
Submitted:
08 December 2023
Posted:
11 December 2023
You are already at the latest version
Abstract
The successful advancement of xenotransplantation has led to the development of highly sensitive detection systems for the screening for potentially zoonotic viruses in donor pigs and preventing their transmission to the recipient. To validate these methods, genetically modified pigs generated for xenotransplantation, numerous minipig and other pig breeds have been tested, thereby increasing our knowledge concerning the pig virome and the distribution of pig viruses. Of particular importance are the porcine cytomegalovirus, a porcine roseolovirus (PCMV/PRV) and the hepatitis E virus genotype 3 (HEV3). PCMV/PRV has been shown to reduce the survival time of pig transplants in non-human primates and was also transmitted in the first pig heart transplantation to a human patient. Here, we determined the sensitivities of our methods to detect PCMV/PRV, HEV3, porcine lymphotropic herpesvirus - 1 (PLHV-1), PLHV-2, PLHV-3, porcine circovirus 2 (PCV2), PCV3, PCV4 and porcine parvovirus 1 (PPV1) efficiently and used the methods to screen indigenous Greek black pigs. Since porcine endogenous retroviruses (PERVs) of type A and B are integrated in the genome of all pigs, but PERV-C is not, the animals were screened for PERV-C and PERV-A/C. Our detection methods were sensitive and detected PCMV/PRV, PLHV-1, PLHV-1, PLHV-3, PVC3 and PERV-C in most animals. PPV1, HEV3, PCV4 and PERV-A/C were not detected. These data are of great interest since the animals are healthy and very resistant to diseases.
Keywords:
xenotransplantation
; virus safety
; porcine cytomegalovirus/porcine roseolovirus (PCMV/PRV)
; hepatitis E virus
; porcine endogenous retroviruses (PERVs)
1. Introduction
The virome of pigs is not well studied [1]. The virome is the total amount of viruses in and on the pig body and also includes the endogenous retroviruses as well as the bacteriophages infecting bacteria present in the pig organisms. Most common in healthy pigs are picornaviruses followed by circoviruses, adenoviruses, and parvoviruses [2]. In the case of diarrhea the percentage of adenoviruses and circoviruses decreased and the percentage of anelloviruses and reoviruses increased [2]. In diarrhoeic faeces samples from 27 Chinese pigs porcine bocavirus-2, a parvovirus, was found in 59 % of the animals, porcine bocavirus-4 (also a parvovirus) in 18%, Torque teno sus virus-2 (TTSuV-2, an anellovirus) in 7%, porcine epidemic diarrhea virus (PEDV, a coronavirus) in 70%, porcine stool associated circular virus (PoSCV, circovirus-like) in 7%, sapovirus (a calicivirus) in 33%, sapelovirus (a picornavirus) in 48%, torovirus (a coronavirus) in 33%, posavirus-1 (a picornavirus) in 40%, porcine astrovirus in 74%, coronavirus in 7%, porcine enterovirus-9 (a picornavirus) in 85%, picobirnavirus (PBV) in 15%, kobuvirus (a picornavirus) in 44% of the 27 animals [3].
In a recent study in the United States, serum samples from healthy show pigs from the years 2018 or 2019 were analyzed by high-throughput sequencing to estimate the virome. Results demonstrated the presence of DNA viral families (Parvoviridae, Circoviridae, and Herpesviridae) and RNA families (Arteriviridae, Flaviviridae, and Retroviridae). Twenty-three viral species were identified, among them were important swine pathogens including porcine reproductive and respiratory syndrome virus (PRRSV), atypical porcine pestivirus, and porcine circovirus (PCV) [4]. The herpesvirus detected was PCMV/PRV, but only one contig. This underlines that next-generation sequencing (NGS) can detect known viruses but has an extremely limited sensitivity. When 36 pooled porcine nasal swabs and blood serum samples collected from both sides of the Dutch-German border region were evaluated, 46 different viral species were detected using viral targeted sequence capture (TSC), compared to 40 viral species with a shotgun metagenomics approach [5]. In contrast, more sensitive methods such as PCR and real-time PCR can detect viruses even with a very low virus load [1].
Studies on the prevalence of porcine viruses were stimulated by the rapid development of xenotransplantation using pig cells, tissues, and organs. Xenotransplantation is under development to alleviate the shortage of human donor organs for the treatment of organ failure. In the last years, remarkable survival times of pig xenotransplants in non-human primates were achieved. In 2022 and 2023 the first two pig hearts were transplanted into patients in Baltimore [6,7]. However, xenotransplantation may be associated with the transmission of porcine viruses which may be zoonotic or xenozoonotic. Viruses are zoonotic when they can cause a disease in healthy humans such as the hepatitis E virus (HEV). Viruses are xenozoonotic when they do not induce a disease in healthy humans but affect the recipient when transmitted with a xenotransplant such as PCMV/PRV [8]. It was shown that the transmission of PCMV/PRV drastically reduced the survival time of pig xenotransplants in non-human primates [9,10]. PCMV/PRV was also transmitted to the first patient in Baltimore and probably contributed to his death [6,11]. To detect potentially zoonotic or xenozoonotic pig viruses, in many laboratories sensitive and specific detection methods were developed (for review see [12,13]). It became clear that for a successful detection of pig viruses not only sensitive and specific detection methods, either PCR-based or cell-based methods or immunological methods, are required. An entire detection system including sample generation, sample preparation, sample origin, time of sampling as well as negative and positive controls is important [12,13]. Using these detection systems, different minipigs such as the Auckland Island pigs [14,15], the Göttingen minipigs [16,17,18,19,20,21,22], Göttingen minipigs with dippity pig syndrome (DPS) [23], the Aachen minipigs [24] and the Mini LEWE pigs [25] as well as genetically modified pigs generated for xenotransplantation [10,26,27,28,29,30,31], Greek pigs with erythema multiforme [32] and wild boars [33,34] were analyzed.
Here, we analyzed the indigenous Greek black pigs (Figure 1) using these methods. This breed is the only traditional indigenous pig breed reared in Greece. Most interestingly, it has its roots in ancient Greece. It is thought that these are the pigs from the Odyssey in the farm of Odysseus with his swineherd Eumaios [35].
Most Greek organic pig farms are located in mountainous or semi-mountainous areas, which is why they don’t have a thick layer of fat like other types of pigs. They are resistant to weather conditions and diseases. Conventional pigs give birth to 12-14 piglets, whereas indigenous Greek black pigs give birth only up to seven. A conventional pig is utilized at the age of 5 months and weight of 110 kg, in contrast, an indigenous Greek black pig is slaughtered at 7–10 months of age, reaching a carcass weight of about 60 kg [36,37]. The animals give delicious pork meat and in some farms they are fed with olives. When the genetic diversity, based on microsatellite analysis, of the Greek black pig was investigated, its genetic uniqueness was demonstrated. Despite their low population size, they have a high degree of genetic variability, which will be useful for breeding programs aimed at maintaining the long-term survival of this ancient breed [36,37].
Twenty-one animals from 4 farms in Greece (Figure 2) were analyzed using real-time PCR for PCMV/PRV, PCV2, PCV3, PCV4, PLHV-1, PLHV-2 and PLHV-3, as well as real-time RT-PCR for HEV. For the detection of PERV-C and PERV-A/C conventional PCRs were used. In addition, 11 animals from two farms were screened for antibodies against PCMV/PRV using a Western blot assay.
2. Materials and Methods
2.1. Animals and tissues
Liver and some spleen tissues were obtained from animals in four different farms in Greece (Figure 2). Farm 1 located near Drama, North Greece (4 samples), farm 2 (3 samples), farm 3 (6 samples) and farm 4 (8 samples), all located near Trikala, Thessaly, Central Greece. In addition, sera were obtained from 11 animals from farms 1 and 4.
2.2. DNA and RNA isolation
DNA and RNA were isolated from the tissues according to the manufacturer´s instructions using the DNeasy Blood & Tissue kit and RNeasy kit (Qiagen, Hilden, Ger-many), respectively. DNA and RNA concentrations were determined using NanoDrop ND-1000 (Thermo Fisher Scientific Inc., Worcester, MA, USA).
2.3. Real-time PCR for the detection of DNA viruses
Real-time PCRs were performed to detect PCMV/PRV, PLHV-1, PLHV-2, PLHV-3, PCV2, PCV3, PCV4 and PPV1 as described previously using specific primers and probes (Table 1) [23,38,39,40,41,42,43]. All protocols were performed using the SensiFAST Probe No-ROX kit (Meridian Bioscience, Cincinnati, OH, USA) in a reaction volume of 16 µl plus 4 µl (100 ng) of DNA template. All real-time PCRs were carried out as duplex PCRs that simultaneously indicate the gene of interest and porcine glyceraldehyde-3-phosphate-dehydrogenase (pGAPDH) as internal control for each sample. Real-time PCR reactions were carried out with a qTOWER3 G qPCR cycler (Analytik Jena, Jena, Germany) and the real-time PCR conditions as previously described [23].
2.4. Real-time reverse transcriptase PCR for the detection of HEV
Real-time reverse transcriptase-PCR (real-time RT-PCR) as described by Jothikumar et al. [44] was carried out to detect hepatitis E virus (HEV). All real-time RT-PCR reactions were performed in a reaction volume of 16 µl using SensiFAST Probe No-ROX One-Step Kit (Meridian Bioscience, Cincinnati, OH, USA) plus 4 µl (100 ng) template RNA. The reaction was performed at the qTOWER3 G qPCR cycler (Analytik Jena, Jena, Germany). The temperature-time profile applied consists of a reverse transcriptase step of 30 minutes at 50°C, followed by an activation step of 15 minutes at 95°C and 45 cycles comprising a step of 10 seconds at 95°C, followed by a step of 20 seconds at 55°C and 15 seconds at 72°C [25].
2.5. Conventional PCR for the detection of PERVs
A conventional PCR was performed to determine the presence of PERV-C. PERV-C was detected using a set of primers with an amplicon length of 288 bp (described as PCR4 in [45]). It was carried out with AmpliTaq DNA Polymerase (Applied Biosystems, Waltham, MA, USA) and was set up with a Biometra TRIO cycler (Analytik Jena, Jena, Germany). The following temperature-time profile was used: 95°C for 10 minutes (activation step), followed by 45 cycles at 95°C for 15 seconds (denaturation), 55°C for 30 seconds (annealing) and 72°C for 30 seconds (extension) and a final extension at 72°C for 5 minutes.
A conventional PCR to determine the presence of human-tropic PERV-A/C, was set up using specific primer pairs (Table 1) [46]. The PERV-A/C long primer mix detects an amplicon of 1266 bp length. It was carried out with AmpliTaq DNA Polymerase (Applied Biosystems, Waltham, MA, USA) and was set up with a Biometra TRIO cycler (Analytik Jena, Jena, Germany). The following temperature-time profile was used: 95°C for 10 minutes (activation step), followed by 45 cycles at 95°C for 15 seconds (denaturation), 55°C for 30 seconds (annealing) and 72°C for 90 seconds (extension) and a final single cycle at 72°C for 5 minutes.
2.6. Determination of the sensitivity
The sensitivity of real-time PCRs for detection of various viruses tested in this study was determined using gene blocks. Briefly, different copy numbers (101 to 109) of gene

Table 1.
Oligonucleotides for the primers and probes used in this study.
| Virus | Primer/Probe | Sequence 5′-3′ | Reference | |
|---|---|---|---|---|
| HEV | JVHEV-Fwd | GGT GGT TTC TGG GGT GAC | Jothikumar et al. 2006 [45] | |
| JVHEV-Rev | AGG GGT TGG TTG GAT GAA | |||
| JVHEV-Probe | 6FAM-TGA TTC TCA GCC CTT CGC-BHQ | |||
| PCMV/PRV | PCMV-Fwd | ACT TCG TCG CAG CTC ATC TGA | Mueller et al. 2002, [38] | |
| PCMV-Rev | GTT CTG GGA TTC CGA GGT TG | |||
| PCMV-Probe | 6FAM-CAG GGC GGC GGT CGA GCT C-BHQ | |||
| PLHV-1 | PLHV-1 (1125)-Fwd | CTC ACC TCC AAA TAC AGC GA | Chmielewicz et al. 2003 [39] | |
| PLHV-1 (1125)-Rev | GCT TGA ATC GTG TGT TCC ATA G | |||
| PLHV-1 (1125)-Probe | 6FAM-CTG GTC TAC TGA ATC GCC GCT AAC AG-TAMR | |||
| PLHV-2 | PLHV-2 (1155)-Fwd | GTC ACC TGC AAA TAC ACA GG | Chmielewicz et al. 2003 [39] | |
| PLHV-2 (1155)-Rev | GGC TTG AAT CGT ATG TTC CAT AT | |||
| PLHV-2 (1155)-Probe | 6FAM-CTG GTC TAC TGA AGC GCT GCC AAT AG-TAMRA | |||
| PLVH-3 | PLHV-3 (210s)-Fwd | AAC AGC GCC AGA AAA AAA GG | McMahon et al. 2006 [40] | |
| PLHV-3 (210as)-Rev | GGA AAG GTA GAA GGT GAA CCA TAA AA | |||
| PLHV-3 (210)-Probe | 6-FAM CCA AAG AGG AAA ATC-MGB | |||
| PCV2 | PCV2 (F2020)-Fwd | CTG AGT CTT TTT TAT CAC TTC GTA ATG GT | Chen et al. 2021, [41] | |
| PCV2 (F2020)-Rev | ACT GCG TTC GAA AAC AGT ATA TAC GA | |||
| PCV2 (F2020)-Probe | 6FAM-TTA AGT GGG GGG TCT TTA AGA TTA AAT TCT CTG AAT TGT-BHQ2 | |||
| PCV3 | PCV3-Fwd | AGT GCT CCC CAT TGA ACG | Palinski et al. 2017 [42] | |
| PCV3-Rev | ACA CAG CCG TTA CTT CAC | |||
| PCV3-Probe | 6FAM-ACC CCA TGG CTC AAC ACA TAT GAC C-BHQ1 | |||
| PCV4 | PCV4 (F2020)-Fwd | ATT ATT AAA CAG ACT TTA TTT GTG TCA TCA CTT | Chen et al. 2021 [41] | |
| PCV4 (F2020)-Rev | ACA GGG ATA ATG CGT AGT GAT CAC T | |||
| PCV4 (F2020)-Probe | 6FAM-ATA CTA CAC TTG ATC TTA GCC AAA AGG CTC GTT GA-BHQ1 | |||
| PPV1 | PPV1-Fwd | CAG AAT CAG CAA CCT CAC CA | Opriessnig et al. 2011 [43] | |
| PPV1-Rev | GCT GCT GGT GTG TAT GGA AG | |||
| PPV1-Probe | 6FAM-TGC AAG CTT/ZEN/AAT GGT CGC ACT AGA CA-BHQ1 | |||
| pGAPDH | pGAPDH-Fwd | ACA TGG CCT CCA AGG AGT AAG A | Duvigneau et al. 2005 [47] | |
| pGAPDH-Rev | GAT CGA GTT GGG GCT GTG ACT | |||
| pGAPDH-Probe | HEX-CCA CCA ACC CCA GCA AGA G-BHQ1 | |||
| PERV-C | PERV-envC-Fwd | GAT TAG AAC TGG AAG CCC CAA GTG CTC T | Kaulitz et al., 2013 [45] | |
| PERV-envC-Rev | TCT GAT CCA GAA GTT ATG TTA GAG GAT GGT | |||
| PERV-A/C | PERV-A env VRBF-Fwd | CCT ACC AGT TAT AAT CAA TTT AAT TAT GGC | Wood et al. 2004 [46] | |
| PERV-C env TMR-Rev | CTC AAA CCA CCC TTG AGT AGT TTC C | |||
Fwd = forward primer, Rev = reverse primer.
2.7. Western blot to detect antibodies against PCMV/PRV
3. Results
3.1. Sensitivity of the assays
To determine the sensitivity of the real-time PCRs, gBlock gene fragments (Integrated DNA Technologies, IDT, Coralville, IA, USA) were used as described [25]. The gBlocks comprised the virus-specific oligosequences corresponding to the primer and probe, which are separated by spacers. The spacers are non-functional oligosequences of eight to fifteen base pairs that are used as placeholders sequences. In additon to the viral gBlocks we used also a gBlock containing the primer and probe sequences of the porcine GAPDH. Full sequences and characterization of the gBlocks are given in [25]. Using these gBlocks the sensitivity of the real-time PCR was determined (Supplementary Figure 1) and compared with the sensitivity of previously published real-time PCRs (Table 2). The sensitivity of the real-time PCRs ranged between 1 and 10 copies (Table 2).
3.2. Screening for herpesviruses: PCMV/PRV, PLHV-1, PLHV-2, PLHV-3
When the animals were tested for these herpesviruses, 16 of 21 animals were positive for PCMV/PRV in liver tissues, 12 animals were positive for PLHV-1, 15 animals were positive for PLHV-2, and all were positive for PLHV-3 (Table 3). When spleen tissues were tested similar results were obtained (Table 4).
3.3. Screening for circoviruses and PPV-1
3.4. Screening for RNA viruses. HEV3
All animals were free from HEV3 (Table 3).
3.4. Screening for PERVs
Since all pigs harbor PERV-A and PERV-B in their genome, we tested only for PERV-C using a primer pair described previously (PCR4 in [45]). 11 of 21 animals were positive for PERV-C using this PCR (Table 3, Figure 3). When the PERV-C positive animals were tested for PERV-A/C, this recombinant virus was not found (Table 3).
Table 2.
Sensitivity of different PCR-based methods detecting pig viruses.
| Virus | Method | Sensitivity (copy number per 100 ng DNA) |
Sensitivity R2 | Reference |
|---|---|---|---|---|
| PCMV/PRV | conventional PCR | 15 copies | Morozov et al., 2016 [51] | |
| nested PCR | 5 copies | |||
| real-time PCR | 2 copies | |||
| real-time PCR | 20 copies | Mueller et al., 2002 [38] | ||
| real-time PCR | 10 copy | 0.9964 | this manuscript | |
| HEV | real-time RT-PCR | 10 | Jothikumar et al., 2006 [44] | |
| real-time RT-PCR | 150-200 | Morozov et al., 2015 [17] | ||
| real-time RT-PCR | 10 copy | 0.9962 | this manuscript | |
| PCV2 | multiplex | 101/µl | Zhou et al., 2022 [52] | |
| real-time PCR | 1 copy | 0.9935 | this manuscript | |
| PCV3 | real-time PCR | 10 copies | 0.9906 | this manuscript |
| PCV4 | real-time PCR | 100 copies | 0.9906 | this manuscript |
| PLHV-1 | real-time PCR | 20 copies | Chmielewicz et al. 2003 [39] | |
| real-time PCR | 1 copy | 0.9964 | this manuscript | |
| PLHV2 | real-time PCR | 20 copies | Chmielewicz et al. 2003 [39] | |
| real-time PCR | 1 copy | 0.9953 | this manuscript | |
| PLHV3 | real-time PCR | 1 copies | 0.9983 | this manuscript |
| PPV1 | real-time PCR | 10 copy | 0.9961 | this manuscript |
3.5. Western blot assay to detect antibodies against PCMV/PRV
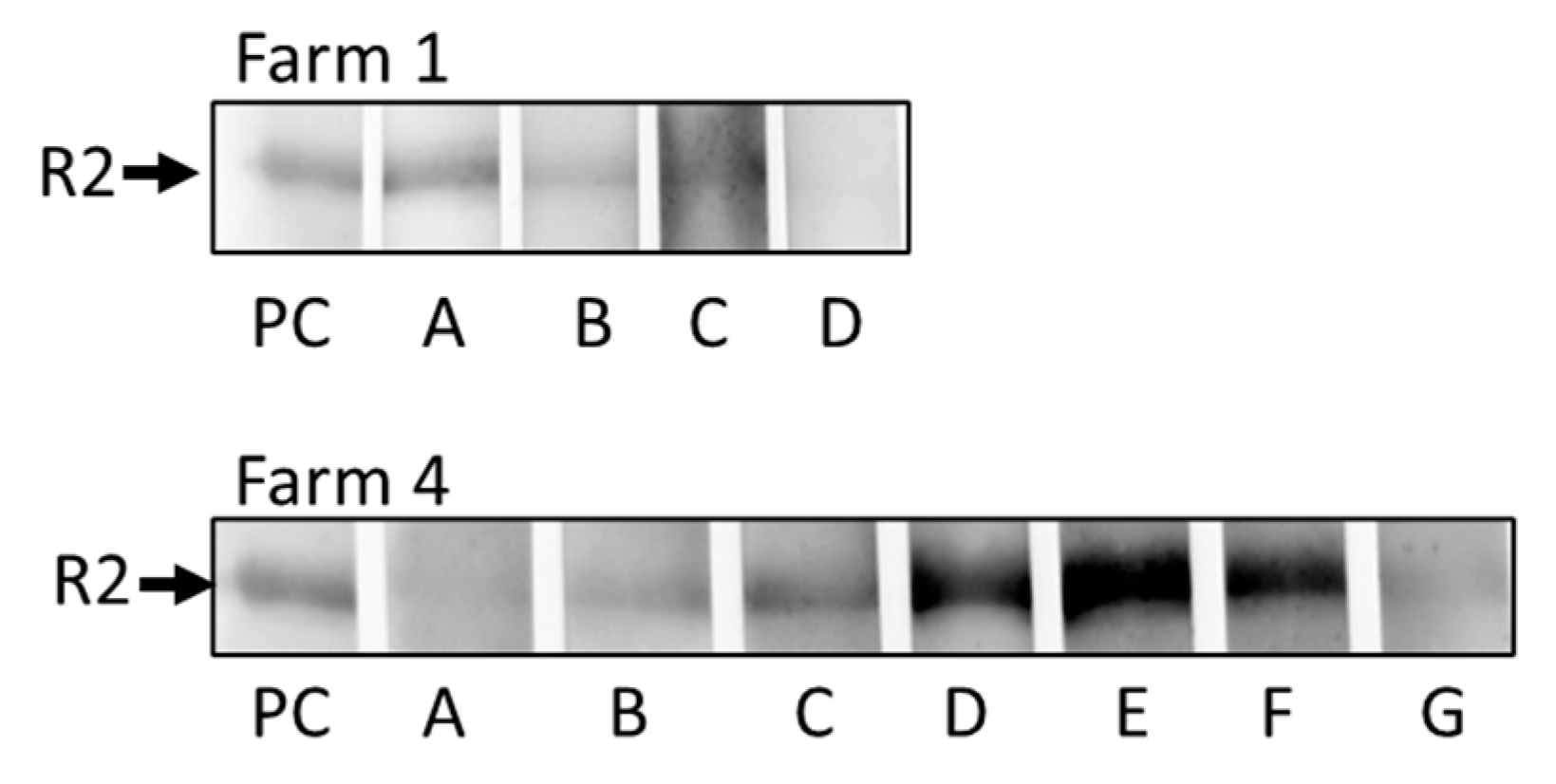
A Western blot analysis of sera from animals from Farms 1 and 4 was performed using the recombinant C-terminal fragment of gB of PCMV/PRV [48]. The tested animals were not the same as the animals tested using the PCR, but in the same age (4 animals from farm 1, 8-9 months, 7 animals from farm 4, 10-11 months). All sera reacted positive (Figure 4), however, with strong differences in the extent of the band. The serum from animal D from farm 1 seems to be negative in Figure 4 but using a longer exposition time (2 seconds instead of 9 milliseconds) these animals were found positive.
4. Discussion
In order to evaluate the potency of our improved detection methods developed for the effective screening of viruses potentially posing a risk for xenotransplantation, indigenous Greek black pigs were thoroughly screened. First of all, they were tested for PCMV/PRV, which had been shown to reduce the survival time of pig transplants in non-human primates significantly [9,10]. PCMV/PRV was also transmitted in the first transplantation of a pig heart into a patient in Baltimore [6,11]. Since the symptoms in the baboons with PCMV/PRV-positive transplants are like the symptoms in the Baltimore patient, the virus obviously contributed to the death of the patient. The real-time PCR developed by Mueller at al. [38] was modified and performed as a duplex real-time PCR detecting simultaneously porcine GAPDH [25]. Furthermore, gBlocks comprising the virus-specific oligosequences corresponding to the primers and probes were used as positive control and for the standard curves (Supplementary Figures 1). Using this real-time PCR, we detected 16 positive animals out of 21 (76%). In Farm 4 all animals were infected and in this farm the animals with the highest virus load (ct values around 29) were found.
When sera from animals from Farms 1 and 4 were analyzed in a Western blot using a recombinant C-terminal fragment of gB of PCMV/PRV, all tested animals were reacting positive (Figure 4). Some animals had a very faint reaction, e.g., animal D from farm 1 and animals A and G from farm 4. Animals D, E, and F from farm 4 had a very strong reaction. The result is comparable with a Western blot testing of animals from a German slaughterhouse [48]. The R2 fragment was shown immunodominant in the gB protein [48] and gave similar results when compared with an ELISA using synthetic peptides corresponding to the R2 sequence [53,54]. The assay was used repeatedly to determine the antibody response in different pig breeds [31,49].
When we started the investigation, we did expect a very low number of viruses due to the seclusion of the farms. However, the opposite was observed. This was a great advantage for our investigations because the detection methods can only be tested if viruses are present. Despite the high number of detected viruses, the animals were healthy (the samples were collected from the slaughterhouse).
PLHV-3 was found in all tested indigenous Greek black pigs. This is a similar prevalence compared with other investigations: When 5 pigs in each of 22 farms in Ireland were screened for PLHV, every farm harbored animals infected with PLHV-1 (55%) and 82% of farms scored positive for the presence of PLHV-2 and PLHV-3, respectively [40]. PLHV-1 was the most prevalent of the three species, followed by PLHV-3 and PLHV-2. Coinfections with two or even three viruses were reported. Despite the high prevalence of these viruses, until now, no association between PLHVs and any pig diseases had been described [55]. However, we recently described the finding of PLHV-3 in pigs with dippity pig syndrome [23] and Greek pigs with erythema multiforme [32]. Whether porcine lymphotropic herpesviruses, especially PLHV-3, pose a risk for xenotransplantation, is unclear. The transmission of PCMV/PRV to the progeny can easily be prevented by caesarean section, which is not the case with PLHV. In one study, piglets obtained by somatic cell nuclear transfer (SCNT) and derived via caesarean section, were screened
Table 3.
Screening for pig viruses in liver of indigenous Greek black pigs (mean ct values).
| Animal |
Age (months) |
PCMV/PRV | PLHV-1 |
PLHV -2 |
PLHV -3 |
PPV-1 | PCV2 | PCV3 | PCV4 | HEV | PERV-C | PERV-A/C | |
| Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time RT-PCR | PCR | PCR | |||
| Farm 1 | |||||||||||||
| 1 | 8-9 | n.d. | 33.75 | 33.31 | 28.49 | n.d. | 31.1 | n.d. | n.d. | n.d. | + | - | |
| 2 | 8-9 | 34.31 | n.d. | 28.74 | 28.09 | n.d. | 31.35 | 32.85 | n.d. | n.d. | + | - | |
| 3 | 8-9 | n.d. | n.d. | 27.33 | 34.24 | n.d. | 30.35 | n.d. | n.d. | n.d. | - | - | |
| 4 | 8-9 | 33.49 | n.d. | 27.55 | 26.72 | n.d. | 27.58 | 34.02 | n.d. | n.d. | + | - | |
| Farm 2 | |||||||||||||
| 1 | 11-12 | n.d. | 33.51 | n.d. | 29.91 | n.d. | 30.12 | n.d. | n.d. | n.d. | + | - | |
| 2 | 11-12 | 33.56 | 32.87 | n.d. | 22.74 | n.d. | 32.43 | n.d. | n.d. | n.d. | - | - | |
| 3 | 11-12 | 34.92 | 32.33 | 30.57 | 33.8 | n.d. | 34.52 | 25.17 | n.d. | n.d. | + | - | |
| Farm 3 | |||||||||||||
| 1 | 4 | 33.44 | n.d. | 31.15 | 22.86 | n.d. | 18.66 | 29.69 | n.d. | n.d. | - | - | |
| 2 | 36 | 35.32 | n.d. | 31.98 | 32.19 | n.d. | 33.37 | 29.45 | n.d. | n.d. | - | - | |
| 3 | 4 | 32.00 | n.d. | 32.34 | 27.27 | n.d. | 32.8 | 28.02 | n.d. | n.d. | + | - | |
| 4 | 4 | 31.94 | 28.99 | 27.04 | 36.44 | n.d. | 23.54 | n.d. | n.d. | n.d. | - | - | |
| 5 | 5 | n.d. | 33.46 | n.d. | 31.8 | n.d. | 34.51 | n.d. | n.d. | n.d. | + | - | |
| 6 | 5 | n.d. | 32.44 | n.d. | 34.33 | n.d. | 34.58 | n.d. | n.d. | n.d. | + | - | |
| Farm 4 | |||||||||||||
| 1 | 10-11 | 29.94 | 33.98 | 15.69 | 24.24 | n.d. | 28.99 | n.d. | n.d. | n.d. | - | - | |
| 2 | 10-11 | 29.8 | 28.23 | 25.58 | 24.54 | n.d. | 29.98 | n.d. | n.d. | n.d. | - | - | |
| 3 | 10-11 | 29.42 | 30.22 | n.d. | 31.52 | n.d. | 21.69 | n.d. | n.d. | n.d. | + | - | |
| 4 | 10-11 | 32.60 | n.d. | 29.88 | 27.26 | n.d. | 28.62 | n.d. | n.d. | n.d. | + | - | |
| 5 | 10-11 | 32.47 | n.d. | 28.43 | 20.98 | n.d. | 31.14 | n.d. | n.d. | n.d. | - | - | |
| 6 | 10-11 | 30.21 | 31.38 | 28.85 | 22.51 | n.d. | 26.1 | n.d. | n.d. | n.d. | - | - | |
| 7 | 10-11 | 31.41 | 29.74 | n.d. | 31.79 | n.d. | 23.02 | n.d. | n.d. | n.d. | + | - | |
| 8 | 10-11 | 31.70 | n.d. | 29.83 | 21.95 | n.d. | 26.03 | n.d. | n.d. | n.d. | - | - | |
n.d., not detected; +, positive PCR result; -, negative PCR result.
Table 4.
Comparison of the PCMV virus load in spleen and liver of four pigs in Farm 1.
| Animal | Organ | PCMV | pGAPDH |
|---|---|---|---|
| 1 | spleen | n.d. | 19.10 |
| liver | n.d. | 19.72 | |
| 2 | spleen | 31.34 | 18.58 |
| liver | 34.31 | 19.41 | |
| 3 | spleen | n.d. | 19.57 |
| liver | n.d. | 19.89 | |
| 4 | spleen | 32.32 | 20.00 |
| liver | 33.49 | 19.17 |
n.d., not detected.
using real-time PCR methods. PLHV-3 was detected in five of nine and PLHV-2 in three of nine piglets [56]. In a study transplanting pig kidneys and hearts into immunosuppressed baboons, all donor pigs carried PCMV/PRV, and 55% of them carried PLHV. PCMV was detected in all baboon recipients, but PLHV was not transmitted [57]. PLHV was also not transmitted to baboons through the hearts of eight out of eight genetically modified pigs used for orthotopic pig heart transplantation which were all positive for PLHV-3 [10]. As mentioned, PLHV-3 was also found in pigs suffering from dippity pig syndrome (DPS) [23] and from erythema multiforme [32]. However, it remains unclear whether the virus is involved in the corresponding pathogenesis.
Whereas all animals were positive for PCV2, only 6 animals were positive for PCV3 (Table 3). PCV2 causes porcine circovirus-associated diseases (PCVAD) including subclinical infection (PCV-2-SI), systemic (PCV-2-SD) and reproductive (PCV-2-RD) diseases, and porcine dermatitis and nephropathy syndrome (PDNS) [50,58]. PCV2 was originally identified as the causative agent of Post-Weaning Multisystemic Wasting Syndrome (PMWS) and the respiratory form of PCV2 has been classified as PCV2-associated respiratory disease or PCV2-lung disease (PCV2-LD) [59]. The situation with PCV3, which was also associated with pig diseases, was not clear from the beginning and it was thought that co-infections with other viruses were the reason for these diseases [60]. PCV3 was found in tissues of animals displaying PDNS and reproductive disorders [42]. However, meanwhile it is clear that cloned PCV3 can induce disease in specified pathogen-free (spf) pigs [61,62]. Therefore, is interesting that there are pigs infected with PCV3 without any clinical signs, suggesting that some pig breeds have also a genetic resilience protecting them from the pathogenic properties of PCV3.
PCV4 was described for the first time in China in 2019 [63]. Recently the first detection of PCV4 in Europe, in Spain and in Italy, was reported [64]. Notably, the prevalence of PCV4 was higher in wild boars compared with commercial pigs. The fact that the indigenous Greek black pigs are free from PCV4 demonstrates that the virus has not penetrated into remote Greek regions.
PPV-1 causes infectious infertility [65]. Although this virus is ubiquitous among pigs throughout the world, all indigenous Greek black pigs were free of PPV-1 (Table 3).
At present, HEV3 is the only virus with well-known zoonotic potential [66,67]; all indigenous Greek black pigs were free of HEV3 (Table 3).
Whereas PERV-A and PERV-B are present in the genome of all pigs, PERV-C is not. 11 animals of the 21 tested indigenous Greek black pigs (52%) carried PERV-C in their genome. In some pig breeds even 100% of the animals carry PERV-C (for review see [68]). A low prevalence of PERV-C was found in some farms in the USA (6% up to 41%) [69], and in Chinese miniature pigs (30%) [70,71]. The presence of PERV-C opens the opportunity of a recombination with PERV-A. The resulting recombinant PERV-A/C was characterized by the ability to infect human cells with a high replication rate [72]. PERV-A/C were never found in the germ line, supporting the fact that PERVs are active in living animals, can integrate de novo and recombine [73]. All indigenous Greek black pigs tested were negative for PERV-A/C (Table 3).
The fact that in all indigenous Greek black pigs so many viruses were found even though the animals were healthy, and the samples were taken at the slaughterhouse, is of great interest. It suggests that the animals have a natural resilience to virus infections, obviously they express many antiviral restriction factors protecting them. The situation seems to be similar to that in bats. Bats are recognized as important reservoirs of viruses deadly to other mammals, however, these viral infections are typically nonpathogenic in bats [74]. For example, bats possess more tetherin genes – an antiviral protein, which prevents viral particles from escaping their host cell - than other mammals. Furthermore, some bats encode structurally unique tetherins [75]. Another restriction factor, the tripartite motif-containing protein 5 (TRIM5) was found in multiple copies in bats and TRIM22 was often found duplicated in some bat species, an evolutionary phenomenon not yet observed in any other lineages of mammals [76]. Other bat species possess the largest and most diverse array of APOBEC3 genes, another restriction factor, identified in any mammal reported to date [77]. On the other hand, an excellent immune system may be the reason for the resilience of bats and all indigenous Greek black pigs [78]. Possibly genetic markers could be associated with resistance to infectious diseases. Studies on Italian Large White pigs, wild boars and local breeds indicated that the frequency of the resistance-associated alleles for four polymorphisms was usually higher in local pig breeds, indirectly supporting a higher rusticity of autochthonous breeds than in commercial populations [79]. In a study on indigenous Greek black pigs, the only local pig breed in Greece, it was shown, that this breeds can be the reservoir of interesting genetic variants. In these animals a novel allele in the melanocortin 1 receptor (MC1R) gene was identified, not previously reported in any other pig populations [80]. The novel allele leads to the production of different pigmentation. It was also shown that indigenous Greek black pigs experienced genetic admixture from two sources, wild boars, and cosmopolitan breeds. This situation might raise concerns for the genetic integrity of this animal genetic resource, but on the other hand, this might contribute to within-population genetic variability reducing the problem of inbreeding of the small breed population.
5. Conclusions
Indigenous Greek black pigs were tested for 11 porcine viruses which have relevance for the virus safety of xenotransplantation. The analyses of the sensitivity of the detection methods and the fact that all viruses except for PPV1, PCV4, HEV3 and PERV-A/C were detected, indicates the high sensitivity of the methods. Even though many viruses were detected, the animals were healthy, suggesting that they express active antiviral restriction factors. Further studies are needed to systematically investigate and understand the antiviral resistance of Greek black pigs against common pig viruses.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1-S3: Results of the real-time PCRs using gBlocks: Standard curves indicating the sensitivity.
Author Contributions
Conceptualization, J.D.; methodology, H.J.; validation, H.J.; formal analysis, H.J.; investigation, H.J., L.K., V.P.; resources, V.P., G.P.; data curation, H.J., J.D.; writing—original draft preparation, J.D.; writing—review and editing, H.J., V.P., G.P., L.K., J.D., B.K.; visualization, H.J.; supervision, J.D.; project administration, J.D, B.K.; funding acquisition, J.D, B.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Deutsche Forschungsgemeinschaft, grant number TR127.
Institutional Review Board Statement
All animal procedures regarding animal care, handling and welfare included in the experimental protocol of this study were approved by the Ethical Research Committee of the University of Thessaly (Ethical Approval Code: 104/16.11.2021).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data supporting reported results can be found in the manuscript and the Supplementary material.
Acknowledgments
The authors wish to thank the farmers of Black Greek Pigs as well as the student Konstantinos Stravogiannis for his support during tissue and serum sampling.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Denner, J. The porcine virome and xenotransplantation. Virol. J. 2017, 14, 171. [CrossRef]
- Karlsson, O.E.; Larsson, J.; Hayer, J.; Berg, M.; Jacobson, M. The intestinal eukaryotic virome in healthy and diarrhoeic neonatal piglets. PLoS One. 2016, 11, e0151481. [CrossRef]
- Zhang, B.; Tang, C.; Yue, H.; Ren, Y.; Song, Z. Viral metagenomics analysis demonstrates the diversity of viral flora in piglet diarrhoeic faeces in China. J. Gen. Virol. 2014, 95(Pt 7), 1603-1611. [CrossRef]
- Paim, W.P.; Maggioli, M.F.; Weber, M.N.; Rezabek, G.; Narayanan, S.; Ramachandran, A.; Canal, C.W.; Bauermann, F.V. Virome characterization in serum of healthy show pigs raised in Oklahoma demonstrated great diversity of ssDNA viruses. Virology. 2021, 556, 87-95. [CrossRef]
- Schuele, L.; Lizarazo-Forero, E.; Strutzberg-Minder, K.; Schütze, S.; Löbert, S.; Lambrecht, C.; Harlizius, J.; Friedrich, A.W.; Peter, S.; Rossen, J.W.A.; Couto, N. Application of shotgun metagenomics sequencing and targeted sequence capture to detect circulating porcine viruses in the Dutch-German border region. Transbound. Emerg. Dis. 2022, 69, 2306-2319. [CrossRef]
- Griffith, B.P.; Goerlich, C.E.; Singh, A.K.; Rothblatt, M.; Lau, C.L.; Shah, A.; Lorber, M.; Grazioli, A.; Saharia, K.K.; Hong, S.N.; Joseph, S.M.; Ayares, D.; Mohiuddin, M.M. Genetically Modified Porcine-to-Human Cardiac Xenotransplantation. N. Engl. J. Med. 2022, 387, 35-44. [CrossRef]
- https://www.statnews.com/2023/09/22/pig-heart-transplant-university-of-maryland/ (assessed November 13, 2023).
- Denner, J. Zoonosis and xenozoonosis in xenotransplantation: A proposal for a new classification. Zoonoses Public Health. 2023. [CrossRef]
- Denner, J. Reduction of the survival time of pig xenotransplants by porcine cytomegalovirus. Virol. J. 2018, 15, 171. [CrossRef]
- Denner, J.; Längin, M.; Reichart, B.; Krüger, L.; Fiebig, U.; Mokelke, M.; Radan, J.; Mayr, T.; Milusev, A.; Luther, F.; Sorvillo, N.; Rieben, R.; Brenner, P.; Walz, C.; Wolf, E.; Roshani, B.; Stahl-Hennig, C.; Abicht, J.M. Impact of porcine cytomegalovirus on long-term orthotopic cardiac xenotransplant survival. Sci. Rep. 2020, 10, 17531. [CrossRef]
- Mohiuddin, M.M.; Singh, A.K.; Scobie, L.; Goerlich, C.E.; Grazioli, A.; Saharia, K.; Crossan, C.; Burke, A.; Drachenberg, C.; Oguz, C.; Zhang, T.; Lewis, B.; Hershfeld, A.; Sentz, F.; Tatarov, I.; Mudd, S.; Braileanu, G.; Rice, K.; Paolini, J.F.; Bondensgaard, K.; Vaught, T.; Kuravi, K.; Sorrells, L.; Dandro, A.; Ayares, D.; Lau, C.; Griffith, B.P. Graft dysfunction in compassionate use of genetically engineered pig-to-human cardiac xenotransplantation: a case report. Lancet. 2023, 29,. [CrossRef]
- Denner, J. Virus Safety of Xenotransplantation. Viruses. 2022, 14, 1926. [CrossRef]
- Denner, J. Sensitive detection systems for infectious agents in xenotransplantation. Xenotransplantation. 2020, e12594. [CrossRef]
- Wynyard, S.; Nathu, D.; Garkavenko, O.; Denner, J.; Elliott, R. Microbiological safety of the first clinical pig islet xenotransplantation trial in New Zealand. Xenotransplantation. 2014, 21, 309-23.
- Garkavenko, O.; Dieckhoff, B.; Wynyard, S.; Denner, J.; Elliott, R.B.; Tan, P.L.; Croxson, M.C. Absence of transmission of potentially xenotic viruses in a prospective pig to primate islet xenotransplantation study. J. Med. Virol. 2008, 80, 2046-52. [CrossRef]
- Semaan, M.; Rotem, A.; Barkai, U.; Bornstein, S.; Denner, J. Screening pigs for xenotransplantation: prevalence and expression of porcine endogenous retroviruses in Göttingen minipigs. Xenotransplantation. 2013, 20, 148-56.
- Morozov, V.A.; Morozov, A.V.; Rotem, A.; Barkai, U.; Bornstein, S.; Denner, J. Extended Microbiological Characterization of Göttingen Minipigs in the Context of Xenotransplantation: Detection and Vertical Transmission of Hepatitis E Virus. PLoS One. 2015, 10, e0139893.
- Morozov, V.A.; Ludwig, S.; Ludwig, B.; Rotem, A.; Barkai, U.; Bornstein, S.R.; Denner, J. Islet cell transplantation from Göttingen minipigs to cynomolgus monkeys: analysis of virus safety. Xenotransplantation. 2016, 23, 320-7. [CrossRef]
- Morozov, V.A.; Plotzki, E.; Rotem, A.; Barkai, U.; Denner, J. Extended microbiological characterization of Göttingen minipigs: porcine cytomegalovirus and other viruses. Xenotransplantation. 2016, 23, 490-496.
- Fiebig, U.; Fischer, K.; Bähr, A.; Runge, C.; Schnieke, A.; Wolf, E.; Denner, J. Porcine endogenous retroviruses: Quantification of the copy number in cell lines, pig breeds, and organs. Xenotransplantation. 2018, 25, e12445.
- Krüger, L.; Kristiansen, Y.; Reuber, E.; Möller, L.; Laue, M.; Reimer, C.; Denner, J. A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Göttingen Minipigs. Viruses. 2019, 12, 38.
- Halecker, S.; Krabben, L.; Kristiansen, Y.; Krüger, L.; Möller, L.; Becher, D.; Laue, M.; Kaufer, B.; Reimer, C.; Denner, J. Rare isolation of human-tropic recombinant porcine endogenous retroviruses PERV-A/C from Göttingen minipigs. Virol. J. 2022, 19, 30.
- Jhelum, H.; Grand, N.; Jacobsen, K.R.; Halecker, S.; Salerno, M.; Prate, R.; Krüger, L.; Kristiansen, Y.; Krabben, L.; Möller, L.; Laue, M.; Kaufer, B.; Kaaber, K.; Denner, J. First virological and pathological study of Göttingen Minipigs with Dippity Pig Syndrome (DPS). PLoS One. 2023, 18, e0281521. [CrossRef]
- Plotzki, E.; Heinrichs, G.; Kubícková, B.; Ulrich, R.G.; Denner, J. Microbiological characterization of a newly established pig breed, Aachen Minipigs. Xenotransplantation. 2016, 23, 159-67.
- Halecker, S.; Metzger, J.; Strube, C.; Krabben, L.; Kaufer, B.; Denner, J. Virological and Parasitological Characterization of Mini-LEWE Minipigs Using Improved Screening Methods and an Overview of Data on Various Minipig Breeds. Microorganisms. 2021, 9, 2617.
- Abicht, J.M.; Mayr, T.A.; Reichart, B.; Plotzki, E.; Güthoff, S.; Falkenau, A.; Kind, A.; Denner, J. Hepatic Failure After Pig Heart Transplantation Into a Baboon: No Involvement of Porcine Hepatitis E Virus. Ann. Transplant. 2016, 21, 12-6.
- Morozov, V.A.; Abicht, J.M.; Reichart, B.; Mayr, T.; Guethoff, S.; Denner, J. Active Replication of Porcine Cytomegalovirus (PCMV) Following Transplantation of a Pig Heart into a Baboon despite Undetected Virus in the Donor Pig. Ann. Virol. Res. 2016, 2, 1018.
- Egerer, S.; Fiebig, U.; Kessler, B.; Zakhartchenko, V.; Kurome, M.; Reichart, B.; Kupatt, C.; Klymiuk, N.; Wolf, E.; Denner, J.; Bähr, A. Early weaning completely eliminates porcine cytomegalovirus from a newly established pig donor facility for xenotransplantation. Xenotransplantation. 2018, 25, e12449.
- Fiebig, U.; Abicht, J.M.; Mayr, T.; Längin, M.; Bähr, A.; Guethoff, S.; Falkenau, A.; Wolf, E.; Reichart, B.; Shibahara, T.; Denner, J. Distribution of Porcine Cytomegalovirus in Infected Donor Pigs and in Baboon Recipients of Pig Heart Transplantation. Viruses. 2018, 10, 66.
- Krüger, L.; Längin, M.; Reichart, B.; Fiebig, U.; Kristiansen, Y.; Prinz, C.; Kessler, B.; Egerer, S.; Wolf, E.; Abicht, J.M.; Denner, J. Transmission of Porcine Circovirus 3 (PCV3) by Xenotransplantation of Pig Hearts into Baboons. Viruses. 2019, 11, 650. [CrossRef]
- Halecker, S.; Hansen, S.; Krabben, L.; Ebner, F.; Kaufer, B.; Denner, J. How, where and when to screen for porcine cytomegalovirus (PCMV) in donor pigs for xenotransplantation. Sci. Rep. 2022, 12, 21545. [CrossRef]
- Halecker, S.; Papatsiros, V.; Psalla, D.; Krabben, L.; Kaufer, B.; Denner, J. Virological Characterization of Pigs with Erythema Multiforme. Microorganisms. 2022, 10, 652. [CrossRef]
- Krüger, L.; Stillfried, M.; Prinz, C.; Schröder, V.; Neubert, L.K.; Denner, J. Copy Number and Prevalence of Porcine Endogenous Retroviruses (PERVs) in German Wild Boars. Viruses. 2020, 12, 419.
- Hansen, S.; Menandro, M.L.; Franzo, G.; Krabben, L.; Marino, S.F.; Kaufer, B.; Denner, J. Presence of porcine cytomegalovirus, a porcine roseolovirus, in wild boars in Italy and Germany. Arch. Virol. 2023, 168, 55.
- Homer. The Odyssey. Translation by Ian Johnston. Book Fourteen. Odysseus Meets Eumaeus. Available online: https://www.hellenicaworld.com/Greece/Literature/Homer/en/Odyssey14.html (assessed on July, 27, 2023).
- Michailidou, S.; Kalivas, A.; Ganopoulos, I.; Stea, E.; Michailidis, G.; Tsaftaris, A.; Argiriou, A. A multi-farm assessment of Greek black pig genetic diversity using microsatellite molecular markers. Genet. Mol. Res. 2014, 13, 2752-2765.
- Papakonstantinou, G.I.; Arsenakis, I.; Pourlis, A.; Papatsiros, V.G. Animal Health and Productivity of Organic Greek Pig Farms: The Current Situation and Prospects for Sustainability. Animals. 2023, 13, 2834.
- Mueller, N.J.; Barth, R.N.; Yamamoto, S.; Kitamura, H.; Patience, C.; Yamada, K.; Cooper, D.K.; Sachs, D.H.; Kaur, A.; Fishman, J.A. Activation of cytomegalovirus in pig-to-primate organ xenotransplantation. J. Virol. 2002, 76, 4734-40. [CrossRef]
- Chmielewicz, B.; Goltz, M.; Franz, T.; Bauer, C.; Brema, S.; Ellerbrok, H.; Beckmann, S.; Rziha, H.-J.; Lahrmann, K.-H.; Romero, C.; et al. A novel porcine gammaherpesvirus. Virol. 2003, 308, 317–329. [CrossRef]
- McMahon, K.J.; Minihan, D.; Campion, E.M.; Loughran, S.T.; Allan, G.; McNeilly, F.; Walls, D. Infection of pigs in Ireland with lymphotropic gamma-herpesviruses and relationship to postweaning multisystemic wasting syndrome. Vet. Microbiol. 2006, 116, 60–6.
- Chen, N.; Xiao, Y.; Li, X.; Li, S.; Xie, N.; Yan, X.; Li, X.; Zhu, J. Development and application of a quadruplex real-time PCR assay for differential detection of porcine circoviruses (PCV1 to PCV4) in Jiangsu province of China from 2016 to 2020. Transbound. Emerg. Dis. 2021, 68, 1615–1624. [CrossRef]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J. Virol. 2017, 91, e01879. [CrossRef]
- Opriessnig, T.; Shen, H.G.; Pal, N.; Ramamoorthy, S.; Huang, Y.W.; Lager, K.M.; Beach, N.M.; Halbur, P.G.; Meng, X. A Live-Attenuated Chimeric Porcine Circovirus Type 2 (PCV2) Vaccine Is Transmitted to Contact Pigs but Is Not Upregulated by Concurrent Infection with Porcine Parvovirus (PPV) and Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) and Is Efficacious in a PCV2b-PRRSV-PPV Challenge Model. Clin. Vaccine Immunol. 2011, 18, 1261–1268.
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods. 2006, 131, 65–71.
- Kaulitz, D.; Mihica, D.; Adlhoch, C.; Semaan, M.; Denner, J. Improved pig donor screening including newly identified variants of porcine endogenous retrovirus-C (PERV-C). Arch. Virol. 2013, 158, 341-8.
- Wood, J.C.; Quinn, G.; Suling, K.M.; Oldmixon, B.A.; Van Tine, B.A.; Cina, R.; Arn, S.; Huang, C.A.; Scobie, L.; Onions, D.E.; et al. Identification of Exogenous Forms of Human-Tropic Porcine Endogenous Retrovirus in Miniature Swine. J. Virol. 2004, 78, 2494–2501.
- Duvigneau, J.; Hartl, R.; Groiss, S.; Gemeiner, M. Quantitative simultaneous multiplex real-time PCR for the detection of porcine cytokines. J. Immunol. Methods. 2005, 306, 16–27.
- Plotzki E, Keller M, Ivanusic D, Denner J. A new Western blot assay for the detection of porcine cytomegalovirus (PCMV). J Immunol Methods. 2016, 437, 37-42. [CrossRef]
- Hansen S, Fischer K, Krabben L, Rinke Carrapeiro A, Klinger B, Schnieke A, Kaufer B, Denner J. Detection of porcine cytomegalovirus, a roseolovirus, in pig ovaries and follicular fluid: implications for somatic cells nuclear transfer, cloning and xenotransplantation. Virol J. 2023, 20, 15. [CrossRef]
- Denner J, Mankertz A. Porcine Circoviruses and Xenotransplantation. Viruses. 2017, 9, 83. [CrossRef]
- Morozov, V.A.; Morozov, A.V.; Denner, J. New PCR diagnostic systems for the detection and quantification of the porcine cytomegalovirus (PCMV). Arch. Virol. 2016, 161, 1159–1168. [CrossRef]
- Zou, J.; Liu, H.; Chen, J.; Zhang, J.; Li, X.; Long, Y.; Jiang, Y.; Li, W.; Zhou, B. Development of a TaqMan-Probe-Based Multiplex Real-Time PCR for the Simultaneous Detection of Porcine Circovirus 2, 3, and 4 in East China from 2020 to 2022. Vet. Sci. 2022, 10, 29.
- Fischer N, Gulich B, Keßler B, Längin M, Fishman JA, Wolf E, Boller K, Tönjes RR, Godehardt AW. PCR and peptide based PCMV detection in pig - development and application of a combined testing procedure differentiating newly from latent infected pigs. Xenotransplantation. 2023, 30, e12803. [CrossRef]
- Denner J, Jhelum H, Hansen S, Kaufer BB, Comparison of methods for the detection of porcine cytomegalovirus/roseolovirus in relation to biosafety monitoring of xenotransplantation products. Xenotransplantation, in press.
- Denner J. Porcine Lymphotropic Herpesviruses (PLHVs) and Xenotranplantation. Viruses. 2021;13:1072.
- Hartline, C.B.; Conner, R.L.; James, S.H.; Potter, J.; Gray, E.; Estrada, J.; Tector, M.; Tector, A.J.; Prichard, M.N. Xenotransplantation panel for the detection of infectious agents in pigs. Xenotransplantation 2018, 25, e12427. [CrossRef]
- Mueller, N.J.; Livingston, C.; Knosalla, C.; Barth, R.N.; Yamamoto, S.; Gollackner, B.; Dor, F.J.; Buhler, L.; Sachs, D.H.; Yamada, K.; et al. Activation of porcine cytomegalovirus, but not porcine lymphotropic herpesvirus, in pig-to-baboon xenotransplantation. J. Infect.Dis. 2004, 189, 1628–1633.
- Segalés J, Sibila M. Revisiting Porcine Circovirus Disease Diagnostic Criteria in the Current Porcine Circovirus 2 Epidemiological Context. Vet Sci. 2022, 9, 110.
- Opriessnig T, Meng XJ, Halbur PG. Porcine circovirus type 2 associated disease: update on current terminology, clinical manifestations, pathogenesis, diagnosis, and intervention strategies. J Vet Diagn Investig. 2007, 19, 591–615.
- Faccini S, Barbieri I, Gilioli A, Sala G, Gibelli LR, Moreno A, Sacchi C, Rosignoli C, Franzini G, Nigrelli A. Detection and genetic characterization of Porcine circovirus type 3 in Italy. Transbound. Emerging Dis. 2017, 64, 1661. [CrossRef]
- Jiang, H., Wang, D., Wang, J., Zhu, S., She, R., Ren, X., Tian, J., Quan, R., Hou, L., Li, Z., Chu, J., Guo, Y., Xi, Y., Song, H., Yuan, F., Wei, L., Liu, J., 2019. Induction of Porcine Dermatitis and Nephropathy Syndrome in Piglets by Infection with Porcine Circovirus Type 3. J. Virol. 2019, 93, e02045-18.93. [CrossRef]
- Franzo, G., He, W., Correa-Fiz, F., Li, G., Legnardi, M., Su, S., Segalés, J., 2019b. A Shift in Porcine Circovirus 3 (PCV-3) History Paradigm: Phylodynamic Analyses Reveal an Ancient Origin and Prolonged Undetected Circulation in the Worldwide Swine Population. Adv Sci (Weinh) 2019, 6, 1901004.
- Zhang HH, Hu WQ, Li JY, et al. Novel circovirus species identified in farmed pigs designated as Porcine circovirus 4, Hunan province, China. Transboundary and emerging diseases. 2020, 67, 1057-1061.
- Holgado-Martín R, Arnal JL, Sibila M, Franzo G, Martín-Jurado D, Risco D, Segalés J, Gómez L. First detection of porcine circovirus 4 (PCV-4) in Europe. Virol J. 2023, 20, 230.
- Mengeling W, Lager K, Vorwald A. The effect of porcine parvovirus and porcine reproductive and respiratory syndrome virus on porcine reproductive performance. Anim Reprod Sci. 2000, 60, 199–210. [CrossRef]
- Wang B, Meng XJ. Hepatitis E virus: host tropism and zoonotic infection. Curr Opin Microbiol. 2021, 59, 8-15. [CrossRef]
- Denner J. Xenotransplantation and Hepatitis E virus. Xenotransplantation. 2015, 22, 167-73. [CrossRef]
- Krüger L, Stillfried M, Prinz C, Schröder V, Neubert LK, Denner J. Copy Number and Prevalence of Porcine Endogenous Retroviruses (PERVs) in German Wild Boars. Viruses. 2020, 12, 419. [CrossRef]
- Pal, N.; Baker, R.; Schalk, S.; Scobie, L.; Tucker, A.W.; Opriessnig, T. Detection of porcine endogenous retrovirus (PERV) viremia in diseased versus healthy US pigs by qualitative and quantitative real-time RT-PCR. Transbound. Emerg. Dis. 2011, 58, 344–351.
- Liu, G.; Li, Z.; Pan, M.; Ge, M.; Wang, Y.; Gao, Y. Genetic prevalence of porcine endogenous retrovirus in chinese experimental miniature pigs. Transplant. Proc. 2011, 43, 2762–2769. [CrossRef]
- Wu, J.; Ma, Y.; Lv, M.; Yang, Y.; Guo, Y.; Yu, X.; Tian, K.; Zhang, J. Large-scale survey of porcine endogenous retrovirus in Chinese miniature pigs. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 367–371. [CrossRef]
- Harrison I, Takeuchi Y, Bartosch B, Stoye JP. Determinants of high titer in recombinant porcine endogenous retroviruses. J Virol. 2004, 78, 13871-1389. [CrossRef]
- Scobie L, Taylor S, Wood JC, Suling KM, Quinn G, Meikle S, Patience C, Schuurman HJ, Onions DE. Absence of replication-competent human-tropic porcine endogenous retroviruses in the germ line DNA of inbred miniature Swine. J Virol. 2004, 78, 2502-2509.
- Hayward JA, Tachedjian M, Johnson A, Irving AT, Gordon TB, Cui J, Nicolas A, Smith I, Boyd V, Marsh GA, Baker ML, Wang LF, Tachedjian G. Unique Evolution of Antiviral Tetherin in Bats. J Virol. 2022, 96, e0115222.
- Fernandes AP, Águeda-Pinto A, Pinheiro A, Rebelo H, Esteves PJ. Evolution of TRIM5 and TRIM22 in Bats Reveals a Complex Duplication Process. Viruses. 2022, 14, 345.
- Hayward JA, Tachedjian M, Cui J, Cheng AZ, Johnson A, Baker ML, Harris RS, Wang LF, Tachedjian G. Differential Evolution of Antiretroviral Restriction Factors in Pteropid Bats as Revealed by APOBEC3 Gene Complexity. Mol Biol Evol. 2018, 35, 1626-1637. [CrossRef]
- Weinberg M, Yovel Y. Revising the paradigm: Are bats really pathogen reservoirs or do they possess an efficient immune system? iScience. 2022, 25, 104782.
- Calisher, C.H., Childs, J.E., Field, H.E., Holmes, K.V., and Schountz, T. Bats: important reservoir hosts of emerging viruses. Clin.Microbiol. Rev. 2006, 19, 531–545. [CrossRef]
- Geraci C, et al., Genetic markers associated with resistance to infectious diseases have no effects on production traits and haematological parameters in Italian Large White pigs, Livestock Science 2019, 223, 32-38. [CrossRef]
- Ribani A. et al. Signatures of Admixture and Genetic Uniqueness in the Autochthonous Greek Black Pig Breed Deduced from Gene Polymorphisms Affecting Domestication-Derived Traits. Animals 2023, 13, 1763. [CrossRef]
Figure 1.
Indigenous Greek black pigs on a mountainous pasture in Greece (Photo: Vasileios Papatsiros).
Figure 1.
Indigenous Greek black pigs on a mountainous pasture in Greece (Photo: Vasileios Papatsiros).

Figure 2.
Localization of the farms with indigenous Greek black pigs analyzed here.
Figure 3.
Results of the PCR testing for PERV-C. Animals from all farms were tested, DNA was from the liver. PC, positive control, NTC, negative control.
Figure 3.
Results of the PCR testing for PERV-C. Animals from all farms were tested, DNA was from the liver. PC, positive control, NTC, negative control.
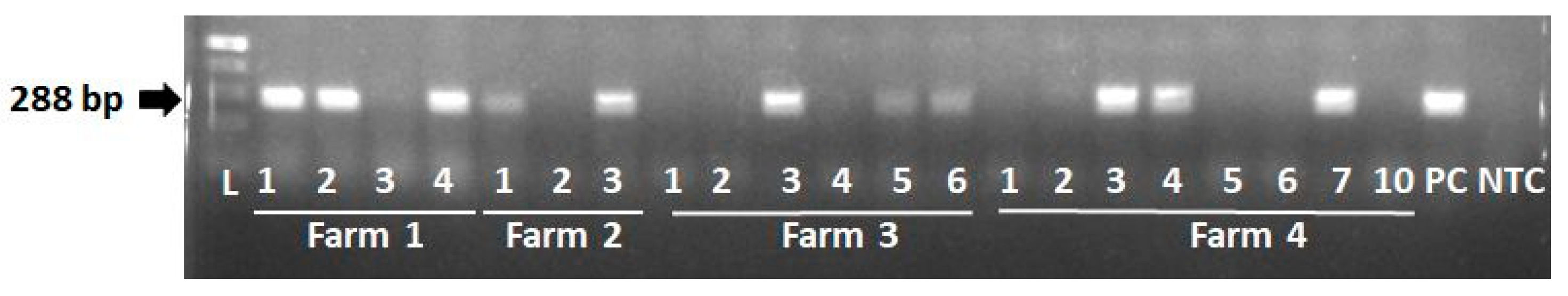
Figure 4.
Results of the Western blot analysis to detect antibodies against PCMV/PRV. Animals A, B, C, and D from farm 1 and animals A to G from farm 4 were tested. PC, positive control.
Figure 4.
Results of the Western blot analysis to detect antibodies against PCMV/PRV. Animals A, B, C, and D from farm 1 and animals A to G from farm 4 were tested. PC, positive control.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



