
Submitted:
30 November 2023
Posted:
01 December 2023
You are already at the latest version
Abstract
Climate change and the increasing frequency of high-temperature events are significant threats to global crop yields. To address this, a comprehensive understanding of how plants respond to heat shock (HS) is essential. Signaling pathways involving calcium (Ca2+), a versatile second messenger in plants, encode information through temporal and spatial variations in ion concentration. Ca2+ is detected by Ca2+-sensing effectors, including channels and binding proteins, which trigger specific cellular responses. At elevated temperatures, the cytosolic concentration of Ca2+ in plant cells increases rapidly, making Ca2+ signals the earliest response to HS. In this review, we discuss the crucial role of Ca2+ signaling in raising plant thermotolerance and we explore its multifaceted contributions to various aspects of the plant HS response.
Keywords:
crop yield
; thermotolerance
; Ca2+ signaling
1. Ca2+ signaling and plant thermotolerance
High temperatures can elevate the intracellular concentration of calcium (Ca2+), a common second messenger in both animal and plant cells. Plants exposed to high temperatures can experience heat shock (HS), which triggers a rise in cytosolic Ca2+ in seedling leaves and which disrupts the oscillations in Ca2+ level associated with pollen tube growth. Ca2+ plays a critical role in maintaining the normal physiological functions of plant cells and is involved in various physiological processes in plants. Additionally, as a ubiquitous second messenger, Ca2+ participates in plant responses to various stressors. Thus, understanding the impact of Ca2+ on plant thermotolerance is essential for the development of heat-resistant crops [3,4].
Ca2+ is a versatile intracellular signal; information is encoded based on temporal and spatial patterns of Ca2+ concentration changes. These patterns are decoded by Ca2+-sensing effectors such as Ca2+-permeable channels and Ca2+-binding proteins to initiate specific cellular responses [4]. The induction of Ca2+ signals represents the most rapid response to elevated temperatures in plants. In one study, the cytosolic Ca2+ concentration peaked within 10–15 min of a sudden temperature increase from 24 to 36°C before returning gradually to baseline as the HS response continued [5].
Ca2+ entry into the cytoplasm is facilitated by several families of protein channels, including cyclic nucleotide-gated channels (CNGCs), glutamate (Glu) receptor-like channels (GLRs), annexins, and mechanosensitive (MS) channels. Each of these channel types plays a crucial role in promoting the influx of Ca2+. Furthermore, plants possess various Ca2+-binding proteins that decode and transmit the primary Ca2+ signal to elicit specific cellular responses. These proteins include calmodulins (CaMs), CaM-like proteins (CMLs), Ca2+-dependent protein kinases (CDPKs or CPKs), Ca2+- and CaM-dependent protein kinases (CCaMKs), calcineurin B-like proteins (CBLs), and CBL-interacting protein kinases (CIPKs). When these proteins bind Ca2+, they undergo conformational changes that allow them to initiate downstream signaling events via interactions with other proteins or molecules. This mechanism enables plants to translate variations in the intracellular concentration of Ca2+ into specific cellular responses, including changes in gene expression, enzyme activity, or ion channel activity [6,7,8].
Recent work has focused on understanding how plants detect Ca2+ increases due to HS, and many studies have identified crucial molecules and signaling pathways involved. Here, we review the latest findings on thermosensing in different crop species.
2. Ca2+-permeable channels perceive elevated temperatures
Presently, the identity of the definitive thermosensor in plants remains elusive. It is hypothesized that Ca2+ channels located in the plasma membrane are crucial players in perceiving elevated temperatures. These channels could be activated directly or indirectly, leading to increased cytoplasmic Ca2+ levels. Consequently, heat-stress sensors may function as Ca2+ channels, directly modulating Ca2+ signaling in response to external stimuli. Alternatively, they may act as regulatory elements that influence the membrane lipid composition, which is closely linked to Ca2+ channel activity. Another possibility is that they operate as receptor-like kinases, initiating an influx of Ca2+ in response to external stimuli.
Genome sequencing has revealed that plants lack typical animal Ca2+ channels such as voltage-dependent Ca2+ channels, transient receptor potential channels, purinergic P2X receptor channels, and cysteine loop channels. Instead, they have expanded families of CNGCs, GLRs, annexins, reduced hyperosmolarity-induced [Ca2+]i increase channels (OSCAs), “Mid1-complementing activity” channels (MCAs), two-pore channels (TPCs), MS-like channels (MSLs), and Piezo channels (MSPs). In the context of HS responses, we will explore several potential candidates among these Ca2+-permeable ion channels (Figure 1 and Table 1).
2.1. Heat sensing via CNGCs
CNGCs are non-specific cation channels that regulate the flow of Ca2+ by binding to ligands such as cAMP and cGMP. CNGCs are activated in response to various abiotic and biotic stresses [9,10]. Plant and animal CNGCs belong to the K+-selective shaker channel family and share similarities in their amino acid sequences and overall structures [11]. They possess cyclic nucleotide-binding domains and one or more CaM-binding domains at their cytosolic N- and C-termini [12,13].
HS induces plasma membrane fluidization, allowing an influx of Ca2+ into the cytoplasm. Furthermore, nucleotide cyclases can elevate cAMP and cGMP levels under HS conditions, further promoting the influx of Ca2+ and activating associated signaling pathways. CNGCs were initially discovered in barley [14]; subsequent studies identified 20 and 16 CNGCs in the genomes of Arabidopsis and rice, respectively [15,16]. Currently, plant CNGCs are known to participate in a range of biological processes [17,18,19,20].
The involvement of CNGCs in plant HS responses was initially suggested through studies of Arabidopsis CNGC2 and Physcomitrella patens CNGCb null mutants. These studies revealed that a loss of CNGC2/CNGCb function enhanced HS tolerance at the seedling stage, accompanied by a moderate increase in the accumulation of heat-responsive proteins compared to wild-type plants [5,20]. Intriguingly, AtCNGC2 dysfunction has been linked to thermosensitivity at the reproductive stage, indicating a developmental stage-dependent role in HS responses [21]. Recent research also revealed differential regulation of AtCNGC4 in shoots and roots under variable temperature conditions [22]. In Arabidopsis, a study employing genetic analysis and whole-cell patch-clamp techniques showed that CNGC6 mediates a heat-induced Ca2+ influx, promoting the expression of genes encoding HS proteins (HSPs). Nitric oxide (NO) and hydrogen peroxide (H2O2) have been identified as downstream partners of CNGC6 that contribute to increased thermotolerance [23,24,25].
In rice, oscngc14 and oscngc16 mutant plants exhibited reduced survival rates, elevated levels of H2O2, and increased cell death under HS conditions. These findings indicate that OsCNGC14 and OsCNGC16 modulate thermotolerance by regulating cytosolic Ca2+ levels in response to HS, underscoring their critical roles in HS responses in rice [26]. Interestingly, of the 16 CNGC genes in rice, 10 were notably upregulated under low-temperature conditions, warranting further investigation into their expression changes in response to heat stress. Additionally, CNGC genes from other plants (e.g., P. patens, Nicotiana tabacum, Brassica oleracea, and Mangifera indica) have been suggested to contribute to HS tolerance, indicating their potential importance in dealing with extreme temperature conditions [27].
2.2. Heat sensing via GLRs
In mammals, ionotropic Glu receptor cation channels (iGluRs) respond to the neurotransmitter Glu, initiating Ca2+ signal cascades [28]. The first evidence of Glu signaling in plants was uncovered by Lam et al. [29], who identified the AtGLR gene family in Arabidopsis. Through sequence analysis and comparisons, homologous Glu receptor variants have been identified in dicot and monocot species [30,31,32]. The Arabidopsis genome contains 20 GLRs, while the rice and poplar genomes harbor 13 and 61 GLRs, respectively [33,34]. Plant GLRs display significant sequence and structural homologies with animal iGluRs, including high degrees of amino acid sequence identity in their channel and ligand-binding domains [35].
Studies on the physiological roles of GLRs have revealed their involvement in diverse processes such as photosynthesis, carbon/nitrogen balance, Ca2+ regulation, root architecture, pollen tube growth, defense signaling, and environmental stress responses [36,37,38,39,40]. For example, GLR3.3 has been implicated in the response of Arabidopsis to pathogen infection [41], while GLR3.5 promotes drought tolerance in faba bean [42]. In tomato, GLR3.3 and GLR3.5 mediate cold stress tolerance by regulating apoplastic H2O2 production and redox homeostasis [43].
Although research on the involvement of GLR genes in plant thermotolerance is limited, some studies indicate a potential role in responding to HT stress. For instance, one study investigated the effect of Glu on the survival of maize seedlings under HS conditions and its impact on Ca2+ signaling [44]. The results showed that pretreatment with Glu enhanced the thermotolerance of maize seedlings, possibly through GLR-mediated Ca2+ signaling. Glu is emerging as a novel signaling molecule with involvement in a wide range of physiological processes in plants. Further exploration is needed to uncover the specific functions of GLRs, particularly in the context of thermotolerance. Additional research methodologies should be employed to elucidate the mechanisms by which GLRs contribute to thermotolerance in plants.
2.3. Heat sensing via annexins
Annexins, an evolutionarily conserved family of proteins present in a variety of organisms, are renowned for their ability to bind to Ca2+ and phospholipids. They play pivotal roles in a plethora of cellular processes [45,46]. Annexins exhibit peroxidase and ATPase/GTPase activities and are associated with the regulation of Ca2+ channels [47]. Their functions encompass a wide range of intracellular and extracellular phenomena, including vesicular trafficking, organization of the membrane–cytoskeleton, exocytosis, endocytosis, phagocytosis, ion channel regulation, and apoptosis [48]. The genomes of Arabidopsis, rice, and wheat contain 8, 10, and 25 annexin genes, respectively [49,50,51]. Annexins feature a conserved protein core domain capable of binding both Ca2+ and phospholipids, alongside an N-terminal domain that varies in sequence and length among different annexins [52].
In plants, annexins participate in environmental stress responses and they play roles in growth, development, and signaling [53]. Some plant annexins regulate the level of cytosolic free Ca2+, and certain annexins can form Ca2+-permeable channels in lipid bilayers or vesicles [54]. Moreover, plant annexins may possess peroxidase activity or ATPase/GTPase activity, each contributing to functional specificity [55]. Annexins play a broad regulatory role in diverse biochemical and cellular processes, including Ca2+ channel regulation, and in plant growth, development, and biotic and abiotic environmental stress responses [55].
Several studies have highlighted the pivotal role of annexins in plant thermotolerance. For example, NnANN1, a heat-induced annexin, was identified in the embryonic axes of the sacred lotus (Nelumbo nucifera Gaertn.) through comparative proteomics. NnANN1 expression increased substantially in response to HT treatment. Ectopic expression of NnANN1 in Arabidopsis enhanced the HS tolerance of transgenic seeds. These seeds exhibited increased peroxidase activity, reduced lipid peroxidation, and reduced reactive oxygen species (ROS) levels compared to wild-type seeds [56]. Among crop plants, the soybean annexin GmANN has been found to promote tolerance to high temperatures and humidity and to improve seed vigor. GmANN-transgenic Arabidopsis seeds displayed enhanced heat resistance and greater seed vitality under HT stress and high humidity compared to wild-type seeds [57]. Additionally, GmANN overexpression in plants led to increased peroxidase activity, decreased lipid peroxidation, and reduced ROS levels compared to wild-type plants. OsANN1, a rice annexin, has been reported to enhance abiotic stress tolerance by modulating antioxidant accumulation. Overexpression of OsANN1 promoted the activities of superoxide dismutase (SOD) and catalase (CAT), which regulate the H2O2 content and redox homeostasis in cells. This suggests the existence of a feedback loop that controls OsANN1 and H2O2 production under conditions of abiotic stress. Elevated OsANN1 expression conferred cellular protection against HS and H2O2, with significant cytosolic localization observed following HT treatment [58]. In another study, a phylogenetic analysis revealed that radish annexins (RsANNs), along with Arabidopsis and rice annexins, clustered into five groups with similar motif patterns. Real-time quantitative PCR showed that most RsANN genes are induced by such abiotic stressors as heat, drought, salinity, oxidation, and abscisic acid (ABA). Furthermore, overexpression of RsANN1a enhanced the growth and thermotolerance, while the knockdown of RsANN1a using artificial microRNAs resulted in decreased survival, of HS-treated Arabidopsis plants [59].
Annexins constitute a diverse multigene family with multifaceted roles in plants; however, our understanding of their functions is still in its infancy. Further research is needed to delve into the potential mechanisms by which annexin genes contribute to thermotolerance in plants.
2.4. Heat sensing via OSCAs
Recently discovered MS Ca2+ channels called OSCAs have the ability to detect and respond to changes in osmotic pressure, regardless of whether it originates externally or internally. OSCAs are essential for regulating the flow of Ca2+ in plants; thus, they play a crucial role in plant growth and adaptation to environmental stress. OSCA1, the first OSCA identified in Arabidopsis thaliana, is responsible for raising cytosolic Ca2+ levels by triggering an influx of Ca2+. In fact, this channel is responsible for the increase in intracellular Ca2+ triggered by multiple stimuli in plants [60]. In total, 15, 11, 12, and 62 OSCA genes have been identified in Arabidopsis [60], rice [61], Zea mays [62], and cotton [63], respectively.
In rice, 11 OsOSCA genes were identified from the Oryza sativa L. Japonica genome database. These genes were categorized into four clades based on their sequence composition and phylogenetic relationships. Their expression profiles were assessed at different stages of vegetative growth, during reproductive development, and in response to osmotic stress. Remarkably, differential expression due to osmotic stress was noted for 10 of the 11 genes [61].
Similarly, in maize, a comprehensive analysis of the ZmOSCA gene family was conducted through bioinformatics and expression studies. A total of 12 OSCA genes were identified from the maize genome database. These genes were classified into four groups (I–IV) based on their sequence composition and phylogenetic relationships. Notably, when maize plants were subjected to HS (40°C), OSCA gene expression exhibited two distinct patterns. Six genes, ZmOSCA1.4, ZmOSCA2.1, ZmOSCA2.2, ZmOSCA2.5, ZmOSCA3.1, and ZmOSCA4.1, displayed a significant increase in expression after 1 h of HT stress, reached a peak, and then underwent rapid downregulation, followed by a gradual increase. Meanwhile, the expression patterns of ZmOSCA1.3, ZmOSCA1.5, and ZmOSCA2.4 were contrary to those of all other maize OSCA family members [64]. These findings strongly suggest that OSCA genes play a crucial role in mediating plant responses to HS.
2.5. The functions of MSLs, MCAs, and MSPs in plants following HT treatment
In addition to OSCAs, plants possess several other types of MS ion channels, including MSLs, MCAs, and MSPs. These channels are permeable to Ca2+ and play crucial roles in enabling plants to respond to mechanical stimuli and changes in osmotic pressure.
MSLs are MS ion channels found in both bacteria and plants. In Arabidopsis, MSL2 and MSL3 are essential to protect plastids against bursting inside leaf epidermal cells during growth under conditions of high osmotic pressure [65]. MSL8 responds to plasma membrane distortion during pollen grain rehydration and germination [66]. Additionally, MSL10 and MSL9 exhibit MS ion channel activity in root protoplasts [67]. In rice, most OsMSL genes are expressed during reproductive growth, suggesting their involvement in plant growth, development, and stress responses [68].
MCAs are plant-specific MS Ca2+-permeable channels found in such plants as Arabidopsis, rice, and N. tabacum [69,70]. In Arabidopsis, two such channels, MCA1 and MCA2, have been identified. MCA1 and MCA2 were shown to mediate a Ca2+ influx in yeast cells and to function in the rapid elevation of cytosolic Ca2+ levels during cold exposure [71]. Wild-type Arabidopsis (Col-0) plants exhibited higher cold-induced cytosolic Ca2+ concentrations compared to mca1 and mca2 mutant plants. Conversely, the double mutant mca1 mca2 showed freezing and chilling sensitivity, indicating that MCA1 and MCA2 confer cold stress tolerance to plants [72].
While MSPs are primarily found in animals, one gene encoding an MSP has been identified in Arabidopsis [73,74]. AtPiezo, an uncharacterized MSP in Arabidopsis and an ortholog of animal MSPs, was recently discovered [75]. AtPiezo was necessary to inhibit a systemic infection by CMV-2aT∆2b or turnip mosaic virus tagged with green fluorescent protein (TuMV-GFP). Viral infection induced AtPiezo expression, especially in the petioles of rosette leaves. This study marks the first demonstration of the biological function of an MSP in plants. Further, it suggests that this mechanism represents a common antiviral strategy since many monocots and dicots possess a single MSP ortholog.
While the direct involvement of these channels in HS responses has not been extensively studied, their potential role in such responses warrants further investigation.

Table 1.
Ca2+ channels that are likely involved in sensing heat.
| Gene type | Organism | Gene symbol | Function | References |
|---|---|---|---|---|
| CNGCs | Physcomitrella patens | CNGCb | Sensitive to heat stress | [5] |
| Arabidopsis thaliana | AtCNGC2 | Sensitive to heat stress at the seedling stage; Tolerance to heat stress at the reproductive stage | [5,20,21] | |
| AtCNGC4 | Tolerance to extreme temperatures; Response to pathogen infection | [22] | ||
| AtCNGC6 | Regulates tolerance to extreme temperatures together with H2O2 and NO | [23,24,25] | ||
| Oryza sativa L. | OsCNGC14 | Tolerance to extreme temperatures | [26] | |
| OsCNGC16 | Tolerance to extreme temperatures | [26] | ||
| GLRs | Arabidopsis thaliana | AtGLR3.3 | Response to pathogen infection | [41] |
| Vicia faba L. | VfGLR3.5 | Tolerance to drought | [42 | |
| Zea mays L. | ZmGLR | Tolerance to heat stress | [44] | |
| Solanum lycopersicum L. | SlGLR3.3 | Tolerance to cold stress by regulating apoplastic H2O2 production and redox homeostasis | [43] | |
| SlGLR3.5 | Tolerance to cold stress by regulating apoplastic H2O2 production and redox homeostasis | [43] | ||
| ANNEXINs | Nelumbo nucifera Gaertn. | NnANN1 | Tolerance to heat stress | [56] |
| Glycine max L. | GmANN | Tolerance to high temperatures and humidity stress | [57] | |
| Oryza sativa L. | OsANA1 | Tolerance to heat shock, H2O2 treatment, and abiotic stress | [58] | |
| Raphanus sativus L. | RsANN | Tolerance to heat, drought, salinity, oxidation, and ABA stress | [59] | |
| OSCAs | Zea mays L. | ZmOSCA1.4 | Gene expression increases in response to heat stress | [64] |
| ZmOSCA2.1 | Gene expression increases in response to heat stress | [64] | ||
| ZmOSCA2.2 | Gene expression increases in response to heat stress | [64] | ||
| ZmOSCA2.5 | Gene expression increases in response to heat stress | [64] | ||
| ZmOSCA3.1 | Gene expression increases in response to heat stress | [64] | ||
| ZmOSCA4.1 | Gene expression increases in response to heat stress | [64] | ||
| ZmOSCA1.3 | Gene expression decreases in response to heat stress | [64] | ||
| ZmOSCA1.5 | Gene expression decreases in response to heat stress | [64] | ||
| ZmOSCA2.4 | Gene expression decreases in response to heat stress | [64] | ||
| MSLs | Arabidopsis thaliana | AtMSL2 | Tolerance to high osmotic stress | [65] |
| AtMSL3 | Tolerance to high osmotic stress | [65] | ||
| AtMSL8 | Response to plasma membrane distortion during pollen grain rehydration and germination | [66] | ||
| AtMSL9 | Exhibits MS ion channel activity | [67] | ||
| AtMSL10 | Exhibits MS ion channel activity | [67] | ||
| Oryza sativa L. | OsMSLs | Responses to plant growth, development, and various stressors | [68] | |
| MCAs | Arabidopsis thaliana | AtMCA1 | Tolerance to cold stress | [72] |
| AtMCA2 | Tolerance to cold stress | [72] | ||
| MSPs | Arabidopsis thaliana | AtPiezo | Response to virus infection | [75] |
3. Ca2+-binding protein involvement in HS responses
Under HS conditions, the intracellular Ca2+ concentration in plants increases, primarily due to an influx of Ca2+ from outside the cell, or release of Ca2+ from intracellular stores [5,76]. This rise serves as a pivotal “signal” that triggers a complex signal transduction cascade. This signal, in conjunction with downstream effector proteins, including CaMs, CMLs, CBLs, and CDPKs/CPKs, serves as a molecular code that must be interpreted by the cell. Together, these proteins create a sophisticated signaling network that transmits stress signals and orchestrates a multitude of responses in plants [77]. In this section, we will concentrate on candidate Ca2+-binding proteins that have been reported to be involved in HS responses in crops (Figure 1 and Table 2).
3.1. CaMs in HS signaling
CaMs, highly conserved sensor proteins containing EF-hand motifs, are found in various plant organelles. In total, 9, 5, and 8 CaM genes have been identified in Arabidopsis [78,79], rice [80], and maize [81], respectively. CaMs are ubiquitously expressed in all eukaryotic cells and possess a dumbbell-shaped structure. They are multifunctional, consisting of two globular lobes at the N- and C-termini connected by a flexible central linker. Each lobe contains a pair of EF-hand motifs capable of binding Ca2+ ions with positive cooperativity [82]. The EF-hand motif in CaM is characterized by a helix-loop-helix (D-X-D) conformation, consisting of 12 amino acid residues, totaling 36 amino acid residues across both motifs. In the D-X-D motif, the 14th and 16th positions are fixed, while the 15th position can be occupied by any amino acid [83]. These motifs are involved in interactions with a diverse array of downstream target proteins, including ion channels, pumps, antiporters, kinases, phosphatases, transcription factors, and enzymes involved in metabolic pathways. In some instances, these motifs function as transcription factors, recognizing and binding to target genes, thereby modulating stress responses in plants.
Several studies have highlighted the critical role of CaMs in plant thermotolerance. In Arabidopsis, which possesses nine CAM genes, the mRNA levels of these genes were examined in root and shoot tissues from seedlings exposed to normal and HS temperatures. The mRNA levels of all of the CAM genes, except for CAM5 in the root and shoot and CAM1 in the shoot, were up-regulated in response to HS treatment [79]. Another study in Arabidopsis revealed reduced thermotolerance in knockout mutants of AtCaM3 after heat treatment at 45°C for 50 min. Conversely, overexpression of AtCaM3 in either an AtCaM3 knockout or wild-type background significantly enhanced thermotolerance. These findings underscore the significance of endogenous AtCaM3 in the Ca2+/CaM HS signal transduction pathway [84]. NO also plays a role in thermotolerance in Arabidopsis seedlings by acting upstream of AtCaM3. Following HS treatment, NO stimulates the DNA-binding activity of HS transcription factors and the accumulation of HSP18.2 through AtCaM3. This suggests that NO functions in HS signaling and acts upstream of AtCaM3 in thermotolerance, dependent on increased HS transcription factor DNA-binding activity and HSP accumulation [85]. Additionally, overexpression of CsCaM3, a CaM gene isolated from the cucumber inbred line “02-8,” has been shown to enhance HT tolerance in cucumber plants [86]. That study found that CsCaM3 transcription was induced by HS or ABA. Overexpression of CsCaM3 in cucumber plants improved their thermotolerance and protected against both oxidative damage and damage to the photosynthesis system by regulating the expression of HT-responsive genes, including those related to chlorophyll catabolism, under HS stress. In rice, HS was found to cause rapid increases in the cytosolic Ca2+ concentration and in the expression and nuclear localization of OsCaM1-1 [87]. These changes are needed to mediate downstream HS-related gene expression, which contributes to the acquisition of thermotolerance in rice. Furthermore, OsCaM1-1 induced the expression of Ca2+/HS-related genes such as AtCBK3, AtPP7, AtHSF, and AtHSP (even at non-inducing temperatures) and it enhanced thermotolerance in transgenic Arabidopsis plants. These findings highlight the significant role of OsCaM1-1 as a mediator of downstream HS signaling.
3.2. CMLs in HS signaling
CMLs differ from CaMs in their length and the number of EF-hand motifs they possess. While CaMs typically contain four EF-hand motifs, CMLs can have varying numbers, ranging from one to six. CMLs are termed “CaM-like” due to their 15% amino acid identity with CaM. These proteins play a crucial role in physiological responses to various stresses, including salinity, drought, heat, and cold. In Arabidopsis, 50 CMLs have been identified, compared to 32 in rice [88].
In Arabidopsis, a rise in external Ca2+ or HS significantly increased the mRNA levels of AtCML12 and AtCML24 [89]. Both stimuli are known to increase cytoplasmic Ca2+, suggesting a role for Ca2+ itself in the regulation of CaM-related genes. In rice, another study demonstrated that the expression of OsMSR2, a CML gene, was strongly up-regulated by a wide range of stresses, including cold, drought, and heat, in different tissues and at different developmental stages [90]. The pronounced induction of OsMSR2 expression by HS suggests its involvement in thermotolerance. Furthermore, SlCML39, a CML gene from tomato, was found to have a negative impact on thermotolerance during germination and seedling growth in A. thaliana [91]. SlCML39 is expressed in various tissues in tomato plants, including leaves, stems, roots, flowers, and fruits. Overexpression of SlCML39 in A. thaliana resulted in reduced germination rates and compromised seedling growth under HT conditions. Thus, SlCML39 may play a regulatory role in plant responses to HT stress.
Recent research has significantly advanced our understanding of the roles of CaMs and CMLs as Ca2+ sensors in plants. CaMs and CMLs serve as central hubs for integrating various signal transduction mechanisms, allowing cells to respond appropriately to multiple environmental stimuli. Future research efforts are expected to address many of the remaining questions concerning the roles of CaMs and CMLs in plant thermotolerance.
3.3. CDPKs in HS signaling
CDPKs, or CPKs, belong to a class of protein kinases that possess four C-terminal EF-hand motifs, which enable them to sense changes in intracellular Ca2+ levels. CDPKs are effector proteins that play pivotal roles in regulating a wide range of physiological processes, including environmental stress responses, in various plant cell types [92]. CDPKs consist of several domains, including a variable N-terminal domain, a serine/threonine protein kinase domain, an autoinhibitory junction domain (JD), and a C-terminal CML regulatory domain (CaMLD) connected by a tether. The CaMLD is composed of four EF-hand Ca2+-binding motifs. At low Ca2+ concentrations, the C-lobe of a CDPK already contains Ca2+ and interacts with the JD, stabilizing its conformation. The JD forms a helical structure that blocks substrate access by being buried within the active site of the kinase domain due to an intramolecular interaction. When the concentration of Ca2+ rises, both the N-lobe and C-lobe of the CaMLD interact with the JD, leading to a substantial conformational change that releases the active site [93].
The Arabidopsis genome contains 34 CDPK genes [94], the tomato genome contains 29 CDPK genes [95], and the maize genome contains 40 CDPK genes [96]. The encoded proteins play important roles in plant growth and development, as well as in environmental stress responses (e.g., HS). In Arabidopsis, inactivation of the autoinhibitory domain of AtCPK1 led to increased tolerance to salt, cold, and heat in AtCPK1-transformed Rubia cordifolia L. cell cultures, implicating AtCPK1 in the HS response [97]. In tomato, LeCPK28 was found to phosphorylate ascorbate peroxidase (APX), enhancing plant thermotolerance [98]. Mutants lacking LeCPK28 exhibited decreased thermotolerance, increased HS-induced accumulation of reactive ROS, elevated protein oxidation levels, and reduced activity of antioxidant enzymes such as APX. Additionally, the tomato CPK gene LeCPK2, which is predominantly expressed in flowers, exhibited high-level expression at 42°C [99]. In maize, ZmCDPK7 plays a role in HS tolerance by interacting with and phosphorylating sHSP17.4 at Ser-44, thereby upregulating its expression. ZmCDPK7 can translocate from the plasma membrane to the cytosol under HS conditions, and it interacts with the respiratory burst oxidase homolog RBOHB, phosphorylating it at Ser-99 [100]. Another maize CDPK gene, ZmCK3, exhibited increased transcription in response to drought, salt, and HS in maize seedlings [101]. Overexpressing ZmCK3 in Arabidopsis improved plant survival under conditions of drought and HS. In foxtail millet, SiCDPK7 was shown to be responsive to extreme temperature stress [102]. SiCDPK7 overexpression enhanced HS tolerance in both Arabidopsis and foxtail millet, with increased transcription of heat and cold stress-responsive genes observed under stressful conditions. In grapevine, the CDPK gene VaCPK29 was shown to be involved in responses to both heat and osmotic stress [103]. VaCPK29 overexpression in Vitis amurensis callus cell cultures and A. thaliana plants increased their tolerance to heat and high mannitol stress, indicating that VaCPK29 positively regulates the responses of grapevine plants to these stressors.
The CDPK family is diverse, with multiple members and isoforms across plant species. Recent whole-genome expression analyses have shed light on the transcriptional regulation of CDPKs in response to various stresses, including HS, in important crop species [95,104,105,106,107,108]. These findings provide valuable insight into the functional roles of CDPKs and their potential utility in enhancing the thermotolerance of crops.
3.4. CBLs and CIPKs in HS signaling
CBLs comprise a distinct family of Ca2+ sensors in plants, with essential roles in Ca2+ signaling pathways. Specifically, they interact with and modulate the activity of CIPKs. CBLs bear resemblance to the calcineurin B-subunit in yeast and neuronal Ca2+ sensors in animals [109]. CBLs possess four EF hands, which are Ca2+-binding motifs found in various proteins. EF hands are structural domains characterized by a D-X-D structure, with the loop region serving as the Ca2+-binding site. These domains allow proteins to function as Ca2+ sensors that can respond to changing levels of intracellular Ca2+. In the case of CBLs, the four EF-hand domains facilitate the capture of Ca2+ ions, enabling them to play pivotal roles in numerous plant physiological processes [110]. Notably, CBLs lack intrinsic kinase activity. To transmit signals, they must form complexes with CIPKs, which are serine/threonine kinases found in plants. CIPKs possess a functional kinase domain but remain in an inactive state due to autoinhibition caused by an interaction between the kinase domain and regulatory domain. An inhibitory motif known as NAF/FISL blocks the active site in CIPKs, preventing substrate binding and subsequent phosphorylation. This autoinhibitory mechanism keeps CIPKs in an inactive state until they are activated by binding to a CBL. Once active, the CIPK can regulate downstream proteins. This activation process is crucial for proper functioning of the CBL–CIPK signaling pathway in plants [111]. When plants encounter stress, such as HS, the intracellular concentration of Ca2+ rises. This increase enables Ca2+ ions to bind to the EF-hand motifs in CBLs. This binding promotes the interaction of CBLs with the NAF/FISL element in CIPKs. Consequently, the CIPKs become active and can participate in stress response pathways by phosphorylating downstream proteins.
While significant progress has been made in understanding the physiological and biochemical functions of CBLs and CIPKs in plant signal transduction and abiotic stress responses, their roles in thermotolerance are less understood. However, some research indicates their involvement in heat resistance. For instance, in rice, the OsCBL8–OsCIPK17 module plays a critical role in conferring resistance to HS. OsCBL8 facilitates the targeting of OsNAC77 and OsJAMYB by OsCIPK17, leading to enhanced resistance to high temperatures and pathogens in rice [112]. AcCIPK5, a CIPK from pineapple, has been shown to confer salt, osmotic stress, and cold tolerance while negatively regulating the HS response in transgenic Arabidopsis plants [113].
The CBL–CIPK network is considered a vital regulatory mechanism that deciphers Ca2+ signals triggered by HS. Further research will shed more light on the roles of CBLs and CIPKs in regulating plant responses to HS.
4. Ca2+ signaling networks mediate plant HS responses
Ca2+ ions are ubiquitous second messengers in eukaryotes, participating in a wide array of signaling pathways and responses to various environmental conditions. As mentioned above, when plants experience HS the cytosolic Ca2+ concentration rises due to an influx of Ca2+ facilitated by membrane-localized Ca2+ permeable cation channels. These ions subsequently bind to Ca2+-binding proteins, initiating signal transmission to their respective downstream pathways. In recent years, substantial progress has been made in understanding the mechanisms related to thermotolerance in plants. These mechanisms include the production of reactive ROS, an increase in NO, activation of mitogen-activated protein kinase (MAPK) signaling, heightened activity of HS factor (HSF) transcription factors, and augmented expression of HSPs. In the following section, we consider several potential downstream pathways of Ca2+ in response to HS in plants (Figure 2).
Table 2.
Ca2+-binding proteins that are likely involved in plant heat responses.
| Gene type | Organism | Gene symbol | Function | References |
|---|---|---|---|---|
| CaMs | Arabidopsis thaliana | AtCaM3 | Tolerance to heat stress | [84,85] |
| Cucumis sativus L. | CsCaM3 | Tolerance to heat stress; Safeguards against oxidative damage | [86] | |
| Oryza sativa L. | OsCaM1-1 | Tolerance to heat stress | [87] | |
| CMLs | Arabidopsis thaliana | AtCML12 | Gene expression significantly increased under heat stress | [89] |
| AtCML24 | Gene expression significantly increased under heat stress | [89] | ||
| Oryza sativa L. | OsMSR2 | Response to cold, drought, and heat stress | [90] | |
| Solanum lycopersicum L. | SlCML39 | Negative impact on high-temperature tolerance | [91] | |
| CDPKs | Arabidopsis thaliana | AtCPK1 | Tolerance to salt, cold, and heat | [97] |
| Lycopersicon esculentum Mill. | LeCPK28 | Tolerance to heat stress | [98] | |
| Lycopersicon esculentum Mill. | LeCPK2 | Tolerance to heat stress | [99] | |
| Zea mays L. | ZmCPK7 | Tolerance to heat stress | [100] | |
| ZmCK3 | Exhibits increased transcription in response to drought, salt, and heat stress | [101] | ||
| Setaria italica | SiCDPK7 | Response to extreme temperature stress | [102] | |
| Vitis amurensis Rupr. | VaCPK29 | Response to heat and osmotic stress | [103] | |
| CBLs | Oryza sativa L. | OsCBL8 | Enhances resistance to high temperatures and pathogens | [112] |
| CIPKs | Oryza sativa L. | OsCIPK17 | Enhances resistance to high temperatures and pathogens | [112] |
| Ananas comosus | AcCIPK5 | Promotes tolerance to salt, osmotic stress, and cold stress while negatively regulating heat stress responses | [113] |
4.1. ROS-mediated signaling
In response to HS, plants rapidly accumulate ROS, which are crucial signaling molecules in various stress responses [114]. One particularly potent ROS, H2O2, is generated in response to the presence of Ca2+ ions. H2O2 is produced during processes such as photorespiration, mitochondrial electron transport, and the beta-oxidation of fatty acids in plants. To maintain cellular homeostasis, the intracellular level of H2O2 is meticulously regulated through the action of enzymes like NADPH oxidase, also known as RBOH [115]. Among these enzymes, RBOHD, an ROS-generating NADPH oxidase located at the plasma membrane, plays a pivotal role in H2O2 production. Its activity is directly linked to an increase in cytosolic Ca2+ and/or CDPK phosphorylation [116]. Notably, RBOH phosphorylation initiates a positive feedback loop that further enhances both Ca2+ and ROS signaling [117]. NADPH oxidase activation results in augmented ROS production in the apoplastic space. These ROS are subsequently transported into the cell through aquaporins, which regulate cellular responses to HS [118].
ROS can trigger downstream signaling pathways associated with the HS response in cells through the involvement of multiprotein bridging factor 1 (MBF1), specific HSFs, and MAPKs. This cascade of events can alter a cell’s redox state, primarily through ROS accumulation. MBF1 family proteins operate as transcription co-factors, bridging the gap between transcription factors and the essential transcription machinery. In the context of plants, MBF1 proteins are integral players in abiotic stress responses, particularly in the context of HS [119]. One study found that the accumulation of MBF1c, a member of the MBF1 family, exhibited delayed kinetics in the ROS-producing rbohd mutant but accelerated kinetics in the ROS-scavenging apx1 mutant. This observation underscores the intricate interplay between ROS production and ROS scavenging and highlights the role of MBF1 proteins in modulating the ROS wave in response to HS [120]. Furthermore, MBF1c regulates the expression of more than 30 HS-related transcripts, including HSFB2a, HSFB2b, and DREB2A, which function upstream of HSFA3 [121]. Under HS conditions, members of the HS transcription factor family, including HSFA2, HSFA4a, and HSFA8, translocate rapidly from the cytosol to the nucleus. Importantly, this translocation is redox state-dependent and is mediated by the formation of an intramolecular disulfide bond within these transcription factors. Notably, the formation of this disulfide bond is reversible and is tightly regulated by the cellular redox state [122,123]. These findings reveal a novel mechanism through which plants can swiftly respond to temperature fluctuations by modulating the subcellular localization of key transcription factors.
ROS can also activate the MAPK phosphorylation cascade. In a study conducted in Arabidopsis leaf cells, H2O2 was shown to be a potent activator of MAPKs [124]. Specifically, H2O2 was able to activate a specific Arabidopsis MAPK kinase kinase, ANP1. This activation set in motion a phosphorylation cascade involving two stress-responsive MAPKs; namely, AtMPK3 and AtMPK6 [124]. Researchers were able to enhance the ability of tobacco plants to tolerate HS by overexpressing a constitutively active version of the ANP1 homolog NPK1 [124]. This manipulation suggests that the activation of ANP1 and subsequent activation of MAPK3 and MAPK6 can bolster plant thermotolerance. Furthermore, activated MAPK3 and MAPK6 play crucial roles in enhancing HSP expression. This occurs via the phosphorylation of HSFA2 and HSFA4a [125,126], demonstrating the central regulatory role of MAPK signaling in both the HS response and the expression of genes crucial for cellular protection against heat-induced damage.
4.2. NO signaling
In addition to ROS, another significant free radical produced in plant cells in response to HS is NO. H2O2 functions upstream of NO in the HS pathway in Arabidopsis seedlings. Following HS exposure, the NO levels in seedlings lacking such specific ROS-producing enzymes as atrbohB, atrbohD, and atrbohB/D were found to be lower than those in wild-type seedlings. To mitigate their heat sensitivity, these deficient seedlings were treated with compounds like sodium nitroprusside or S-nitroso-N-acetylpenicillamine, which partially rescued their thermotolerance [127]. This observation highlights the interplay between H2O2 and NO in regulating plant HS responses. Moreover, H2O2-induced NO was shown to stimulate the activity of antioxidant enzymes, allowing plants to counterbalance excessive H2O2 levels. This inhibited the DNA-binding activity of HSFs and the accumulation of HSPs. This points to a feedback loop between NO and H2O2 that regulates thermotolerance [128]. Additionally, a study found that Arabidopsis CaM3 inhibited NO accumulation and enhanced thermotolerance by directly promoting S-nitrosoglutathione reductase activity. This discovery suggests that feedback inhibition occurs between CaM3 and NO in the context of thermotolerance [129]. These findings highlight the intricate regulatory network that controls the response of plants to HS with the help of ROS and NO.
The primary physiological effect of NO is protein S-nitrosylation, a redox-based post-translational modification. This modification involves the covalent attachment of a NO molecule to a cysteine thiol on target proteins [130]. Numerous proteins that are integral to the HS response in plants undergo S-nitrosylation. This includes various categories of proteins, including HS signaling proteins (CDPK2, CDPK4, CDPK26, CaM, and UVR8), HSPs (HSP70, HSP90, HSP91, HSP88, and HSP60), and enzymes responsible for regulating cellular redox levels (CAT, APX, monodehydroascorbate reductase, SOD, glutathione peroxidase, glutaredoxin, and glutathione S-transferase) [3,130]. S-nitrosylation regulates various aspects of the modified proteins, including their expression levels, stability, subcellular localization, and enzymatic activity. In doing so, S-nitrosylation plays a pivotal role in fine-tuning and coordinating plant HS responses, ensuring the proper functioning of critical proteins involved in stress adaptation and survival.
4.3. HSF–HSP signaling
The synthesis of HSPs represents a crucial protective strategy that enables plants to effectively cope with HS. Within the promoter regions of HSP genes, specific sequences known as HS elements (HSEs; 5′-AGAAnnTTCT-3′) are present. These HSEs are recognized and bound by HSFs, which subsequently regulate the expression of HSP genes. Ca2+ has been implicated in increasing the DNA-binding activity of HSFs through direct interactions [131].
Studies have provided valuable insight into the role of Ca2+–CaM signaling in regulating thermotolerance in plants. Knockout mutants of AtCaM3 or its downstream targets, including AtCBK3 (a protein kinase) and AtPP7 (a protein phosphatase), exhibited decreased HSF activity and reduced HSP synthesis, which impaired the ability of the plants to tolerate high temperatures [84,132,133]. Notably, AtCBK3 and AtPP7 have been reported to interact with HSFs, further underscoring their involvement in the regulation of HS responses. Additionally, overexpression of OsCaM1-1, a rice Ca2+ sensor, in Arabidopsis has been demonstrated to enhance thermotolerance. This enhancement was associated with the elevated expression of HS-responsive genes, including AtCBK3, AtPP7, AtHSF, and AtHSP [87]. Furthermore, as mentioned earlier, HSFA2, HSFA4a, and HSFA8 activity is modulated by ROS induced by fluctuations in intracellular Ca2+ under HS conditions [122,123]. Notably, AtCaM3 has also been shown to increase thermotolerance via NO-mediated HSF activation and HSP accumulation [85]. Together, these findings highlight the intricate involvement of Ca2+–CaM signaling in the regulation of HSF activity and in the accumulation of HSPs, and they stress the significance of this pathway in enhancing the ability of plants to withstand HS.
5. Conclusions and perspectives
Rising temperatures represent an unusual environmental signal that can substantially reduce global crop yields and pose a significant threat to meeting the future demands of the world’s growing population [2,134]. Elevated temperatures can have a range of physiological, biochemical, and developmental consequences for crops [135]. In response to these challenges, plants have developed intricate signaling networks to detect temperature changes, with the rapid induction of Ca2+ signals being one of the earliest responses. This is initiated by an influx of Ca2+ into the cytoplasm through plasma membrane-situated Ca2+ channels. Subsequently, Ca2+ is sensed by Ca2+-binding proteins, which activate a signal transduction cascade.
Despite substantial progress in understanding the plant HS response over the past two decades, many questions remain unanswered. Future research should prioritize the following areas: (1) Integration of existing pathways. There is a need to integrate various fragmented pathways into a unified primary Ca2+ signaling pathway associated with HS. This endeavor could result in a more comprehensive understanding of the role of Ca2+ signaling in heat resistance. (2) Identification of new components. Ongoing efforts should focus on identifying new components that sense Ca2+ signals induced by HS. This continuing exploration will contribute to an improved understanding of the HS response pathway in plants. (3) Ca2+ crosstalk. Understanding the crosstalk between Ca2+-mediated HS responses and other stress signaling pathways is essential. This knowledge can shed light on the broader regulatory network that governs plant responses to multiple stressors.
Ultimately, the transformation of fundamental knowledge into practical applications is of paramount importance. Marker-assisted selection, next-generation molecular breeding, precision breeding, and genome editing techniques represent powerful tools that can enhance the efficiency of plant breeding programs. These techniques will enable breeders to more accurately and swiftly select plants with desirable traits, thereby addressing the challenges posed by rising temperatures and improving crop resilience in the face of environmental stress.
Author Contributions
L.Z. and X.L. developed the concept and drafted the manuscript; X.L. helped write the manuscript; X.K. searched for references and helped prepare the manuscript; L.Z. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Hebei Natural Science Foundations (C2021205013 and C2023205049 to Liqun Zhao, and C2023205031 to Xiaotong Liu).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fitter, A.H.; Fitter, R.S.R. Rapid changes in flowering time in British plants. Science 2002, 296, 1689–1691. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; Durand, J.L.; Elliott, J.; Ewert, F.; Janssens, I.A.; Li, T.; Lin, E.; Liu, Q.; Martre, P.; Müller, C.; Peng, S.; Peñuelas, J.; Ruane, A.C.; Wallach, D.; Wang, T.; Wu, D.; Liu, Z.; Zhu, Y.; Zhu, Z.; Asseng, S. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. 2017, 114, 9326–9331. [Google Scholar] [CrossRef]
- Li, B.; Gao, K.; Ren, H.; Tang, W. Molecular mechanisms governing plant responses to high temperatures. J. Integr. Plant Biol. 2018, 60, 757–779. [Google Scholar] [CrossRef]
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef]
- Finka, A.; Cuendet, A.F.; Maathuis, F.J.; Saidi, Y.; Goloubinoff, P. Plasma membrane cyclic nucleotide gated calcium channels control land plant thermal sensing and acquired thermotolerance. Plant Cell 2012, 24, 3333–3348. [Google Scholar] [CrossRef]
- Kudla, J.; Batistic, O.; Hashimoto, K. Calcium signals: the lead currency of plant information processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Batistič, O.; Kudla, J. Analysis of calcium signaling pathways in plants. Biochim. Biophys. Acta. 2012, 1820, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Pirayesh, N.; Giridhar, M.; Ben Khedher, A.; Vothknecht, U.C.; Chigri, F. Organellar calcium signaling in plants: An update. Biochim. Biophys Acta. Mol. Cell Res. 2021, 1868, 118948. [Google Scholar] [CrossRef] [PubMed]
- Swarbreck, S.M.; Colaço, R.; Davies, J.M. Plant calcium-permeable channels. Plant Physiol. 2013, 163, 514–522. [Google Scholar] [CrossRef]
- Jammes, F.; Hu, H.C.; Villiers, F.; Bouten, R.; Kwak, J.M. Calcium-permeable channels in plant cells. Febs j. 2011, 278, 4262–4276. [Google Scholar] [CrossRef]
- Kaplan, B.; Sherman, T.; Fromm, H. Cyclic nucleotide-gated channels in plants. FEBS Lett 2007, 581, 2237–2246. [Google Scholar] [CrossRef] [PubMed]
- DeFalco, T.A.; Marshall, C.B.; Munro, K.; Kang, H.G.; Moeder, W.; Ikura, M.; Snedden, W.A.; Yoshioka, K. Multiple calmodulin-binding sites positively and negatively regulate Arabidopsis cyclic nucleotide-gated channel 12. Plant Cell 2016, 28, 1738–1751. [Google Scholar] [PubMed]
- Fischer, C.; Kugler, A.; Hoth, S.; Dietrich, P. An IQ domain mediates the interaction with calmodulin in a plant cyclic nucleotide-gated channel. Plant Cell Physiol. 2013, 54, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Schuurink, R.C.; Shartzer, S.F.; Fath, A.; Jones, R.L. Characterization of a calmodulin-binding transporter from the plasma membrane of barley aleurone. Proc. Natl. Acad. Sci. 1998, 95, 1944–1949. [Google Scholar] [CrossRef]
- Mäser, P.; Thomine, S.; Schroeder, J.I.; Ward, J.M.; Hirschi, K.; Sze, H.; Talke, I.N.; Amtmann, A.; Maathuis, F.J.; Sanders, D.; Harper, J.F.; Tchieu, J.; Gribskov, M.; Persans, M.W.; Salt, D.E.; Kim, S.A.; Guerinot, M.L. Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef]
- Bridges, D.; Fraser, M.E.; Moorhead, G.B. Cyclic nucleotide binding proteins in the Arabidopsis thaliana and Oryza sativa genomes. BMC Bioinform. 2005, 6, 6. [Google Scholar] [CrossRef]
- Zhou, L.; Lan, W.; Jiang, Y.; Fang, W.; Luan, S. A calcium-dependent protein kinase interacts with and activates a calcium channel to regulate pollen tube growth. Mol. Plant 2014, 7, 369–376. [Google Scholar] [CrossRef]
- Zhang, S.; Pan, Y.; Tian, W.; Dong, M.; Zhu, H.; Luan, S.; Li, L. Arabidopsis CNGC14 mediates calcium influx required for tip growth in root hairs. Mol. Plant 2017, 10, 1004–1006. [Google Scholar] [CrossRef] [PubMed]
- Chiasson, D.M.; Haage, K.; Sollweck, K.; Brachmann, A.; Dietrich, P.; Parniske, M. A quantitative hypermorphic CNGC allele confers ectopic calcium flux and impairs cellular development. Elife 2017, 6. [Google Scholar]
- Wang, Y.; Kang, Y.; Ma, C.; Miao, R.; Wu, C.; Long, Y.; Ge, T.; Wu, Z.; Hou, X.; Zhang, J.; Qi, Z. CNGC2 Is a Ca2+ Influx Channel That Prevents Accumulation of Apoplastic Ca2+ in the Leaf. Plant Physiol. 2017, 173, 1342–1354. [Google Scholar] [CrossRef]
- Katano, K.; Kataoka, R.; Fujii, M.; Suzuki, N. Differences between seedlings and flowers in anti-ROS based heat responses of Arabidopsis plants deficient in cyclic nucleotide gated channel 2. Plant Physiol. Biochem. 2018, 123, 288–296. [Google Scholar] [CrossRef]
- Thoen, M.P.; Davila Olivas, N.H.; Kloth, K.J.; Coolen, S.; Huang, P.P.; Aarts, M.G.; Bac-Molenaar, J.A.; Bakker, J.; Bouwmeester, H.J.; Broekgaarden, C.; Bucher, J.; Busscher-Lange, J.; Cheng, X.; Fradin, E.F.; Jongsma, M.A.; Julkowska, M.M.; Keurentjes, J.J.; Ligterink, W.; Pieterse, C.M.; Ruyter-Spira, C.; Smant, G.; Testerink, C.; Usadel, B.; van Loon, J.J.; van Pelt, J.A.; van Schaik, C.C.; van Wees, S.C.; Visser, R.G.; Voorrips, R.; Vosman, B.; Vreugdenhil, D.; Warmerdam, S.; Wiegers, G.L.; van Heerwaarden, J.; Kruijer, W.; van Eeuwijk, F.A.; Dicke, M. Genetic architecture of plant stress resistance: multi-trait genome-wide association mapping. New Phytol. 2017, 213, 1346–1362. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Han, X.W.; Wu, J.H.; Zheng, S.Z.; Shang, Z.L.; Sun, D.Y.; Zhou, R.G.; Li, B. A heat-activated calcium-permeable channel - Arabidopsis cyclic nucleotide-gated ion channel 6 - is involved in heat shock responses. Plant J. 2012, 70, 1056–1069. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, J.; Ai, L.; Wu, D.; Li, B.; Zhang, L.; Zhao, L. Cyclic nucleotide-gated ion channel 6 mediates thermotolerance in Arabidopsis seedlings by regulating hydrogen peroxide production via cytosolic calcium ions. Front. Plant Sci. 2021, 12, 708672. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, X.; Li, B.; Zhao, L. Cyclic nucleotide-gated ion channel 6 mediates thermotolerance in Arabidopsis seedlings by regulating nitric oxide production via cytosolic calcium ions. BMC Plant Biol. 2019, 19, 368. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Lu, S.; Li, Z.; Cheng, J.; Hu, P.; Zhu, T.; Wang, X.; Jin, M.; Wang, X.; Li, L.; Huang, S.; Zou, B.; Hua, J. Cyclic nucleotide-gated ion channels 14 and 16 promote tolerance to heat and chilling in rice. Plant Physiol. 2020, 183, 1794–1808. [Google Scholar] [CrossRef]
- Nawaz, Z.; Kakar, K.U.; Saand, M.A.; Shu, Q.Y. Cyclic nucleotide-gated ion channel gene family in rice, identification, characterization and experimental analysis of expression response to plant hormones, biotic and abiotic stresses. BMC Genom. 2014, 15, 853. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.L. Structural biology of glutamate receptor ion channel complexes. Curr. Opin. Struct. Biol. 2016, 41, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.M.; Chiu, J.; Hsieh, M.H.; Meisel, L.; Oliveira, I.C.; Shin, M.; Coruzzi, G. Glutamate-receptor genes in plants. Nature 1998, 396, 125–126. [Google Scholar] [CrossRef]
- Lacombe, B.; Becker, D.; Hedrich, R.; DeSalle, R.; Hollmann, M.; Kwak, J.M.; Schroeder, J.I.; Le Novère, N.; Nam, H.G.; Spalding, E.P.; Tester, M.; Turano, F.J.; Chiu, J.; Coruzzi, G. The identity of plant glutamate receptors. Science 2001, 292, 1486–1487. [Google Scholar] [CrossRef]
- Li, J.; Zhu, S.; Song, X.; Shen, Y.; Chen, H.; Yu, J.; Yi, K.; Liu, Y.; Karplus, V.J.; Wu, P.; Deng, X.W. A rice glutamate receptor-like gene is critical for the division and survival of individual cells in the root apical meristem. Plant Cell 2006, 18, 340–349. [Google Scholar] [CrossRef]
- Dietrich, P.; Anschütz, U.; Kugler, A.; Becker, D. Physiology and biophysics of plant ligand-gated ion channels. Plant Biol. 2010, 12 (Suppl. 1), 80–93. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.C.; Mukherjee, A. Computational analysis of the glutamate receptor gene family of Arabidopsis thaliana. J. Biomol. Struct. Dyn. 2017, 35, 2454–2474. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.M.; Mäser, P.; Schroeder, J.I. Plant ion channels: gene families, physiology, and functional genomics analyses. Annu. Rev. Physiol. 2009, 71, 59–82. [Google Scholar] [CrossRef] [PubMed]
- Davenport, R. Glutamate receptors in plants. Ann. Bot. 2002, 90, 549–557. [Google Scholar] [CrossRef]
- Salvador-Recatalà, V. New roles for the glutamate receptor-like 3.3, 3.5, and 3.6 genes as on/off switches of wound-induced systemic electrical signals. Plant Signal Behav. 2016, 11, e1161879. [Google Scholar] [CrossRef]
- Green, M.N.; Gangwar, S.P.; Michard, E.; Simon, A.A.; Portes, M.T.; Barbosa-Caro, J.; Wudick, M.M.; Lizzio, M.A.; Klykov, O.; Yelshanskaya, M.V.; Feijó, J.A.; Sobolevsky, A.I. Structure of the Arabidopsis thaliana glutamate receptor-like channel GLR3.4. Mol. Cell 2021, 81, 3216–3226. [Google Scholar] [CrossRef]
- Kong, D.; Hu, H.C.; Okuma, E.; Lee, Y.; Lee, H.S.; Munemasa, S.; Cho, D.; Ju, C.; Pedoeim, L.; Rodriguez, B.; Wang, J.; Im, W.; Murata, Y.; Pei, Z.M.; Kwak, J.M. L-Met activates Arabidopsis GLR Ca2+ channels upstream of ROS production and regulates stomatal movement. Cell Rep. 2016, 17, 2553–2561. [Google Scholar] [CrossRef]
- Xue, N.; Zhan, C.; Song, J.; Li, Y.; Zhang, J.; Qi, J.; Wu, J. The glutamate receptor-like 3.3 and 3.6 mediate systemic resistance to insect herbivores in Arabidopsis. J. Exp. Bot. 2022, 73, 7611–7627. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Fichman, Y.; Mittler, R. Vascular bundles mediate systemic reactive oxygen signaling during light stress. Plant Cell 2020, 32, 3425–3435. [Google Scholar] [CrossRef]
- Manzoor, H.; Kelloniemi, J.; Chiltz, A.; Wendehenne, D.; Pugin, A.; Poinssot, B.; Garcia-Brugger, A. Involvement of the glutamate receptor AtGLR3.3 in plant defense signaling and resistance to Hyaloperonospora arabidopsidis. Plant J. 2013, 76, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, R.; Mori, I.C.; Kamizono, N.; Shichiri, Y.; Shimatani, T.; Miyata, F.; Honda, K.; Iwai, S. Glutamate functions in stomatal closure in Arabidopsis and fava bean. J. Plant Res. 2016, 129, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Duan, H.; Liu, Y.; Sun, X.; Zhao, J.; Lin, H. Patellin protein family functions in plant development and stress response. J. Plant Physiol. 2019, 234–235, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.G.; Ye, X.Y.; Qiu, X.M. Glutamate signaling enhances the heat tolerance of maize seedlings by plant glutamate receptor-like channels-mediated calcium signaling. Protoplasma 2019, 256, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, J.C.; Laohavisit, A.; Macpherson, N.; Webb, A.; Brownlee, C.; Battey, N.H.; Davies, J.M. Annexins: multifunctional components of growth and adaptation. J. Exp. Bot. 2008, 59, 533–544. [Google Scholar] [CrossRef]
- Davies, J.M. Annexin-mediated calcium signalling in plants. Plants 2014, 3, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Gorecka, K.M.; Konopka-Postupolska, D.; Hennig, J.; Buchet, R.; Pikula, S. Peroxidase activity of annexin 1 from Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2005, 336, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Gerke, V.; Creutz, C.E.; Moss, S.E. Annexins: linking Ca2+ signalling to membrane dynamics. Nat. Rev. Mol. Cell Biol. 2005, 6, 449–461. [Google Scholar] [CrossRef]
- Clark, G.B.; Sessions, A.; Eastburn, D.J.; Roux, S.J. Differential expression of members of the annexin multigene family in Arabidopsis. Plant Physiol. 2001, 126, 1072–1084. [Google Scholar] [CrossRef]
- Jami, S.K.; Clark, G.B.; Ayele, B.T.; Roux, S.J.; Kirti, P.B. Identification and characterization of annexin gene family in rice. Plant Cell Rep. 2012, 31, 813–825. [Google Scholar] [CrossRef]
- Xu, L.; Tang, Y.; Gao, S.; Su, S.; Hong, L.; Wang, W.; Fang, Z.; Li, X.; Ma, J.; Quan, W.; Sun, H.; Li, X.; Wang, Y.; Liao, X.; Gao, J.; Zhang, F.; Li, L.; Zhao, C. Comprehensive analyses of the annexin gene family in wheat. BMC Genom. 2016, 17, 415. [Google Scholar] [CrossRef]
- Gerke, V.; Moss, S.E. Annexins: from structure to function. Physiol. Rev. 2002, 82, 331–371. [Google Scholar] [CrossRef]
- Saad, R.B.; Ben Romdhane, W.; Ben Hsouna, A.; Mihoubi, W.; Harbaoui, M.; Brini, F. Insights into plant annexins function in abiotic and biotic stress tolerance. Plant Signal Behav. 2020, 15, 1699264. [Google Scholar] [CrossRef]
- Laohavisit, A.; Davies, J.M. Annexins. New Phytol. 2011, 189, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, Y.; Bian, Y.; Ren, Y.; Xu, X.; Zhou, F.; Ding, H. A critical review on plant annexin: Structure, function, and mechanism. Plant Physiol. Biochem. 2022, 190, 81–89. [Google Scholar] [CrossRef]
- Chu, P.; Chen, H.; Zhou, Y.; Li, Y.; Ding, Y.; Jiang, L.; Tsang, E.W.; Wu, K.; Huang, S. Proteomic and functional analyses of Nelumbo nucifera annexins involved in seed thermotolerance and germination vigor. Planta 2012, 235, 1271–1288. [Google Scholar] [CrossRef]
- Wei, J.; Shen, Y.; Zhao, H.; Liu, X.; Jia, Y.; Yu, X.; Ma, H. GmANN, a glutathione S-transferase-interacting annexin, is involved in high temperature and humidity tolerance and seed vigor formation in transgenic Arabidopsis. Plant Cell 2019, 138, 583–595. [Google Scholar] [CrossRef]
- Qiao, B.; Zhang, Q.; Liu, D.; Wang, H.; Yin, J.; Wang, R.; He, M.; Cui, M.; Shang, Z.; Wang, D.; Zhu, Z. A calcium-binding protein, rice annexin OsANN1, enhances heat stress tolerance by modulating the production of H2O2. J. Exp. Bot. 2015, 66, 5853–5866. [Google Scholar] [CrossRef]
- Shen, F.; Ying, J.; Xu, L.; Sun, X.; Wang, J.; Wang, Y.; Mei, Y.; Zhu, Y.; Liu, L. Characterization of annexin gene family and functional analysis of RsANN1a involved in heat tolerance in radish (Raphanus sativus L.). Physiol. Mol. Biol. Plants 2021, 27, 2027–2041. [Google Scholar] [CrossRef]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; Johnson, D.M.; Swift, G.B.; He, Y.; Siedow, J.N.; Pei, Z.M. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yuan, F.; Wen, Z.; Li, Y.; Wang, F.; Zhu, T.; Zhuo, W.; Jin, X.; Wang, Y.; Zhao, H.; Pei, Z.M.; Han, S. Genome-wide survey and expression analysis of the OSCA gene family in rice. BMC Plant Biol. 2015, 15, 261. [Google Scholar] [CrossRef]
- Ding, S.; Feng, X.; Du, H.; Wang, H. Genome-wide analysis of maize OSCA family members and their involvement in drought stress. PeerJ 2019, 7, e6765. [Google Scholar] [CrossRef]
- Yang, X.; Xu, Y.; Yang, F.; Magwanga, R.O.; Cai, X.; Wang, X.; Wang, Y.; Hou, Y.; Wang, K.; Liu, F.; Zhou, Z. Genome-wide identification of OSCA gene family and their potential function in the regulation of dehydration and salt stress in Gossypium hirsutum. J. Cotton Res. 2019, 2, 11. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Li, B.; Hou, L.; Yu, J.; Jia, C.; Wang, Z.; Chen, S.; Zhang, M.; Qin, J.; Cao, N.; Cui, J.; Shi, W. Preliminary expression analysis of the OSCA gene family in Maize and their involvement in temperature stress. Int. J. Mol. Sci. 2022, 23, 13658. [Google Scholar] [CrossRef]
- Haswell, E.S.; Meyerowitz, E.M. MscS-like proteins control plastid size and shape in Arabidopsis thaliana. Curr. Biol. 2006, 16, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, E.S.; Haswell, E.S. The tension-sensitive ion transport activity of MSL8 is critical for its function in pollen hydration and germination. Plant Cell Physiol. 2017, 58, 1222–1237. [Google Scholar] [CrossRef]
- Peyronnet, R.; Haswell, E.S.; Barbier-Brygoo, H.; Frachisse, J.M. AtMSL9 and AtMSL10: Sensors of plasma membrane tension in Arabidopsis roots. Plant Signal. Behav. 2008, 3, 726–729. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, M.; Sun, J.; Mao, X.; Wang, J.; Wang, J.; Liu, H.; Zheng, H.; Zhen, Z.; Zhao, H.; Zou, D. Genome-wide characterization and identification of trihelix transcription factor and expression profiling in response to abiotic stresses in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2019, 20, 251. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Katagiri, T.; Shinozaki, K.; Qi, Z.; Tatsumi, H.; Furuichi, T.; Kishigami, A.; Sokabe, M.; Kojima, I.; Sato, S.; Kato, T.; Tabata, S.; Iida, K.; Terashima, A.; Nakano, M.; Ikeda, M.; Yamanaka, T.; Iida, H. Arabidopsis plasma membrane protein crucial for Ca2+ influx and touch sensing in roots. Proc. Natl. Acad. Sci. 2007, 104, 3639–3644. [Google Scholar] [CrossRef] [PubMed]
- Kurusu, T.; Yamanaka, T.; Nakano, M.; Takiguchi, A.; Ogasawara, Y.; Hayashi, T.; Iida, K.; Hanamata, S.; Shinozaki, K.; Iida, H.; Kuchitsu, K. Involvement of the putative Ca²⁺-permeable mechanosensitive channels, NtMCA1 and NtMCA2, in Ca²⁺ uptake, Ca²⁺-dependent cell proliferation and mechanical stress-induced gene expression in tobacco (Nicotiana tabacum) BY-2 cells. J. Plant Res. 2012, 125, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, T.; Nakagawa, Y.; Mori, K.; Nakano, M.; Imamura, T.; Kataoka, H.; Terashima, A.; Iida, K.; Kojima, I.; Katagiri, T.; Shinozaki, K.; Iida, H. MCA1 and MCA2 that mediate Ca2+ uptake have distinct and overlapping roles in Arabidopsis. Plant Physiol. 2010, 152, 1284–1296. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Renhu, N.; Naito, M.; Nakamura, A.; Shiba, H.; Yamamoto, T.; Suzaki, T.; Iida, H.; Miura, K. Ca2+-permeable mechanosensitive channels MCA1 and MCA2 mediate cold-induced cytosolic Ca2+ increase and cold tolerance in Arabidopsis. Sci. Rep. 2018, 8, 550. [Google Scholar] [CrossRef] [PubMed]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Kurusu, T.; Kuchitsu, K.; Nakano, M.; Nakayama, Y.; Iida, H. Plant mechanosensing and Ca2+ transport. Trends Plant Sci. 2013, 18, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tong, X.; Liu, S.Y.; Chai, L.X.; Zhu, F.F.; Zhang, X.P.; Zou, J.Z.; Wang, X.B. Genetic analysis of a Piezo-like protein suppressing systemic movement of plant viruses in Arabidopsis thaliana. Sci. Rep. 2019, 9, 3187. [Google Scholar] [CrossRef] [PubMed]
- Knight, H. Calcium signaling during abiotic stress in plants. Int. Rev. Cytol. 2000, 195, 269–324. [Google Scholar] [PubMed]
- Mohanta, T.K.; Yadav, D.; Khan, A.L.; Hashem, A.; Abd Allah, E.F.; Al-Harrasi, A. Molecular players of EF-hand containing calcium signaling event in plants. Int. J. Mol. Sci. 2019, 20, 1476. [Google Scholar] [CrossRef]
- Braam, J.; Davis, R.W. Rain-, wind-, and touch-induced expression of calmodulin and calmodulin-related genes in Arabidopsis. Cell 1990, 60, 357–364. [Google Scholar] [CrossRef]
- Al-Quraan, N.A.; Locy, R.D.; Singh, N.K. Expression of calmodulin genes in wild type and calmodulin mutants of Arabidopsis thaliana under heat stress. Plant Physiol. Biochem. 2010, 48, 697–702. [Google Scholar] [CrossRef]
- Choi, M.S.; Kim, M.C.; Yoo, J.H.; Moon, B.C.; Koo, S.C.; Park, B.O.; Lee, J.H.; Koo, Y.D.; Han, H.J.; Lee, S.Y.; Chung, W.S.; Lim, C.O.; Cho, M.J. Isolation of a calmodulin-binding transcription factor from rice (Oryza sativa L.). J. Biol. Chem. 2005, 280, 40820–40831. [Google Scholar] [CrossRef]
- Hardoim, P.R.; de Carvalho, T.L.; Ballesteros, H.G.; Bellieny-Rabelo, D.; Rojas, C.A.; Venancio, T.M.; Ferreira, P.C.; Hemerly, A.S. Genome-wide transcriptome profiling provides insights into the responses of maize (Zea mays L.) to diazotrophic bacteria. Plant and Soil. 2020, 451, 121–143. [Google Scholar] [CrossRef]
- Gifford, J.L.; Walsh, M.P.; Vogel, H.J. Structures and metal-ion-binding properties of the Ca2+-binding helix-loop-helix EF-hand motifs. Biochem. J. 2007, 405, 199–221. [Google Scholar] [CrossRef]
- Kundu, P.; Nehra, A.; Gill, R.; Tuteja, N.; Gill, S.S. Unraveling the importance of EF-hand-mediated calcium signaling in plants. S. AFR. J. BOT. 2022, 148, 615–633. [Google Scholar] [CrossRef]
- Zhang, W.; Zhou, R.G.; Gao, Y.J.; Zheng, S.Z.; Xu, P.; Zhang, S.Q.; Sun, D.Y. Molecular and genetic evidence for the key role of AtCaM3 in heat-shock signal transduction in Arabidopsis. Plant Physiol. 2009, 149, 1773–1784. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Y.; Zhou, S.; Wang, L.; Cheng, Y.; Zhao, L. Nitric oxide functions as a signal and acts upstream of AtCaM3 in thermotolerance in Arabidopsis seedlings. Plant Physiol. 2010, 153, 1895–1906. [Google Scholar] [CrossRef]
- Yu, B.; Yan, S.; Zhou, H.; Dong, R.; Lei, J.; Chen, C.; Cao, B. Overexpression of CsCaM3 improves high temperature tolerance in cucumber. Front. Plant Sci. 2018, 9, 797. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Luo, D.L.; Vignols, F.; Jinn, T.L. Heat shock-induced biphasic Ca2+ signature and OsCaM1-1 nuclear localization mediate downstream signalling in acquisition of thermotolerance in rice (Oryza sativa L.). Plant Cell Environ. 2012, 35, 1543–1557. [Google Scholar] [CrossRef] [PubMed]
- McCormack, E.; Tsai, Y.C.; Braam, J. Handling calcium signaling: Arabidopsis CaMs and CMLs. Trends Plant Sci. 2005, 10, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Braam, J. Regulated expression of the calmodulin-related TCH genes in cultured Arabidopsis cells: induction by calcium and heat shock. Proc. Natl. Acad. Sci. 1992, 89, 3213–3216. [Google Scholar] [CrossRef]
- Xu, G.Y.; Rocha, P.S.; Wang, M.L.; Xu, M.L.; Cui, Y.C.; Li, L.Y.; Zhu, Y.X.; Xia, X. A novel rice calmodulin-like gene, OsMSR2, enhances drought and salt tolerance and increases ABA sensitivity in Arabidopsis. Planta 2011, 234, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Qian, Y.; Fang, Y.; Ji, Y.; Sheng, J.; Ge, C. Characteristics of SiCML39, a tomato calmodulin-like gene, and its negative role in high temperature tolerance of Arabidopsis thaliana during germination and seedling growth. Int. J. Mol. Sci. 2021, 22, 11479. [Google Scholar] [CrossRef]
- Atif, R.M.; Shahid, L.; Waqas, M.; Ali, B.; Rashid, M.A.R.; Azeem, F.; Nawaz, M.A.; Wani, S.H.; Chung, G. Insights on calcium-dependent protein kinases (CPKs) signaling for abiotic stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 5298. [Google Scholar] [CrossRef]
- Yip Delormel, T.; Boudsocq, M. Properties and functions of calcium-dependent protein kinases and their relatives in Arabidopsis thaliana. New Phytol. 2019, 224, 585–604. [Google Scholar] [CrossRef]
- Cheng, S.H.; Willmann, M.R.; Chen, H.C.; Sheen, J. Calcium signaling through protein kinases. The Arabidopsis calcium-dependent protein kinase gene family. Plant Physiol. 2002, 129, 469–485. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Lv, X.; Xia, X.; Zhou, J.; Shi, K.; Yu, J.; Zhou, Y. Genome-wide identification and expression analysis of calcium-dependent protein kinase in tomato. Front. Plant Sci. 2016, 7, 469. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Lv, W.; Jiang, S.; Zhang, D.; Cai, G.; Pan, J.; Li, D. Genome-wide identification and expression analysis of calcium-dependent protein kinase in maize. BMC Genom. 2013, 14, 433. [Google Scholar] [CrossRef] [PubMed]
- Veremeichik, G.N.; Shkryl, Y.N.; Gorpenchenko, T.Y.; Silantieva, S.A.; Avramenko, T.V.; Brodovskaya, E.V.; Bulgakov, V.P. Inactivation of the auto-inhibitory domain in Arabidopsis AtCPK1 leads to increased salt, cold and heat tolerance in the AtCPK1-transformed Rubia cordifolia L cell cultures. Plant Physiol. Biochem. 2021, 159, 372–382. [Google Scholar] [CrossRef]
- Hu, Z.; Li, J.; Ding, S.; Cheng, F.; Li, X.; Jiang, Y.; Yu, J.; Foyer, C.H.; Shi, K. The protein kinase CPK28 phosphorylates ascorbate peroxidase and enhances thermotolerance in tomato. Plant Physiol. 2021, 186, 1302–1317. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.J.; Su, H.S.; Li, W.J.; Zhang, Z.L. Expression profiling of a novel calcium-dependent protein kinase gene, LeCPK2, from tomato (Solanum lycopersicum) under heat and pathogen-related hormones. Biosci. Biotechnol. Biochem. 2009, 73, 2427–2431. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Du, H.; Wang, Y.; Wang, H.; Yang, S.; Li, C.; Chen, N.; Yang, H.; Zhang, Y.; Zhu, Y.; Yang, L.; Hu, X. The calcium-dependent protein kinase ZmCDPK7 functions in heat-stress tolerance in maize. J. Integr. Plant Biol. 2021, 63, 510–527. [Google Scholar] [CrossRef]
- Wang, C.-T.; Song, W. ZmCK3, a maize calcium-dependent protein kinase gene, endows tolerance to drought and heat stresses in transgenic Arabidopsis. J. Plant Biochem. 2014, 23, 249–256. [Google Scholar] [CrossRef]
- Wei, J.-T.; Hou, Z.-H.; Wang, Y.; Hao, J.-M.; Wang, J.; Wang, W.; Wang, W.; Wang, D.-M.; Xu, Z.-S.; Song, X.; Wang, F.; Li, R. Foxtail millet SiCDPK7 gene enhances tolerance to extreme temperature stress in transgenic plants. Environ. Exp. Bot. 2023, 207, 105197. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V.; Khristenko, V.S.; Aleynova, O.A. The calcium-dependent protein kinase gene VaCPK29 is involved in grapevine responses to heat and osmotic stresses. Plant Growth Regula. 2017, 82, 79–89. [Google Scholar] [CrossRef]
- Zhang, K.; Han, Y.T.; Zhao, F.L.; Hu, Y.; Gao, Y.R.; Ma, Y.F.; Zheng, Y.; Wang, Y.J.; Wen, Y.Q. Genome-wide identification and expression analysis of the CDPK gene family in grape, vitis spp. BMC Plant Biol. 2015, 15, 164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, W.Z.; Zhang, Y.; Deng, M.; Niu, F.; Yang, B.; Wang, X.; Wang, B.; Liang, W.; Deyholos, M.K.; Jiang, Y.Q. Identification, expression and interaction analyses of calcium-dependent protein kinase (CPK) genes in canola (Brassica napus L.). BMC Genom. 2014, 15, 211. [Google Scholar]
- Wu, P.; Wang, W.; Duan, W.; Li, Y.; Hou, X. Comprehensive analysis of the CDPK-SnRK superfamily genes in chinese cabbage and its evolutionary implications in plants. Front. Plant Sci. 2017, 8, 162. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, M.; Lu, L.; He, M.; Qu, W.; Xu, Q.; Qi, X.; Chen, X. Genome-wide analysis and expression of the calcium-dependent protein kinase gene family in cucumber. Mol. Genet. Genom. 2015, 290, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Cheng, J.; Yan, Y.; Xiao, Z.; Li, J.; Mou, S.; Qiu, A.; Lai, Y.; Guan, D.; He, S. Genome-wide identification and expression analysis of calcium-dependent protein kinase and its closely related kinase genes in Capsicum annuum. Front. Plant Sci. 2015, 6, 737. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, J.K. A calcium sensor homolog required for plant salt tolerance. Science 1998, 280, 1943–1945. [Google Scholar] [CrossRef] [PubMed]
- Luan, S. The CBL-CIPK network in plant calcium signaling. Trends Plant Sci. 2009, 14, 37–42. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Mahiwal, S.; Nambiar, D.M.; Pandey, G.K. CBL-CIPK module-mediated phosphoregulation: facts and hypothesis. Biochem. J. 2020, 477, 853–871. [Google Scholar] [CrossRef]
- Gao, C.; Lu, S.; Zhou, R.; Wang, Z.; Li, Y.; Fang, H.; Wang, B.; Chen, M.; Cao, Y. The OsCBL8-OsCIPK17 module regulates seedling growth and confers resistance to heat and drought in rice. Int. J. Mol. Sci. 2022, 23, 12451. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Greaves, J.G.; Jakada, B.H.; Fakher, B.; Wang, X.; Qin, Y. AcCIPK5, a pineapple CBL-interacting protein kinase, confers salt, osmotic and cold stress tolerance in transgenic Arabidopsis. Plant Sci. 2022, 320, 111284. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; He, R.J.; Xie, Q.L.; Zhao, X.H.; Deng, X.M.; He, J.B.; Song, L.; He, J.; Marchant, A.; Chen, X.Y.; Wu, A.M. Ethylene response factor 74 (ERF74) plays an essential role in controlling a respiratory burst oxidase homolog D (RbohD)-dependent mechanism in response to different stresses in Arabidopsis. New Phytol. 2017, 213, 1667–1681. [Google Scholar] [CrossRef] [PubMed]
- Saxena, I.; Srikanth, S.; Chen, Z. Cross talk between H2O2 and interacting signal molecules under plant stress response. Front. Plant Sci. 2016, 7, 570. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, Y.; Kaya, H.; Hiraoka, G.; Yumoto, F.; Kimura, S.; Kadota, Y.; Hishinuma, H.; Senzaki, E.; Yamagoe, S.; Nagata, K.; Nara, M.; Suzuki, K.; Tanokura, M.; Kuchitsu, K. Synergistic activation of the Arabidopsis NADPH oxidase AtrbohD by Ca2+ and phosphorylation. J. Biol. Chem. 2008, 283, 8885–8892. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Kaya, H.; Kawarazaki, T.; Hiraoka, G.; Senzaki, E.; Michikawa, M.; Kuchitsu, K. Protein phosphorylation is a prerequisite for the Ca2+-dependent activation of Arabidopsis NADPH oxidases and may function as a trigger for the positive feedback regulation of Ca2+ and reactive oxygen species. Biochim. Biophys. Acta. 2012, 1823, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Møller, A.L.; Kristiansen, K.A.; Schulz, A.; Møller, I.M.; Schjoerring, J.K.; Jahn, T.P. Specific aquaporins facilitate the diffusion of hydrogen peroxide across membranes. J. Biol. Chem. 2007, 282, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Jaimes-Miranda, F.; Chávez Montes, R.A. The plant MBF1 protein family: a bridge between stress and transcription. J. Exp. Bot. 2020, 71, 1782–1791. [Google Scholar] [CrossRef]
- Suzuki, N.; Miller, G.; Salazar, C.; Mondal, H.A.; Shulaev, E.; Cortes, D.F.; Shuman, J.L.; Luo, X.; Shah, J.; Schlauch, K.; Shulaev, V.; Mittler, R. Temporal-spatial interaction between reactive oxygen species and abscisic acid regulates rapid systemic acclimation in plants. Plant Cell 2013, 25, 3553–3569. [Google Scholar] [CrossRef]
- Suzuki, N.; Sejima, H.; Tam, R.; Schlauch, K.; Mittler, R. Identification of the MBF1 heat-response regulon of Arabidopsis thaliana. Plant J. 2011, 66, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Giesguth, M.; Sahm, A.; Simon, S.; Dietz, K.J. Redox-dependent translocation of the heat shock transcription factor AtHSFA8 from the cytosol to the nucleus in Arabidopsis thaliana. FEBS Lett. 2015, 589, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Mittler, R. Could heat shock transcription factors function as hydrogen peroxide sensors in plants? Ann. Bot. 2006, 98, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Kovtun, Y.; Chiu, W.L.; Tena, G.; Sheen, J. Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants. Proc. Natl. Acad. Sci. 2000, 97, 2940–2945. [Google Scholar] [CrossRef] [PubMed]
- Evrard, A.; Kumar, M.; Lecourieux, D.; Lucks, J.; von Koskull-Döring, P.; Hirt, H. Regulation of the heat stress response in Arabidopsis by MPK6-targeted phosphorylation of the heat stress factor HsfA2. PeerJ 2013, 1, e59. [Google Scholar] [CrossRef]
- Pérez-Salamó, I.; Papdi, C.; Rigó, G.; Zsigmond, L.; Vilela, B.; Lumbreras, V.; Nagy, I.; Horváth, B.; Domoki, M.; Darula, Z.; Medzihradszky, K.; Bögre, L.; Koncz, C.; Szabados, L. The heat shock factor A4A confers salt tolerance and is regulated by oxidative stress and the mitogen-activated protein kinases MPK3 and MPK6. Plant Physiol. 2014, 165, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, Y.; Jia, L.; Chu, H.; Zhou, S.; Chen, K.; Wu, D.; Zhao, L. Hydrogen peroxide acts upstream of nitric oxide in the heat shock pathway in Arabidopsis seedlings. Plant Physiol. 2014, 164, 2184–2196. [Google Scholar] [CrossRef]
- Wu, D.; Chu, H.; Jia, L.; Chen, K.; Zhao, L. A feedback inhibition between nitric oxide and hydrogen peroxide in the heat shock pathway in Arabidopsis seedlings. Plant Growth Regul. 2015, 75, 503–509. [Google Scholar] [CrossRef]
- Jia, D.; Liu, Y.H.; Zhang, B.; Ji, Z.Y.; Wang, Y.X.; Gao, L.L.; Ma, R.Y. Induction of heat shock protein genes is the hallmark of egg heat tolerance in Agasicles hygrophila (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2020, 113, 1972–1981. [Google Scholar] [CrossRef]
- Hu, J.; Huang, X.; Chen, L.; Sun, X.; Lu, C.; Zhang, L.; Wang, Y.; Zuo, J. Site-specific nitrosoproteomic identification of endogenously S-nitrosylated proteins in Arabidopsis. Plant Physiol. 2015, 167, 1731–1746. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Brandizzi, F.; Benning, C.; Larkin, R.M. A membrane-tethered transcription factor defines a branch of the heat stress response in Arabidopsis thaliana. Proc. Natl. Acad. Sci. 2008, 105, 16398–16403. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.T.; Li, G.L.; Chang, H.; Sun, D.Y.; Zhou, R.G.; Li, B. Calmodulin-binding protein phosphatase PP7 is involved in thermotolerance in Arabidopsis. Plant Cell Environ. 2007, 30, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.T.; Gao, F.; Li, G.L.; Han, J.L.; Liu, D.L.; Sun, D.Y.; Zhou, R.G. The calmodulin-binding protein kinase 3 is part of heat-shock signal transduction in Arabidopsis thaliana. Plant J. 2008, 55, 760–773. [Google Scholar] [CrossRef]
- Wheeler, T.; von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef]
- Zhu, T.; De Lima, C.F.F.; De Smet, I. The heat is on: how crop growth, development and yield respond to high temperature. J. Exp. Bot. 2021, 72, 7359–7373. [Google Scholar] [CrossRef]
Figure 1.
Ca2+ channels involved in sensing heat. (By Figdraw).
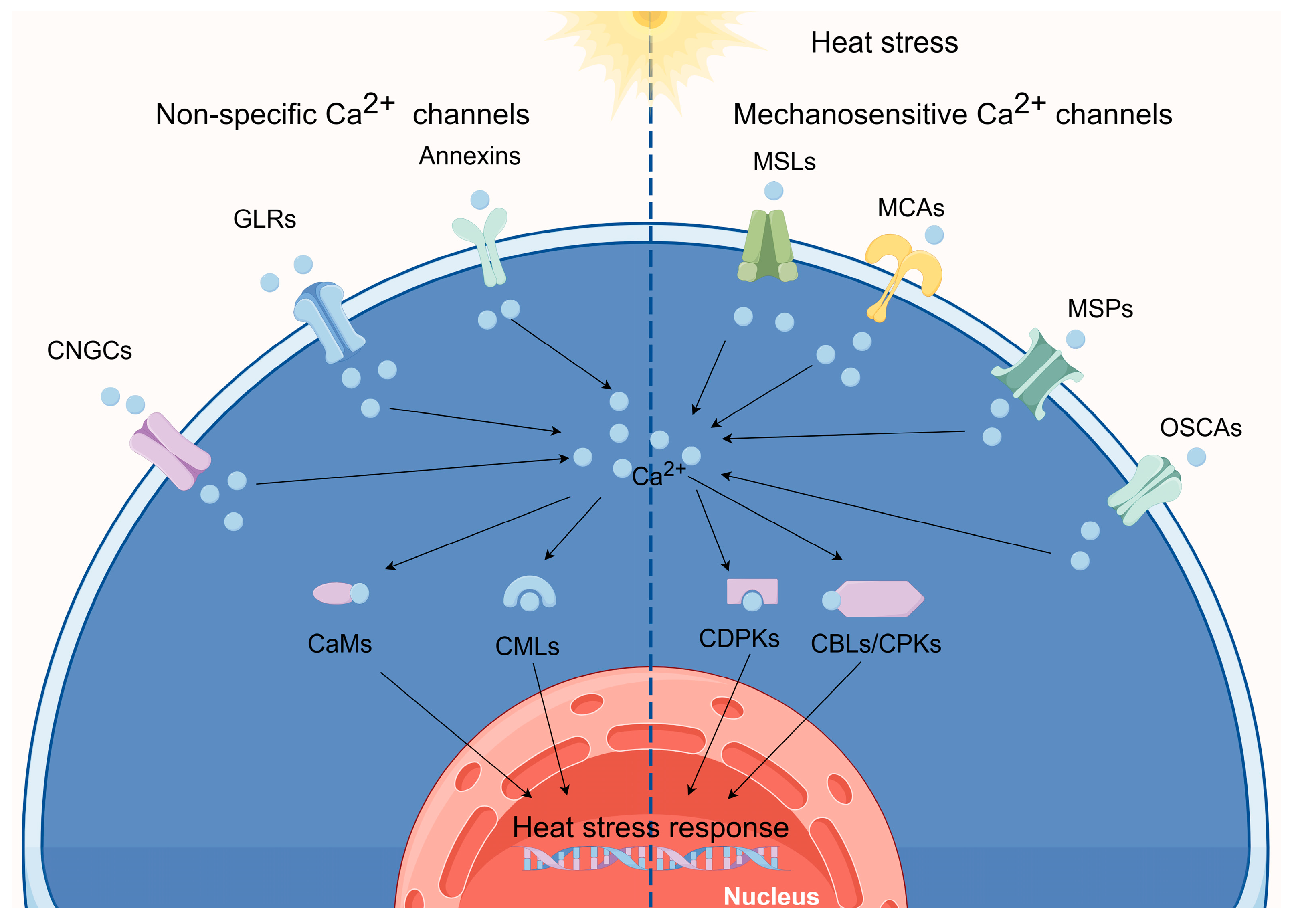
Figure 2.
Ca2+ signaling pathway under HS. (By Figdraw).
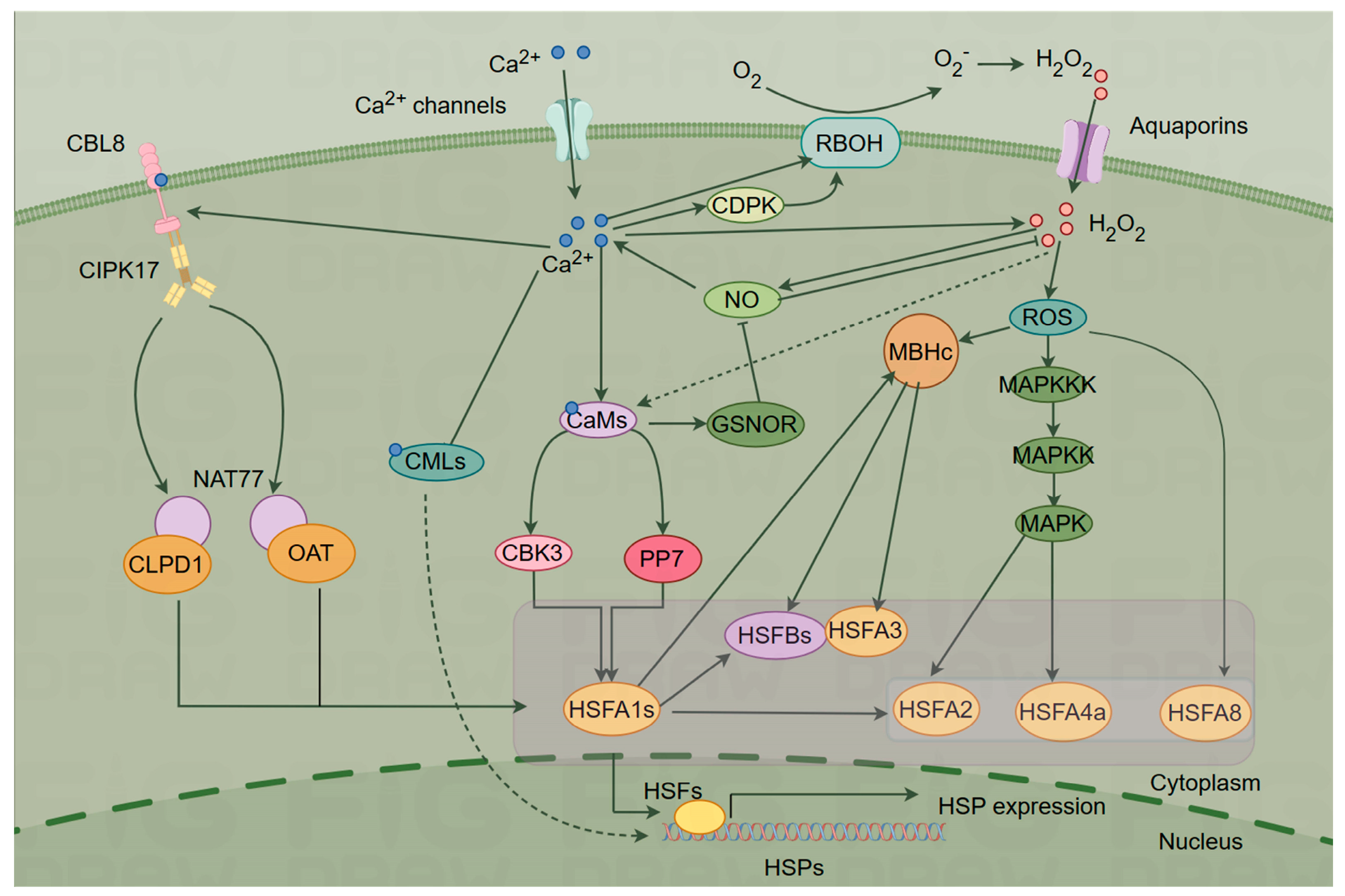
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



