Submitted:
29 November 2023
Posted:
30 November 2023
You are already at the latest version
Abstract
In the absence of information on Borrelia species causing Lyme borreliosis (LB) in Serbia, we were interested in the pathogenic potential of local strains of Borrelia burgdorferi sensu lato isolated from ticks. The susceptibility of B. burgdorferi sensu lato isolates to human complement was tested in vitro. A total of 31 strains were analysed by serum susceptibility testing - 12 Borrelia afzelii, 12 Borrelia lusitaniae, three Borrelia bavariensis, two Borrelia garinii, and two Borrelia valaisiana. The serum-resistant strains (B. afzelii, B. bavariensis and B. valaisiana) expressing OspA and OspB or OspA and OspC proteins might be related to the different pathogenic potential of the specific Borrelia strains and their ability to cause different clinical manifestations of LB in humans. Among the B. lusitaniae strains from Serbia, two were significantly more motile than others (all p < 0.05) and equally motile as human isolate (both p > 0.05), and since they express OspC, their pathogenic potential remains unclear. For further studies, it is of particular interest to explore the mechanism of resistance of B. valaisiana and to explain the fair motility of B. lusitaniae, a predominant Borrelia species in Serbia, in the presence of human complement.
Keywords:
Borrelia
; human complement
; resistance
1. Introduction
Lyme borreliosis (LB), a tick-borne infectious disease caused by spirochetes of the Borrelia burgdorferi sensu lato (s.l.) complex, is the most widespread vector-borne disease in the temperate climates of the Northern Hemisphere [1,2]. Currently, 21 described Borrelia species with considerable variability in host and vector associations and geographical distribution are assigned to the B. burgdorferi s.l. complex [3]. Only a subset of B. burgdoferi s.l. species are known to cause disease in humans. In North America, LB is predominantly caused by the Borrelia burgdorferi sensu stricto (s.s.), whereas in Europe the overall diversity of B. burgdorferi s.l. is higher and LB is caused by Borrelia afzelii, Borrelia garinii, Borrelia bavariensis (formerly B. garinii OspA type 4), Borrelia spielmanii and, more rarely, Borrelia burgdorferi s.s., while Borrelia valaisiana, Borrelia lusitaniae and Borrelia bissettii have only been identified in isolated cases and are recognised as potential pathogens [1,2]. Infection with different Borrelia species leads to different clinical manifestations, but the skin lesion - erythema migrans is the most common [1,2]. In persistent infection, B. afzelii usually remains localised in the skin, B. garinii and B. bavariensis are usually associated with nervous system disorders, while B. burgdorferi s.s. is often associated with the development of arthritis [1,2].
The pathogenicity of specific Borrelia strains depends on their invasiveness, antigenic variability, lymphocyte stimulation, and resistance to host complement [4], which is enabled by differential expression of outer surface proteins (Osp) and their variability. In vitro studies have shown that B. afzelii, B. bavariensis, B. spielmanii and B. burgdorferi s.s. are resistant to complement-mediated killing, B. bissettii is intermediate serum resistant, while B. garinii, B. valaisiana, and are highly susceptible to complement-mediated killing (4). Differences in serum susceptibility have also been found between strains within the B. valaisiana and B. garinii species [5,6,7].
While B. valaisina, B. afzelii, and B. garinii are the most prevalent species in ticks throughout Europe [8], studies on the presence of B. burgdorferi s.l. in ticks from Serbia [9,10,11] indicated the dominance of B. lusitaniae followed by B. afzelii, B. bavariensis, B. garinii, B. valaisiana and B. burgdorferi s.s. The region of Serbia and the Balkan Peninsula is characterised by a high prevalence and diversity of Borrelia species in Ixodes ricinus ticks [12], but the reported incidence of LB in Serbia between 2013 and 2019 was 6.83-13.32/100.00 inhabitants [13], which is much lower compared to regions with a similar prevalence of Borrelia in ticks [14,15]. In Germany and Slovenia, for example, the annual incidence rate in 2014 was 111/100,000 and 188.7/100,000, respectively [14]; in Lithuania, 99.9/100,000; and in Finland, 61/100,000. In France, the annual incidence rate between 2009 and 2017 was 53/100,000; in the United Kingdom, 12.1/100,000; and in Italy, there were only 1.24 new cases per 1 million population between 2000 and 2015 [15]. The most common clinical manifestation of LB in Serbia is erythema migrans, which occurs in 93.21% of cases, followed by neurological, musculoskeletal, and cardiac manifestations in 2.80, 2.46, and 1.10% of cases, respectively [16]. However, apart from several studies on the presence and diversity of B. burgdorferi s.l. in ticks and animal hosts [17,18], there are no data on the exact Borrelia species that cause LB in humans. Against this background, we wanted to find out which Borrelia strains have the potential to cause LB by testing the susceptibility of different local strains isolated from I. ricinus ticks to human complement in vitro. As B. lusitaniae species predominate in both Serbia and Mediterranean countries and the pathogenic potential is still unclear, we were interested in comparing the susceptibility of available B. lusitaniae strains from Serbia, Spain, and Portugal to human complement and their pathogenic potential in vitro.
2. Materials and Methods
2.1. Borrelia Strains
The thirty-one B. burgdorferi s.l. strains were included in the study - 30 isolated from I. ricinus ticks and one strain isolated from human skin. The twenty-seven Borrelia strains (12 B. afzelii, eight B. lusitaniae, three B. bavariensis, two B. garinii, and two B. valaisiana) were isolated from I. ricinus ticks from different localities in Serbia and previously described by Ćakić and colleagues [9]. Four external B. lusitaniae strains (Heavy, Listu, PotiB2, and PoHL1) were provided from Spain and Portugal and described previously [19]. All strains tested are listed in Table 1.
2.2. Protein Analysis
Protein characterisation was performed for a total of 29 Borrelia strains - 27 Borrelia strains isolated from I. ricinus ticks from Serbia [9] and two Borrelia strains isolated from I. ricinus ticks from Spain [20,21]. Cultivation of the Borrelia strains, extraction of the Borrelia proteins and analysis of the obtained proteins using SDS-PAGE were performed as previously described [22,23]. Briefly, proteins were separated by SDS-PAGE with 12% gel, stained afterwards with Coomassie brilliant blue. At least three electrophoreses were performed for each strain. The molecular masses of the proteins were determined based on their position in the gel compared to the molecular mass standard. Discrimination between strains with respect to the presence of Osps (Osp-positive, Osp-weakly positive, or Osp-negative) was determined based on band intensity using the Gel Doc software (BioRad, Germany) described previously [22]. Based on the results obtained, the Borrelia strains were classified into protein profile groups (hereafter referred to as PPGs) distinguished by the presence or absence of OspA, OspB and OspC.
The protein profiles of two Borrelia strains from Portugal used in this study (one isolated from the I. ricinus tick and one isolated from human skin) were described by Zeidner and colleagues (24) and Collares-Pereira and colleagues [25].
2.3. Collection of Sera
Serum samples from eight volunteer blood donors were pooled for normal human serum (NHS), filtered through 0.22 µm syringe filters (Lab Logistics Group GmbH, Meckenheim, Germany), immediately aliquoted and frozen at -80°C and thawed only once before use. Heat-inactivated serum (HIS) was generated by incubating NHS at 56°C for 45 minutes and used as a complement control [6,26]. Serum samples were tested for the presence of anti-B. burgdorferi s.l. IgM and IgG, and antibodies were all negative using an ELISA assay (Euroimmun, Germany). To confirm the results, the sera were tested with the anti-Borrelia EUROLINE Western blot test (Euroimmun, Germany) and all samples were negative.
2.4. Ethical Statements
All healthy subjects gave written informed consent at the Institute of Blood Transfusion and Haemobiology, Military Medical Academy, Belgrade, Republic of Serbia (Decision number 1130-8, November 20, 2015) The study complied with the Declaration of Helsinki and was approved by the Ethics Committee of the Military Medical Academy, Belgrade, Republic of Serbia.
2.5. Serum Susceptibility
Before performing the serum susceptibility test, all Borrelia strains (31 strains), previously stored in biobanks, were recultured in Barbour-Stoenner-Kelly-H medium (BSK-H) (Sigma-Aldrich, St Louis, MO, U.S.A.) at 33°C until they reached a concentration of 1-2 x 107 cells/ml (26). The number of Borrelia cells/mL was determined by dark-field microscopy using the Neubauer counting chamber (Brand Gmbh & Co. KG, Wertheim, Germany) as described before [26].
The serum susceptibility test was performed by counting the spirochetes under the dark-field microscope after an incubation period of one and three hours, as previously described by Wagemakers and colleagues [26]. Briefly, in a 96-well microtitre plate, 25 μL of each Borrelia culture and 25 μl of NHS or HIS were mixed, and the plate was sealed and incubated at 33°C for one and three hours. After incubation, 5 μl of the suspension from each well was examined under a dark-field microscope. Samples were blinded and 100 Borrelia per well were counted as either motile or immotile. The loss of motility of spirochetes in NHS wells compared with HIS was indicative of complement-mediated killing and inactivation of spirochetes (susceptibility of Borrelia strains to NHS), while the presence of viable, motile spirochetes in NHS compared with HIS was indicative of resistance of spirochetes to human complement [26]. For each strain of Borrelia, the experiment was repeated at least three times and then the median percentage of the number of viable and motile spirochetes was determined. To obtain consistent results and minimise the possibility of errors, the counting of motile and immotile spirochetes under the dark-field microscope was always performed by the same experienced person.
2.6. Statistical Analysis
A Kruskal-Wallis test was performed to identify a difference in motility between Borrelia species and between Borrelia strains within species observed in the serum susceptibility tests performed. The significance of the difference between the motility of two strains and two species in NHS and in two test conditions (NHS and HIS) for each Borrelia strain after one and three hours of incubation was analysed using a Mann-Whitney test. All statistical tests were considered statistically significant at the 0.05 probability level. Statistical analyses were performed using the program PASW Statistics 18 (IBM, Armonk, New York, USA). The graphs were created with Excel (Microrosft 365).
3. Results
3.1. Protein Analysis
In this study, the protein profiles of 27 Borrelia strains isolated from I. ricinus ticks from Serbia and two Borrelia strains isolated from I. ricinus ticks from Spain were determined (Table S1, Figure S1, Figure S2, Figure S3, Figure S4, Figure S5). The molecular mass of OspA, OspB and OspC was calculated using data obtained from repeated electrophoreses and aforementioned software. The results were the same for all electrophoreses performed (Table S1). The protein profiles of two Borrelia strains from Portugal (PotiB2 and PoHL1) have been described previously, but the molecular masses have not been defined [24,25]. Some proteins were found weakly positive (Osp-weakly positive), but the number of these strains was small. Only one isolate was weakly OspB-positive (RS 235_13cd) (Table S1, Figure S1), while two isolates (RS 164_11g and RS 226_10d) were weakly OspC-positive (Table S1, Figure S3, Figure S5).
The presence of specific Osp in relation to Borrelia strains is shown in Table 2 and Table 3. Among the Osps, OspA-positive isolates were found most frequently (97%), followed by OspC-positive (65%), while OspB-positive were found in almost half of the strains (55%) (Table 2).
The presence or absence of Osps in the tested strains resulted in five protein profile groups (PPGs I-V). The results are shown in Table 3. All three proteins (OspA, OspB and OspC) were present in 14/31 (45%) strains, OspA and OspB were simultaneously present in 2/31 (7%), while OspA and OspC were simultaneously present in 6/31 (19%) strains; only OspA was present in 8/31 (26%) strains, while only OspB was present in 1/31 (3%) (Table 3).
3.2. Serum Susceptibility
Serum susceptibility test was performed for 31 strains of Borrelia tested - 27 strains of Borrelia from Serbia (12 B. afzelii, eight B. lusitaniae, three B. bavariensis, two B. garinii, and two B. valaisiana) and four B. lusitaniae from Portugal and Spain. A range and median percentage of viable, motile spirochetes in the presence of NHS and heat-inactivated serum (HIS) for each Borrelia strain after one hour (1h) and three hours (3h) of incubation are presented in Table 1. The most motile Borrelia species in the presence of NHS after three hours of incubation were B. bavariensis (median percentage of viable motile spirochetes=100%, range=96-100%), B. afzelii (median=100%, range=90-100%) and B. valaisiana (median=60%; range=52-89%), followed by B. lusitaniae (median=1%, range=0-50%) and B. garinii (median=0%, range=0-2%) (Table 1).
Based on the comparison of median percentage of viable motile spirochetes in the presence of NHS and HIS, all tested B. afzelii strains (12/12), B. bavariensis strains (3/3), and one of two B. valaisiana strains (RS 164_12b) were resistant to NHS after one and three hours of incubation, while all tested B. garinii strains (2/2) and B. lusitaniae strains (12/12) and another B. valaisiana strain (RS 224_10b) were susceptible to NHS after one and three hours of incubation (Table 1, Figure1).
3.3. Protein Profiles of Analyzed Strains and Susceptibility to Human Complement
Considering the protein profiles of the strains tested and the susceptibility to human complement, resistance to NHS was found after one and three hours of incubation in 13 (10 B. afzelii strains and three B. bavariensis strains) of 14 Borrelia strains assigned to PPG I (all tested proteins present), two (both B. afzelii strains) of two Borrelia strains assigned to PPG II (OspC absent), and one (one B. valaisiana strain) of six Borrelia strains assigned to PPG III (OspB absent) (Table 1, Table 3). Five (two B. garinii strains and three B. lusitaniae strains) of six Borrelia strains assigned to PPG III (OspB absent), eight (one B. valaisiana strain and seven B. lusitaniae strains) of eight Borrelia strains assigned to PPG IV (OspB and OspC absent), one (B. lusitaniae strain) of one Borrelia strain assigned to PPG V (OspA and OspC absent) and one (B. lusitaniae strain) of 14 Borrelia strains assigned to PPG I (all tested proteins present) were susceptible to NHS after one and three hours of incubation (Table 1, Table 3).
Two of three B. lusitaniae strains (median percentage of motile RS 226_10d strain=20%, range=3-45%, and median percentage of motile RS 76_12a strain=15%, range=5-20%) from Serbia (PPG III) were more motile than other B. lusitaniae (strains belong to PPGs III, IV and V) and equally motile as the human isolate from Portugal (PPG I) (median percentage of motile PoHL1 strain=32%, range=27-50%) after three hours of incubation in the presence of NHS (Table 1, Table 3, Figure 1B). One B. valasiana strain (RS 164_12b) assigned to PPG III (OspB absent) (median percentage of motile strain= 89%, range=85-91% after one hour of incubation and median percentage of motile strain=87%, range=77-89% after three hours of incubation) was resistant to NHS, while another B. valasiana strain (RS 224_10b) assigned to PPG IV (OspB and OspC absent) (median percentage of motile strain= 75%, range=53-81% after one hour of incubation and median percentage of motile strain =60%, range=52-76% after three hours of incubation) was susceptible to NHS (Table 1, Table 3, Figure 1).
4. Discussion
The diversity of B. burgdorferi s.l. strains circulating in tick-mammal cycles in certain regions exceed the diversity of strains that can cause LB in humans. The ability of specific Borrelia strains to resist clearance by the host complement system is an important step for persistence in any host, including humans.
In this study, we were able to demonstrate differences between the Borrelia strains examined in terms of their susceptibility to NHS in vitro (Table 1, Figure 1). The method we chose to identify local Borrelia strains with pathogenic potential was a serum susceptibility test based on the counting of motile and non-motile spirochetes under the dark-field microscope after one- and three-hours of incubation [26]. Counting of Borrelia by dark-field microscopy is a time-consuming and subjective method, but still considered the gold standard for determining motile spirochetes [27]. The results of this study showed that all B. afzelii, B. bavariensis, and one B. valaisiana strain tested were motile and viable in the presence of NHS suggesting resistance to human complement, pathogenic potential, and the ability to cause LB in humans. In contrast, all tested B. lusiataniae, two B. garinii, and one B. valaisiana strain were partially or mostly immotile in the presence of NHS, suggesting susceptibility to complement and non-pathogenic potential of these strains.
Although the serum susceptibility pattern of most Borrelia strains is consistent with pathogenicity in humans, there are two exceptions [4] - B. garinii, which is known to frequently cause LB and is usually associated with nervous system disorders and B. valaisina [1,2]. The underlying molecular mechanisms of how B. garinii circumvent complement-mediated killing are largely unknown [4] and since both B. garinii strains analyzed in our study were susceptible to human serum, we are also unable to answer why the local Borrelia strains, which are among the proven human pathogenic species, are serum-susceptible.
The pathogenic potential of B. valaisiana is still unclear [1,2]. There is evidence that B. valaisiana (previously referred to as genomic groups VS116 and M19) may also cause LB, although only skin biopsies or cerebrospinal fluid (CSF) samples from patients have been positive for B. valaisiana DNA [28,29]. In vitro, B. valaisiana has been shown to be a serum-susceptible species [4], although Schwab and colleagues [5] have shown that B. valaisiana strains differ in their susceptibility to human serum. It appears that serum-resistant B. valaisiana strains have different molecular mechanisms for inhibiting complement activation, independent of the recruitment of complement regulators or by inactivation of central complement components. The exact molecular mechanism is still unclear. In our study, one of two B. valaisiana strains examined was serum-resistant, suggesting that it can potentially cause LB.
Another B. valaisiana strain tested was serum-susceptible but statistically significantly more motile than all B. garinii and B. lusitaniae strains in the presence of NHS, indicating intermediate serum resistance of this strain. In our study, the B. valaisiana strain is the second to show resistance to human complement, having already been found in the study by Schwab and colleagues [5]. Further studies are needed to identify the mechanism behind the complement resistance of B. valaisiana strains circulating in the region.
Borrelia lusitaniae is also considered a potentially pathogenic species [1,2]. The isolation of B. lusitaniae from chronic human skin lesions from Portugal [25] adds to the evidence that this species may cause LB. In the present study, we demonstrated that all B. lusitaniae strains tested from Serbia, Portugal, and Spain were serum-susceptible. However, two B. lusitaniae strains from Serbia (RS 226_10d and RS 76_12a) and the strain isolated from human skin (PoHL1) were statistically significantly more motile (in the presence of NHS) than other B. lusitaniae strains and fairly motile. The similarity of activity between the local Borrelia strains and the human isolate from Portugal in the presence of NHS may suggest that some B. lusitaniae strains circulating in the region have the potential to overcome complement and cause LB in humans.
The heterogeneity of Borrelia Osps has implications for the pathogenesis and therapy of infection and is of great importance for serological testing and vaccine development [30]. OspA is a protein common to Borrelia strains and has been interpreted as an adhesive molecule required for the attachment of Borrelia to proteins, including proteins of the tick's intestinal cells [31]. OspB has also been interpreted as crucial for the adherence to the tick gut and survival of Borrelia within ticks, but not crucial for Borrelia transmission [32,33], while the difference in the presence of OspB between species may support findings on the function of OspB as an adhesive molecule that could anchor Borrelia in the skin and thus prevent the spread of Borrelia [22]. In the present study, the expression of the Osps studied ranged from absent to highly expressed and based on the expression patterns, the strains were classified into five PPGs (I to V) (Table 3, Figure S1, Figure S2, Figure S3, Figure S4, Figure S5). We observed a clear separation of strains with specific protein profiles with respect to resistance to NHS. All serum-resistant strains were assigned to PPGs I and II, while serum-susceptible strains were assigned to PPGs III, IV and V. The only exceptions to this rule are the serum-susceptible human B. lusitaniae isolate (PoHL1), which belongs to PPG I and was quite motile after three hours of incubation in the presence of NHS, and a serum-resistant B. valaisiana, which belongs to PPG III (Table 1, Table 3, Figure 1).
All serum-resistant strains expressed all Osps or OspA and OspB, except for one B. valasiana strain (RS 164_12b) that expressed OspA and OspC (Table 1, Table 3, Figure 1), indicating a possible significant role of simultaneous expression of OspA and OspB or OspA and OspC for the pathogenic potential of local Borrelia strains. On the other hand, strains that were serum-susceptible expressed OspA and OspC, exclusively expressed OspA or exclusively expressed OspB (Table 1, Table 3, Figure 1).
In our study, the same methodology (all conditions were identical) was used to determine the protein profile of local Borrelia isolates from ticks as in the study by Glinšek and colleagues [22]. The authors determined the protein profile of B. afzelii and B. garinii isolated from human material (skin and CSF) and found that B. afzelii and B. garinii strains expressing OspC were associated with both skin and neurological manifestations of LB compared to strains expressing only OspA and OspB, which were predominantly associated with skin manifestations and mostly determined to be B. afzelii. In our study, a total of 65% (20/31) of the Borrelia strains tested expressed OspC (Table 2, Table 3), of which 14 strains (10 B. afzelii, three B. bavariensis and one B. valaisiana) were resistant to human complement (Table 1, Figure 1), indicating a substantial number of local Borrelia strains with potential for dissemination and neurotropism. Despite the fact that the true identity of the Borrelia species causing LB in humans in Serbia is unknown, as isolates from human materials are lacking at present, it is expected that the same strains present in ticks, such as the isolates examined in the current work, may also be transmitted to humans. Thus, our results based on expressed Osps and resistance to human complement of isolates from ticks may be highly relevant to public health.
5. Conclusions
In conclusion, our results demonstrated that the heterogeneity regarding the presence or absence of Osps of analyzed strains may correspond to different susceptibility to human complement. Since that all serum-reisistant strains (B. afzelii, B. bavariensis and one B. valaisana) expressed OspA and OspB or OspA and OspC, these combinations of proteins may have a role in identification of pathogenic potential of Borrelia strains and ones that express these proteins have potential for dissemination and neurotropism and ability to cause different clinical manifestations of LB in humans. Further research needs to elucidate the mechanisms of resistance of B. valaisiana strains and to explain observed fair motility of B. lusitaniae strains.
Supplementary Materials
Supplementary Materials contain one table and five figures: Table S1 Molecular mass (kDa) of outer surface proteins (OspA, OspB, OspC) in Borrelia burgdorferi sensu lato strains from Serbia, Spain, and Portugal; Figure S1 Protein profiles of seven out of 12 Borrelia afzelii isolates from Serbia. Lane MW - molecular size marker (BioRad, Germany). The strains depicted herein belong to protein profile group I (OspA, OspB and OspC are present in strains RS 32_12b, RS 168_11g, RS 168_11c, RS 232_13b, and RS 235_13cd), and protein profile group II (OspA and OspB are present, OspC is missing in strains RS 230_13c and RS 166_12a); Figure S2 Protein profiles of five out of 12 Borrelia afzelii isolates from Serbia. Lane MW - molecular size marker (BioRad, Germany). The strains depicted herein belong to protein profile group I (OspA, OspB and OspC are present in strains RS 71_11a, RS 164_11a, RS 163_11i, RS 167_11f, and RS 168_11a); Figure S3 Protein profiles of two out of two Borrelia garinii (Bg) and three out of three Borrelia bavariensis (Bb) isolates from Serbia. Lane MW - molecular size marker (BioRad, Germany). The strains depicted herein belong to protein profile group I (OspA, OspB and OspC are present in strains RS 220_10e, RS 160_13e, and RS 163_11h ) and protein profile group III (OspA and OspC are present, OspB is missing in strains RS 164_11g and RS 226_10a); Figure S4 Protein profiles of two out of eight Borrelia lusitaniae and two out of two Borrelia valaisiana isolates from Serbia. Lane MW - molecular size marker (BioRad, Germany). The strains depicted herein belong to protein profile group III (OspA and OspC are present, OspB is missing in strain RS 164_12b) and protein profile group IV (OspA is present, OspB and OspC are missing in strains RS 224_10b, RS 221_10c, and RS 222_10d); Figure S5 Protein profiles of six out of eight Borrelia lusitaniae isolates from Serbia. Lane MW - molecular size marker (BioRad, Germany). The strains depicted herein belong to protein profile group group III (OspA and OspC are present, OspB is missing in strains RS 77_12b, RS 226_10d, and RS 76_12a) and IV (OspA is present, OspB and OspC are missing in RS 167_11c, RS 162_11b, and RS 167_11b).
Author Contributions
All authors contributed to this study. Experimental operation and data analysis were performed by G.V., S.Ć., D.M., R.S., A.S. and E.R.; G.V. and S.T. wrote the manuscript; S.Ć., D.M. and R.S commented on the manuscript. supervision: S.T. and E.RS. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Ministry of Science, Technological Development and Innovation, Republic of Serbia (contract number 451-03-47/2023-01/200015) and an Erasmus Mundus action 2 program Basileus V scholarship for the exchange of PhD students, contract number 2013–2549 (European Union).
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stanek, G.; Wormser, G.P.; Gray, J.; Strle, F. Lyme borreliosis. Lancet. 2012;379:461–73.
- Stanek G, Strle F. Lyme borreliosis–from tick bite to diagnosis and treatment. FEMS Microbiol. Rev. 2018, 42, 233–258.
- Margos, G.; Fingerle, V.; Reynolds, E.S. Borrelia bavariensis: vector switch, niche invasion, and geographical spread of a tick–borne bacterial parasite. Front. Ecol. Evol. 2019, 7:1–20.
- Kraiczy, P. Hide and Seek: How Lyme Disease spirochetes overcome complement attack. Front. Immunol. 2016, 7, 385.
- Schwab, J.; Hammerschmidt, C.; Richter, D.; Skerka, C.; Matuschka, F.R.; Wallich, R.; Zipfel, P.F.; Kraiczy, P. Borrelia valaisiana resist complement–mediated killing independently of the recruitment of immune regulators and inactivation of complement components. PLoS One. 2013, 8, e53659.
- Bhide, M.R. ; Travnicek, M.; Levkutova, M.; Curlik, J.; Revajova, V.; Levkut, M. Sensitivity of Borrelia genospecies to serum complement from different animals and human: a host–pathogen relationship. FEMS Immunol. Med. Microbiol. 2005, 43,165–172.
- van Dam, A,P.; Oei, A.; Jaspars, R.; Fijen, C.; Wilske, B.; Spanjaard, L.; Dankert, J. Complement–mediated serum sensitivity among spirochetes that cause Lyme disease. Infect. Immun. 1997, 65, 1228–1236.
- Estrada-Peña, A,; Cutler, S.; Potkonjak, A.; Vassier-Tussaut, M.; Van Bortel, W.; Zeller, H.; Fernández-Ruiz, N.; Mihalca, A.D. An updated meta–analysis of the distribution and prevalence of Borrelia burgdorferi s.l. in ticks in Europe. Int. J. Health. Geogr. 2018, 17, 41.
- Ćakić, S.; Veinović, G.; Cerar, T.; Mihaljica, D.; Sukara, R.; Ružić-Sabljić, E.; Tomanović, T. Diversity of Lyme borreliosis spirochetes isolated from ticks in Serbia. Med. Vet. Entomol. 2019, 33, 512–520.
- Milutinović, M.; Masuzawa, T.; Tomanović, S.; Radulović, Z.; Fukui, T.; Okamoto, Y. Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum, Francisella tularensis and their co–infections in hostseeking Ixodes ricinus ticks collected in Serbia. Exp. Appl. Acarol. 2008, 45, 171–183.
- Potkonjak, A.; Kleinerman, G.; Gutiérrez, R.; Savić, S.; Vračar, V.; Nachum-Biala, Y.; Jurišić, A.; Rojas, A.; Petrović, A.; Ivanović, I.; Harrus, S.; Baneth, G. Occurrence of Borrelia burgdorferi sensu lato in Ixodes ricinus ticks with first identification of Borrelia miyamotoi in Vojvodina, Serbia. Vector. Borne. Zoonotic. Dis. 2016, 16, 631-635.
- Strnad, M.; Hönig, V.; Růžek, D.; Grubhoffer, L.; Rego, R.O.M. Europe-wide meta-analysis of Borrelia burgdorferi sensu lato prevalence in questing Ixodes ricinus ticks. Appl. Environ. Microbiol. 2017, 83, e00609-17.
- Institute for Public Health of Serbia. Available from: https://www.batut.org.rs/download/izvestaji/Godisnji%20izvestaj%20zarazne%20bolesti%202017.pdf.
- Lohr, B.; Fingerle, V.; Norris, D.E.; Hunfeld, K.P. Laboratory diagnosis of Lyme borreliosis: Current state of the art and future perspectives. Crit. Rev. Clin. Lab. Sci. 2018, 55, 219–245.
- Petrulionienė, A.; Radzišauskienė, D.; Ambrozaitis, A.; Čaplinskas, S.; Paulauskas, A.; Venalis, A. Epidemiology of Lyme disease in a highly endemic European zone. Medicina (Kaunas). 2020, 56, 115.
- Mladenović, J. Epidemiological aspects of Lyme disease on the theritory of Belgrade city. Doctoral dissertation, Faculty of Medicine, University of Nis, Serbia. 2014; 64 [in Serbian].
- Sukara, R.; Chochlakis, D.; Ćirović, D.; Penezić, A.; Mihaljica, D.; Ćakić, S.; Valčić, M.; Tselentis, Y.; Psaroulaki, A.; Tomanović, S . Golden jackals (Canis aureus) as hosts for ticks and tick-borne pathogens in Serbia. Ticks. Tick. Borne. Dis. 2018, 9, 1090-1097.
- Sukara, R.; Juwaid, S.; Ćirović, D.; Penezić, A.; Mihaljica, D.; Veinović, G.; Radojičić, S.; Hodžić, A.; Duscher, G.G.; Tomanović, S. Candidatus Neoehrlichia sp. (FU98) and Borrelia burgdorferi sensu lato in red foxes (Vulpes vulpes) from Serbia. Acta Veterinaria-Beograd. 2019, 69, 312-324.
- Ruzić-Sabljić, E.; Zore, A.; Strle, F. Characterization of Borrelia burgdorferi sensu lato isolates by pulsed-field gel electrophoresis after MluI restriction of genomic DNA. Res. Microbiol. 2008, 159, 441–448.
- Barral, M.; García-Pérez, A.L.; Juste, R.A.; Hurtado, A.; Escudero, R.; Sellek, R.E.; Anda, P. Distribution of Borrelia burgdorferi sensu lato in Ixodes ricinus (Acari: Ixodidae) ticks from the Basque Country, Spain. J. Med. Entomol. 2002, 39, 177–184.
- Escudero, R.; Barral, M.; Pérez, A.; Vitutia, M.M.; García-Pérez, A.L.; Jiménez, S.; Sellek R.E.; Anda, P. Molecular and pathogenic characterization of Borrelia burgdorferi sensu lato isolates from Spain. J. Clin. Microbiol. 2000, 38, 4026–4033.
- Glinšek, U.; Udovič, T.; Cerar, T.; Strle, F.; Ružić–Sabljić, E. Protein profile determination of Borrelia afzelii and Borrelia garinii isolated from skin and cerebrospinal fluid. World. J. Microbiol. Biotechnol. 2009, 25, 1287–1296.
- Ruzić-Sabljić, E.; Maraspin, V.; Lotric-Furlan, S.; Jurca, T.; Logar, M.; Pikelj-Pecnik, A.; Strle, F. Characterization of Borrelia burgdorferi sensu lato strains isolated from human material in Slovenia. Wien. Klin. Wochenschr. 2002, 114, 544–550.
- Zeidner, N.S.; Núncio, M.S.; Schneider, B,S.; Gern, L.; Piesman, J.; Otilia Brandão, O.; Filipe, A.R. A portuguese isolate of Borrelia lusitaniae induces disease in C3H/HeN mice. J. Med. Microbiol. 2001, 50, 1055–1060.
- Collares-Pereira, M.; Couceiro, S.; Franca, I.; Kurtenbach, K.; Schäfer, S.M.; Vitorino, L.; Gonçalves, L.; Baptista, S.; Vieira, M.L.; Cunha, C First isolation of Borrelia lusitaniae from a human patient. J. Clin. Microbiol. 2004, 42, 1316–138.
- Wagemakers, A.; Oei, A.; Fikrig, M.M.; Miellet, W.R.; Hovius, J.W. The relapsing fever spirochete Borrelia miyamotoi is cultivable in a modified Kelly–Pettenkofer medium, and is resistant to human complement. Parasit. Vectors. 2014, 7, 418.
- Koetsveld, J.; Draga, R.O.P.; Wagemakers, A.; Manger, A.; Oei, A.; Visser, C.E.; Hovius, J.W. In vitro susceptibility of the relapsing-fever spirochete Borrelia miyamotoi to antimicrobial agents. Antimicrob. Agents. Chemother. 2017, 61, e00535-17.
- Diza, E.; Papa, A.; Vezyri, E.; Tsounis, S.; Milonas, I.; Antoniadis, A. Borrelia valaisiana in cerebrospinal fluid. Emerg. Infect. Dis. 2004, 10, 1692–1693.
- Rijpkema, S.G.T.; Tazelaar, D.J.; Molkenboer, M,J,C,H.; Noordhoek, G,T.; Plantinga, G.; Schouls, L.M.; Schellekens, J.F.P. Detection of Borrelia afzelii, Borrelia burgdorferi sensu stricto, Borrelia garinii and group VS116 by PCR in skin biopsies of patients with erythema migrans and acrodermatitis chronica atrophicans. Clin. Microbiol. Infect. 1997, 3, 109–116.
- Ružić-Sabljić, E.; Strle, F.; Cimperman, J.; Maraspin, V.; Lotrič-Furlan, S.; Pleterski-Rigler, D. Characterisation of Borrelia burgdorferi sensu lato strains isolated from patients with skin manifestations of Lyme borreliosis residing in Slovenia. J. Med. Microbiol. 2000, 49, 47–53.
- Fingerle, V.; Laux, H.; Munderloh, U.G.; Schulte–Spechtel, U.; Wilske, B. Differential expression of outer surface proteins A and C by individual Borrelia burgdorferi in different genospecies. Med. Microbiol. Immunol. 2000, 189, 59–66.
- Neelakanta, G.; Li, X.; Pal, U.; Liu, X.; Beck, D.S.; DePonte, K.; Fish, D.; Kantor, F.S.; Fikrig, E. Outer surface protein B is critical for Borrelia burgdorferi adherence and survival within Ixodes ticks. PLoS Pathog. 2007, 3, e33.
- Yang, X.F.; Pal, U.; Alani, S.M.; Fikrig, E.; Norgard, M.V. Essential role for OspA/B in the life cycle of the Lyme disease spirochete. J. Exp. Med. 2004, 199, 641–648.
Figure 1.
Serum susceptibiliy of 31 different Borrelia burgdorferi sensu lato strains - comparing median of motile and viable spirochetes in normal human serum (NHS) versus heat-inactivated serum (HIS) after one hour (A) and after three hours of incubation (B). For each Borrelia strain motility between NHS and HIS was compared using a Mann-Whitney test: no significant difference in the motility of each B. afzelii, B. bavariensis and B. valaisiana (RS 164 _12b) between NHS and HIS after one and three hours of incubation (all p > 0.05); significant difference in the motility of each B. lusitaniae, B. garinii and B. valaisiana (RS 224_10b) strain between NHS and HIS after one and after three hours of incubation (all p < 0.05). A Kruskal-Wallis test: significant difference in the motility between Borrelia strains within single species in the presence of NHS after one and three hours of incubation (both p < 0.05). The Mann-Whitney test (in the presence of NHS): significant difference in the motility between B. afzelii (RS 164_11a and RS 168_11g) and other B. afzelii strains and between two B. valaisiana strains (RS 164 _12b and RS 224_10b) after one and three hours of incubation (all p < 0.05); significant difference in the motility between B. lusiataniae (RS 226_10d, RS 167_11c, RS 162 _11b and RS 76_12a, PoHL1, Listu) and other B. lusitaniae strains (all p < 0.05) after one hour of incubation and significant difference in the motility between B. lusitaniae (RS 226_10d, RS 76 _12a, PoHL1) and other B. lusitaniae strains (all p < 0.05) after three hours of incubation; no significant difference in the motility between tick isolate from Serbia (RS 226 _10d ) and human isolate from Portugal (PoHL1) (p > 0.05) and between tick isolate from Serbia (RS 76 _12a) and human isolate from Portugal (PoHL1) (p > 0.05) after three hours of incubation. In the presence of NHS, after one and three hours of incubation, B. valaisiana (RS 224_10b) significantly less motile (all p < 0.05) than each B. afzelii, B. bavariensis, and another B. valaisiana strain (RS 164_12b) but significantly more motile (all p < 0.05) than all B. lusianiae and B. garinii strains.  B. afzelii in HIS;
B. afzelii in HIS;  B. afzelii in NHS;
B. afzelii in NHS;  B. garinii in HIS;
B. garinii in HIS;  B. garinii in NHS;
B. garinii in NHS;  B. bavariensis in HIS;
B. bavariensis in HIS; B. bavariensis in NHS;
B. bavariensis in NHS;  B. valaisiana in HIS;
B. valaisiana in HIS;  B. valaisiana in NHS;
B. valaisiana in NHS;  B. lusitaniae in HIS;
B. lusitaniae in HIS;  B. lusitaniae in NHS.
B. lusitaniae in NHS.
B. afzelii in HIS; B. afzelii in NHS; B. garinii in HIS; B. garinii in NHS; B. bavariensis in HIS; B. bavariensis in NHS; B. valaisiana in HIS; B. valaisiana in NHS; B. lusitaniae in HIS; B. lusitaniae in NHS.
Figure 1.
Serum susceptibiliy of 31 different Borrelia burgdorferi sensu lato strains - comparing median of motile and viable spirochetes in normal human serum (NHS) versus heat-inactivated serum (HIS) after one hour (A) and after three hours of incubation (B). For each Borrelia strain motility between NHS and HIS was compared using a Mann-Whitney test: no significant difference in the motility of each B. afzelii, B. bavariensis and B. valaisiana (RS 164 _12b) between NHS and HIS after one and three hours of incubation (all p > 0.05); significant difference in the motility of each B. lusitaniae, B. garinii and B. valaisiana (RS 224_10b) strain between NHS and HIS after one and after three hours of incubation (all p < 0.05). A Kruskal-Wallis test: significant difference in the motility between Borrelia strains within single species in the presence of NHS after one and three hours of incubation (both p < 0.05). The Mann-Whitney test (in the presence of NHS): significant difference in the motility between B. afzelii (RS 164_11a and RS 168_11g) and other B. afzelii strains and between two B. valaisiana strains (RS 164 _12b and RS 224_10b) after one and three hours of incubation (all p < 0.05); significant difference in the motility between B. lusiataniae (RS 226_10d, RS 167_11c, RS 162 _11b and RS 76_12a, PoHL1, Listu) and other B. lusitaniae strains (all p < 0.05) after one hour of incubation and significant difference in the motility between B. lusitaniae (RS 226_10d, RS 76 _12a, PoHL1) and other B. lusitaniae strains (all p < 0.05) after three hours of incubation; no significant difference in the motility between tick isolate from Serbia (RS 226 _10d ) and human isolate from Portugal (PoHL1) (p > 0.05) and between tick isolate from Serbia (RS 76 _12a) and human isolate from Portugal (PoHL1) (p > 0.05) after three hours of incubation. In the presence of NHS, after one and three hours of incubation, B. valaisiana (RS 224_10b) significantly less motile (all p < 0.05) than each B. afzelii, B. bavariensis, and another B. valaisiana strain (RS 164_12b) but significantly more motile (all p < 0.05) than all B. lusianiae and B. garinii strains. B. afzelii in HIS; B. afzelii in NHS; B. garinii in HIS; B. garinii in NHS; B. bavariensis in HIS; B. bavariensis in NHS; B. valaisiana in HIS; B. valaisiana in NHS; B. lusitaniae in HIS; B. lusitaniae in NHS.
B. afzelii in HIS; B. afzelii in NHS; B. garinii in HIS; B. garinii in NHS; B. bavariensis in HIS; B. bavariensis in NHS; B. valaisiana in HIS; B. valaisiana in NHS; B. lusitaniae in HIS; B. lusitaniae in NHS.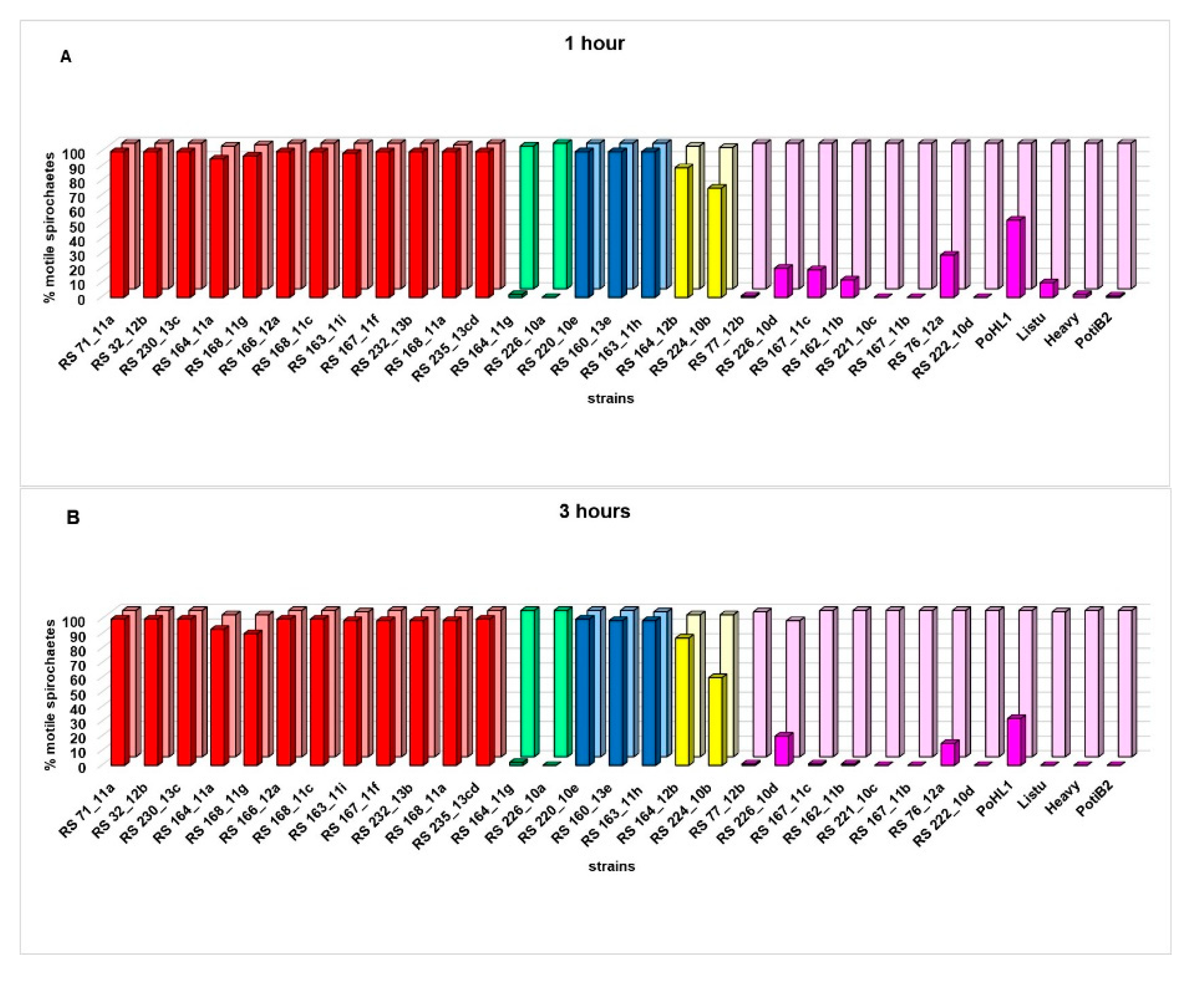

Table 1.
A range and median percentage of motile spirochetes for each Borrelia burgdorferi sensu lato strains in the presence of normal human serum (NHS) and heat-inactivated serum (HIS) after one (1h) and three hours (3h) of incubation.
Table 1.
A range and median percentage of motile spirochetes for each Borrelia burgdorferi sensu lato strains in the presence of normal human serum (NHS) and heat-inactivated serum (HIS) after one (1h) and three hours (3h) of incubation.
| Strain | Region (host) | NHS (1h) Median (range)a % |
HIS (1h) Median (range) % |
p- Value |
NHS (3h) Median (range) % |
HIS (3h) Median (range) % |
p- Value |
Susceptibility to NHS |
|---|---|---|---|---|---|---|---|---|
| Borrelia valaisana | ||||||||
| RS 164_12b | Serbia (tick) | 89 (85-91) | 98 (98-98) | >0.05 | 87 (77-89) | 97 (96-97) | >0.05 | Rb |
| RS 224_10b | Serbia (tick) | 75 (53-81) | 97 (96-99) | <0.05 | 60 (52-76) | 97 (95-97) | <0.05 | Sc |
| median (range) | 83 (53-91) | 98 (96-99) | 76.5 (52-89) | 97 (95-97) | ||||
| Borrelia afzelii | ||||||||
| RS 71_11a | Serbia (tick) | 100 (100-100) | 100 (100-100) | >0.05 | 100 (100-100) | 100 (100-100) | >0.05 | R |
| RS 32_12b | Serbia (tick) | 100 (100-100) | 100 (100-100) | >0.05 | 100 (100-100) | 100 (100-100) | >0.05 | R |
| RS 230_13c | Serbia (tick) | 100 (100-100) | 100 (100-100) | >0.05 | 100 (100-100) | 100 (100-100) | >0.05 | R |
| RS 164_11a | Serbia (tick) | 95 (94-98) | 98 (98-98) | >0.05 | 93 (90-93) | 97 (97-97) | >0.05 | R |
| RS 168_11g | Serbia (tick) | 97 (97-98) | 99 (98-99) | >0.05 | 90 (88-94) | 97 (94-97) | >0.05 | R |
| RS 166_12a | Serbia (tick) | 100 (100-100) | 100 (100-100) | >0.05 | 100 (100-100) | 100 (100-100) | >0.05 | R |
| RS 168_11c | Serbia (tick) | 100 (100-100) | 100 (100-100) | >0.05 | 100 (100-100) | 100 (100-100) | >0.05 | R |
| RS 163_11i | Serbia (tick) | 99 (98-100) | 100 (99-100) | >0.05 | 99 (97-100) | 99 (98-100) | >0.05 | R |
| RS 167_11f | Serbia (tick) | 100 (99-100) | 100 (100-100) | >0.05 | 99 (98-100) | 100 (99-100) | >0.05 | R |
| RS 232_13b | Serbia (tick) | 100 (99-100) | 100 (100-100) | >0.05 | 99 (98-100) | 100 (99-100) | >0.05 | R |
| RS 168_11a | Serbia (tick) | 100 (98-100) | 99 (100-100) | >0.05 | 99 (98-100) | 100 (99-100) | >0.05 | R |
| RS 235_13cd | Serbia (tick) | 100 (100-100) | 100 (100-100) | >0.05 | 100 (100-100) | 100 (100-100) | >0.05 | R |
| median (range) | 100 (94-100) | 100 (98-100) | 100 (90-100) | 100 (94-100) | ||||
| Borrelia garinii | ||||||||
| RS 164_11g | Serbia (tick) | 2 (0-3) | 98 (97-99) | <0.05 | 2 (0-2) | 98 (97-99) | <0.05 | S |
| RS 226_10a | Serbia (tick) | 0 (0-0) | 100 (100-100) | <0.05 | 0 (0-0) | 100 (100-100) | <0.05 | S |
| median (range) | 0 (0-3) | 99.5 (97-100) | 0 (0-2) | 99.5 (97-100) | ||||
| Borrelia bavariensis | ||||||||
| RS 220_10e | Serbia (tick) | 100 (100-100) | 100 (100-100) | >0.05 | 100 (100-100) | 100 (100-100) | >0.05 | R |
| RS 160_13e | Serbia (tick) | 100 (100-100) | 100 (100-100) | >0.05 | 99 (96-100) | 99 (99-99) | >0.05 | R |
| RS 163_11h | Serbia (tick) | 100 (100-100) | 100 (100-100) | >0.05 | 99 (99-100) | 100 (100-100) | >0.05 | R |
| median (range) | 100 (100-100) | 100 (100-100) | 100 (96-100) | 100 (99-100) | ||||
| Borrelia lusitaniae | ||||||||
| RS 77_12b | Serbia (tick) | 1 (0-2) | 100 (100-100) | <0.05 | 1 (0-1) | 99 (99-100) | <0.05 | S |
| RS 226_10d | Serbia (tick) | 20 (4-50) | 100 (92-100) | <0.05 | 20 (3-45) | 93 (91-94) | <0.05 | S |
| RS 167_11c | Serbia (tick) | 19 (15-20) | 100 (100-100) | <0.05 | 1 (0-1) | 100 (100-100) | <0.05 | S |
| RS 162_11b | Serbia (tick) | 12 (3-14) | 100 (100-100) | <0.05 | 1 (1-2) | 100 (100-100) | <0.05 | S |
| RS 221_10c | Serbia (tick) | 0 (0-7) | 100 (100-100) | <0.05 | 0 (0-0) | 100 (100-100) | <0.05 | S |
| RS 167_11b | Serbia (tick) | 0 (0-0) | 100 (100-100) | <0.05 | 0 (0-0) | 100 (98-100) | <0.05 | S |
| RS 76_12a | Serbia (tick) | 29 (26-35) | 100 (100-100) | <0.05 | 15 (5-20) | 100 (100-100) | <0.05 | S |
| RS 222_10d | Serbia (tick) | 0 (0-0) | 100 (100-100) | <0.05 | 0 (0-0) | 100 (100-100) | <0.05 | S |
| PoHL1 | Portugal (human) | 53 (47-70) | 100 (100-100) | <0.05 | 32 (27-50) | 100 (80-100) | <0.05 | S |
| Listu | Spain (tick) | 10 (5-18) | 100 (99-100) | <0.05 | 0 (0-3) | 99 (98-100) | <0.05 | S |
| Heavy | Spain (tick) | 2 (0-5) | 100 (100-100) | <0.05 | 0 (0-1) | 100 (100-100) | <0.05 | S |
| PotiB2 | Portugal (tick) | 1 (1-3) | 100 (100-100) | <0.05 | 0 (0-1) | 100 (100-100) | <0.05 | S |
| median (range) | 4.5 (0-70) | 100 (92-100) | 1 (0-50) | 100 (91-100) |
p-values under 0.05 are in bold. a Median percentage of viable, motile spirochetes and the range were determined for each Borrelia strains in normal human serum (NHS) and heat-inactivated serum (HIS) after one and three hours of incubations. For each strain experiment was repeated at least three times. b R-Borrelia strains resistant to NHS after one and three hours of incubation: presence of viable, motile spirochetes in NHS compared with HIS indicates resistance of Borrelia strains to NHS. c S- Borrelia strains susceptible to NHS after one and three hours of incubation: loss of motility of spirochetes in NHS compared with HIS indicates susceptibility of Borrelia strains to NHS.
Table 2.
Frequency of presence of outer surface proteins (OspA, OspB, and OspC) in Borrelia burgdorferi sensu lato strains.
Table 2.
Frequency of presence of outer surface proteins (OspA, OspB, and OspC) in Borrelia burgdorferi sensu lato strains.
| Species | OspA | OspB | OspC |
|---|---|---|---|
| B. afzelii | 12 (100%) | 12 (100%) | 10 (83%) |
| B. garinii | 2 (100%) | 2 (100%) | |
| B. bavariensis | 3 (100%) | 3 (100%) | 3 (100%) |
| B. valaisiana | 2 (100%) | 1 (50%) | |
| B. lusitaniae | 11 (92) | 2 (17%) | 4 (33%) |
| All | 30 (97%) | 17 (55%) | 20 (65%) |
Table 3.
Frequency of present outer surface proteins (OspaA, OspB, and OspC) and protein profile group (I-V).
Table 3.
Frequency of present outer surface proteins (OspaA, OspB, and OspC) and protein profile group (I-V).
| Present protein (Protein profile group) |
B. afzelii No. 12 |
B. garinii No. 2 |
B. bavariensis No. 3 |
B. valaisiana No. 2 |
B. lusitaniae No. 12 |
All No. 31 |
|---|---|---|---|---|---|---|
| OspA, OspB, and OspC (I) |
10 (83%) RS 71_11a RS 32_12b RS 164_11a RS 168_11g RS 168_11c B RS 167_11f RS 232_13b RS 168_11a RS 235_13cd |
3 (100%) RS 220_10e RS 160_13e RS 163_11h |
1 (8%) PoHL1 |
14 (45%) | ||
|
Osp A and OspB (II) |
2 (17%) RS 230_13c RS 166_12a |
2 (7%) |
||||
| OspA and OspC (III) |
2 (100%) RS 164_11g RS 226_10a |
1 (50%) RS 164_12b |
3 (25%) RS 77_12b RS 226_10d RS 76_12a |
6 (19%) | ||
|
OspA (IV) |
1 (50%) RS 224_10b |
7 (59%) RS 167_11c RS 162_11b RS 221_10c RS 167_11b RS 222_10d Listu Heavy |
8 (26%) |
|||
| OspB (V) |
1 (8%) PotiB2 |
1 (3%) |
Borrelia strains resistant to normal human serum are in bold.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



