC of synthetic origin is expected to contain some impurities due to residual amounts of starting materials and reagents as well as side products of the reactions. The various methods of analysis and detection of impurities present in the marketed products will be discussed. The pharmacological effects of Δ8-THC, including its interaction with CB1 and CB2 cannabinoid receptors in comparison with Δ9-THC, will be reviewed.
Keywords:
Δ8-THC
; isolation
; chemistry
; analysis
; pharmacology
1. Introduction
Cannabis sativa is one of the oldest plants used by humans either as a source of food, textile fiber, or for medical applications [1]. The medical uses of C. sativa started in Asia and the Middle East during the sixth century B.C. then it was introduced to the Western world in the nineteenth century [2,3]. The plant has been used for the treatment of many conditions including inflammation, pain, depression, nausea, glaucoma, and neuralgia [4]. The medical use of cannabis was not flourishing due to variations in chemical composition and quality, until the first isolation of the main psychoactive component, Δ9-tetrahydrocannabinol (Δ9-THC) [5].
Due to the variation in the concentration of Δ9-THC in the many cannabis plant varieties, three phenotypes were recognized, mainly for legal classification reasons. The drug-type or phenotype I must contain a higher concentration of Δ9-THC than the non-psychoactive cannabidiol (CBD) [6,7]. Phenotypes II and III contain a higher percentage of CBD (more than 0.5%). Phenotype II must contain at least 0.3% of Δ9-THC in the female plant, while phenotype III has Δ9-THC less than 0.3% [6,7]. More than 125 cannabinoids- oxygen-containing C21 aromatic hydrocarbons- were identified from C. sativa grown in different places around the globe [8]. As an increasing number of cannabinoid related structures were synthesized, the term “Phytocannabinoids” was given to the natural cannabinoids found in the cannabis plant [9]. Based on their chemical structure, phytocannabinoids were classified into 11 categories including the (-)-Δ9- trans -tetrahydrocannabinol (Δ9-THC) and (-)-Δ8-trans-tetrahydrocannabinol (Δ8-THC) types among others. Other types include, cannabidiol (CBD), cannabinol (CBN), cannabichromene (CBC), cannabigerol (CBG), cannabicyclol (CBL), cannabielsoin (CBE), cannabinodiol (CBND), cannabitriol (CBT), and Miscellaneous [6]. Δ9-THC expresses its pharmacological effects including hypolocomotion, hypothermia, catalepsy, and analgesia via activation of the CB1 receptor [10]. Δ9-THC also possesses neuroprotective, antispasmodic, and anti-inflammatory actions mediated through the activation of CB2 and PPAR receptors [11]. Δ9-THC is reported to have anti-proliferative potential against breast cancer cells and lung metastases [12,13]. Experimentally, Δ9-THC can inhibit the cell growth of many tumours such as melanoma, myeloma, hepatocellular carcinoma, and leukemia both in vitro and in vivo [14,15].
In 1985, the U.S. Food and Drug Administration (FDA) approved an oral drug product containing synthetic Δ9-THC (Marinol® capsules) to manage nausea and vomiting resulting from chemotherapy. Seven years later, it was approved by FDA as an appetite stimulant in HIV/AIDS patients [16]
Despite of the therapeutic activities of Δ9-THC, it may also result in serious adverse effects such as tachycardia and anxiety [17,18]. Furthermore, Δ9-THC may produce euphoria, cognitive impairments, and transient negative emotional states such as paranoia [19,20,21]. Reported data indicated that acute administration of Δ9-THC may induce transient psychosis in healthy volunteers in addition to cognitive impairments and electroencephalography (EEG) patterns comparable to psychosis [22]. The use of Δ9-THC may lead to disability or loss of sense of time and distance intervals [23]. Users of Δ9-THC may also suffer from panic reactions, disconnected thoughts, disturbing changes in perception, feelings of depersonalization, delusions, and hallucinatory experiences [24].
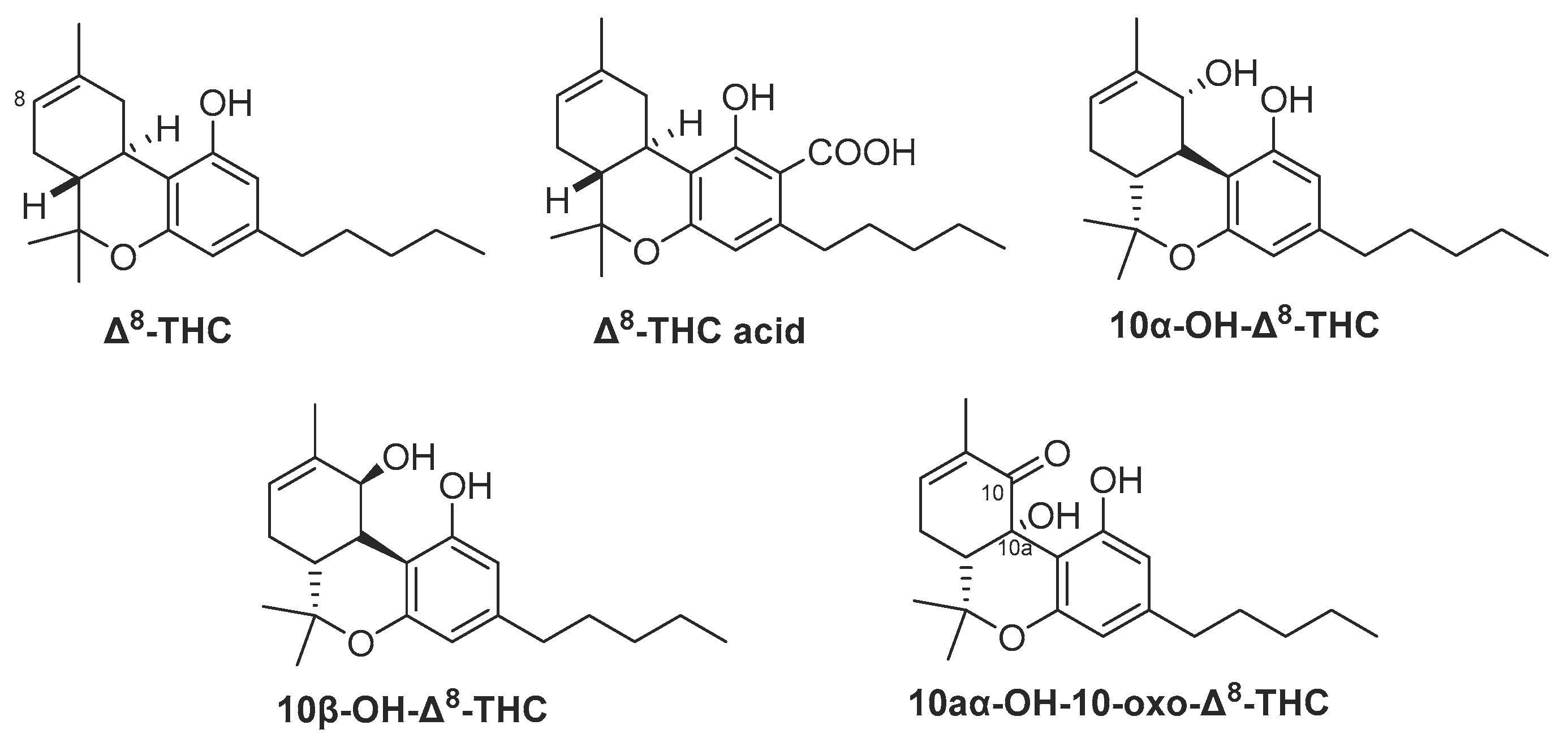
Δ8-THC is another phytocannabinoid present in much smaller amounts compared to Δ9-THC [6]. Δ8-THC was first isolated from marijuana (dried leaves and flowers of cannabis) grown in Maryland, USA [25]. Δ8-THC is the positional isomer of Δ9-THC and represents one of the minor groups of cannabinoids with the same chemical skeleton of tetrahydrocannabinoids. In 1975, Krejcí et al. reported on the characterization of the carboxylic acid precursor of Δ8-THC (Δ8-trans-tetrahydrocannabinolic acid A) (Δ9-THC acid A) from cannabis of Czechoslovakian origin [26]. Three members of the same skeleton were isolated from high-potency C. sativa grown at the University of Mississippi, USA, namely: 10α-hydroxy-Δ8-tetrahydrocannabinol, 10β-hydroxy-Δ8-tetrahydrocannabinol and 10aα-hydroxy-10-oxo-Δ8-tetrahydrocannabinol [27,28]. In the plant, Δ8-THC is typically formed from nonenzymatic isomerization of Δ9-THC and/or cyclization of CBD [29]. Using chemical synthesis, Δ8-THC is much easier to produce than Δ9-THC [30]. Additionally, compounds of the Δ8-type are much more stable than their Δ9-counterparts. Indicative of its chemical stability, Δ8-THC was detected in a burial tomb in Jerusalem dating back to the fourth century B.C [31]. Pharmacologically, it acts as a partial agonist to CB1 and CB2 cannabinoid receptors similar to Δ9-THC but with lower activity on both receptors [32]. The psychoactivity of Δ8-THC was claimed to be about 75% that of Δ9-THC [33]. The improved stability of Δ8-THC and its derivatives compared to Δ9-THC, coupled with the ease of its synthesis qualifies Δ8-THC to be a better lead compound for exploring the undiscovered aspects of cannabinoid receptors [34].
The availability and sale of Δ8-THC, obtained mainly from synthetic sources, has increased in the United States after the passing of the Farm Bill in 2018 [35]. One report indicated that Δ8-THC was more effective and better tolerated than Δ9-THC [36]. Based on the fact that Δ8-THC in commercial products is chemically synthesized in an unregulated market, the presence of impurities and byproducts as well as residual solvents is often likely. Some marketed Δ8-THC products were analyzed for the presence of such contaminants and several compounds such as 9β-hydroxy-hexahydrocannabinol (9β-OH-HHC) and its isomer, 9α-OH-HHC were identified. However, other impurities are still unknown [37].
The legal situation of Δ8-THC in the USA varies from one state to another [35]. The American FDA received more than 100 reports on adverse effects among recreational users between December 2020, and February 2022. Among these reported adverse effects, 55% required intervention such as evaluation by emergency medical services or hospitalization [38]. Since these products are not approved nor evaluated for safety and/or analyzed for content, the cause of these adverse effects cannot be currently attributed to Δ8-THC alone. This review presents all the currently available data regarding the isolation, synthesis, analysis, and biological profile and regulation of Δ8-THC and its products.
2. Isolation of Δ8-THC and its derivatives
Δ8-THC was first isolated by Hively et al. in 1966 from the petroleum ether extract of the flowering tops and leaves of fresh cannabis grown in Maryland (USA) [25]. This extract was fractionated using silicic acid column chromatography eluted with benzene and the column fractions rich in Δ8-THC were combined and further purified on silicic acid Thin thin-layer chromatography (TLC) plates impregnated in silver nitrate using benzene as an eluent. The same research group also isolated Δ8-THC along with Δ9-THC from another sample of cannabis of Mexican origin and found that Δ8-THC constituted up to 10% of the total tetrahydrocannabinol content of the fresh marijuana samples of the above two cannabis varieties [25]. The chemical structure of Δ8-THC was identified by its total synthesis from cannabidiol (CBD) after treatment with p-toluene sulfonic acid using a previously reported method [39] and comparing the spectroscopic data of the product (IR, UV, NMR, and optical rotation) with those of Δ9-THC. Catalytic hydrogenation of both Δ8-THC and Δ9-THC gave two colorless resinous compounds with identical IR spectra and optical rotation indicating that both compounds have the same stereochemistry [25].
The isolated Δ8-THC was named Δ6-THC, while, Δ9-THC was named Δ1-THC based on the monoterpene numbering system that was dominant around the time (Figure 1).
In 1975, Hanuš and Krejči reported the isolation of Δ8-THC acid (Figure 2) from C. sativa of Czechoslovak origin [40]. The dried flowering tops were extracted with petroleum ether and the extract was treated with 2% Na2CO3 solution followed by acidification with cold 5% H2SO4, then extracted with ether to produce an acid fraction containing cannabinoid acids. This fraction was purified over a silica gel column eluted with chloroform to yield a cannabinoid acids mixture, which was esterified by diazomethane and further purified on a silica gel column using n-hexane: ether (2:1) as a mobile phase to afford Δ8-THC acid methyl ester. Identification of this methyl ester was performed by IR, UV, and NMR spectroscopic analysis as well as EI-Mass spectrometry [40].
From high potency C. sativa (≥ 20% w/w Δ9-THC) grown at the University of Mississippi, USA, three minor hydroxylated derivatives of Δ8-THC were isolated in 2015, and identified as 10aα-OH-10-oxo-Δ8-THC, 10α-OH-Δ8-THC and 10β-OH-Δ8-THC (Figure 2). The dried buds were sequentially extracted with hexanes, CH2Cl2, EtOAc, EtOH, EtOH/H2O, and H2O. The hexanes extract was fractionated on successive silica gel columns followed by purification on semi-preparative reversed-phase HPLC using ACN: H2O (75: 25) to afford 10aα-OH-10-oxo-Δ8-THC along with other minor oxygenated cannabinoid derivatives [28]. The CH2Cl2 and EtOAc extracts were combined and subjected to a silica gel Vacuum Liquid Chromatography column (VLC) on silica gel eluted with a gradient mixture of EtOAc/hexanes to give nine fractions (A-I). Fraction D was chromatographed on a silica gel column and then further purified on a flash C18 column using 60% aqueous MeOH as a mobile phase. Two cannabinoids, 10α-OH- Δ8-THC, and 10β-OH-Δ8-THC, were finally isolated using a silica HPLC column, isocratically eluted with 10% EtOAc/n-hexane [27].
Figure 2.
Chemical structure Δ8-THC and its derivatives isolated from cannabis.
3. Is Δ8-THC a Natural Secondary Metabolite or an Artifact?
Upon its initial isolation in 1966, Hively et al. claimed that Δ8-THC is a natural metabolite and not an artifact produced during the extraction and isolation procedures. The researchers performed two experiments to support their assumption; first, they subjected pure Δ9-THC to chromatography using the same methods described for marijuana extract. They discovered that no Δ8-THC was formed [25]. Second, they extracted 2-years old Spanish marijuana and 3-years old Mexican marijuana with petroleum ether, and only Δ9-THC was detected and no Δ8-THC based on TLC analysis [25]. Later, Pars and Razadan (1971) studied the stability of synthetic Δ8-THC and Δ9-THC at 80˚C. Δ8-THC showed no change in the content, while Δ9-THC was totally decomposed to CBN only and no Δ8-THC was observed as indicated by GC analysis [41].
Turner et al. (1973) reported a 2-year stability study of the four cannabinoids, Δ9-THC, Δ9-THC acid A, CBD, and CBN in dried cannabis plant samples stored at five different temperatures (-18°C, 4˚C, 22°C, 37°C, and 50°C). GLC was used to analyze the content of each cannabinoid in all samples every 10 weeks for 104 weeks. At the end of the study, it was concluded that the concentration of Δ9-THC, and Δ9-THC acid A was decreased as the storage temperature increased, while, the CBN content increased with increasing temperature except it decomposed at 80°C after 50 weeks. CBD content did not change at any of the temperatures. Also, no change in the Δ8-THC was observed in this study [42]. In the same year, the same research group reported that chloroform is an efficient extraction solvent for C. sativa compared to other 7 solvents (benzene, pentane, hexane, petroleum ether, ethanol, acetone and ether) and the cannabinoids present in this extract (Δ9-THC, CBD, CBC, and CBN) were stable for 144 hrs based on GC analysis (before and after silylation) [43]. Δ8-THC was detected in trace amounts but when the GC column contains a free hydroxyl group or the extraction solvent contains H2O, the content of Δ8-THC increased. The researchers postulated that Δ8-THC is an artifact formed during the extraction procedure or GC analysis.
Based on the above experiments, further research is needed to answer the question, “Is Δ8-THC a natural (biosynthetically produced) cannabinoid or an artifact?”.
4. Synthesis of Δ8-THC
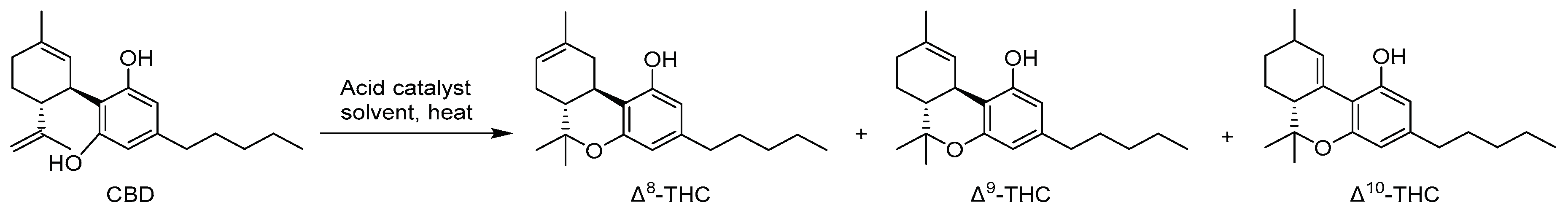
Since the concentration of the naturally occurring Δ8-THC in the cannabis plant is exceedingly low, its extraction holds little economic viability due to the substantial associated expenses [44,45]. Consequently, almost all the Δ8-THC on the market today is synthetically produced, predominantly from the chemical conversion of cannabidiol (CBD) [46]. CBD can be readily transformed into Δ8-THC through acid-catalyzed cyclization reaction [47,48], as shown in Figure 3. The use of common household chemicals to obtain the same chemical transformation is readily accessible, and numerous internet sources provide comprehensive step-by-step descriptions of this conversion [36].
5. Analysis of Δ8-THC
The small difference in the chemical structure of the two isomers of tetrahydrocannabinol has a noticeable effect on the three-dimensional shape of the molecule, which may account for the differences in the pharmacological and psychological effects described [49]. Δ8-THC shares the same mechanism of action as Δ9-THC as a partial agonist of the cannabinoid CB1 receptor but with less potency [33,49,50]. The increasing availability of products containing Δ8-THC and its isomers, which are erroneously classified as hemp, emphasize the need for precise and accurate analytical methods to evaluate its existence and concentration in various matrices.
5.1. Analysis of Δ8-THC in cannabis biomass and cannabis-derived products
Liquid chromatography coupled with photodiode array detection (HPLC-PDA) is commonly used for the analysis of Δ8-THC in samples from herbal cannabis and/or cannabis-derived products (extracts, oils, concentrates, and hash). In contrast, liquid chromatography coupled to mass spectrometry (LC-MS) is the preferred method for the determination of Δ8-THC and its metabolites in biological fluids. Additional analytical techniques including gas chromatography-mass spectrometry (GC-MS), gas chromatography with flame ionization detector (GC/FID), and quantitative nuclear magnetic resonance (QNMR) were also reported for Δ8-THC analysis.
HPLC-PDA methods are very often utilized in association with the quantification of Δ8-THC in complex extracts from C. Sativa. In 1979, Masoud et al. reported the very first method for the analysis of Δ8-THC in Mexican cannabis extracts using HPLC with electrochemical detection [51]. More recently, Correia et al. (2023) developed and validated an HPLC-PDA method for the quantification of Δ8-THC, in addition to Δ9-THC, CBD, CBN, Δ9-THCA, and CBDA in cannabis products, following an ultrasound-assisted solid-liquid extraction protocol [52].
Recently, an HPLC method was validated for the analysis of Δ8-THC and other 14 cannabinoids in eight cannabis plant material samples of four chemovars (high THC, high CBD, THC/CBD, and high CBG) and two E-cigarettes. Δ8-THC content (%w/w) was in the range of 0.036—0.60 in high THC, high CBD, THC/CBD chemovars, while it was not detected in both CBG chemovar and cannabis e-cigarettes [53].
An HPLC-PDA method was developed by Duffy et al. in response to the E-cigarette or Vaping Product Use-Associated Lung Injury (EVALI) outbreak in New York State. The analyzed samples from cannabis vaporizer cartridges contained abnormally high amounts of Δ8-THC whereas medical marijuana products typically do not. Δ8-THC amounts ranged from 9.6%-98%w/w in the analyzed vaping products. Moreover, untargeted analysis by GC-MS and LC-HRMS was performed for the determination of diluents and other cartridge components in order to further understand the occurrence of EVALI [54].
Using standards in the validation phase, Δ8-THC could be baseline separated from ten other cannabinoids using acetonitrile (with 0.05% formic acid) and water (with 0.05% formic acid) as a mobile phase in a gradient mode [55]. This study used an Ultrahigh-Performance Liquid Chromatograph coupled with a Photodiode Array and single quadrupole Mass Spectrometry detectors (UPLC-PDA-MS) for its analysis in different C. sativa samples including leaves, flower buds, and hashish. However, only a negligible amount of Δ8-THC was found in one hashish sample [55].
Different analytical techniques have been employed for the evaluation of cannabinoid content including Δ8-THC in hemp seed oil, and hemp distillate. In 2019, Križman et al. developed an HPLC-PDA method, applying isocratic elution at 37 °C for the analysis of hemp plant samples. However, only one plant sample analyzed contained a detectable amount of Δ8-THC [56]. Based on the author's claim, the use of 275 nm detection wavelength gave a significantly better sensitivity (signal-to-noise ratio) than the conventional 228 nm wavelength commonly used for cannabinoid analysis. In the total of four hemp samples, Δ8-THC was detected and quantified in only one sample at a concentration of 0.06 % w/w [56].
A UPLC-PDA method was developed and validated by Song et al. (2022) for the determination of 16 cannabinoids including Δ8-THC in different hemp-derived concentrates. The mobile phase composition was optimized by studying different percentages of acetonitrile and formic acid concentrations in water. The best resolution between Δ9-THC and Δ8-THC peaks was achieved using 0.028% formic acid and 73% acetonitrile [57]. Method specificity was validated by quadrupole time-of-flight (Q-TOF) mass spectrometry. Only two out of nine hemp concentrates analyzed contained Δ8-THC, namely, one type of Δ8-THC distillate and one type of Δ8-THC hemp shatter where Δ8-THC contents were 70.96% and 72.32% (w/w), respectively [57].
Generally, LC-MS/MS techniques are used for analyzing Δ8-THC in complex cannabis matrices such as blood samples or food products. The technique cannot distinguish Δ8-THC from interfering cannabinoids such as Δ9-THC, CBD, CBC, and CBL, based on the analyte's mass-to-charge ratio (m/z) as they exhibit nearly identical m/z values and share matching fragmentation spectra [58]. As a result, for accurate analysis of Δ8-THC, a full chromatographic separation has to be achieved.
Christinat et al. (2020) developed and validated an LC-MS/MS method for the quantification of 15 phytocannabinoids including Δ8-THC in different food products. The mobile phase consisted of 0.1% formic acid solution in water (solvent A) and acetonitrile (solvent B) in a gradient elution mode. In 14 out of 23 hemp-based food products, Δ8-THC amounts were <1 mg/kg, while in one CBD oil sample contained as much as 4.96 mg/kg [59].
An ESI-LC/MS method was reported for the metabolic profiling of phytocannabinoids in cannabis, where cannabinoid concentration from ethanolic extraction was determined using deuterated cannabinoids (Δ8-THC-d9, Δ9-THC-d9, CBD-d3, and CBN-d3) as internal standards. The identified Δ8-THC amount was below the limit of quantification of the reported method (1.25 ng/mL) [60].
In 2012, Trofin et al. developed a GC-MS method for the determination of cannabinoids with baseline separation of Δ8-THC and Δ9-THC, in different types of herbal cannabis. The analysis was performed using a fused silica capillary column and a temperature program from 150°C to 280°C [61].
5.2. Analysis of Δ8-THC, impurities, and possible contaminants in commercial consumer products
In the United States, commercial products containing Δ8-THC in high concentrations are gaining popularity, since the 2018 Farm Bill made tremendous changes to the regulation of hemp products [62]. A wide variety of Δ8-THC products are being sold on the shelves of shops, gas stations or from online sources.
These products are manufactured from CBD derived from CBD-rich hemp flowers, mainly based on the acid cyclization of cannabidiol (CBD) as the reactant (Figure 4). The cyclization reaction categorizes the resulting Δ8-THC by some scientists as a synthetic cannabinoid, thereby designating it as a controlled substance [47]. However, the discussion on the legality of Δ8-THC is ongoing for now. According to the stated arguments, Δ8-THC in commercial products is typically a synthetic product, consequently, there are valid concerns regarding the presence of impurities in these products with unknown effects on the human body. A recent FDA consumer warning update (2022) addressed several concerns with the chemical manufacturing process mentioned above [38]. Some manufacturers may use potentially unsafe household chemicals to make Δ8-THC through this chemical synthesis process. Additional undisclosed chemicals may be used to change the color of the final product or to increase shelf life. The final Δ8-THC product may have potentially harmful by-products (contaminants) and there is uncertainty with respect to other potential contaminants that may be present or produced depending on the composition of the raw starting materials. When consumed or inhaled, contaminants from the synthetic starting materials as well as byproducts of the synthetic products of Δ8-THC, can be harmful.
Δ8-THC marketed products were found to contain unnatural cannabinoids suggesting these were of synthetic origin [37].
Figure 4.
Synthesis of Δ8-THC from CBD (yielding small amounts of Δ9-THC and Δ10-THC).
As a result of the synthetic production methods and the lack of regulatory requirements, hemp-derived products containing Δ8-THC and other isomers are at greater risk of contamination and adulteration than herbal cannabis products sourced from licensed cannabis producers for medical or adult use. By the year 2020, the Interim Final Rule (2020 IFR) issued by the Drug Enforcement Administration (DEA) confirmed that hemp-derived THC products are not legalized by the 2018 Farm Bill [63]. As a result, the increased proportion of Δ8-THC and impurities present in various non-traditional marijuana-containing products such as vape cartridges, extracts, and edibles has become an important analytical challenge.
In order to address this emerging class of Δ8-THC containing consumer products, different analytical techniques have been reported in the literature. Radwan et al. (2023) isolated and characterized impurities found in synthesized Δ8-THC raw material in the US.A. using GC-MS, one Dimension and two Dimension Nuclear Magnetic Resonance spectroscopic (1D and 2D NMR) analysis as well as High-Resolution Electron Spray Ionization Mass Spectrometry (HR-ESI-MS) [37]. The identified compounds included, olivetol, Δ4,8-iso-tetrahydrocannabinol (Δ4,8-iso-THC), iso-tetrahydrocannabifuran (iso-THCBF), cannabidiol (CBD), Δ8-iso tetrahydrocannabinol (Δ8-iso-THC), Δ8-cis-iso-tetrahydrocannabinol (Δ8-cis-iso-THC), 4,8-epoxy-iso tetrahydrocannabinol (4,8-epoxy-iso-THC), Δ9-THC, 8-hydroxy-iso-THC (8-OH-iso-THC), 9α-hydroxyhexahydrocannabinol (9α-OH-HHC), and 9β- hydroxyhexahydrocannabinol (9β-OH-HHC) (Figure 5).
The presence of the precursor molecule olivetol suggests a fully synthetic Δ8-THC in these products. This hypothesis is supported by some studies that reported the synthesis of tetrahydrocannabinols from olivetol as a precursor [64,65]. The presence of olivetol was also reported by Meehan-Atrash and Rahman (2021) as a contaminant in 27 Δ8-THC products [66]. The study found that none of the analyzed products had accurate labeling of Δ8-THC content (within a range of ± 20 % of the concentration on the label). Moreover, all the products contained reaction by-products in addition to olivetol, including Δ4,8-iso-THC, 9-ethoxyhexahydrocannabinol (9-EtO-HHC), Δ9-THC, and a previously unidentified cannabinoid (iso-THCBF, Figure 5) in addition to heavy metals [66]. Quantitative NMR and GC-MS methodology were also used to analyze the major components of 27 e-cigarette products advertising Δ8-THC. All products were found to contain at least some undisclosed diluents and/or reaction side-products from the conversion of CBD to Δ8-THC. Among others, triethyl citrate (diluent; present in 7 out of 27 samples), olivetol (likely the starting material of the chemical synthesis in 22/27 samples), and Δ8-THC -iso-THC (a byproduct of the acid-catalyzed CBD cyclization; 27/27 samples) were identified (Figure 5).
In addition, the previously unknown cannabinoid (5aR,9aS)-5a-isopropyl-8-methyl-3-pentyl-5a,6,7,9a-tetrahydrodibenzo[b,d]furan-1-ol or iso-THCBF was found (Figure 5). It was present in nearly all products tested but was not quantifiable in products containing the diluent triethyl citrate due to spectral overlap. Analysis of heavy metals by ICP-MS confirmed detectable levels of magnesium, chromium, nickel, copper, zinc, mercury, lead, and silicon [66]. Moreover, QNMR indicated that Δ8-THC levels can vary as much as 40% from the labeled value, suggestive of poor testing capabilities or perhaps even falsified results. For one of the brands tested, the average of the sums of Δ8-THC and Δ8-iso-THC for each product was not significantly different from the average reported Δ8-THC content suggesting the analysis method (HPLC-UV as stated in the certificate of analysis) cannot discriminate between the two compounds [66].
Ray et al. (2022) used 1H-NMR, HPLC-UV, and HPLC-MS to determine unknown impurities in Δ8-THC consumer products purchased online or from local retailers from the U.S.A. [67]. Ten Δ8-THC productes including distillates and vaporizers were analyzed during the study. The results indicated that the tested samples contain a range of impurities in concentrations far beyond what is declared on certificates of analysis for these products. Impurities were identified as compounds resulting from low-quality CBD (extracted from hemp used as starting material) as well as known side reaction products from the cyclization reaction used to convert CBD into Δ8-THC. The identified compounds included cannabidivarin (CBDV) and cannabidihexol (CBDH). The study concluded that the problem with these products is threefold: impure CBD starting material, poor post-reaction purification, and inadequate separation of closely related cannabinoids during laboratory analysis of the final product [67].
In one study, in addition to Δ8-THC, the cannabinoids Δ9-THC, CBD, 9(S/R)- Δ6a,10a-THC, and (6aR,9R)- Δ10-THC were identified in an e-cigarette cartridge. All of these are isomers of Δ9-THC. The identification was based on retention time and fragmentation patterns compared with reference materials. Moreover, some unknown components were also detected within the vape oil of the same e-cigarette [45].
The main cause of the presence of THC isomers is probably due to exposure of cannabis source materials, such as cannabis concentrates or converted hemp materials, to chemical and thermal treatments during the manufacturing process. Using GC-MS and HPLC-PDA analyses, some isomers have been identified in many of the vaping liquids and distillates including Δ8-THC, Δ9-THC, 9R-Δ6a,10a-THC, 9S-Δ6a,10a-THC, 6aR,9R-Δ10-THC, 6aR,9S-Δ10-THC, Δ6a,10a-THC, exo-THC, Δ6a,10a-tetrahydrocannabivarin (Δ6a,10-THCV), THCA, Δ9-THCV, Δ8-THCV, CBD, CBDV, CBN, CBG, and CBC [68]. Significant amounts of Δ8-THC were found in the analyzed vaping liquids and distillates ranging from 0.4-79% w/w, while Δ9-THC amounts ranged from 1.4-93.1 %w/w as determined by GC-MS analysis. In addition, low amounts of the Δ8-THCV, Δ9-THCV, and CBDV were determined. The chemical structures of THC isomers are depicted in Figure 6.
5.3. Analysis of Δ8-THC and Δ9-THC metabolites in different biological matrices.
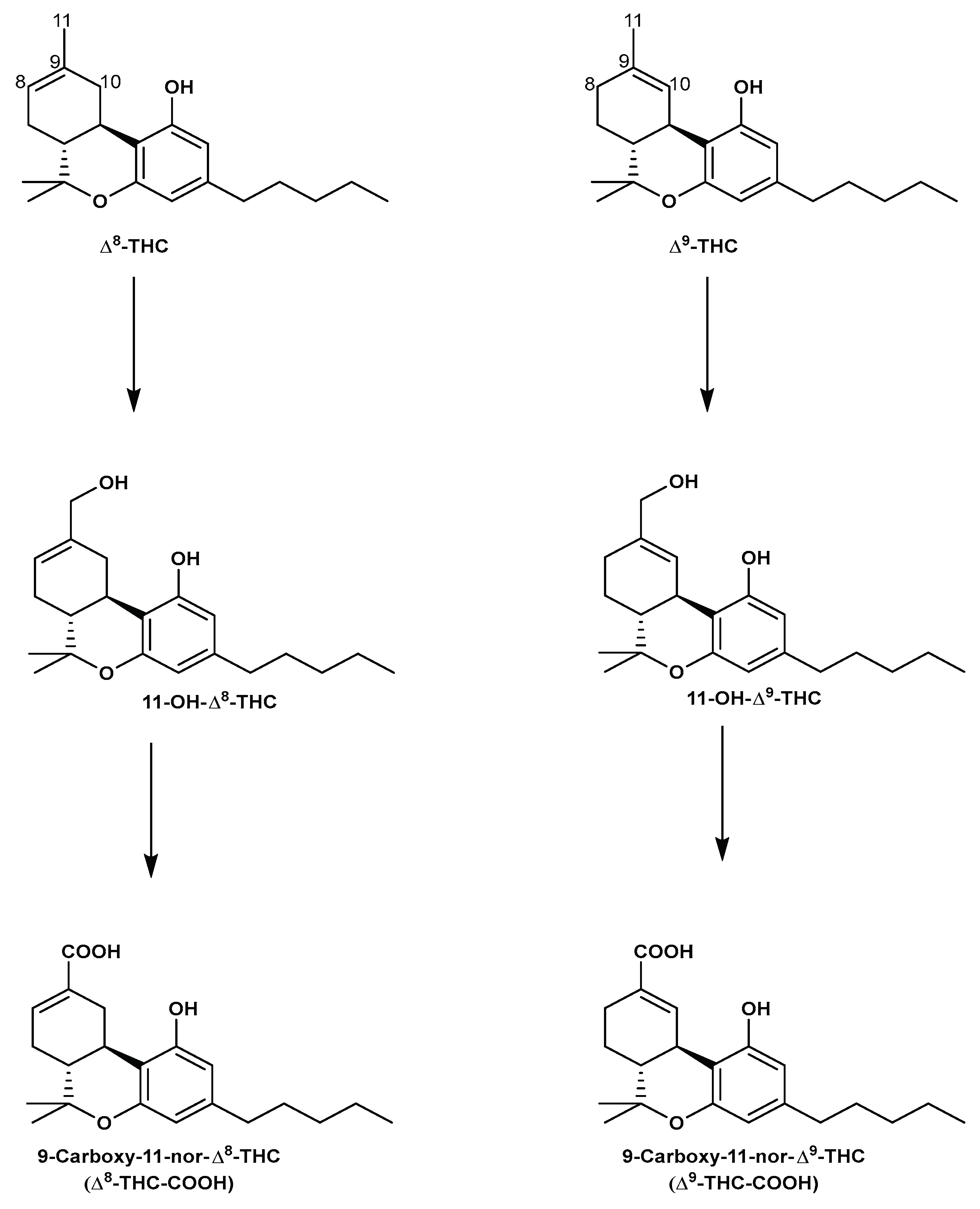
In human liver microsomes, Δ8-THC is enzymatically metabolized similar to Δ9-THC. In brief, Δ8-THC is oxidized to the pharmacologically active, 11-hydroxy-Δ8-tetrahydrocannabinol (11-OH-Δ8-THC) [69,70], and then further to the non-psychoactive 9-carboxy-11-nor-Δ8-tetrahydrocannabinol (Δ8-THC-COOH), which is finally conjugated to form its corresponding glucuronide before being excreted, mainly in urine [71,72,73]. Figure 7 shows the metabolic scheme for Δ8-THC and Δ9-THC.
The terminal elimination half-life of Δ8-THC has not yet been reported in humans. The terminal half-life of Δ9-THC is up to 4 days in chronic users [71]. This long half-life is attributed to the slow release of the highly lipophilic Δ9-THC from fat tissue to plasma. Given the similar distribution to fat tissue, it is expected that Δ8-THC will have a similarly long terminal half-life, which means that Δ8-THC metabolites may be detected for days or even weeks after the last consumption [71]. Although human data on the metabolism of Δ8-THC is limited, some information has been learned from cases involving accidental overdose [74].
At the Karolinska University Laboratory in Sweden, and during routine LC-MS/MS analysis of the human Δ9-THC metabolite Δ9-THC-COOH in urine, an unidentified peak was detected with a slightly different retention time than Δ9-THC-COOH. Using an optimized LC-HRMS method with better separation between the two peaks, the unknown peak was confirmed to be Δ8-THC COOH [75].
Pharmacokinetic studies were conducted using the LC-MS method for the determination of Δ8-THC and its metabolite Δ8-THC-COOH in guinea pig plasma. A liquid-liquid extraction protocol was developed to extract the target analytes from guinea pig plasma using acetonitrile: ethyl acetate (50:50, v/v) [76].
Figure 7.
The metabolic pathway of Δ8-THC and Δ9-THC.
Rzeppa et al. (2021) used HPLC-MS/MS for the analysis of Δ8-THC-COOH as a less common metabolite compared to Δ9-THC-COOH. The authors also carried out a GC-MS/MS for simultaneous analysis of Δ8-THC-COOH. The content of Δ8-THC-COOH in doping samples ranged from 0.05 to 2.83% [44]. These findings are in line with those made by in vitro experiments using human liver microsomes [71].
LC-QTOF/MS was used for the analysis of Δ9-THC and Δ8-THC in the serum and urine of an accidentally intoxicated 2-year-old girl with Δ8-THC infused gummies [77]. The analysis confirmed the presence of Δ8-THC and the absence of Δ9-THC in the gummy. Plasma concentrations of Δ8-THC and Δ8-THC-COOH were 107.6 ng/mL and 746.5 ng/mL, respectively, while urine analysis exhibited 1550 ng/mL of Δ8-THC-COOH. The study revealed that the symptoms observed in individuals exposed to Δ8-THC closely resemble the established effects of Δ9-THC. These resemblances pose challenges for clinicians attempting to pinpoint the exact substance that has been ingested.
Another LC-MS/MS method was developed for the analysis of Δ8-THC and Δ9-THC and their metabolites in blood. Analyte extraction was carried out using 10 mM phosphate-buffered saline, 1 % phosphoric acid, and mixed solvent (80: 10: 10 hexane/ethyl acetate/methyl-tert-butyl-ether) [45].
Analysis of 1,504 urine specimens with a positive immunoassay Δ9-THC initial test using a liquid chromatography-tandem mass spectrometry (LC-MS-MS) showed the presence of Δ8-THC-COOH in 378 samples (15 ng/mL, cutoff), compared to 1,144 specimens containing Δ9-THC-COOH [78].
Isomeric separation was achieved between Δ8-THC-COOH and Δ9-THC-COOH by using an automated online μSPE-LC-MS/MS method. Δ8-THC-COOH were detected in 54 out of 78 urine samples. An Acquity UPLC HSS T3 (2.1 x 100 mm, 1.8 µm, Waters, Milford, MA, USA) column was used for chromatographic separation. The mobile phase consisted of 0.05 % acetic acid in water and methanol with gradient elution [79].
For many years, oral fluid has typically been used for drug testing. Different extraction methods were employed for Δ8-THC analysis in oral fluid. In order to avoid strong acidic conditions preventing CBD conversion to Δ8-THC, LC-MS/MS methods using solid phase extraction and solid phase microextraction methods were optimized for the measurement of cannabinoids including Δ8-THC in oral fluid (Table 1) [80,81,82].
An HPLC-MS/MS method was developed for the analysis of Δ8-THC, Δ9-THC, CBD, and 10 additional cannabinoids and their metabolites in oral fluid (including 11-OH-Δ9-THC, Δ9-THC-COOH, THCV, CBDV, cannabidiorcol (CBD-C1), CBC, CBN, and CBG [80]. Baseline separation between Δ8-THC and Δ9-THC peaks was achieved using a CORTECS® Cl8 analytical column at 26°C under a gradient elution of the mobile phase (A) 0.1 % formic acid in water/acetonitrile (95:5, v/v) and (B) 0.1 % formic acid in acetonitrile. In 11 out of the 200 oral fluid samples, Δ8-THC was detected at concentrations ranging from 0.2-339.5 ng/mL. In the same study, two multiple reaction monitoring transitions (MRMs) were monitored for Δ8-THC; a quantifier; 315.1→123.0 (m/z) and a qualifier 315.1→135.1 (m/z), and only one MRM transition was monitored for the deutrated internal standard of Δ8-THC (Δ8-THC-d9); 324.21→123.l (m/z) for the analysis of Δ8-THC. The results showed that only one sample contained Δ8-THC and its concentration was as high as 925.7 ng/mL, suggesting possible mucosa saturation [80].
A 2D Ultra-Fast Liquid Chromatography tandem mass spectrometry (UFLC-MS-MS) method was developed and validated to separate Δ8-THC and Δ9-THC isomers and their metabolites in human blood. A liquid-liquid extraction protocol was used for extracting the target cannabinoids from whole blood. Complete chromatographic separation of Δ8-THC-COOH from Δ9-THC-COOH was achieved over a run time of 10 min. The authors claimed that "this is the first report of a method that successfully quantitates these primary cannabinoids in blood specimens where significant concentrations of both Δ8-THC and Δ9-THC isomers are present" [45].
5.4. Overestimation of Δ9-THC -COOH Levels: A Special Concern
Due to structural similarities, the existence of Δ8-THC-COOH in a urine specimen could potentially negatively interfere with the testing process for Δ9-THC-COOH. Consequently, LC-MS-based analytical methods may experience identification problems with the two isomers, unless they are chromatographically well separated. This is because they have the same molecular mass and Δ8-THC-COOH also showed the same most abundant MS transitions, thereby often meeting the MS acceptance requirements for a positive identification as Δ9-THC-COOH. There have been several studies demonstrating interfering peaks during the detection and/or quantitation of Δ9-THC-COOH by LC-MS/MS or GC-MS [80].
Hadener et al. (2017) expressed special concern about the potential overestimation of Δ9-THC-COOH blood levels due to the existence of its Δ8 isomer, using LC-ESI-MS/MS. This could contribute to inconsistencies in Δ9-THC-COOH concentrations reported by different laboratories [83].
A drug screening by immunoassay is typically the first step in drug testing for both forensic and clinical samples. The cross reactivity immunoassay of Δ8-THC-COOH in the EMIT II phase was assesed at a cutoff of 20 ng/mL. The authors carried out a confirmatory GC-MS method to prevent false positive results of overestimation of Δ9-THC-COOH. Moreover, the reported GC-MS method succeeded in the separation and quantification of both Δ9-THC-COOH and Δ8-THC-COOH as TMS derivatives using relative retention time compared to Δ9-THC-COOH-d9 as the internal standard [84].
A baseline separation of Δ8-THC-COOH and Δ9-THC-COOH peaks has been achieved using GC-MS analysis. Although Δ8-THC-COOH and Δ9-THC-COOH show common fragmentation ions, they were eluted 0.05 min apart, resulting in baseline resolution. The developed method was then applied for the analysis of Δ8-THC-COOH and Δ9-THC-COOH in postmortem urine samples. A total of 26 out of 194 postmortem urine samples were positive for Δ8-THC-COOH with six samples only positive for Δ8-THC-COOH without the presence of Δ9-THC-COOH [85]. A summary of the analysis of Δ8-THC and its metabolites in different biological matrices is displayed in Table 1.

Table 1.
Analysis of Δ8-THC and its metabolites in different biological matrices.
| Matrix | Analytical technique (Analyte) |
Sample preparation | MRMs | Internal Standard (IS) | Reference |
|---|---|---|---|---|---|
| Oral fluid | HPLC-MS/MS (Δ8-THC) |
SPE: I 0 µL of IS+ 400 µL of 2% phosphoric acid+ 400 µL of oral fluid samples, washed with 400 µL of water/methanol (95:5, v/v) and eluted by 400 µL of acetonitrile/methanol (90: 10, v/v) | Quantifier 315.1→123.0 (m/z) a qualifier 315.1→135.1 (m/z) |
d9-Δ8-THC, d3-Δ9-THC, d3-CBD and d3-CBN. d9-Δ8-THC: 324.21→123.1 (m/z) |
[80] |
| Whole blood and serum |
LC-ESI-MS/MS (Δ8-THC-COOH) |
200 µL blood samples+ 20 µL of IS+ 600 µL of ACN, mixing, and centrifugation, evaporated to dryness+ reconstituted in 200 µL of ACN/ H2O/ Formic acid, 60/40/0.1: v/v/v | (m/z, 345→327 (quantifier), 345→299 (qualifier) |
THC-d3 and 11-OH-THC-d3, 10ng, THC-COOH-d3 |
[83] |
| Human urine |
GC/MS (Δ8-THC-COOH) |
after derivatization and Cannabinoid immunoassay |
m/z 488→473, 371 +ve (SIM). |
Δ9-THC-COOH | [84] |
| Postmortem urine |
GC/MS (Δ8-THC-COOH) |
SPE after alkaline hydrolysis and derivatization using BSTFA with I% trimethylchlorosilane |
(Δ9-THC-COOH: m/z, 371,473 and 488; Δ8-THC-COOH: m/z 488, 473, and 432) |
d9-Δ9-THC-COOH: (m/z 374, 476, and 491) |
[85] |
5.5. Stability of Δ8-THC and its metabolites
A recent study on the stability for the analysis of Δ8-THC and its metabolites; 11-OH-Δ8-THC and Δ8-THC-COOH in drug-free urine was reported. The study was carried out at different pHs (4.5, 7, and 9) and temperature conditions (4°C, 20°C and 45°C) for 28 days using an Abbott Architect Plus c4000 Autoanalyzer. The Lin-Zhi enzymatic immunoassay method at the cut-off concentration of 25 ng/mL was used for the analysis. Samples were analyzed daily in the first week and once a week for the remaindar of the study or until the traget analyte below the cut-off concentration of 25 ng/mL [86].
The study revealed that the response of Δ8-THC varied significantly depending on pH and temperature conditions. At 4°C, Δ8-THC instrument response decreased by about 30% over 7 days, irrespective of pH. Furthermore, no notable declines in response observed for Δ8-THC at 4°C, regardless of pH, for the remainder of the study. However, at 20°C, regardless of pH, Δ8-THC instrument response decreased by more than 70% over 7 days. By day 14, Δ8-THC was undetectable at 20°C, regardless of pH. Additionally, Δ8-THC was not detectable after a single day at 45°C, regardless of pH [86].
During the period of 7 days, the instrument responses for 11-OH-Δ8-THC decreased by 10-15%, 15-20%, and 40-60% at 4°C , 20°C, and 45°C, respectively, regardless of pH. However, at 4°C, there was no significant decline in the 11-OH-Δ8-THC instrument response for the remaindar of the study, regardless of pH. By day 14, 11-OH-Δ8-THC was not detectable at pH 4.5 and 45°C. However, at 20°C and 45°C, the instrument responses for 11-OH-Δ8-THC declined by 10-20% and 40-60%, respectively, for the remainder of the study, regardlessof pH. As for Δ8-THC-COOH, at pH=4.5 the instrument response decreased by 40-70% whille at pH=7 and pH=9 the response decreased by 0-5% regardlesss of temperature. For the remainder of the study and at pH=7 and pH= 9, no significant decline in Δ8-THC-COOH response was observed [86].
In another stability study, it was suggested that the effectiveness of behavioural modification in rats might depend on the stability of Δ8-THC. Specifically, when pure Δ8-THC was used, it led to the fastest reduction in avoidance behaviour. On the other hand, when partially deteriorated Δ8-THC was employed, it didn't have a significant impact on avoidance extinction. However, it's important to note that this observation could be influenced by factors like tolerance and the possibility of withdrawal symptoms arising [87]. Inside the human body, Δ8-THC was rapidly and extensively metabolized by the liver into the l1-hydroxy metabolite, shedding light on the assumption that the active form of Δ8-THC may be the 11-hydroxy metabolites [88].
6. Pharmacology of Δ8-THC
6.1. CB1 and CB2 activation
Both cannabinoid binding receptors, CB1 and CB2, are G protein-coupled receptors that are part of the endocannabinoid system interacting and responding to cannabinoids [89]. CB1 receptors are primarily located in the central nervous system and affect functions like mood and appetite. Their activation is associated with the psychoactive effects of cannabis. CB2 receptors mainly constitute a part of the immune system and modulate inflammation and immune cell activity without causing psychoactive effects [90]. Both CB receptors play a role in the body's response to cannabinoids, whether they are produced by the body (endocannabinoids) or those from external sources [91].
Interestingly, Δ8-THC exhibits the ability to competitively attach to the orthosteric sites of both CB1 and CB2 receptors, with Ki values falling within the nanomolar (nM) range. In detail, Govaerts and colleagues documented binding affinities in both murine tissues and cultured cells, that were genetically modified to express human cannabinoid receptors, of 251 nM and 417 nM against CB1 and CB2 receptors, respectively [92]. Husni et al. [32], and Radwan et al. [27] independently reported affinity values of 78 nM and 12 nM against CB1 and CB2 receptors, respectively. On the other hand, Nadipuram and colleagues reported close affinity values of 28.5 nM and 25.0 nM against the same receptors, respectively [93]. Similarly, in their study focused on the synthesis of specific ligands for the CB2 receptor, Huffman’s research team reported an affinity value for Δ8-THC of 44 nM [94]. In contrast, when considering mouse CB1 and CB2 receptors, Δ8-THC exhibited binding affinities, as measured by Ki values, within a range of 39.3 nM to 179 nM [92,95,96,97,98,99,100]. The antiemetic effects of 5 mg/kg and 10 mg/kg Δ8-THC could be counteracted by a CB1 antagonist, but not by a CB2 antagonist. It is important to highlight that Δ8-THC exhibited superior effectiveness in preventing vomiting compared to Δ9-THC, despite its lower affinity for CB1 and CB2 receptors [101].
6.2. Neurotransmitters’ levels and activities
Several manuscripts have investigated how Δ8-THC influences the levels of acetylcholine in order to elucidate its impact on the central nervous system. The effects of Δ8-THC on the hippocampus were found to be dose-dependent, and this was linked to alterations in acetylcholine turnover in the striatum through a stereospecificity-affected turnover [102,103]. Conversely, another study didn’t find convincing proof to suggest any intrinsic pharmacological impact of the Δ8-THC on monoamine oxidase (MAO) activity [104]. Noticeable reductions in dopamine and serotonin concentrations were observed in both the hypothalamus and the hippocampus, respectively. Meanwhile, norepinephrine levels tended to rise in both the hypothalamus and hippocampus [105]. Observable reductions in serotonin concentration were also detected in another study upon the uptake of Δ8-THC [106]. Δ8-THC exposure and stressful environment may lead to increased dopamine reuptake in the striatum, potentially influencing cannabis-induced behaviours under stress [107]. Furthermore, Δ8-THC induced a dual-phase impact on the absorption and discharge of dopamine and norepinephrine in both the corpus striatum and hypothalamic areas of the brain [108]. Sagratella and colleagues undertook several investigations on rabbits to examine the potential role of diminished GABA transmission [109]. Other studies have indicated that Δ8-THC inhibits adenylate cyclase and subsequently cAMP in a dose-dependent manner, though this mechanism is not thought to be the origin of behavioural alterations [110].
6.3. Cognitive functions
When Δ8-THC is administered before a decision-making-based test, it can lead to discriminative behaviour, indicating alterations in a state-dependent manner. The study explored the induction and reversal of memory and learning impairment as a means to differentiate between substances, achieved by altering the 'correct' choice [111]. Experiments conducted by Jarbe et al. at various doses collectively suggest that Δ8-THC enhanced the abilities of rats to pass the T-maze. Individuals subjected to higher doses of Δ8-THC required fewer trials to acquire the maze-learned behaviour [112,113]. This result was in agreement with another study that identified the discriminative stimulation effects of Δ8-THC [114]. However, in contrast, Doty et al. found that rats subjected to phencyclidine (PCP) were responsive to Δ8-THC. Although this didn't directly assess the potential discriminative effect of Δ8-THC itself, these authors did find that Δ8-THC altered the dose–response curve of PCP's discriminative stimulus. The mechanism of this interaction remains unclear and likely involves multiple factors. Interestingly, this is one of the few effects where Δ8-THC did not exhibit a clear dose dependency [115].
Evaluating the effects of Δ8-THC on pigeons suggested a dose-dependent effect of Δ8-THC on pigeons trained with Δ9-THC, thus mirroring similar findings reported in rats [116]. Utilizing the fixed behavioural test-resistance to the extinction of avoidance responses in rats, pure Δ8-THC led to the quickest cessation of avoidance [87]. Examining different behavioural symptoms, Δ8-THC exhibits activity in humans similar to Δ9-THC. When assessed through both oral and intravenous administration, it demonstrates a relative potency ratio of 2:3. Doses ranging from 1 to 6 mg of Δ8-THC intravenously induce a broad spectrum of effects reminiscent of cannabis [33].
6.4. Analgesic and hypothermic activities
In a short-term study, dose-dependent pain relief, and body temperature decrease, were observed in rats after Δ8-THC administration. These effects were noticeable within the range of doses of Δ8-THC and accompanied by complete tolerance [117]. In the vaporized form, employing the tail withdrawal assay in male rats, a mixture containing three parts CBD to one part Δ8-THC produced an immediate analgesic effect when compared to vapor from the control vehicle [118]. In the same study, immediately after exposure, the mixture induced a reduction in body temperature, leading to a hypothermic effect compared to exposure to the control vehicle [118].
Upon oral ingestion, Δ8-THC exhibited an analgesic effect that was dose-dependent when measured by the hot-plate method. Interestingly, this analgesic effect was found to be comparable in potency to Δ9-THC but both were notably less potent than morphine [119]. Conversely, when administered intrathecally rather than orally, Δ8-THC was reported to be less potent than Δ9-THC employing the tail-flick test [120]. In a combination study, to understand the mechanism of analgesic action, while morphine pretreatment did amplify the analgesic effect, administering naloxone to mice beforehand did not impede it. This suggests that the pain-relieving effects brought about by Δ8-THC do not arise from a direct interaction with the opiate receptor [120].
Another study demonstrated the interactions in the use of Δ8-THC and opioids within intracellular systems. The potential shared points of interaction include Gi/o protein-coupled receptors. These receptors lead to a decrease in both calcium entry into neurons and the calcium content. This subsequently results in a reduction of cAMP levels and induces hyperpolarization of neurons. These effects are facilitated through the involvement of both ATP- and apamin-sensitive potassium channels [121]. El-Alfy et al. were able to corroborate and establish statistical significance for an existing, though less potent, dose-dependent antinociceptive effect of Δ8-THC relative to Δ9-THC when assessing for antidepressant-like properties [122]. In a more comprehensive investigation, Welch and his team administered CB1 and CB2 selective antagonist (SR141716A) alongside Δ8-THC using the tail-flick test to uncover any potential analgesic role through the cannabinoid receptors. The authors noted varying degrees of antagonism when switching between routes of administration for both SR141716A and Δ8-THC. This implies a potentially intricate role of cannabinoid receptors involving various subtypes and distinct localization throughout the central nervous system [123].
6.5. Antiepileptic activities
A statistically significant dose-dependent reduction in audiogenic response score, indicating diminished seizure activity, was observed in female rats at all three doses of Δ8-THC administered with an effective dose for complete seizure inhibition of 6.5 mg/kg, exceeding the upper limit of the trial protocol [124]. Another research explored diverse methods including kindling to provoke seizures, a higher intraperitoneal dose of 10 mg/kg of Δ8-THC was required to significantly reduce seizure manifestations, with significant outcomes defined as > 25% protection of the population [125]. In a different experimental animal model, seizures were induced in Senegalese baboons through phototherapy and kindling. Observations revealed dose-dependent prevention and reduction of kindled seizures from electrical stimulation, but no such effect was observed with light-induced seizures after intraperitoneal injections of Δ8-THC [126].
The same research group once again employed kindling and a similar seizure staging model, this time assessing another animal model: cats. In contrast to their previous findings, Δ8-THC demonstrated no significant effect on suppressing or reducing seizure behaviour during any stage of kindling in cats at doses ranging from 0.5–4 mg/kg [127]. An alternative study utilized cobalt-epileptic rats observing the activity at the epileptic focus suggesting that the potential anticonvulsant activity of Δ8-THC may not be consistent across different types of seizures and could potentially exacerbate convulsant activity in seizures arising from other causes [128]. Interestingly, both Δ8-THC and Δ9-THC were determined to have similar relative potency in anticonvulsant activity for PTZ-induced seizures in mice, contrary to previous studies in rats. Additionally, the same study showed that Δ8-THC exhibited an additional protective effect against seizures when combined with phenobarbital, but not with diphenylhydantoin [129].
6.6. Cardiovascular activities
In a short-term study, dose-dependent effects on the reduction of heart rate were observed in rats while complete tolerance to the decrease in heart rate was established within only 13 days [117]. In a conflicting study, Gong and colleagues conducted a three-part experiment examining bronchodilation and tolerance to Δ8-THC. They observed dose-dependent responses. These responses included statistically significant inducible bronchodilation and tachycardia [130]. Smiley KA observed arrhythmia and reduced contractility on perfused rat hearts [131]. A subsequent in vitro study, in rats, investigated changes in contractility by measuring nitric oxide production recording a significant reduction [132]. Another study suggested alterations in the activity of monoamine oxidase enzymes in the hearts of live rats as a potential mechanism for bronchodilation and tachycardia . An early study that compared intravenous and intra-arterial Δ8-THC in rats mentioned a potential role as a peripheral vasoconstrictor with intra-arterial administration, while the intravenous administration led to immediate increases in blood pressure, followed by subsequent drop and a decrease in heart rate [133].
6.7. Gastro-intestinal tract activities
Δ8-THC, when administered orally, led to dose-dependent reductions in intestinal motility in mice. This effect was assessed by monitoring the transit of a charcoal meal through the digestive tract. Specifically, as the dosage of Δ8-THC increased, there was a corresponding decrease in the pace at which the charcoal meal moved through the intestines [119]. In a study targeted the food intake, a very low dose of Δ8-THC (0.001 mg/kg) caused increased food consumption and a tendency to improve cognitive function, without cannabimimetic side effects suggesting that the low dose might serve as a potential therapeutic agent in the treatment of weight disorders [104]. Inversely, rats experienced weight loss, decreased intake, bradycardia, and decreased body temperature after administration of Δ8-THC in another study; however, partial or complete tolerance developed to many of these physiologic changes [130]. Emesis prevention in radiation-treated shrews showed a statistically significant dose-dependent response at single doses of 5 mg/kg and 10 mg/kg of Δ8-THC. The prevention of emesis could be reversed with a CB1, but not with a CB2 antagonist. It's noteworthy that Δ8-THC demonstrated greater efficacy in preventing emesis compared to Δ9-THC, despite having a lower binding affinity for CB1 and CB2 receptors presenting a unique scenario [101]. In another study on emesis prevention using oral Δ8-THC administration every 6 hours for 24 hours in eight children undergoing multiple cycles of antineoplastic regimens for hemolytic malignancies over 8 months, the study found that administering Δ8-THC over 24 hours effectively prevented emesis on the day of treatment and for the subsequent two days [134]. On the other hand, a recent retrospective case report suggests that frequent oral administration of Δ8-THC may be associated with cannabinoid hyperemesis syndrome indicating a paradoxical, dose-dependent, and time-dependent effect. The factors that could be involved in such results include a potential biphasic or antagonist effect at higher doses, in addition to the receptor downregulation and subsequent alterations in the endocannabinoid system [74].
6.8. Anticancer activities
Several scientific in vitro and in vivo studies have explored the potential anti-cancer effects of Δ8-THC. These investigations aim to understand how this compound may impact different kinds of cancer cells and their proliferation. To provide a clear overview of these findings, a selection of studies has been compiled and organized in the following Table 2 for easy reference and analysis.
The diverse array of studies outlined in Table 2 collectively underscores the multifaceted antitumor properties of Δ8-THC. The findings consistently reveal its ability to impede critical processes such as DNA synthesis, cancer cell growth, and cellular respiration across a spectrum of cell types, including murine leukemia cells, human cancer cells, and lymphoma cells. Notably, Δ8-THC demonstrates promise in inhibiting multidrug resistance in certain cancer cells and induces a range of cellular responses, from apoptosis and autophagy to modulation of molecular markers associated with cell cycle progression. Furthermore, in vivo experiments on mice carrying lung adenocarcinoma suggest that Δ8-THC may not only retard tumor growth but also enhance the life span of treated animals. These cumulative results support the potential therapeutic relevance of Δ8-THC in combating cancer through diverse mechanisms, warranting further exploration and clinical investigation.
Table 2.
Anticancer activities of Δ8-THC.
| Examined cells, organs or system | Type of Study | Results | Reference |
|---|---|---|---|
| Mice, Lewis lung cells, L1210 leukemia cells, and bone marrow cells |
In vitro and In vivo | Δ8-THC showed a dosage-dependent reduction in DNA synthesis. | [135] |
| L1210 murine leukemia.. | In vivo | Among the compounds examined, Δ8-THC exhibited the highest potency, with a remarkable 99% inhibition of DNA synthesis. |
[136] |
| Mice and L1210 murine leukemia |
In vivo and in vitro | Δ8--THC didn’t Inhibit cancer cells’ respiration. | [137] |
| Human cells which metabolize polycyclic hydrocarbon carcinogens |
In vitro | Δ8-THC caused a dose-dependent inhibition of cancer cell growth in addition to a dose-dependent inhibition of [3H]thymidine, [3H]uridine and [3H]leucine incorporation. | [138] |
| Neuroblastoma cell membranes |
In vitro | Δ8-THC inhibited adenylate cyclase in plasma membranes | [139] |
| Human mdr1-gene transfected mouse lymphoma cells | In vitro | Δ8-THC exhibited membrane-associated antitumor effects and reversal of multidrug resistance. |
[140] |
| Human oral Tu183 cancer cells | In vitro | Δ8-THC exhibited dose-dependent potent inhibition against cancer cellular respiration |
[141] |
| Human oral cancer cell | In vitro | Δ8-THC promoted apoptosis and autophagy. Furthermore, it hindered cell migration and invasion. It decreased the production of reactive oxygen and increased levels of glutathione and its expression. it downregulated the expressions of cyclin D1, p53, NOXA, PUMAα, and DRAM, but upregulated the expressions of p21 and H2AX. |
[142] |
| Mice carrying Lewis lung carcinoma |
In vivo | Δ8-THC led to a dose-dependent retardation of tumor growth. Δ8-THC increased the life span of the treated mice and decreased primary tumor size. | [143,144] |
6.9. Immunomodulatory activities
Numerous scientific investigations have focused on understanding the immunomodulatory properties of Δ8-THC. These studies aim to unravel how this compound can influence the immune system. To facilitate a comprehensive grasp of these findings, a subset of these studies has been thoughtfully consolidated and presented in the forthcoming Table 3 for easy reference and analysis.
Table 3.
Immunomodulatory activities of Δ8-THC.
| Examined cells, organs, or system |
Study Type | Results | Reference |
|---|---|---|---|
| Lymphocytes | In vitro | Δ8-THC led to a dose-dependent inability of lymphocytes to integrate the [3H] thymidine. |
[145] |
| T and B lymphocyte | In vitro | Δ8-THC inhibited the mitogen-induced T and B lymphocyte proliferation. |
[146] |
| Mouse macrophage J774-1 cells |
In vitro | Δ8-THC induced cell death of J774-1 cells in a concentration- and/or exposure time-dependent manner. Associated with vacuole formation, chromatin condensation, cell swelling, and nuclear fragmentation. |
[147] |
| BALB/c mice, Plasma and Spleen |
In vivo | Δ8-THC demonstrated a notable inhibition of direct hemolytic plaque-forming cells in the spleen on day 4. |
[148] |
| Guinea pig, Skin | In vivo | moderate (Grade III) sensitizers, causing allergic contact dermatitis. |
[149] |
| Rats, Brain | In vivo | Suppression of autoimmune encephalomyelitis by Δ8-THC was noticed and attributed to its influence on the secretion of corticosterone. |
[150] |
| Rats, Brain | In vivo | Δ8-THC reduced lever-pressing behaviour by activating the arachidonic acid cascade, leading to an elevated production of prostaglandin E2 in the brain through the CB1 receptor. |
[151] |
| Rats, Brain | In vivo | Δ8-THC suppressed lever-pressing behaviour through the activation of the prostanoid EP3 receptor by elevation of prostaglandin E2. |
[152] |
| Human B-lymphoblastoid and mouse fibroblast cell | In vitro | Δ8-THC showed antioxidative effect by preventing serum-deprived cell death that induced by anhydroretinol. | [153] |
In summary, the diverse array of studies examining the effects of Δ8-THC across different cells, organs, and systems underscores its multifaceted immunomodulatory impact. Δ8-THC demonstrates a range of outcomes, from inhibiting lymphocyte proliferation and inducing cell death in macrophages to influencing immune responses in vivo. Notably, it exhibits both immunosuppressive effects, as observed in the reduction of hemolytic plaque-forming cells and the suppression of autoimmune encephalomyelitis, and modulatory effects on behavior, evidenced by the regulation of lever-pressing behavior through intricate signaling pathways in the brain. Additionally, Δ8-THC appears to antagonize oxidative stress, suggesting potential protective effects on B lymphocytes and fibroblasts. The presented findings collectively highlight the complex and context-dependent nature of Δ8-THC’s interactions within the biological systems studied, emphasizing the need for further research to elucidate its therapeutic immunomodulatory potential and potential adverse effects.
6.10. Ocular activities
The mechanism of Δ8-THC in corneal injury treatment was found to be mediated by cannabinoid receptors, as a study observed reduced pain and inflammation after corneal injury in mice with topical administration of Δ8-THC [154]. Interstingly, additional lipid-based vehicles for topical preparation of different Δ8-THC-containing topical formulations enhanced corneal penetration and minimized systemic absorption [155]. Similar results were obtained after the application of a submicron emulsion as an ocular vehicle for Δ8-THC as a treatment for intraocular pressure in rabbits [156]. An early study involving eight rabbits observed a decrease in the intraocular pressure following intravenous administration of Δ8-THC, but this effect was not dose-dependent [157]. In another study, topical Δ8-THC did eventually lead to a reduction in intraocular pressure in rabbits [158]. Further studies by the same team indicated that the effect was likely not centrally mediated or a result of the drug crossing the blood–brain barrier [159].
6.11. Locomotor activities
In the vaporized form, a concentration of 10 mg of Δ8-THC initially induced hyperlocomotion in male rats within the first 30 minutes of the session, followed by a hypo locomotor effect later in the session [118]. In mice, both Δ8-THC and Δ9-THC demonstrated a notable decrease in spontaneous activity, leading to reduced awareness. They also caused a loss of muscle coordination, diminished sensorimotor responses, and elicited a hunched posture and gait reminiscent of the effects induced by narcotic analgesics. Additionally, when administered intravenously, both Δ8-THC and Δ9-THC exhibited a quicker onset of activity compared to intracerebral administration [88].
6.12. Fertility affecting activities
In an investigation into its anti-androgenic properties, Δ8-THC demonstrated remarkable effectiveness as an inhibitor of mitochondrial O2 consumption within human sperm cells. This finding underscores the significant impact that Δ8-THC can exert on the metabolic processes crucial for sperm function and reproductive physiology [160]. In another study, rats that were exposed chronically to Δ8-THC displayed significantly reduced body weight and lower levels of follicle-stimulating hormone and luteinizing hormone in comparison to the control group. However, it is worth noting that these hormone levels did recover once the drug was discontinued. The precise clinical implications of these effects on fertility remain uncertain and warrant further investigation [161].
6.13. Antidepressant activities
The antidepressant effects of Δ8-THC were evaluated, using the automated mouse-forced swim and tail suspension tests. Interestingly, doses of 1.25, 2.5, and 5 mg/kg administered intraperitoneally demonstrated a U-shaped dose-response in terms of their antidepressant action [122].
6.14. Toxicity of Δ8-THC
Following the receipt of 104 reports detailing adverse events in individuals who had consumed Δ8-THC products between December 1, 2020, and February 28, 2022, the Food and Drug Administration (FDA) released a report underscoring the potential hazards linked with Δ8-THC products and emphasized the necessity for regulatory oversight and public awareness. The FDA report highlights five major significant concerns regarding Δ8-THC products [38]. Firstly, these products have not undergone FDA assessment or approval for safe usage, potentially posing risks to public health. There are worries about inconsistent formulations, incorrect labelling, and varying Δ8-THC levels. Some products may be misleadingly labelled as "hemp products," potentially leading consumers to underestimate their psychoactive effects. Secondly, the report expresses apprehension about products claiming therapeutic benefits without FDA approval. This could endanger consumers, as the safety and efficacy of such products have not been confirmed. The data presented indicates possible adverse events, particularly among pediatric patients, emphasizing the need for caution. Thirdly, Δ8-THC shares similar psychoactive effects with Δ9-THC, indicating a comparable level of impairment. Fourthly, concerns are raised about the manufacturing process, which may involve potentially harmful chemicals and lead to contaminants in the final product. Finally, manufacturers' packaging Δ8-THC products in ways appealing to children is a notable concern, as this could lead to unintentional exposure [38].
On September 14, 2020, the Centers for Disease Control and Prevention (CDC) announced an alert, warning healthcare workers and the public about the recent rise in adverse health events related to Δ8-THC consumption. It highlights that emergency room visits mentioning Δ8-THC as the primary complaint were mostly concentrated in southern states without legal adult-use marijuana access. In these cases, Δ8-THC products are often marketed as hemp products, potentially leading to accidental intoxication due to consumer confusion. The CDC advises caution, noting that Δ8-THC products can be intoxicating, may have misleading labels, and should be kept away from children [162].
In the same context, chemists expressed serious concerns over impurities in Δ8-THC, as many Δ8-THC products available may not be entirely pure, containing other cannabinoids in addition to uncertain by-products with unclear health effects. The lack of oversight and testing raises safety worries. Regulators are struggling to manage Δ8-THC’s sales and chemists stress the importance of enhanced regulation and transparency in production. They also caution against potential risks stemming from unregulated synthesis methods [163]. In light of this, the findings of a recent study combining techniques such as NMR, HPLC, and MS revealed that the tested products contain not only 10 different Δ8-THC related structures but also several impurities, present in concentrations significantly higher than what is stated on the certificates of analysis for these products [67].
A recent investigation into self-reported effects linked to Δ8-THC revealed similarities to those reported during acute cannabis intoxication. The most commonly reported adverse events were related to psychiatric disorders, followed by respiratory, thoracic, and mediastinal issues, as well as nervous system problems. The most frequently cited symptoms in these reports included feelings of anxiety, coughing, and paranoia [164]. This study supported a prior investigation that detailed cases of two individuals, aged 19 and 20, experiencing acute psychiatric distress after consuming a Δ8-THC product. These cases demonstrate a conceivable link in time between the consumption of products containing Δ8-THC and the emergence of manic or psychotic symptoms, implying a potential relationship between dosage and response [165]. In pediatric cases, the clinical manifestations resulting from exposure to Δ8-THC encompass a spectrum of effects, which extend beyond but are not confined to alterations in mental status. These include presentations such as episodes reminiscent of seizures and irregularities in vital signs [166].
In an early assessment of tolerance to Δ8-THC, after chronic administration and mice sacrifice, the livers demonstrated significant increases in tyrosine aminotransferase activity with acute administration; however, little-to-no increased activity was noted after 8 and 12 weeks of chronic administration of Δ8-THC [167]. Additionally, the Birmingham School of Medicine has reported recently a notable surge in the frequency of emergency department admissions, both in their own hospital and in several others, attributed to the utilization of Δ8-THC [168]. In a study on the antiemetic effects of oral Δ8-THC in eight children undergoing hemolytic malignancies over an 8-month period, the authors observed minimal incidence of anxiolytic or psychotropic effects, taking into consideration the challenges in both identifying and self-reporting these symptoms in young children [134].
6.15. Tolerance
It is worth noting that tolerance to the biological effects of Δ8-THC has been a recurring observation in several scientific publications. These studies have consistently pointed out that over time, the response to Δ8-THC can undergo significant alterations. This phenomenon has been extensively documented, shedding light on the complex nature of how the body interacts with this compound. Mice became completely tolerant to the hypothermic effects and partially tolerant to extended phenobarbital-induced sleeping times and catalepsy within 38 days of daily intravenous Δ8-THC administration [169,170]. Moreover, Δ8-THC -tolerant mice were cross-tolerant to the body temperature-lowering effects of chlorpromazine but not to morphine or pentobarbital [171,172]. When considering morphine interactions, a study did find that morphine-tolerant mice treated with Δ8-THC induced heightened catalepsy rather than cross-tolerance [173]. Furthermore, the development of cross-tolerance to the prolongation of pentobarbital-induced sleep by Δ8-tetrahydrocannabinol and 11-hydroxy- Δ8-tetrahydrocannabinol was reported in mice [170]. In a study exploring the reducing power of Δ8-THC against seizure manifestations, chronic administration of intraperitoneal 15 mg/kg Δ8-THC rapidly led to tolerance in preventing seizure activity. Only two intraperitoneal injections were required to induce tolerance [125]. At elevated levels of Δ8-THC, the ability to counteract epileptic activity seems to diminish in Senegalese baboons following neural discharge at the targeted stimulation site [126]. In a short-term study regarding the cardiovascular system, a relationship between the administered dose (2 mg/kg) of Δ8-THC and a decrease in heart rate was evident. Notably, a complete tolerance to the heart rate reduction occurred in just 13 days [117]. The immunomodulatory effects of 60 mg/kg of Δ8-THC against direct haemolytic plaque-forming cells in the mice spleen showed a state of reduced responsiveness (hyporesponsiveness) developed when a pretreatment regimen was employed [148].
6.16. Withdrawal activities
Some research articles have focused on the potential of Δ8-THC withdrawal. In general, the decrease in exploratory behaviour with short-term and long-term administration of Δ8-THC supports the CNS depressant effects. Significant changes in rat’s defecation (potentially indicative of mood changes) and grooming behaviours (potentially indicative of withdrawal symptoms) were noted [180]. In morphine-dependent mice, increased jumping and forepaw tremors upon administration of naloxone were considered consistent with withdrawal. The administration of Δ8-THC was able to inhibit these behavioural modifications. Also, high doses of Δ8-THC induced hyperactivity in morphine-dependent mice rather than ataxia/catalepsy [174].
The upregulation of the CB1 receptors coupled with the mitigation of withdrawal symptoms with CB1 stimulation indicates that dysregulation of the endocannabinoid system may have a role in opiate-induced withdrawal [174]. A contradictory study showed that a single administration of 10 mg/kg of Δ8-THC was insufficient to suppress naloxone-induced morphine withdrawal [175]. Abrupt cessation in rats after chronic administration of Δ8-THC for 30 days increased spontaneous yawning in a similar way to that of opioid withdrawal [176]. On the other hand, the acute administration portion of the study identified that Δ8-THC reduced yawning induced by dopaminergic and cholinergic agonists [176]. Further investigations into the potential dopaminergic modulation of Δ8-THC may clarify any potential for inducing overuse-associated behaviours.
Modulation of the arachidonic acid pathway and the stimulation of the prostaglandin EP3 receptors may also be involved in dependence and withdrawal [176]. Furthermore, with this induced prostaglandin synthesis, the rats’ brain reduces natural production and becomes dependent on THC for prostaglandin maintenance [176]. The injection of 10 mg/kg diclofenac, an inhibitor of prostaglandin synthesis, intraperitoneally 30 minutes before the synthetic CB1 antagonist, SR 141716A, induced withdrawal symptoms in mice that were naïve to cannabinoids. These signs closely resembled those of Δ8-THC withdrawal symptoms. Conversely, the administration of 10 mg/kg Δ8-THC i.p., 15 minutes before SR 141716A, prevented the manifestation of these withdrawal signs. These findings imply that Abrupt cessation of Δ8-THC may cause a decrease in prostaglandin levels that contributes to the withdrawal syndrome [177]. Anggadiredj and colleagues' study suggested that the potential for Δ8-THC to provoke withdrawal symptoms is not yet well understood and warrants further research [177].
7. Conclusion
This review has provided a comprehensive exploration of the chemical, analytical, and pharmacological aspects of Δ8-THC. The review shed light on its emergence as a psychoactive cannabinoid with increased popularity, driven by its superior stability and the easy synthetic procedure compared to the more well-known Δ9-THC. The paper delved into the methods of extraction, purification, and structure elucidation of Δ8-THC, addressing the crucial question of whether it is a natural compound or an artifact. Additionally, strategies for the chemical synthesis and analyses of Δ8-THC were presented, along with considerations of potential impurities associated with synthetic manufacturing. The discussion on methods of analysis and detection of impurities in marketed products serves as a valuable guide for researchers, industry professionals, and regulatory bodies. The pharmacological effects of Δ8-THC were thoroughly discussed, focusing on its interaction with CB1 and CB2 cannabinoid receptors as well as other physiological targets. This in-depth analysis offers a comprehensive understanding of the specific effects elicited by Δ8-THC. As the interest in Δ8-THC continues to grow, this review may serve as a valuable resource for researchers, practitioners, and policymakers in cannabis-related fields. Further research and ongoing scrutiny are warranted to better understand the compound's properties, implications, and potential applications.
Author Contributions
All authors contributed to the writing, preparation, review and editing of this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Prince Sattam Bin Abdulaziz University for supporting the study via the project number (PSAU/2023/01/009).
Data Availability Statement
Not applicable
Acknowledgments
M.S.A.-K. is thankful to Prince Sattam Bin Abdulaziz University for supporting the study via the project number (PSAU/2023/01/009).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kriese, U.; Schumann, E.; Weber, W.; Beyer, M.; Brühl, L.; Matthäus. Oil content, tocopherol composition and fatty acid patterns of the seeds of 51 Cannabis sativa L. genotypes. Euphytica 2004, 137, 339–351. [Google Scholar] [CrossRef]
- Doyle, E.; Spence, A. Cannabis as a medicine? Br. J. Anaesth. 1995, 74, 359–361. [Google Scholar] [PubMed]
- Zuardi, A.W. History of cannabis as a medicine: a review. Braz. J. Psychiatry. 2006, 28, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Crocq, M.-A. History of cannabis and the endocannabinoid system. Dialogues Clin. Neurosci. 2020, 22, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Gaoni, Y.; Mechoulam, R. Isolation, structure, and partial synthesis of an active constituent of hashish. J. Am. Chem. Soc. 1964, 86, 1646–1647. [Google Scholar] [CrossRef]
- ElSohly, M.A.; Radwan, M.M.; Gul, W.; Chandra, S.; Galal, A. Phytochemistry of Cannabis sativa L. In Phytocannabinoids: Unraveling the Complex Chemistry and Pharmacology of Cannabis sativa; Kinghorn, A.D., Falk, H., Gibbons, S., Kobayashi, J.i., Eds.; Springer International Publishing: Cham, 2017; pp. 1–36. [Google Scholar]
- Turner, C.E.; Elsohly, M.A.; Boeren, E.G. Constituents of Cannabis sativa L. XVII. A review of the natural constituents. J. Nat. Prod. 1980, 43, 169–234. [Google Scholar] [CrossRef] [PubMed]
- Radwan, M.M.; Chandra, S.; Gul, S.; ElSohly, M.A. Cannabinoids, phenolics, terpenes and alkaloids of cannabis. Molecules 2021, 26, 2774. [Google Scholar] [CrossRef]
- Pertwee, R.G. Pharmacological Actions of Cannabinoids. In Cannabinoids, Pertwee, R.G., Ed.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2005; pp. 1–51. [Google Scholar]
- Maccarrone, M. Missing pieces to the endocannabinoid puzzle. Trends Mol. Med. 2020, 26, 263–272. [Google Scholar] [CrossRef]
- Alves, P.; Amaral, C.; Teixeira, N.; Correia-da-Silva, G. Cannabis sativa: Much more beyond Δ9-tetrahydrocannabinol. Pharmacol. Res. 2020, 157, 104822. [Google Scholar] [CrossRef]
- Preet, A.; Ganju, R.; Groopman, J. Δ9-Tetrahydrocannabinol inhibits epithelial growth factor-induced lung cancer cell migration in vitro as well as its growth and metastasis in vivo. Oncogene 2008, 27, 339–346. [Google Scholar] [CrossRef]
- Caffarel, M.M.; Andradas, C.; Mira, E.; Pérez-Gómez, E.; Cerutti, C.; Moreno-Bueno, G.; Flores, J.M.; García-Real, I.; Palacios, J.; Mañes, S. Cannabinoids reduce ErbB2-driven breast cancer progression through Akt inhibition. Mol. Cancer 2010, 9, 1–11. [Google Scholar] [CrossRef]
- Laezza, C.; Pagano, C.; Navarra, G.; Pastorino, O.; Proto, M.C.; Fiore, D.; Piscopo, C.; Gazzerro, P.; Bifulco, M. The endocannabinoid system: A target for cancer treatment. Int. J. Mol. Sci. 2020, 21, 747. [Google Scholar] [CrossRef]
- Pagano, C.; Navarra, G.; Coppola, L.; Bifulco, M.; Laezza, C. Molecular mechanism of cannabinoids in cancer progression. Int. J. Mol. Sci. 2021, 22, 3680. [Google Scholar] [CrossRef]
- Institute of Medicine (US). Development of Cannabinoid Drugs. In Marijuana and medicine: Assessing the science base, Joy JE, W.S.J., Benson JA Jr.,, Eds.; National Academies Press Washington (DC), USA, 1999; Volume 1999; pp. 146-168.
- Zeiger, J.S.; Haberstick, B.C.; Corley, R.P.; Ehringer, M.A.; Crowley, T.J.; Hewitt, J.K.; Hopfer, C.J.; Stallings, M.C.; Young, S.E.; Rhee, S.H. Subjective effects to marijuana associated with marijuana use in community and clinical subjects. Drug Alcohol Depend. 2010, 109, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Crean, R.D.; Crane, N.A.; Mason, B.J. An evidence based review of acute and long-term effects of cannabis use on executive cognitive functions. J. Addict. Med. 2011, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Curran, V.H.; Brignell, C.; Fletcher, S.; Middleton, P.; Henry, J. Cognitive and subjective dose-response effects of acute oral Δ 9-tetrahydrocannabinol (THC) in infrequent cannabis users. Psychopharmacology 2002, 164, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Serrano, M.J.; Pérez-García, M.; Verdejo-García, A. What are the specific vs. generalized effects of drugs of abuse on neuropsychological performance? Neurosci. Biobehav. Rev. 2011, 35, 377–406. [Google Scholar] [CrossRef] [PubMed]
- D'Souza, D.C.; Perry, E.; MacDougall, L.; Ammerman, Y.; Cooper, T.; Wu, Y.-t.; Braley, G.; Gueorguieva, R.; Krystal, J.H. The psychotomimetic effects of intravenous delta-9-tetrahydrocannabinol in healthy individuals: implications for psychosis. Neuropsychopharmacology 2004, 29, 1558–1572. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.M.; Di Forti, M. Cannabis and psychosis: what degree of proof do we require? Biol. Psychiatry 2016, 79, 514–515. [Google Scholar] [CrossRef]
- Grotenhermen, F. Review of unwanted actions of cannabis and THC. In Cannabis and cannabinoids. Pharmacology, toxicology, and therapeutic potential; Russo, E.B., Grotenhermen, Franjo, Eds.; Haworth Press: New York, 2002; pp. 233–247. [Google Scholar]
- Leweke, F.M. Acute effects of cannabis and the cannabinoids. In Cannabis and Cannabinoids. Pharmacology, Toxicology and Therapeutic Potential; Russo, E.B., Grotenhermen, Franjo, Eds.; The Haworth Integrative Healing Press: New York, 2002; pp. 249–256. [Google Scholar]
- Hively, R.L.; Mosher, W.A.; Hoffmann, F.W. Isolation of trans-Δ6-tetrahydrocannabinol from marijuana. J. Am. Chem. Soc. 1966, 88, 1832–1833. [Google Scholar] [CrossRef] [PubMed]
- Hanuš, L.; Krejcí, Z. Isolation of two new cannabinoid acids from Cannabis sativa L. of Czechoslovak origin. Acta Univ Olomuc, Fac Med 1975, 74, 161–166. [Google Scholar]
- Radwan, M.M.; ElSohly, M.A.; El-Alfy, A.T.; Ahmed, S.A.; Slade, D.; Husni, A.S.; Manly, S.P.; Wilson, L.; Seale, S.; Cutler, S.J. Isolation and pharmacological evaluation of minor cannabinoids from high-potency Cannabis sativa. J. Nat. Prod. 2015, 78, 1271–1276. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Ross, S.A.; Slade, D.; Radwan, M.M.; Khan, I.A.; ElSohly, M.A. Minor oxygenated cannabinoids from high potency Cannabis sativa L. Phytochemistry 2015, 117, 194–199. [Google Scholar] [CrossRef]
- Hanuš, L.O.; Meyer, S.M.; Muñoz, E.; Taglialatela-Scafati, O.; Appendino, G. Phytocannabinoids: a unified critical inventory. Nat. Prod. Rep. 2016, 33, 1357–1392. [Google Scholar] [CrossRef]
- Mechoulam, R.; Braun, P.; Gaoni, Y. Stereospecific synthesis of (-)-. DELTA. 1-and (-)-. DELTA. 1 (6)-tetrahydrocannabinols. J. Am. Chem. Soc. 1967, 89, 4552–4554. [Google Scholar] [CrossRef]
- Zlas, J.; Stark, H.; Seligman, J.; Levy, R.; Werker, E.; Breuer, A.; Mechoulam, R. Early medical use of cannabis. Nature 1993, 363, 215–215. [Google Scholar] [CrossRef]
- Husni, A.S.; McCurdy, C.R.; Radwan, M.M.; Ahmed, S.A.; Slade, D.; Ross, S.A.; ElSohly, M.A.; Cutler, S.J. Evaluation of phytocannabinoids from high-potency Cannabis sativa using in vitro bioassays to determine structure–activity relationships for cannabinoid receptor 1 and cannabinoid receptor 2. Med. Chem. Res. 2014, 23, 4295–4300. [Google Scholar] [CrossRef]
- Hollister, L.E.; Gillespie, H. Delta-8-and delta-9-tetrahydrocannabinol; Comparison in man by oral and intravenous administration. Clin. Pharmacol. Ther. 1973, 14, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Stern, E.; Lambert, D.M. Medicinal chemistry endeavors around the phytocannabinoids. Chem. Biodivers. 2007, 4, 1707–1728. [Google Scholar] [CrossRef] [PubMed]
- Leas, E.C.; Nobles, A.L.; Shi, Y.; Hendrickson, E. Public interest in∆ 8-Tetrahydrocannabinol (delta-8-THC) increased in US states that restricted∆ 9-Tetrahydrocannabinol (delta-9-THC) use. Int. J. Drug Policy 2022, 101, 103557. [Google Scholar] [CrossRef] [PubMed]
- Babalonis, S.; Raup-Konsavage, W.M.; Akpunonu, P.D.; Balla, A.; Vrana, K.E. Δ8-THC: legal status, widespread availability, and safety concerns. Cannabis Cannabinoid Res. 2021, 6, 362–365. [Google Scholar]
- Radwan, M.M.; Wanas, A.S.; Gul, W.; Ibrahim, E.A.; ElSohly, M.A. Isolation and Characterization of Impurities in Commercially Marketed Δ8-THC Products. J. Nat. Prod. 2023, 86, 822–829. [Google Scholar] [CrossRef]
- The U.S. Food and Drug Administration. 5 Things to know about Delta-8-Tetrahydrocannabinol-Delta-8-THC. Available online: https://www.fda.gov/consumers/consumer-updates/5-things-know-about-delta-8-tetrahydrocannabinol-delta-8-thc (accessed on November 19).
- Adams, R. Marihuana: harvey lecture, February 19, 1942. Bull N Y Acad Med 1942, 18, 705. [Google Scholar]
- Krejcí, Z.; Šantavý, F. Isolation of two new cannabinoid acids from Cannabis sativa L. of Czechoslovak origin. Acta Univ Olomuc, Fac Med 1975, 74, 161–166. [Google Scholar]
- Pars, H.G.; Razdan, R.K. Tetrahydrocannabinols and synthetic analogs. Ann. N. Y. Acad. Sci. 1971, 191, 15–22. [Google Scholar] [CrossRef]
- Turner, C.E.; Hadley, K.W.; Fetterman, P.S.; Doorenbos, N.J.; Quimby, M.W.; Waller, C. Constituents of Cannabis sativa L. IV: Stability of cannabinoids in stored plant material. J. Pharm. Sci. 1973, 62, 1601–1605. [Google Scholar] [CrossRef] [PubMed]
- Turner, C. , Hadley KW, Davis KH,. Constituents of Cannabis sativa L. V: Stability of an analytical sample extracted with chloroform. Acta Pharm. Jucoslavica 1973, 23, 89–94. [Google Scholar]
- Rzeppa, S.; Große, J.; Rautenberg, C.; Thieme, D.; Lepp, T.; Vallimäe, H.; Keiler, A.M. Emergence of the less common cannabinoid Δ8-Tetrahydrocannabinol in a doping sample. Drug Test Anal. 2021, 13, 1936–1943. [Google Scholar] [CrossRef] [PubMed]
- Chan-Hosokawa, A.; Nguyen, L.; Lattanzio, N.; Adams, W.R. Emergence of delta-8 tetrahydrocannabinol in DUID investigation casework: method development, validation and application. J. Anal. Toxicol. 2022, 46, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Leas, E.C. The hemp loophole: a need to clarify the legality of delta-8-THC and other hemp-derived tetrahydrocannabinol compounds. Am. J. Public Health 2021, 111, 1927–1931. [Google Scholar] [CrossRef] [PubMed]
- Marzullo, P.; Foschi, F.; Coppini, D.A.; Fanchini, F.; Magnani, L.; Rusconi, S.; Luzzani, M.; Passarella, D. Cannabidiol as the substrate in acid-catalyzed intramolecular cyclization. J. Nat.Prod. 2020, 83, 2894–2901. [Google Scholar] [CrossRef]
- Gaoni, Y.; Mechoulam, R. Hashish—VII: The isomerization of cannabidiol to tetrahydrocannabinols. Tetrahedron 1966, 22, 1481–1488. [Google Scholar] [CrossRef]
- Razdan, R.K. Chemistry and structure-activity relationships of cannabinoids: An overview. In The cannabinoids: Chemical, pharmacologic, and therapeutic aspects; Agurell, S., Dewey, W.L., Willette, R.E., Eds.; Academic Press: New York, U.S.A., 1984; pp. 63–78. [Google Scholar]
- Pertwee, R.G.; Cascio, M.G. Known pharmacological actions of delta-9-tetrahydrocannabinol and of four other chemical constituents of cannabis that activate cannabinoid receptors; Oxford University Press: Oxford, UK, 2014; Volume 115, pp. 137–156. [Google Scholar]
- Masoud, A.; Wingard, D. High performance liquid chromatography with electrochemical detection. 1: Separation of Cannabis constituents and quantitation of Δ9 tetrahydrocannabinol. J. High Resol. Chromatogr. 1979, 2, 118–122. [Google Scholar] [CrossRef]
- Correia, B.; Ahmad, S.M.; Quintas, A. Determination of phytocannabinoids in cannabis samples by ultrasound-assisted solid-liquid extraction and high-performance liquid chromatography with diode array detector analysis. J. Chromatogr. A. 2023, 1705, 464191. [Google Scholar] [CrossRef] [PubMed]
- Elhendawy, M.A.; Radwan, M.M.; Ibrahim, E.A.; Wanas, A.S.; Chandra, S.; Godfrey, M.; ElSohly, M.A. Validation and Quantitation of Fifteen Cannabinoids in Cannabis and Marketed Products Using High-Performance Liquid Chromatography-Ultraviolet/Photodiode Array Method. Cannabis Cannabinoid Res. 2023. [Google Scholar] [CrossRef]
- Duffy, B.C.; Li, L.; Lu, S.; Dittmar, M.A.; Delaney-Baldwin, E.; Durocher, L.A.; Spink, D.C. Chemotyping of∆ 8-THC-Containing e-Liquids Analyzed during the 2019–2020 New York State EVALI Investigation. J. Anal. Toxicol. 2022, 46, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Avula, B.; ElSohly, M.A.; Radwan, M.M.; Wang, M.; Wanas, A.S.; Mehmedic, Z.; Khan, I.A. Quantitative determination of Δ9-THC, CBG, CBD, their acid precursors and five other neutral cannabinoids by UHPLC-UV-MS. Planta Med. 2018, 84, 260–266. [Google Scholar] [CrossRef]
- Križman, M. A simplified approach for isocratic HPLC analysis of cannabinoids by fine tuning chromatographic selectivity. Eur. Food Res. Technol. 2020, 246, 315–322. [Google Scholar] [CrossRef]
- Song, L.; Carlson, S.; Valenzuela, G.; Chao, M.; Pathipaka, S.B. Development of a validated method for rapid quantification of up to sixteen cannabinoids using ultra-high-performance liquid chromatography diode-array detector with optional electrospray ionization time-of-flight mass spectrometry detection. J. Chromatogr. A. 2022, 1670, 462953. [Google Scholar] [CrossRef]
- Madden, O.; Walshe, J.; Kishore Patnala, P.; Barron, J.; Meaney, C.; Murray, P. Phytocannabinoids-An Overview of the Analytical Methodologies for Detection and Quantification of Therapeutically and Recreationally Relevant Cannabis Compounds. Critical Reviews in Analytical Chemistry 2023, 53, 211–231. [Google Scholar] [CrossRef]
- Christinat, N.; Savoy, M.-C.; Mottier, P. Development, validation and application of a LC-MS/MS method for quantification of 15 cannabinoids in food. Food Chem. 2020, 318, 126469. [Google Scholar] [CrossRef] [PubMed]
- Berman, P.; Futoran, K.; Lewitus, G.M.; Mukha, D.; Benami, M.; Shlomi, T.; Meiri, D. A new ESI-LC/MS approach for comprehensive metabolic profiling of phytocannabinoids in Cannabis. Sci. Rep. 2018, 8, 14280. [Google Scholar] [CrossRef] [PubMed]
- Trofin, I.G.; Vlad, C.C.; Noja, V.V.; Dabija, G. Identification and characterization of special types of herbal cannabis. UPB Sci. Bull., Ser. B 2012, 74, 119–130. [Google Scholar]
- United States H.R.2. Agriculture Improvement Act of 2018. In: 115th United States Congress. Available online: https://www.congress.gov/bill/115th-congress/house-bill/2 (accessed on December 6).
- Drug Enforcement Administration (DEA), D.o.J. Implementation of the Agriculture Improvement Act of 2018. Available online: https://www.federalregister.gov/documents/2020/08/21/2020-17356/implementation-of-the-agriculture-improvement-act-of-2018 (accessed on November 20).
- Razdan, R.K.; Dalzell, H.C.; Handrick, G.R. Hashish. X. Simple one-step synthesis of (-)-. DELTA. 1-tetrahydrocannabinol (THC) from p-mentha-2, 8-dien-1-ol and olivetol. J. Am. Chem. Soc. 1974, 96, 5860–5865. [Google Scholar] [CrossRef]
- Malkov, A.V.; Kočovský, P. Tetrahydrocannabinol revisited: Synthetic approaches utilizing molybdenum catalysts. Collect. Czechoslov. Chem. Commun. 2001, 66, 1257–1268. [Google Scholar] [CrossRef]
- Meehan-Atrash, J.; Rahman, I. Novel Δ8-tetrahydrocannabinol vaporizers contain unlabeled adulterants, unintended byproducts of chemical synthesis, and heavy metals. Chem. Res. Toxicol. 2021, 35, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Ray, C.L.; Bylo, M.P.; Pescaglia, J.; Gawenis, J.A.; Greenlief, C.M. Delta-8 tetrahydrocannabinol product impurities. Molecules 2022, 27, 6924. [Google Scholar] [CrossRef]
- Ciolino, L.A.; Ranieri, T.L.; Brueggemeyer, J.L.; Taylor, A.M.; Mohrhaus, A.S. EVALI vaping liquids part 1: GC-MS cannabinoids profiles and identification of unnatural THC isomers. Front. Chem. 2021, 9, 746479. [Google Scholar] [CrossRef]
- Burstein, S.H.; Menezes, F.; Williamson, E.; MECHOULAM, R. Metabolism of Δ1 (6)-Tetrahydro-cannabinol, an active marihuana constituent. Nature 1970, 225, 87–88. [Google Scholar] [CrossRef]
- Watanabe, K.; Yamaori, S.; Funahashi, T.; Kimura, T.; Yamamoto, I. Cytochrome P450 enzymes involved in the metabolism of tetrahydrocannabinols and cannabinol by human hepatic microsomes. Life Sci 2007, 80, 1415–1419. [Google Scholar] [CrossRef]
- Huestis, M.A. Human cannabinoid pharmacokinetics. Chem. Biodivers. 2007, 4, 1770. [Google Scholar] [CrossRef] [PubMed]
- Tagen, M.; Klumpers, L.E. Review of delta-8-tetrahydrocannabinol (Δ8-THC): Comparative pharmacology with Δ9-THC. Br. J. Pharmacol. 2022, 179, 3915–3933. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Yamamoto, I.; Oguri, K.; Yoshimura, H. Metabolic disposition of delta 8-tetrahydrocannabinol and its active metabolites, 11-hydroxy-delta 8-tetrahydrocannabinol and 11-oxo-delta 8-tetrahydrocannabinol, in mice. Drug Metab. Dispos. 1981, 9, 261–264. [Google Scholar] [PubMed]
- Rosenthal, J.; Howell, M.; Earl, V.; Malik, M. Cannabinoid hyperemesis syndrome secondary to delta-8 THC use. Am. J. Med. 2021, 134, e582–e583. [Google Scholar] [CrossRef] [PubMed]
- Helander, A.; Johansson, M.; Andersson, A.; Villén, T. Analytical and medico-legal problems linked to the presence of delta-8-tetrahydrocannabinol (delta-8-THC): Results from urine drug testing in Sweden. Drug Test Anal. 2022, 14, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Valiveti, S.; Hammell, D.C.; Earles, D.C.; Stinchcomb, A.L. LC–MS method for the estimation of Δ8-THC and 11-nor-Δ8-THC-9-COOH in plasma. J. Pharm. Biomed. Anal. 2005, 38, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Akpunonu, P.; Baum, R.A.; Reckers, A.; Davidson, B.; Ellison, R.; Riley, M.; Trecki, J.; Gerona, R. Sedation and acute encephalopathy in a pediatric patient following ingestion of delta-8-tetrahydrocannabinol gummies. Am. J. Case Rep. 2021, 22, e933488–933481. [Google Scholar] [CrossRef]
- Vikingsson, S.; Hart, E.D.; Winecker, R.E.; Cone, E.J.; Kuntz, D.J.; Clark, M.; Jacques, M.; Hayes, E.D.; Flegel, R.R. Prevalence of∆ 8-tetrahydrocannabinol carboxylic acid in workplace drug testing. J. Anal. Toxicol. 2023, 47, 719–725. [Google Scholar] [CrossRef]
- Sim, Y.E.; Kim, J.W.; Ko, B.J.; Kim, J.Y.; Cheong, J.C.; Pyo, J. Determination of urinary metabolites of cannabidiol, Δ8-tetrahydrocannabinol, and Δ9-tetrahydrocannabinol by automated online μSPE–LC–MS/MS method. J. Chromatogr. B 2023, 1214, 123568. [Google Scholar] [CrossRef]
- Lin, L.; Amaratunga, P.; Reed, J.; Huang, P.; Lemberg, B.L.; Lemberg, D. Quantitation of Δ8-THC, Δ9-THC, cannabidiol and 10 other cannabinoids and metabolites in oral fluid by HPLC–MS-MS. J. Anal. Toxicol. 2022, 46, 76–88. [Google Scholar] [CrossRef]
- Coulter, C.; Wagner, J.R. Cannabinoids in oral fluid: limiting potential sources of cannabidiol conversion to Δ9-and Δ8-tetrahydrocannabinol. J. Anal. Toxicol. 2021, 45, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Milman, G.; Barnes, A.J.; Schwope, D.M.; Schwilke, E.W.; Darwin, W.D.; Goodwin, R.S.; Kelly, D.L.; Gorelick, D.A.; Huestis, M.A. Disposition of cannabinoids in oral fluid after controlled around-the-clock oral THC administration. Clin. Chem. 2010, 56, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Hädener, M.; König, S.; Fabritius, M.M.; Weinmann, W. Using THC-COOH cut-off concentrations for assessing cannabis consumption frequency: A recently detected THC-COOH isomer poses an important analytical problem. Toxichem Krimtech 2017, 84, 168. [Google Scholar]
- Baird, S.N.; Frazee III, C.C.; Garg, U. Evaluation of a Delta-9-Tetrahydrocannabinol Carboxylic Acid (Δ9-THC-COOH) Immunoassay and a Gas Chromatography–Mass Spectrometry (GC-MS) Method for the Detection of Delta-8-Tetrahydrocannabinol Carboxylic Acid (Δ8-THC-COOH). J. Lab. Med 2023, 8, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Crosby, M.M.; Truver, M.T.; Chronister, C.W.; Hoyer, J.L.; Kinsey, A.M.; Brogan, S.C.; Goldberger, B.A. Identification of 11-nor-∆ 8-Tetrahydrocannabinol-9-Carboxylic Acid in Postmortem Urine. J. Anal. Toxicol. 2023, 47, 481–487. [Google Scholar] [CrossRef]
- Pokhai, A.A.; Poklis, J.L.; Williams, G.R.; Wolf, C.E. The preanalytical stability of emerging cannabinoid analogs (delta-8 THC and its metabolites, delta-10 THC and carboxy-HHC) in urine. J. Anal. Toxicol. 2023, 47, 726–731. [Google Scholar] [CrossRef]
- Morin, M.M.; Baum, M. Effects of delta8 tetrahydrocannabinol (THC) on avoidance extinction in rats. Psychon. Bull. Rev. 1986, 24, 385–387. [Google Scholar] [CrossRef]
- Christensen, H.; Freudenthal, R.; Gidley, J.; Rosenfeld, R.; Boegli, G.; Testino, L.; Brine, D.; Pitt, C.; Wall, M. Activity of Δ8-and Δ9-tetrahydrocannabinol and related compounds in the mouse. Science 1971, 172, 165–167. [Google Scholar] [CrossRef]
- Pertwee, R.G.; Ross, R. Cannabinoid receptors and their ligands. Prostaglandins Leukot. Essent. Fatty Acids 2002, 66, 101–121. [Google Scholar] [CrossRef]
- Mackie, K. Cannabinoid receptors: where they are and what they do. J. Neuroendocrinol. 2008, 20, 10–14. [Google Scholar] [CrossRef]
- Mechoulam, R.; Parker, L.A. The endocannabinoid system and the brain. Annu. Rev. Psychol. 2013, 64, 21–47. [Google Scholar] [CrossRef]
- Govaerts, S.J.; Hermans, E.; Lambert, D.M. Comparison of cannabinoid ligands affinities and efficacies in murine tissues and in transfected cells expressing human recombinant cannabinoid receptors. Eur. J. Pharm. Sci. 2004, 23, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Nadipuram, A.K.; Krishnamurthy, M.; Ferreira, A.M.; Li, W.; Moore II, B.M. Synthesis and testing of novel classical cannabinoids: exploring the side chain ligand binding pocket of the CB1 and CB2 receptors. Bioorg. Med. Chem. 2003, 11, 3121–3132. [Google Scholar] [CrossRef]
- Huffman, J.W.; Liddle, J.; Yu, S.; Aung, M.M.; Abood, M.E.; Wiley, J.L.; Martin, B.R. 3-(1′, 1′-Dimethylbutyl)-1-deoxy-Δ8-THC and related compounds: synthesis of selective ligands for the CB2 receptor. Bioorg. Med. Chem. 1999, 7, 2905–2914. [Google Scholar] [CrossRef] [PubMed]
- Hirst, R.A.; Almond, S.L.; Lambert, D.G. Characterisation of the rat cerebella CB1 receptor using SR141716A, a central cannabinoid receptor antagonist. Neurosci. Lett. 1996, 220, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.R.; Compton, D.R.; Semus, S.F.; Lin, S.; Marciniak, G.; Grzybowska, J.; Charalambous, A.; Makriyannis, A. Pharmacological evaluation of iodo and nitro analogs of Δ8-THC and Δ9-THC. Pharmacol. Biochem. Behav.r 1993, 46, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Huffman, J.W.; Miller, J.R.; Liddle, J.; Yu, S.; Thomas, B.F.; Wiley, J.L.; Martin, B.R. Structure–activity relationships for 1′, 1′-dimethylalkyl-Δ8-tetrahydrocannabinols. Bioorg. Med. Chem. 2003, 11, 1397–1410. [Google Scholar] [CrossRef]
- Busch-Petersen, J.; Hill, W.A.; Fan, P.; Khanolkar, A.; Xie, X.-Q.; Tius, M.A.; Makriyannis, A. Unsaturated side chain β-11-hydroxyhexahydrocannabinol analogs. J. Med. Chem 1996, 39, 3790–3796. [Google Scholar] [CrossRef]
- Charalambous, A.; Lin, S.; Marciniak, G.; Banijamali, A.; Friend, F.; Compton, D.; Martin, B.; Makriyannis, A. Pharmacological evaluation of halogenated Δ8-THC analogs. Pharmacol. Biochem. Behav. 1991, 40, 509–512. [Google Scholar] [CrossRef]
- Papahatjis, D.P.; Nahmias, V.R.; Andreou, T.; Fan, P.; Makriyannis, A. Structural modifications of the cannabinoid side chain towards C3-aryl and 1′, 1′-cycloalkyl-1′-cyano cannabinoids. Bioorganic & medicinal chemistry letters 2006, 16, 1616–1620. [Google Scholar]
- Darmani, N.A.; Janoyan, J.J.; Crim, J.; Ramirez, J. Receptor mechanism and antiemetic activity of structurally-diverse cannabinoids against radiation-induced emesis in the least shrew. Eur. J. Pharmacol. 2007, 563, 187–196. [Google Scholar] [CrossRef]
- Domino, E.F. Cannabinoids and the cholinergic system. J Clin Pharmacol. 1981, 21, 249S–255S. [Google Scholar] [CrossRef]
- Tripathi, H.; Vocci, F.; Brase, D.; Dewey, W. Effects of cannabinoids on levels of acetylcholine and choline and on turnover rate of acetylcholine in various regions of the mouse brain. J. Drug Alcohol Res. 1987, 7, 525–532. [Google Scholar]
- Clarke, D.; Jandhyala, B. Acute and chronic effects of tetrahydrocannabinols on monoamide oxidase activity: possible vehicle/tetrahydrocannabinol interactions. Res. commun. chem. pathol. pharmacol. 1977, 17, 471–480. [Google Scholar] [PubMed]
- Avraham, Y.; Ben-Shushan, D.; Breuer, A.; Zolotarev, O.; Okon, A.; Fink, N.; Katz, V.; Berry, E.M. Very low doses of Δ8-THC increase food consumption and alter neurotransmitter levels following weight loss. Pharmacol. Biochem. Behav. 2004, 77, 675–684. [Google Scholar] [CrossRef]
- Darmani, N.A. Cannabinoids of diverse structure inhibit two DOI-induced 5-HT2A receptor-mediated behaviors in mice. Pharmacol. Biochem. Behav.r 2001, 68, 311–317. [Google Scholar] [CrossRef]
- MacLean, K.I.; Littleton, J.M. Environmental stress as a factor in the response of rat brain catecholamine metabolism to Δ8-tetrahydrocannabinol. Eur. J. Pharmacol. 1977, 41, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Poddar, M.; Dewey, W. Effects of cannabinoids on catecholamine uptake and release in hypothalamic and striatal synaptosomes. J. Pharmacol. Exp. Ther. 1980, 214, 63–67. [Google Scholar]
- Sagratella, S.; de Carolis, A.S.; Longo, V. EEG interaction between delta 8-tetrahydrocannabinol and some sedative-anxiolytic drugs. Is the anxiogenic effect of cannabis related to an action on the GABAergic system? Adv. biochem. psychopharmacol. 1986, 41, 203–209. [Google Scholar]
- Little, P.J.; Martin, B.R. The effects of Δ9-tetrahydrocannabinol and other cannabinoids on cAMP accumulation in synaptosomes. Life Sci. 1991, 48, 1133–1141. [Google Scholar] [CrossRef]
- JÄrbe, T.U.; Henriksson, B.G. Effects of Δ 8-THC, and Δ 9-THC on the acquisition of a discriminative positional habit in rats: The transitions between normal and tetrahydrocannabinol-induced states on reversal learning. Psychopharmacologia 1973, 31, 321–332. [Google Scholar] [CrossRef]
- Järbe, T.; Johansson, J.; Henriksson, B. Characteristics of tetrahydrocannabinol (THC)-produced discrimination in rats. Psychopharmacologia 1976, 48, 181–187. [Google Scholar] [CrossRef]
- JÄrbe, T.U.; Henriksson, B.G. Discriminative response control produced with hashish, tetrahydrocannabinols (Δ 8-THC and Δ 9-THC), and other drugs. Psychopharmacologia 1974, 40, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bueno, O.; Carlini, E.; Finkelfarb, E.; Suzuki, J.S. Δ 9-Tetrahydrocannabinol, ethanol, and amphetamine as discriminative stimuli-generalization tests with other drugs. Psychopharmacologia 1976, 46, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Doty, P.; Dykstra, L.A.; Picker, M.J. Discriminative stimulus effects of phencyclidine: pharmacologically specific interactions with Δ9-and Δ8-tetrahydrocannabinol. Drug Alcohol Depend. 1994, 35, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Järbe, T.U.; Henriksson, B.G.; Ohlin, G.C. Delta9-THC as a discriminative cue in pigeons: effects of delta8-THC, CBD, and CBN. Arch. Int. Pharmacodyn. Ther. 1977, 228, 68–72. [Google Scholar] [PubMed]
- Hine, B.; Torrelio, M.; Gershon, S. Analgesic, heart rate, and temperature effects of Δ8-THC during acute and chronic administration to conscious rats. Pharmacology 1977, 15, 65–72. [Google Scholar] [CrossRef]
- Durbin, D.J.; King, J.M.; Stairs, D.J. Behavioral Effects of Vaporized Delta-8 Tetrahydrocannabinol, Cannabidiol, and Mixtures in Male Rats. Cannabis Cannabinoid Res. 2023. [Google Scholar] [CrossRef] [PubMed]
- Chesher, G.; Dahl, C.; Everingham, M.; Jackson, D.; Marchant-Williams, H.; Starmer, G. The effect of cannabinoids on intestinal motility and their antinociceptive effect in mice. Br. J. Pharmacol. 1973, 49, 588–594. [Google Scholar] [CrossRef]
- Welch, S.P.; Stevens, D.L. Antinociceptive activity of intrathecally administered cannabinoids alone, and in combination with morphine, in mice. J. Pharmacol. Exp. Ther. 1992, 262, 10–18. [Google Scholar]
- Welch, S.P.; Thomas, C.; Patrick, G.S. Modulation of cannabinoid-induced antinociception after intracerebroventricular versus intrathecal administration to mice: possible mechanisms for interaction with morphine. J. Pharmacol. Exp. Ther. 1995, 272, 310–321. [Google Scholar]
- El-Alfy, A.T.; Ivey, K.; Robinson, K.; Ahmed, S.; Radwan, M.; Slade, D.; Khan, I.; ElSohly, M.; Ross, S. Antidepressant-like effect of Δ9-tetrahydrocannabinol and other cannabinoids isolated from Cannabis sativa L. Pharmacol. Biochem. Behav. 2010, 95, 434–442. [Google Scholar] [CrossRef]
- Welch, S.P.; Huffman, J.W.; Lowe, J. Differential blockade of the antinociceptive effects of centrally administered cannabinoids by SR141716A. J. Pharmacol. Exp. Ther. 1998, 286, 1301–1308. [Google Scholar]
- Consroe, P.F.; Man, D.P.; Chin, L.; Picchioni, A.L. Reduction of audiogenic seizure by Δ8-and Δ9-tetrahydrocannabinols. J. Pharm. Pharmacol. 1973, 25, 764–765. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, M.E.; McCaughran Jr, J.A.; Wada, J.A. Antiepileptic and prophylactic effects of tetrahydrocannabinols in amygdaloid kindled rats. Epilepsia 1978, 19, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Wada, J.A.; Osawa, T.; Corcoran, M.E. Effects of tetrahydrocannabinols on kindled amygdaloid seizures and photogenic seizures in Senegalese baboons, Papio papio. Epilepsia 1975, 16, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Wada, J.A.; Wake, A.; Sato, M.; Corcoran, M.E. Antiepileptic and prophylactic effects of tetrahydrocannabinols in amygdaloid kindled cats. Epilepsia 1975, 16, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Brenda, K.C.; Charles, L.; Charles, R.C. Effects of marihuana cannabinoids on seizure activity in cobalt-epileptic rats. Pharmacol. Biochem. Behav. 1982, 16, 573–578. [Google Scholar]
- Chandradhar, D.; Raymond, D.H. Anticonvulsant activities of Δ-8 and Δ-9 tetrahydrocanabinol and uridine. Toxicol. Appl. Pharmacol. 1975, 31, 452–458. [Google Scholar]
- Johansson, J.; Jarbe, T.; Henriksson, B. Acute and subchronic influences of tetrahydrocannabinols on water and food intake, body weight, and temperature in rats. T.-I.-T. J. Life Sci. 1975, 5, 17–27. [Google Scholar]
- Smiley, K.; Karler, R.; Turkanis, S. Effects of cannabinoids on the perfused rat heart. Res. commun. chem. pathol. pharmacol. 1976, 14, 659–675. [Google Scholar]
- Coffey, R.G.; Yamamoto, Y.; Snella, E.; Pross, S. Tetrahydrocannabinol inhibition of macrophage nitric oxide production. Biochem. Pharmacol. 1996, 52, 743–751. [Google Scholar] [CrossRef]
- Adams, M.; Earnhardt, J.; Dewey, W.; Harris, L. Vasoconstrictor actions of delta8-and delta9-tetrahydrocannabinol in the rat. J. Pharmacol. Exp. Ther. 1976, 196, 649–656. [Google Scholar]
- Abrahamov, A.; Abrahamov, A.; Mechoulam, R. An efficient new cannabinoid antiemetic in pediatric oncology. Life Sci. 1995, 56, 2097–2102. [Google Scholar] [CrossRef]
- Carchman, R.A.; Harris, L.S.; Munson, A.E. The inhibition of DNA synthesis by cannabinoids. Cancer Res. 1976, 36, 95–100. [Google Scholar]
- Tucker, A.N.; Friedman, M.A. Effects of cannabinoids on L1210 murine leukemia. 1. Inhibition of DNA synthesis. Res. commun. chem. pathol. pharmacol. 1977, 17, 703–714. [Google Scholar] [PubMed]
- Tucker, A.N.; Friedman, M.A. Effects of cannabinoids on L1210 murine leukemia. III. Inhibition of respiration. Res. commun. chem. pathol. pharmacol. 1979, 23, 327–332. [Google Scholar] [PubMed]
- Green, L.G.; Stein, J.L.; Stein, G.S. A decreased influence of cannabinoids on macromolecular biosynthesis and cell proliferation in human cells which metabolize polycyclic hydrocarbon carcinogens. Anticancer Res. 1983, 3, 211–217. [Google Scholar] [PubMed]
- Howlett, A.C.; Fleming, R.M. Cannabinoid inhibition of adenylate cyclase. Pharmacology of the response in neuroblastoma cell membranes. Mol. Pharmacol. 1984, 26, 532–538. [Google Scholar] [PubMed]
- Molnár, J.; Szabó, D.; Pusztai, R.; Mucsi, I.; Berek, L.; Ocsovszki, I.; Kawata, E.; Shoyama, Y. Membrane associated antitumor effects of crocine-, ginsenoside- and cannabinoid derivates. Anticancer Res. 2000, 20, 861–867. [Google Scholar] [PubMed]
- Whyte, D.A.; Al-Hammadi, S.; Balhaj, G.; Brown, O.M.; Penefsky, H.S.; Souid, A.K. Cannabinoids inhibit cellular respiration of human oral cancer cells. Pharmacology 2010, 85, 328–335. [Google Scholar] [CrossRef]
- Semlali, A.; Beji, S.; Ajala, I.; Rouabhia, M. Effects of tetrahydrocannabinols on human oral cancer cell proliferation, apoptosis, autophagy, oxidative stress, and DNA damage. Arch. Oral Biol. 2021, 129, 105200. [Google Scholar] [CrossRef]
- Munson, A.E.; Harris, L.S.; Friedman, M.A.; Dewey, W.L.; Carchman, R.A. Antineoplastic Activity of Cannabinoids2. JNCI: J. Natl. Cancer Inst. 1975, 55, 597–602. [Google Scholar] [CrossRef]
- Friedman, M.A. In vivo effects of cannabinoids on macromolecular biosynthesis in Lewis lung carcinomas. Cancer Biochem. Biophys. 1977, 2, 51–54. [Google Scholar]
- Nahas, G.G.; Morishima, A.; Desoize, B. Effects of cannabinoids on macromolecular synthesis and replication of cultured lymphocytes. Fed. Proc. 1977, 36, 1748–1752. [Google Scholar] [PubMed]
- Schwarz, H.; Blanco, F.J.; Lotz, M. Anadamide, an endogenous cannabinoid receptor agonist inhibits lymphocyte proliferation and induces apoptosis. J. Neuroimmunol. 1994, 55, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Yamaori, S.; Ishii, H.; Chiba, K.; Yamamoto, I.; Watanabe, K. Δ-Tetrahydrocannabinol induces cytotoxicity in macrophage J774-1 cells: involvement of cannabinoid receptor 2 and p38 MAPK. Toxicology 2013, 314, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Loveless, S.; Harris, L.; Munson, A. Hyporesponsiveness to the immunosuppressant effects of delta-8-tetrahydrocannabinol. J. Immunopharmacol. 1981, 3, 371–384. [Google Scholar] [CrossRef]
- Watson, E.S.; Murphy, J.C.; Turner, C.E. Allergenic properties of naturally occurring cannabinoids. J. Pharm. Sci. 1983, 72, 954–955. [Google Scholar] [CrossRef] [PubMed]
- Wirguin, I.; Mechoulam, R.; Breuer, A.; Schezen, E.; Weidenfeld, J.; Brenner, T. Suppression of experimental autoimmune encephalomyelitis by cannabinoids. Immunopharmacol. 1994, 28, 209–214. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Shoyama, Y.; Watanabe, S.; Yamamoto, T. Behavioral suppression induced by cannabinoids is due to activation of the arachidonic acid cascade in rats. Brain Res. 2001, 889, 149–154. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Kubota, T.; Watanabe, S.; Yamamoto, T. Activation of brain prostanoid EP3 receptors via arachidonic acid cascade during behavioral suppression induced by Delta8-tetrahydrocannabinol. J. Neurochem. 2004, 88, 148–154. [Google Scholar] [CrossRef]
- Chen, Y.; Buck, J. Cannabinoids protect cells from oxidative cell death: a receptor-independent mechanism. J. Pharmacol. Exp. Ther. 2000, 293, 807–812. [Google Scholar]
- Thapa, D.; Cairns, E.A.; Szczesniak, A.-M.; Toguri, J.T.; Caldwell, M.D.; Kelly, M.E. The cannabinoids Δ8THC, CBD, and HU-308 act via distinct receptors to reduce corneal pain and inflammation. Cannabis cannabinoid Res. 2018, 3, 11–20. [Google Scholar] [CrossRef]
- Punyamurthula, N.S.; Adelli, G.R.; Gul, W.; Repka, M.A.; ElSohly, M.A.; Majumdar, S. Ocular disposition of ∆8-tetrahydrocannabinol from various topical ophthalmic formulations. AAPS PharmSciTech 2017, 18, 1936–1945. [Google Scholar] [CrossRef]
- Muchtar, S.; Almog, S.; Torracca, M.; Saettone, M.; Benita, S. A submicron emulsion as ocular vehicle for delta-8-tetrahydrocannabinol: effect on intraocular pressure in rabbits. Ophthalmic Res. 1992, 24, 142–149. [Google Scholar] [CrossRef]
- Elsohly, M.; Harland, E.; Murphy, J.; Wirth, P.; Waller, C. Cannabinoids in glaucoma: a primary screening procedure. J. Clin. Pharmacol. 1981, 21, 472S–478S. [Google Scholar] [CrossRef] [PubMed]
- Elsohly, M.A.; Harland, E.C.; Benigni, D.A.; Waller, C.W. Cannabinoids in glaucoma II: the effect of different cannabinoids on intraocular pressure of the rabbit. Curr. Eye Res. 1984, 3, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Green, K.; Symonds, C.M.; Oliver, N.W.; Elijah, R.D. Intraocular pressure following systemic administration of cannabinoids. Curr. Eye Res. 1982, 2, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Badawy, Z.S.; Chohan, K.R.; Whyte, D.A.; Penefsky, H.S.; Brown, O.M.; Souid, A.-K. Cannabinoids inhibit the respiration of human sperm. Fertil. Steril. 2009, 91, 2471–2476. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Elbracht, C. Effect of tetrahydrocannabinols on pubertal body weight spurt and sex hormones in developing male rats. Res. Exp. Med. 1983, 182, 95–104. [Google Scholar] [CrossRef]
- Knopf, A. CDC and FDA warn of delta-8 THC harms. Alcoholism & Drug Abuse Weekly 2021, 33, 7–7. [Google Scholar]
- Erickson, B.E. Delta-8-THC craze concerns chemists. Chem. Eng. News 2021, 99, 25–28. [Google Scholar]
- Leas, E.C.; Harati, R.M.; Satybaldiyeva, N.; Morales, N.E.; Huffaker, S.L.; Mejorado, T.; Grant, I. Self-reported adverse events associated with∆ 8-Tetrahydrocannabinol (Delta-8-THC) Use. J Cannabis Res. 2023, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.J.; Swenson, C.; Fishkin, I.; Malanga, A. Now I know my CBDs; cases of psychiatric admissions after delta-8-tetrahydrocannabinol (delta-8-THC, Δ⁸-THC) product usage. Psychiatry Res. Case Rep. 2023, 2, 100166. [Google Scholar] [CrossRef]
- Shaker, K.; Nillas, A.; Ellison, R.; Martin, K.; Trecki, J.; Gerona, R.; Aldy, K. Delta-8-tetrahydrocannabinol exposure and confirmation in four pediatric patients. J Med Toxicol. 2023, 19, 190–195. [Google Scholar] [CrossRef]
- Wrenn, J.; Friedman, M. Effects of chronic administration of delta8-and delta9-tetrahydro-cannabinol on hepatic tyrosine aminotransferase activity in mice. Arch. Int. Pharmacodyn. Ther. 1978, 235, 4–8. [Google Scholar]
- Bozman, M.E.; Manoharan, S.V.R.R.; Vasavada, T. Marijuana variant of concern: delta 8-tetrahydrocannabinol (delta-8-THC, Δ8-THC). Psychiatry Res. Case Rep. 2022, 1, 100028. [Google Scholar] [CrossRef]
- Watanabe, K.; Narimatsu, S.; Yamamoto, I.; Yoshimura, H. Difference in tolerance development to hypothermia and pentobarbital-induced sleep prolongating effect of 11-hydroxy-Δ8-tetrahydrocannabinol and 11-oxo-Δ8-tetrahydrocannabinol in mice. Eur. J. Pharmacol. 1982, 77, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Yamamoto, I.; Yoshimura, H. Development of tolerance and cross-tolerance to the cataleptogenic effects of Δ8-tetrahydrocannabinol and 11-hydroxy-Δ8-tetrahydrocannabinol in mice. Eur. J. Pharmacol. 1983, 94, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, I.; Watanabe, K.; Narimatsu, S.; Hamajima, K.; Yoshimura, H. Cross-tolerance to the hypothermic effect of Δ8-tetrahydrocannabinol 11-hydroxy-Δ8-tetrahydrocannabinol and chlorpromazine in the mouse. Eur. J. Pharmacol. 1985, 111, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Narimatsu, S.; Yamamoto, I.; Watanabe, K.; Yoshimura, H. Change in hypothermia and catalepsy induced by cannabinoids or morphine in mice tolerant to these substances. Eur. J. Pharmacol. 1987, 141, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Sjödén, P.-O.; Järbe, T.U.; Henriksson, B.G. Effects of long-term administration and withdrawal of tetrahydrocannabinols (Δ8-THC and Δ9-THC) on open-field behavior in rats. Pharm. Biochem. Behav. 1973, 1, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Hagiwara, Y.; Tanaka, H.; Sugiura, T.; Waku, K.; Shoyama, Y.; Watanabe, S.; Yamamoto, T. Endogenous cannabinoid, 2-arachidonoylglycerol, attenuates naloxone-precipitated withdrawal signs in morphine-dependent mice. Brain Res. 2001, 909, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, H.N. Time course of the effects of naturally occurring cannabinoids on morphine abstinence syndrome. Pharm. Biochem. Behav. 1978, 8, 7–11. [Google Scholar] [CrossRef]
- Nakamura-Palacios, E.M.; Bueno, O.F.A.; Takahashi, R.N.; Tufik, S. Acute or chronic effects of cannabinoids on spontaneous or pharmacologically induced yawning in rats. Pharm. Biochem. Behav. 2002, 74, 205–212. [Google Scholar] [CrossRef]
- Anggadiredja, K.; Yamaguchi, T.; Tanaka, H.; Shoyama, Y.; Watanabe, S.; Yamamoto, T. Decrease in prostaglandin level is a prerequisite for the expression of cannabinoid withdrawal: a quasi abstinence approach. Brain Res. 2005, 1066, 201–205. [Google Scholar] [CrossRef]
Figure 1.
Numbering systems of tetrahydrocannabinols according to the monoterpene (past) and dibenzopyrane (current) numbering systems.
Figure 1.
Numbering systems of tetrahydrocannabinols according to the monoterpene (past) and dibenzopyrane (current) numbering systems.
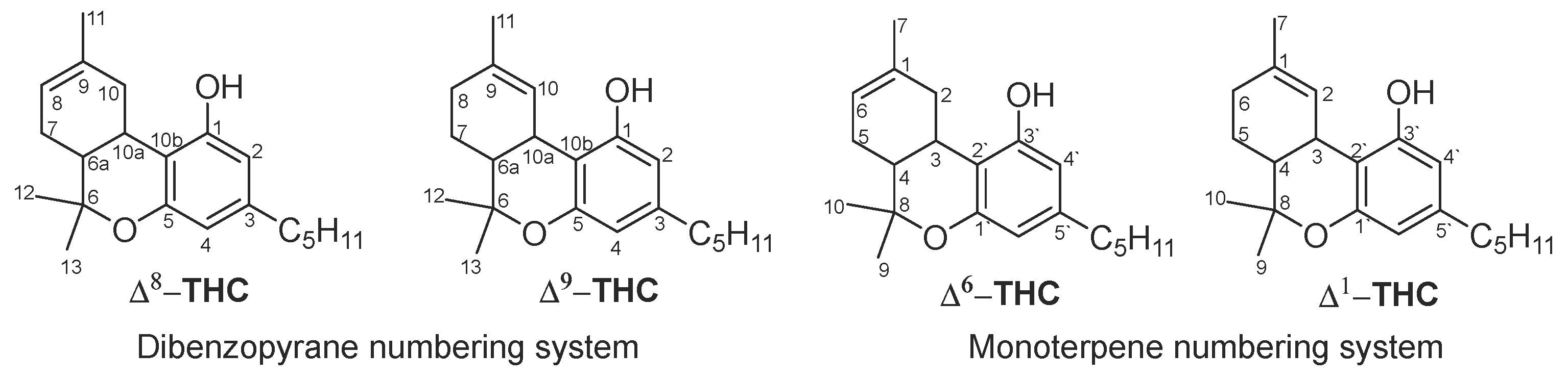
Figure 3.
The synthetic pathway of Δ8-THC.
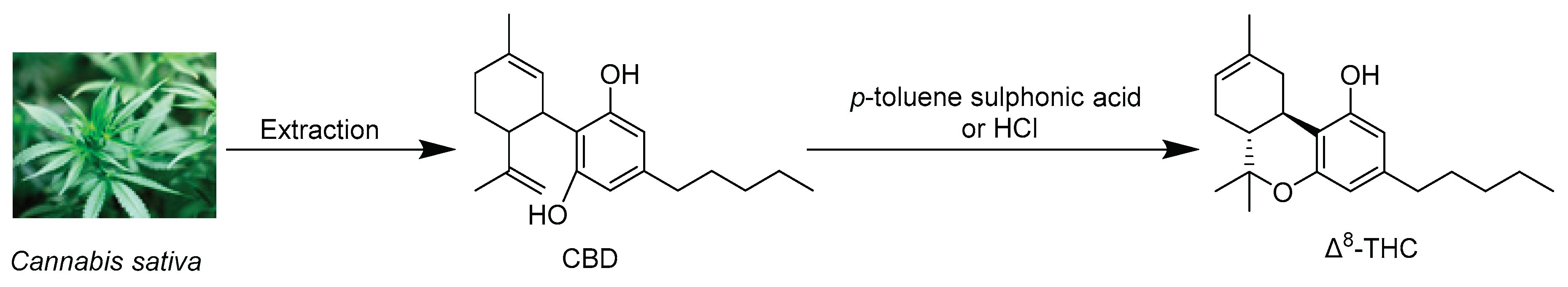
Figure 5.
Chemical structures of Δ8-THC and its related impurities found in Δ8-THC commercial products.
Figure 5.
Chemical structures of Δ8-THC and its related impurities found in Δ8-THC commercial products.
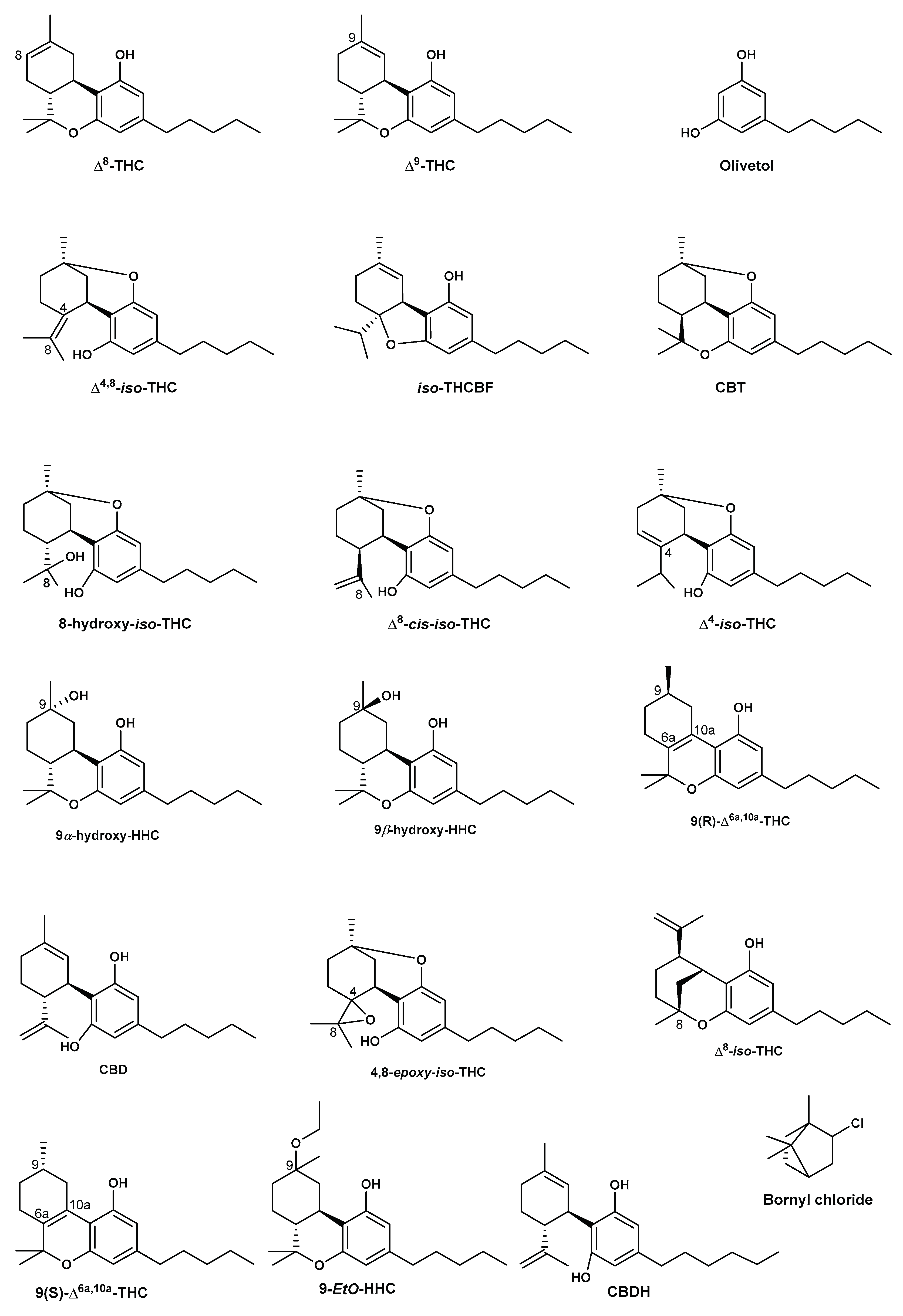
Figure 6.
The chemical structures of tetrahydrocannabinol (THC) isomers [68].
Figure 6.
The chemical structures of tetrahydrocannabinol (THC) isomers [68].
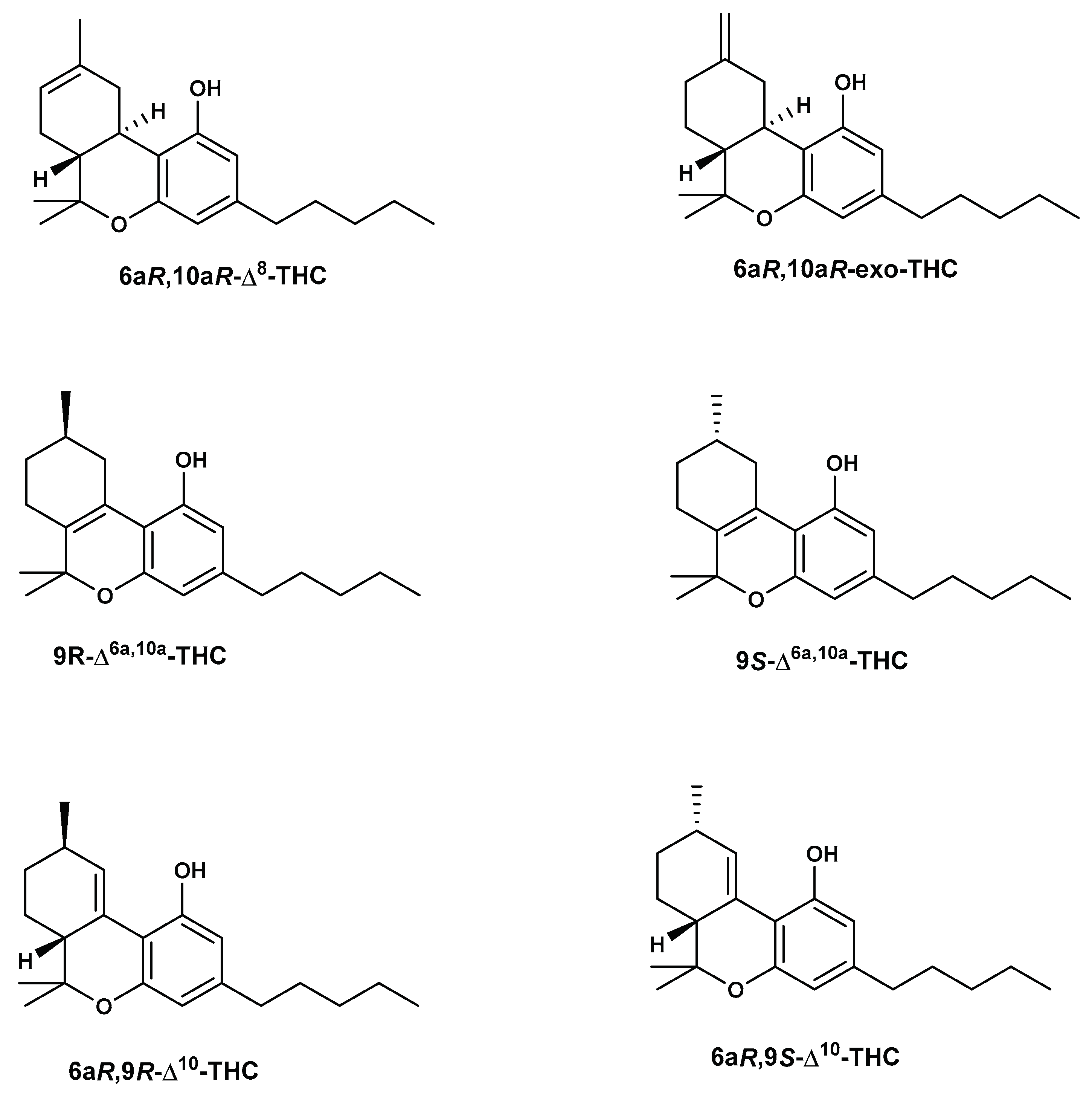
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



