
Submitted:
05 December 2023
Posted:
07 December 2023
You are already at the latest version
Abstract
Microbial competition within plant tissues affects invading pathogens' fitness. Metabolomics is a great tool for studying their biochemical interactions by identifying accumulated metabolites. Xylella fastidiosa, a Gram-negative bacterium causing Pierce's disease (PD) in grapevines, secretes various virulence factors including cell wall degrading enzymes, adhesion proteins, and quorum sensing molecules. These factors, along with outer membrane vesicles, contribute to its pathogenicity. Previous studies demonstrated that co-inoculating X. fastidiosa with Paraburkholderia phytofirmans strain PsJN suppressed PD symptoms. Here, we further investigated the interaction between the phytopathogen and the endophyte by analyzing the exometabolome of wild-type X. fastidiosa and a diffusible signaling factor (DSF) mutant lacking quorum sensing, cultivated with 20% P. phytofirmans spent media. LC-MS and MAGI were used to detect and map metabolites to genomes revealing a total of 121 metabolites, of which 25 were further investigated. These metabolites potentially relate to host adaptation, virulence, and pathogenicity. Notably, this study presents the first comprehensive profile of X. fastidiosa in the presence of P. phytofirmans spent media. The results highlight that P. phytofirmans and the absence of a functional quorum sensing affect the ratios of glutamine to glutamate (Gln:Glu) in X. fastidiosa. Additionally, two compounds with plant metabolism and growth properties, 2-Aminoisobutyric acid and Gibberellic Acid, were downregulated when X. fastidiosa interacted with P. phytofirmans. These findings suggest that P. phytofirmans-mediated disease suppression involves modulation of the exometabolome of X. fastidiosa, impacting plant immunity.
Keywords:
Xylella fastidiosa
; Paraburkholderia phytofirmans
; metabolomics
; phytopathogen
; liquid chromatography mass-spectrometry
; MAGI
1. Introduction
Deciphering molecular aspects of the interaction between Xylella fastidiosa and its plant hosts can provide important clues about disease development [1]. One key aspect leading to disease is its ability to modulate colonization behavior in plants and in insect vector transmission through quorum sensing (QS) mechanisms [2,3]. For this X. fastidiosa uses a family of modified fatty acids known as diffusible signaling factors (DSFs) that control individual and collective behavior and expression of virulence factors [4]. Many aspects contribute to the complexity of the disease at different molecular levels. These include the bacterium's capacity to become systemic, its ability to evade detection by the host immune system, and the subsequent overreaction of the plant immune system, causing, among other effects, plant water deprivation [1,5,6,7].
Microorganisms, including symbiotic bacteria, have the capacity to influence their host behavior by regulating the synthesis of specific compounds, thus fulfilling the metabolic and protein requirements of their host organisms [8,9,10]. Nevertheless, niche competition among microorganisms – a naturally environmental occurrence –, such as a spatial dispute between a pathogen and an endophyte, often alters the fitness dynamics within the host [11,12]. Nutrient competition, nutrient provision, toxin secretion, and competitor predation – which are some relational possibilities among microorganisms and their hosts – can be unveiled by metabolite level alteration assessed by metabolomics [10,13,14,15,16].
X. fastidiosa causes diseases in several plant hosts of economic importance to world agriculture, including grapevines, almonds, citrus, and more recently olive trees [1,6,17,18]. This bacterium colonizes exclusively the lumen of xylem vessels of its plant hosts and is transmitted by insect vectors like sharpshooters throughout the Americas and a spittlebug Philaenus spumarius in Europe [19,20,21]. X. fastidiosa produces biofilm and secretes several virulence factors such as cell wall degrading enzymes (CWDE), and lipases/esterases, among others [22,23,24]. X. fastidiosa does not have the Type 3 Secretion System (T3SS), which in most bacterial pathogens is responsible for the delivery of effectors/virulence factors inside host cells [25,26,27], thus reflecting the lifestyle of this phytopathogen that colonizes xylem vessels, consisted mostly tissue consists mostly of dead lignified cells [28].
Paraburkholderia phytofirmans strain PsJN [29], known as a grapevine endophyte, multiplies within grapevines, similar to X. fastidiosa [30,31]. Co-inoculation with X. fastidiosa results in a significant reduction of the phytopathogen population and disease symptoms as previously reported for other pathogens [32]. This effect involves priming innate disease resistance pathways in grapevines leading to reduced symptoms when infected with X. fastidiosa [33]. Recently, in a subsequent study, P. phytofirmans PsJN, when topically applied with a surfactant, effectively controlled Pierce's Disease (PD) in grapevines, demonstrating systemic protection against Xylella fastidiosa infection, although the extent of protection was spatially limited, with potential implications for this endophyte use as a biological control agent [34].
Exometabolomics (or footprint), i.e., analysis of secreted metabolites, is a very effective approach to track signatures of microorganisms, e.g., contamination signatures [35]. Integrating metabolomics and genomics helps to track the origins of major trends in metabolite levels in different species, but also while interacting with each other [36,37]. In this sense, the characterization of the exometabolome is a useful tool to further understand the responses of X. fastidiosa to its environment and/or interactions with other organisms. In addition, the analysis of the exometabolome has many analytical advantages over the analysis of intracellular compounds due to their lower turnover, higher stability, and, consequently, a greater reproducibility of the metabolic footprint [38,39].
Here, we show that P. phytofirmans spent media is capable of disturbing X. fastidiosa mature biofilm formation in vitro. Additionally, we demonstrate the changes in the X. fastidiosa exometabolome triggered by P. phytofirmans spent media, providing further insights into the effect on QS and disease development.
2. Materials and Methods
Maintenance and Cultivation of Bacteria
Bacterial strains used in this work are listed in Table 1:
X. fastidiosa was grown in PD3 medium (2 g/L starch; soytone 2 g/L; tryptone 4 g/L; 1 g/L sodium citrate; succinate 1 g/L; 10 mg/L hemin chloride; MgSO4 .7H2O 1 g/L; KH2PO4 1 g/L; K2HPO4 1.5 g/L) for 7 days. The WT and ΔrpfF strains were maintained in a PD3-agar medium. For ΔrpfF, PD3-agar had kanamycin added to a final concentration of 50 μg/ml.
For culture in liquid medium, after 7 days of culture in PD3-agar bacteria were transferred to 100 mL of PD3 medium and maintained at 28 °C at 100 rpm for up to 7 days. Cultures were started with OD600nm = 0.05.
P. phytofirmans PsJN was selected from a plate of King’s B medium [44] containing rifampicin (KBR) at 100 µg/mL. Next, P. phytofirmans was transferred to 100 ml of PD3. X. fastidiosa and P. phytofirmans were grown at 28 °C and 100 rpm. Fresh culture was started from frozen stocks at –80 °C.
Alternatively, WT and ΔrpfF strains were grown in PD3 supplemented with 20% of P. phytofirmans spent media. P. phytofirmans PsJN grown for 1 day in PD3 medium was centrifugated at 4000 g for 30 min, 4 °C, the supernatant was collected, adjusted to pH 4, and subjected to vacuum filtration through a 0.22 μm membrane. The eluate was added to a regular PD3 medium to a final concentration of 20% and used for subsequent experiments.
X. fastidiosa Biofilm Measurement
An aliquot (500 μL) of the P. phytofirmans PsJN spent medium eluate was added to 2.5 ml of PD3 liquid culture and then used to grow WT X. fastidiosa for 7 days at 28°C and 200 rpm. After this period the biofilm formed was quantified by staining with the crystal violet method as previously described [45]. Briefly, the biofilm deposited at the air-medium interface of the Temecula1 strain was washed with distilled water, stained for 20 min with 0.1% crystal violet reagent, washed again with distilled water, and then taken up in acetone: ethanol (6:4). Quantification was performed by measuring the absorbance at 595 nm (Abs595nm).
Analysis of Metabolites of the Supernatant of X. fastidiosa Cultures by Liquid Chromatography Coupled to Mass Spectrometry (LC-MS)
Samples (1 mL) of spent medium from cultures and sterile control medium were centrifuged in 1.5 mL microcentrifuge tubes at 1700 g for 5 min. Supernatants, containing extracellular metabolites, were lyophilized and then resuspended in LC-MS grade methanol (300 μL). Resuspended samples were centrifuged again to pellet salts. Supernatants were dried under vacuum (Savant SpeedVac Plus SC110A) and resuspended in 500 μL of LC-MS grade methanol containing a mixture of internal standards (25 μM of 13C-15N-L-phenylalanine, 2-Amino-3-bromo-5-methylbenzoic acid, 3,6-dihydroxy-4-methylpyridazine, d4-lysine, d5-Benzoic acid, and 9 anthracene carboxylic acid). The samples were filtered through a 0.22 μm microcentrifuge filtration device and transferred to 1.5 mL borosilicate glass vials (Agilent) for LC-MS analysis as previously described [63]. Extraction blanks (blank microcentrifuge tubes taken through the entire extraction procedure to check for contaminants introduced during sample preparation) were included in the analysis. A quality control mixture (a defined mixture of common metabolites) and injection blanks (pure methanol) were also analyzed at the beginning and ends of each run to ensure no drift in retention times or signal abundances and no signs of column fouling or metabolite carryover. For polar metabolite analysis, an Agilent 1290 LC system equipped with a ZIC-pHILIC column (150 mm × 1 mm, 3.5 μm 100 Å, Merck Sequant) was used for metabolite separation with the following LC conditions: solvent A, 5 mM ammonium acetate; solvent B, 9:1 acetonitrile:H2O with 5 mM ammonium acetate; timetable: 0 min at 100% B, 1.5 min at 100% B, 21 min at 0% B, 27 min at 0% B, 33 min at 100% B, and 45 min at 100% B; 0.8 mL/min; column compartment temperature of 40 °C. Mass spectrometry analyses were performed using a 6550 quadrupole time-of-flight mass spectrometer. Mass Spectrometry data were collected in a mass range of 30-1200 m/z, drying gas rate of 11 L/min, and gas temperature of 290 °C. Nebulizer pressure was set at 30 psi and capillary voltage was 3500 V. Data were acquired in both positive (+) and negative (–) polarity. The extracted samples were obtained in triplicate for both biological and technical replicates. Additionally, three replicate samples from sterile control media and eleven samples from extraction blanks were processed under the same conditions. A total of 62 injections were performed, including 3-11 replicate injections.
LCMS Data Processing and Analysis
Mass Hunter qualitative analysis software (Agilent, Santa Clara, CA, USA) was used to inspect the raw data. Then, the raw data obtained were parsed using Python scripts within MetAtlas [46], which was used to extract out putative metabolite peaks using an in-house metabolite database containing m/z and retention time data [46,47]. Next, the LC-MS data were analyzed using a custom code in Perl. Each of the detected peaks was evaluated to assign a level of confidence in the identification of the compound. Compounds that were identified had a corresponding retention time and m/z for a pure standard using the same methods as above. Negative and positive polarity mode lists of compounds were retrieved and merged, and compounds present in the negative mode were given preference over the detection in positive polarity mode. In order to confirm the identity of a compound, we checked the RT difference (i.e., abs[RTstandard – RTµexperiment]) and the ppm error mass (i.e., [(abs[m/zstandard – m/zµexperiment])/m/zstandard] ×106) (Supplementary Table S1). Compounds with RT difference ≤ 0.05 and ppm error mass ≤ 6 were considered with their identities confirmed (Supplementary Table 1) [48]. Compounds that did not pass one or both criteria (up to ppm error mass ≤ 15) were considered putative. Next, compound abundance was normalized by strains OD in each biological replicate (Supplementary Table 2).
For further comparisons among conditions, it was considered expressed in the supernatants the compound that satisfied the following criteria: [ µisample - µiblank] - [ µimedium - µiblank] > 0. Where µ stands for “average of peak area value” and i stands for any detected compound. Finally, an ANOVA test was conducted to acknowledge variability across all conditions (Supplementary Table S2). Compounds with a p-value ≤ 0.05 were considered statistically significant. Pairwise comparison among conditions was performed and a t-test with a p-adjusted value ≤ 0.05 was calculated for each compound. Compounds with statistical significance between two conditions were considered differentially expressed metabolites (DEM) (Supplementary Table S3). Extracted ion chromatograms (EIC) were generated, one representative EIC of each extract or exometabolome was used for building a summary panel for each discussed compound and all the remaining EIC from all biological and technical replicates were made available in the supplementary material (Supplementary Figures S1–S3).
A Python library for the Venn diagram was used to highlight unique and shared compounds across the conditions tested. These were referred to as Xf (X. fastidiosa WT Temecula1), ΔrpfF (X. fastidiosa ΔrpfF), Xfsm (X. fastidiosa Temecula1 WT cultivated in PD3 media supplemented with 20% P. phytofirmans spent media), ΔrpfFsm (X. fastidiosa ΔrpfF cultivated in PD3 media supplemented with 20% P. phytofirmans spent media), and P. phytofirmans PsJN (Pp). A matrix was built from three biological replicates of Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp. This matrix was given as an input to a multivariate data analysis in a principal coordinate analysis (PCoA) [49], taking as a parameter the Euclidean distance between the samples. To compute this dataset, we used the Python library skbio, and the library emperor to build the PCoA plot. Permanova and permdisp statistical analysis were made with the PCoA input data using skbio modules permanova and permdisp, respectively, and the information was added later to the PCoA plot.
A heatmap based on clustering of the compounds across conditions was generated with the Python libraries matplotlib and seaborn. The expression values of all compounds in all conditions were normalized by a z-score. A color code and symbols were added to compound names to indicate identification level and statistical significance.
Integration of Metabolites and Gene Set for Functional Annotation of Exometabolomes
Whole genome annotation (in fasta format) from Xylella fastidiosa Temecula1 and Paraburkholderia phytofirmans PsJN together with a list of InChiKey identifiers from all the compounds assigned as expressed in previous steps were given as input for a container (docker) of MAGI v1.0 [50,51] for metabolite and gene integration. The output spreadsheets were filtered by metabolites expressed in each given condition. Next, all genes annotated by MAGI were retrieved and duplicates were filtered out. Each gene list representing an exometabolome was submitted to the software OmicsBox [52] for functional annotation using gene ontology. Enrichment analysis was performed, annotations with p-value adj ≤ 0.05 were collected and the values were log2 transformed and ranked. The top 5 under and over-enriched annotations from each condition were collected for comparison. Matplotlib and seaborn Python libraries were used to build a bar plot.
Bioinformatic Analysis
Expressed metabolites were compared in box plot and bar plot, coded by matplotlib and seaborn Python libraries. Pairwise significance comparison was performed by the Python library statannotations, t-test argument was used. When necessary precomputed p-adjusted values for pairwise comparison were annotated later (Supplementary Table S2).
Metabolic reactions of nicotinamide, nicotinate, and biotin were freely redrawn from annotated X. fastidiosa and P. phytofirmans Kegg pathways. Illustrations of gene operons were rebuilt based on the synteny presented in the browser IMG/DoE [53]. Mummer [54] was used for the global alignment of genomic sequences.
For the metabolome and transcriptome integration, a bubble plot was created using matplotlib and seaborn Python libraries. The metabolites' expression values were 2z-score transformed in order to maximize the expression differences and keep all values positive.
Cooperative Metabolic Interactions between X. fastidiosa and P. phytofirmans
In order to analyze the potential ecological microbial interactions [55] between X. fastidiosa and P. phytofirmans, a reverse ecology analysis was applied to predict the ecological structure of this symbiotic relationship [56].
For defining microbe-microbe cooperative and competitive potential in a pairwise manner, a local and customized NetSeed version [57] was used (to include the metabolic complementarity index and effective metabolic overlap index) [58, 59]. The metabolic complementarity index between two species measures the ratio (range 0–1) of acquired compounds in one species that are found in the metabolic network of the other species and are not used by it. The effective metabolic overlap index of two species is the fraction of compounds required by both species and is, therefore, a measure of potential nutritional competition [58] with values ranging from 0 (no competition) to 1 (high competition).
3. Results
P. phytofirmans Interacts with X. fastidiosa through its Exometabolome
In a prior study, Baccari and colleagues [33] demonstrated that direct contact between X. fastidiosa and P. phytofirmans was not necessary for reducing X. fastidiosa titer loads in xylem vessels. Here we explore the indirect interaction between X. fastidiosa and P. phytofirmans, including the QS-insensitive mutant ΔrpfF. Following initial trials (unpublished data), we established a specific condition for subsequent experiments. We used PD3 culture media supplemented with 20% P. phytofirmans PD3 spent media, which noticeably affected the X. fastidiosa phenotype. This impact was particularly evident in terms of biofilm intensity and deposition, as measured by the Crystal Violet assay (Figure 1A).
During these experiments, we cultivated both Xf and ΔrpfF in regular PD3 culture media or supplemented with 20% P. phytofirmans PsJN spent media (Xf sm or ΔrpfFsm), and Pp. The biofilm formation of Xf sm and ΔrpfFsm increased by 39% and 128%, respectively (Figure 1B). Moreover, a larger amount of bacterial biomass was visually observed at the medium/air interface of Xf sm and ΔrpfFsm compared to Xf and ΔrpfF (Figure 1A). As PD3 supplemented with 20% P. phytofirmans spent media led to quantifiable changes in the biofilm formation profile of both bacterial strains in in vitro cultures, we hypothesized that the secreted metabolome (exometabolomes) of P. phytofirmans triggers a disturbance in the exometabolomes of the tested Xylella strains.
Exometabolome Variation among X. fastidiosa Strains in Response to P. phytofirmans
Subsequently, we investigated the exometabolomes of Xf and Pp to characterize the qualitative and quantitative signatures of compounds. By employing an internal reference library, we were able to identify a total of 131 compounds across all strains and experimental conditions (Supplementary Tables S1–S3). Referring to the detailed criteria for detection outlined in the Methods section, we established a threshold and inferred the detection of 121 compounds among the entire spectrum of strains and conditions examined (Supplementary Table S3) [48]. Specifically, within Xf, Xfsm, ΔrpfF, ΔrpfFsm, and Pp, we detected 114, 119, 118, 115, and 48 compounds, respectively (Figure 2A).
There is a shared core of 43 compounds present in all five exometabolomes. Focusing exclusively on the X. fastidiosa strains and treatments (Xf, Xfsm, ΔrpfF, and ΔrpfFsm), the Venn diagram highlights a core of 112 compounds. Notably, the Venn diagram also unveils a distinct subset of exclusive compounds (69) unique to X. fastidiosa (Xf, Xfsm, ΔrpfF, and ΔrpfFsm). The preeminent class of compounds within the core exometabolome belongs to "nucleotides and derivatives" (comprising 32.6% of the total). Among the exclusively expressed compounds found in X. fastidiosa strains, amino acids and derivatives dominate (52.2%). It is noteworthy that nicotinamide stands as the sole compound exclusively identified in Pp exometabolome, explored in further detail in the subsequent section.
Utilizing Principal Coordinate Analysis (PCoA), we discerned a distinct clustering pattern within the exometabolomes, initially delineating separation based on bacterial species (Xf versus Pp), and subsequently segregating according to strains and treatments (Xf versus ΔrpfF, and Xf sm versus ΔrpfFsm) (Figure 2B). This analysis yielded the formation of three primary and discernible clusters: (1) Pp, (2) Xf sm, ΔrpfF, and ΔrpfFsm, and (3) Xf. Notably, the implementation of Permanova and Permdisp analyses yielded statistically significant outcomes, denoted by p-values of <0.05 and <0.005, respectively, with respect to the distribution of samples.
Drawing from the outcomes of the PCoA, we postulate that the metabolites originating from Pp exert an influence on Xf exometabolomes, leading to a convergence in profile akin to that of the ΔrpfF metabolome. Interestingly, while ΔrpfF remains unaffected, as evidenced by the similarity to ΔrpfFsm samples, this transformative effect is conspicuously absent. The PCoA not only highlights sample correlation but also offers a platform for constructing a working hypothesis. In essence, the assimilation of Pp spent media by Xf potentially results in a convergence of exometabolomic profiles, aligning Xf more closely with the characteristics of ΔrpfF, at least on the exometabolome level. A noteworthy observation is the similarity apparent in the sample distribution between ΔrpfF and ΔrpfFsm. The distribution of exometabolome samples, as represented in Figure 2b, guides our hypothesis that the interaction of Xf with Pp spent media draws it near to the phenotype of ΔrpfF.
Finally, we generated a heatmap illustrating the expression levels of compounds, normalized by 2z-score. To enhance our comprehension of the functional attributes, variability, and expression levels of the metabolites, we organized the identified compounds into groups based on PubChem classes [60]. These groups correspond to various chemical classes, including carbohydrates, amino acids (and derivatives), nucleotides (and derivatives), carboxylic acids, pyridines, amines, sulfurs, terpenes, polyols, parabens, and vitamins. Noteworthy are the compounds that serve as intermediates within metabolic pathways and the citric acid cycle (such as α-ketoglutarate, fumarate, lactate, and succinate), along with secondary metabolites (including suberic acid and shikimic acid), present in the exometabolomes of both X. fastidiosa and P. phytofirmans (Figure 2C and Supplementary Table S3).
Within Xf, 75 metabolites demonstrated significantly upregulated expression levels compared to Pp, ΔrpfF, Xf sm, and ΔrpfFsm. Of these, 44% were amino acids and derivatives. Similarly, Xf sm exhibited a majority of upregulated metabolites (5), accounting for 60% of amino acids and derivatives. Conversely, ΔrpfF and ΔrpfFsm displayed 8 and 12 upregulated compounds, respectively, with 50% and 41,7% being carboxylic acids. In contrast, the Pp exometabolome exhibited notably low metabolite detection, particularly when contrasted with those identified in Xf exometabolomes. Of the detected metabolites in Pp, 10 exhibited upregulation when compared to their counterparts in Xf exometabolomes, of which 50% were nucleotides or nucleotide derivatives.
The heatmap construction encompassed all compounds detected above the threshold of blank (MeOH) and culture media (PD3 or PD3sm), totaling 121 compounds. Our annotation approach included two additional layers of information—statistical significance, reflecting variance among conditions, and the level of identification. The heatmap (Figure 2C) encompasses compounds that met the criteria of standard confirmation through RT and m/z ppm, and annotations also included assigning compounds demonstrating statistical significance (p < 0.05) based on ANOVA analysis (Supplementary Tables S2 and S3). A notable feature of the heatmap is the emergence of two distinct clusters: one primarily composed of compounds highly expressed in Pp, and the other predominantly featuring compounds highly expressed in Xf strains. There exists a subset of compounds that are more highly expressed in Xf sm, ΔrpfF, and ΔrpfFsm in contrast to Xf. Upon a comprehensive review of the overall detected metabolites, we focused the investigation on a subset of 25 compounds (amino acids, vitamins, and plant hormones), as detailed in the subsequent sections.
X. fastidiosa Secretes High Amounts of Amino Acids and Vitamins
First, we chose to further investigate compounds that are well known for their role in interaction between bacteria and some species of insect hosts [61]. For such, we examined amino acid levels among the strains and conditions.
The nutritional status of amino acids within the exometabolomes of Xf, ΔrpfF, Xf sm, ΔrpfFsm, and Pp was assessed, primarily utilizing the ratio of glutamine (GLN) to glutamate (GLU). This ratio, commonly employed to gauge nitrogen (N) status in various eukaryotic cells, has been linked to N limitation when <0.2 and indicative of N-replete cells when >0.5 [10,62]. The computed median GLN:GLU ratios within the exometabolomes unveiled variations: Xf, ΔrpfF, Xf sm, and ΔrpfFsm exometabolomes all exhibited ratios predicting N-replete conditions, spanning from 0.52:1 in Xf to 8.33:1 in ΔrpfF (Figure 3A). Notably, Gln and Glu were not detected within the exometabolome of Pp under the assessed conditions. The 0.5 threshold for N-replete conditions was drawn in the plot, with all conditions (Xf, Xf sm, ΔrpfF, and ΔrpfFsm) surpassing this threshold—except for Pp, which lies below, and Xf, which teeters at the threshold. Importantly, our analysis did not identify any statistically significant differences between Xf, Xf sm, ΔrpfF, and ΔrpfFsm.
In order to provide a comprehensive understanding of compound levels within the experimental framework, we constructed a panel of EIC encompassing various controls, including the blank (MeOH), and culture media controls, PD3 and PD3sm (Figure 3B,C; Supplementary Figures S1 A–T). These control conditions were juxtaposed with the investigated states: Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp. As anticipated, control conditions (culture media PD3 and PD3sm) exhibited detectable amino acid levels. Upon scrutinizing Xf, Xf sm, ΔrpfF, and ΔrpfFsm, a noticeable pattern emerges: amino acid levels, whether pertaining to nEAA or EAA, consistently surpass those found in the blank samples and even exceed the amino acid concentrations in the culture media controls. This observation aligns with the knowledge that PD3, given its protein-rich composition, inherently contains a source of amino acids. Upon closer examination, this elevated amino acid content within Xf, Xf sm, ΔrpfF, and ΔrpfFsm challenges conventional assumptions, indicating a plausible role as an insect-vector decoy within its presumed biological function. In stark contrast, the levels of EAA and nEAA in Pp are notably diminished, bordering on complete absence.
While the visual inspection of differences in the levels of essential amino acids (EAA) and non-essential amino acids (nEAA) does not consistently yield pronounced distinctions, we undertook individual assessments of these two amino acid categories. The nature of this evaluation displayed variability across species, strains, and treatments, with notable statistical significance emerging primarily between amino acid quantifications in Xf and Xf sm. Specifically, variations were evident in tyrosine, serine, proline, glutamine, aspartate, asparagine, alanine, and glycine within the nEAA, and valine, methionine, lysine, leucine, isoleucine, and arginine within the EAA. Similarly, only histidine exhibited a discernible difference between ΔrpfF and ΔrpfFsm (Figure 3B,C). Notably, within the exometabolome of Pp, neither EAA nor nEAA were detected.
Given the disparities in ionization among amino acids, merging them into a unified value for comparison across diverse conditions is not viable. Nonetheless, a consistent observation emerges: essential amino acids (EAA) exhibit higher individual abundance within Xf, Xf sm, ΔrpfF, and ΔrpfFsm in comparison to non-essential amino acids (nEAA).
We also investigated levels of complex B vitamin detected in the tested exometabolomes. These compounds are relevant to the relationship between insect and host plant. Our findings illustrate the accumulation of nicotinamide in Pp, while it is notably absent in Xf, Xf sm, ΔrpfF, and ΔrpfFsm—this absence is evident across both normalized values (peak area intensity by OD) and EIC profiles, including comparison with controls. Conversely, the presence of nicotinic acid follows an inverse pattern (Figure 4A–F, Supplementary Figure S2A–C). The product of pncA, namely nicotinic acid, exhibits high expression in Xf, along with downregulation and relatively consistent levels in Xf sm, ΔrpfF, and ΔrpfFsm. Biotin, on the other hand, was not detected in Xf, ΔrpfF, or Pp. In contrast, its presence was identified at relatively low levels in Xf sm and ΔrpfFsm. The pathways remain conserved in both X. fastidiosa and P. phytofirmans (Figure 4G,H).
Two Plant Hormones are Exclusively Secreted by X. fastidiosa
We conducted a comparative analysis of the levels of two compounds, 2-aminoisobutyric acid (AIB) and gibberellic acid (GA), detected within the exometabolomes of Xf, ΔrpfF, Xf sm, ΔrpfFsm, and Pp. These compounds possess implications for the interaction between X. fastidiosa and its plant hosts, potentially functioning as pathogen metabolite effectors. AIB, a non-proteinogenic amino acid, exists in other two isomeric forms: β-aminobutyric acid (BABA), known to induce plant disease resistance, and γ-aminobutyric acid (GABA), a neurotransmitter in animals that is also produced in plants, likely serving a signaling role. GABA was detected highly abundant in xylem sap of grapevines infected with X. fastidiosa . Gibberellic acid (GA), on the other hand, functions as a hormone in both plants and fungi.
Across the conditions assessed in our metabolomics analysis, AIB and GA exhibit notably similar profiles. Both compounds form three distinct clusters: (1) Xf, (2) Xf sm, ΔrpfF, and ΔrpfFsm, and (3) Pp (Figure 5A,B, Supplementary Figure S3A,B). Comparatively, AIB and GA levels prove significantly higher in Xf than in any other strain or treatment. Xf sm displays reduced AIB and GA expression, akin to the observed pattern in ΔrpfF. Conversely, ΔrpfFsm exhibits no significant alteration in AIB and GA expression compared to ΔrpfF. Notably, AIB remains undetected in the Pp exometabolome, while GA levels range from 6.63% to 26.89% of those detected in Xf, ΔrpfF, Xf sm, and ΔrpfFsm exometabolomes.
Chromatograms illustrate the contrast between AIB levels in Pp, which appear lower than those in controls, while AIB levels in Xf consistently remain elevated (Figure 5C). GA is generally detected at low levels across all conditions, including Xf. However, both Xf and ΔrpfF exhibit levels 2-to-3 times higher than controls. Xf sm and ΔrpfFsm exhibit increases of approximately 100% and 50%, respectively, compared to the culture medium controls. Notably, Pp's GA levels mirror those of the controls (Figure 5C and D).
We investigated the potential synthesis of AIB and GA by X. fastidiosa or P. phytofirmans through the search for gene clusters for their synthesis. We examined the presence of homologous genes or clusters akin to the reported aib operon from Rhodococcus wratislaviensis [63] and the Ga operon from Xanthomonas oryzae pv oryzicola [9, 64]. While we identified orthologs of certain genes associated with AIB or GA synthesis (or metabolism) in X. fastidiosa or P. phytofirmans, we did not find them organized in operons as originally annotated in the genomes of Rhodococcus wratislaviensis and Xanthomonas oryzae pv oryzicola, respectively (Supplementary Tables S14 and S15).
Exometabolome, genome, and transcriptome integration for X. fastidiosa and P. phytofirmans
Firstly, we used NetSeed [57] — a tool for assessing metabolic network topology and determining the set of exogenously acquired compounds — to assess the metabolic networks of X. fastidiosa and P. phytofirmans predicting them as profiles in cooperation or competition. The analysis indicated a low degree of complementarity (< 0.32) and a moderate-to-high level of competition (> 0.49) between X. fastidiosa and P. phytofirmans.
In a second approach, using MAGI v.1.0 [50] we searched for a connection between annotated genes in X. fastidiosa or P. phytofirmans genomes and the detected compounds, respectively, in Xf, Xf sm, ΔrpfF, ΔrpfFsm or Pp (Table 2, Table 3 and Supplementary Tables S4–S8). Out of the 121 identified compounds given as input to MAGI, 118 compounds returned results in gene-to-compound connection information from X. fastidiosa or P. phytofirmans. Specifically, all surveyed amino acids (both nEAA and EAA) exhibited connections with high reciprocity scores (≥2), i.e., compounds and annotated genes are both linked to the same reactions or metabolic pathways from the consulted databases RheA [65] or MetaCyc [66]. CDS coding for amino acid metabolism was previously annotated in the genomes of X. fastidiosa [27] and P. phytofirmans [43]. Results for AIB showed a high reciprocity score with 18 and 11 genes in X. fastidiosa and P. phytofirmans, respectively (Supplementary Tables S4–S7). Conversely, GA displayed a notably low reciprocity score with the only two associated genes in X. fastidiosa (PD0286, PD0716) and likewise with the three genes (Bphyt_2421, Bphyt_2594, and Bphyt_0368) in P. phytofirmans (Supplementary Table S8), suggesting a limited likelihood of synthesis by these genes and, respective reactions. In P. phytofirmans, Bphyt_5413 is linked to nicotinamide metabolism. In X. fastidiosa, PD0043, PD1071, and PD1494 are linked to biotin metabolism and PD0393 and PD1310 to nicotinic acid metabolism.
Next, the resulting genes from MAGI analysis as described above were used for functional annotation via gene ontology [67,68], through OmicsBox (Supplementary Tables S9–S13). From all the annotated functions, we focused on the top five over-enriched and top 5 under-enriched functions (Figure 6A). Despite minor variations in ΔrpfF, Xf sm, and ΔrpfFsm—especially when compared to Xf—over-represented functions generally appeared common across those conditions, such as "ligase activity, forming carbon-nitrogen bonds," "oxidoreductase activity, acting on NAD(P)H," and "ligase activity" (log2FC > 5). Functions like "Vitamin B6 binding," "pyridoxal phosphate binding," and "tricarboxylic acid cycle" were enriched in Xf, ΔrpfF, Xf sm, and ΔrpfFsm, but under-enriched in Pp. Conversely, "CoA-ligase activity" and "fatty acid ligase activity" were highly over-represented in Pp (log2FC > 5), whereas they were not enriched in Xf, ΔrpfF, Xf sm, or ΔrpfFsm (Figure 6A).
Finally, we lined up our metabolome data with a previously published RpfF-Regulon, i.e., transcriptomic data obtained with microarray technology [3]. We did not directly and statistically integrate metabolome and transcriptome data, but, instead, we tracked genes directly linked to the detected compounds (obtained from MAGI) and cross-referenced them with the published differentially expressed genes (DEGs) in the RpfF-Regulon (Figure 6B). A consistent pattern exists where genes downregulated in the ΔrpfF strain were associated with lower levels of metabolites detected in ΔrpfF compared to Xf, including 2-Aminoisobutyric acid, cAMP, cGMP, Phenylalanine, and Tyrosine. In contrast, the upregulated DEGs in the ΔrpfF strain showed no pattern at the metabolite level. Instead, metabolite expression within Xf sm, ΔrpfF, and ΔrpfFsm differed when compared to Xf, signifying a lack of regularity but rather an upregulation of specific metabolites in these conditions. Metabolites cross-checked as upregulated in metabolome and as a consequence of upregulated genes in RpfF-regulon include Choline, Cysteine, 2-deoxyAdenosine-5’-monophosphate, Guanosine-5’-monophosphate, Citraconic acid, Asparagine, Glutamic acid, Threonine, Serine, Homoserine, Uridine-5’-monophosphate, Uridine, Thymidine, 2-deoxyUridine, Cellobiose, and Galactose.
4. Discussion
In prior studies, it was demonstrated that the co-inoculation of P. phytofirmans along with X. fastidiosa leads to a reduction in the symptoms of leaf scorching induced by X. fastidiosa in grapevines. This ameliorative effect was attributed to the activation of plant defense genes, specifically involving pathways related to salicylic acid and ethylene [33]. It was further reported that the topical application of P. phytofirmans with a surfactant, was efficient and sufficient for PD control in grapevines, even though the protection was spatially limited [34].
In our investigation, we observe that the cultivation of X. fastidiosa in PD3 medium supplemented with 20% spent media from P. phytofirmans (Xf sm and ΔrpfFsm) results in a perturbation of the X. fastidiosa biofilm structure. In particular, the biofilm exhibits an increased volume and extends across the air-liquid media interface, deviating from the usual compacted ring pattern observed in the liquid culture of Xf. Additionally, Baccari and colleagues [33] previously highlighted a synergistic impact arising from the concurrent infection of X. fastidiosa and P. phytofirmans, leading to the activation of plant defense mechanisms. However, in that work, there was no exploration of a direct interaction between P. phytofirmans and X. fastidiosa, at the metabolomic level, that could provide insights into their relevance for conventional virulence and plant colonization.
While comparing metabolite profiles, we found a considerable disparity in the number of identified metabolites in Pp in comparison to Xf, Xf sm, ΔrpfF, and ΔrpfFsm. The limited abundance of detected metabolites in Pp, coupled with reduced variability among replicates, contributes to its distinct distribution pattern observed in the Principal Coordinate Analysis (PCoA). While replicates of the Xf exometabolome exhibit close clustering, there is noticeable variability within the Xf sm, ΔrpfF, and ΔrpfFsm exometabolomes, although statistical analyses such as Permdisp and Permanova confirm significance across all conditions. The absence of a functional quorum sensing (QS) system in the ΔrpfF strain is mirrored in its altered metabolite expression profile, akin to findings in other omics analyses such as transcriptomics and outer membrane vesicle (OMV) proteome [3,69]. Notably, the exometabolome profile of Xf sm shows greater similarity to that of ΔrpfF as opposed to Xf. Nevertheless, ΔrpfFsm exometabolomes maintain an overall likeness to ΔrpfF. Consequently, we report that Pp affects Xf sm exometabolome, although this effect is notably diminished in the impaired DSF-producing strain (ΔrpfFsm).
X. fastidiosa is known to exhibit a hypersecretory trait, particularly regarding virulence proteins and extracellular vesicles (EVs), as established in prior reports [23,24,69,70]. Expanding on this, our study targeted the characterization of X. fastidiosa exometabolome, focusing on low molecular weight compounds. Our findings indicate that, in overall comparison among the 5 different groups, Xf has a higher expression of compounds. Pairwise comparison of Xf versus Xf sm, ΔrpfF, ΔrpfFsm, or Pp shows a similar pattern, with, at least 80% of compounds being upregulated in Xf. It remains an area for further investigation to determine whether the quorum sensing system interferes with the ability of X. fastidiosa to secrete metabolites. The high secretion of certain metabolites, as will be discussed below, could benefit X. fastidiosa from the plant defense mechanisms as the phytopathogen is still not established in a higher population under the influence of the QS system [2].
Another point to highlight, ΔrpfFsm exometabolome suggests a less responsive profile to P. phytofirmans spent media than Xf sm. Detected metabolite levels in ΔrpfF/ΔrpfFsm are more similar to the levels in Xf/Xf sm (e.g., they had 114 compounds detected in common, from which only 2 were DEM downregulated in ΔrpfF/ΔrpfFsm). Taken together, these observations indicate that at least partially, the detected responses to P. phytofirmans occur by disturbance of DSF signaling in Xf. Here we were limited to metabolites being detected through the LCMS method in a HILIC column. Other coupled MS analyses could complement our findings, e.g., using a c18 column (focused on non-polar and large compounds) or through a surface-based MS technique such as NIMS, which would increase the detected metabolites as seen for other models [71,72].
From the exometabolomes of Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp, two sets of compounds were further studied: amino acids (EAA and nEAA), vitamin B complex, and hormones. The average GLN:GLU ratios are highly similar in Xf and ΔrpfF (0.52 and 0.57, respectively). These values are regarded in the literature as (> 0.5) indicative of N-replete cells [62]. The average GLN:GLU ratios in Xf sm and ΔrpfFsm show an increased trend from treatments without contact with P. phytofirmans spent media, although without statistical significance. Secondly, the EAA and nEAA overall levels were inspected using their chromatogram abundance profiles. EAA are more abundant than nEAA in Xf when compared to Xf sm. It is noteworthy that a high ratio of EAA:nEAA was reported in the bacteriomes of four xylem-feeding insects [10]. We speculate that X. fastidiosa exometabolomes showing especially high EAA might be an indication of attractiveness to insect vectors. Indeed, studies in the field that had simultaneously evaluated the xylem of host plants and insects report that adults chose plants with high amino acid concentrations [19]. The levels of EAA and nEAA are significantly different between Xf and ΔrpfF. Another point, a study conducted by Daugherty and colleagues [73] indicated that the insect prefers plants with lower symptoms than plants with disease symptoms. In fact, the ΔrpfF mutant reaches faster a systemic infection in grapevines compared to the WT strain and ΔrpfF has a lower fitness for being less transmitted by the insect vector when compared to the WT strain. Moreover, ΔrpfF has a restricted ability to form cell aggregates, which is the successful state of X. fastidiosa when acquired by the insect vector feeding on xylem sap, as reported for WT strain [41]. In this case, it remains to be tested whether the lower level of amino acids secreted by the ΔrpfF would add to its worse tendency to be acquired by the insect. P. phytofirmans had very low or totally absent detection of amino acids in its exometabolome under the conditions here tested. Nevertheless, P. phytofirmans has all the predicted pathways for amino acid production according to its genome annotation [43].
Similar to amino acids, compounds within the Vitamin B complex hold particular significance for insect-borne phytopathogens [74]. Our metabolomics analysis unveiled the presence of nicotinamide, nicotinic acid, and biotin. Notably, the interplay between nicotinamide and nicotinic acid mirrors the pattern of coding sequences present or absent in X. fastidiosa and P. phytofirmans. The enzyme nicotinamidase is predicted to convert the compound nicotinamide into nicotinic acid (both being isoforms of B3 vitamin). In fact, in X. fastidiosa genome is predicted the enzyme pncA (nicotinamidase) is, whereas P. phytofirmans genome there is no homolog of a nicotinamidase gene predicted. This conversion was indeed corroborated by our metabolomics analysis (Figure 3, Supplementary Figure S4). Symbiont bacteria, e.g., Wolbachia and Baumannia, have been reported to convert precursors and provide B vitamins to their insect xylem-sap feeding hosts [74]. Nevertheless, vitamin B3 is usually not among the B vitamins this symbiont bacteria are able to yield. It remains to be further investigated if the X. fastidiosa provision of this specific isoform of B vitamin increases the host plant attractiveness towards the insect vector. We also detected biotin, another compound from vitamin B, in Xf sm and ΔrpfFsm. Nevertheless, there was a low level of expression in both exometabolomes and a low level of identification.
AIB is reported as the immediate precursor of ethylene in higher plants. Specifically, AIB inhibits ethylene production because it acts through competitive inhibition of the conversion of 1-aminocyclopropane-1-carboxylic acid to ethylene [75]. Very little is known about AIB metabolism in bacteria, although it is reported that the AIB catabolism in Rhodococcus wratislaviensis C31-06 leads to its conversion into α-methyl-D-serine, and after other downstream metabolic conversions, it results in pyruvic acid [63]. In turn, gibberellic acid is a well-known plant growth promoter [76]. In bacteria, it seems the role of gibberellins is linked to pathogenicity, where secreted gibberellins could act as virulence factors, most probably by suppressing jasmonic acid formation and, ultimately, impairing the host defense response [64], as reported in the rice pathogen Xanthomonas oryzae pv. oryzicola [77].
The identity of compounds AIB and GA was confirmed by comparing with standards by MS. Nevertheless, operons for AIB and GA synthesis are absent in X. fastidiosa and P. phytofirmans as seen in other bacteria, it remains to be further investigated which genes are directly linked with their synthesis in X. fastidiosa or P. phytofirmans. As mentioned before, GABA was highly detected in the xylem sap of grapevines infected with X. fastidiosa [15]. Also, the fact that is well established that these hormones have a function on plant growth and development makes it even more interesting to address the role of AIB and GA as virulence factors for X. fastidiosa during disease progression. AIB and GA do not seem to have a synergistic effect therefore it remains unclear how their concomitant secretion would benefit X. fastidiosa. AIB has a higher expression than GA (3,444,408.4 intensity×OD-1 versus 506,103.7 intensity×OD-1, respectively) and in case its function prevails it would most probably induce an impairment of host plant growth and X. fastidiosa could take advantage of that by a slower response from the host plant in its vasculature system. The identification and characterization of changes in specialized metabolites caused by X. fastidiosa in plant hosts are still in the early stages. However, collectively, these findings present a promising outlook on the reprogramming of metabolism following the interaction of plants with the phytopathogen [78]. For instance, in infected Leccino plants, higher amounts of salicylic acid was observed compared to Cellina di Nardò plants. Leccino variety develops milder symptoms compared to those observed on the Cellina di Nardò variety [79]. Although it has been described a GA cluster in Burkholderia species [80], a closely related species, such a gene cluster was not found in the P. phytofirmans genome (Supplementary Figure S4).
Reverse ecology analysis revealed generally low complementarity (< 0.32) and medium-to-high competition scores (> 0.49) for X. fastidiosa towards P. phytofirmans and vice-versa as stated in the results section. Therefore, X. fastidiosa and P. phytofirmans are predicted competitive bacterial species. This has implications for the search for a microorganism that can act as a biocontrol for the diseases caused by X. fastidiosa, as seen in the natural occurrence of other endophytes, e.g., M. mesophilicum in orange trees [81].
Overrepresented Gene Ontology (GO) annotations reveal distinctions not only between Xf and Pp but also with Xf sm, ΔrpfF, and ΔrpfFsm. Collectively, these findings suggest that P. phytofirmans induces a metabolic perturbation in Xf sm, rendering it more similar to ΔrpfF. This observation is further supported by the overall similarity between ΔrpfFsm and ΔrpfF. The specific metabolite or set of metabolites produced by P. phytofirmans, which is not the subject of investigation in this study and appears not to be synthesized by a ΔrpfF ortholog (as reported by Baccari and colleagues [33]), may still be exerting a disruptive effect on quorum sensing. Notably, quorum sensing disruption appears to be an effective strategy for disease suppression. For instance, in a study both tobacco, a model host, and orange tree, a natural host, that produces DSF have shown increased resistance to X. fastidiosa. The disruption in signaling confounds bacterial behavior and hinders disease development [82].
Overall, we demonstrated that metabolomics can be used for the prospection of X. fastidiosa footprint. The secretion of EAA and nEAA, vitamin B complex, and the ratio between Gln:Glu are influenced by P. phytofirmans spent media and also, by the absence of a functional QS signaling. Finally, metabolites with plant metabolism and growth properties, AIB and GA, were detected, which are down-regulated in X. fastidiosa when in contact with P. phytofirmans spent media or in the absence of QS signaling. To our knowledge, this is the first report of its exometabolome in response to another endophyte.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: List of EICs from Essential Amino Acids and Non Essential Amino Acids in all conditions and replicates; Figure S2: List of EICs from Nicotinic Acid, Nicotinamide and Biotin in all conditions and replicates; Figure S3: List of EICs from Isoaminobutyric Acid and Gibberellic Acid in all conditions and replicates; Figure S4: Comparison between Burkholderia cepacia genome to its close related species, P. phytofirmans; Table S1: Summary of metabolites standard comparison; Table S2: List of metabolites with presence-absence detection analysis; Table S3: Final list of confirmed metabolites and pairwise statistical analysis; Table S4: MAGI Xf's genome-metabolome integration; Table S5: MAGI Xf sm' genome-metabolome integration; Table S6: MAGI ΔrpfF's genome-metabolome integration; Table S7: MAGI ΔrpfFsm' genome-metabolome integration; Table S8: MAGI Pp's genome-metabolome integration; Table S9: Xf's genome-metabolome extracted gene ontology enrichment; Table S10: Xf sm' genome-metabolome extracted gene ontology enrichment; Table S11: ΔrpfF's genome-metabolome extracted gene ontology enrichment; Table S12: ΔrpfFsm' genome-metabolome extracted gene ontology enrichment; Table S13: Pp's genome-metabolome extracted gene ontology enrichment; Table S14: AIB operon Rhodococcus wratislaviensis and relative homologous genes in X. fastidiosa and P. phytofirmans; Table S15: GA operon in Xanthomonas oryzae pv oryzicola and relative homologous genes in X. fastidiosa and P. phytofirmans.
Author Contributions
Conceptualization: A.M.d.S., A.L., O.R.F.-J. Methodology: O.R.F.-J, A.L., D.B., J.M-J, S.M.K., C.B., B.P.B. Computing resources: D.B., J.M-J, B.P.B. Data curation: O.R.F.-J, A.L., D.B., J.M-J, S.M.K., B.P.B. Formal analysis: O.R.F.-J., A.L., P.A.Z., S.K., B.P.B. Visualization: O.R.F.-J, P.A.Z, A.M.d.S. Writing—original draft preparation: O.R.F.-J., P.A.Z. Writing—review and editing: O.R.F.-J, P.A.Z, A.M.d.S. Supervision: A.M.d.S, T.R.N., S.E.L. Funding acquisition: A.M.d.S, T.R.N., and S.E.L. All authors read, provided critical review, and approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by São Paulo Research Foundation (FAPESP), grant number 08/11703-4, and by Coordination for the Improvement of Higher Education Personnel (CAPES), by the Program Sandwich-Doctorate Abroad (PDSE), grant number 99999.009713/2014-00. D.B. received a fellowship from CAPES (88887.285065/2018-00 and 88887.318125/2019-00). A.M.d.S. received a research fellowship award 309182/2016-6 from the National Council for Scientific and Technological Development (CNPq). A.L and S.M.K were supported by the Office of Science, Office of Biological and Environmental Research, of the US Department of Energy, Award No. DE-SC0012627. T.R.N. and B.P.B. were partially supported by the former award and the U.S. Department of Energy Joint Genome Institute (https://ror.org/04xm1d337), a DOE Office of Science User Facility, is supported by the Office of Science of the U.S. Department of Energy operated under Contract No. DE-AC02-05CH11231.
Data Availability Statement
X. fastidiosa and P. phytofirmans genomic sequences were accessed from the GenBank RefSeq database at NCBI (National Center for Biotechnology Information).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Sicard, A.; Zeilinger, A. R.; Vanhove, M.; Schartel, T. E.; Beal, D. J.; Daugherty, M. P.; Almeida, R. P. P., Xylella fastidiosa: Insights into an Emerging Plant Pathogen. Annual Review of Phytopathology, Vol 56 2018, 56, 181-202.
- Chatterjee, S.; Almeida, R. P.; Lindow, S., Living in two worlds: the plant and insect lifestyles of Xylella fastidiosa. Annu Rev Phytopathol 2008, 46, 243-71. [CrossRef]
- Wang, N.; Li, J. L.; Lindow, S. E., RpfF-dependent regulon of Xylella fastidiosa. Phytopathology 2012, 102, (11), 1045-53. [CrossRef]
- Ionescu, M.; Yokota, K.; Antonova, E.; Garcia, A.; Beaulieu, E.; Hayes, T.; Iavarone, A. T.; Lindow, S. E., Promiscuous Diffusible Signal Factor Production and Responsiveness of the Xylella fastidiosa Rpf System. MBio 2016, 7, (4).
- Roper, C.; Castro, C.; Ingel, B., Xylella fastidiosa: bacterial parasitism with hallmarks of commensalism. Curr Opin Plant Biol 2019, 50, 140-147. [CrossRef]
- Rapicavoli, J.; Ingel, B.; Blanco-Ulate, B.; Cantu, D.; Roper, C., Xylella fastidiosa: an examination of a re-emerging plant pathogen. Mol Plant Pathol 2018, 19, (4), 786-800. [CrossRef]
- Morris, C. E.; Moury, B., Revisiting the Concept of Host Range of Plant Pathogens. Annual Review of Phytopathology, Vol 57, 2019 2019, 57, 63-90.
- Parniske, M., Uptake of bacteria into living plant cells, the unifying and distinct feature of the nitrogen-fixing root nodule symbiosis. Curr Opin Plant Biol 2018, 44, 164-174. [CrossRef]
- Nagel, R.; Turrini, P. C.; Nett, R. S.; Leach, J. E.; Verdier, V.; Van Sluys, M. A.; Peters, R. J., An operon for production of bioactive gibberellin A(4) phytohormone with wide distribution in the bacterial rice leaf streak pathogen Xanthomonas oryzae pv. oryzicola. New Phytol 2017, 214, (3), 1260-1266. [CrossRef]
- Ankrah, N. Y. D.; Wilkes, R. A.; Zhang, F. Q.; Aristilde, L.; Douglas, A. E., The Metabolome of Associations between Xylem-Feeding Insects and their Bacterial Symbionts. Journal of Chemical Ecology 2020, 46, (8), 735-744. [CrossRef]
- Yang, J.; Masoudi, A.; Li, H.; Gu, Y.; Wang, C.; Wang, M.; Yu, Z.; Liu, J., Microbial community structure and niche differentiation under different health statuses of Pinus bungeana in the Xiong'an New Area in China. Front Microbiol 2022, 13, 913349. [CrossRef]
- De Silva, N. I.; Brooks, S.; Lumyong, S.; Hyde, K. D., Use of endophytes as biocontrol agents. Fungal Biol Rev 2019, 33, (2), 133-148. [CrossRef]
- Saldanha, L. L.; Allard, P. M.; Dilarri, G.; Codesido, S.; Gonzalez-Ruiz, V.; Queiroz, E. F.; Ferreira, H.; Wolfender, J. L., Metabolomic- and Molecular Networking-Based Exploration of the Chemical Responses Induced in Citrus sinensis Leaves Inoculated with Xanthomonas citri. J Agr Food Chem 2022. [CrossRef]
- Ryffel, F.; Helfrich, E. J. N.; Kiefer, P.; Peyriga, L.; Portais, J. C.; Piel, J.; Vorholt, J. A., Metabolic footprint of epiphytic bacteria on Arabidopsis thaliana leaves. Isme J 2016, 10, (3), 632-643. [CrossRef]
- Zaini, P. A.; Nascimento, R.; Gouran, H.; Cantu, D.; Chakraborty, S.; Phu, M.; Goulart, L. R.; Dandekar, A. M., Molecular Profiling of Pierce's Disease Outlines the Response Circuitry of Vitis vinifera to Xylella fastidiosa Infection. Front Plant Sci 2018, 9, 771. [CrossRef]
- Chen, X. L.; Sun, M. C.; Chong, S. L.; Si, J. P.; Wu, L. S., Transcriptomic and Metabolomic Approaches Deepen Our Knowledge of Plant-Endophyte Interactions. Frontiers in Plant Science 2022, 12. [CrossRef]
- Cariddi, C.; Saponari, M.; Boscia, D.; De Stradis, A.; Loconsole, G.; Nigro, F.; Porcelli, F.; Potere, O.; Martelli, G. P., Isolation of a Xylella fastidiosa strain infecting olive and oleander in Apulia, Italy. J Plant Pathol 2014, 96, (2), 425-429.
- Desprez-Loustau, M.-L.; Balci, Y.; Cornara, D.; Gonthier, P.; Robin, C.; Jacques, M.-A., Is Xylella fastidiosa a serious threat to European forests? Forestry: An International Journal of Forest Research 2020, 94, (1), 1-17.
- Krugner, R.; Sisterson, M. S.; Backus, E. A.; Burbank, L. P.; Redak, R. A., Sharpshooters: a review of what moves Xylella fastidiosa. Austral Entomol 2019, 58, (2), 248-267. [CrossRef]
- Saponari, M.; Loconsole, G.; Cornara, D.; Yokomi, R. K.; De Stradis, A.; Boscia, D.; Bosco, D.; Martelli, G. P.; Krugner, R.; Porcelli, F., Infectivity and transmission of Xylellua fastidiosa by Philaenus spumarius (Hemiptera: Aphrophoridae) in Apulia, Italy. J Econ Entomol 2014, 107, (4), 1316-9. [CrossRef]
- Huang, W. J.; Reyes-Caldas, P.; Mann, M.; Seifbarghi, S.; Kahn, A.; Almeida, R. P. P.; Béven, L.; Heck, M.; Hogenhout, S. A.; Coaker, G., Bacterial Vector-Borne Plant Diseases: Unanswered Questions and Future Directions. Molecular Plant 2020, 13, (10), 1379-1393. [CrossRef]
- Roper, M. C.; Greve, L. C.; Warren, J. G.; Labavitch, J. M.; Kirkpatrick, B. C., Xylella fastidiosa requires polygalacturonase for colonization and pathogenicity in Vitis vinifera grapevines. Molecular Plant-Microbe Interactions 2007, 20, (4), 411-419.
- Nascimento, R.; Gouran, H.; Chakraborty, S.; Gillespie, H. W.; Almeida-Souza, H. O.; Tu, A.; Rao, B. J.; Feldstein, P. A.; Bruening, G.; Goulart, L. R.; Dandekar, A. M., The Type II Secreted Lipase/Esterase LesA is a Key Virulence Factor Required for Xylella fastidiosa Pathogenesis in Grapevines (vol 6, 18598, 2016). Sci Rep-Uk 2016, 6.
- Feitosa, O. R.; Stefanello, E.; Zaini, P. A.; Nascimento, R.; Pierry, P. M.; Dandekar, A. M.; Lindow, S. E.; da Silva, A. M., Proteomic and Metabolomic Analyses of Xylella fastidiosa OMV-Enriched Fractions Reveal Association with Virulence Factors and Signaling Molecules of the DSF Family. Phytopathology 2019, 109, (8), 1344-1353. [CrossRef]
- Block, A.; Li, G. Y.; Fu, Z. Q.; Alfano, J. R., Phytopathogen type III effector weaponry and their plant targets. Current Opinion in Plant Biology 2008, 11, (4), 396-403. [CrossRef]
- Kvitko, B. H.; Collmer, A., Discovery of the Hrp Type III Secretion System in Phytopathogenic Bacteria: How Investigation of Hypersensitive Cell Death in Plants Led to a Novel Protein Injector System and a World of Inter-Organismal Molecular Interactions Within Plant Cells. Phytopathology 2023. [CrossRef]
- Van Sluys, M. A.; Monteiro-Vitorello, C. B.; Camargo, L. E.; Menck, C. F.; Da Silva, A. C.; Ferro, J. A.; Oliveira, M. C.; Setubal, J. C.; Kitajima, J. P.; Simpson, A. J., Comparative genomic analysis of plant-associated bacteria. Annu Rev Phytopathol 2002, 40, 169-89. [CrossRef]
- De La Fuente, L.; Merfa, M. V.; Cobine, P. A.; Coleman, J. J., Pathogen Adaptation to the Xylem Environment. Annu Rev Phytopathol 2022, 60, 163-186. [CrossRef] [PubMed]
- Sawana, A.; Adeolu, M.; Gupta, R. S., Molecular signatures and phylogenomic analysis of the genus Burkholderia: proposal for division of this genus into the emended genus Burkholderia containing pathogenic organisms and a new genus Paraburkholderia gen. nov. harboring environmental species. Front Genet 2014, 5, 429. [CrossRef] [PubMed]
- Sessitsch, A.; Coenye, T.; Sturz, A. V.; Vandamme, P.; Barka, E. A.; Salles, J. F.; Van Elsas, J. D.; Faure, D.; Reiter, B.; Glick, B. R.; Wang-Pruski, G.; Nowak, J., Burkholderia phytofirmans sp. nov., a novel plant-associated bacterium with plant-beneficial properties. International journal of systematic and evolutionary microbiology 2005, 55, 1187-1192.
- Mitter, B.; Petric, A.; Shin, M. W.; Chain, P. S.; Hauberg-Lotte, L.; Reinhold-Hurek, B.; Nowak, J.; Sessitsch, A., Comparative genome analysis of Burkholderia phytofirmans PsJN reveals a wide spectrum of endophytic lifestyles based on interaction strategies with host plants. Front Plant Sci 2013, 4, 120. [CrossRef] [PubMed]
- Miotto-Vilanova, L.; Jacquard, C.; Courteaux, B.; Wortham, L.; Michel, J.; Clement, C.; Barka, E. A.; Sanchez, L., Burkholderia phytofirmans PsJN Confers Grapevine Resistance against Botrytis cinerea via a Direct Antimicrobial Effect Combined with a Better Resource Mobilization. Front Plant Sci 2016, 7, 1236. [CrossRef] [PubMed]
- Baccari, C.; Antonova, E.; Lindow, S., Biological Control of Pierce's Disease of Grape by an Endophytic Bacterium. Phytopathology 2019, 109, (2), 248-256. [CrossRef] [PubMed]
- Lindow, S.; Koutsoukis, R.; Meyer, K. M.; Baccari, C., Control of Pierce's disease of grape with Paraburkholderia phytofirmans PsJN in the field. Phytopathology 2023. [CrossRef] [PubMed]
- Sue, T.; Obolonkin, V.; Griffiths, H.; Villas-Boas, S. G., An Exometabolomics Approach to Monitoring Microbial Contamination in Microalgal Fermentation Processes by Using Metabolic Footprint Analysis. Appl Environ Microb 2011, 77, (21), 7605-7610. [CrossRef]
- Drenos, F., Mechanistic insights from combining genomics with metabolomics. Curr Opin Lipidol 2017, 28, (2), 99-103. [CrossRef]
- Liu, R.; Bao, Z. X.; Zhao, P. J.; Li, G. H., Advances in the Study of Metabolomics and Metabolites in Some Species Interactions. Molecules 2021, 26, (11).
- Villas-Boas, S. G.; Noel, S.; Lane, G. A.; Attwood, G.; Cookson, A., Extracellular metabolomics: a metabolic footprinting approach to assess fiber degradation in complex media. Analytical biochemistry 2006, 349, (2), 297-305. [CrossRef]
- Villas-Boas, S. G.; Mas, S.; Akesson, M.; Smedsgaard, J.; Nielsen, J., Mass spectrometry in metabolome analysis. Mass Spectrom Rev 2005, 24, (5), 613-46. [CrossRef] [PubMed]
- Newman, K. L.; Almeida, R. P.; Purcell, A. H.; Lindow, S. E., Use of a green fluorescent strain for analysis of Xylella fastidiosa colonization of Vitis vinifera. Appl Environ Microbiol 2003, 69, (12), 7319-27. [CrossRef] [PubMed]
- Newman, K. L.; Almeida, R. P. P.; Purcell, A. H.; Lindow, S. E., Cell-cell signaling controls Xylella fastidiosa interactions with both insects and plants. Proceedings of the National Academy of Sciences of the United States of America 2004, 101, (6), 1737-1742. [CrossRef] [PubMed]
- Frommel, M. I.; Nowak, J.; Lazarovits, G., Growth Enhancement and Developmental Modifications of in Vitro Grown Potato (Solanum tuberosum spp. tuberosum) as Affected by a Nonfluorescent Pseudomonas sp. Plant Physiol 1991, 96, (3), 928-36.
- Weilharter, A.; Mitter, B.; Shin, M. V.; Chain, P. S. G.; Nowak, J.; Sessitsch, A., Complete Genome Sequence of the Plant Growth-Promoting Endophyte Burkholderia phytofirmans Strain PsJN. J Bacteriol 2011, 193, (13), 3383-3384. [CrossRef]
- King, E. O.; Ward, M. K.; Raney, D. E., Two simple media for the demonstration of pyocyanin and fluorescin. J Lab Clin Med 1954, 44, (2), 301-7.
- Zaini, P. A.; De La Fuente, L.; Hoch, H. C.; Burr, T. J., Grapevine xylem sap enhances biofilm development by Xylella fastidiosa. FEMS microbiology letters 2009, 295, (1), 129-134. [CrossRef] [PubMed]
- Bowen, B. P.; Northen, T. R., Dealing with the unknown: metabolomics and metabolite atlases. J Am Soc Mass Spectrom 2010, 21, (9), 1471-6. [CrossRef] [PubMed]
- Yao, Y.; Sun, T.; Wang, T.; Ruebel, O.; Northen, T.; Bowen, B. P., Analysis of Metabolomics Datasets with High-Performance Computing and Metabolite Atlases. Metabolites 2015, 5, (3), 431-42. [CrossRef] [PubMed]
- Sumner, L. W.; Amberg, A.; Barrett, D.; Beale, M. H.; Beger, R.; Daykin, C. A.; Fan, T. W. M.; Fiehn, O.; Goodacre, R.; Griffin, J. L.; Hankemeier, T.; Hardy, N.; Harnly, J.; Higashi, R.; Kopka, J.; Lane, A. N.; Lindon, J. C.; Marriott, P.; Nicholls, A. W.; Reily, M. D.; Thaden, J. J.; Viant, M. R., Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, (3), 211-221. [CrossRef]
- Gower, J. C., Some Distance Properties of Latent Root and Vector Methods Used in Multivariate Analysis. Biometrika 1966, 53, 325-&. [CrossRef]
- Erbilgin, O.; Rubel, O.; Louie, K. B.; Trinh, M.; Raad, M.; Wildish, T.; Udwary, D.; Hoover, C.; Deutsch, S.; Northen, T. R.; Bowen, B. P., MAGI: A Method for Metabolite Annotation and Gene Integration. ACS Chem Biol 2019, 14, (4), 704-714. [CrossRef] [PubMed]
- Merkel, D., Docker: lightweight linux containers for consistent development and deployment. Linux journal 2014, 2014, 2.
- Gotz, S.; Garcia-Gomez, J. M.; Terol, J.; Williams, T. D.; Nagaraj, S. H.; Nueda, M. J.; Robles, M.; Talon, M.; Dopazo, J.; Conesa, A., High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res 2008, 36, (10), 3420-3435. [CrossRef] [PubMed]
- Grigoriev, I. V.; Nordberg, H.; Shabalov, I.; Aerts, A.; Cantor, M.; Goodstein, D.; Kuo, A.; Minovitsky, S.; Nikitin, R.; Ohm, R. A.; Otillar, R.; Poliakov, A.; Ratnere, I.; Riley, R.; Smirnova, T.; Rokhsar, D.; Dubchak, I., The genome portal of the Department of Energy Joint Genome Institute. Nucleic Acids Res 2012, 40, (Database issue), D26-32. [CrossRef]
- Delcher, A. L.; Phillippy, A.; Carlton, J.; Salzberg, S. L., Fast algorithms for large-scale genome alignment and comparison. Nucleic Acids Res 2002, 30, (11), 2478-2483. [CrossRef] [PubMed]
- Michelini, S.; Balakrishnan, B.; Parolo, S.; Matone, A.; Mullaney, J. A.; Young, W.; Gasser, O.; Wall, C.; Priami, C.; Lombardo, R.; Kussmann, M., A reverse metabolic approach to weaning: in silico identification of immune-beneficial infant gut bacteria, mining their metabolism for prebiotic feeds and sourcing these feeds in the natural product space. Microbiome 2018, 6, (1), 171. [CrossRef]
- Levy, R.; Borenstein, E., Reverse Ecology: from systems to environments and back. Adv Exp Med Biol 2012, 751, 329-45. [PubMed]
- Carr, R.; Borenstein, E., NetSeed: a network-based reverse-ecology tool for calculating the metabolic interface of an organism with its environment. Bioinformatics 2012, 28, (5), 734-5. [CrossRef]
- Levy, R.; Borenstein, E., Metabolic modeling of species interaction in the human microbiome elucidates community-level assembly rules. Proc Natl Acad Sci U S A 2013, 110, (31), 12804-9. [CrossRef]
- Kreimer, A.; Doron-Faigenboim, A.; Borenstein, E.; Freilich, S., NetCmpt: a network-based tool for calculating the metabolic competition between bacterial species. Bioinformatics 2012, 28, (16), 2195-7. [CrossRef]
- Kim, S.; Chen, J.; Cheng, T. J.; Gindulyte, A.; He, J.; He, S. Q.; Li, Q. L.; Shoemaker, B. A.; Thiessen, P. A.; Yu, B.; Zaslavsky, L.; Zhang, J.; Bolton, E. E., PubChem 2023 update. Nucleic Acids Res 2022. [CrossRef]
- Bennett, G. M.; Moran, N. A., Heritable symbiosis: The advantages and perils of an evolutionary rabbit hole. Proc Natl Acad Sci U S A 2015, 112, (33), 10169-76. [CrossRef]
- Flynn, K. J.; Dickson, D. M. J.; Al-Amoudi, O. A., The ratio of glutamine:glutamate in microalgae: a biomarker for N-status suitable for use at natural cell densities. Journal of Plankton Research 1989, 11, (1), 165-170. [CrossRef]
- Hibi, M.; Fukuda, D.; Kenchu, C.; Nojiri, M.; Hara, R.; Takeuchi, M.; Aburaya, S.; Aoki, W.; Mizutani, K.; Yasohara, Y.; Ueda, M.; Mikami, B.; Takahashi, S.; Ogawa, J., A three-component monooxygenase from Rhodococcus wratislaviensis may expand industrial applications of bacterial enzymes. Commun Biol 2021, 4, (1).
- Salazar-Cerezo, S.; Martinez-Montiel, N.; Garcia-Sanchez, J.; Perez, Y. T. R.; Martinez-Contreras, R. D., Gibberellin biosynthesis and metabolism: A convergent route for plants, fungi and bacteria. Microbiol Res 2018, 208, 85-98. [CrossRef] [PubMed]
- Bansal, P.; Morgat, A.; Axelsen, K. B.; Muthukrishnan, V.; Coudert, E.; Aimo, L.; Hyka-Nouspikel, N.; Gasteiger, E.; Kerhornou, A.; Neto, T. B.; Pozzato, M.; Blatter, M. C.; Ignatchenko, A.; Redaschi, N.; Bridge, A., Rhea, the reaction knowledgebase in 2022. Nucleic Acids Res 2022, 50, (D1), D693-D700. [CrossRef]
- Caspi, R.; Billington, R.; Keseler, I. M.; Kothari, A.; Krummenacker, M.; Midford, P. E.; Ong, W. K.; Paley, S.; Subhraveti, P.; Karp, P. D., The MetaCyc database of metabolic pathways and enzymes - a 2019 update. Nucleic Acids Res 2020, 48, (D1), D445-D453. [CrossRef]
- Ashburner, M.; Ball, C. A.; Blake, J. A.; Botstein, D.; Butler, H.; Cherry, J. M.; Davis, A. P.; Dolinski, K.; Dwight, S. S.; Eppig, J. T.; Harris, M. A.; Hill, D. P.; Issel-Tarver, L.; Kasarskis, A.; Lewis, S.; Matese, J. C.; Richardson, J. E.; Ringwald, M.; Rubin, G. M.; Sherlock, G.; Consortium, G. O., Gene Ontology: tool for the unification of biology. Nat Genet 2000, 25, (1), 25-29. [CrossRef] [PubMed]
- Gene Ontology, C.; Aleksander, S. A.; Balhoff, J.; Carbon, S.; Cherry, J. M.; Drabkin, H. J.; Ebert, D.; Feuermann, M.; Gaudet, P.; Harris, N. L.; Hill, D. P.; Lee, R.; Mi, H.; Moxon, S.; Mungall, C. J.; Muruganugan, A.; Mushayahama, T.; Sternberg, P. W.; Thomas, P. D.; Van Auken, K.; Ramsey, J.; Siegele, D. A.; Chisholm, R. L.; Fey, P.; Aspromonte, M. C.; Nugnes, M. V.; Quaglia, F.; Tosatto, S.; Giglio, M.; Nadendla, S.; Antonazzo, G.; Attrill, H.; Dos Santos, G.; Marygold, S.; Strelets, V.; Tabone, C. J.; Thurmond, J.; Zhou, P.; Ahmed, S. H.; Asanitthong, P.; Luna Buitrago, D.; Erdol, M. N.; Gage, M. C.; Ali Kadhum, M.; Li, K. Y. C.; Long, M.; Michalak, A.; Pesala, A.; Pritazahra, A.; Saverimuttu, S. C. C.; Su, R.; Thurlow, K. E.; Lovering, R. C.; Logie, C.; Oliferenko, S.; Blake, J.; Christie, K.; Corbani, L.; Dolan, M. E.; Drabkin, H. J.; Hill, D. P.; Ni, L.; Sitnikov, D.; Smith, C.; Cuzick, A.; Seager, J.; Cooper, L.; Elser, J.; Jaiswal, P.; Gupta, P.; Jaiswal, P.; Naithani, S.; Lera-Ramirez, M.; Rutherford, K.; Wood, V.; De Pons, J. L.; Dwinell, M. R.; Hayman, G. T.; Kaldunski, M. L.; Kwitek, A. E.; Laulederkind, S. J. F.; Tutaj, M. A.; Vedi, M.; Wang, S. J.; D'Eustachio, P.; Aimo, L.; Axelsen, K.; Bridge, A.; Hyka-Nouspikel, N.; Morgat, A.; Aleksander, S. A.; Cherry, J. M.; Engel, S. R.; Karra, K.; Miyasato, S. R.; Nash, R. S.; Skrzypek, M. S.; Weng, S.; Wong, E. D.; Bakker, E.; Berardini, T. Z.; Reiser, L.; Auchincloss, A.; Axelsen, K.; Argoud-Puy, G.; Blatter, M. C.; Boutet, E.; Breuza, L.; Bridge, A.; Casals-Casas, C.; Coudert, E.; Estreicher, A.; Livia Famiglietti, M.; Feuermann, M.; Gos, A.; Gruaz-Gumowski, N.; Hulo, C.; Hyka-Nouspikel, N.; Jungo, F.; Le Mercier, P.; Lieberherr, D.; Masson, P.; Morgat, A.; Pedruzzi, I.; Pourcel, L.; Poux, S.; Rivoire, C.; Sundaram, S.; Bateman, A.; Bowler-Barnett, E.; Bye, A. J. H.; Denny, P.; Ignatchenko, A.; Ishtiaq, R.; Lock, A.; Lussi, Y.; Magrane, M.; Martin, M. J.; Orchard, S.; Raposo, P.; Speretta, E.; Tyagi, N.; Warner, K.; Zaru, R.; Diehl, A. D.; Lee, R.; Chan, J.; Diamantakis, S.; Raciti, D.; Zarowiecki, M.; Fisher, M.; James-Zorn, C.; Ponferrada, V.; Zorn, A.; Ramachandran, S.; Ruzicka, L.; Westerfield, M., The Gene Ontology knowledgebase in 2023. Genetics 2023, 224, (1).
- Ionescu, M.; Zaini, P. A.; Baccari, C.; Tran, S.; da Silva, A. M.; Lindow, S. E., Xylella fastidiosa outer membrane vesicles modulate plant colonization by blocking attachment to surfaces. Proceedings of the National Academy of Sciences of the United States of America 2014, 111, (37), E3910-8. [CrossRef] [PubMed]
- Smolka, M. B.; Martins, D.; Winck, F. V.; Santoro, C. E.; Castellari, R. R.; Ferrari, F.; Brum, I. J.; Galembeck, E.; Coletta, H. D.; Machado, M. A.; Marangoni, S.; Novello, J. C., Proteome analysis of the plant pathogen Xylella fastidiosa reveals major cellular and extracellular proteins and a peculiar codon bias distribution. Proteomics 2003, 3, (2), 224-237. [CrossRef]
- Jacoby, R. P.; Martyn, A.; Kopriva, S., Exometabolomic Profiling of Bacterial Strains as Cultivated Using Arabidopsis Root Extract as the Sole Carbon Source. Molecular Plant-Microbe Interactions 2018, 31, (8), 803-813.
- Louie, K. B.; Bowen, B. P.; Cheng, X.; Berleman, J. E.; Chakraborty, R.; Deutschbauer, A.; Arkin, A.; Northen, T. R., "Replica-extraction-transfer" nanostructure-initiator mass spectrometry imaging of acoustically printed bacteria. Anal Chem 2013, 85, (22), 10856-62. [CrossRef]
- Daugherty, M. P.; Rashed, A.; Almeida, R. P. P.; Perring, T. M., Vector preference for hosts differing in infection status: sharpshooter movement and Xylella fastidiosa transmission. Ecol Entomol 2011, 36, (5), 654-662. [CrossRef]
- Douglas, A. E., The B vitamin nutrition of insects: the contributions of diet, microbiome and horizontally acquired genes. Curr Opin Insect Sci 2017, 23, 65-69. [CrossRef] [PubMed]
- Wang, H.; Zhi, W.; Qu, H.; Lin, H.; Jiang, Y., Application of alpha-aminoisobutyric acid and beta-aminoisobutyric acid inhibits pericarp browning of harvested longan fruit. Chem Cent J 2015, 9, (1), 54. [CrossRef] [PubMed]
- Phinney, B. O., Growth Response of Single-Gene Dwarf Mutants in Maize to Gibberellic Acid. Proceedings of the National Academy of Sciences of the United States of America 1956, 42, (4), 185-189. [CrossRef] [PubMed]
- Lu, X.; Hershey, D. M.; Wang, L.; Bogdanove, A. J.; Peters, R. J., An ent-kaurene-derived diterpenoid virulence factor from Xanthomonas oryzae pv. oryzicola. New Phytologist 2015, 206, (1), 295-302. [CrossRef] [PubMed]
- Vergine, M.; Nicolì, F.; Sabella, E.; Aprile, A.; De Bellis, L.; Luvisi, A., Secondary Metabolites in Plant Interaction. Pathogens 2020, 9, (9).
- Novelli, S.; Gismondi, A.; Di Marco, G.; Canuti, L.; Nanni, V.; Canini, A., Plant defense factors involved in Olea europaea resistance against Xylella fastidiosa infection. J Plant Res 2019, 132, (3), 439-455. [CrossRef] [PubMed]
- Joo, G. J.; Kang, S. M.; Hamayun, M.; Kim, S. K.; Na, C. I.; Shin, D. H.; Lee, I. J., Burkholderia sp KCTC 11096BP as a newly isolated gibberellin producing bacterium. Journal of Microbiology 2009, 47, (2), 167-171. [CrossRef]
- Azevedo, J. L.; Araujo, W. L.; Lacava, P. T., The diversity of citrus endophytic bacteria and their interactions with Xylella fastidiosa and host plants. Genetics and Molecular Biology 2016, 39, (4), 476-491. [CrossRef]
- Caserta, R.; Souza-Neto, R. R.; Takita, M. A.; Lindow, S.; Souza, A., Ectopic expression of Xylella fastidiosa rpfF conferring production of diffusible signal factor in transgenic tobacco and citrus alters pathogen behavior and reduces disease severity. Molecular plant-microbe interactions : MPMI 2017.
Figure 1.
Biofilm increase in X. fastidiosa in response to exposure of P. phytofirmans secretome. A. Temecula1 WT or ΔrpfF strains were grown in PD3 medium for 7 days with (Xf sm and ΔrpfFsm) or without (Xf and ΔrpfF) 20% v/v of a 1-day PD3 medium P. phytofirmans (Pp) spent media. A. Glass tube image of the stained air-liquid interface of biofilm formation. B. Biofilm quantification was performed by measurement of crystal violet staining (Abs595nm). Arrows indicate biofilm ring in the tubes. Error bars indicate the standard error of triplicate assays. Significantly different biofilm quantification was calculated using a t-test (ns: p ≤ 1.0×100, *: 1.0×10-2 < p ≤ 5.0×10-2, **: 1.0×10-3 < p ≤ 1.0×10-2).
Figure 1.
Biofilm increase in X. fastidiosa in response to exposure of P. phytofirmans secretome. A. Temecula1 WT or ΔrpfF strains were grown in PD3 medium for 7 days with (Xf sm and ΔrpfFsm) or without (Xf and ΔrpfF) 20% v/v of a 1-day PD3 medium P. phytofirmans (Pp) spent media. A. Glass tube image of the stained air-liquid interface of biofilm formation. B. Biofilm quantification was performed by measurement of crystal violet staining (Abs595nm). Arrows indicate biofilm ring in the tubes. Error bars indicate the standard error of triplicate assays. Significantly different biofilm quantification was calculated using a t-test (ns: p ≤ 1.0×100, *: 1.0×10-2 < p ≤ 5.0×10-2, **: 1.0×10-3 < p ≤ 1.0×10-2).
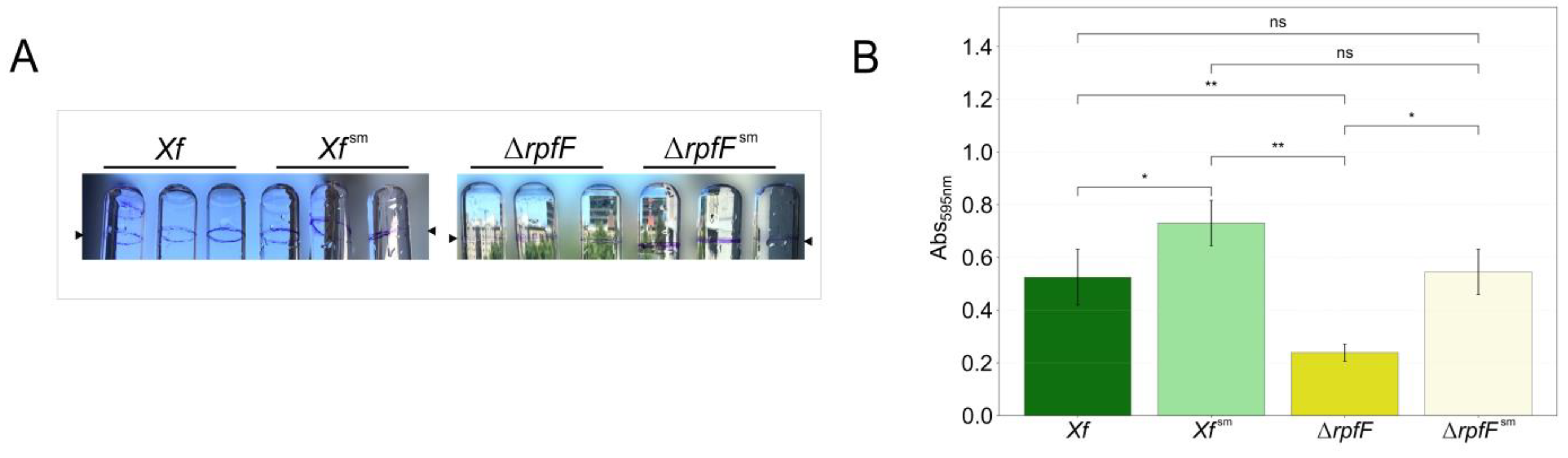
Figure 2.
X. fastidiosa and P. phytofirmans exometabolomes. A. Venn Diagram of Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes. Shared or exclusive metabolites detected in the Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes are displayed. B. Principal Coordinate Analysis (PCoA) of Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes. Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp were cultured in PD3 medium for 7 days without (dark yellow and dark green circles) or with (light yellow and light green circles) 20% supernatant of P. phytofirmans PsJN; Pp was cultured in PD3 medium for 1 day (red circles). Each circle represents three technical replicates with 116 putative-identified metabolites. Permanova and Permidisp statistical analysis with 999 permutations were performed, the statistical significances are shown in the figure. C. Heatmap representing the abundance of metabolites in the Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes. Each one of the 116 compounds detected in this study is indicated in the columns, each respective strain is indicated in the rows. The color code highlights the abundance of the metabolites in the conditions tested, ranging from low (blue), and medium (white) to high (red) expression. An additional column was added with the respective compound class as stated in PubChem. Two main clusters were highlighted, in green and red boxes, as they contain the compounds highly expressed in X. fastidiosa and P. phytofirmans, respectively. A z-score normalization was performed for each compound across the different conditions. Three biological replicates of each species and treatments were used for creating this heatmap. Compounds in black were confirmed by standard run comparison (regarding RT and m/z ppm), compounds in orange have a lower level of confirmation (either or both RT and m/z ppm), and compounds in red could not be confirmed by comparison RT. Low confidence compounds were therefore considered as putative (possibly tautomer of the compound assigned). (*) indicates ANOVA statistical significance across conditions.
Figure 2.
X. fastidiosa and P. phytofirmans exometabolomes. A. Venn Diagram of Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes. Shared or exclusive metabolites detected in the Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes are displayed. B. Principal Coordinate Analysis (PCoA) of Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes. Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp were cultured in PD3 medium for 7 days without (dark yellow and dark green circles) or with (light yellow and light green circles) 20% supernatant of P. phytofirmans PsJN; Pp was cultured in PD3 medium for 1 day (red circles). Each circle represents three technical replicates with 116 putative-identified metabolites. Permanova and Permidisp statistical analysis with 999 permutations were performed, the statistical significances are shown in the figure. C. Heatmap representing the abundance of metabolites in the Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes. Each one of the 116 compounds detected in this study is indicated in the columns, each respective strain is indicated in the rows. The color code highlights the abundance of the metabolites in the conditions tested, ranging from low (blue), and medium (white) to high (red) expression. An additional column was added with the respective compound class as stated in PubChem. Two main clusters were highlighted, in green and red boxes, as they contain the compounds highly expressed in X. fastidiosa and P. phytofirmans, respectively. A z-score normalization was performed for each compound across the different conditions. Three biological replicates of each species and treatments were used for creating this heatmap. Compounds in black were confirmed by standard run comparison (regarding RT and m/z ppm), compounds in orange have a lower level of confirmation (either or both RT and m/z ppm), and compounds in red could not be confirmed by comparison RT. Low confidence compounds were therefore considered as putative (possibly tautomer of the compound assigned). (*) indicates ANOVA statistical significance across conditions.
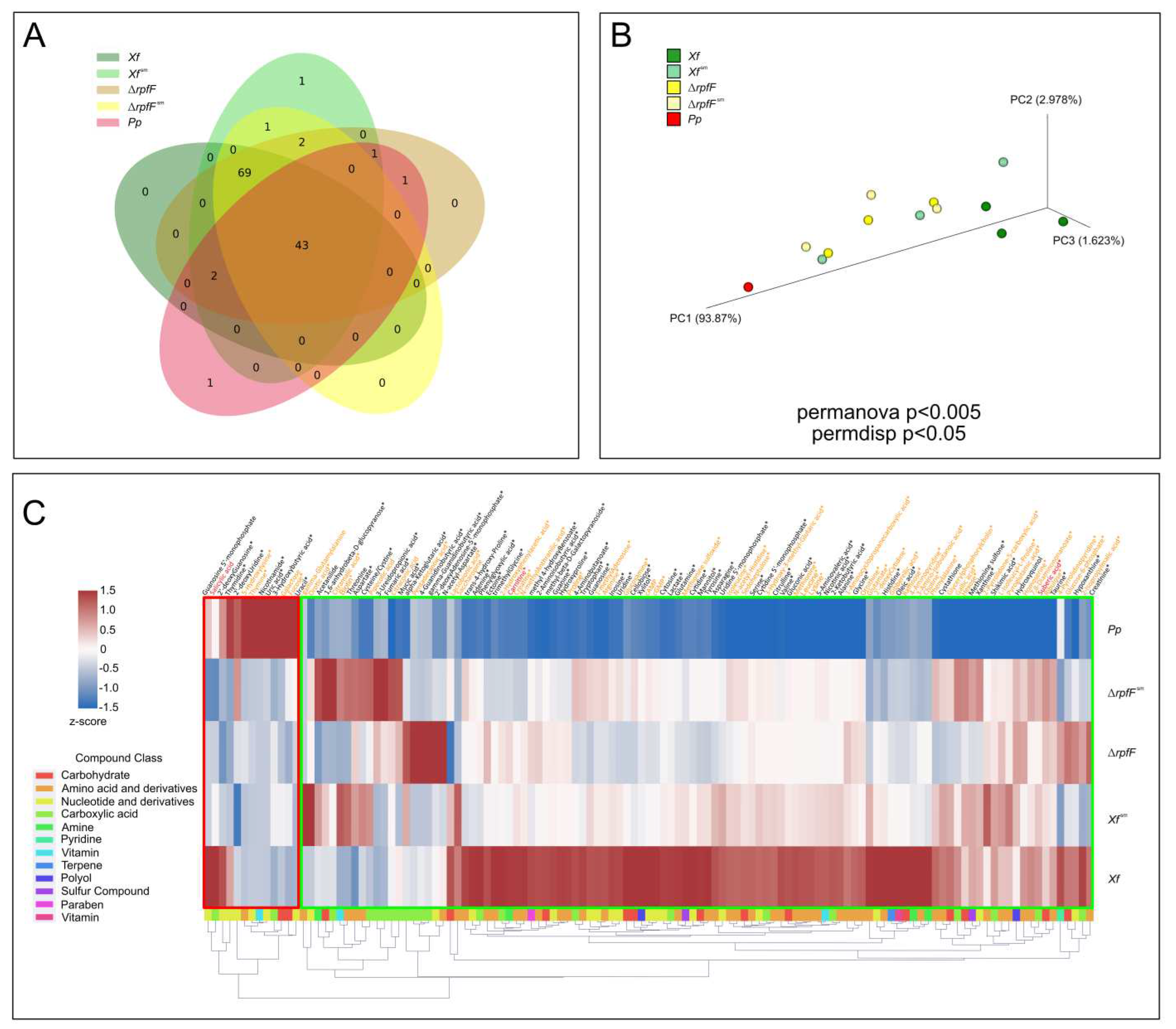
Figure 3.
Nitrogen status of Xf, ΔrpfF, Xfsm, ΔrpfFsm, and Pp exometabolomes. A. glutamine:glutamate ratio. Average expression of Glutamine and Glutamate, from 3 biological replicates and 3 technical replicates, were used to calculate the gln:glu ratio. Dotted line indicates the “0.5”, a ratio indicating cellular N-replete. B. EAA and C. nEAA EICs of representative assessments in blank (MeOH), PD3, PD3sm, Xf, ΔrpfF, Xf sm, ΔrpfFsm, and Pp extracts or exometabolomes.
Figure 3.
Nitrogen status of Xf, ΔrpfF, Xfsm, ΔrpfFsm, and Pp exometabolomes. A. glutamine:glutamate ratio. Average expression of Glutamine and Glutamate, from 3 biological replicates and 3 technical replicates, were used to calculate the gln:glu ratio. Dotted line indicates the “0.5”, a ratio indicating cellular N-replete. B. EAA and C. nEAA EICs of representative assessments in blank (MeOH), PD3, PD3sm, Xf, ΔrpfF, Xf sm, ΔrpfFsm, and Pp extracts or exometabolomes.
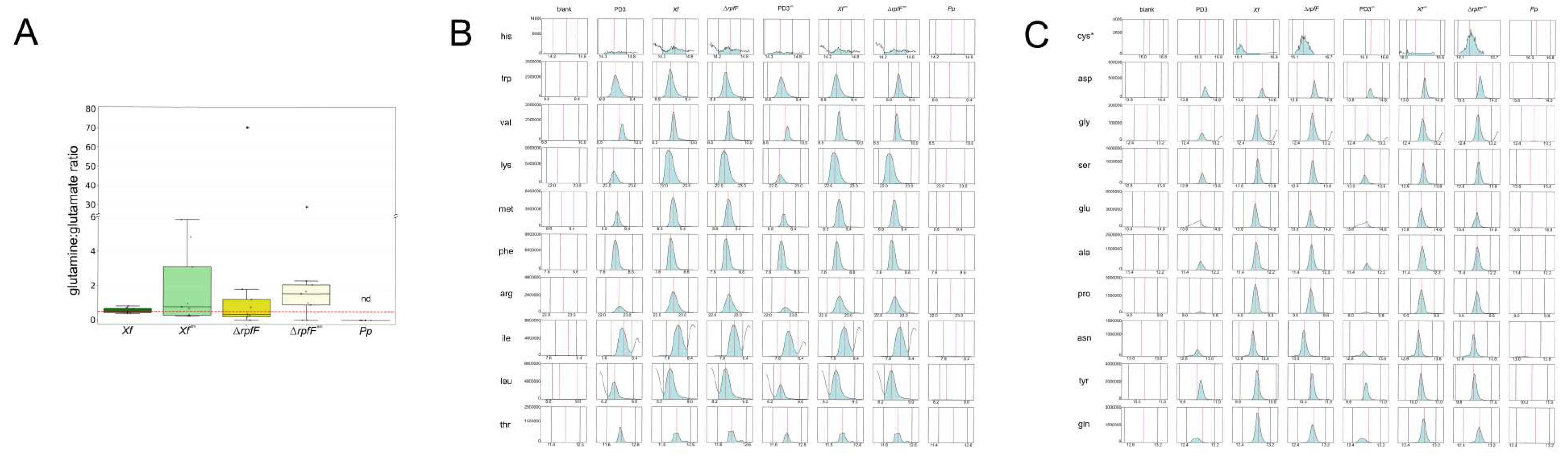
Figure 4.
Vitamin B compounds expression in Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes. A. Nicotinamide, B. Nicotinic acid and C. Biotin expression among the exometabolomes tested. Nicotinic acid is downregulated in Xf sm to ΔrpfF and ΔrpfFsm levels. Absence of the enzyme pyrazinamidase/nicotinamidase leads to Nicotinamide accumulation in Pp exometabolomes, there was no detection of this compound in X. fastidiosa strains. Biotin is detected only in Xf sm and ΔrpfFsm at very low levels. Significantly different expression by p-adjust value of pairwise comparison calculated by t-test (ns: p > 0.05, *: p ≤ 5.0×10-2, **: p ≤ 5.0×10-3, ***: p ≤ 5.0×10-4, ****: p ≤ 5.0×10-5). D. Nicotinamide, E. Nicotinic acid, and F. Biotin EIC. EICs of representative assessments in blank (MeOH), PD3, PD3sm, Xf, ΔrpfF, Xf sm, ΔrpfFsm, and Pp extracts or exometabolomes. Main reactions in “nicotinate and nicotinamide metabolism” and in “biotin metabolism” according to KEGG pathways (www.kegg.jp/pathway/) in G. X. fastidiosa and H. P. phytofirmans, respectively.
Figure 4.
Vitamin B compounds expression in Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes. A. Nicotinamide, B. Nicotinic acid and C. Biotin expression among the exometabolomes tested. Nicotinic acid is downregulated in Xf sm to ΔrpfF and ΔrpfFsm levels. Absence of the enzyme pyrazinamidase/nicotinamidase leads to Nicotinamide accumulation in Pp exometabolomes, there was no detection of this compound in X. fastidiosa strains. Biotin is detected only in Xf sm and ΔrpfFsm at very low levels. Significantly different expression by p-adjust value of pairwise comparison calculated by t-test (ns: p > 0.05, *: p ≤ 5.0×10-2, **: p ≤ 5.0×10-3, ***: p ≤ 5.0×10-4, ****: p ≤ 5.0×10-5). D. Nicotinamide, E. Nicotinic acid, and F. Biotin EIC. EICs of representative assessments in blank (MeOH), PD3, PD3sm, Xf, ΔrpfF, Xf sm, ΔrpfFsm, and Pp extracts or exometabolomes. Main reactions in “nicotinate and nicotinamide metabolism” and in “biotin metabolism” according to KEGG pathways (www.kegg.jp/pathway/) in G. X. fastidiosa and H. P. phytofirmans, respectively.
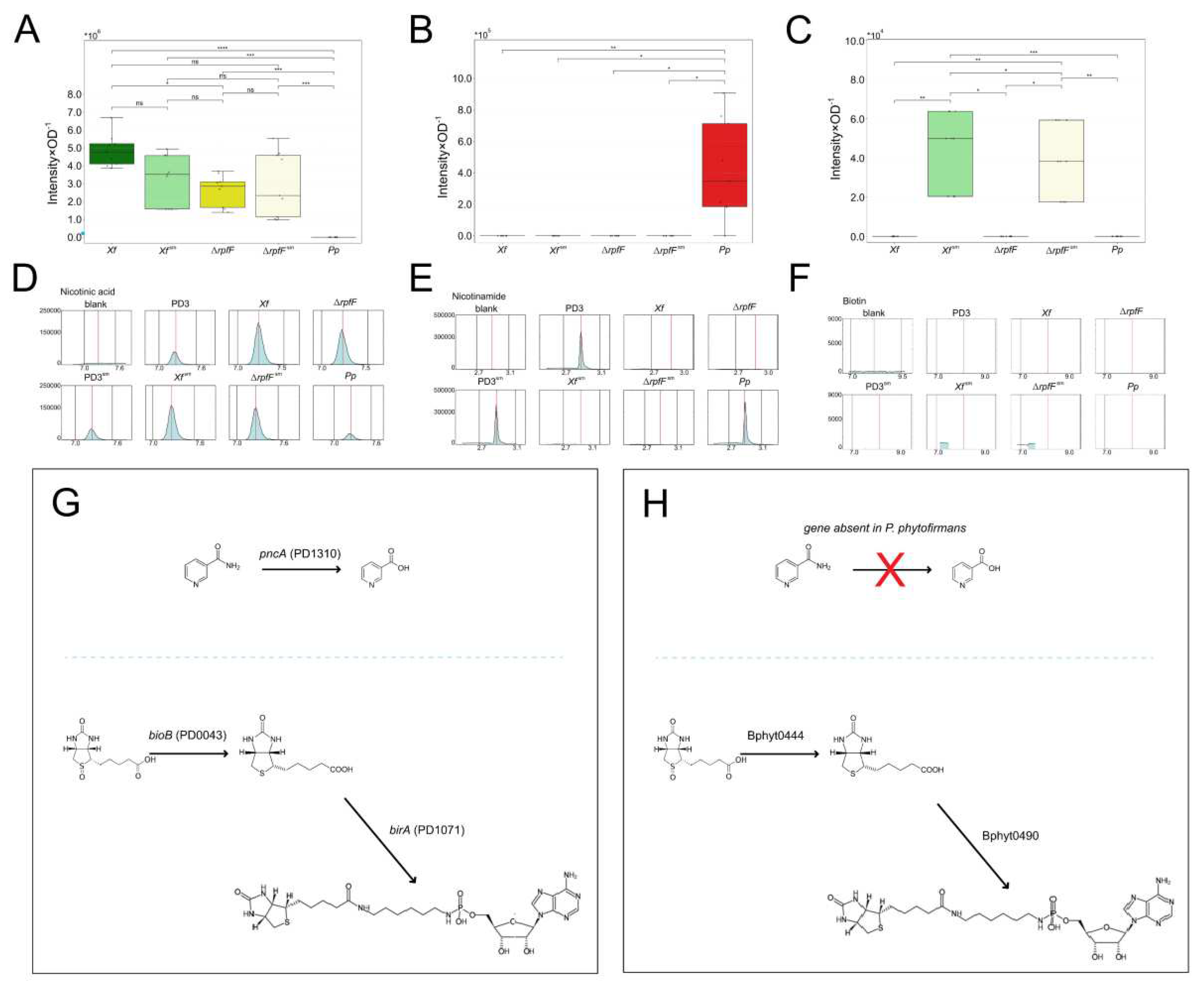
Figure 5.
AIB and GA expression in Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes. A. AIB and B. GA expression. AIB and B. GA are downregulated in Xf sm to ΔrpfF and ΔrpfFsm levels. Significantly different expression by p-adjust value of pairwise comparison calculated by t-test (ns: p > 0.05, *: p ≤ 5.0×10-2, **: p ≤ 5.0×10-3, ***: p ≤ 5.0×10-4, ****: p ≤ 5.0×10-5). C. AIB and D. GA EICs. EICs of representative assessments in blank (MeOH), PD3, PD3sm, Xf, ΔrpfF, Xf sm, ΔrpfFsm, and Pp extracts or exometabolomes.
Figure 5.
AIB and GA expression in Xf, Xf sm, ΔrpfF, ΔrpfFsm, and Pp exometabolomes. A. AIB and B. GA expression. AIB and B. GA are downregulated in Xf sm to ΔrpfF and ΔrpfFsm levels. Significantly different expression by p-adjust value of pairwise comparison calculated by t-test (ns: p > 0.05, *: p ≤ 5.0×10-2, **: p ≤ 5.0×10-3, ***: p ≤ 5.0×10-4, ****: p ≤ 5.0×10-5). C. AIB and D. GA EICs. EICs of representative assessments in blank (MeOH), PD3, PD3sm, Xf, ΔrpfF, Xf sm, ΔrpfFsm, and Pp extracts or exometabolomes.
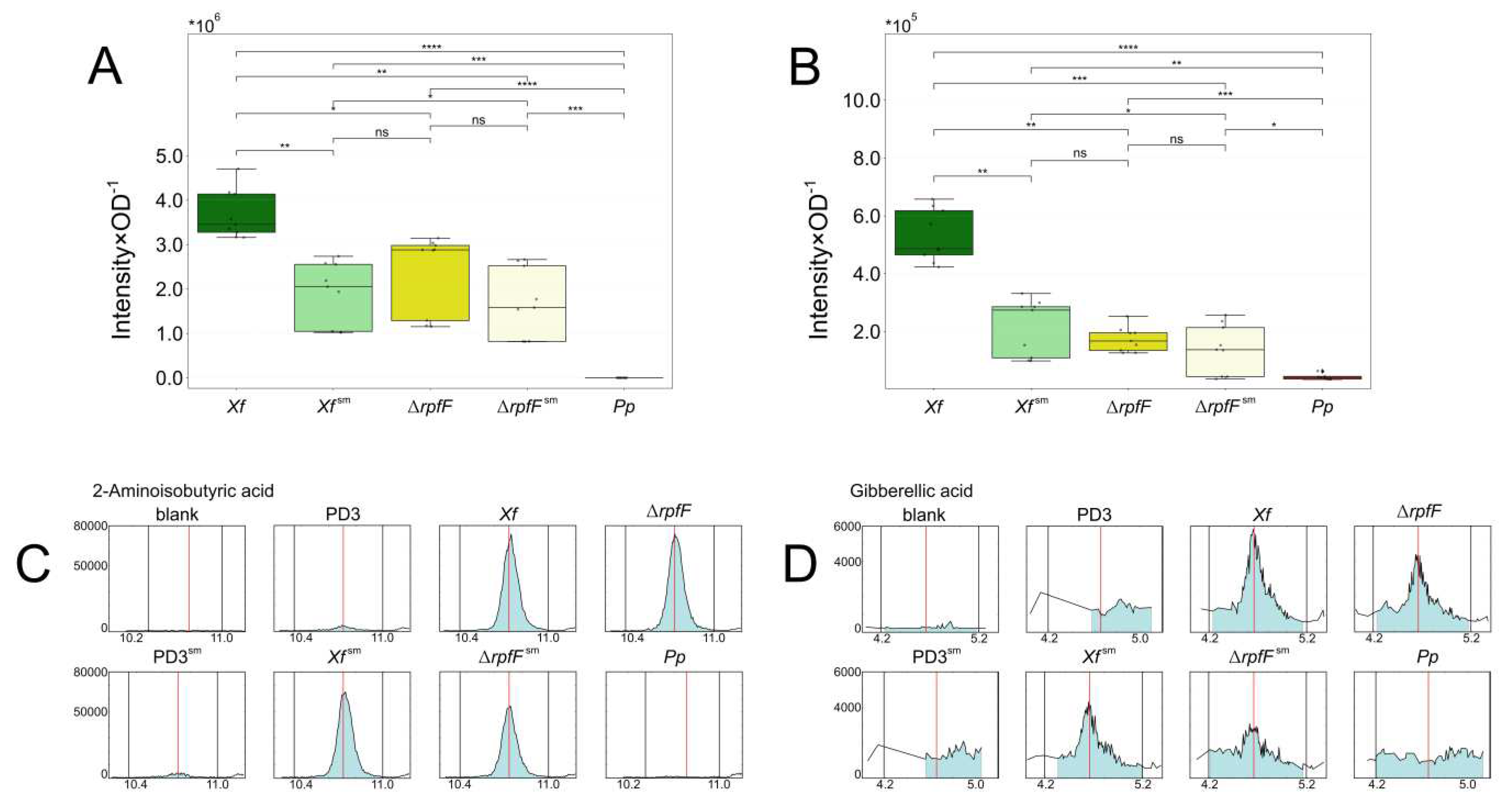
Figure 6.
Integration of metabolomics data with functional genomics and RpfF regulon (transcriptomic). A. GO Enrichment Analysis performed with genes retrieved from MAGI analysis. GO Annotations displayed are the top 5 over- and underrepresented in the 5 conditions tested, all the showed GO Annotations passed the p-value adjusted < 0.05 criteria. B. Public data from RpfF Regulon was crossed with genes predicted to be directly involved with the synthesis of the compounds detected in the exometabolomes. Color code, blue-to-red, indicates down and up-regulated genes in the RpfF Regulon. The shape size of circles indicates the expression level in peak size of compounds detected among the conditions tested. Peak size numbers were normalized into squared z-scores in order to maximize expression level differences among conditions. Dark gray dashed-circles around dots or “bubbles” indicate DEM in relation to Xf.
Figure 6.
Integration of metabolomics data with functional genomics and RpfF regulon (transcriptomic). A. GO Enrichment Analysis performed with genes retrieved from MAGI analysis. GO Annotations displayed are the top 5 over- and underrepresented in the 5 conditions tested, all the showed GO Annotations passed the p-value adjusted < 0.05 criteria. B. Public data from RpfF Regulon was crossed with genes predicted to be directly involved with the synthesis of the compounds detected in the exometabolomes. Color code, blue-to-red, indicates down and up-regulated genes in the RpfF Regulon. The shape size of circles indicates the expression level in peak size of compounds detected among the conditions tested. Peak size numbers were normalized into squared z-scores in order to maximize expression level differences among conditions. Dark gray dashed-circles around dots or “bubbles” indicate DEM in relation to Xf.
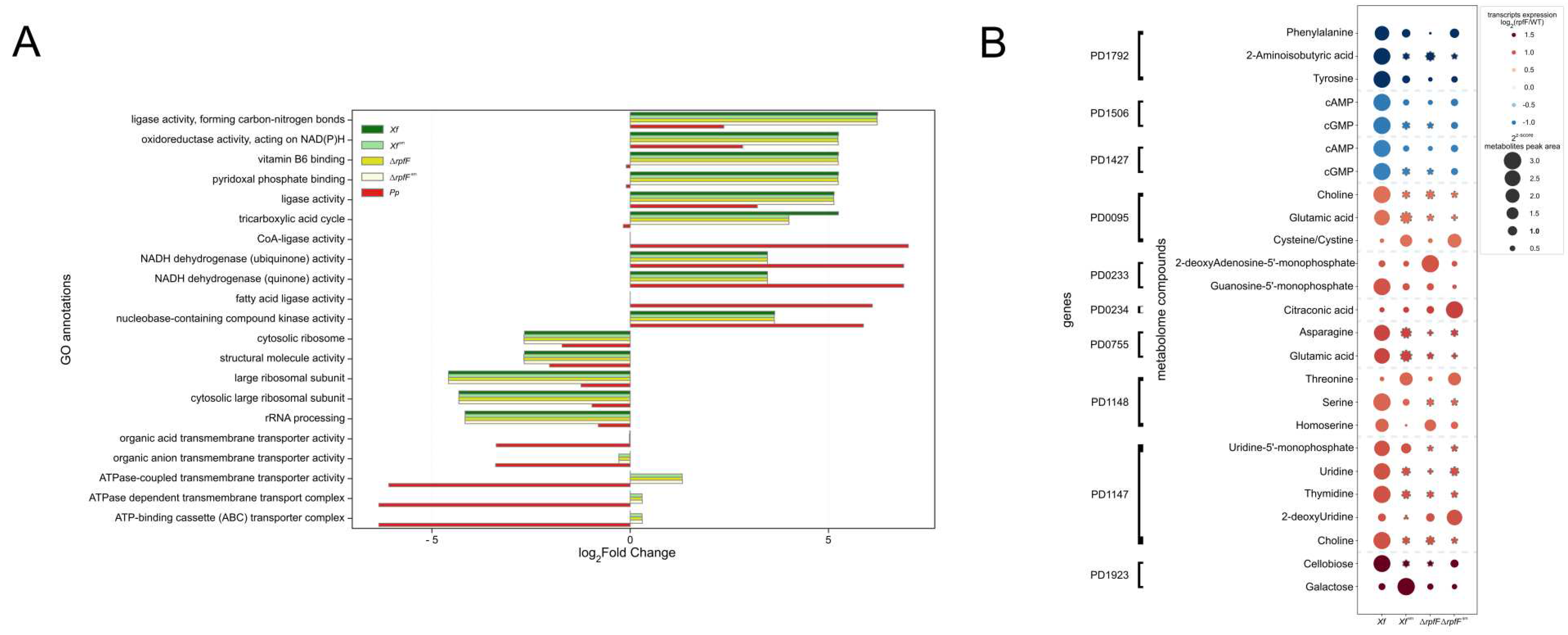

Table 1.
List of bacteria and strains used in this work.
| Bacteria | Strain | Original Host | Origin | Reference |
|---|---|---|---|---|
| Xylella fastidiosa | Temecula1 Wild Type (WT) |
Vitis vinifera | Temecula, California, EUA | [27,40] |
| ΔrpfF | - | - | [41] | |
| Paraburkholderia phytofirmans | PsJN | Allium cepa | Ontario, Canada | [42,43] |
Table 2.
Top-scored genes-to-compounds in X. fastidiosa according to MAGI results.
| compound name | gene id | reciprocal score | e-score reaction-to-gene | database id reaction-to-gene | e-score gene-to-reaction | database id gene-to-reaction | MAGI score |
|---|---|---|---|---|---|---|---|
| 2-Aminoisobutyric acid | PD0094 | 2.00 | 200.00 | ALANINE--TRNA-LIGASE-RXN | 200.00 | ALANINE--TRNA-LIGASE-RXN | 1.58 |
| PD1696 | 2.00 | 200.00 | RXN-16659 | 200.00 | RXN-16659 | 1.58 | |
| 2.00 | 200.00 | RXN-16649 | 200.00 | RXN-16649 | 1.58 | ||
| PD1823 | 2.00 | 200.00 | RHEA:20249 | 200.00 | RHEA:20249 | 1.58 | |
| PD1864 | 2.00 | 200.00 | RHEA:11224 | 200.00 | RHEA:11224 | 1.58 | |
| PD1865 | 2.00 | 200.00 | RHEA:23374 | 200.00 | RHEA:23374 | 1.58 | |
| Alanine | PD1864 | 2.00 | 200.00 | RHEA:11224 | 200.00 | RHEA:11224 | 6.33 |
| PD1823 | 2.00 | 200.00 | RHEA:20249 | 200.00 | RHEA:20249 | 6.33 | |
| PD0094 | 2.00 | 200.00 | ALANINE--TRNA-LIGASE-RXN | 200.00 | ALANINE--TRNA-LIGASE-RXN | 6.33 | |
| Arginine | PD0116 | 2.00 | 34.19 | ARGININE--TRNA-LIGASE-RXN | 32.31 | ARGININE--TRNA-LIGASE-RXN | 4.01 |
| Asparagine | PD1947 | 2.00 | 200.00 | ASPARAGINE--TRNA-LIGASE-RXN | 200.00 | ASPARAGINE--TRNA-LIGASE-RXN | 6.33 |
| Aspartate | PD0089 | 2.00 | 200.00 | RHEA:12228 | 200.00 | RHEA:12228 | 6.33 |
| 2.00 | 200.00 | RHEA:11375 | 200.00 | RHEA:11375 | 6.33 | ||
| PD0166 | 2.00 | 200.00 | RHEA:22630 | 200.00 | RHEA:22630 | 6.33 | |
| PD0291 | 2.00 | 200.00 | RHEA:10932 | 200.00 | RHEA:10932 | 6.33 | |
| PD0868 | 2.00 | 200.00 | RHEA:25877 | 200.00 | RHEA:25877 | 6.33 | |
| PD0946 | 2.00 | 200.00 | ASPARTATE--TRNA-LIGASE-RXN | 200.00 | ASPARTATE--TRNA-LIGASE-RXN | 6.33 | |
| PD1273 | 2.00 | 200.00 | RHEA:23777 | 200.00 | RHEA:23777 | 6.33 | |
| PD1274 | 2.00 | 200.00 | RHEA:20015 | 200.00 | RHEA:20015 | 6.33 | |
| PD1627 | 2.00 | 200.00 | RHEA:15753 | 200.00 | RHEA:15753 | 6.33 | |
| Biotin | PD0043 | 2.00 | 140.61 | 2.8.1.6-RXN | 145.10 | 2.8.1.6-RXN | 5.80 |
| PD1071 | 2.00 | 58.42 | RHEA:31118 | 62.94 | RHEA:31118 | 4.66 | |
| 2.00 | 58.42 | BIOTINLIG-RXN | 62.94 | BIOTINLIG-RXN | 4.66 | ||
| PD1494 | 2.00 | 167.61 | DETHIOBIOTIN-SYN-RXN | 165.73 | DETHIOBIOTIN-SYN-RXN | 1.51 | |
| Cysteine | PD0655 | 2.00 | 200.00 | RHEA:13285 | 200.00 | RHEA:13285 | 6.33 |
| PD1812 | 2.00 | 156.15 | RHEA:20400 | 154.24 | RHEA:20400 | 5.93 | |
| PD1841 | 2.00 | 143.27 | ACSERLY-RXN | 137.99 | ACSERLY-RXN | 5.77 | |
| PD0118 | 2.00 | 134.13 | RHEA:19398 | 132.24 | RHEA:19398 | 5.71 | |
| 2.00 | 134.13 | RHEA:25159 | 132.24 | RHEA:25159 | 5.71 | ||
| PD0690 | 2.00 | 133.98 | RXN0-308 | 132.09 | RXN0-308 | 5.71 | |
| PD0287 | 2.00 | 126.84 | CYSTEINE--TRNA-LIGASE-RXN | 124.08 | CYSTEINE--TRNA-LIGASE-RXN | 5.62 | |
| Gibberellic acid | PD0286 | 0.01 | 0.89 | RHEA:36115 | 7.41 | RHEA:25891 | 0.43 |
| PD0716 | 0.01 | 0.54 | RHEA:36115 | 159.33 | RHEA:13804 | 0.38 | |
| Nicotinic acid | PD0393 | 2.00 | 200.00 | RHEA:36166 | 200.00 | RHEA:36166 | 6.33 |
| PD1310 | 2.00 | 42.72 | RHEA:14545 | 40.84 | RHEA:14545 | 4.26 | |
| Glutamic acid | PD0650 | 2.00 | 200.00 | RHEA:17130 | 200.00 | RHEA:17130 | 6.33 |
| PD0654 | 2.00 | 200.00 | RHEA:18052 | 200.00 | RHEA:18052 | 6.33 | |
| PD0399 | 2.00 | 200.00 | RHEA:18633 | 200.00 | RHEA:18633 | 6.33 | |
| PD2062 | 2.00 | 200.00 | RHEA:11613 | 200.00 | RHEA:11613 | 6.33 | |
| 2.00 | 200.00 | RHEA:15504 | 200.00 | RHEA:15504 | 6.33 | ||
| PD1358 | 2.00 | 200.00 | RHEA:16574 | 200.00 | RHEA:16574 | 6.33 | |
| 2.00 | 200.00 | RHEA:14329 | 200.00 | RHEA:14329 | 6.33 | ||
| PD1266 | 2.00 | 200.00 | RHEA:23746 | 200.00 | RHEA:23746 | 6.33 | |
| PD0089 | 2.00 | 200.00 | RHEA:12228 | 200.00 | RHEA:12228 | 6.33 | |
| PD0839 | 2.00 | 200.00 | RHEA:24385 | 200.00 | RHEA:24385 | 6.33 | |
| PD1447 | 2.00 | 200.00 | GMP-SYN-GLUT-RXN | 200.00 | GMP-SYN-GLUT-RXN | 6.33 | |
| PD1848 | 2.00 | 200.00 | GLURS-RXN | 200.00 | GLURS-RXN | 6.33 | |
| PD1026 | 2.00 | 200.00 | RHEA:45804 | 200.00 | RHEA:45804 | 6.33 | |
| 2.00 | 200.00 | RHEA:16170 | 200.00 | RHEA:16170 | 6.33 | ||
| PD0851 | 2.00 | 200.00 | RHEA:14908 | 200.00 | RHEA:14908 | 6.33 | |
| PD0785 | 2.00 | 200.00 | RHEA:15136 | 200.00 | RHEA:15136 | 6.33 | |
| PD2063 | 2.00 | 200.00 | RHEA:12131 | 200.00 | RHEA:12131 | 6.33 | |
| 2.00 | 200.00 | RHEA:32192 | 200.00 | RHEA:32192 | 6.33 | ||
| 2.00 | 200.00 | GLUTAMATE-SYNTHASE-FERREDOXIN-RXN | 200.00 | GLUTAMATE-SYNTHASE-FERREDOXIN-RXN | 6.33 | ||
| PD0398 | 2.00 | 200.00 | RHEA:18633 | 200.00 | RHEA:18633 | 6.33 | |
| PD0170 | 2.00 | 200.00 | RHEA:21735 | 200.00 | RHEA:21735 | 6.33 | |
| PD0296 | 2.00 | 200.00 | RHEA:14879 | 200.00 | RHEA:14879 | 6.33 | |
| PD0541 | 2.00 | 200.00 | RHEA:15890 | 200.00 | RHEA:15890 | 6.33 | |
| PD0110 | 2.00 | 200.00 | RHEA:13239 | 200.00 | RHEA:13239 | 6.33 | |
| PD0655 | 2.00 | 200.00 | RHEA:13285 | 200.00 | RHEA:13285 | 6.33 | |
| Glutamine | PD1447 | 2.00 | 200.00 | GMP-SYN-GLUT-RXN | 200.00 | GMP-SYN-GLUT-RXN | 6.33 |
| PD0584 | 2.00 | 200.00 | GLUTAMINE--TRNA-LIGASE-RXN | 200.00 | GLUTAMINE--TRNA-LIGASE-RXN | 6.33 | |
| PD2063 | 2.00 | 200.00 | GLUTAMATE-SYNTHASE-FERREDOXIN-RXN | 200.00 | GLUTAMATE-SYNTHASE-FERREDOXIN-RXN | 6.33 | |
| Glycine | PD1750 | 2.00 | 200.00 | RHEA:15482 | 200.00 | RHEA:15482 | 6.33 |
| PD0620 | 2.00 | 200.00 | GCVMULTI-RXN | 200.00 | GCVMULTI-RXN | 6.33 | |
| 2.00 | 200.00 | GCVP-RXN | 200.00 | GCVP-RXN | 6.33 | ||
| PD1810 | 2.00 | 200.00 | GCVMULTI-RXN | 200.00 | GCVMULTI-RXN | 6.33 | |
| 2.00 | 200.00 | GCVP-RXN | 200.00 | GCVP-RXN | 6.33 | ||
| PD0827 | 2.00 | 200.00 | RHEA:17453 | 200.00 | RHEA:17453 | 6.33 | |
| PD0704 | 2.00 | 200.00 | RHEA:19938 | 200.00 | RHEA:19938 | 6.33 | |
| PD0844 | 2.00 | 200.00 | RHEA:13557 | 200.00 | RHEA:13557 | 6.33 | |
| PD0773 | 2.00 | 200.00 | RXN0-7068 | 200.00 | RXN0-7068 | 6.33 | |
| 2.00 | 200.00 | RXN0-7082 | 200.00 | RXN0-7082 | 6.33 | ||
| PD0841 | 2.00 | 170.04 | GLYCINE--TRNA-LIGASE-RXN | 168.01 | GLYCINE--TRNA-LIGASE-RXN | 6.06 | |
| PD0840 | 2.00 | 164.75 | GLYCINE--TRNA-LIGASE-RXN | 162.87 | GLYCINE--TRNA-LIGASE-RXN | 6.02 | |
| Histidine | PD1267 | 2.00 | 200.00 | RHEA:20641 | 200.00 | RHEA:20641 | 6.33 |
| PD1772 | 2.00 | 60.65 | RHEA:20641 | 58.79 | RHEA:20641 | 4.66 | |
| PD1270 | 2.00 | 26.76 | HISTIDINE--TRNA-LIGASE-RXN | 24.84 | HISTIDINE--TRNA-LIGASE-RXN | 3.76 | |
| Isoleucine | PD1437 | 2.00 | 200.00 | ISOLEUCINE--TRNA-LIGASE-RXN | 200.00 | ISOLEUCINE--TRNA-LIGASE-RXN | 6.33 |
| Leucine | PD1230 | 2.00 | 200.00 | LEUCINE--TRNA-LIGASE-RXN | 200.00 | LEUCINE--TRNA-LIGASE-RXN | 6.33 |
| Lysine | PD0404 | 2.00 | 200.00 | LYSINE--TRNA-LIGASE-RXN | 200.00 | LYSINE--TRNA-LIGASE-RXN | 6.33 |
| PD1514 | 2.00 | 69.38 | RXN-1961 | 78.45 | RXN-1961 | 4.86 | |
| PD2000 | 2.00 | 55.90 | RHEA:15944 | 53.31 | RHEA:15944 | 1.14 | |
| Methionine | PD1590 | 2.00 | 200.00 | METHIONINE--TRNA-LIGASE-RXN | 200.00 | METHIONINE--TRNA-LIGASE-RXN | 6.33 |
| 2.00 | 200.00 | RXN-16165 | 200.00 | RXN-16165 | 6.33 | ||
| Phenylalanine | PD1911 | 2.00 | 200.00 | PHENYLALANINE--TRNA-LIGASE-RXN | 200.00 | PHENYLALANINE--TRNA-LIGASE-RXN | 6.33 |
| PD0665 | 2.00 | 200.00 | RXN-15898 | 200.00 | RXN-15898 | 1.58 | |
| Proline | PD1635 | 2.00 | 200.00 | PROLINE--TRNA-LIGASE-RXN | 200.00 | PROLINE--TRNA-LIGASE-RXN | 6.33 |
| Serine | PD0612 | 2.00 | 200.00 | RHEA:26437 | 200.00 | RHEA:26437 | 6.33 |
| 2.00 | 200.00 | RHEA:10532 | 200.00 | RHEA:10532 | 6.33 | ||
| PD1750 | 2.00 | 200.00 | RHEA:15482 | 200.00 | RHEA:15482 | 6.33 | |
| PD1318 | 2.00 | 200.00 | RXN0-2161 | 200.00 | RXN0-2161 | 6.33 | |
| 2.00 | 200.00 | SERINE--TRNA-LIGASE-RXN | 200.00 | SERINE--TRNA-LIGASE-RXN | 6.33 | ||
| PD0613 | 2.00 | 200.00 | RHEA:26437 | 200.00 | RHEA:26437 | 6.33 | |
| 2.00 | 200.00 | RHEA:10532 | 200.00 | RHEA:10532 | 6.33 | ||
| Threonine | PD1916 | 2.00 | 200.00 | THREONINE--TRNA-LIGASE-RXN | 200.00 | THREONINE--TRNA-LIGASE-RXN | 6.33 |
| Tryptophan | PD0612 | 2.00 | 200.00 | RHEA:10532 | 200.00 | RHEA:10532 | 6.33 |
| 2.00 | 200.00 | RHEA:26437 | 200.00 | RHEA:26437 | 6.33 | ||
| PD0613 | 2.00 | 200.00 | RHEA:10532 | 200.00 | RHEA:10532 | 6.33 | |
| 2.00 | 200.00 | RHEA:26437 | 200.00 | RHEA:26437 | 6.33 | ||
| PD1650 | 2.00 | 47.82 | TRYPTOPHAN--TRNA-LIGASE-RXN | 45.82 | TRYPTOPHAN--TRNA-LIGASE-RXN | 4.38 | |
| Tyrosine | PD0665 | 2.00 | 200.00 | RXN-15898 | 200.00 | RXN-15898 | 6.33 |
| PD0132 | 2.00 | 33.23 | TYROSINE--TRNA-LIGASE-RXN | 31.35 | TYROSINE--TRNA-LIGASE-RXN | 3.98 | |
| Valine | PD0102 | 2.00 | 200.00 | VALINE--TRNA-LIGASE-RXN | 200.00 | VALINE--TRNA-LIGASE-RXN | 6.33 |
Table 3.
Top-scored genes-to-compounds in P. phytofirmans according to MAGI results.
| compound name | gene id | reciprocal score | e-score reaction-to-gene | database id reaction-to-gene | e-score gene-to-reaction | database id gene-to-reaction | MAGI score |
|---|---|---|---|---|---|---|---|
| Cysteine | Bphyt_3930 | 2.00 | 200.00 | RHEA:13285 | 200.00 | RHEA:13285 | 6.33 |
| Bphyt_4072 | 2.00 | 200.00 | RXN-17172 | 200.00 | RXN-17172 | 6.33 | |
| Bphyt_2579 | 2.00 | 200.00 | RXN-15881 | 200.00 | RXN-15881 | 6.33 | |
| 2.00 | 200.00 | RXN0-308 | 200.00 | RXN0-308 | 6.33 | ||
| Bphyt_3068 | 2.00 | 200.00 | RHEA:28784 | 200.00 | RHEA:28784 | 6.33 | |
| Bphyt_0110 | 2.00 | 200.00 | RHEA:13285 | 200.00 | RHEA:13285 | 6.33 | |
| Bphyt_3953 | 2.00 | 200.00 | RHEA:13285 | 200.00 | RHEA:13285 | 6.33 | |
| Bphyt_2579 | 2.00 | 177.70 | RXN-14385 | 175.79 | RXN-14385 | 6.13 | |
| Bphyt_2510 | 2.00 | 175.50 | CYSTEINE--TRNA-LIGASE-RXN | 171.77 | CYSTEINE--TRNA-LIGASE-RXN | 6.10 | |
| Gibberellic acid | Bphyt_5231 | 0.01 | 18.39 | RXN-14318 | 39.82 | RXN-16827 | 0.23 |
| 0.01 | 18.39 | RXN-14317 | 39.82 | RXN-16827 | 0.23 | ||
| 0.01 | 18.39 | RXN-7617 | 39.82 | RXN-16827 | 0.23 | ||
| Nicotinamide | Bphyt_5413 | 2.00 | 200.00 | RHEA:16150 | 200.00 | RHEA:16150 | 6.33 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



