Submitted:
12 September 2023
Posted:
14 September 2023
You are already at the latest version
Abstract
Colistin is used in treatment against gram-negative bacteria. It causes toxic effects due to oxidative stress in a dose-dependent manner. Naringin is a flavonoid with strong antioxidant properties found naturally in many plants, especially citrus fruits. This study aimed to determine the protective effects of NRG in CLS-induced sciatic nerve injury. Thirty-five male Spraque rats were randomly divided into five groups: control, NRG, CLS, CLS+NRG50, and CLS+NRG100. CLS was administered orally at a 15mg/kg for seven days. NRG was administered orally at 50 or 100 mg/kg for seven days. CLS caused toxic damage in sciatic nerve tissue by increasing MDA and decreasing antioxidant capacity (SOD, CAT, GPx, and GSH); increasing NF-κB, TNF-α, nNOS release; increasing apoptotic factor (Caspase-3); decreasing HO-1 and nCAM; and inducing 8-OHdG, MAPK and GFAP. NRG, on the other hand, reversed all these changes in sciatic nerve tissue and reduced CLS-induced oxidative stress damage, inflammatory damage and apoptotic damage. These effects were more effective at the 100mg/kg dose of NRG than at the 50mg/kg dose. In sciatic nerve tissue, CLS-induced sciatic nerve toxicity with increased oxidative stress, inflammation, and apoptotic damage. NRG showed a protective effect against CLS-induced sciatic nerve toxicity.
Keywords:
apoptosis
; colistin
; inflammation
; naringin
; oxidative stress
; sciatic nerve
1. Introduction
Colistin (Polymyxin E, CLS) is an antibiotic drug used in the treatment of infections caused by gram-negative bacteria such as Escherichia coli since the 1960s [1]. When CLS acts on bacteria, it interacts with anionic lipopolysaccharides in the bacterial membrane, translocates calcium and magnesium, and neutralizes the bacteria [2]. The use of CLS also produces toxic effects. Among these toxic effects, two important side effects such as neurotoxicity and nephrotoxicity lead to dose limitation or discontinuation of treatment [3]. In previous studies in mice, CLS (15mg/kg, 7 days, i.p.) was reported to cause damage to brain and sciatic nerve tissues [4,5]. In studies of CLS, there is increasing evidence that oxidative stress and mitochondrial damage underlie the mechanism of neurotoxicity [6].
Compounds found naturally in plants, which stand out with their antioxidant properties, protect tissues in the body against damage [7]. Flavonoids are ingredients that are taken daily in the diet and are of interest in research due to their key effects such as anti-oxidant and anti-inflammatory [8]. Naringin (NRG) is a flavanone glycoside derived from the flavanone of naringenin. It occurs naturally in many plants, especially citrus fruits [9]. NRG have gained importance in traditional medicine due to their non-addictive and non-toxic nature [10]. Studies have reported that NRG exhibits antioxidant properties by increasing the activities of antioxidants in different tissues. In addition, NRG is also protective against nitrosative stress by inhibiting the activity of nitric oxide synthase enzyme (eNOS) and removing nitric oxide-derived free radicals [11]. It has also been found to have antioxidant and anti-inflammatory effects in studies on neurodegenerative disorders [12]. The amount of NRG in grapefruit averages 17 mg/100 g of edible fruit. However, studies have reported that taking NRG in the amount of 400 mg/day increases antioxidant capacity in the body [13].
The aim of this study was to investigate the effects of CLS on oxidative stress, inflammatory and apoptotic damage pathways in rat sciatic nerve tissues and the protective properties of NRG against these effects of CLS.
2. Results
2.1. Oxidant and Antioxidant Status Findings
It was found that SOD, CAT and GPx were inhibited and antioxidant activity decreased by decreasing GSH in CLS compared to the control (p<0.05). In addition, lipid peroxidation was manifested by an increase in MDA (p<0.05). Antioxidant enzyme activities and GSH increased and MDA decreased in CLS+NRG50 and CLS+NRG-100 compared to CLS (p<0.05). NRG showed more pronounced effects at a dose of 100mg/kg (p<0.05). (Figure 1).
2.2. Inflammation Markers findings
Inflammation-related NF-κB, TNF-α, and nNOS in sciatic nerve tissues were analyzed by ELISA. There was a increase in NF-κB, TNF-α, and nNOS in the CLS compared to the control (p<0.05). CLS+NRG50 and CLS+NRG100 showed a decrease in all these parameters compared to CLS (p<0.05). When different doses of NRG were compared, 100 mg/kg was found to be more effective (p<0.05) (Figure 2).
2.3. Findings of HO-1 and 8−OHdG Levels
Compared to the control, HO-1 level decreased in the CLS, while 8-OHdG level increased (p<0.05). With NRG administration, this situation was reversed in CLS+NRG50 and CLS+NRG100 compared to CLS (p<0.05). When different doses of NRG were compared, 100 mg/kg was found to be more effective (p<0.05) (Figure 3).
2.4. Findings of MAPK14, nCAM and GFAP Levels
In CLS, nCAM was inhibited and MAPK and GFAP were increased compared to control (p<0.05). In CLS+NRG50 and CLS+NRG100, nCAM levels increased and MAPK and GFAP levels decreased compared to CLS (p<0.05). NRG showed more pronounced effects at 100mg/kg dose (p<0.05) (Figure 4).
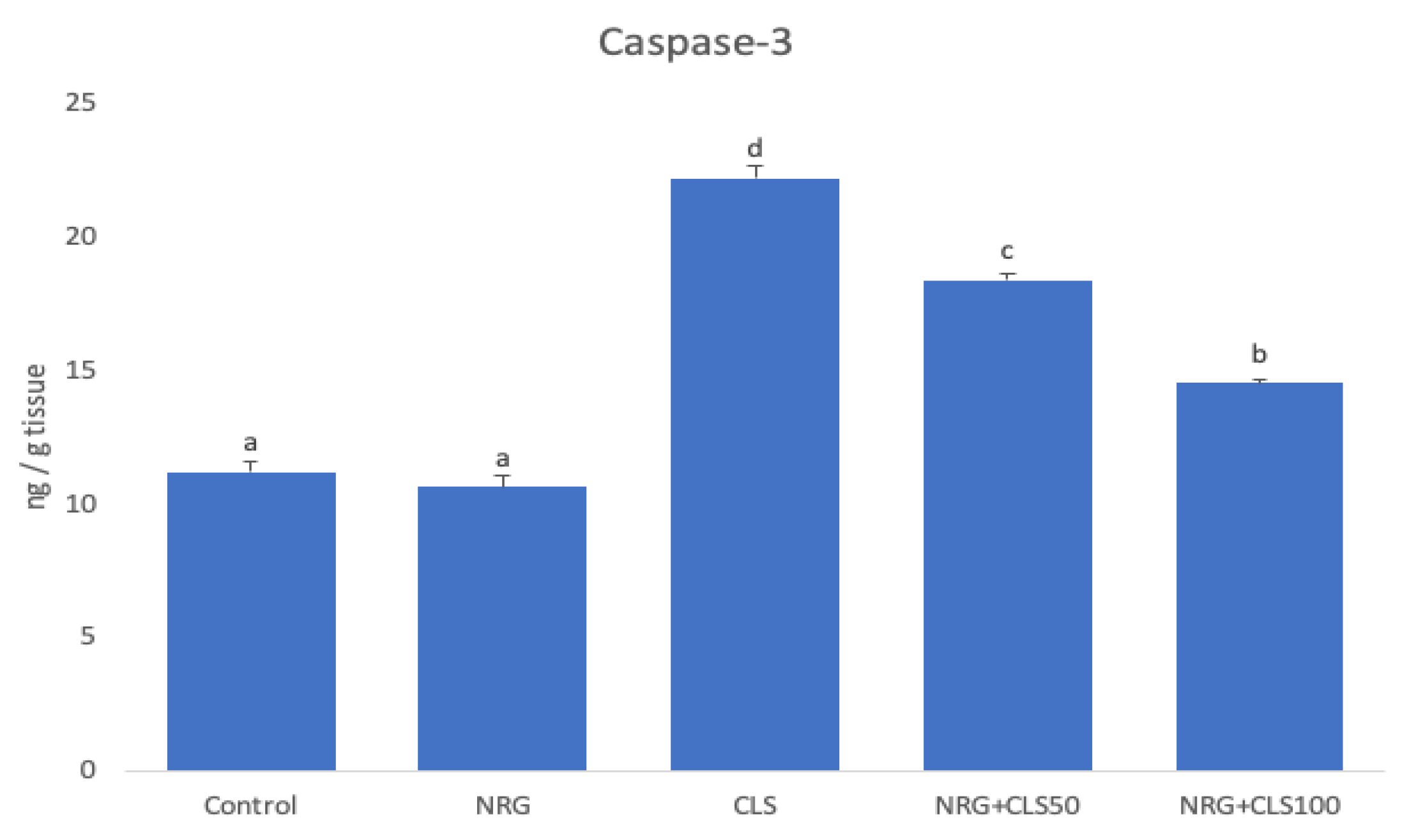
2.5. Apoptotic Markers Findings
In CLS group, there was an increase in Caspase-3 (p<0.05). In CLS+NRG50 and CLS+NRG100 groups, the change in Caspase-3 was reversed (p<0.05). NRG showed more pronounced effects at a dose of 100mg/kg (Figure 5).
3. Discussion
CLS is a drug used to treat drug-resistant gram-negative bacteria but has a dose-dependent toxic effect. It is hypothesized that oxidative stress plays a role in its toxic effects on tissues [14]. There are many antioxidants that reduce oxidative stress by inhibiting ROS production [15]. Flavonoids are naturally occurring compounds found in many plants such as fruits, vegetables, cereals, teas, etc. and have numerous biological effects, including neuroprotective effects [16]. NRG is an active flavanone glycoside found in plants, especially in citrus group plants, and stands out with its antioxidant properties [17]. Therefore, in the present study, the effects of NRG on CLS-induced sciatic nerve tissue toxicity in rats were investigated.
While the balance disrupted as a result of ROS increase causes oxidative stress to damage tissues, antioxidant defense systems protect tissues against this damage [18]. MDA, an indicator of lipid peroxidation, occurs as a result of peroxidation of fatty acids containing three or more double bonds and affects ion exchange across cell membranes [19,20]. Antioxidant enzymes SOD, CAT and GPx are the first line of defense against oxidative stress damage caused by ROS [21]. Another line of defense, GSH, is a tripeptide that acts as the main redox buffer in the cell in oxidative stress damage [22]. In this study, CLS increased oxidative stress by increasing MDA levels and decreasing SOD, CAT and GPx activity and GSH levels in sciatic nerve tissues. NRG administration, on the other hand, increased antioxidant activity by decreasing CLS-induced increased MDA levels and increasing antioxidant enzyme activities and GSH levels. This antioxidant activity was stronger, especially at a 100 mg/kg dose.
If oxidative stress is prolonged, inflammation also occurs [23]. During inflammation, NF-κB is the first transcription factor to be induced in tissues and accelerates the inflammatory process by stimulating pro-inflammatory cytokines such as TNF-α [24]. The triggering of NO production due to alterations in NOS isoforms activates pathological processes such as shock and pain associated with neurotoxicity. nNOS is one of the 3 isoforms of NOS [13]. In the current study, CLS caused inflammatory damage in sciatic nerve tissues by increasing NF-κB, TNF-α and nNOS. NRG administration, on the other hand, reduced CLS-induced inflammatory damage by reducing NF-κB, TNF-α and nNOS. NRG could emerge as an effective therapeutic agent in sciatic nerve tissue toxicity caused by inflammation due to CLS exposure.
MAPK14 is an important member of the MAPK family that is stimulated in cells as a result of increased ROS [13]. MAPK14 induces IκB to become phophorylated and subsequently causes IκB dissociation from NF-κB [25]. nCAM is a glycoprotein released in the central nervous system and is involved in cell-cell and cell-extracellular matrix interactions [26]. GFAP is an intermediate filament protein located in glial cells (astrocytes, ependymal cells, etc.). GFAP increases with the occurrence of damage and toxic conditions in neuronal cells [27]. In the present study, CLS increased MAPK14 and GFAP levels, while decreasing nCAM levels. NRG administration reversed this situation, decreased MAPK14 and GFAP levels and provided support to the nCAM activity.
8-OHdG is one of the indicators of oxidative DNA damage caused by increased free radicals [28]. HO-1 activity has been shown to regulate key cellular activities such as inflammation and apoptosis [29]. HO-1 plays an active role in cellular defense against cellular damage and shows protective properties, especially by preventing ROS-induced damage [13]. In this study, CLS increased the level of 8-OHdG, an indicator of oxidative DNA damage, and decreased the level of protective HO-1 in sciatic nerve tissues. NRG administration reversed this situation, decreased DNA damage and provided support to the defense system.
Intracellular increase in ROS may trigger cellular aging and apoptosis by damaging macromolecules [27]. Apoptosis (programmed cell death) is an essential physiological process that destroys and removes damaged or dangerous cells in the body. In healthy tissue cells, cellular stress or injury is required to trigger apoptosis [30]. The increase of oxygen radicals negatively affects cellular activities related to intracellular signaling, such as the apoptotic pathway [31]. While caspase family members are inactive in normal healthy cells, they are activated when the death signal arrives. Caspase-3, the most important caspase, is also known as executioner caspase and is pro-apoptotic [32]. In the this study, levels of Caspase-3, which is an apoptotic factor, increased with CLS exposure in sciatic nerve tissues. When NRG was administered together with CLS, the opposite effect was observed and NRG exhibited antiapoptotic properties. Therefore, NRG may be an effective agent against apoptosis in CLS-induced sciatic nerve tissue toxicity.
4. Materials and Methods
4.1. Chemicals
CLS was purchased from Koçak Pharmaceuticals (Colimycin, 150 mg/flakon, Istanbul/Türkiye). NRG and all other chemicals (analytical purities) were obtained from Sigma Chemical Co. (St. Louis, USA).
Experimental Procedure
Thirty-five male rats (Sprague dawley, 220-250g, 10-12 weeks) obtained from Atatürk University Experimental Animal Center (Erzurum, Türkiye) were used in the experiments. Rats were housed under standard laboratory conditions (12-h light and dark cycle, ventilation, 23±2oC, standard cage). Unlimited access to food and drinking water was provided. Rats were randomly divided into 5 groups (n=7). CLS and NRG doses were determined from the literature [3,33].
1- Control (CNT): Saline was given orally and intraperitoneally (i.p.) for seven days
2- Naringin (NRG): NRG was given orally at a dose of 100 mg/kg once daily for 7 days.
3- Colistin (CLS): CLS was given intraperitoneally (i.p.) at a dose of 15 mg/kg once daily for 7 days.
4- Colistin+Naringin 50 (CLS+NRG50): For 7 days, CLS was given i.p. at a dose of 15 mg/kg followed by NRG 50 mg/kg orally.
5- Colistin+Naringin 100 (CLS+NRG100): For 7 days, CLS was given i.p. at a dose of 15 mg/kg followed by NRG 100 mg/kg orally.
4.2. Collection of samples
The sciatic nerve tissue was removed 24 hours after the last drug administration (day 8), separated from the surrounding tissues, washed in physiologic saline solution and stored.
4.3. Lipid peroxidation analysis
To determine the level of malondialdehyde (MDA) in sciatic nerve tissues, 532 nm absorbance was measured after reaction with thiobarbituric acid. For MDA analysis of sciatic nerve tissues, homogenization was performed according to the previous method [34]. For the analysis of MDA levels, the method in the literature was used [35].
4.4. Antioxidant analysis
Catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPx) activities and glutathione (GSH) levels were analyzed to analyze the antioxidant status of sciatic nerve tissue. For antioxidants markers analysis of sciatic nerve tissues, homogenization was performed according to the previous method [7]. SOD [36], CAT [37], GPx [38], and GSH [39] were determined using the literature. Lowry et al. method was used for protein analysis [40].
4.5. Analysis of Inflammatory Markers
Cytokine production in sciatic nerve tissue was determined by ELISA using commercial kits in accordance with the procedure. Supernatants obtained from homogenates prepared with phosphate buffer (pH 7.4, 0.1 M) were used in the analysis. Nuclear Factor kappa B (NF-κB), Tumor necrosis factor alpha (TNF-α), neuronal nitric oxide synthase (nNOS) levels were determined from sciatic nerve tissue using a rat ELISA kit (NF-κB and TNF-α: Sunred, China; nNOS: YL Biont, China).
4.6. Analysis of MAPK14, nCAM and GFAP Levels
Mitogen-activated protein kinase 14 (MAPK14), neural cell adhesion molecule (nCAM), and Glial fibrillary acidic protein (GFAP) in sciatic nerve tissue was determined by ELISA using commercial kits in accordance with the procedure. Supernatants obtained from homogenates prepared with phosphate buffer (pH 7.4, 0.1 M) were used in the analysis. MAPK14, nCAM, and GFAP levels were determined from sciatic nerve tissue using a rat ELISA kit (Sunred, China).
4.7. Analysis of HO-1 and 8−OHdG Levels
Heme oxygenase-1 (HO-1), and 8-hydroxy-2′-deoxyguanosine (8−OHdG) in sciatic nerve tissue was determined by ELISA using commercial kits in accordance with the procedure. Supernatants obtained from homogenates prepared with phosphate buffer (pH 7.4, 0.1 M) were used in the analysis. HO-1 and 8−OHdG levels were determined from sciatic nerve tissue using a rat ELISA kit (Sunred, China).
4.8. Analysis of Apoptotic Marker
Supernatants obtained from homogenates prepared with phosphate buffer (pH 7.4, 0.1 M) were used for apoptotic damage level analysis in sciatic nerve tissue. Caspase-3 levels were determined from sciatic nerve tissue using a rat ELISA kit (YL Biont, China).
4.9. Statistical analysis
Statistical analysis of the data obtained from sciatic nerve tissues was performed with SPSS 20.0 (IBM, NY) program. One-way ANOVA and Tukey's post hoc tests were used for comparison between groups. Data are presented as mean±SE. Statistical significance was accepted: p<0.05.
5. Conclusions
In conclusion, CLS caused toxic effects by increasing inflammation, oxidative stress, and apoptotic damage levels in sciatic nerve tissue. On the other hand, NRG was found to reduce the toxic effect by decreasing all these damages. It can be concluded that administration of NRG, especially at the level of 100mg/kg, will be much more effective in preventing sciatic nerve tissue damage in terms of all these pathways and improving the quality of life of the patient.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was approved by Atatürk University Experimental Animal Ethics Committee (2023/08/119).
Informed Consent Statement
Not applicable.
Data Availability Statement
None.
Acknowledgments
None.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aksu EH, Kandemir FM, Küçükler S, Mahamadu A. Improvement in colistin-induced reproductive damage, apoptosis, and autophagy in testes via reducing oxidative stress by chrysin. J Biochem Mol Toxicol 2018 ; 32(11) : e22201. [CrossRef]
- Aksu EH, Kandemir FM, Küçükler S. The effects of hesperidin on colistin-induced reproductive damage, autophagy, and apoptosis by reducing oxidative stress. Andrologia 2021 ; 53(2) : e13900. [CrossRef]
- Çelik H, Kandemir FM, Caglayan C, Özdemir S, Çomaklı S, Kucukler S, et al. Neuroprotective effect of rutin against colistin-induced oxidative stress, inflammation and apoptosis in rat brain associated with the CREB/BDNF expressions. Mol Biol Rep 2020 ; 47(3) : 2023-2034. [CrossRef]
- Dai C, Li J, Lin W, Li G, Sun M, Wang F, et al. Electrophysiology and ultrastructural changes in mouse sciatic nerve associated with colistin sulfate exposure. Toxicol Mech Methods 2012 ; 22(8) : 592-6. [CrossRef]
- Dai C, Li J, Li J. New insight in colistin induced neurotoxicity with the mitochondrial dysfunction in mice central nervous tissues. Exp Toxicol Pathol 2013 ; 65(6) : 941-8. [CrossRef]
- Dai C, Tang S, Biao X, Xiao X, Chen C, Li J. Colistin induced peripheral neurotoxicity involves mitochondrial dysfunction and oxidative stress in mice. Mol Biol Rep 2019 ; 46(2) : 1963-72. [CrossRef]
- Caglayan C, Kandemir FM, Ayna A, Gür C, Küçükler S, Darendelioğlu E. Neuroprotective effects of 18β-glycyrrhetinic acid against bisphenol A-induced neurotoxicity in rats: involvement of neuronal apoptosis, endoplasmic reticulum stress and JAK1/STAT1 signaling pathway. Metab Brain Dis 2022 ; 37(6) : 1931-40. [CrossRef]
- Varışlı B, Caglayan C, Kandemir FM, Gür C, Ayna A, Genç A, et al. Chrysin mitigates diclofenac-induced hepatotoxicity by modulating oxidative stress, apoptosis, autophagy and endoplasmic reticulum stress in rats. Mol Biol Rep 2023 ; 50(1) : 433-42. [CrossRef]
- Ghanbari-Movahed M, Jackson G, Farzaei MH, Bishayee A. A Systematic Review of the Preventive and Therapeutic Effects of Naringin Against Human Malignancies. Front Pharmacol 2021 ; 12: 639840. [CrossRef]
- Akintunde JK, Akintola TE, Adenuga GO, Odugbemi ZA, Adetoye RO, Akintunde OG. Naringin attenuates Bisphenol-A mediated neurotoxicity in hypertensive rats by abrogation of cerebral nucleotide depletion, oxidative damage and neuroinflammation. Neurotoxicology 2020 ; 81: 18-33. [CrossRef]
- Kaur G, Prakash A. Involvement of the nitric oxide signaling in modulation of naringin against intranasal manganese and intracerbroventricular β-amyloid induced neurotoxicity in rats. J Nutr Biochem 2020 ; 76 : 108255. [CrossRef]
- Garabadu D, Agrawal N. Naringin Exhibits Neuroprotection Against Rotenone-Induced Neurotoxicity in Experimental Rodents. Neuromolecular Med. 2020 ; 22(2) : 314-30. [CrossRef]
- Semis HS, Kandemir FM, Caglayan C, Kaynar O, Genc A, Arıkan SM. Protective effect of naringin against oxaliplatin-induced peripheral neuropathy in rats: A behavioral and molecular study. J Biochem Mol Toxicol 2022 ; 36(9) : e23121. [CrossRef]
- Hanedan B, Ozkaraca M, Kirbas A, Kandemir FM, Aktas MS, Kilic K, et al. Investigation of the effects of hesperidin and chrysin on renal injury induced by colistin in rats. Biomed Pharmacother 2018 ; 108 : 1607-16. [CrossRef]
- Akaras N, Abuc OO, Koc K, Bal T, Geyikoglu F, Atilay H, et al. (1 → 3)-β-d-glucan enhances the toxicity induced by Bortezomib in rat testis. J Food Biochem 2020 ; 44(3) : e13155. [CrossRef]
- Thayumanavan G, Jeyabalan S, Fuloria S, Sekar M, Ravi M, Selvaraj LK, et al. Silibinin and naringenin against bisphenol a-ınduced neurotoxicity in Zebrafish model-potential flavonoid molecules for new drug design, development, and therapy for neurological disorders. Molecules 2022 ; 27(8) : 2572. [CrossRef]
- Kandemir FM, Kucukler S, Caglayan C, Gur C, Batil AA, Gülçin İ. Therapeutic effects of silymarin and naringin on methotrexate-induced nephrotoxicity in rats: Biochemical evaluation of anti-inflammatory, antiapoptotic, and antiautophagic properties. J. Food Biochem 2017; 41(5) , e12398. [CrossRef]
- Sagliyan A, Benzer F, Kandemir FM, Gunay C, Han MC, Ozkaraca M. Beneficial effects of oral administrations of grape seed extract on healing of surgically induced skin wounds in rabbits. Revue Méd. Vét 2012 ; 163(1) : 11-17.
- Ömür AD, Kandemir FM, Yıldırım BA, Akman O, Şenocak EA, Eldutar E, et al. Protective effect of dandelion (Taraxacum officinale) extract Against gentamicin-induced reproductive damage in male rats. Kafkas Univ. Vet. Fak 2016 ; 22 : 929-36.
- Ileriturk M, Kandemir O, Akaras N, Simsek H, Genc A, Kandemir FM. Hesperidin has a protective effect on paclitaxel-induced testicular toxicity through regulating oxidative stress, apoptosis, inflammation and endoplasmic reticulum stress. Reprod Toxicol 2023 ; 118 : 108369. [CrossRef]
- Ileritürk M., Kandemir Ö. Protective Effect of Rutin on Malathion-induced Gastric Toxicity: Evaluation of Oxidative Stress, Inflammation and Apoptosis. F.U. Vet. J. Health Sci 2023 ; 37(2) : 139-45.
- Şimşek H, Akaras N, Gür C, Küçükler S, Kandemir FM. Beneficial effects of Chrysin on Cadmium-induced nephrotoxicity in rats: Modulating the levels of Nrf2/HO-1, RAGE/NLRP3, and Caspase-3/Bax/Bcl-2 signaling pathways. Gene 2023 ; 875 : 147502. [CrossRef]
- Gur C, Akarsu SA, Akaras N, Tuncer SC, Kandemir FM. Carvacrol reduces abnormal and dead sperm counts by attenuating sodium arsenite-induced oxidative stress, inflammation, apoptosis, and autophagy in the testicular tissues of rats. Environ Toxicol 2023; 38(6) : 1265-76. [CrossRef]
- Simsek H, Akaras N. Acacetin ameliorates acetylsalicylic acid-induced gastric ulcer in rats by interfering with oxidative stress, inflammation, and apoptosis. Int J Med Biochem 2023; 6(2) : 96-103. [CrossRef]
- Akaras N, Kandemir FM, Şimşek H, Gür C, Aygörmez S. Antioxidant, Antiinflammatory, and Antiapoptotic Effects of Rutin in Spleen Toxicity Induced by Sodium Valproate in Rats. TJNS 2023 ; 12(2) : 138-44.
- Yardım A, Kucukler S, Özdemir S, Çomaklı S, Caglayan C, Kandemir FM, et al. Silymarin alleviates docetaxel-induced central and peripheral neurotoxicity by reducing oxidative stress, inflammation and apoptosis in rats. Gene 2021 ; 769 : 145239. [CrossRef]
- Celik H, Kucukler S, Ozdemir S, Comakli S, Gur C, Kandemir FM, et al. Lycopene protects against central and peripheral neuropathy by inhibiting oxaliplatin-induced ATF-6 pathway, apoptosis, inflammation and oxidative stress in brains and sciatic tissues of rats. Neurotoxicology 2020 ; 80 : 29-40. [CrossRef]
- Yardım A, Kandemir FM, Çomaklı S, Özdemir S, Caglayan C, Kucukler S, et al. Protective Effects of Curcumin Against Paclitaxel-Induced Spinal Cord and Sciatic Nerve Injuries in Rats. Neurochem Res 2021 ; 46(2) : 379-95. [CrossRef]
- Gur C, Kandemir FM, Caglayan C, Satıcı E. Chemopreventive effects of hesperidin against paclitaxel-induced hepatotoxicity and nephrotoxicity via amendment of Nrf2/HO-1 and caspase-3/Bax/Bcl-2 signaling pathways. Chem Biol Interact 2022 ; 365 : 110073. [CrossRef]
- Akcılar R, Akcılar A, Koçak C, Koçak FE, Bayat Z, Şimşek H, et al. Effects of Ukrain on intestinal apoptosis caused by ischemia-reperfusion injury in rats. Int J Clin Exp Med 2015 ; 8(12) : 22158-66.
- Akaras N, Bal T, Atilay H, Selli J, Halici MB. Protective effects of agomelatine on testicular damage caused by bortezomib. Biotech Histochem 2017 ; 92(8) : 552-59. [CrossRef]
- Şimşek H, Demiryürek Ş, Demir T, Atabay HD, Çeribasi AO, Bayraktar R, et al. Assessment of expressions of Bcl-XL, b-FGF, Bmp-2, Caspase-3, PDGFR-α, Smad1 and TGF-β1 genes in a rat model of lung ischemia/reperfusion. Iran J Basic Med Sci 2016 ; 19(2) : 209-14.
- Temel Y, Çağlayan C, Ahmed BM, Kandemir FM, Çiftci M. The effects of chrysin and naringin on cyclophosphamide-induced erythrocyte damage in rats: biochemical evaluation of some enzyme activities in vivo and in vitro. Naunyn Schmiedebergs Arch Pharmacol 2021 ; 394(4) : 645-54. [CrossRef]
- Semis HS, Kandemir FM, Kaynar O, Dogan T, Arikan SM. The protective effects of hesperidin against paclitaxel-induced peripheral neuropathy in rats. Life Sci 2021; 287 : 120104. [CrossRef]
- Placer ZA, Cushman LL, Johnson BC. Estimation of product of lipid peroxidation (malonyl dialdehyde) in biochemical systems. Anal Biochem 1966 ; 16(2) : 359-64. [CrossRef]
- Sun Y, Oberley LW, Li Y. A simple method for clinical assay of superoxide dismutase. Clin Chem 1988 ; 34(3) : 497-500. [CrossRef]
- Aebi H. Catalase in vitro. Methods Enzymol 1984 ; 105 : 121-26.
- Lawrence RA, Burk RF. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem Biophys Res Commun 1976 ; 71(4) : 952-58. [CrossRef]
- Sedlak J, Lindsay RH. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman's reagent. Anal Biochem. 1968; 25(1):192-205. [CrossRef]
- Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951; 193(1):265-75. [CrossRef]
Figure 1.
Effects of CLS and NRG administrations on oxidant and antioxidant markers in sciatic nerve tissues of rats. Values are given as mean ± SE. Different letters indicate statistical difference: *p<0.05.
Figure 1.
Effects of CLS and NRG administrations on oxidant and antioxidant markers in sciatic nerve tissues of rats. Values are given as mean ± SE. Different letters indicate statistical difference: *p<0.05.
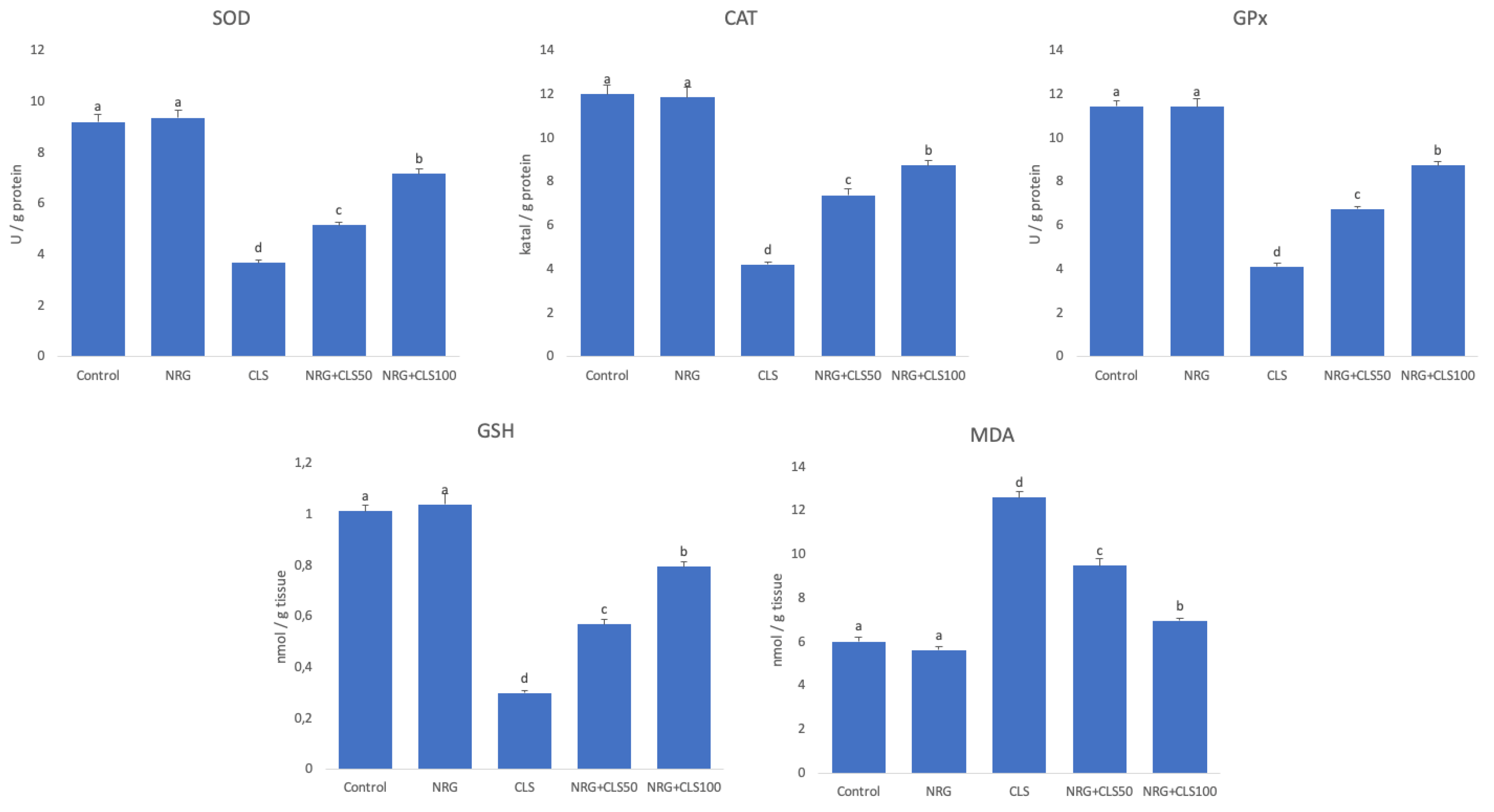
Figure 2.
Effects of CLS and NRG administrations on NF-κB, TNF-α, and nNOS levels in sciatic nerve tissues of rats. Values are given as mean ± SE. Different letters indicate statistical difference: *p<0.05.
Figure 2.
Effects of CLS and NRG administrations on NF-κB, TNF-α, and nNOS levels in sciatic nerve tissues of rats. Values are given as mean ± SE. Different letters indicate statistical difference: *p<0.05.
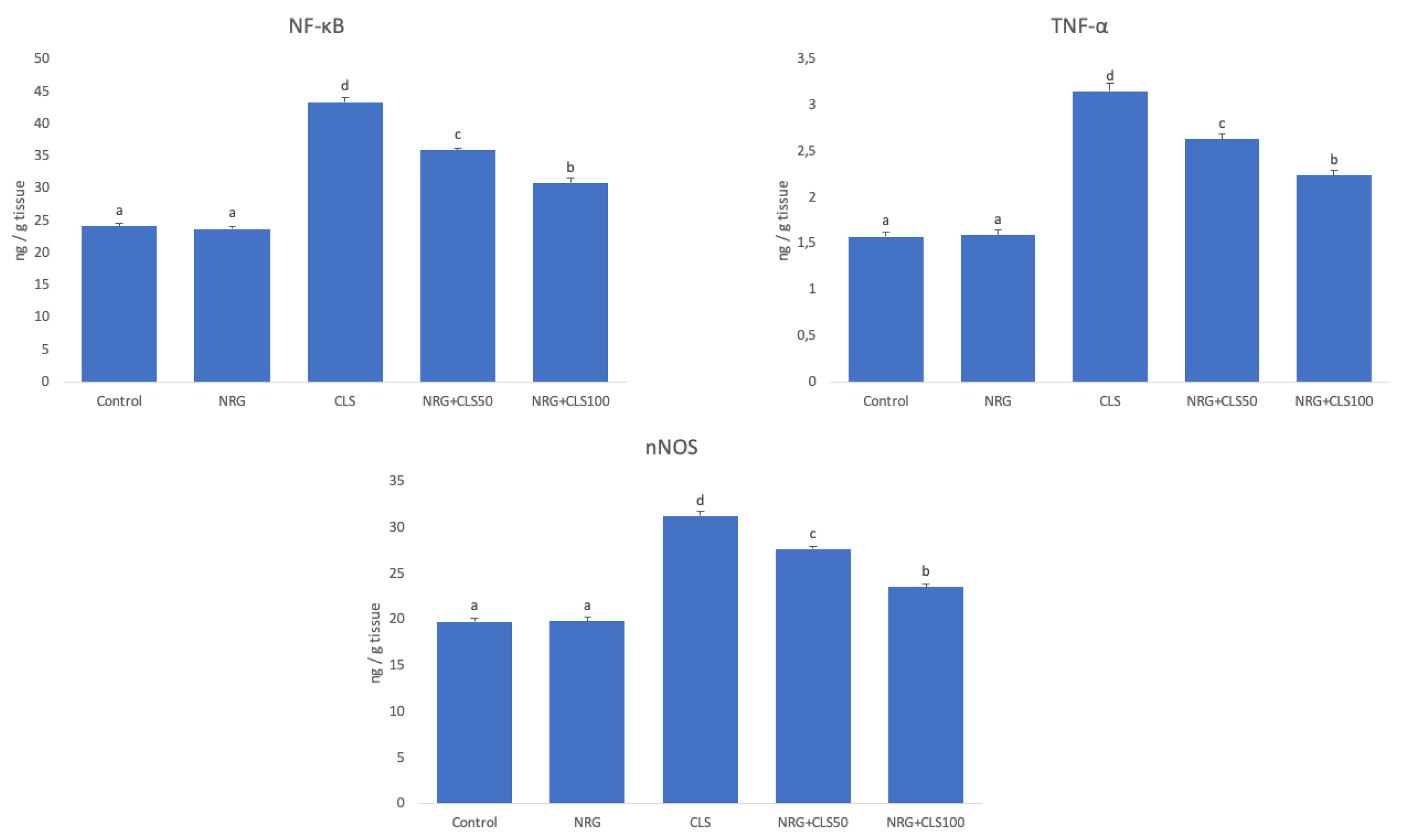
Figure 3.
Effects of CLS and NRG administrations on HO-1 and 8-OHdG levels in sciatic nerve tissues of rats. Values are given as mean ± SE. Different letters indicate statistical difference: *p<0.05.
Figure 3.
Effects of CLS and NRG administrations on HO-1 and 8-OHdG levels in sciatic nerve tissues of rats. Values are given as mean ± SE. Different letters indicate statistical difference: *p<0.05.
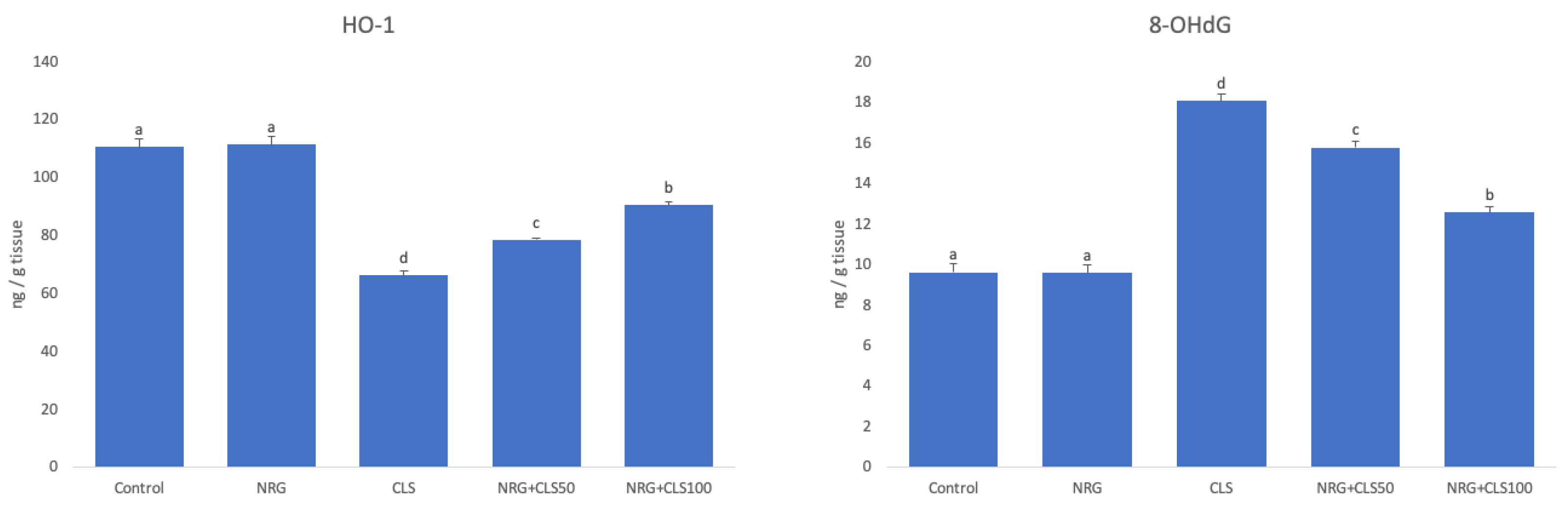
Figure 4.
Effects of CLS and NRG administrations on MAPK, nCAM and GFAP levels in sciatic nerve tissues of rats. Values are given as mean ± SE. Different letters indicate statistical difference: *p<0.05.
Figure 4.
Effects of CLS and NRG administrations on MAPK, nCAM and GFAP levels in sciatic nerve tissues of rats. Values are given as mean ± SE. Different letters indicate statistical difference: *p<0.05.
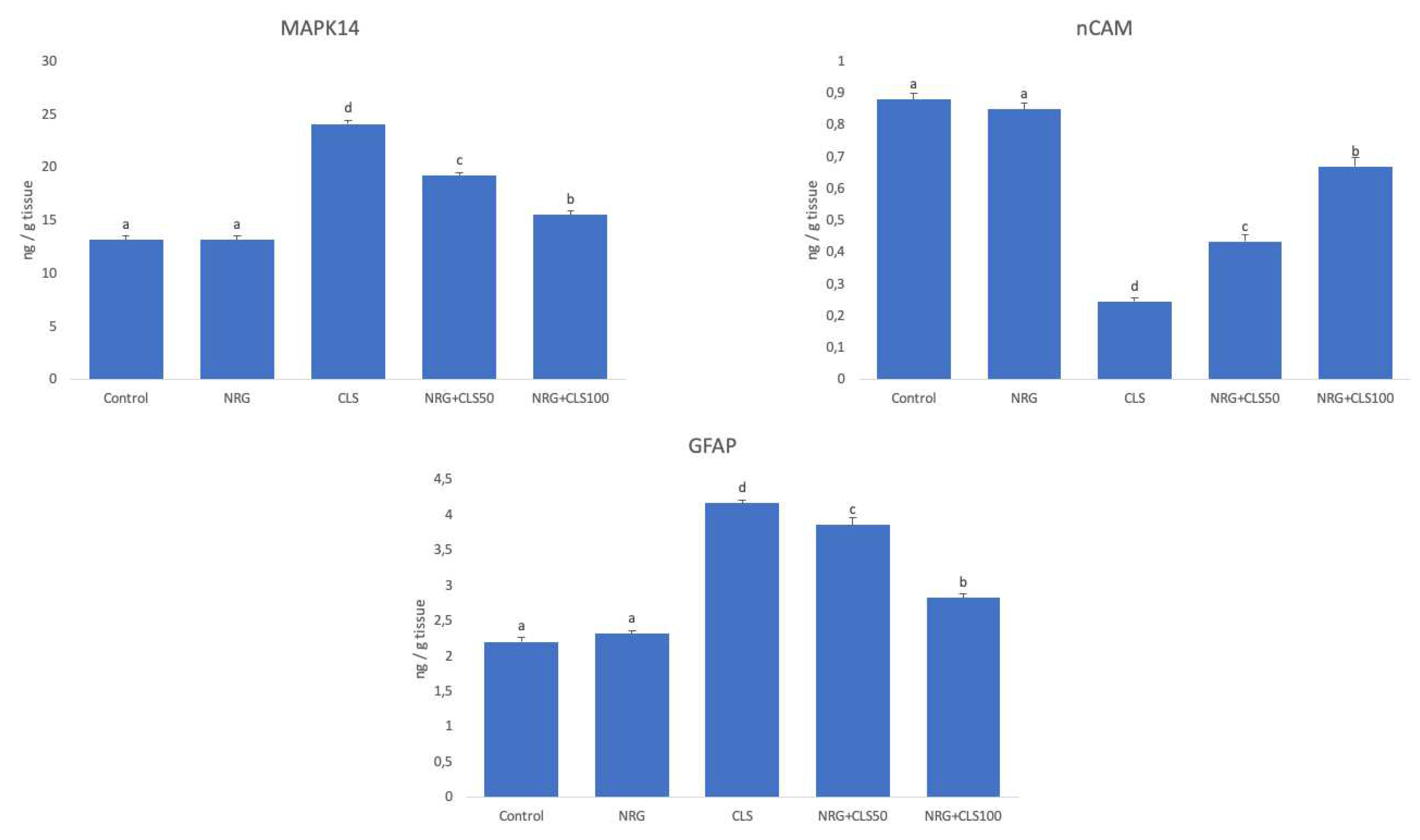
Figure 5.
Effects of CLS and NRG administrations on Caspase-3 levels in sciatic nerve tissues of rats.
Figure 5.
Effects of CLS and NRG administrations on Caspase-3 levels in sciatic nerve tissues of rats.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



