Submitted:
09 January 2024
Posted:
10 January 2024
You are already at the latest version
Abstract
The modification of GFP-CaaL sensor proteins by geranylgeranyl groups in plants requires an active plastidial mevalonate (MVA)-independent pathway. As a consequence, fosmidomycin, a specific inhibitor of 1-deoxy-D-xylulose (DX)-5 phosphate reductoisomerase/DXR, the second enzyme in this so-called methylerythritol phosphate (MEP) pathway, impedes protein prenylation, which results in GFP-labeling of nuclei instead of the plasma membrane. In tobacco cells, a visual screen of conditions that allowed membrane localization of GFP-CaaL proteins in the presence of fosmidomycin, pinpointed jasmonic acid methyl esther (Me-JA) as a chemical able to overcome the inhibition in a gradual manner. Using Arabidopsis protein prenyltransferase loss-of-function mutant lines expressing GFP-CaaL proteins, we demonstrated that Me-JA induces a modification in protein substrate recognition by protein farnesyltransferase (PFT). Similar to Me-JA, farnesol and MVA also alter the protein substrate specificity of PFT, whereas DX and geranylgeraniol have limited or no effect. Our data indicate that stimulation of the MVA pathway adjusts the protein substrate specificity of PFT, which is corroborated with metabolic cross-talk of the origin of the prenyl group used to modify the protein. To illustrate how activation of the MVA pathway impacting PFT substrate specificity benefits a plant under stress conditions, a model is proposed.
Keywords:
metabolic cross-talk
; isoprenoid biosynthesis
; isoprenylated proteins
; jasmonic acid methyl esther/Me-JA
; MEP pathway
; MVA pathway
1. Introduction
Type I protein prenylation is a post-translational process common to eukaryotes that adds a hydrophobic farnesyl (F) or geranylgeranyl (GG) moiety to proteins harboring a C-terminus CaaX (C = cyst; a = aliphatic; X = C-terminus amino acid) motif. Modified proteins attain new traits mainly gaining in hydrophobicity. Thus, prenylation enables interactions with membrane structures, with other proteins for example in mega-complexes, or changes the conformation of the target protein. It relies on two metalloenzymes that have to discern two co-substrates: the CaaX protein and a prenyl diphosphate. These protein prenyltransferases (PPTs) are heterodimers with a common α-subunit and a distinct and specific β-subunit. Accordingly, protein farnesyltransferase (PFT; EC 2.5.1.58) catalyzes a thioetherification between the cysteine and farnesyl using farnesyl diphosphate (FPP) as a co-substrate, while type-I protein geranylgeranyltransferase (PGGT-I; EC 2.5.1.59) uses geranylgeranyl diphosphate (GGPP). It had been accepted that the nature of the “X” amino acid guides the enzyme’s reaction to either a transfer of farnesyl (farnesylation) catalyzed by PFT or geranylgeranyl (geranylgeranylation) catalyzed by PGGT-I [1]. Although in practice, very few prenylated proteins have been characterized in plants, it is clear that CaaX-motif proteins are involved in many cellular processes. They particularly participate to plant development and plant stress responses [2,3]. The use and characterization of loss-of-function mutants have been fundamental in these investigations. In this matter, Arabidopsis thaliana T-DNA insertion mutants in the PLP gene encoding the α-subunit, in the ERA1 gene (enhanced response to ABA-1) encoding the β-subunit of PFT and in the GGB gene encoding the β-subunit of PGGT have been described [4,5,6,7]. The ggb mutant plant lacks any evident developmental phenotype, but shows enhanced response to abscisic acid (ABA) in guard cells leading to stomatal closure [7]. Compared to ggb plants, the phenotypes of an Arabidopsis era1 mutant plant are more marked with an enhanced response to ABA in guard cells, but also in seed germination. They are characterized by a growth delay and reduced fertility under standard growth conditions, as well. In addition, era1 mutants also show enlarged meristems and supranumerary floral organs, as well larger seeds [8,9,10,11,12]. Moreover, compared to wild-type plants, era1 loss-of-function plants display different sensibilities towards environmental factors with clear drought and heat tolerance phenotypes [4,5,13,14,15,16,17]. The phenotypes of the plp mutant [6] and the era1ggb double mutant [7] are comparable to that of era1, but are significantly more marked. Both mutant plants produce severely hypertrophied meristems, additional floral organs, and show stunted growth and significantly reduced stature than the wild type [6,7].
To attain biological activity, CaaX-proteins must be modified by the prenyl group, a modification that ensures the proper localization within the cell. In addition, a proteolysis releasing the “aaX” peptide and carboxymethylation of the cysteine are described to be important steps for the biological activity, at least for some proteins [18]. Since FPP and GGPP are central isoprenoid precursors, shared by different metabolic branches in the pathway, PPTs must compete with other enzymes for the availability of these substrates. For instance, in Nicotiana tabacum, phytoalexins, including capsidiol, lubimin, rishitin, solavetivone or phytuberin [19], are sesquiterpenoids derived from FPP. In a similar way, GGPP is for instance the precursor of plastidial pigments such as carotenoids and chlorophylls. It is noteworthy to mention that concentrations of prenyl diphosphates needed for protein modifications are much lower than those needed for sesquiterpenoid or pigment biosynthesis. The way PPTs get their part of the cake must be closely connected to metabolic regulation, cellular compartmentation and affinity of enzymes to their substrate. However, the molecular mechanisms by which such a regulation occurs in cellula is very poorly understood, most of data being generated from experiments performed in vitro [20]. The modulation of PPT activity can be achieved at two distinct levels: either on an extension of protein substrates used specifically by an enzyme or by using alternative prenyl diphosphate substrates to modify the protein. The CaaX consensus motif (containing three variable residues) is by definition not very stringent and has over years constantly been extended and reevaluated [21]. A further degree of flexibility, especially in plants, is achieved by the use of the second substrate, the prenyl diphosphate. Protein prenylation in mammals and yeast relies exclusively on the classical mevalonic acid (MVA) isoprenoid biosynthesis pathway [1]. In contrast, plants are unique in that they use two isoprenoid biosynthesis pathways in parallel, with the potential for exchanging metabolic intermediates. This flexibility is known as metabolic cross-talk for the biosynthesis of isoprenoids (see [22]). In practice, modifications with these hydrophobic prenyl groups depend on isoprenoid precursors (FPP or GGPP) biosynthesized either through the classical MVA pathway [23,24,25,26] or the plastidial methylerythritol phosphate (MEP) pathway [27,28,29]. To study regulation of protein prenylation in vivo, this characteristic is of major advantage in determing metabolic origins of prenyl groups used to modify prenylated proteins [29]. In fact, the use of specific inhibitors to the synthesis of GGPP (from the MEP pathway) or FPP (from the MVA pathway) enables discrimination of the metabolic origin under specific conditions. Furthermore, instead of being neosynthesized, FPP can be recycled through a process known as the “FPP salvage pathway”, implemented for modifying a new series of proteins (for review see [30]). Plants generate also a broader range of prenyl diphosphates than other organisms engaged in protein prenylation [31]. Chain length and saturation degree of those prenyl diphosphates fluctuate [32]. Interestingly, some of those (e.g. dolichols, phytyl residues) have been proposed to be incorporated into prenylated protein [25]. Jointly, these singularities in plants consent to an adjustment of precursor supplies, according to specific biosynthetic needs. This strategy offers flexibility by providing precursors that allow cells to simultaneously biosynthesize a variety of different isoprenoid compounds according to their needs.
Despite the extensive investigation of metabolic cross-talk for the biosynthesis of isoprenoid metabolites [22,33,34,35], the situation regarding the supply of FPP and GGPP utilized in protein modification is scarcely described. Tobacco BY-2 cells have been extensively used as a model plant to study protein prenylation in plants, this for several reasons. They divide rapidly and thus are metabolically very active, but also because the catalytical activities of PPTs are rather high [23]. Under standard culture conditions, this cell suspension uses exclusively plastidial MEP-derived GGPP to modify a chimeric GFP-CaM-CVIL line, that can be defined as a prenylable GFP-based sensor [28]. This property is characterized by a reallocation of the fluorescent signal from the plasma membrane into the nucleus, when MEP pathway specific inhibitors, like fosmidomycin or oxoclomazone block geranylgeranylation of the protein [28,36]. Thus, the PPT catalyzing the transfer of GGPP to the cysteinyl residue uses specifically a MEP-derived C20 substrate and is unable to substitute this substrate by an MVA-derived prenyl group. The question arising is to determine of whether under specific conditions, prenylation of this protein can be achieved using MVA-derived isoprenyl units? A switch at this level would imply the existence of specific signals initiating metabolic cross-talk in the cell. Yet, in this context, it has been shown that farnesol, known as an enhancer of plant HMGR activity [37], functions as a compound shifting the metabolic origin of the prenylation substrates from the MEP to the MVA pathway, this to modify the GFP-CaM-CVIL sensor protein [38]. The mechanism by which this switch occurs is unknown and needs further investigations. The identification of cross-talk inducers within cellular systems would aid in the establishment of optimal conditions for the accumulation of valuable isoprenoid metabolites in plants. Thus, we screened for chemicals being able to allow protein prenylation in tobacco BY-2 cells under restrictive GGPP biosynthesis conditions, with a special emphasis given on hormonal regulation. The effect of the stress hormone jasmonic acid methyl esther (methyl-jasmonate, Me-JA) that was identified as a key element in the induction of such a metabolic switch, was carefully investigated and analyzed to figure out of whether the prenylation activity switches from the use of a MEP pathway-derived metabolite to that of an MVA-derived.
2. Results
2.1. Me-JA Promotes Membrane Localization of Prenylated MEP-Derived CaaL-Box Proteins in the Presence of Fosmidomycin
Our previous studies support the view that under standard growth conditions, the MEP pathway supplies prenyl diphosphate used to modify GFP-CaaL-like proteins in tobacco cells and plants [28,29,36,38,39]. This in mind, the objective of this study was to identify conditions under which the MVA pathway could possibly be activated to take over the supply of prenyl diphosphates in order to modify a protein substrate typically dedicated to PGGT. To do so, the prenylation sensor protein GFP-CaM-CVIL expressed in tobacco BY-2 cell line under the control of an inducible dexamethasone promoter has been used [28].
Under standard growth conditions, the protein is geranylgeranylated and localizes to the plasma membrane (Figure 1a, CONTROL). When treated with the MEP pathway specific inhibitor fosmidomycin, which blocks the plastidial GGPP biosynthesis by inhibiting the second enzyme in the MEP pathway (1-deoxy-D-xylulose 5-phosphate reductoisomerase), the fluorescent protein labels the nucleus triggered by a NLS motif (Figure 1a, FOS). This indicates a deficiency in prenylation [28]. Using fosmidomycin-treated cells, we ran an optical screen for compounds preventing nuclear localization of the fluorescent GFP-BD-CVIL protein in the presence of both fosmidomycin and the additional chemical. The candidate compound Me-JA (50 µM) restored plasma membrane-specific labeling in the presence of fosmidomycin (Figure 1a, FOS + MeJA). Since cells were co-treated with rather high concentrations of fosmidomycin (100 µM), it can be inferred that a MEP pathway-independent prenylation has been activated. In order to evaluate the reproducibility of the effect of Me-JA, statistical analyses were conducted (Figure 1b). To demonstrate that the effect is independent of the nature of the prenylated protein, in addition to GFP-CaM-CVIL, we also used a second tobacco BY-2 cell line expressing GFP-ROP6-CSIL. ROP6-CSIL, is not only prenylated, but is also acylated [40]. Me-JA (20 µM), as the only chemical added to the culture had no significant effect on GFP-CaaL prenylation. Fosmidomycin (100 µM) delocalizes proteins into the nucleus (> 87%), and in combination with Me-JA, full membrane localization (93% and 65%) was significantly reached (Figure 1b). In order to evaluate the minimum concentration of Me-JA required to achieve protein prenylation, a concentration range from 0.5 µM up to 20 µM was examined (Figure 1c). Concentrations as low as 0.5 µM Me-JA lead to significant relocalization of GFP-CaM-CVIL into the plasma membrane (Figure 1c). In a dose dependent manner, the higher the concentration of Me-JA is, the better the membrane localization is detected. Other phytohormones have been tested, but in the presence of ABA, IAA, NPA or kinetin, inhibition with fosmidomycin remained efficient (Supplemental material Figure S1). On the other hand, ethephon was identified as a hit. We wondered if ethylene might behave in a synergy with MeJA, as both are as defense signaling hormones [41]. Ethephon is capable of overcoming inhibition by fosmidomycin at 100 µM (Figure 1d). In the presence of Me-JA (1 µM), the effect is additive. We can therefore conclude that ethylene acts synergistically with Me-JA.
2.2. PFT Modifies GFP-CaM-CVIL in the Presence of Methyl-Jasmonate
Me-JA-induced relocalization of GFP-CaM-CVIL in the presence of fosmidomycin raises new questions, especially what enzyme is responsible for catalyzing the transfer of a prenyl group in treated cells? To answer this question, we analyzed Arabidopsis KO mutant plants, in which either PGGT (era1 lines) is expressed alone, or PFT (ggb lines). Lines expressing GFP-CaM-CVIL under the control of a dexamethasone-inducible promoter were generated. The protein has been expressed in WT Arabidopsis Col0 plants, in the PFT loss-of-function era1 mutant line and in the ggb mutant line, in which PGGT is lacking. The induced subcellular localization of GFP-CaM-CVIL has been monitored by confocal microscopy in leaves (surrounded by a green rectangle) and roots (surrounded by a yellow rectangle) of two-to-three weeks-old plantlets (Figure 2). As expected, the membrane localization observed in root and epidermic cells of Col0 (wild-type/WT) or era1 lines is consistent with PGGT being the active enzyme catalyzing GFP-CaM-CVIL modification under standard growth conditions (Figure 2). Furthermore, a nuclear localization of GFP-CaM-CVIL expressed in ggb mutant lines indicates that in the absence of an active PGGT, PFT is unable to support the prenylation of the sensor protein (Figure 2). This mutant line, which is the only one with a nuclear localization of GFP-CaM-CVIL, has been used to test the effect of Me-JA. In contrast to untreated ggb plants, the protein localizes mainly in the plasma membrane in the presence of Me-JA (30 µM), with only a negligible nuclear labeling (surrounded by a blue rectangle, Figure 2). Hence, in the presence of Me-JA, PFT which is the active enzyme in ggb plants, recognizes GFP-CaM-CVIL as a protein substrate. From this, it can be deduced that in the presence of Me-JA, PFT recognizes the GFP-CaM-CVIL substrate typically dedicated to PGGT, and that Me-JA seems to induce a signal directing PFT substrate specificity. The question arising is what kind of signal might be responsible of this functional enzymatic adjustment?
2.3. Me-JA Endorses Protein Prenylation Capacity in Tobacco
It is well known that Me-JA induces both the production of MVA- and MEP pathway-derived metabolites [22,42,43]. With this in mind, we hypothesized that Me-JA may affect the metabolic flux leading to the biosynthesis of prenyl diphosphates used to modify proteins. To test this hypothesis, we analyzed the subcellular localization of GFP-CaM-CVIL in tobacco (Nicotiana tabacum L. var. xanthi) plants expressed under the control of a dexamethasone-inducible promoter. Tobacco plants can be elicited in order to induce isoprenoid metabolite production. We established experimental conditions by using cellulase-induced elicitation, which enhances the MVA biosynthesis via HMGR induction, ultimately leading the biosynthesis of capsidiol, a sesquiterpenoid phytoalexin [39]. Previously, we routinely used 0.5% cellulase to set up elicitation experiences [39], and tested here if the treatment possibly modifies the capacity of the prenylation process?
Similar to the GFP pattern of tobacco BY-2 cells or Arabidopsis leaves and roots, the untreated tobacco epidermic cells display the same plasma membrane labeling (Figure 3a,d). In this experiment, we opted to block both the MVA and MEP path7ways simultaneously in order to prevent the biosynthesis of isoprenoid precursors. Thus, we determined minimal concentrations of fosmidomycin (FOS/blocking the MEP pathway) and mevinolin (MV/blocking the MVA pathway) leading to the delocalization of GFP-CaM-CVIL into the nucleus. By evaluating the nuclear localization in combined FOS and MV treatments, we selected mevinolin (5 µM) and fosmidomycin (100 µM) as reference concentrations (FOS100/MV5) being sufficient to block protein prenylation in leaf disks. It has to be noted here that without inhibition of the MVA pathway, at 300 µM, fosmidomycin delocalize the protein into the nucleus, while mevinolin even at very high concentrations cannot. To test if stress increases the prenylation capacity in tobacco leaf disks, cellulase and Me-JA treatments were used to evaluate membrane localization in FOS100/MV5 treated tobacco leaf-disks (Figure 3b,3e). The membrane localization was restored when inhibition of the prenylation was combined with cellulase treatments (Figure 3c). In a similar manner, a treatment with Me-JA restored prenylation inhibition (Figure 3f). It is noteworthy that above the concentrations of FOS100/MV5, reversion does not work anymore, which points toward an increase of the metabolic pool used to prenylate the protein.
These results suggest that restoring of protein prenylation is due to an excess of prenyl diphosphate substrate generated by the stress, which would allow to compete with the inhibition and permit membrane localization again. Cellulase-induced elicitation leads to stimulation of HMGR synthesis and activity, and produced MVA is used for the inducible production of metabolites [39]. We therefore hypothesized that MVA production could act as a signal to reverse inhibition by fosmidomycin in tobacco BY-2 cells (Figure 1).
2.4. GFP-CaM-CVIL Prenylation under Restrictive Conditions Is Correlated with Mobilization of the MVA Pathway
Next, to support our assertions, we examined the effect of another compound known to boost HMGR activity by acting as an elicitor. As mentioned earlier, Fol has been described as a chemical stimulating HMGR activity both at a transcriptional and translational level in tobacco BY-2 cells [37]. This results in the rise of MVA levels, the enzyme product. At the same time, Fol is a substrate of specific kinases catalyzing the formation of FPP in the realm of the FPP salvage pathway [30]. In addition, Fol can overcome fosmidomycin-induced effect in tobacco BY-2 cells, but not when cells were inhibited with fosmidomycin and mevinolin simultaneously [38]. On this basis, we tested the ability of Fol to induce membrane localization of GFP-CaM-CVIL expressed in Arabidopsis ggb lines, where only PFT is active (Figure 4a). At 20 µM, Fol revived partially membrane localization of GFP-CaM-CVIL protein in the Arabidopsis ggb background. On the other hand, in geranylgeraniol (GGol, 20 µM)-treated seedlings the localization remained nuclear (Figure 4a). This implies that in the presence of Fol, PFT, which is the sole type-I PPT enzyme being active in an Arabidopsis ggb background, is recognizing GFP-CaM-CVIL as a protein substrate. It can be proposed here, that Fol is not directly used as a substrate, but rather the compound induces the production of MVA by activation of HMGR [37,38].
To evaluate if possibly an increase in metabolic pool might interfere with the prenylation capacity, we carried out PPT enzyme assays using cell-free protein extracts isolated from cells previously pretreated with different metabolites: MVA or Fol to mimic an increase in the MVA pathway, and GGol or 1-deoxy-D-xylulose (DX) to mimic an increase in MEP-derived metabolites (Figure 4b). An increase in “cold” prenyl diphosphate substrates is responsible for diluting the prenylation capacity of the enzyme activity, which is based on the use of radioactive substrates. Figure 4b illustrates this isotopic dilution. It can be observed that the metabolic contribution of all added chemicals decreases the prenylation capacity of crude extracts in the presence of radiolabeled FPP, but also in the presence of radiolabeled GGPP (Figure 4b). It is therefore not possible to conclude whether the protein is modified by FPP or GGPP. However, overall, the activity of an FPP-mediated modification of GFP-CaM-CVIL remains higher than that of GGPP-mediated GFP-CaM-CVIL in protein extracts isolated from 24h tobacco cells (Figure 4b), pointing towards a possible modification with MVA-derived FPP.
To further investigate the potential role of MVA in modifying PFT behavior in plants under stress conditions, we first checked whether MVA production had been promoted under our experimental conditions. Thus, we estimated the production of phytosterols, which are the main MVA-derived metabolites produced in tobacco cells [44]. Cells were prepared as described in Figure 1b and a total sterol fraction (saponified fraction) has been isolated and quantified by GC-FID (Figure 5a). Under those growth conditions, no significant increase in sterol production could be observed. In fact, a slight drop in sterols levels is observed in the presence of MeJA. To understand this incoherent pattern, we quantified HMGR levels expressed in these same cells. HMGR protein quantification was realized by western-blotting using microsomal fractions isolated from cells grown as described in Figure 1b and an antibody raised against the catalytical domain of Nt-HMGR2 [39]. Again, in 24h-old cells, regardless of treatment, the level of HMGR was not significantly different (Figure 5b). We then speculated that HMGR levels have already been stimulated and tested if subculturing one week-old cells into new MS-medium could possibly induce HMGR production and mask probable Me-JA effects (Figure 5c). Interestingly, the amount of HMGR is strongly induced after 24-h and gradually decreases over time, consistent with the quantification of enzyme activity realized previously [27]. Results described in Figure 1a-c suggest that Me-JA does not stimulate the accumulation of HMGR protein in tobacco BY-2 cells and does not promote an MVA-derived accumulation of total phytosterols sterols under those growth conditions. That would mean that the signal we are trying to identify may be independent of the global HMGR protein stimulation.
If MeJA does not induce HMGR accumulation, it can be hypothesized that MVA is maybe better incorporation into prenylated protein in the presence of the phytohormone? Thus, we examined whether, after treatment with Me-JA, radiolabeled-MVA is more efficiently incorporated into proteins separated on an SDS-PAGE (Figure 5d). A slight increase in the radiolabeled signal was indeed observed. More interestingly, MVA re-established membrane localization in ggb::CVIL plants, just like what we observed after treating plantlets with Me-JA (Figure 5e). Also, the supplementation with DX, a precursor of the MEP pathway, induces membrane localization to some extent (Figure 5e). It has however to be mentioned here that the incorporation of DX into the MEP pathway depends upon the activity of a xylulose kinase catalyzing the formation of DXP [45]. Thus, comparing effects of MVA and DX can be misleading. Nevertheless, according to these observations, at least an increased MVA concentrations can favor reactivation of protein prenylation in ggb mutant lines and modify PFT selectivity for GFP-CaM-CVIL.
To foster the idea that MVA is a link to enable prenylation by Me-JA, we treated reversed tobacco BY-2 cells (FOS + Me-JA) with mevinolin (MV, 10 µM), a competitive inhibitor of HMGR, blocking MVA production. Co-treatments restored protein localization back into the nucleus, as observed with FOS alone (Figure 6a). This result confirms that cells, in the presence of Me-JA, used an MVA-derived precursor when GGPP was no longer available to modify GFP-CaM-CVIL. MV is unable to inhibit the prenylation of GFP-CaM-CVIL, this in the presence or absence of Me-JA, indicating that the MEP needs to be blocked in order to force tobacco cells to utilize the MVA pathway for the biosynthesis of prenyl diphosphate substrates (Figure 6a). Finally, we tested if MVA could act in synergy with Me-JA in re-establishing membrane localization of fosmidomycin/mevinolin-treated tobacco BY-2 cells (Figure 6b). We blocked both isoprenoid biosynthesis pathway, to be sure to block all endogenous substrate supplies. A concentration range of MVA from 50 to 1000 µM has been tested in the presence (+) or absence (-) of Me-JA (20 µM). Up to 200 µM, Me-JA acts in synergy with MVA. At MVA concentrations ≥ 500 µM, MVA alone is capable of reverting the isoprenoid biosynthesis inhibition (Figure 6b). Overall, our observations allude that the heightened activation of the MVA pathway might be a trigger for altering the substrate specificity of PFT observed in the presence of Me-JA.
3. Discussion
3.1. Modulation of PFT Substrate Specificities by Me-JA
The results presented in this article lead to the conclusion that Me-JA treatment tunes PFT protein substrate selectivity, by allowing an MVA-dependent prenylation of a substrate typically assigned to the activity of PGGT. This implies that the recognition of a prenylation motif dedicated to an enzyme is extended under specific physiological conditions requiring well-defined signals. However, at this point, it cannot be excluded that another prenyltransferase may be activated to catalyze the modification. Indeed, other types of protein prenylation have been described in eukaryotes. A second class of enzymes (PGGT-II; EC 2.5.1.60) catalyzes the modification of RAB proteins on two C-terminus cysteines using 2 geranylgeranyl groups [2]. Very recently, a third class of enzymes have been described in animals. It is proposed that this family catalyzes the transfer of a geranylgeranyl onto pre-liminary farnesylated proteins such as YKT6 (UniProtKB-O15498) or FBXL2 (UniProtKB-Q9UKC9) both imposing an additional cysteine creating a CCaaX motif [46,47].
Jasmonates are linolenic acid-derived phytohormones acting as elicitors inducing defense responses [48]. This phytohormone impacts the activity of different kind of enzymes, mainly by stimulating their gene expression [49,50]. Me-JA is known to induce the production of various metabolites, including isoprenoid compounds. This is how triterpene saponins are induced in Medicago truncatula [51], the sesquiterpene artemisinin in Artemisia annua L [52] or monoterpenes in Picea glauca [53]. Together, specialized metabolism is activated, while ROS production can be circumscribed and growth controlled [54,55,56]. Furthermore, in Catharanthus roseus, the production of monoterpene indole alkaloids (MIA) induced by the jasmonate signaling pathway, is controlled by protein geranylgeranylation [57,58], with a paramount contribution from the plastidial CrGGPPS2 [59]. In a similar way, MEP-derived protein prenylation is mandatory for the production of MVA-derived sesquiterpenoid phytoalexins after elicitation of tobacco plants with cellulase [39]. These studies demonstrate a clear correlation between induction of the production of specialized metabolites required for defense responses and protein prenylation in plants.
Although knowledge remains limited, the regulation of eukaryotic protein prenylation in is better characterized in human, animals and yeast than in plants [21]. Yet, it has been described that , on a molecular level, a tryptophane at position 110 should be important for selectivity against GGPP (https://www.uniprot.org/uniprotkb/Q38920/entry). Because PPTs have to process a multitude of substrates, it is essential that catalytical properties are precisely controlled within the cell. It is accepted that in vitro, the recombinant Arabidopsis PFT recognizes GFP-CaM-CVIL, the protein substrate known as a PGGT-specific substrate, in similar affinity ranges than its dedicated substrate (GFP-CaM-CVIM), but the turn-over is seven times lower [13]. Enzyme activity regulations in planta, have so far not been characterized. Here, we demonstrated that GFP-CaM-CVIL stays unmodified when expressed in the ggb mutant line where only PFT is active. Thus, PFT appears as being unable to modify GFP-CaM-CVIL under standard growth conditions in Arabidopsis and shows higher substrate selectivity in planta than in vitro. However, the capacity of PFT to modify GFP-CaM-CVIL can apparently be initiated following Me-JA treatment. Therefore, it is possible that the microenvironment of enzymes is highly dynamic and requires constant readjustment. At this point, it can be proposed that MeJA modulates the cellular environment controlling PFT activity and substrate specificity. Jasmonate-mediated signal responses are known to involve and modulate post-translational modifications (PTMs) [60]. In this context, we can mention phosphorylation in the common the α-subunit increasing enzyme activity [21]. For example, there is evidence that human enzyme activity can be enhanced by insulin, following phosphorylation of serine residues in the α-subunit [21]. Our case study does not provide clear indications that PFT activity has been stimulated, but rather that substrate recognition has been modified. Currently, it is unknown whether PTMs can modulate substrate specificity and no investigations have not been conducted for any enzyme yet.
3.2. MVA as a Sensor for Cross-Talk between MVA and MEP Pathway?
In this study, it has been observed that the substrate specificity of PFT can be modulated under stress conditions, with Me-JA serving as an enhancer and MVA possibly being a central metabolite connecting both. It appears that MVA is a key component in the modification of the capacity to modify GFP-CaM-CVIL proteins, however it cannot be excluded at this point that such functions could be assigned to MVA-derived metabolites.
MVA is a central compound to enhance the biosynthesis of isoprenoid compounds [61], also for supposedly MEP-derived metabolites [22,34,62,63]. In plants, the number of different isoforms of HMGRs responsible for MVA biosynthesis can reach almost 10 different proteins [64], most of them with specific functions and different spatio-temporal patterns. The enhancement of a particular HMGR isoform, which is devoted to protein prenylation, may not be quantifiable through western-blotting when employing an antibody that recognizes all forms. The existence of a MVA metabolon being carefully regulated has been proposed [65,66]. Those dynamic supramolecular complexes would drive a specific MVA-derived metabolite production under particular conditions, by substrate channeling. MVA has even functions beyond isoprenoid biosynthesis, like sugar and amino acids uptake in human cells [67]. Regulation of the MVA biosynthesis pathway is also related to energy sensors. For instance, extracellular ATP activates the metabolism [68], while the energy sensor AKIN10-GRIK1 inhibits HMGR via protein phosphorylation [69]. If MVA by itself can be sensed has so far not been described, but it could be a conceivable possibility in the context of protein prenylation regulation.
Nevertheless, the implication of MVA in the regulation of plant protein prenylation is undeniable. MVA changes in protein profile of small GTP-binding proteins [70]. In the same molecular range, we observed a slight increase in MVA incorporation in MeJA-treated cells. Such proteins are involved in defense signal-transduction pathways [71]. It should be pointed out, however, that not all of those proteins are prenylated. The importance of membrane localization of regulatory element in phytohormone signaling pathways have been described. This is for instance the case of myristoylated calcium-dependent protein kinases [72]. Protein prenylation should be crucial upstream of jasmonate signaling as VIGS-silencing of the PGGT β-subunit in Catharanthus cells and plants negatively impacts the expression of Me-JA-induced transcription factors [58,59]. More recently, it has been proposed that some Catharanthus Rho of Plant (ROP) proteins acts as transcriptional activators within the nucleus [73].
3.3. A Working Model of How Plants Benefit from Modifying PFT Substrate Selectivity
The fact that, in the presence of Me-JA, MVA-derived prenyl diphosphate is used to modify GFP-CaM-CVIL sensor proteins raises a number of interrogations. One question at hand is whether it is advantageous to modify the substrate specificity of a PPT? It is likely that the importance of protein prenylation in plant stress responses [2], is part of the answer. Because enzyme-catalyzed reactions are limited by substrate supply, such a strategy involving two enzymes and two metabolic pathways, could promote more efficiently biocatalysis by requisitioning substrates from a wider range of sources. Thus, by using a substrate generated by the MVA pathwa, rather than, or in addition, to the MEP pathway to modify a CaaL-motif protein important for stress-response, would make it possible to do it more effectively.
We propose a model for prenylation of GFP-CaaL proteins, in which a stress-induced signal modifies PFT protein substrate specificity (Figure 7). In unstressed tissues, PGGT is the active enzyme responsible in modifying CaaL-like proteins, using a plastidial MEP-derived prenyl diphosphate group (GGPP). This assessment is supported by the nuclear localization of GFP-CaM-CVIL expressed in Arabidopsis ggb plants and the loss of the membrane localization after fosmidomycin-treatments.
The stress-induced signal might be correlated with an increase in the amount of MVA or an MVA-derived metabolite. It is anticipated that under stress conditions, PPTs become less stringent to dedicated protein substrates, which would increase the capacity to modify proteins needed in signaling pathways supporting stress responses. Abiotic stresses related to bites or wounding induce some signals leading potentially in Me-JA production that will modify PFT substrate specificity. As a result, both PFT and PGGT can modify the same protein, which result in an increase of the prenylation capacity and potentially in the way the protein is modified (FPP vs. GGPP).
4. Materials and Methods
4.1. Chemical Materials
All-trans-farnesol, all-trans-geranylgeraniol, farnesyl diphosphate, geranylgeranyl diphosphate, dexamethasone, fosmidomycin, methyl-jasmonate, ethephon and mevalonolactone were purchased from Sigma-Aldrich (Saint Quentin Fallavier). Cellulase RS was obtained from Yakult Pharmaceutical Industry (Japan) and mevinolin was a kind gift from Drs. M. Greenspan and A.W. Alberts (Merck Sharp and Dohme). Before use, lactone forms of mevinolin and mevalonolactone were converted into their free salt forms as described by [28]. 1-deoxy-D-xylulose (DX) was obtained from AlsaChim (Illkirch Graffenstaden, France). [1-3H]Farnesyl diphosphate (60 Ci/mmol) and [1-3H]geranylgeranyl diphosphate (60 Ci/mmol) used for protein prenyltransferase assays was purchased from American Radiolabeled Chemicals, Inc and (R,S)-[2-14C]mevalonolactone was purchased from Amersham. Analytical grade solvents were purchased from Sigma, Carlo Erba, or Fischer.
4.2. DNA Constructs, Biological Materials and Culture Conditions
Two sensors for protein prenylation have been used in this study: GFP-CaM-CVIL, a GFP protein fusion with the C-terminus side of rice Calmoduline 61 (GenBank: AAA98933) and GFP-ROP6-CSIL, a fusion protein of GFP with Rho of Plant ROP6 (GenBank: OAP00270). Both coding sequences have been cloned into pTA7001 and are under the control of a dexamethasone inducible promoter [74]. The pTA-GFP-CaM-CVIL construct was generated as previously indicated [28]. To construct pTA-GFP-ROP6-CSIL, rop6 has been amplified by PCR using using ROP6-F 5’-GCAGAGCTCAGTGCTTCAAGGTTTATCAAG-3’ and ROP6-R 5’-TCTTCTAGATTCAGAGTATAGAACAACCTTTCTGAGATTTTCTC-3’ as a primer set and using as template, cDNA prepared from Arabidopsis seedlings [45]. The fragment was cloned between SacI and XbaI restriction sites, in frame of GFP of the pMRC-GFP vector [75]. The XhoI and SpeI restriction sites were incorporated by PCR using the primer set XhoIGFP-F 5’- CCGCTCGAGGGATGGTGAGCAAGGGCGAGGAGC-3’ and SpeIROP6-R GGACTAGTCCTCAGAGTATAGAACAACCTTTC-3’ to subclone GFP-ROP6-CSIL into pTA7001.
Suspension-cultured tobacco (Nicotiana tabacum) cv. BY-2 cells were maintained in modified Murashige and Skoog medium as previously described [76]. The generation of transgenic tobacco BY-2 cells expressing GFP-CaaL proteins under the control of the inducible dexamethasone promoter were obtained by co-culturing 10 days-old calli with pTA-GFP-CaM-CVIL or pTA-GFP-ROP6-CSIL-transformed Agrobacterium tumefaciens LBA4404.pBBR1MCS-5.virGN54D strain [77]. After a selection of hygromycin-resistant calli (30 µM), a homogenous cell suspension was regenerated. Protein expression is induced with 15 µM dexamethasone.
All Arabidopsis thaliana mutants and transgenic lines are in the Columbia-0 (Col0) ecotype background. After ethanol and bleach-sterilization, seeds were set to germinate on half MES-Murashige and Skoog medium supplemented with 2% sucrose and 0.8% agar. Transformed Col0 plants(Col0::GFP-CaM-CVIL) and T-DNA loss of function mutant ggb-2 plants [7] (ggb::GFP-CaM-CVIL) expressing GFP-CaM-CVIL, were obtained from Prof. D.N. Crowell (Idaho State University, Pocatello, USA). Lines have been transformed by floral dip with pTA-GFP-CaM-CVIl, construct. The T-DNA insertion farnesyltransferase β-subunit loss of function era1-9 mutant was obtained from the Arabidopsis Biological Resource Center under the reference SALK_110517 (https://www.arabidopsis.org/servlets/TairObject?type=polyallele&id=500194245). Because, transformation by floral dip was inefficient, era1.9::pTA-GFP-CaM-CVIL was obtained by crossing era1.9 with WT::pTA-GFP-CaM-CVIL. For transformation of era1.9, Col0::GFP-CaM-CVIL and era1.9 plantlets were transferred to soil and grown under short days conditions (16-h-light (22°C)/8-h-dark (18°C) photoperiod). Father era1.9 plants were crossed with mother Col0::GFP-CaM-CVIL plants and hygromycin (30 µM)-resistant plants were screened for GFP expression after induction with dexamethasone (30 µM). Transformed homozygote era1 plants were selected by PCR genotyping using era5F 5’-ACCTACTGTGGTTTGGCTGC-3’, era5R 5’-CAACAACGGGTCATGCTGCT-3’ and Lb1b 5’-TGGCAGGATATATTGTGGTG -3’.
Tobacco (Nicotiana tabacum) wild-type (var. xanthi line SH6) was grown as axenic shoot culture on hormone-free MS-medium (Duchefa, The Netherlands). Transformation was conducted by co-cultivation Agrobacterium tumefaciens GV3101 harboring the construct pTA-GFP-CaM-CVIL and 5-6 weeks old leaf-disks [78]. Following dexamethasone induction (30 µM), three independent-hygromycin-resistant (30 µM) transformed F1 generation plants were selected by confocal microscopy subsequent analyses for fluorescence emission.
4.3. Microscopy Analyses
For tobacco microscopy analyses, GFP-CaaL protein are expressed by adding dexamethasone (15 µM for tobacco cells, 30 µM for plantlets, during 18 h) to biological material pretreated with chemicals (3 h for tobacco cells, 6 hours for plantlets). This step is necessary to observe GFP-CaaL prenylation statues according to isoprenoid pool availabilities. Tobacco leaf disks (1-cm diameter) were punched from 5-6 weeks-old plants and positioned adaxial side upward onto MS-medium water and treated as indicated in Figure 3. Axenic Arabidopsis 2-leaves plantlets were transferred to MS-medium and treated for 24 h in 12-wells plates, before protein expression is induced by adding 200 µL of MS-medium supplemented with 30 µM dexamethasone. Tobacco BY-2 cells (>300) were analyzed in two independent measurements and classified by visual selection into the following categories: plasma membrane-localized, nucleus-localized, or both. The efficiency of cellular-treatments with inhibitors was evaluated using chi-squared tests against the null hypothesis being true. Significant differences were assigned to p-values <0.01. Acquisitions of images were realized with a Zeiss LSM700 confocal microscope equipped with a Plan-Apochromat 20x/0.8 M27 objective using a 488 nm laser (15%) for excitation. Fluorescence emission was revealed by using the SP555 filter and the Main Dichroic beam splitter MBS 405/488/555/639. Acquisitions were handled using ZEN 2009 software (Carl Zeiss; http://www.zeiss.com/), exported as Tagged Image Files and processed with Adobe Photoshop 5.0 software (Adobe Systems).
4.4. Phytosterol Analyses
Phytosterol extraction were realized as previously described by [79] using 100 mg (dry weight) of tobacco BY-2 cells. Experiences were realized in biological triplicates and sterols were quantified using α-cholestane as a standard and calculated as a percentage taking the mean of total phytosterol content in control cells as a reference.
4.5. Protein Quantification, Enzyme Assays and Fluorography
Microsomal protein fractions prepared as previously described [76]. Protein concentration was quantified by a Bradford method using BSA as a standard and the manufacturer’s protocol (BioRad, France). HMGR contained in 40 µg was revealed and quantified by western blotting using a polyclonal rabbit antibody raised against the soluble domain of N. tabacum HMGR2 [39]. Protein prenyltransferase enzyme assays using recombinant GFP-CaM-CVIL or GFP-CaM-SVIL were earlier described [39]. For the fluorogram, tobacco BY-2 cells (500 µL of 1 week-old cells diluted 5-times in MS-medium) were incubated with 1 µCi of [14C]MVA for 24-h at 22°C, on a tube rotator . Cells were recovered by centrifugation and washed twice with PBS buffer. Proteins were extracted with 200 µL TCA (10%) and precipitated by adding 800 µL cold acetone 80%. The pellet obtained was dried under vacuum and recovered in 75 µL of Laemmli loading buffer. A 12 % SDS-PAGE gel was loaded with mixture. The fixed gel was activated with Amplify from Amersham and exposed to a preflashed X-ray film for 1 month, before being scanned and proceeded with Adobe Photoshop 5.0 software (Adobe Systems).
5. Conclusions
Here, we demonstrated that PPT substrate specificity is modulable under phytohormonal orchestration in planta by a so far unknown mechanism. This study provided evidence that Me-JA impairs the ability of PFT, an enzyme known to transfer farnesyl moieties, to specifically use a protein substrate typically dedicated to its sister enzyme PGGT-I. These results suggest that prenylated proteins might possibly be modified by different prenyl groups depending on the physiological conditions, the plant growth. The possibility to regulate PPT activities and substrate specificities in planta offers a way to interplay with the use of two pathways for prenyl diphosphate biosynthesis, with MVA conceivably acting as a sensor metabolite underpinning this dialog. In addition, this pool-driven flexibility of enzyme substrate specificity, may be essential in plant stress responses. Me-JA is an important inducer employed to produce valuable secondary metabolites in plants [80]. Understanding the regulation of protein prenylation will provide a better understanding of plants' ability to produce valuable specialized metabolites and identify new strategies for their accumulation, for instance, by using both isoprenoid biosynthesis pathways.
Author Contributions
Conceptualization, QC, Al.H., T.J.B. and An.H.; methodology, Q.C., Al.H., P.D., P.M., M.H. and An.H.; validation, Q.C., Al.H. and An.H.; formal analysis, An.H..; investigation, Q.C., Al.H., P.D., P.M., M.H., An.H..; data curation, An.H.; writing—original draft preparation, An.H..; writing—review and editing, T.J.B., H.S. and An.H..; visualization, Q.C., Al.H., P.D., An.H..; supervision, C.V.S., T.J.B., H.S. and An.H..; project administration, An.H..; funding acquisition, C.V.S., T.J.B., H.S. and An.H.
Funding
This research was funded by the Centre National de la Recherche Scientifique (C.N.R.S.), the Université de Strasbourg (UdS) and received no extra external funding. The Microscopy and cellular imaging platform was supported by the CNRS, Université de Strasbourg, Région Alsace, and Association pour la Recherche sur le Cancer for equipment cofinancing. QC, AlH and PD were supported by a predoctoral fellowship (Contrat Doctoral) from the French Ministry of higher Education, Research and Innovation.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank the late Prof. Dring N. Crowell for transformation of Col0 and ggb-2 lines and Monique Schmitz for her help in Nicotiana tabacum plant transformation. We are grateful to Prof. N. Chua (Rockefeller University, New York, NY) for the pTA7001 vector.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fu, H.W.; Casey, P.J. Enzymology and biology of CaaX protein prenylation. Recent Prog. Horm. Res. 1999, 54, 315–342. [Google Scholar]
- Hála, M.; Žárský, V. Protein prenylation in plant stress responses. Molecules 2019, 24, 3906. [Google Scholar] [CrossRef] [PubMed]
- Hemsley, P.A. The importance of lipid modified proteins in plants. New Phytol. 2015, 205, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.; Ghassemian, M.; Bonetta, D.; Cooney, S.; McCourt, P. A protein farnesyl transferase involved in abscisic acid signal transduction in Arabidopsis. Science 1996, 273, 1239–1241. [Google Scholar] [CrossRef] [PubMed]
- Pei, Z.M.; Ghassemian, M.; Kwak, C.M.; McCourt, P.; Schroeder, J.I. Role of farnesyltransferase in ABA regulation of guard cell anion channels and plant water loss. Science 1998, 282, 287–290. [Google Scholar] [CrossRef]
- Running, M.P.; Lavy, M.; Sternberg, H.; Galichet, A.; Gruissem, W.; Hake, S.; Ori, N.; Yalovsky, S. Enlarged meristems and delayed growth in plp mutants result from lack of CaaX prenyltransferases. Proc. Natl. Acad. Sci. USA 2004, 101, 7815–7820. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.D.; Chary, S.N.; Chernoff, E.A.; Zeng, Q.; Running, M.P.; Crowell, D.N. Protein geranylgeranyltransferase I is involved in specific aspects of abscisic acid and auxin signaling in Arabidopsis. Plant Physiol. 2005, 139, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Running, M.P.; Fletcher, J.C.; Meyerowitz, E.M. The WIGGUM gene is required for proper regulation of floral meristem size in Arabidopsis. Development 1998, 125, 2545–2553. [Google Scholar] [CrossRef]
- Bonetta, D.; Bayliss, P.; Sun, S.; Sage, T.; McCourt, P. Farnesylation is involved in meristem organization in Arabidopsis. Planta 2000, 211, 182–190. [Google Scholar] [CrossRef]
- Yalovsky, S.; Rodríguez-Concepción, M.; Bracha, K.; Toledo-Ortiz, G.; Gruissem, W. Prenylation of the floral transcription factor APETALA1 modulates its function. Plant Cell 2000, 12, 1257–1266. [Google Scholar] [CrossRef]
- Ziegelhoffer, E.C.; Medrano, L.J.; Meyerowitz, E.M. Cloning of the Arabidopsis WIGGUM gene identifies a role for farnesylation in meristem development. Proc. Natl. Acad. Sci. USA 2000, 97, 7633–7638. [Google Scholar] [CrossRef] [PubMed]
- Vergès, V.; Dutilleul, C.; Godin, B.; Collet, B.; Lecureuil, A.; Rajjou, L.; Guimaraes, C.; Pinault, M.; Chevalier, S.; Giglioli-Guivarc’h, N.; Ducos, E. Protein farnesylation takes part in Arabidopsis seed development. Front. Plant Sci. 2021, 12, 620325. [Google Scholar] [CrossRef]
- Andrews, M.; Huizinga, D.H.; Crowell, D.N. The CaaX specificities of Arabidopsis protein prenyltransferases explain era1 and ggb phenotypes. BMC Plant Biol. 2010, 10, 118. [Google Scholar] [CrossRef]
- Barghetti, A.; Sjögren, L.; Floris, M.; Paredes, E.B.; Wenkel, S.; Brodersen, P. Heat-shock protein 40 is the key farnesylation target in meristem size control, abscisic acid signaling, and drought resistance. Genes Dev. 2017, 31, 2282–2295. [Google Scholar] [CrossRef] [PubMed]
- Brady, S.M.; Sarkar, S.F.; Bonetta, D.; McCourt, P. The ABSCISIC ACID INSENSITIVE 3 (ABI3) gene is modulated by farnesylation and is involved in auxin signaling and lateral root development in Arabidopsis. Plant J. 2003, 34, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.R.; Wang, L.C.; Lin, Y.R.; Weng, C.P.; Yeh, C.H.; Wu, S.J. The Arabidopsis heat-intolerant 5 (hit5)/enhanced response to aba 1 (era1) mutant reveals the crucial role of protein farnesylation in plant responses to heat stress. New Phytol. 2017, 213, 1181–1193. [Google Scholar] [CrossRef]
- Yalovsky, S.; Kulukian, A.; Rodríguez-Concepción, M.; Young, C.A.; Gruissem, W. Functional requirement of plant farnesyltransferase during development in Arabidopsis. Plant Cell 2000, 12, 1267–1278. [Google Scholar] [CrossRef]
- Bracha-Drori, K.; Shichrur, K.; Lubetzky, T.C.; Yalovsky, S. Functional analysis of Arabidopsis postprenylation CaaX processing enzymes and their function in subcellular protein targeting. Plant Physiol. 2008; 148, 119–143. [Google Scholar] [CrossRef]
- Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A. Ergosterol-induced sesquiterpenoid synthesis in tobacco cells. Molecules 2012, 17, 1698–1715. [Google Scholar] [CrossRef]
- Palsuledesai, C.C.; Distefano, M.D. Protein prenylation: enzymes, therapeutics, and biotechnology applications. ACS Chem. Biol. 2015, 10, 51–62. [Google Scholar] [CrossRef]
- Jung, D.; Bachmann, H.S. Regulation of protein prenylation. Biomed. Pharmacother. 2023, 164, 114915. [Google Scholar] [CrossRef]
- Hemmerlin, A.; Harwood, J.L.; Bach, T.J. A raison d'être for two distinct pathways in the early steps of plant isoprenoid biosynthesis? Prog. Lipid Res. 2012, 51, 95–148. [Google Scholar] [CrossRef] [PubMed]
- Randall, S.K.; Marshall, M.S.; Crowell, D.N. Protein isoprenylation in suspension-cultured tobacco cells. Plant Cell 1993, 5, 433–442. [Google Scholar] [CrossRef]
- Morehead, T.A.; Biermann, B.J.; Crowell, D.N.; Randall, S.K. Changes in protein isoprenylation during the growth of suspension-cultured tobacco cells. Plant Physiol. 1995, 109, 277–284. [Google Scholar] [CrossRef]
- Swiezewska, E.; Thelin, A.; Dallner, G.; Andersson, B.; Ernster, L. Occurrence of prenylated proteins in plant cells. Biochem. Biophys. Res. Commun. 1993, 192, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Galichet, A.; Gruissem, W. Developmentally controlled farnesylation modulates AtNAP1; 1 function in cell proliferation and cell expansion during Arabidopsis leaf development. Plant Physiol. 2006, 142, 1412–1426. [Google Scholar] [CrossRef]
- Hemmerlin, A.; Hoeffler, J.-F.; Meyer, O.; Tritsch, D.; Kagan, I.A.; Grosdemange-Billiard, C.; Rohmer, M.; Bach, T.J. Crosstalk between the cytosolic mevalonate and the plastidial methylerythritol phosphate pathways in tobacco Bright Yellow-2 cells. J. Biol. Chem. 2003, 278, 26666–26676. [Google Scholar] [CrossRef]
- Gerber, E.; Hemmerlin, A.; Hartmann, M.; Heintz, D.; Hartmann, M.A.; Mutterer, J.; Rodríguez-Concepción, M.; Boronat, A.; Van Dorsselaer, A.; Rohmer, M.; Crowell, D.N.; Bach, T.J. The plastidial 2-C-methyl-D-erythritol 4-phosphate pathway provides the isoprenyl moiety for protein geranylgeranylation in tobacco BY-2 cells. Plant Cell 2009, 21, 285–300. [Google Scholar] [CrossRef]
- Hartmann, M.; Hemmerlin, A.; Gas-Pascual, E.; Gerber, E.; Tritsch, D.; Rohmer, M.; Bach, T.J. The effect of MEP pathway and other inhibitors on the intracellular localization of a plasma membrane-targeted, isoprenylable GFP reporter protein in tobacco BY-2 cells. F1000Res. 2013, 2, 170. [Google Scholar] [CrossRef] [PubMed]
- Hemmerlin, A. Phosphorylation of metabolites involved in salvage pathways for isoprenoid biosynthesis in plants. Kinases Phosphatases 2023, 1, 151–166. [Google Scholar] [CrossRef]
- Nagel, R.; Schmidt, A.; Peters, R.J. Isoprenyl diphosphate synthases: the chain length determining step in terpene biosynthesis. Planta 2019, 249, 9–20. [Google Scholar] [CrossRef]
- Swiezewska, E.; Danikiewicz, W. Polyisoprenoids: structure, biosynthesis and function. Prog. Lipid Res. 2005, 44, 235–258. [Google Scholar] [CrossRef]
- Lipko, A.; Swiezewska, E. Isoprenoid generating systems in plants—a handy toolbox how to assess contribution of the mevalonate and methylerythritol phosphate pathways to the biosynthetic process. Prog. Lipid Res. 2016, 63, 70–92. [Google Scholar] [CrossRef] [PubMed]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Wille, A.; Zimmermann, P.; Vranová, E.; Fürholz, A.; Laule, O.; Bleuler, S.; Hennig, L.; Prelić, A.; von Rohr, P.; Thiele, L.; Zitzler, E.; Gruissem, W.; Bühlmann, P. Sparse graphical Gaussian modeling of the isoprenoid gene network in Arabidopsis thaliana. Genome Biol. 2004, 5, R92. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Gas-Pascual, E.; Hemmerlin, A.; Rohmer, M.; Bach, T.J. Development of an image-based screening system for inhibitors of the plastidial MEP pathway and of protein geranylgeranylation. F1000Res. 2015, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Hemmerlin, A.; Bach, T.J. Farnesol induced cell death and stimulation of 3-hydroxy-3-methylglutaryl coenzyme A reductase activity in tobacco cv. Bright Yellow-2 cells. Plant Physiol. 2000, 123, 1257–1268. [Google Scholar] [CrossRef]
- Huchelmann, A.; Brahim, M.S.; Gerber, E.; Tritsch, D.; Bach, T.J.; Hemmerlin, A. Farnesol-mediated shift in the metabolic origin of prenyl groups used for protein prenylation in plants. Biochimie 2016, 127, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Huchelmann, A.; Gastaldo, C.; Veinante, M.; Zeng, Y.; Heintz, D.; Tritsch, D.; Schaller, H.; Rohmer, M.; Bach, T.J.; Hemmerlin, A. S-Carvone suppresses cellulase-induced capsidiol production in Nicotiana tabacum by interfering with protein isoprenylation. Plant Physiol. 2014, 164, 935–950. [Google Scholar] [CrossRef] [PubMed]
- Yalovsky, S. Protein lipid modifications and the regulation of ROP GTPase function. J. Exp. Bot. 2015, 66, 1617–1624. [Google Scholar] [CrossRef]
- Zhu, Z. Molecular basis for jasmonate and ethylene signal interactions in Arabidopsis. J. Exp. Bot. 2014, 65, 5743–5748. [Google Scholar] [CrossRef]
- Hampel, D.; Mosandl, A.; Wüst, M. Induction of de novo volatile terpene biosynthesis via cytosolic and plastidial pathways by methyl jasmonate in foliage of Vitis vinifera L. J. Agric. Food Chem. 2005, 53, 2652–2657. [Google Scholar] [CrossRef]
- Wei, Q.; Lan, K.; Liu, Y.; Chen, R.; Hu, T.; Zhao, S.; Yin, X.; Xie, T. Transcriptome analysis reveals regulation mechanism of methyl jasmonate-induced terpenes biosynthesis in Curcuma wenyujin. PLoS One 2022, 17, e0270309. [Google Scholar] [CrossRef]
- Wentzinger, L.F.; Bach, T.J.; Hartmann, M.A. Inhibition of squalene synthase and squalene epoxidase in tobacco cells triggers an up-regulation of 3-hydroxy-3-methylglutaryl coenzyme A reductase. Plant Physiol. 2002, 130, 334–346. [Google Scholar] [CrossRef]
- Hemmerlin, A.; Tritsch, D.; Hartmann, M.; Pacaud, K.; Hoeffler, J.-F.; Van Dorsselaer, A.; Rohmer, M.; Bach, T.J. A cytosolic Arabidopsis D-xylulose kinase catalyzes the phosphorylation of 1-deoxy-D-xylulose into a precursor of the plastidial isoprenoid pathway. Plant Physiol. 2006, 142, 441–457. [Google Scholar] [CrossRef]
- Kuchay, S.; Wang, H.; Marzio, A.; Jain, K.; Homer, H.; Fehrenbacher, N.; Philips, M.R.; Zheng, N.; Pagano, M. GGTase3 is a newly identified geranylgeranyltransferase targeting a ubiquitin ligase. Nat. Struct. Mol. Biol. 2019, 26, 628–636. [Google Scholar] [CrossRef]
- Shirakawa, R.; Goto-Ito, S.; Goto, K.; Wakayama, S.; Kubo, H.; Sakata, N.; Trinh, D.A.; Yamagata, A.; Sato, Y.; Masumoto, H.; Cheng, J.; Fujimoto, T.; Fukai, S.; Horiuchi, H. A SNARE geranylgeranyltransferase essential for the organization of the Golgi apparatus. EMBO J. 2020, 39, e104120. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates and octadecanoids: signals in plant stress responses and development. Prog. Nucleic Acid Res. Mol. Biol. 2002, 72, 165–221. [Google Scholar] [CrossRef]
- van der Fits, L.; Memelink, J. ORCA3, a jasmonate-responsive transcriptional regulator of plant primary and secondary metabolism. Science 2000, 289, 295–297. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, W.; Zhang, Y.; Zhang, X.; Lang, D.; Zhang, X. The roles of methyl jasmonate to stress in plants. Funct. Plant Biol. 2018, 46, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Mertens, J.; Pollier, J.; Vanden Bossche, R.; Lopez-Vidriero, I.; Franco-Zorrilla, J.M.; Goossens, A. The bHLH transcription factors TSAR1 and TSAR2 regulate triterpene saponin biosynthesis in Medicago truncatula. Plant Physiol. 2016, 170, 194–210. [Google Scholar] [CrossRef]
- Yu, Z.X.; Li, J.X.; Yang, C.Q.; Hu, W.L.; Wang, L.J.; Chen, X.Y. The jasmonate-responsive AP2/ERF transcription factors AaERF1 and AaERF2 positively regulate artemisinin biosynthesis in Artemisia annua L. Mol. Plant 2012, 5, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Perreca, E.; Eberl, F.; Santoro, M.V.; Wright, L.P.; Schmidt, A.; Gershenzon, J. Effect of drought and methyl jasmonate treatment on primary and secondary isoprenoid metabolites derived from the MEP pathway in the white spruce Picea glauca. Int. J. Mol. Sci. 2022, 23, 3838. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.T.; Murthy, H.N.; Park, S.Y. Methyl jasmonate induced oxidative stress and accumulation of secondary metabolites in plant cell and organ cultures. Int. J. Mol. Sci. 2020, 21, 716. [Google Scholar] [CrossRef] [PubMed]
- Sasaki-Sekimoto, Y.; Taki, N.; Obayashi, T.; Aono, M.; Matsumoto, F.; Sakurai, N.; Suzuki, H.; Yokota Hirai, M.; Noji, M.; Saito, K.; Masuda, T.; Takamiya, K.I.; Shibata, D.; Ohta, H. Coordinated activation of metabolic pathways for antioxidants and defence compounds by jasmonates and their roles in stress tolerance in Arabidopsis. Plant J. 2005, 44, 653–668. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.I.; Pandian, S.; Rakkammal, K.; Largia, M.J.V.; Thamilarasan, S.K.; Balaji, S.; Zoclanclounon, Y.A.B.; Shilpha, J.; Ramesh, M. Jasmonates in plant growth and development and elicitation of secondary metabolites: An updated overview. Front. Plant Sci. 2022, 13, 942789. [Google Scholar] [CrossRef] [PubMed]
- Courdavault, V.; Burlat, V.; St-Pierre, B.; Giglioli-Guivarc’h, N. Proteins prenylated by type I protein geranylgeranyltransferase act positively on the jasmonate signalling pathway triggering the biosynthesis of monoterpene indole alkaloids in Catharanthus roseus. Plant Cell Rep. 2009, 28, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Courdavault, V.; Thiersault, M.; Courtois, M.; Gantet, P.; Oudin, A.; Doireau, P.; St-Pierre, B.; Giglioli-Guivarc’h, N. CaaX-prenyltransferases are essential for expression of genes involved in the early stages of monoterpenoid biosynthetic pathway in Catharanthus roseus cells. Plant Mol. Biol. 2005, 57, 855–870. [Google Scholar] [CrossRef]
- Kumar, S.R.; Rai, A.; Bomzan, D.P.; Kumar, K.; Hemmerlin, A.; Dwivedi, V.; Godbole, R.C.; Barvkar, V.; Shanker, K.; Shilpashree, H.B.; Bhattacharya, A.; Smitha, A.R.; Hedge, N.; Nagegowda, D.A. A plastid-localized bona fide geranylgeranyl diphosphate synthase plays a necessary role in monoterpene indole alkaloid biosynthesis in Catharanthus roseus. Plant J. 2020, 103, 248–265. [Google Scholar] [CrossRef] [PubMed]
- Lacchini, E.; Goossens, A. Combinatorial control of plant specialized metabolism: mechanisms, functions, and consequences. Annu. Rev. Cell Dev. Biol. 2020, 36, 291–313. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.; Hemmerlin, A.; Bach, T.J.; Chye, M.L. The potential of the mevalonate pathway for enhanced isoprenoid production. Biotechnol. Adv. 2016, 34, 697–713. [Google Scholar] [CrossRef]
- Opitz, S.; Nes, W.D.; Gershenzon, J. Both methylerythritol phosphate and mevalonate pathways contribute to biosynthesis of each of the major isoprenoid classes in young cotton seedlings. Phytochemistry 2014, 98, 110–119. [Google Scholar] [CrossRef]
- Schneider, M.M.; Hampp, R.; Ziegler, H. Envelope permeability to possible precursors of carotenoid biosynthesis during chloroplast-chromoplast transformation. Plant Physiol. 1977, 1977. 60, 518–520. [Google Scholar] [CrossRef]
- Li, W.; Liu, W.; Wei, H.; He, Q.; Chen, J.; Zhang, B.; Zhu, S. Species-specific expansion and molecular evolution of the 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGR) gene family in plants. PLoS One 2014, 9, e94172. [Google Scholar] [CrossRef]
- Chappell, J. Biochemistry and molecular biology of the isoprenoid biosynthetic pathway in plants. Annu. Rev. Plant Biol. 1995, 46, 521–547. [Google Scholar] [CrossRef]
- Gutensohn, M.; Hartzell, E.; Dudareva, N. Another level of complexity: The role of metabolic channeling and metabolons in plant terpenoid metabolism. Front. Plant Sci. 2022, 13, 954083. [Google Scholar] [CrossRef]
- Gong, L.; Xiao, Y.; Xia, F.; Wu, P.; Zhao, T.; Xie, S.; Wang, R.; Wen, Q.; Zhou, W.; Xu, H.; Zhu, L.; Zheng, Z.; Yang, T.; Chen, Z.; Duan, Q. The mevalonate coordinates energy input and cell proliferation. Cell Death Dis. 2019, 10, 327. [Google Scholar] [CrossRef]
- Cho, S.H.; Tóth, K.; Kim, D.; Vo, P.H.; Lin, C.H.; Handakumbura, P.P.; Ubach, A.R.; Evans, S.; Paša-Tolić, L.; Stacey, G. Activation of the plant mevalonate pathway by extracellular ATP. Nat. Commun. 2022, 13, 450. [Google Scholar] [CrossRef]
- Robertlee, J.; Kobayashi, K.; Suzuki, M.; Muranaka, T. AKIN 10, a representative Arabidopsis SNF 1-related protein kinase 1 (SnRK1), phosphorylates and downregulates plant HMG-CoA reductase. FEBS lett. 2017, 591, 1159–1166. [Google Scholar] [CrossRef]
- Hemmerlin, A.; Brown, S.C.; Bach, T.J. Function of mevalonate in tobacco cell proliferation. Acta Bot. Gall. 1999, 146, 85–100. [Google Scholar] [CrossRef]
- Sano, H.; Ohashi, Y. Involvement of small GTP-binding proteins in defense signal-transduction pathways of higher plants. Proc. Natl. Acad. Sci. USA 1995, 92, 4138–4144. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Huang, W. Calcium-dependent protein kinases in phytohormone signaling pathways. Int. J. Mol. Sci. 2017, 18, 2436. [Google Scholar] [CrossRef]
- Bomzan, D.P.; Sharma, A.; Cruz, P.L.; Carqueijeiro, I.; Bellenger, L.; Rai, A.; Kumar Thippesh, A.; Venkatesha, S.C.; Parihar, D.; Ducos, E.; Courdavault, V.; Nagegowda, D.A. Rho of plant GTPases with geranylgeranylation motif modulate monoterpene indole alkaloid biosynthesis in Catharanthus roseus. bioRxiv 2023, 2023–09. [Google Scholar] [CrossRef]
- Aoyama, T.; Chua, N.H. A glucocorticoid-mediated transcriptional induction system in transgenic plants. Plant J. 1997, 11, 605–612. [Google Scholar] [CrossRef]
- Rodríguez-Concepción, M.; Yalovsky, S.; Zik, M.; Fromm, H.; Gruissem, W. The prenylation status of a novel plant calmodulin directs plasma membrane or nuclear localization of the protein. EMBO J. 1999, 1999. 18, 1996–2007. [Google Scholar] [CrossRef]
- Hemmerlin, A.; Bach, T.J. Effects of mevinolin on cell cycle progression and viability of tobacco BY-2 cells. Plant J. 1998, 14, 65–74. [Google Scholar] [CrossRef]
- van der Fits, L.; Deakin, E.A.; Hoge, J.H.C.; Memelink, J. The ternary transformation system: constitutive virG on a compatible plasmid dramatically increases Agrobacterium-mediated plant transformation. Plant Mol. Biol. 2000, 43, 495–502. [Google Scholar] [CrossRef]
- Gallois, P.; Marinho, P. Leaf disk transformation using Agrobacterium tumefaciens - expression of heterologous genes in tobacco. Methods Mol. Biol. 1995, 49, 39–48. [Google Scholar]
- Darnet, S.; Martin, L.; Mercier, P.; Bracher, F.; Geoffroy, P.; Schaller, H. Inhibition of phytosterol biosynthesis by azasterols. Molecules 2020, 25, 1111. [Google Scholar] [CrossRef]
- Namdeo, A.G. Plant cell elicitation for production of secondary metabolites: A review. Pharmacogn. Rev. 2007, 1, 69–79. [Google Scholar]
Figure 1.
Jasmonic acid methyl esther restores inhibition of GFP-CaaL protein prenylation by fosmidomycin. (a) Confocal pictures of control, fosmidomycin (100 µM) and fosmidomycin/MeJA (100 µM/20 µM) cells expressing GFP-CaM-CVIL under the control of dexamethasone inducible promoter. Seven days-old cells (stationary phase) were diluted 5 times in new MS medium and cultivated for 3 hours in the presence of inhibitors, before induction with 30 µM dexamethasone. (b) Cells expressing GFP-CaM-CVIL have been categorized in 2 classes: cells without a nucleus localization and cells in which nuclei are labelled. 3 independent set of experiments were performed where at least 300 localizations were evaluated. A box and Whisker plot was used to illustrate results. Groups with different letters have significantly different means (p ≤ 0.005) as calculated by a Bonferroni-Holm test following a one-way ANOVA. (c) Heat maps showing dose-dependent effect of Me-JA and the synergic effect with ethephon (d). The more the color is green, the better membrane localization is effective. Cells with membrane localizations (M), were count.
Figure 1.
Jasmonic acid methyl esther restores inhibition of GFP-CaaL protein prenylation by fosmidomycin. (a) Confocal pictures of control, fosmidomycin (100 µM) and fosmidomycin/MeJA (100 µM/20 µM) cells expressing GFP-CaM-CVIL under the control of dexamethasone inducible promoter. Seven days-old cells (stationary phase) were diluted 5 times in new MS medium and cultivated for 3 hours in the presence of inhibitors, before induction with 30 µM dexamethasone. (b) Cells expressing GFP-CaM-CVIL have been categorized in 2 classes: cells without a nucleus localization and cells in which nuclei are labelled. 3 independent set of experiments were performed where at least 300 localizations were evaluated. A box and Whisker plot was used to illustrate results. Groups with different letters have significantly different means (p ≤ 0.005) as calculated by a Bonferroni-Holm test following a one-way ANOVA. (c) Heat maps showing dose-dependent effect of Me-JA and the synergic effect with ethephon (d). The more the color is green, the better membrane localization is effective. Cells with membrane localizations (M), were count.
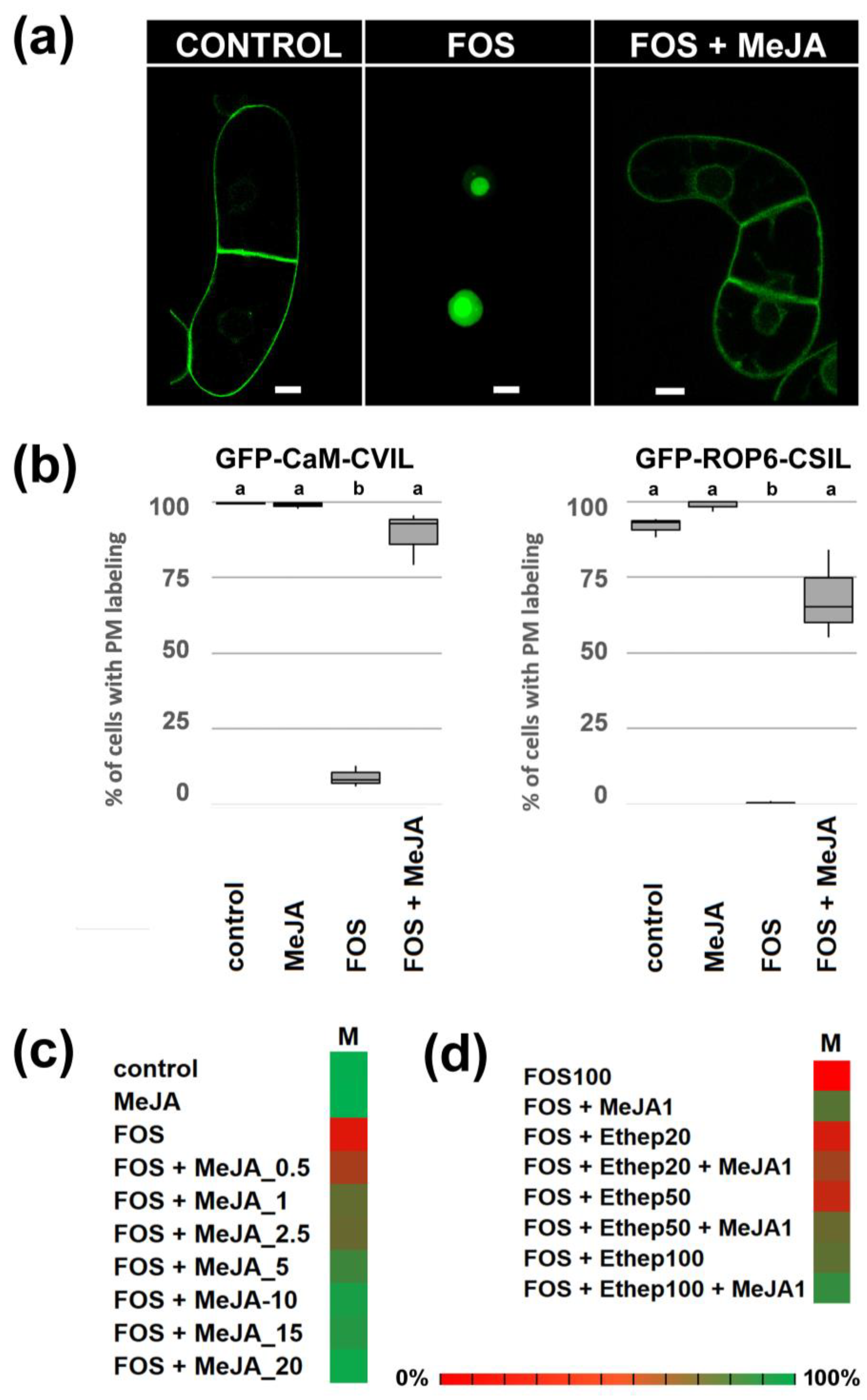
Figure 2.
Subcellular localization of GFP-CaM-CVIL protein expressed in Arabidopsis lines and activation of protein prenylation by Ma-JA in ggb lines. Col0, era1, and ggb lines have been transformed with pTA-GFP-CaM-CVIL constructs (CVIL) and leaves (green scare) or roots (yellow scare) have been observed by confocal microscopy. The WT Arabidopsis background retain both PFT and PGGT activities, while loss of function era1 lines, only PGGT-I is expressed, and in ggb lines only PFT remains active (green letters). In ggb::CVIL lines the protein cannot be modified, indicating that PFT alone cannot modify GFP-CaM-CVIL under standard growth conditions (red letters). In the presence of Me-JA, the membrane localization of the GFP modified protein is again observed in ggb plants, suggesting an active PFT (green letters). The white bar corresponds to 50 µm. On the left, the Arabidopsis plantlet has been created using biorender (https://www.biorender.com/).
Figure 2.
Subcellular localization of GFP-CaM-CVIL protein expressed in Arabidopsis lines and activation of protein prenylation by Ma-JA in ggb lines. Col0, era1, and ggb lines have been transformed with pTA-GFP-CaM-CVIL constructs (CVIL) and leaves (green scare) or roots (yellow scare) have been observed by confocal microscopy. The WT Arabidopsis background retain both PFT and PGGT activities, while loss of function era1 lines, only PGGT-I is expressed, and in ggb lines only PFT remains active (green letters). In ggb::CVIL lines the protein cannot be modified, indicating that PFT alone cannot modify GFP-CaM-CVIL under standard growth conditions (red letters). In the presence of Me-JA, the membrane localization of the GFP modified protein is again observed in ggb plants, suggesting an active PFT (green letters). The white bar corresponds to 50 µm. On the left, the Arabidopsis plantlet has been created using biorender (https://www.biorender.com/).
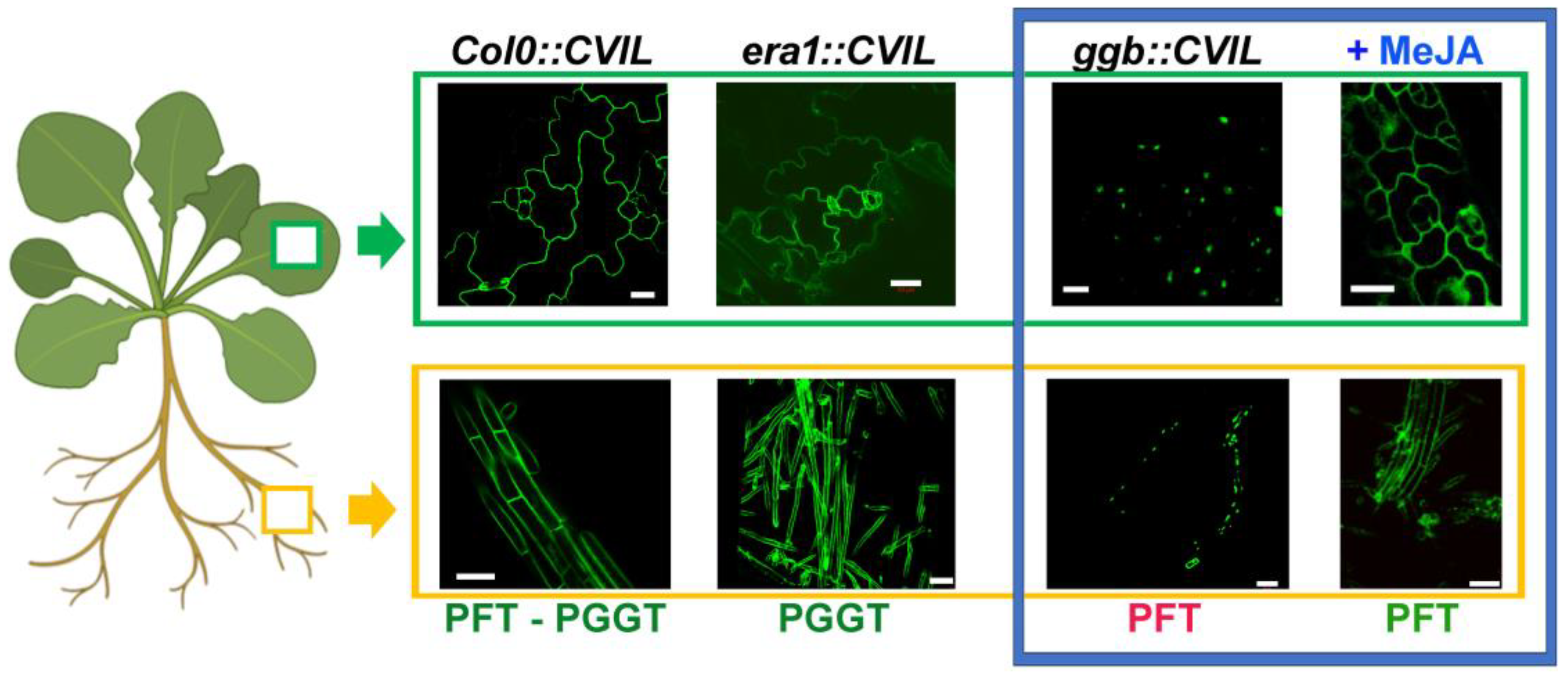
Figure 3.
Treatment with Methyl-jasmonate increases the biosynthetic capacity of protein prenylation in Nicotiana tabacum leaf-disks. (a) and (d) control; (b) and (e) fosmidomycin (100 µM) and mevinolin (5 µM) treatments. (c) represents (b) conditions in the presence of 0.5% cellulase and (f) (e) condition in the presence of 30 µM Me-JA. Leaf-disks were pretreated for 6h with chemicals as indicated below before GFP-CaM-CVIL expressed was induced with 30 µM dexamethasone. During the whole experiment, plant materials were incubated in the dark at 22°C. The white bar represents 20 µm.
Figure 3.
Treatment with Methyl-jasmonate increases the biosynthetic capacity of protein prenylation in Nicotiana tabacum leaf-disks. (a) and (d) control; (b) and (e) fosmidomycin (100 µM) and mevinolin (5 µM) treatments. (c) represents (b) conditions in the presence of 0.5% cellulase and (f) (e) condition in the presence of 30 µM Me-JA. Leaf-disks were pretreated for 6h with chemicals as indicated below before GFP-CaM-CVIL expressed was induced with 30 µM dexamethasone. During the whole experiment, plant materials were incubated in the dark at 22°C. The white bar represents 20 µm.
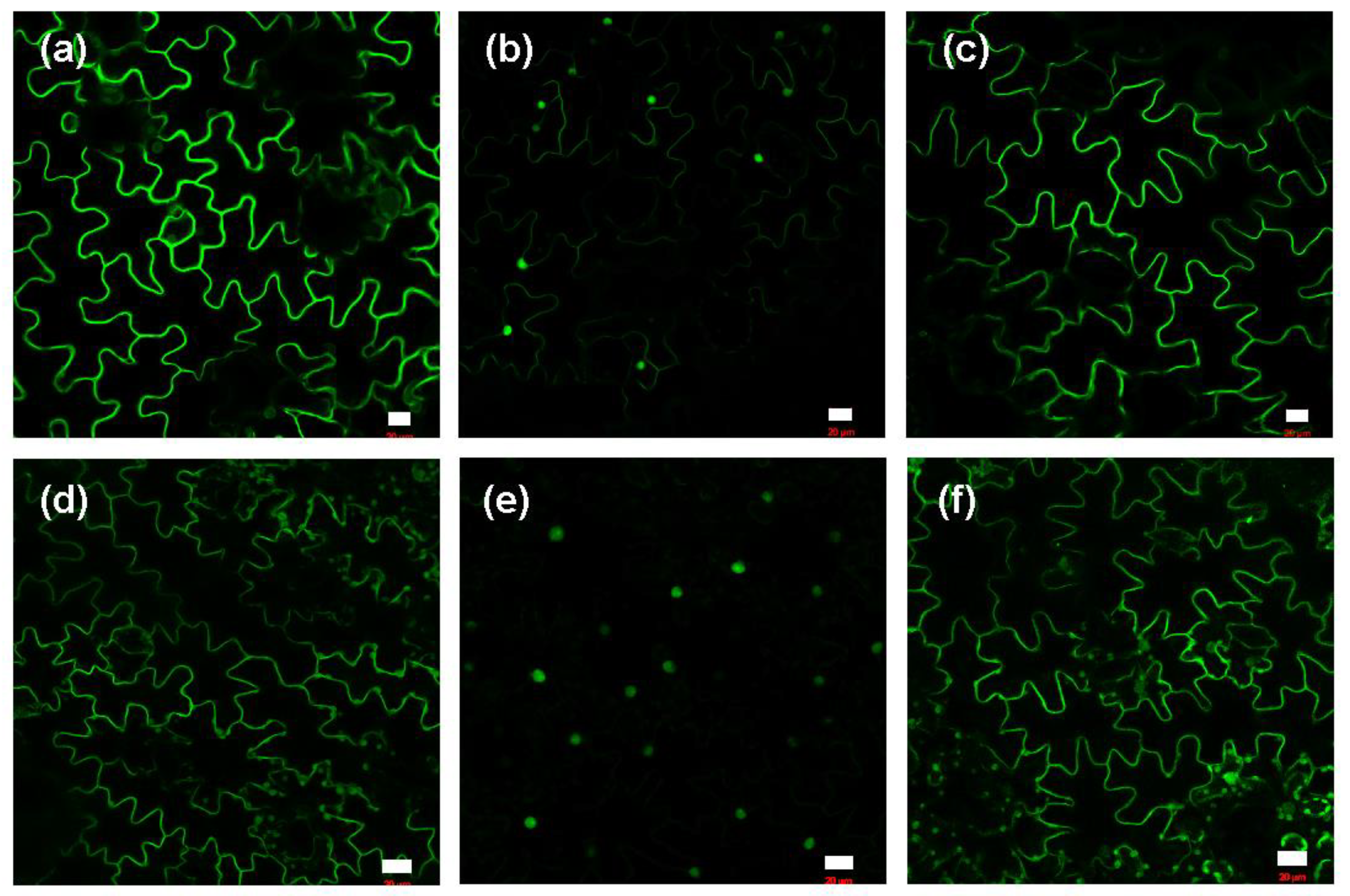
Figure 4.
(a) Fol-induced protein prenylation in leaves of Arabidopsis ggb::CVIL mutant plants transformed with pTA-GFP-CaM-CVIL. Subcellular localization of GFP-CaM-CVIL was evaluated by confocal microscopy. The white bar corresponds to 20 µm. (b) Protein prenyltransferase enzyme assays using radiolabeled prenyl diphosphates (0.25 mCi/assay), purified GFP-CaaX protein substrates (35 µM), and 15 ug tobacco BY-2 cell-free protein extracts to evaluate prenylation capacity by FPP and GGPP. Tobacco By-2 cells were pretreated with MVA (2 mM), Fol (20 µM), GGol (20 µM) or DX (2 mM) for 24h as compared to untreated cells (NT), before protein extracts were isolated. Radiolabeled FPP or GGPP have been used as prenyl diphosphate donner and recombinant GFP-CaM-CVIL or the non-prenylable GFP-CaM-SVIL as protein substrate. The absence of a radiolabeled protein band for GFP-CaM-SVIL indicates that the signal is specific to prenylation of proteins.
Figure 4.
(a) Fol-induced protein prenylation in leaves of Arabidopsis ggb::CVIL mutant plants transformed with pTA-GFP-CaM-CVIL. Subcellular localization of GFP-CaM-CVIL was evaluated by confocal microscopy. The white bar corresponds to 20 µm. (b) Protein prenyltransferase enzyme assays using radiolabeled prenyl diphosphates (0.25 mCi/assay), purified GFP-CaaX protein substrates (35 µM), and 15 ug tobacco BY-2 cell-free protein extracts to evaluate prenylation capacity by FPP and GGPP. Tobacco By-2 cells were pretreated with MVA (2 mM), Fol (20 µM), GGol (20 µM) or DX (2 mM) for 24h as compared to untreated cells (NT), before protein extracts were isolated. Radiolabeled FPP or GGPP have been used as prenyl diphosphate donner and recombinant GFP-CaM-CVIL or the non-prenylable GFP-CaM-SVIL as protein substrate. The absence of a radiolabeled protein band for GFP-CaM-SVIL indicates that the signal is specific to prenylation of proteins.
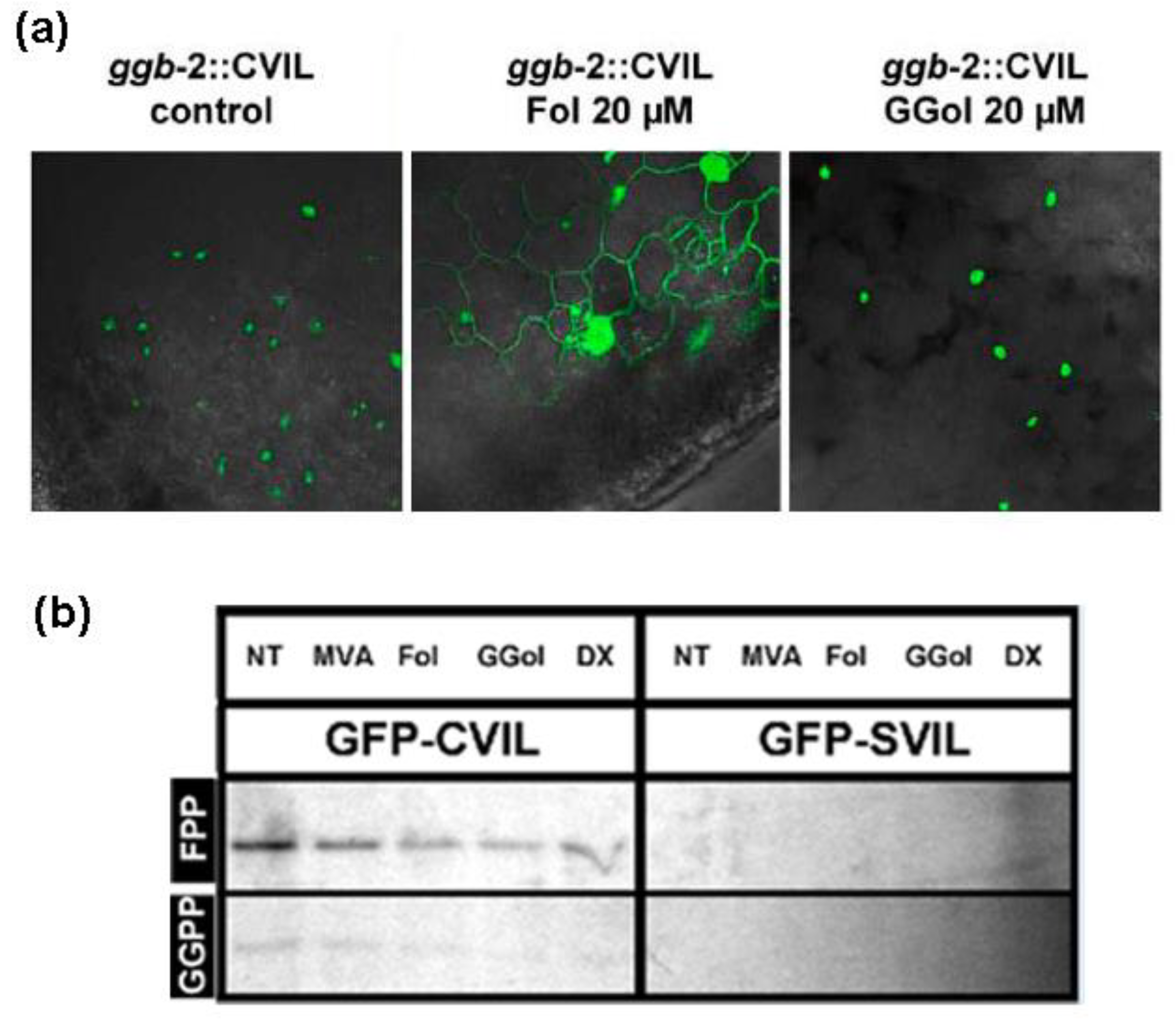
Figure 5.
Stimulation of HMGR and MVA production by Methyl which modifies PFT protein substrate specificity. (a) Quantification of total phytosterol isolated from untreated cells (C), cells treated with Me-JA (30 µM), with fosmidomycin (FOS 100 µM) and MeJA/FOS have been deposited in each well. Groups have no significantly different means (p ≤ 0.005) as calculated by a Bon-ferroni-Holm test following a one-way ANOVA. (b) Quantification of HMGR protein by western-blotting using @HMGR2cat. 40 µg of a microsomal protein fraction from tobacco BY-2 cells treated as described in (a) have been charged in each well. (c) Quantification of HMGR in tobacco BY-2 cells during growth curve. Cells were diluted 5-fold in MS medium and grown for 24-hours, 48 hours, 72 hours, 96 hours and 120 hours before collected to make microsomal fractions. (d) [14C]MVA (1 µCi) incorporation into prenylated proteins of tobacco BY-2 cells treated (+) or not (-) with 20 µM MeJA during 24 hours. The picture represents a fluorogram of SDS-PAGE-separated total protein extracts. (e) Subcellular localization of GFP-CaM-CVIL protein expressed in Arabidopsis ggb lines grown for 48 h in the presence of MVA (2 mM), or DX (2 mM), before protein expression is induced with dexamethasone (30 µM).
Figure 5.
Stimulation of HMGR and MVA production by Methyl which modifies PFT protein substrate specificity. (a) Quantification of total phytosterol isolated from untreated cells (C), cells treated with Me-JA (30 µM), with fosmidomycin (FOS 100 µM) and MeJA/FOS have been deposited in each well. Groups have no significantly different means (p ≤ 0.005) as calculated by a Bon-ferroni-Holm test following a one-way ANOVA. (b) Quantification of HMGR protein by western-blotting using @HMGR2cat. 40 µg of a microsomal protein fraction from tobacco BY-2 cells treated as described in (a) have been charged in each well. (c) Quantification of HMGR in tobacco BY-2 cells during growth curve. Cells were diluted 5-fold in MS medium and grown for 24-hours, 48 hours, 72 hours, 96 hours and 120 hours before collected to make microsomal fractions. (d) [14C]MVA (1 µCi) incorporation into prenylated proteins of tobacco BY-2 cells treated (+) or not (-) with 20 µM MeJA during 24 hours. The picture represents a fluorogram of SDS-PAGE-separated total protein extracts. (e) Subcellular localization of GFP-CaM-CVIL protein expressed in Arabidopsis ggb lines grown for 48 h in the presence of MVA (2 mM), or DX (2 mM), before protein expression is induced with dexamethasone (30 µM).
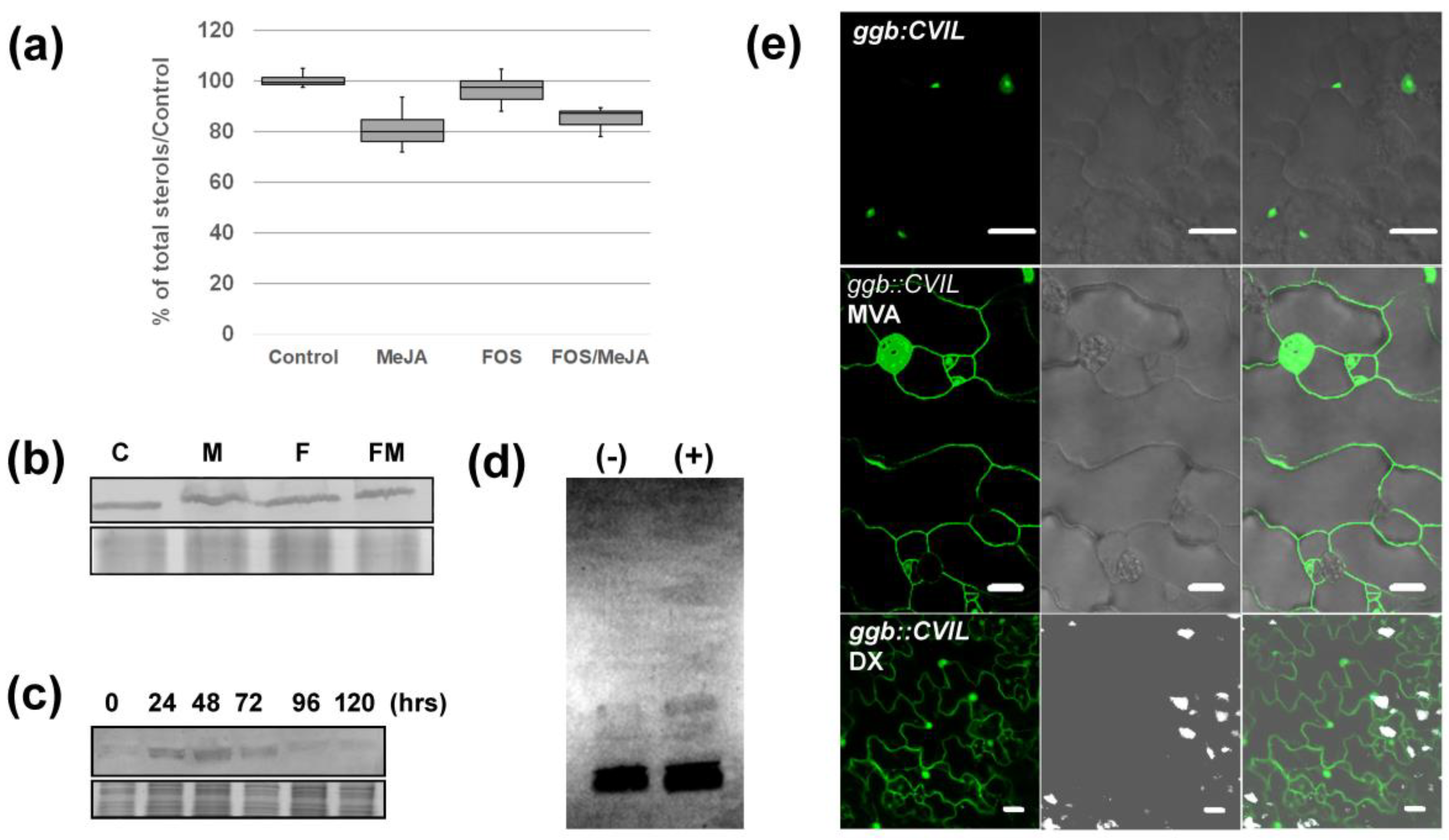
Figure 6.
MVA as a candidate for providing prenyl groups in Me-JA/FOS- treated tobacco BY-2 cells. In each experiment, cells were pretreated for 3 hours with chemicals as indicated, before protein expression was induced with dexamethasone (15 µM). (a) Mevinolin (MV 10 µM) inhibits Me-JA (20 µM) -mediated reversion of FOS (100 µM). Treated with FOS + MeJA + MV, the nuclear localization of GFP-CaM-CVIL expressed in tobacco BY-2 cells is revived. MVA (2 mM) reverses, MV-induced inhibition of protein prenylation. Distribution of GFP fluorescence in membrane (M, black) membrane/nucleus (M+N, white) and nucleus (N gray) are indicated in percentage and chi-squared tests against the null hypothesis being true was used to assign for significant differences to p-values <0.01, which are indicated by different letters. (b) Synergetic effect of increasing concentrations of MVA with (+) or without (-) Me-JA (20 µM) in FOS/MV (100 µM / 5 µM)-treated tobacco BY-2 cells. Color scale from red (0%) to green (100 %) indicates the percentage of cells with membrane localization.
Figure 6.
MVA as a candidate for providing prenyl groups in Me-JA/FOS- treated tobacco BY-2 cells. In each experiment, cells were pretreated for 3 hours with chemicals as indicated, before protein expression was induced with dexamethasone (15 µM). (a) Mevinolin (MV 10 µM) inhibits Me-JA (20 µM) -mediated reversion of FOS (100 µM). Treated with FOS + MeJA + MV, the nuclear localization of GFP-CaM-CVIL expressed in tobacco BY-2 cells is revived. MVA (2 mM) reverses, MV-induced inhibition of protein prenylation. Distribution of GFP fluorescence in membrane (M, black) membrane/nucleus (M+N, white) and nucleus (N gray) are indicated in percentage and chi-squared tests against the null hypothesis being true was used to assign for significant differences to p-values <0.01, which are indicated by different letters. (b) Synergetic effect of increasing concentrations of MVA with (+) or without (-) Me-JA (20 µM) in FOS/MV (100 µM / 5 µM)-treated tobacco BY-2 cells. Color scale from red (0%) to green (100 %) indicates the percentage of cells with membrane localization.
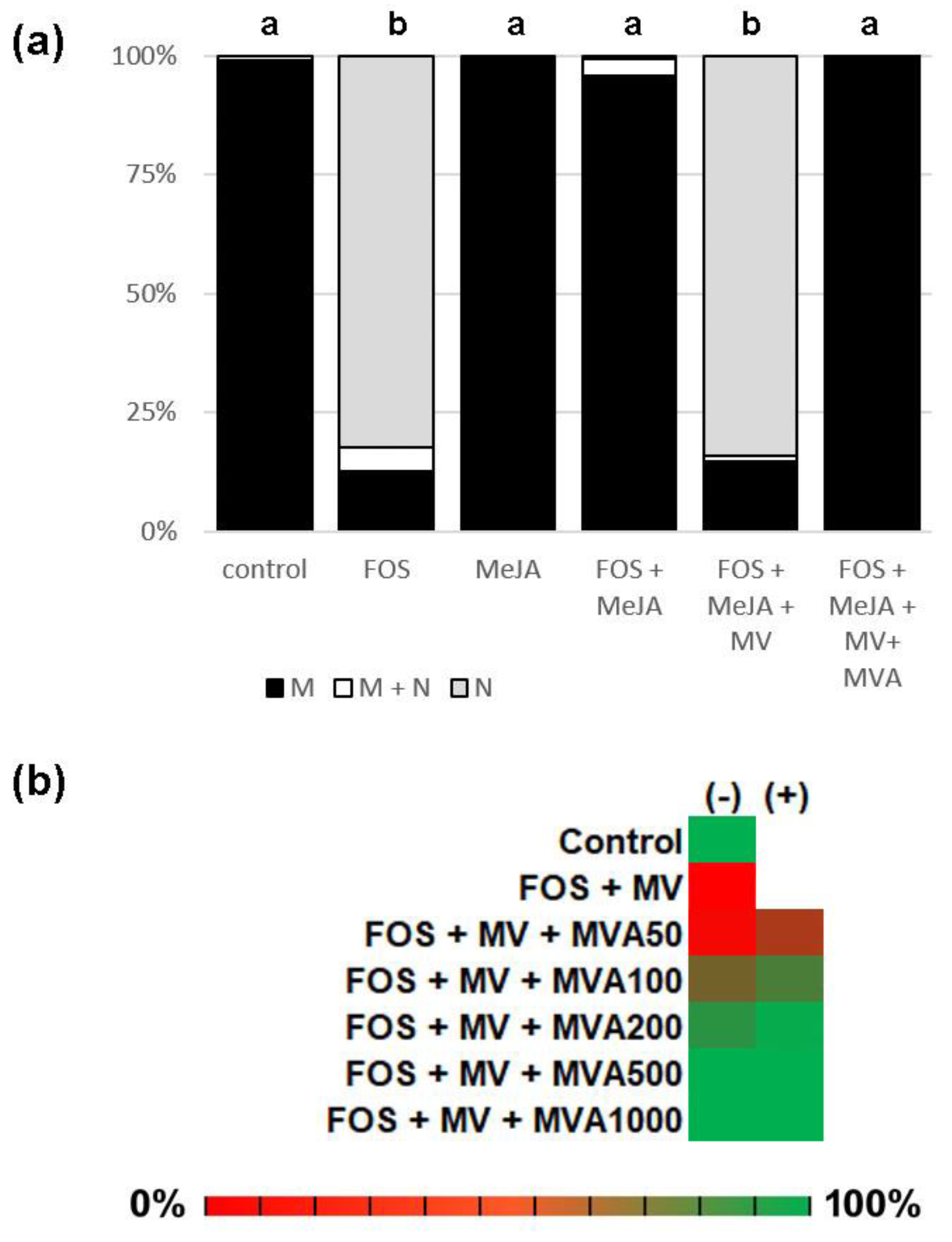
Figure 7.
A working model of the role of jasmonic acid methyl esther (Me-JA) in the activation of protein prenylation through isoprenoid metabolic cross-talk. Under standard growth conditions, the protein is modified with a plastidial methylerythritol phosphate (1) pathway-derived geranylgeranyl group and is labeling the plasma membrane [28]. Unprenylated, the protein localizes in the cell nucleus. The absence of prenylation is attributed to the inhibition of the MEP pathway by fosmidomycin (3), or the absence of PGGT in the Arabidopsis ggb KO mutant. Treatment with Me-JA (4) leads to the stimulation of the mevalonic acid (MVA) (2) pathway through activation of HMGR, to improve the capacity for protein prenylation in plants under stress conditions. This somehow modifies substrate specificity of PFT, which recognizes GFP-CaM-CVIL under those conditions. The increase of prenylation capacity allows a reversion of fosmidomycin inhibition, and a capacity of PFT to recognize a PGGT protein substrate.
Figure 7.
A working model of the role of jasmonic acid methyl esther (Me-JA) in the activation of protein prenylation through isoprenoid metabolic cross-talk. Under standard growth conditions, the protein is modified with a plastidial methylerythritol phosphate (1) pathway-derived geranylgeranyl group and is labeling the plasma membrane [28]. Unprenylated, the protein localizes in the cell nucleus. The absence of prenylation is attributed to the inhibition of the MEP pathway by fosmidomycin (3), or the absence of PGGT in the Arabidopsis ggb KO mutant. Treatment with Me-JA (4) leads to the stimulation of the mevalonic acid (MVA) (2) pathway through activation of HMGR, to improve the capacity for protein prenylation in plants under stress conditions. This somehow modifies substrate specificity of PFT, which recognizes GFP-CaM-CVIL under those conditions. The increase of prenylation capacity allows a reversion of fosmidomycin inhibition, and a capacity of PFT to recognize a PGGT protein substrate.
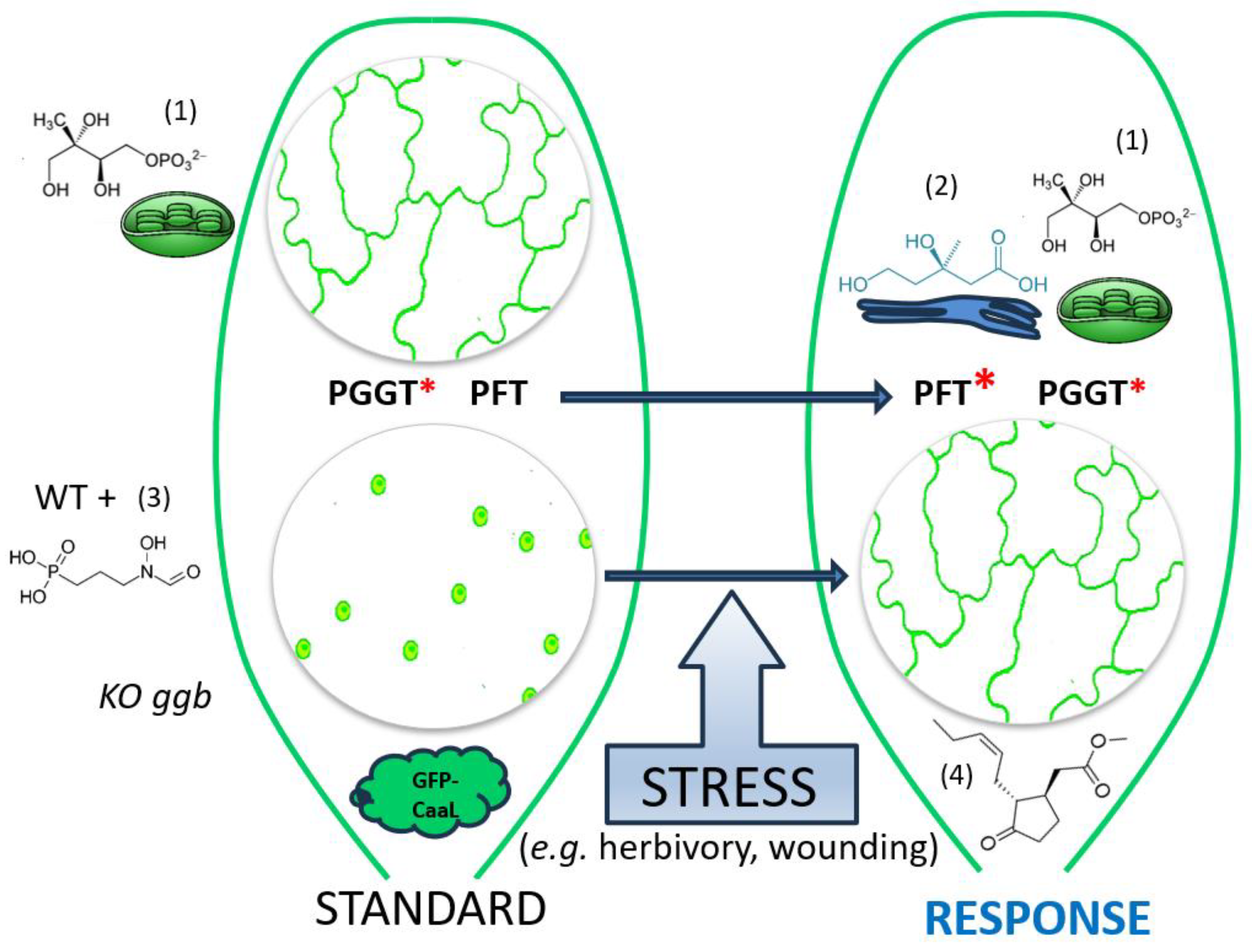
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



