Submitted:
06 August 2023
Posted:
07 August 2023
You are already at the latest version
Abstract
: Molecular targeting strategies have been used for years to control cancer progression and are often based on targeting various enzymes involved in metabolic pathways. Keeping that in mind, it is essential to know the role of each enzyme in a particular metabolic pathway. In this review, we are providing in-depth knowledge of various enzymes such as ceramidase, sphingosine kinase, sphingomyelin synthase, dihydroceramide desaturase, and ceramide synthase, which are associated with multiple types of cancer that depend on ceramide metabolism. Focus has also been given to discussing physicochemical properties of well-studied inhibitors of natural products origin and their related structures for these enzymes. Targeting ceramide metabolism exhibited promise in mono and combination therapies at preclinical stages to prevent cancer progression and paved the way for the significance of sphingolipid metabolism in cancer treatments. Targeting ceramide metabolizing enzymes will help the medicinal chemist to design potent and selective small molecules for treating cancer progression at various levels.
Keywords:
Natural Products and related small molecules
; Sphingolipids
; Ceramide
; Ceramide Synthase
; Anti-cancer therapies
1. Introduction
Sphingolipids (SLs) are key modulators of physiologic processes including cell cycle, apoptosis, angiogenesis, stress, and inflammation [1]. Among SLs, ceramides and sphingosine-1-phosphate (S1P) are the most studied (Figure 1) and often exert opposing biological functions. Cancer cells have been shown to have a shift in the balance between proapoptotic ceramide and cancer-promoting S1P. This phenomenon is associated with pancreatic cancer progression and poor therapeutic outcomes [2,3]. Biochemical dysregulation of sphingolipid metabolism can be used as a biomarker and prognostic factor in pancreatic cancer [4,5,6]. Further studies suggested that ceramide generation and accumulation is a critical determinant of facilitating apoptosis in pancreatic cancer cells in response to cytotoxic agents, including gemcitabine (GMZ) [7], which highlights the significance of manipulating these pathways to overcome resistance of pancreatic cancer to current therapies. It is understandable why ceramide and its biosynthetic derivatives are so important to rapidly dividing cells like cancer cells, because ceramide is a basic unit of stable lipid membranes that supports transmembrane functionality and integrity [8].
Given the broad biological significance of sphingolipids, this review has focused on updating the medicinal chemistry approaches utilized towards increasing the cellular ceramide levels. The pharmacological goal is to inhibit enzymatic functions that increase cytotoxic ceramide, thus inducing cellular apoptosis. A knowledge of the pathway and its associated enzymes paves the way for identifying medicinal chemistry approaches targeting these enzymes, which could help overcome chemotherapy resistant cancer cells by exploring combination therapies that target the ceramide metabolism pathway.
2. Ceramide biosynthesis pathway
Central to sphingolipids is ceramide, which constitutes the hydrophobic backbone of all complex sphingolipids (e.g., glycosphingolipids (GS), sphingomyelin (SM), cerebrosides, gangliosides) and structurally consists of a fatty acyl of variable chain lengths bound to an amino group of a sphingoid base. The fatty acyl chains are in general saturated or monounsaturated and might contain an OH group linked to C-2 or to the terminal carbon atom (α- and ω-hydroxy fatty acids, Figure 1). Among ceramide-containing sphingolipids, those containing long (C16–20) and very long (C22–24) acyl chains are the most abundant in mammalian cells, but ceramides with longer acyl chains (C26–36) are also found in epidermal keratinocytes and male germ cells during their differentiation and maturation [8].
Ceramide is biosynthesized starting from L-Serine in the de novo synthetic pathway (Figure 1). Its biosynthesis involves cellular serine palmitoylation using the cofactor activated palmitoyl CoA by serine palmitoyl transferase resulting in 3-keto sphingosine. This is a rate determining step in the biosynthesis of sphingolipids. Cellular enantioselective reduction of the ketone catalyzed by ketosphingosine reductase allows the required 1,2 anti-amino alcohol system to be formed. N-palmitoylation of ketosphingosine followed by desaturation permits the trans alkene of ceramide. The structural and stereochemical core of ceramide has the inherent chirality of L-serine, N- palmitoylation and lipophilic alkyl chain modifications. There are different types of ceramides based on the side chain substitutions on the polar head group both the N- alkyl and the alkyl side chains.
The ceramide metabolic pathway is very dynamic and can result in various sphingolipids being synthesized to accommodate the cellular needs and enhancing cell signaling pathways. Ceramide metabolizing enzymes are cell fate specific and expressed based on the physiological role of the cell. Cancer cells tend to upregulate enzymes that promote production of sphingolipids and cell membrane stability. The remainder of the review will highlight the importance of six different classes of sphingolipid metabolizing enzymes: ceramidase, sphingosine kinase, sphingomyelin synthase, 3-ketosphinganine reductase, dihydroceramide desaturase, and ceramide synthases.
3. Ceramidase
Ceramidases (CDases) are a group of ceramide-metabolizing enzymes that hydrolyze ceramide to sphingosine. Sphingosine is then further metabolized to sphingosine-1-phosphate (S1P) by sphingosine kinase 1 or 2. In humans, currently, there are five known CDases genes. These CDases expressed by these genes can be divided into three categories depending on the pH required for their optimal catalytic activity: (i) acid ceramidase (encoded by ASAH1 gene), (ii) neutral ceramidase (encoded by ASAH2 gene), and (iii) alkaline ceramidase (encoded by ACER1, ACER2, ACER3 genes) [10].
3.1. Acid Ceramidase
Acid ceramidase (AC)/ASAH1 is also called N-acylsphingosine amidohydrolase. AC has a molecular weight of 50 kDa and requires a pH of 4.2 – 4.3 for its optimal activity [11]. Under these acidic conditions the byproduct sphingosine amine exits as ammonium species providing the active site tolerance for this charged functional group. The 3D crystal structure of AC is in Figure 2 and was derived from the Protein Data Bank (PDB). AC hydrolyzes the amide bond in unsaturated ceramides with C6-C16 acyl chains [12]. It is mainly localized in lysosomes & maintains intralysosomal ceramide homeostasis [13]. AC is expressed ubiquitously and has a higher expression in the heart & kidney [14]. The KM value of AC is determined to be 389 to 413 µM as determined using 14C-labeled and BODIPY-conjugated C-12 ceramide substrate, N-lauroylsphingosine [15]. A sphingolipid activator protein, Saposin D, is responsible for the enzymatic activity of AC as evidenced by the ceramide accumulation that occurs in the absence of Saponin D [16]. Furthermore, the altered function of mutated AC, the overexpression of normal AC, and the dysregulation of activity have highlighted the importance of its role in sphingolipid metabolism. The dysregulation of AC is associated with a wide range of diseases, thereby suggesting that AC could be an attractive therapeutic target for drug discovery.
3.2. Role of AC in Pathological Conditions
Lysosome architecture involves the basic unit sphingolipid. Both Farber disease (FD) and spinal muscular atrophy with progressive myoclonic epilepsy (SMA-PME) are rare lysosomal storage disorders. They are caused by a missense mutation in the ASAH1 gene resulting in the absence or decrease of the ACDase enzyme. There have been fewer than 200 cases of FD & SMA-PME. With no cure for ACDase deficiency, gene therapy & enzyme replacement therapy is under development [17]. Asah1 knockout in mice results in embryonic lethality [18]. Examples of AC inhibitors are shown in Figure 3. All the inhibitors exhibit a polar head group with a lipophilic tail. The experimental calculated logP depicting the lipophilicity supports this observation. Acid ceramidase is implicated in several cancers as discussed below.
3.2.1. Prostate Cancer (PC)
The role of AC in tumor proliferation and resistance to treatment has been studied in depth in prostate cancer. Prostate cancer cells develop resistance by upregulating AC. Clonogenicity and cytotoxicity assays confirm that radio sensitization can be established by the genetic downregulation of AC with small interfering RNA (siRNA). Also, the radiation-induced AC upregulation creates cross-resistance to chemotherapy. The use of the small molecule AC inhibitor, LCL385, sensitizes primary prostatic carcinoma cell line (PPC-1) cells to radiation [19]. In addition, the same cell line preferentially upregulates AC resulting in increased radiation resistance and proliferation. The use of AC inhibitor, LCL521, acts as a radiosensitizer preventing relapse [20]. This was validated with immunohistochemical studies in these tissues where higher levels of AC were observed after radiation treatment failure than in irradiation-naïve cancer, intraepithelial neoplasia, or benign tissue.
In addition to AC inhibition in an animal xenograft model producing radio-sensitization, it also prevented relapse. Higher IHC expression of AC in primary prostate cancers is associated with more advanced disease stage. In a model derived from PC-3 prostate cancer cells, the highly tumorigenic, metastatic and chemo-resistant PC-3/Mc clone expressed higher levels of AC than the non-metastatic PC-3/S clone. Stable knockdown of Asah1 in PC- 3/Mc cells resulted in an accumulation of ceramide ,a reduction of clonogenic potential, and inhibition of tumorigenesis and lung metastases [21]. Treatment of DU-145 cells with acid ceramidase inhibitor LCL204, a lysosomotropic analog of B13, induces apoptosis in a cathepsin-dependent manner. Upon treatment with this inhibitor, destabilization of membranous lysosomes and the release of lysosomal proteases into the cytosol lead to mitochondria depolarization and caspase activation resulting in apoptosis [22]. Both PC-3 and DU 145 are considered to be hormone-refractory human prostate cancer cell lines. In both of these cell lines, combination treatment with fenretinide (4-HPR), a ceramide-generating anticancer agent, and DM102, a novel synthetic AC inhibitor, resulted in a considerable decrease a cell viability and the combination was more effective than either treatment alone. In the PC-3 cell line, the treatment induces apoptosis through ROS generation. However, another AC inhibitor, N-oleoylethanolamine (NOE), in combination with 4-HPR does not result in synergistic activity [23]. In another study using PPC1 and DU145 prostate cancer cells, AC overexpression results in increased lysosomal density and autophagy and increased expression of the motor protein KIF5B, contributing to lysosomal stability. AC overexpression, in addition to increasing radiation & chemotherapy resistance, increases stress resistance. Although increased lysosomal density in these cells makes them more sensitive to any therapeutic agents targeting lysosomes, providing a new therapeutic target [24].
3.2.2. Head and Neck Cancer (HNC)
AC was overexpressed in four of six primary tumor tissues and six of nine cell lines in HNC. AC also contributed to decreased cisplatin sensitivity. Pharmacological (with N-oleoyl-ethanolamine) or genetic (with short hairpin RNA) inhibition of AC enhanced cisplatin-induced HNC cell death, by increasing ceramide and activating other proapoptotic proteins [25]. In mouse squamous cell carcinoma SCCVII, the inhibition of AC with the small molecule inhibitor LCL521 significantly decreased the survivability after photodynamic therapy by effectively restricting regulatory T lymphocytes and myeloid-derived suppressor cell activity [26]. The combination of photodynamic therapy and dasatinib also decreases SCCVII cell survivability by decreasing AC, leading to increased ceramide activating caspase-3 induced apoptosis [27]. In SCC-1, over expression of AC increased resistance to Fas-induced cell death, which was reversible using specific AC siRNA. The AC inhibitor LCL 204 sensitizes HNSCC cell lines to Fas-induced apoptosis both in vitro and in a xenograft model in vivo, providing an option for combination therapy [28].
3.2.3. Melanoma
The largest organ of human body, skin, has a very complex structure with four main layers. Sphingolipids are found throughout each layer, which maintain the functions of this organ. AC expression is significantly higher in normal human melanocytes and proliferative melanoma cell lines, compared with other skin cells (keratinocytes and fibroblasts) and non-melanoma cancer cells. Melanoma cells exhibit lower amounts of ceramide by downregulating the de novo synthesis pathway. The AC inhibitor ARN14988 in combination with 5-Fluoro Uracil increases cytotoxicity in the proliferative melanoma cell lines by increasing ceramide and reducing S1P levels [29]. In human A375 melanoma cells, dacarbazine (DTIC) decreases ACdase activity by reactive oxygen species-dependent activation of cathepsin B-mediated degradation of the enzyme. Downregulating AC expression increased ceramide level & sensitivity to DTIC providing a therapeutic tool for the treatment of metastatic melanoma [30]. The deletion of the gene encoding for AC, ASAH1, in human A375 melanoma cells, using CrispR-Cas9-mediated gene editing, showed a significantly greater accumulation of long-chain saturated ceramides that are substrates for AC in ASAH1-null cells. The cells lose the ability to undergo self-renewal [31]. Our hypothesis on this observation correlates with ceramide flux and its function.
3.2.4. Myeloid Leukemia
Primary acute myeloid leukemia (AML) cells have higher expression of AC which is essential for AML blast survival. In AML cell lines, increased levels of AC induced higher expression of anti-apoptotic Mcl-1 protein, increased S1P, and decreased ceramide. Treatment with AC inhibitor LCL204 induces ceramide accumulation and decreases Mcl-1. The overall survival of C57BL/6 mice engrafted with leukemic C1498 cells increased significantly with LCL204 treatment while significantly decreasing the disease burden in NSG mice engrafted with primary human AML cells [32]. IFN regulatory factor 8 (IRF8) is a key transcription factor for myeloid cell differentiation. Without IRF8, hematopoietic cells in human myeloid leukemia patient rapidly proliferate and remain undifferentiated. Thus, acting as a tumor suppressor by promoting cell differentiation, IRF8 expression is frequently lost in myeloid leukemias. One of the repressive transcriptional targets of IRF8 is AC; consequently, as IRF8 is lost AC expression increases, solidifying its role as tumor suppressor. In chronic myelogenous leukemia (CML), overexpression of IRF8 repressed AC expression, resulting in C16 ceramide accumulation and increased sensitivity of CML cells to FasL-induced apoptosis. AC expression is significantly higher in cells derived from IRF8-deficient mice. In these cells, inhibition of ACdase activity or application of exogenous ceramide sensitizes the cells to FasL-induced apoptosis [33].
3.2.5. Non-small cell lung cancer (NSCLC)
In NSCLC cells with acquired resistance to ChoKα inhibitors also display increased levels of ASAH1. Inhibition of AC synergistically sensitizes lung cancer cells to the antiproliferative effect of ChoKα inhibitors, which opens up a new therapeutic option for combinatorial treatments of ChoKα inhibitors and AC inhibitors [34].
3.2.6. Breast Cancer
Higher expression of AC has been observed in ER-positive and luminal-A-like breast cancer. High expression of AC in invasive breast cancer is associated with improved prognosis and reduced incidence of recurrence in preinvasive ductal carcinoma in situ (DCIS) [35]. The effect of AC inhibitor (DM102 & NOE) in combination with C6-ceramide (C6-cer), a cell-permeable analog of ceramide, has been studied in three different breast cancer cell lines MDA-MB-231, MCF-7, and BT-474 cells. Although, as single agents, both C6-cer & DM102 are moderately cytotoxic, their combination induced synergistic decreases in viability. In MDA-MB-231 cells, apoptosis is induced by caspase 3/7 activation and poly (ADP-ribose) polymerase (PARP) cleavage. In the same cell line, C6-cer/DM102 increases ROS levels and results in mitochondrial membrane depolarization. Furthermore, C6-cer/DM102 combination is antagonistic in BT-474 cells, suggesting different molecular mechanisms being cell type-specific. AC expression is correlated to Human Epidermal Growth Factor Receptor 2 (HER2) status [36]. Another study shows that AC inhibitor, ceranib-2, induces apoptosis in MDA-MB-231 and MCF-7 by activating stress-activated protein kinase/c-Jun N-terminal kinase and mitogen-activated protein kinase apoptotic pathways by inhibiting the antiapoptotic pathway [37]. Ceranib-2 exhibits similar effects in prostate cancer cell lines too [38].
3.2.7. Ovarian Cancer (OC)
In an immunohistochemical analysis study of 112 ovarian cancer patients, low AC expression has been correlated with tumor progression in ovarian cancer. This analysis contradicts the concept of AC being a cancer progression promoter suggesting AC is involved in alternative pathways in different cancers, which requires further investigation [39].
3.2.8. Hepatobiliary Cancers
AC is downregulated by the chemotherapeutic agent daunorubicin in human (HepG2) and mouse (Hepa1c17) hepatoma cell lines as well as in primary cells from murine liver tumors, but not in cultured mice. Genetic (small interfering RNA) or pharmacological inhibition of AC with N-oleoylethanolamine (NOE) sensitized the cell lines to daunorubicin-induced cell death, preceded by structural mitochondrial changes, stimulation of reactive oxygen species generation and cytochrome c and caspase-3 activation. In vivo siRNA treatment targeting ACDase reduced tumor growth in liver tumor xenografts of HepG2 cells and enhanced daunorubicin therapy, providing a potential therapeutic target for liver cancer [40].
The key chemotherapeutic agent in pancreatic cancer, gemcitabine, exhibits different efficacy due to polymorphism in the expression of enzymes that regulate its metabolism. [41] Deoxycytidine kinase (dCK), which phosphorylates gemcitabine, activates the drug, while cytidine deaminase (CDA) inactivates gemcitabine by deamination [6]. In MIA PaCa-2 and PANC-1 pancreatic cancer cell lines, the novel ceramide analog AL6 inhibits cell growth, induces apoptosis, and synergistically enhances the cytotoxic activity of gemcitabine. AL6 also increases the gene expression of the gemcitabine-activating enzyme deoxycytidine kinase (dCK), improving the efficacy of gemcitabine. This study suggests the use of AL6 & gemcitabine combination therapy for pancreatic cancer [41].
3.2.9. Colon Cancer
Colorectal adenocarcinoma tissues have higher ASAH1 expression compared to the adjacent normal colonic mucosa. In HCT 116 colon cancer cells there is an inverse correlation between the AC expression and p-53 functional activity. Inhibition of AC by carmofur in HTCT 116 CELLS, significantly increased the anti-proliferative, pro-apoptotic, anti-migratory, and anti-clonogenic effects of oxaplatin [42]. In the human colon cancer cell line, AC inhibitor, ceranib-2 increases apoptosis by increasing ASAH1 mRNA expression and reducing TNF-R1 expression [43].
4. Sphingosine Kinase
Sphingosine-1-phosphate (S1P) is a bioactive sphingolipid that regulates the growth, survival, and migration of several cell types. S1P is a ligand for five transmembrane G-protein coupled receptors, S1P1-5, and for several intracellular targets such as histone deacetylases 1 and 2 [44]. Cellular biosynthesis of S1P occurs through phosphorylation of Sphingosine (Sph) catalyzed by sphingosine kinases. Sphingosine, an effector molecule is biosynthesized by ceramidase activity on the central lipid ceramide. Sphingosine kinases exist in two isoforms SphK1 and SphK2 encoded by unlinked genes. The crystal structure of Sphingosine Kinase 1 from Protein Data Bank is shown in Figure 4. Pharmacologically, ceramide and sphingosine are associated with growth arrest and apoptosis. On the contrary, S1P is associated with pro-survival roles [45]. SphKs and S1P have been implicated in a variety of disease states including cancer, [46,47] sickle cell disease, [48,49] atherosclerosis, [50,51] asthma, [52,53] diabetes, fibrosis, [54] among others.
Although SphK1 and SphK2 share a high degree of homology, they differ in size, localization, distribution, and intracellular roles [55,56]. While double knockout studies in mice suggest that SphKs are the sole source of S1P, some functional redundancy exists as SphK1 or SphK2 null mice are viable and fertile [57].
The biological significance of sphingosine kinases has encouraged academia and pharmaceutical companies to target SphKs for therapeutic value. Initial drug discovery efforts resulted in SphK1 potent inhibitors, complimented by the availability of SphK1 crystal structure. Despite that no crystal structure for SphK2 is available, potent SphK2 inhibitors have been developed through homology modeling of SphK1 and strategies aimed toward competitive inhibition. Shown in Figure 5 are the representative sphingosine kinase 2 inhibitors with moderate potency and selectivity. ABC294640 was the first Sphingosine Kinase II inhibitor with Ki 10 μM [58] that has been deployed in a variety of disease models which include ulcerative colitis, [59] Crohn’s disease, [60] ischemia/reperfusion injury, [61] osteoarthritis, [60] colon cancer, [62] colorectal cancer [63]. However, ABC294640 has recently been reported to have an off-target effect of acting as a tamoxifen-like molecule with the estrogen receptor [64]. Other inhibitors, namely SG-12, [65] (R)-FTY720-OMe, [66] K145, [67] and VT-ME6 [68] exhibit optimal potency and selectivity.
Based on Structure-Activity Relationship (SAR) studies, isoform-selective SphK2 inhibitors with improved potency and half-life in mice. These inhibitors include SLR080811, [71] SLP120701, [72] SLM6031434, [73] and SLC5091592 [74], VT 20dd [69] were recently reported in Figure 6.SLR080811 with a Ki of 13.3 μM and 1.3 μM for SphK1 and SphK2 respectively is under study for colorectal cancer resistance to 5-fluorouracil. [71]. An important finding from these studies was the observation of elevation of S1P levels in the mice upon pharmacological inhibition of SphK2. Extensive SAR studies of SLR080811 resulted in azetidine, SLP120701 with an improved half-life to 8 hours in mice [72]. Modifications in the tail region further improved sphingosine kinase II selectivity with analog SLM6031434 [73], and lipophilic tail substituted naphthalene-oxy analog, SLC5091592 [74]. These analogs showed improved SphK2 selectivity and potency comparable to second-generation SphK2 inhibitors in Figure 6.
As evidence, sphingosine kinases targeting scaffolds tail regions can be modified towards transitioning towards kinase specific and selectivity.This study was shown in, aminothiazole based dual and sphingosine kinase 2 inhibitor development 20dd in Figure 6 with improved potency, selectivity and in vivo features [69]. A pubmed search for probing the anticancer potential for these analogs was not reported so far but with a range of selectivity, specificity towards sphingosine kinases warrants to explore these analogs.
5. Sphingomyelin Synthase
Sphingomyelin Synthase (SMS) is responsible for generating Sphingomyelin (SM) and diacylglycerol (DAG) by transferring the phosphocholine from phosphatidylcholine onto the primary hydroxyl group of ceramides [75,76]. Thus, SMS is also biologically important as it regulates the levels of ceramide and DAGs resulting in bioactive lipids [77]. In the de novo synthesis pathway, it starts with L-serine and palmitoyl CoA to give 3- Ketosphinganine, which then undergoes a series of enzymatic reactions to yield ceramide on the cytoplasmic side of the endoplasmic reticulum (ER) membrane. Ceramide is then transferred to the Golgi compartment in a non-vesicle way by ceramide transfer protein (CERT). There SMS transfers the phosphocholine headgroup from phosphatidylcholine (PC) to ceramide yielding SM and DAG. The sphingomyelin produced in this step is then sorted into cell membranes by either vesicle traffic or protein-facilitated transportation [78]. It is noteworthy to mention that SM is the basic component of lipid raft. Lipid rafts are important microdomains of cell membranes that provide a platform for many receptors and transport proteins. The SMS gene family consists of three members - sphingomyelin synthase 1 (SGMS1), sphingomyelin synthase 2 (SGMS2), and sterile alpha motif domain containing (SAMD8) encoding their respective proteins: SMS1, SMS2, and SMS-related protein (SMSr). Even though SMSr contains high homology with SMS 1 and SMS2, it does not have any SM synthase activity [76]. SMS1 and SMS2 are localized in the trans-Golgi network, where SM is synthesized from ceramide, which is transported from the endoplasmic reticulum (ER) to the Golgi by the Ceramide Transporter Protein (CERT) [1]. SMSs are present in all tissues, and SMS1 is the principal contributor to the SMS activity in most cells. Both of the isoforms share 57% of sequential identity and are conserved in mammals [76]. In SMS1, a sterile alpha motif (SAM) is present, which takes part in protein-protein interaction, which is not present in SMS2. SMSs contain 6 transmembrane regions with both N- and C- termini exposed into the cytosol.
5.1. Biological Significance of Sphingomyelin Synthase
The formation of sphingomyelin is essential for cell growth and survival. In a mouse lymphoid cell line deficient in SM synthase activity, loss of SMS activity halted cell growth in serum-free conditions, which could be restored by supplemental exogenous SM or heterologous expression of SMS1 [79]. Different studies have correlated the up and down-regulation of SM synthase activity to mitogenic and pro-apoptotic signaling in different mammalian cell types. [80,81,82] Although the cellular pathway for the effects of SMS is unclear, it can exert its effect through the following mechanisms: 1) SM accumulation in the plasma membrane, and its affinity for sterols contributes to the rigidity of the cell membrane; 2) SM accumulation in the plasma membrane acts as a source for a number of other sphingolipids, which are catalyzed by acidic or neutral sphingomyelinases (SMases) [83,84]. Ceramide, sphingosine, and sphingosine 1-phosphate are all potential SM metabolites and have proven to be significant regulators of cellular function like cell proliferation, differentiation, and apoptosis. [2,85] The microdomain formation of SM in Golgi apparatus plays a role in the sorting process of different sphingolipids. [86] SM synthesis can act as a source of DAG in trans Golgi network, thus facilitating the protein kinase D recruitment leading to the formation of transport carrier proteins [87]. SM synthesis regulates the cellular levels of both pro-apoptotic factor ceramide and mitogenic factor DAG directly impacting cell proliferation [88,89]. Some of the SMS inhibitors which are marine environment origin, natural product like analogs resemblance with SMS substrate are depicted in Figure 7.
6. 3-Ketosphinganine Reductase
3-Ketosphinganine reductase (KSR) mediates the reduction of ketosphinganine to sphinganine (Sa). In certain cancer cells (HGC27, T98G and U87MG), 3-ketosphinganine (KSa) and its deuterated analog at C4 (d2KSa) are metabolized to produce high levels of dihydrosphingolipids [90]. Although the function of KSR have been studied in yeast and plants, the role of it in human pathology lacks supportive findings of its role.
7. Dihydroceramide Desaturase
Dihydroceramide Δ4-desaturase (DES) is the member of the desaturase family, which converts the dihydrosphingosine backbone within ceramide into a sphingosine backbone [91]. The first step is utilizing molecular oxygen to introduce a hydroxyl group into the C4 position of the dihydrosphingosine backbone, which is then followed by a dehydration reaction producing a double bond in the C4-C5 position of dihydroceramide, with the aid of NADPH [92,93,94]. The only difference between dihydroceramide and ceramide is, ceramide has a double bond at C4-C5 position. In mammals, two gene isoforms named DES1 and DES2 have recently been identified [95]. DES1 gene contains multiple transmembrane domains and a recent study shows that it requires myristoylation on its N-terminus for full activity [95,96]. DES1 is localized in the ER membrane where it has access to newly synthesized dihydroceramide species [91]. 4-hydroxyceramide is an intermediate reaction product in the conversion of dihydroceramide to ceramide which is also known as phytoceramide. In plants and yeast, it is the predominant ceramide species. In mammals, DES1 is found in all tissues and only converts dihydroceramide species into fully desaturated ceramide; whereas DES2, is capable of creating either phytoceramide or ceramide from dihydroceramide precursors [97,98]. DES2 is highly expressed in skin, intestine and kidney [97]. The deletion of DES1 and DES2 shifts sphingolipid synthesis pathway toward the sphingolipid lacking the double bond introduced by DES1 and DES2, such as dhS1P, dhSph, dhsphingomyelin (dhSM) and especially dhCer [99]. In Des1−/− mice, the inability to form ceramide leads to highly elevated dihydroceramide, low levels of ceramide, multi-organ dysfunction, and failure to thrive. [100] Ceramide has distinct signaling properties from dihydroceramide and phytoceramide, suggesting that most cells have evolved to recognize ceramide as a more significant determinant to initiate a cellular response [101].
7.1. Role of Dihydroceramides in various disease
7.1.1. DhCer in Brain Diseases
Increased dhCer levels have been observed after hypoxia and subarachnoid hemorrhage. [102,103]. Both of these studies suggest the involvement of dhCer in the mechanisms of disease in oxygen deprivation states such as stroke. Altered dhCer levels have also been noted in studies related to certain neuronal diseases such as luekodystrophia [9], Alzheimer’s [104], and Huntington’s disease (HD) [105]. The association of dhCer with the progression of degenerative brain diseases and other brain-related diseases makes it a potential target as a biomarker.
7.1.2. DhCer in Cardiovascular Disease
DhCers were found to be increased in both human atherosclerotic plaques and rat models of hypercholesterolaemia [106,107]. DhCer also correlates with the release of macrophage inflammatory protein 1β (MIP-1β). Although the role of dhCer in plaque stability is debatable, since the extracellular addition of dhCer to human aortic smooth muscle cells did not cause apoptosis, whereas ceramide did [108]. Apart from these studies increased dhCer levels have been found in patients with rheumatoid arthritis [109] and in doxorubicin-induced cardiac toxicity [110]. All these studies suggest the role of dhCer as a marker for cardiac pathology.
7.1.3. DhCer in Cancer Therapy
As ceramide has been studied for its apoptotic property, in most of these studies, dhCer has been considered as a precursor to ceramide [107,111,112]. Although some studies have focused on dhCer’s potential role in cancer cell autophagy, [113,114,115]in cancer induced bone pain [116], and cell cytotoxicity [117]. The fluctuation in the dhCer and ceramide levels in cancer cells seem to differ according to the site of origin of the cancer. For example, in cancerous tissue of human endometrial cells the level of dhCer was increased 3- to 4.6-fold, and Cer and S1P were increased 1.6- to 1.9-fold [118] whereas in melanoma cells, dhCers and Cers were significantly lowered compared to non-malignant melanocytes [119]. Recent studies have focused on the gatekeeper enzyme, DES1, a new target for cancer therapy for a better understanding of the pathological effects of dhCer in cancer. 4-HPR-fenretinide, a DES1 inhibitor, is currently being studied for different types of cancers including peripheral T-cell lymphomas and solid tumorsIn SMS-KCNR neuroblastoma cells, 4-HPR-fenretinide directly inhibits DES1 with an IC50 of 2.32 µM. Inhibition of SK sensitizes cells to 4-HPR-fenretinide’s cytotoxic effects due to increased level of dhCers [120]. The possible interaction between 4-HPR-fenretinide inhibition of DES1 and SK activity has been supported by few other studies [115,121,122,123]. In cancer cell lines like HEK293, MCF 7, A549 and SMS-KCNR cells, oxidative stress can also inhibit DES1 followed by increased level of dhCers.[124] Increasing the exogenous dhCer levels induce autophagy in T98G, U87MG glioblastoma cells [114] and DU145 cells [113] and reduces proliferation in castration-resistant prostate cancer cells [125]. In human gastric cancer cell line, HGC-27, dhCers exert autophagic effects when DES1 is inhibited by XM462 and resveratrol, resulting in higher levels of dhCer [115]. DhCer only induces autophagy when the de novo sphingolipid biosynthesis pathway is altered; in studies where there were both dhCers and ceramides level increased together, apoptosis occurred instead of autophagy [126]. DES1 assists to the advancement of metastasis in prostate cancer cells, [127] and esophageal carcinoma, possibly by increased cyclin D1 expression as a result of NF-кB activation [128]. These studies suggest the potential of increasing dhCer levels to increase autophagy and inhibiting metastasis through DES1 inhibition as promising targets for cancer therapy. A list of DES1 inhibitors which are natural products and related molecules which are resemblance with sphingolipid structure are shown in Figure 8 and Figure 9 respectively [97].
8. (Dihydro)ceramide Synthase
Dihydrosphingosine (DHSph) is further acylated by six different (dihydro)ceramide synthases. In mammals, six distinct (dihydro)ceramide synthases abbreviated as CerS1-6 have been identified and are encoded by six distinct genes [129,130]. In sphingolipid metabolism, no other step has as many genes devoted to it as (dihydro)ceramide synthesis suggesting that each different CerS has distinct functions.
9. Ceramide Synthases
Ceramide synthases are a group of enzymes which play a central role in sphingolipid metabolism by catalyzing the formation of ceramides from sphingoid bases and acyl-CoA substrates. So far, six CerSs (CerS1–CerS6) have been identified and each of them has a unique characteristic which will been discussed below.
9.1. Ceramide Synthase 1
Studies have shown Ceramide Synthase 1 (CerS1) to prefer stearoyl CoA as a substrate producing the long-chain C18-ceramide [131]. In humans, CerS1 expression has been detected in glioblastoma cells [132], lung cells [132] and in brain tissue [131]. Studies have shown upregulated expression of CerS1 in the anterior cingulate cortex in post-mortem brain tissue from Parkinson`s disease patients [133]. In Parkinson`s disease patients, the C16:0-, C18:0-, C20:0-, C22:0- and C24:1-ceramides concentration level is elevated in plasma, indicating the involvement of other CerS isoforms in the development of the disease [134]. CerS1 is also linked to the autoimmune disorder multiple sclerosis, a neuronal disease characterized by demyelination of neurons. In the spontaneous relapse-remitting EAE (experimental autoimmune encephalomyelitis), CerS1 expression in the lumber spinal cord is decreased [135]. CerS1 is also associated with the development of obesity. In liver microsomes of high-fat diet-induced obese (DIO) mice, an upregulation of CerS1 expression is shown due to high-fat diet [136].
CerS1 has been identified to play a role in the pathogenesis of head and neck squamous cell carcinoma (HNSCC). Studies have substantiated correlations between reduced C18-ceramide in HNSCC tumors and increased lymphovascular invasion, nodal metastasis, and higher tumor stages [137]. In A549 human lung adenocarcinoma cells, C18-ceramide generated by overexpressed CerS1 represses the promoter activity of human telomerase reverse transcriptase (hTERT) [137]. Human breast tumors exhibit increased CerS1 mRNA levels when compared with normal breast tissue, and it was correlated with poor prognosis of the patients [138]. In human colorectal cancer (CRC) tissue compared with nontumor colonic tissue, elevated CerS1 mRNA levels were observed; however, this was accompanied by a reduction in C18-ceramide levels [139].
In neuroblastoma cells, CerS1 downregulation results in ER stress and pro-apoptotic signaling. [140] In human glioma tissue, C18-ceramide levels are lower than in control tissue, and overexpression of CerS1 or exogenous C18-ceramide trigger ER stress, lethal autophagy, and cell death in glioma cell lines [141]. These studies stipulate that CerS1 and its product C18-ceramide can exhibit anti-proliferative effects in different cancer cell lines and tissues.
9.2. Ceramide Synthase 2
Ceramide Synthase 2 (CerS2) utilizes C20-C26 acyl CoA species and is responsible for very long chain ceramide species [142]. CerS2 has a substrate specificity towards C20:0-, C22:0-, C24:0-, C24:1-, and C26:0-acyl-CoAs. Its KM towards sphinganine is 4.8 ± 0.4 μM. In human, CerS2 is expressed in kidney, liver, brain, heart, placenta, lung, breast tissue, skeletal muscle, testis, intestine, and adipose tissue. [143,144,145,146] This broad and quantitatively strong tissue distribution of CerS2 indicates its prominent role among the CerS isoforms and the importance of CerS2-derived very long-chain ceramides for basal cellular sphingolipid metabolism. Due to the wide distribution and its distinct genomic features, the CERS2 gene has been described as a potential housekeeping gene in mammalian cells. The CerS2 protein is localized in the ER.
CerS2 is strongly associated to the development of multiple sclerosis. In the experimental autoimmune encephalomyelitis (EAE) model, CerS2 knockdown has a protective effect, possibly due to an impaired migration of neutrophils into the CNS [147]. In the spontaneous relapse-remitting EAE mouse model, CerS2 expression decreased in the lumbar spinal cord [135]. CerS2 is also linked to the chronic neurodegenerative Alzheimer`s disease. In an Alzheimer`s disease model, there was increased expression of CerS2 in brain tissue, which lead to apoptosis in glial cells. [148] In progressive myoclonic epilepsy (PME) patients, heterozygous deletion of CerS2 in fibroblasts have been observed, which suggests that a reduced CerS2 level led to PME development [149]. CerS2 plays a significant role in CNS development and pathological conditions.
Several studies have supported the role of CerS2 as a tumor suppressor protein and in maintaining cell and tissue integrity. In human HCC tissue, low expression of CerS2 correlates with tumor progression and poor prognosis [150]. In breast cancer patients, inverse relationships between CerS2 expression and tumor progression, lymph node metastasis, and HER2 expression were discovered [151]. CerS2 overexpression inhibits proliferation and triggers cell cycle arrest and apoptosis in a p21/p53-dependent manner in papillary thyroid cancer cells [152]. Decreased level of CerS2 inhibits tumor growth and metastasis in meningioma, [153] bladder cancer [154,155,156] and prostate cancer [157,158].
9.3. Ceramide Synthase 3
CerS3 prefers middle and long chain acyl CoAs and generates C18:0-ceramide and longer chain ceramides [159,160]. In human tissue, CerS3 is expressed in keratinocytes, and shows high expression in kidney and liver with moderate expression in brain, heart, skeletal muscle, placenta, and lung [161,162].
Mutation of CerS3 has been reported as a reason for autosomal recessive congenital ichthyosis (ARCI), a keratinization disorder [163]. CerS3 mRNA is reduced to 70% in these patient’s skin. Another study supported the cause by showing a splice mutation in CerS3 leading to a reduced number of very long-chain ceramides in the skin, which are essential for epidermal differentiation, an essential process for maintenance of epidermal barrier function. [164] There is a lack of data on CerS3 expression in cancer, possibly due to a restricted expression of CerS3 in the mammalian body, the limited availability of specific antibodies, and the lethality of CerS3 knockout mice. One study reported decreased CerS3 mRNA levels in human breast tumors compared with normal breast tissue. CerS3 is the only ceramide synthase which is downregulated compared to significantly upregulated CerS2, 4, 5, and 6 [138].
9.4. Ceramide Synthase 4
Ceramide Synthase 4 (CerS4) exhibits a substrate specificity towards C18:0- and C20:0-acyl-CoAs. In humans, it is expressed in kidney tissue (renal papillae, medulla, cortex) [165] and breast tissue [145].
In an Alzheimer`s disease mouse model, upregulation of CerS4 expression and increased C20:0- and C24:0-ceramide in the hippocampal brain tissue was observed. [148] In human liver cancer tissue, CerS4 is upregulated at the mRNA and protein level and promotes liver cancer cell proliferation associated with NF-κB signaling [166]. In human breast cancer tissue, there is higher CerS4 mRNA expression compared to healthy breast tissue [167]. CerS4 expression is higher in estrogen receptor (EsR)-positive tumors than in EsR-negative tumors [168]. It is possible that the increase in the ceramide synthesis by CerS and other CerSs might promote breast and colorectal cancer cell growth through a disturbed cellular sphingolipid homeostasis. Moreover, breast cancer patients with higher mRNA expression of CerS4, along with CerS1 and CerS5, show a worse prognosis than those with low CerS expression levels [138].
9.5. Ceramide Synthase 5
Ceramide Synthase 5 (Cers5) prefers palmitoyl CoA as substrate and generates predominantly C16-ceramide species [169] and is expressed in human lung, [144] kidney (renal papillae, medulla, cortex) [165] and breast tissue [145]. The KM towards sphinganine is 1.8 +/− 0.4 μM and is expressed in moth ER and nucleus. Study has shown a mild upregulation of CerS5 in the lumber spinal cord in spontaneous relapse-remitting EAE mice model [135]. In addition, CerS5 mRNA expression is elevated in patient-derived colorectal cancer (CRC) tissue in comparison to normal colonic mucosa, [170,171,172] and CerS5 can be used as a marker for CRC [172]. Another study where data from a reverse phase protein microarray using epithelium-enriched, human CRC tissue samples were used, revealed high CerS5 protein expression is associated with autophagy-regulating protein signaling network, in contrast to low CerS5 levels with an apoptosis-related proteomic network [173]. In human neuroglioma tissue, elevated expression of CerS5 mRNA and protein levels were observed when compared with normal nervous ganglion tissue [174]. These studies suggest the correlation between high CerS5 expression and tumor cell proliferation and cancer progression in CRC, breast cancer, and other malignancies.
9.6. Ceramide Synthase 6
Ceramide Synthase 6 (CerS6) has a substrate specificity for C14:0- and C16:0 acyl-CoAs and its KM towards sphinganine is about 2.0 +/− 0.6 μM. It is mainly localized at the ER. In human it is expressed in kidney (renal papillae, medulla, cortex) [165] and breast tissue [145].
Increased CerS6 expression is observed in neutrophils isolated from blood [147] and in macrophages and astroglia in the lumbar spinal cord [175] in progressive, chronic experimental autoimmune encephalomyelitis (EAE) mouse model. In spontaneous relapse-remitting EAE, over expression of CerS6 in macrophages was observed [135].
While comparing with corresponding healthy tissues, an abnormal higher CerS6 is observed in colorectal cancer (CRC) [170,171] and breast cancer [138,167,176,177]. Also, CerS6 expression is higher in estrogen receptor (EsR)-positive breast tumors than in EsR-negative tumors [167,168,178]. A similar kind of pattern was observed in gastric cancer. CerS6 overexpression corelates with poor patient survival and CerS6 knockdown decreases proliferation, migration, and invasion of gastric cancer cells. The proposed mechanism is the downregulation of suppressor of cytokine signaling 2 (SOCS2) by overexpressed CerS6, leading to the activation of JAK-STAT signaling, followed by enhanced expression of genes involved in cell cycle progression (cyclins A and B) and metastasis (MMP-2 and -9) [179,181].
10. Conclusions
Fingolimod and Miglustat are FDA approved medications related to endogenous sphingolipids biochemistry. Targeting enzymes involved in sphingolipid biosynthesis, metabolism and catabolism showed promising hits for drug discovery efforts based on sphingolipidomics. Currently, there are several molecules in clinical studies pushing our understanding of the sphingolipid biology in several disease states, mainly cancer. As discussed, a functional lipid raft is made of sphingolipids. Small molecules, biologics targeting the signaling proteins embedded in these lipid rafts and the relevance of these proteins with sphingolipd flux is fertile for future investigations. So far, the strategies aimed at increasing cellular ceramide has opened an avenue for perturbing cellular ceramide biosynthesis and metabolism. Of the several hallmarks of cancer that promote chemoresistance, targeting ceramide metabolizing enzymes appears to be a promising drug target. The molecular objective is to increase cellular ceramide, and this has opened up a new avenue towards targeting resistant cancers based on membrane trafficking. In fact, using a combination approach along with an FDA approved regimens has promising applications.
Funding
This research was supported, in part, by funds from Biomedical and Pharmaceutical Sciences, College of Pharmacy, Idaho State University; National Science Foundation (NSF) MRI grant CHE-2019074; American Association of Colleges of Pharmacy NIA award – 2023; Office of Research – ISU (LIGS33); Pardee Foundation-Pardee Grant; Institute for Modeling Collaboration and Innovation, University of Idaho, COBRE: P20GM104420 for chemdraw software.
Acknowledgments
We wish to thank Dr. Apurbha Dutta, School of Pharmacy, The University of Kansas, and Dr. Webster L. Santos, Chemistry Department, Virginia Tech for their continued intellectual support in sphingolipid biochemistry; collaborator Dr. Ali Aghazadeh Habashi; and Drs. Amy E. Bryant, Marvin K. Schulte, Kavita Sharma, Biomedical and Pharmaceutical Sciences, Idaho State University, Pocatello, ID. for their interest and support.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| SL | Sphingolipid |
| S1P | Sphingosine-1-phosphate |
| CDases | Ceramidases |
| AC | Acid ceramidase |
| AML | Acute myeloid leukemia |
| HNC | Head and Neck Cancer |
| SMS | Sphingomyelin synthase |
| DES | Dihydroceramide desaturase |
| DHSph | Dihydrosphingosine |
| CerS | Ceramide synthase |
References
- Adada, M.; Luberto, C.; Canals, D. Inhibitors of the sphingomyelin cycle: Sphingomyelin synthases and sphingomyelinases. Chem. Phys. Lipids 2016, 197, 45–59. [Google Scholar] [CrossRef]
- Ogretmen, B.; Hannun, Y.A. Biologically active sphingolipids in cancer pathogenesis and treatment. Nat. Rev. Cancer 2004, 4, 604–616. [Google Scholar] [CrossRef]
- Beckham, T.H.; Elojeimy, S.; Cheng, J.C.; Turner, L.S.; Hoffman, S.R.; Norris, J.S.; Liu, X. Targeting sphingolipid metabolism in head and neck cancer: rational therapeutic potentials. Expert Opin. Ther. Targets 2010, 14, 529–539. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, B.; Zhang, J.; Zhang, R.; Zhang, Q. Short-chain C6 ceramide sensitizes AT406-induced anti-pancreatic cancer cell activity. Biochem. Biophys. Res. Commun. 2016, 479, 166–172. [Google Scholar] [CrossRef]
- Beckham, T.H.; Lu, P.; Jones, E.E.; Marrison, T.; Lewis, C.S.; Cheng, J.C.; Ramshesh, V.K.; Beeson, G.; Beeson, C.C.; Drake, R.R.; et al. LCL124, a Cationic Analog of Ceramide, Selectively Induces Pancreatic Cancer Cell Death by Accumulating in Mitochondria. Experiment 2012, 344, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Peters, G.; van der Wilt, C.; van Moorsel, C.; Kroep, J.; Bergman, A.; Ackland, S. Basis for effective combination cancer chemotherapy with antimetabolites. Pharmacol. Ther. 2000, 87, 227–253. [Google Scholar] [CrossRef] [PubMed]
- Modrak, D.E.; Leon, E.; Goldenberg, D.M.; Gold, D.V. Ceramide Regulates Gemcitabine-Induced Senescence and Apoptosis in Human Pancreatic Cancer Cell Lines. Mol. Cancer Res. 2009, 7, 890–896. [Google Scholar] [CrossRef]
- Castro, B.M.; Prieto, M.; Silva, L.C. Ceramide: A simple sphingolipid with unique biophysical properties. Prog. Lipid Res. 2014, 54, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Edvardson, S.; Yi, J.K.; Jalas, C.; Xu, R.; Webb, B.D.; Snider, J.; Fedick, A.; Kleinman, E.; Treff, N.R.; Mao, C.; et al. Deficiency of the alkaline ceramidase ACER3 manifests in early childhood by progressive leukodystrophy. J. Med Genet. 2016, 53, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A. and L.M. Obeid, Many ceramides. J Biol Chem, 2011. 286(32): p. 27855-62.
- Schulze, H.; Schepers, U.; Sandhoff, K. Overexpression and mass spectrometry analysis of mature human acid ceramidase. Biol. Chem. 2007, 388, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Obeid, L.M. Ceramidases: regulators of cellular responses mediated by ceramide, sphingosine, and sphingosine-1-phosphate. Biochim. et Biophys. Acta (BBA) - Mol. Cell Biol. Lipids 2008, 1781, 424–434. [Google Scholar] [CrossRef]
- Gatt, S. Enzymatic hydrolysis of sphingolipids. I. Hydrolysis and synthesis of ceramides by an enzyme from rat brain. J. Biol. Chem. 1966, 241, 3724–30. [Google Scholar] [CrossRef]
- Li, C.-M.; Park, J.-H.; He, X.; Levy, B.; Chen, F.; Arai, K.; Adler, D.A.; Disteche, C.M.; Koch, J.; Sandhoff, K.; et al. The Human Acid Ceramidase Gene (ASAH): Structure, Chromosomal Location, Mutation Analysis, and Expression. Genomics 1999, 62, 223–231. [Google Scholar] [CrossRef]
- He, X. , et al., Purification and characterization of recombinant, human acid ceramidase. Catalytic reactions and interactions with acid sphingomyelinase. J Biol Chem, 2003. 278(35): p. 32978-86.
- Azuma, N.; Obrien, J.; Moser, H.; Kishimoto, Y. Stimulation of Acid Ceramidase Activity by Saposin D. Arch. Biochem. Biophys. 1994, 311, 354–357. [Google Scholar] [CrossRef]
- Yu, F.P.S.; Amintas, S.; Levade, T.; Medin, J.A. Acid ceramidase deficiency: Farber disease and SMA-PME. Orphanet J. Rare Dis. 2018, 13, 1–19. [Google Scholar] [CrossRef]
- Li, C.M. , et al., Insertional mutagenesis of the mouse acid ceramidase gene leads to early embryonic lethality in homozygotes and progressive lipid storage disease in heterozygotes. Genomics, 2002. 79(2): p. 218-24.
- Mahdy, A.E.; Cheng, J.C.; Li, J.; Elojeimy, S.; Meacham, W.D.; Turner, L.S.; Bai, A.; Gault, C.R.; McPherson, A.S.; Garcia, N.; et al. Acid Ceramidase Upregulation in Prostate Cancer Cells Confers Resistance to Radiation: AC Inhibition, a Potential Radiosensitizer. Mol. Ther. 2009, 17, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.C.; Bai, A.; Beckham, T.H.; Marrison, S.T.; Yount, C.L.; Young, K.; Lu, P.; Bartlett, A.M.; Wu, B.X.; Keane, B.J.; et al. Radiation-induced acid ceramidase confers prostate cancer resistance and tumor relapse. J. Clin. Investig. 2013, 123, 4344–4358. [Google Scholar] [CrossRef] [PubMed]
- Camacho, L. , et al., Acid ceramidase as a therapeutic target in metastatic prostate cancer[S]. Journal of Lipid Research, 2013. 54(5): p. 1207-1220.
- Holman, D.H.; Turner, L.S.; El-Zawahry, A.; Elojeimy, S.; Liu, X.; Bielawski, J.; Szulc, Z.M.; Norris, K.; Zeidan, Y.H.; Hannun, Y.A.; et al. Lysosomotropic acid ceramidase inhibitor induces apoptosis in prostate cancer cells. Cancer Chemother. Pharmacol. 2007, 61, 231–242. [Google Scholar] [CrossRef]
- Gouazé-Andersson, V.; Flowers, M.; Karimi, R.; Fabriás, G.; Delgado, A.; Casas, J.; Cabot, M.C. Inhibition of acid ceramidase by a 2-substituted aminoethanol amide synergistically sensitizes prostate cancer cells to N-(4-hydroxyphenyl) retinamide. Prostate 2010, 71, 1064–1073. [Google Scholar] [CrossRef]
- Turner, L.S.; Cheng, J.C.; Beckham, T.H.; E Keane, T.; Norris, J.S.; Liu, X. Autophagy is increased in prostate cancer cells overexpressing acid ceramidase and enhances resistance to C6 ceramide. Prostate Cancer Prostatic Dis. 2010, 14, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.-L.; Park, J.Y.; Kim, E.H.; Jang, H.J. Targeting acid ceramidase sensitises head and neck cancer to cisplatin. Eur. J. Cancer 2015, 52, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Korbelik, M.; Banáth, J.; Zhang, W.; Saw, K.M.; Szulc, Z.M.; Bielawska, A.; Separovic, D. Interaction of acid ceramidase inhibitor LCL521 with tumor response to photodynamic therapy and photodynamic therapy-generated vaccine. Int. J. Cancer 2016, 139, 1372–1378. [Google Scholar] [CrossRef] [PubMed]
- Separovic, D.; Breen, P.; Boppana, N.B.; VAN Buren, E.; Joseph, N.; Kraveka, J.M.; Rahmaniyan, M.; Li, L.; Gudz, T.I.; Bielawska, A.; et al. Increased killing of SCCVII squamous cell carcinoma cells after the combination of Pc 4 photodynamic therapy and dasatinib is associated with enhanced caspase-3 activity and ceramide synthase 1 upregulation. Int. J. Oncol. 2013, 43, 2064–2072. [Google Scholar] [CrossRef]
- Elojeimy, S.; Liu, X.; Mckillop, J.C.; El-Zawahry, A.M.; Holman, D.H.; Cheng, J.Y.; Meacham, W.D.; Mahdy, A.E.; Saad, A.F.; Turner, L.S.; et al. Role of Acid Ceramidase in Resistance to FasL: Therapeutic Approaches Based on Acid Ceramidase Inhibitors and FasL Gene Therapy. Mol. Ther. 2007, 15, 1259–1263. [Google Scholar] [CrossRef]
- Realini, N. , et al., Acid Ceramidase in Melanoma: expression, localization, and effects of pharmacological inhibition*. Journal of Biological Chemistry, 2016. 291(5): p. 2422-2434.
- Bedia, C.; Casas, J.; Andrieu-Abadie, N.; Fabriàs, G.; Levade, T. Acid Ceramidase Expression Modulates the Sensitivity of A375 Melanoma Cells to Dacarbazine. J. Biol. Chem. 2011, 286, 28200–28209. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.; Realini, N.; La Ferla, M.; Passalacqua, I.; Matteoli, G.; Ganesan, A.; Pistello, M.; Mazzanti, C.M.; Piomelli, D. Complete Acid Ceramidase ablation prevents cancer-initiating cell formation in melanoma cells. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Tan, S.-F.; Liu, X.; Fox, T.E.; Barth, B.M.; Sharma, A.; Turner, S.D.; Awwad, A.; Dewey, A.; Doi, K.; Spitzer, B.; et al. Acid ceramidase is upregulated in AML and represents a novel therapeutic target. Oncotarget 2016, 7, 83208–83222. [Google Scholar] [CrossRef]
- Hu, X.; Yang, D.; Zimmerman, M.; Liu, F.; Yang, J.; Kannan, S.; Burchert, A.; Szulc, Z.; Bielawska, A.; Ozato, K.; et al. IRF8 Regulates Acid Ceramidase Expression to Mediate Apoptosis and Suppresses Myelogeneous Leukemia. Cancer Res 2011, 71, 2882–2891. [Google Scholar] [CrossRef] [PubMed]
- Molina, A.R.d. , et al., Acid Ceramidase as a Chemotherapeutic Target to Overcome Resistance to the Antitumoral Effect of Choline Kinase α Inhibition. Current Cancer Drug Targets, 2012. 12(6): p. 617-624.
- Sänger, N.; Ruckhäberle, E.; Györffy, B.; Engels, K.; Heinrich, T.; Fehm, T.; Graf, A.; Holtrich, U.; Becker, S.; Karn, T. Acid ceramidase is associated with an improved prognosis in both DCIS and invasive breast cancer. Mol. Oncol. 2014, 9, 58–67. [Google Scholar] [CrossRef]
- Flowers, M.; Fabriás, G.; Delgado, A.; Casas, J.; Abad, J.L.; Cabot, M.C. C6-Ceramide and targeted inhibition of acid ceramidase induce synergistic decreases in breast cancer cell growth. Breast Cancer Res. Treat. 2011, 133, 447–458. [Google Scholar] [CrossRef]
- Vethakanraj, H.S.; Sesurajan, B.P.; Padmanaban, V.P.; Jayaprakasam, M.; Murali, S.; Sekar, A.K. Anticancer effect of acid ceramidase inhibitor ceranib-2 in human breast cancer cell lines MCF-7, MDA MB-231 by the activation of SAPK/JNK, p38 MAPK apoptotic pathways, inhibition of the Akt pathway, downregulation of ERα. Anti-Cancer Drugs 2018, 29, 50–60. [Google Scholar] [CrossRef]
- Kus, G.; Kabadere, S.; Uyar, R.; Kutlu, H.M. Induction of apoptosis in prostate cancer cells by the novel ceramidase inhibitor ceranib-2. Vitr. Cell. Dev. Biol. - Anim. 2015, 51, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Hanker, L.C.M.; Karn, T.M.; Holtrich, U.M.; Gätje, R.; Rody, A.; Heinrich, T.M.; Ruckhäberle, E.M.; Engels, K.M. Acid Ceramidase (AC)—A Key Enzyme of Sphingolipid Metabolism—Correlates With Better Prognosis in Epithelial Ovarian Cancer. Int. J. Gynecol. Pathol. 2013, 32, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.; París, R.; Villanueva, A.; Llacuna, L.; García-Ruiz, C.; Fernández-Checa, J.C. Pharmacological inhibition or small interfering RNA targeting acid ceramidase sensitizes hepatoma cells to chemotherapy and reduces tumor growth in vivo. Oncogene 2006, 26, 905–916. [Google Scholar] [CrossRef]
- Giovannetti, E.; Leon, L.G.; Bertini, S.; Macchia, M.; Minutolo, F.; Funel, N.; Alecci, C.; Giancola, F.; Danesi, R.; Peters, G.J. Study of Apoptosis Induction and Deoxycytidine Kinase/Cytidine Deaminase Modulation in the Synergistic Interaction of a Novel Ceramide Analog and Gemcitabine in Pancreatic Cancer Cells. Nucleosides, Nucleotides Nucleic Acids 2010, 29, 419–426. [Google Scholar] [CrossRef]
- Klobučar, M.; Grbčić, P.; Pavelić, S.K.; Jonjić, N.; Visentin, S.; Sedić, M. Acid ceramidase inhibition sensitizes human colon cancer cells to oxaliplatin through downregulation of transglutaminase 2 and β1 integrin/FAK−mediated signalling. Biochem. Biophys. Res. Commun. 2018, 503, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Baspinar, M.; Ozyurt, R.; Kus, G.; Kutlay, O.; Ozkurt, M.; Erkasap, N.; Kabadere, S.; Yasar, N.F.; Erkasap, S. Effects of ceranib-2 on cell survival and TNF-alpha in colon cancer cell line. Bratisl. Med J. 2017, 118, 391–393. [Google Scholar] [CrossRef]
- Hait, N.C.; Allegood, J.; Maceyka, M.; Strub, G.M.; Harikumar, K.B.; Singh, S.K.; Luo, C.; Marmorstein, R.; Kordula, T.; Milstien, S.; et al. Regulation of Histone Acetylation in the Nucleus by Sphingosine-1-Phosphate. Science 2009, 325, 1254–1257. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, S.; Milstien, S. The outs and the ins of sphingosine-1-phosphate in immunity. Nat. Rev. Immunol. 2011, 11, 403–415. [Google Scholar] [CrossRef]
- Kunkel, G.T.; Maceyka, M.; Milstien, S.; Spiegel, S. Targeting the sphingosine-1-phosphate axis in cancer, inflammation and beyond. Nat. Rev. Drug Discov. 2013, 12, 688–702. [Google Scholar] [CrossRef]
- Truman, J.-P.; García-Barros, M.; Obeid, L.M.; Hannun, Y.A. Evolving concepts in cancer therapy through targeting sphingolipid metabolism. Biochim. et Biophys. Acta (BBA) - Mol. Cell Biol. Lipids 2013, 1841, 1174–1188. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, M. , WANTED: Natural-Born Sickler. Science Translational Medicine, 2014. 6(240): p. 240ec101.
- Heeney, M.; Renella, R.; Y, Z.; V, B.; A, S.; K, S.; W, W.; W, Z.; C, N.; C, L.; et al. Faculty Opinions recommendation of Elevated sphingosine-1-phosphate promotes sickling and sickle cell disease progression. . 2014, 124. [Google Scholar] [CrossRef]
- Balakumar, P.; Kaur, T.; Singh, M. Potential target sites to modulate vascular endothelial dysfunction: Current perspectives and future directions. Toxicology 2008, 245, 49–64. [Google Scholar] [CrossRef]
- Machida, T.; Hamaya, Y.; Izumi, S.; Hamaya, Y.; Iizuka, K.; Igarashi, Y.; Minami, M.; Levi, R.; Hirafuji, M. Sphingosine 1-Phosphate Inhibits Nitric Oxide Production Induced by Interleukin-1β in Rat Vascular Smooth Muscle Cells. Experiment 2008, 325, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, R.; Bertolino, A.; Terlizzi, M.; Iacono, V.M.; Maiolino, P.; Cirino, G.; Roviezzo, F.; Pinto, A. B Cell Depletion Increases Sphingosine-1-Phosphate–Dependent Airway Inflammation in Mice. Am. J. Respir. Cell Mol. Biol. 2015, 52, 571–583. [Google Scholar] [CrossRef]
- Price, M.M.; Oskeritzian, C.A.; Falanga, Y.T.; Harikumar, K.B.; Allegood, J.C.; Alvarez, S.E.; Conrad, D.; Ryan, J.J.; Milstien, S.; Spiegel, S. A specific sphingosine kinase 1 inhibitor attenuates airway hyperresponsiveness and inflammation in a mast cell–dependent murine model of allergic asthma. J. Allergy Clin. Immunol. 2012, 131, 501–511. [Google Scholar] [CrossRef]
- Pyne, N.J. G. Dubois, and S. Pyne, Role of sphingosine 1-phosphate and lysophosphatidic acid in fibrosis. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids, 2013. 1831(1): p. 228-238.
- Maceyka, M.; Sankala, H.; Hait, N.C.; Le Stunff, H.; Liu, H.; Toman, R.; Collier, C.; Zhang, M.; Satin, L.S.; Merrill, A.H.; et al. SphK1 and SphK2, Sphingosine Kinase Isoenzymes with Opposing Functions in Sphingolipid Metabolism. PEDIATRICS 2005, 280, 37118–37129. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, H.A.; Pitson, S.M. Roles, regulation and inhibitors of sphingosine kinase 2. FEBS J. 2013, 280, 5317–5336. [Google Scholar] [CrossRef]
- Mizugishi, K.; Yamashita, T.; Olivera, A.; Miller, G.F.; Spiegel, S.; Proia, R.L. Essential Role for Sphingosine Kinases in Neural and Vascular Development. Mol. Cell. Biol. 2005, 25, 11113–11121. [Google Scholar] [CrossRef]
- French, K.J.; Zhuang, Y.; Maines, L.W.; Gao, P.; Wang, W.; Beljanski, V.; Upson, J.J.; Green, C.L.; Keller, S.N.; Smith, C.D. Pharmacology and Antitumor Activity of ABC294640, a Selective Inhibitor of Sphingosine Kinase-2. Experiment 2010, 333, 129–139. [Google Scholar] [CrossRef]
- Maines, L.W.; Fitzpatrick, L.R.; French, K.J.; Zhuang, Y.; Xia, Z.; Keller, S.N.; Upson, J.J.; Smith, C.D. Suppression of Ulcerative Colitis in Mice by Orally Available Inhibitors of Sphingosine Kinase. Dig. Dis. Sci. 2007, 53, 997–1012. [Google Scholar] [CrossRef] [PubMed]
- Maines, L.W.; Fitzpatrick, L.R.; Green, C.L.; Zhuang, Y.; Smith, C.D. Efficacy of a novel sphingosine kinase inhibitor in experimental Crohn’s disease. Inflammopharmacology 2010, 18, 73–85. [Google Scholar] [CrossRef]
- Shi, Y.; Rehman, H.; Ramshesh, V.K.; Schwartz, J.; Liu, Q.; Krishnasamy, Y.; Zhang, X.; Lemasters, J.J.; Smith, C.D.; Zhong, Z. Sphingosine kinase-2 inhibition improves mitochondrial function and survival after hepatic ischemia–reperfusion. J. Hepatol. 2012, 56, 137–145. [Google Scholar] [CrossRef]
- Chumanevich, A.A.; Poudyal, D.; Cui, X.; Davis, T.; Wood, P.A.; Smith, C.D.; Hofseth, L.J. Suppression of colitis-driven colon cancer in mice by a novel small molecule inhibitor of sphingosine kinase. Carcinog. 2010, 31, 1787–1793. [Google Scholar] [CrossRef]
- Xun, C.; Chen, M.-B.; Qi, L.; Tie-Ning, Z.; Peng, X.; Ning, L.; Zhi-Xiao, C.; Li-Wei, W. Targeting sphingosine kinase 2 (SphK2) by ABC294640 inhibits colorectal cancer cell growth in vitro and in vivo. J. Exp. Clin. Cancer Res. 2015, 34, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Antoon, J.W.; White, M.D.; Meacham, W.D.; Slaughter, E.M.; Muir, S.E.; Elliott, S.; Rhodes, L.V.; Ashe, H.B.; Wiese, T.E.; Smith, C.D.; et al. Antiestrogenic Effects of the Novel Sphingosine Kinase-2 Inhibitor ABC294640. Endocrinology 2010, 151, 5124–5135. [Google Scholar] [CrossRef]
- Hara-Yokoyama, M.; Terasawa, K.; Ichinose, S.; Watanabe, A.; Podyma-Inoue, K.A.; Akiyoshi, K.; Igarashi, Y.; Yanagishita, M. Sphingosine kinase 2 inhibitor SG-12 induces apoptosis via phosphorylation by sphingosine kinase 2. Bioorganic Med. Chem. Lett. 2013, 23, 2220–2224. [Google Scholar] [CrossRef]
- Lim, K.G.; Sun, C.; Bittman, R.; Pyne, N.J.; Pyne, S. (R)-FTY720 methyl ether is a specific sphingosine kinase 2 inhibitor: Effect on sphingosine kinase 2 expression in HEK 293 cells and actin rearrangement and survival of MCF-7 breast cancer cells. Cell. Signal. 2011, 23, 1590–1595. [Google Scholar] [CrossRef]
- Liu, K.; Guo, T.L.; Hait, N.C.; Allegood, J.; Parikh, H.I.; Xu, W.; Kellogg, G.E.; Grant, S.; Spiegel, S.; Zhang, S. Biological Characterization of 3-(2-amino-ethyl)-5-[3-(4-butoxyl-phenyl)-propylidene]-thiazolidine-2,4-dione (K145) as a Selective Sphingosine Kinase-2 Inhibitor and Anticancer Agent. PLOS ONE 2013, 8, e56471. [Google Scholar] [CrossRef] [PubMed]
- Raje, M.R.; Knott, K.; Kharel, Y.; Bissel, P.; Lynch, K.R.; Santos, W.L. Design, synthesis and biological activity of sphingosine kinase 2 selective inhibitors. Bioorganic Med. Chem. 2011, 20, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Childress, E.S.; Kharel, Y.; Brown, A.M.; Bevan, D.R.; Lynch, K.R.; Santos, W.L. Transforming Sphingosine Kinase 1 Inhibitors into Dual and Sphingosine Kinase 2 Selective Inhibitors: Design, Synthesis, and in Vivo Activity. J. Med. Chem. 2017, 60, 3933–3957. [Google Scholar] [CrossRef]
- Schnute, M.E.; McReynolds, M.D.; Carroll, J.; Chrencik, J.; Highkin, M.K.; Iyanar, K.; Jerome, G.; Rains, J.W.; Saabye, M.; Scholten, J.A.; et al. Discovery of a Potent and Selective Sphingosine Kinase 1 Inhibitor through the Molecular Combination of Chemotype-Distinct Screening Hits. J. Med. Chem. 2017, 60, 2562–2572. [Google Scholar] [CrossRef]
- Kharel, Y.; Raje, M.; Gao, M.; Gellett, A.M.; Tomsig, J.L.; Lynch, K.R.; Santos, W.L. Sphingosine kinase type 2 inhibition elevates circulating sphingosine 1-phosphate. Biochem. J. 2012, 447, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Patwardhan, N.N.; Morris, E.A.; Kharel, Y.; Raje, M.R.; Gao, M.; Tomsig, J.L.; Lynch, K.R.; Santos, W.L. Structure−Activity Relationship Studies and in Vivo Activity of Guanidine-Based Sphingosine Kinase Inhibitors: Discovery of SphK1- and SphK2-Selective Inhibitors. J. Med. Chem. 2015, 58, 1879–1899. [Google Scholar] [CrossRef] [PubMed]
- Kharel, Y.; Morris, E.A.; Congdon, M.D.; Thorpe, S.B.; Tomsig, J.L.; Santos, W.L.; Lynch, K.R. Sphingosine Kinase 2 Inhibition and Blood Sphingosine 1-Phosphate Levels. Experiment 2015, 355, 23–31. [Google Scholar] [CrossRef]
- Congdon, M.D.; Kharel, Y.; Brown, A.M.; Lewis, S.N.; Bevan, D.R.; Lynch, K.R.; Santos, W.L. Structure–Activity Relationship Studies and Molecular Modeling of Naphthalene-Based Sphingosine Kinase 2 Inhibitors. ACS Med. Chem. Lett. 2016, 7, 229–234. [Google Scholar] [CrossRef]
- Futerman, A.H.; Stieger, B.; Hubbard, A.L.; E Pagano, R. Sphingomyelin synthesis in rat liver occurs predominantly at the cis and medial cisternae of the Golgi apparatus. PEDIATRICS 1990, 265, 8650–8657. [Google Scholar] [CrossRef]
- Huitema, K.; van den Dikkenberg, J.; Brouwers, J.F.H.M.; Holthuis, J.C.M. Identification of a family of animal sphingomyelin synthases. EMBO J. 2004, 23, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Albi, E.; Lazzarini, R.; Magni, M.V. Reverse sphingomyelin-synthase in rat liver chromatin. FEBS Lett. 2003, 549, 152–156. [Google Scholar] [CrossRef]
- Taniguchi, M. and T. Okazaki, The role of sphingomyelin and sphingomyelin synthases in cell death, proliferation and migration-from cell and animal models to human disorders. Biochim Biophys Acta, 2014. 1841(5): p. 692-703.
- Yamaoka, S.; Miyaji, M.; Kitano, T.; Umehara, H.; Okazaki, T. Expression Cloning of a Human cDNA Restoring Sphingomyelin Synthesis and Cell Growth in Sphingomyelin Synthase-defective Lymphoid Cells. Perspect. Surg. 2004, 279, 18688–18693. [Google Scholar] [CrossRef] [PubMed]
- Luberto, C. and Y.A. Hannun, Sphingomyelin Synthase, a Potential Regulator of Intracellular Levels of Ceramide and Diacylglycerol during SV40 Transformation: does sphingomyelin synthase account for the putative phosphatidylcholine-specific phospholipase c? *. Journal of Biological Chemistry, 1998. 273(23): p. 14550-14559.
- Riboni, L. , et al., Basic Fibroblast Growth Factor-induced Proliferation of Primary Astrocytes: evidence for the involvement of sphingomyelin biosynthesis *. Journal of Biological Chemistry, 2001. 276(16): p. 12797-12804.
- Hannun, Y.A.; Obeid, L.M. The Ceramide-centric Universe of Lipid-mediated Cell Regulation: Stress Encounters of the Lipid Kind. PEDIATRICS 2002, 277, 25847–25850. [Google Scholar] [CrossRef]
- Wiegmann, K.; Schütze, S.; Machleidt, T.; Witte, D.; Krönke, M. Functional dichotomy of neutral and acidic sphingomyelinases in tumor necrosis factor signaling. Cell 1994, 78, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Adam-Klages, S.; Adam, D.; Wiegmann, K.; Struve, S.; Kolanus, W.; Schneider-Mergener, J.; Krönke, M. FAN, a Novel WD-Repeat Protein, Couples the p55 TNF-Receptor to Neutral Sphingomyelinase. Cell 1996, 86, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef]
- Holthuis, J.C.M.; Pomorski, T.; Raggers, R.J.; Sprong, H.; Van Meer, G.; Zheng, T.; Li, W.; Altura, B.T.; Shah, N.C.; Altura, B.M.; et al. The Organizing Potential of Sphingolipids in Intracellular Membrane Transport. Physiol. Rev. 2001, 81, 1689–1723. [Google Scholar] [CrossRef]
- Baron, C.L.; Malhotra, V. Role of Diacylglycerol in PKD Recruitment to the TGN and Protein Transport to the Plasma Membrane. Science 2002, 295, 325–328. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. The Ceramide-centric Universe of Lipid-mediated Cell Regulation: Stress Encounters of the Lipid Kind. PEDIATRICS 2002, 277, 25847–25850. [Google Scholar] [CrossRef]
- Hampton, R.Y.; Morand, O.H. Sphingomyelin Synthase and PKC Activation. Science 1989, 246, 1050–1050. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez, Y.F.; González, J.; Bedia, C.; Casas, J.; Abad, J.L.; Delgado, A.; Fabrias, G. 3-Ketosphinganine provokes the accumulation of dihydroshingolipids and induces autophagy in cancer cells. Mol. Biosyst. 2016, 12, 1166–1173. [Google Scholar] [CrossRef]
- Cadena, D.L.; Kurten, R.C.; Gill, G.N. The Product of the MLD Gene Is a Member of the Membrane Fatty Acid Desaturase Family: Overexpression of MLD Inhibits EGF Receptor Biosynthesis, Biochemistry 1997, 36, 6960–6967. [Google Scholar] [CrossRef] [PubMed]
- Geeraert, L.; Mannaerts, G.P.; VAN Veldhoven, P.P. Conversion of dihydroceramide into ceramide: involvement of a desaturase. Biochem. J. 1997, 327, 125–132. [Google Scholar] [CrossRef]
- Michel, C. , et al., Characterization of ceramide synthesis. A dihydroceramide desaturase introduces the 4,5-trans-double bond of sphingosine at the level of dihydroceramide. J Biol Chem, 1997. 272(36): p. 22432-7.
- Savile, C.K. G. Fabriàs, and P.H. Buist, Dihydroceramide delta(4) desaturase initiates substrate oxidation at C-4. J Am Chem Soc, 2001. 123(19): p. 4382-5.
- Beauchamp, E. , et al., Myristic acid increases the activity of dihydroceramide Delta4-desaturase 1 through its N-terminal myristoylation. Biochimie, 2007. 89(12): p. 1553-61.
- Ternes, P. , et al., Identification and characterization of a sphingolipid delta 4-desaturase family. J Biol Chem, 2002. 277(28): p. 25512-8.
- Casasampere, M.; Ordoñez, Y.F.; Pou, A.; Casas, J. Inhibitors of dihydroceramide desaturase 1: Therapeutic agents and pharmacological tools to decipher the role of dihydroceramides in cell biology. Chem. Phys. Lipids 2016, 197, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, Y.; Kihara, A.; Igarashi, Y. Identification of the human sphingolipid C4-hydroxylase, hDES2, and its up-regulation during keratinocyte differentiation. FEBS Lett. 2004, 563, 93–97. [Google Scholar] [CrossRef]
- Siddique, M.M.; Bikman, B.T.; Wang, L.; Ying, L.; Reinhardt, E.; Shui, G.; Wenk, M.R.; Summers, S.A. Ablation of Dihydroceramide Desaturase Confers Resistance to Etoposide-Induced Apoptosis In Vitro. PLOS ONE 2012, 7, e44042. [Google Scholar] [CrossRef]
- Holland, W.L.; Brozinick, J.T.; Wang, L.-P.; Hawkins, E.D.; Sargent, K.M.; Liu, Y.; Narra, K.; Hoehn, K.L.; Knotts, T.A.; Siesky, A.; et al. Inhibition of Ceramide Synthesis Ameliorates Glucocorticoid-, Saturated-Fat-, and Obesity-Induced Insulin Resistance. Cell Metab. 2007, 5, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef]
- Testai, F.D.; Kilkus, J.P.; Berdyshev, E.; Gorshkova, I.; Natarajan, V.; Dawson, G. Multiple sphingolipid abnormalities following cerebral microendothelial hypoxia. J. Neurochem. 2014, 131, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Testai, F.D.; Xu, H.-L.; Kilkus, J.; Suryadevara, V.; Gorshkova, I.; Berdyshev, E.; Pelligrino, D.A.; Dawson, G. Changes in the metabolism of sphingolipids after subarachnoid hemorrhage. J. Neurosci. Res. 2015, 93, 796–805. [Google Scholar] [CrossRef]
- Mielke, M.M.; Haughey, N.J.; Bandaru, V.V.R.; Weinberg, D.D.; Darby, E.; Zaidi, N.; Pavlik, V.; Doody, R.S.; Lyketsos, C.G. Plasma Sphingomyelins are Associated with Cognitive Progression in Alzheimer's Disease. J. Alzheimer's Dis. 2011, 27, 259–269. [Google Scholar] [CrossRef]
- Di Pardo, A.; Basit, A.; Armirotti, A.; Amico, E.; Castaldo, S.; Pepe, G.; Marracino, F.; Buttari, F.; Digilio, A.F.; Maglione, V. De novo Synthesis of Sphingolipids Is Defective in Experimental Models of Huntington's Disease. Front. Neurosci. 2017, 11, 698. [Google Scholar] [CrossRef]
- González-Peña, D.; Checa, A.; de Ancos, B.; Wheelock, C.E.; Sánchez-Moreno, C. New insights into the effects of onion consumption on lipid mediators using a diet-induced model of hypercholesterolemia. Redox Biol. 2017, 11, 205–212. [Google Scholar] [CrossRef]
- Maeng, H.J. , et al., Celecoxib-mediated activation of endoplasmic reticulum stress induces de novo ceramide biosynthesis and apoptosis in hepatoma HepG2 cells mobilization. BMB Rep, 2017. 50(3): p. 144-149.
- Edsfeldt, A.; Dunér, P.; Ståhlman, M.; Mollet, I.G.; Asciutto, G.; Grufman, H.; Nitulescu, M.; Persson, A.F.; Fisher, R.M.; Melander, O.; et al. Sphingolipids Contribute to Human Atherosclerotic Plaque Inflammation. Arter. Thromb. Vasc. Biol. 2016, 36, 1132–1140. [Google Scholar] [CrossRef]
- Fang, L.; Mundra, P.A.; Fan, F.; Galvin, A.; Weir, J.M.; Wong, G.; Chin-Dusting, J.; Cicuttini, F.; Meikle, P.; Dart, A.M. Plasma lipidomic profiling in patients with rheumatoid arthritis. Metabolomics 2016, 12, 136. [Google Scholar] [CrossRef]
- Ji, R.; Chang, J.Y.; Liao, X.; Zhang, X.; Kennel, P.; Castillero, E.; Brunjes, D.; Akashi, H.; Homma, S.; Goldberg, I.; et al. Abstract 17320: Inhibition of Ceramide Synthesis Preserves Cardiac Function and Increases Survival in Doxorubicin-induced Cardiomyopathy. Circulation 2015, 132. [Google Scholar] [CrossRef]
- Park, M.A. , et al., Vorinostat and Sorafenib Increase CD95 Activation in Gastrointestinal Tumor Cells through a Ca2+-De novo Ceramide-PP2A-Reactive Oxygen Species–Dependent Signaling Pathway. Cancer Research, 2010. 70(15): p. 6313.
- Gencer, E.B.; Ural, A.U.; Avcu, F.; Baran, Y. A novel mechanism of dasatinib-induced apoptosis in chronic myeloid leukemia; ceramide synthase and ceramide clearance genes. Ann. Hematol. 2011, 90, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Kollmeyer, J.; Symolon, H.; Momin, A.; Munter, E.; Wang, E.; Kelly, S.; Allegood, J.C.; Liu, Y.; Peng, Q.; et al. Ceramides and other bioactive sphingolipid backbones in health and disease: Lipidomic analysis, metabolism and roles in membrane structure, dynamics, signaling and autophagy. Biochim. et Biophys. Acta (BBA) - Biomembr. 2006, 1758, 1864–1884. [Google Scholar] [CrossRef] [PubMed]
- Casasampere, M.; Ordóñez, Y.F.; Casas, J.; Fabrias, G. Dihydroceramide desaturase inhibitors induce autophagy via dihydroceramide-dependent and independent mechanisms. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 2017, 1861, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, P.; Munoz-Olaya, J.M.; Gagliostro, V.; Casas, J.; Ghidoni, R.; Fabriàs, G. Dihydroceramide intracellular increase in response to resveratrol treatment mediates autophagy in gastric cancer cells. Cancer Lett. 2009, 282, 238–243. [Google Scholar] [CrossRef]
- Grenald, S.A.; Doyle, T.M.; Zhang, H.; Slosky, L.M.; Chen, Z.; Largent-Milnes, T.M.; Spiegel, S.; Vanderah, T.W.; Salvemini, D. Targeting the S1P/S1PR1 axis mitigates cancer-induced bone pain and neuroinflammation. Pain 2017, 158, 1733–1742. [Google Scholar] [CrossRef]
- Holliday, M.W. , et al., C22:0- and C24:0-dihydroceramides confer mixed cytotoxicity in T-cell acute lymphoblastic leukemia cell lines. PLoS One, 2013. 8(9): p. e74768.
- Knapp, P.; Baranowski, M.; Knapp, M.; Zabielski, P.; Błachnio-Zabielska, A.U.; Górski, J. Altered sphingolipid metabolism in human endometrial cancer. Prostaglandins Other Lipid Mediat. 2010, 92, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Realini, N. , et al., Acid Ceramidase in Melanoma: expression, localization, and effects of pharmacological inhibition. J Biol Chem, 2016. 291(5): p. 2422-34.
- Illuzzi, G.; Bernacchioni, C.; Aureli, M.; Prioni, S.; Frera, G.; Donati, C.; Valsecchi, M.; Chigorno, V.; Bruni, P.; Sonnino, S.; et al. Sphingosine Kinase Mediates Resistance to the Synthetic Retinoid N-(4-Hydroxyphenyl)retinamide in Human Ovarian Cancer Cells. J. Biol. Chem. 2010, 285, 18594–18602. [Google Scholar] [CrossRef]
- Noack, J.; Choi, J.; Richter, K.; Kopp-Schneider, A.; Régnier-Vigouroux, A. A sphingosine kinase inhibitor combined with temozolomide induces glioblastoma cell death through accumulation of dihydrosphingosine and dihydroceramide, endoplasmic reticulum stress and autophagy. Cell Death Dis. 2014, 5, e1425–e1425. [Google Scholar] [CrossRef]
- Valsecchi, M.; Aureli, M.; Mauri, L.; Illuzzi, G.; Chigorno, V.; Prinetti, A.; Sonnino, S. Sphingolipidomics of A2780 human ovarian carcinoma cells treated with synthetic retinoids. J. Lipid Res. 2010, 51, 1832–1840. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Maurer, B.J.; Liu, Y.-Y.; Wang, E.; Allegood, J.C.; Kelly, S.; Symolon, H.; Liu, Y.; Merrill, A.H.; Gouazé-Andersson, V.; et al. N-(4-Hydroxyphenyl)retinamide increases dihydroceramide and synergizes with dimethylsphingosine to enhance cancer cell killing. Mol. Cancer Ther. 2008, 7, 2967–2976. [Google Scholar] [CrossRef] [PubMed]
- Idkowiak-Baldys, J.; Apraiz, A.; Li, L.; Rahmaniyan, M.; Clarke, C.J.; Kraveka, J.M.; Asumendi, A.; Hannun, Y.A. Dihydroceramide desaturase activity is modulated by oxidative stress. Biochem. J. 2010, 427, 265–274. [Google Scholar] [CrossRef]
- Venant, H.; Rahmaniyan, M.; Jones, E.E.; Lu, P.; Lilly, M.B.; Garrett-Mayer, E.; Drake, R.R.; Kraveka, J.M.; Smith, C.D.; Voelkel-Johnson, C. The Sphingosine Kinase 2 Inhibitor ABC294640 Reduces the Growth of Prostate Cancer Cells and Results in Accumulation of Dihydroceramides In Vitro and In Vivo. Mol. Cancer Ther. 2015, 14, 2744–2752. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Corbacho, M.J. , et al., Tumor Necrosis Factor-α (TNFα)-induced Ceramide Generation via Ceramide Synthases Regulates Loss of Focal Adhesion Kinase (FAK) and Programmed Cell Death. J Biol Chem, 2015. 290(42): p. 25356-73.
- McNair, C.; Urbanucci, A.; Comstock, C.E.S.; A Augello, M.; Goodwin, J.F.; Launchbury, R.; Zhao, S.G.; Schiewer, M.J.; Ertel, A.; Karnes, J.; et al. Cell cycle-coupled expansion of AR activity promotes cancer progression. Oncogene 2016, 36, 1655–1668. [Google Scholar] [CrossRef]
- Zhou, W.; Ye, X.-L.; Sun, Z.-J.; Ji, X.-D.; Chen, H.-X.; Xie, D. Overexpression of degenerative spermatocyte homolog 1 up-regulates the expression of cyclin D1 and enhances metastatic efficiency in esophageal carcinoma Eca109 cells. Mol. Carcinog. 2009, 48, 886–894. [Google Scholar] [CrossRef]
- Pewzner-Jung, Y. S. Ben-Dor, and A.H. Futerman, When do Lasses (longevity assurance genes) become CerS (ceramide synthases)?: Insights into the regulation of ceramide synthesis. J Biol Chem, 2006. 281(35): p. 25001-5.
- Lahiri, S. , et al., Kinetic characterization of mammalian ceramide synthases: determination of K(m) values towards sphinganine. FEBS Lett, 2007. 581(27): p. 5289-94.
- Venkataraman, K. , et al., Upstream of growth and differentiation factor 1 (uog1), a mammalian homolog of the yeast longevity assurance gene 1 (LAG1), regulates N-stearoyl-sphinganine (C18-(dihydro)ceramide) synthesis in a fumonisin B1-independent manner in mammalian cells. J Biol Chem, 2002. 277(38): p. 35642-9.
- Sridevi, P.; Alexander, H.; Laviad, E.L.; Pewzner-Jung, Y.; Hannink, M.; Futerman, A.H.; Alexander, S. Ceramide synthase 1 is regulated by proteasomal mediated turnover. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2009, 1793, 1218–1227. [Google Scholar] [CrossRef]
- Abbott, S.K.; Li, H.; Muñoz, S.S.; Knoch, B.; Batterham, M.; Murphy, K.E.; Halliday, G.M.; Garner, B. Altered ceramide acyl chain length and ceramide synthase gene expression in Parkinson's disease. Mov. Disord. 2013, 29, 518–526. [Google Scholar] [CrossRef]
- Mielke, M.M.; Maetzler, W.; Haughey, N.J.; Bandaru, V.V.R.; Savica, R.; Deuschle, C.; Gasser, T.; Hauser, A.-K.; Gräber-Sultan, S.; Schleicher, E.; et al. Plasma Ceramide and Glucosylceramide Metabolism Is Altered in Sporadic Parkinson's Disease and Associated with Cognitive Impairment: A Pilot Study. PLOS ONE 2013, 8, e73094. [Google Scholar] [CrossRef]
- Eberle, M.; Ebel, P.; Wegner, M.-S.; Männich, J.; Tafferner, N.; Ferreiros, N.; Birod, K.; Schreiber, Y.; Krishnamoorthy, G.; Willecke, K.; et al. Regulation of ceramide synthase 6 in a spontaneous experimental autoimmune encephalomyelitis model is sex dependent. Biochem. Pharmacol. 2014, 92, 326–335. [Google Scholar] [CrossRef]
- Cinar, R. , et al., Hepatic cannabinoid-1 receptors mediate diet-induced insulin resistance by increasing de novo synthesis of long-chain ceramides. Hepatology, 2014. 59(1): p. 143-53.
- Karahatay, S.; Thomas, K.; Koybasi, S.; Senkal, C.E.; ElOjeimy, S.; Liu, X.; Bielawski, J.; Day, T.A.; Gillespie, M.B.; Sinha, D.; et al. Clinical relevance of ceramide metabolism in the pathogenesis of human head and neck squamous cell carcinoma (HNSCC): Attenuation of C18-ceramide in HNSCC tumors correlates with lymphovascular invasion and nodal metastasis. Cancer Lett. 2007, 256, 101–111. [Google Scholar] [CrossRef]
- Moro, K.; Kawaguchi, T.; Tsuchida, J.; Gabriel, E.; Qi, Q.; Yan, L.; Wakai, T.; Takabe, K.; Nagahashi, M. Ceramide species are elevated in human breast cancer and are associated with less aggressiveness. Oncotarget 2018, 9, 19874–19890. [Google Scholar] [CrossRef]
- Zhao, L.; Spassieva, S.D.; Jucius, T.J.; Shultz, L.D.; Shick, H.E.; Macklin, W.B.; Hannun, Y.A.; Obeid, L.M.; Ackerman, S.L. A Deficiency of Ceramide Biosynthesis Causes Cerebellar Purkinje Cell Neurodegeneration and Lipofuscin Accumulation. PLOS Genet. 2011, 7, e1002063. [Google Scholar] [CrossRef] [PubMed]
- Vanni, N.; Fruscione, F.; Ferlazzo, E.; Striano, P.; Robbiano, A.; Traverso, M.; Sander, T.; Falace, A.; Gazzerro, E.; Bramanti, P.; et al. Impairment of ceramide synthesis causes a novel progressive myoclonus epilepsy. Ann. Neurol. 2014, 76, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wen, L.; Zhu, F.; Wang, Y.; Xie, Q.; Chen, Z.; Li, Y. Overexpression of ceramide synthase 1 increases C18-ceramide and leads to lethal autophagy in human glioma. Oncotarget 2017, 8, 104022–104036. [Google Scholar] [CrossRef] [PubMed]
- Laviad, E.L. , et al., Characterization of ceramide synthase 2: tissue distribution, substrate specificity, and inhibition by sphingosine 1-phosphate. J Biol Chem, 2008. 283(9): p. 5677-84.
- Pan, H.; Qin, W.-X.; Huo, K.-K.; Wan, D.-F.; Yu, Y.; Xu, Z.-G.; Hu, Q.-D.; Gu, K.T.; Zhou, X.-M.; Jiang, H.-Q.; et al. Cloning, Mapping, and Characterization of a Human Homologue of the Yeast Longevity Assurance Gene LAG1. Genomics 2001, 77, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Petrache, I.; Kamocki, K.; Poirier, C.; Pewzner-Jung, Y.; Laviad, E.L.; Schweitzer, K.S.; Van Demark, M.; Justice, M.J.; Hubbard, W.C.; Futerman, A.H. Ceramide Synthases Expression and Role of Ceramide Synthase-2 in the Lung: Insight from Human Lung Cells and Mouse Models. PLOS ONE 2013, 8, e62968. [Google Scholar] [CrossRef]
- Schiffmann, S.; Sandner, J.; Birod, K.; Wobst, I.; Angioni, C.; Ruckhäberle, E.; Kaufmann, M.; Ackermann, H.; Lötsch, J.; Schmidt, H.; et al. Ceramide synthases and ceramide levels are increased in breast cancer tissue. Carcinog. 2009, 30, 745–752. [Google Scholar] [CrossRef]
- Laviad, E.L. , et al., Characterization of Ceramide Synthase 2: tissue distribution, substrate specificity, and inhibition by sphingosine 1-phosphate*. Journal of Biological Chemistry, 2008. 283(9): p. 5677-5684.
- Barthelmes, J.; de Bazo, A.M.; Pewzner-Jung, Y.; Schmitz, K.; Mayer, C.A.; Foerch, C.; Eberle, M.; Tafferner, N.; Ferreirós, N.; Henke, M.; et al. Lack of ceramide synthase 2 suppresses the development of experimental autoimmune encephalomyelitis by impairing the migratory capacity of neutrophils. Brain, Behav. Immun. 2015, 46, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Silva, J.; Dasgupta, S.; Bieberich, E. Long-chain ceramide is elevated in presenilin 1 (PS1M146V) mouse brain and induces apoptosis in PS1 astrocytes. Glia 2008, 56, 449–456. [Google Scholar] [CrossRef]
- Mosbech, M.; Olsen, A.S.B.; Neess, D.; Ben-David, O.; Klitten, L.L.; Larsen, J.; Sabers, A.; Vissing, J.; Nielsen, J.E.; Hasholt, L.; et al. Reduced ceramide synthase 2 activity causes progressive myoclonic epilepsy. Ann. Clin. Transl. Neurol. 2014, 1, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Wang, T.; Yang, C.; Jin, G.; Gu, D.; Deng, X.; Wang, C.; Qin, W.; Jin, H. Co-expression of LASS2 and TGF-β1 predicts poor prognosis in hepatocellular carcinoma. Sci. Rep. 2016, 6, 32421. [Google Scholar] [CrossRef]
- Fan, S.; Niu, Y.; Tan, N.; Wu, Z.; Wang, Y.; You, H.; Ke, R.; Song, J.; Shen, Q.; Wang, W.; et al. LASS2 enhances chemosensitivity of breast cancer by counteracting acidic tumor microenvironment through inhibiting activity of V-ATPase proton pump. Oncogene 2012, 32, 1682–1690. [Google Scholar] [CrossRef]
- Zeng, F. , et al., Overexpression of LASS2 inhibits proliferation and causes G0/G1 cell cycle arrest in papillary thyroid cancer. Cancer Cell International, 2018. 18(1): p. 151.
- Ke, R.-H.; Wang, Y.; Mao, Y.; Zhang, J.; Xiong, J. Decreased expression of LASS2 is associated with worse prognosis in meningiomas. J. Neuro-Oncology 2014, 118, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, J.; Zuo, Y.; Ding, M.; Yan, R.; Yang, D.; Ke, C. Expression and prognostic significance of a new tumor metastasis suppressor gene LASS2 in human bladder carcinoma. Med Oncol. 2011, 29, 1921–1927. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, W.; Zuo, Y.; Ding, M.; Ke, C.; Yan, R.; Zhan, H.; Liu, J.; Wang, J. miR-9 promotes cell proliferation and inhibits apoptosis by targeting LASS2 in bladder cancer. Tumor Biol. 2015, 36, 9631–9640. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, H.; Xiong, T.; Zou, R.; Tang, Z.; Wang, J. The role of LASS2 in regulating bladder cancer cell tumorigenicity in a nude mouse model. Oncol. Lett. 2017, 14, 5149–5156. [Google Scholar] [CrossRef]
- Xu, X. J. You, and F. Pei, Silencing of a novel tumor metastasis suppressor gene LASS2/TMSG1 promotes invasion of prostate cancer cell in vitro through increase of vacuolar ATPase activity. Journal of Cellular Biochemistry, 2012. 113(7): p. 2356-2363.
- Xu, X. , et al., Silencing of LASS2/TMSG1 enhances invasion and metastasis capacity of prostate cancer cell. Journal of Cellular Biochemistry, 2014. 115(4): p. 731-743.
- Mizutani, Y.; Kihara, A.; Igarashi, Y. LASS3 (longevity assurance homologue 3) is a mainly testis-specific (dihydro)ceramide synthase with relatively broad substrate specificity. Biochem. J. 2006, 398, 531–538. [Google Scholar] [CrossRef]
- Levy, M.; Futerman, A.H. Mammalian ceramide synthases. IUBMB Life 2010, 62, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, Y.; Kihara, A.; Chiba, H.; Tojo, H.; Igarashi, Y. 2-Hydroxy-ceramide synthesis by ceramide synthase family: enzymatic basis for the preference of FA chain length. J. Lipid Res. 2008, 49, 2356–2364. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.-F.; Tao, Z.; Yan, Z.-Q.; Yang, S.-L.; Gong, Y. Molecular cloning, characterisation and tissue-specific expression of human LAG3, a member of the novel Lag1 protein family. DNA Seq. 2003, 14, 79–86. [Google Scholar] [CrossRef]
- Eckl, K.-M.; Tidhar, R.; Thiele, H.; Oji, V.; Hausser, I.; Brodesser, S.; Preil, M.-L.; Önal-Akan, A.; Stock, F.; Müller, D.; et al. Impaired Epidermal Ceramide Synthesis Causes Autosomal Recessive Congenital Ichthyosis and Reveals the Importance of Ceramide Acyl Chain Length. J. Investig. Dermatol. 2013, 133, 2202–2211. [Google Scholar] [CrossRef]
- Radner, F.P. , et al., Mutations in CERS3 cause autosomal recessive congenital ichthyosis in humans. PLoS Genet, 2013. 9(6): p. e1003536.
- Marsching, C.; Rabionet, M.; Mathow, D.; Jennemann, R.; Kremser, C.; Porubsky, S.; Bolenz, C.; Willecke, K.; Gröne, H.-J.; Hopf, C.; et al. Renal sulfatides: sphingoid base-dependent localization and region-specific compensation of CerS2-dysfunction. J. Lipid Res. 2014, 55, 2354–2369. [Google Scholar] [CrossRef]
- Chen, J.; Li, X.; Ma, D.; Liu, T.; Tian, P.; Wu, C. [Corrigendum] Ceramide synthase-4 orchestrates the cell proliferation and tumor growth of liver cancer in vitro and in vivo through the nuclear factor-κB signaling pathway. Oncol. Lett. 2021, 21, 1–1. [Google Scholar] [CrossRef]
- Schiffmann, S.; Sandner, J.; Birod, K.; Wobst, I.; Angioni, C.; Ruckhäberle, E.; Kaufmann, M.; Ackermann, H.; Lötsch, J.; Schmidt, H.; et al. Ceramide synthases and ceramide levels are increased in breast cancer tissue. Carcinog. 2009, 30, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Ruckhäberle, E.; Rody, A.; Engels, K.; Gaetje, R.; von Minckwitz, G.; Schiffmann, S.; Grösch, S.; Geisslinger, G.; Holtrich, U.; Karn, T.; et al. Microarray analysis of altered sphingolipid metabolism reveals prognostic significance of sphingosine kinase 1 in breast cancer. Breast Cancer Res. Treat. 2007, 112, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Riebeling, C. , et al., Two mammalian longevity assurance gene (LAG1) family members, trh1 and trh4, regulate dihydroceramide synthesis using different fatty acyl-CoA donors. J Biol Chem, 2003. 278(44): p. 43452-9.
- Chen, L.; Chen, H.; Li, Y.; Li, L.; Qiu, Y.; Ren, J. Endocannabinoid and ceramide levels are altered in patients with colorectal cancer. Oncol. Rep. 2015, 34, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.W.; Park, W.; Min, H.; Kwon, T.K.; Baek, S.K.; Hwang, I.; Kim, S.; Park, J. Altered mRNA expression levels of the major components of sphingolipid metabolism, ceramide synthases and their clinical implication in colorectal cancer. Oncol. Rep. 2018, 40, 3489–3500. [Google Scholar] [CrossRef]
- Kijanka, G.; Hector, S.; Kay, E.W.; Murray, F.; Cummins, R.; Murphy, D.; MacCraith, B.D.; Prehn, J.H.M.; Kenny, D. Human IgG antibody profiles differentiate between symptomatic patients with and without colorectal cancer. Gut 2009, 59, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, S.; Sheehan, K.M.; Espina, V.; O'Grady, A.; Cummins, R.; Kenny, D.; Liotta, L.; O'Kennedy, R.; Kay, E.W.; Kijanka, G.S. High CerS5 expression levels associate with reduced patient survival and transition from apoptotic to autophagy signalling pathways in colorectal cancer. J. Pathol. Clin. Res. 2014, 1, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Li, F.; Han, Z.; Yuan, W.; Cao, L.; Deng, Y.; Peng, X.; Chen, F.; Fan, X.; Liu, X.; et al. LASS5 Interacts with SDHB and Synergistically Represses p53 and p21 Activity. Curr. Mol. Med. 2016, 16, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Schiffmann, S.; Ferreiros, N.; Birod, K.; Eberle, M.; Schreiber, Y.; Pfeilschifter, W.; Ziemann, U.; Pierre, S.; Scholich, K.; Grösch, S.; et al. Ceramide Synthase 6 Plays a Critical Role in the Development of Experimental Autoimmune Encephalomyelitis. J. Immunol. 2012, 188, 5723–5733. [Google Scholar] [CrossRef]
- Erez-Roman, R.; Pienik, R.; Futerman, A.H. Increased ceramide synthase 2 and 6 mRNA levels in breast cancer tissues and correlation with sphingosine kinase expression. Biochem. Biophys. Res. Commun. 2010, 391, 219–223. [Google Scholar] [CrossRef]
- Makoukji, J.; Raad, M.; Genadry, K.; El-Sitt, S.; Makhoul, N.J.; Aldin, E.S.; Nohra, E.; Jabbour, M.; Sangaralingam, A.; Chelala, C.; et al. Association between CLN3 (Neuronal Ceroid Lipofuscinosis, CLN3 Type) Gene Expression and Clinical Characteristics of Breast Cancer Patients. Front. Oncol. 2015, 5, 215. [Google Scholar] [CrossRef]
- Ruckhäberle, E.; Holtrich, U.; Engels, K.; Hanker, L.; Gätje, R.; Metzler, D.; Karn, T.; Kaufmann, M.; Rody, A. Acid ceramidase 1 expression correlates with a better prognosis in ER-positive breast cancer. Climacteric 2009, 12, 502–513. [Google Scholar] [CrossRef]
- Uen, Y.-H. , et al., Ceramide synthase 6 predicts the prognosis of human gastric cancer: It functions as an oncoprotein by dysregulating the SOCS2/JAK2/STAT3 pathway. Molecular Carcinogenesis, 2018. 57(12): p. 1675-1689.
- Minamoto, S.; Ikegame, K.; Ueno, K.; Narazaki, M.; Naka, T.; Yamamoto, H.; Matsumoto, T.; Saito, H.; Hosoe, S.; Kishimoto, T. Cloning and Functional Analysis of New Members of STAT Induced STAT Inhibitor (SSI) Family: SSI-2 and SSI-3. Biochem. Biophys. Res. Commun. 1997, 237, 79–83. [Google Scholar] [CrossRef]
Figure 1.
Ceramide biosynthesis and metabolism pathway.
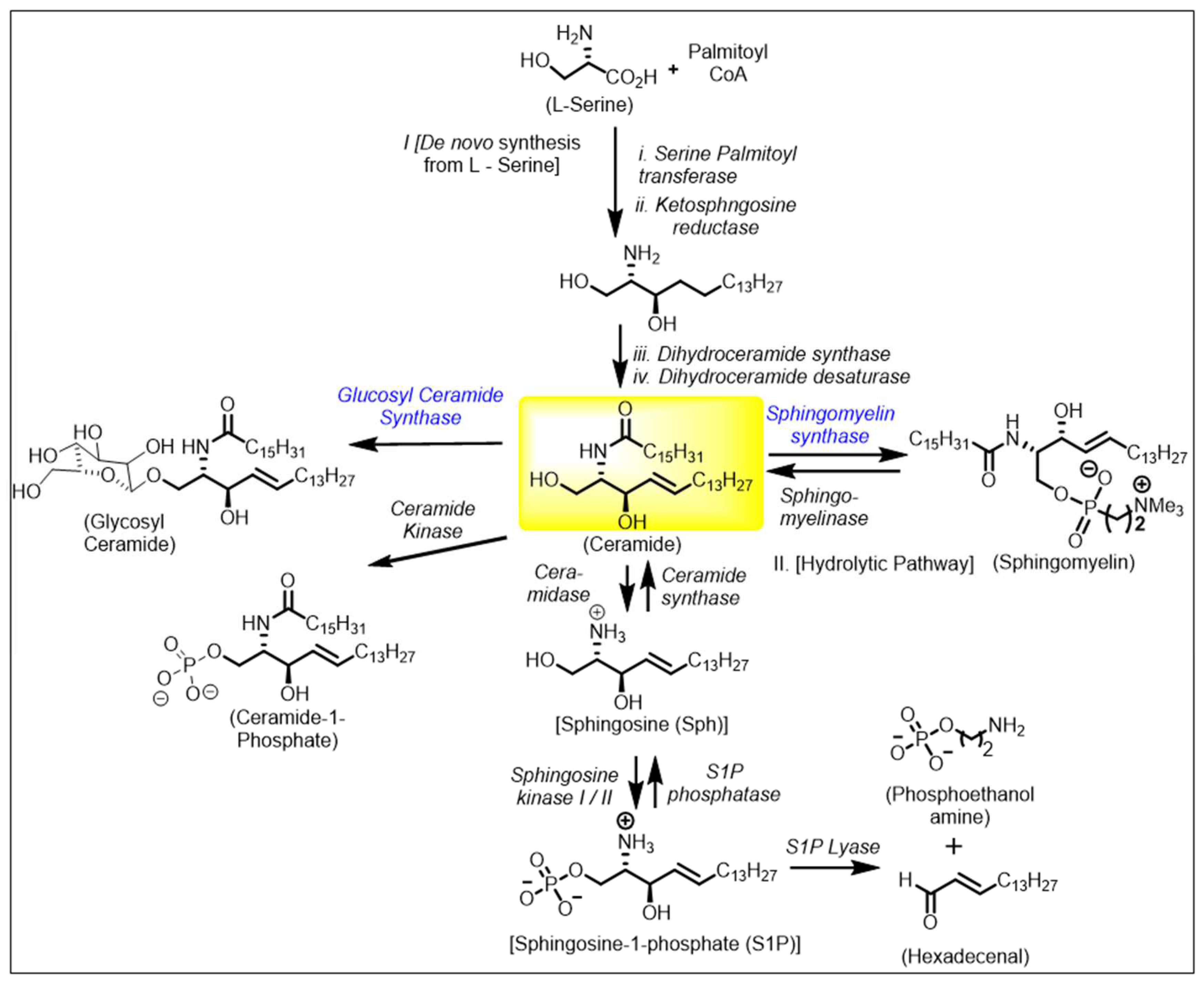
Figure 2.
3D crystal structure of human acid ceramidase (ASAH1, aCDase).
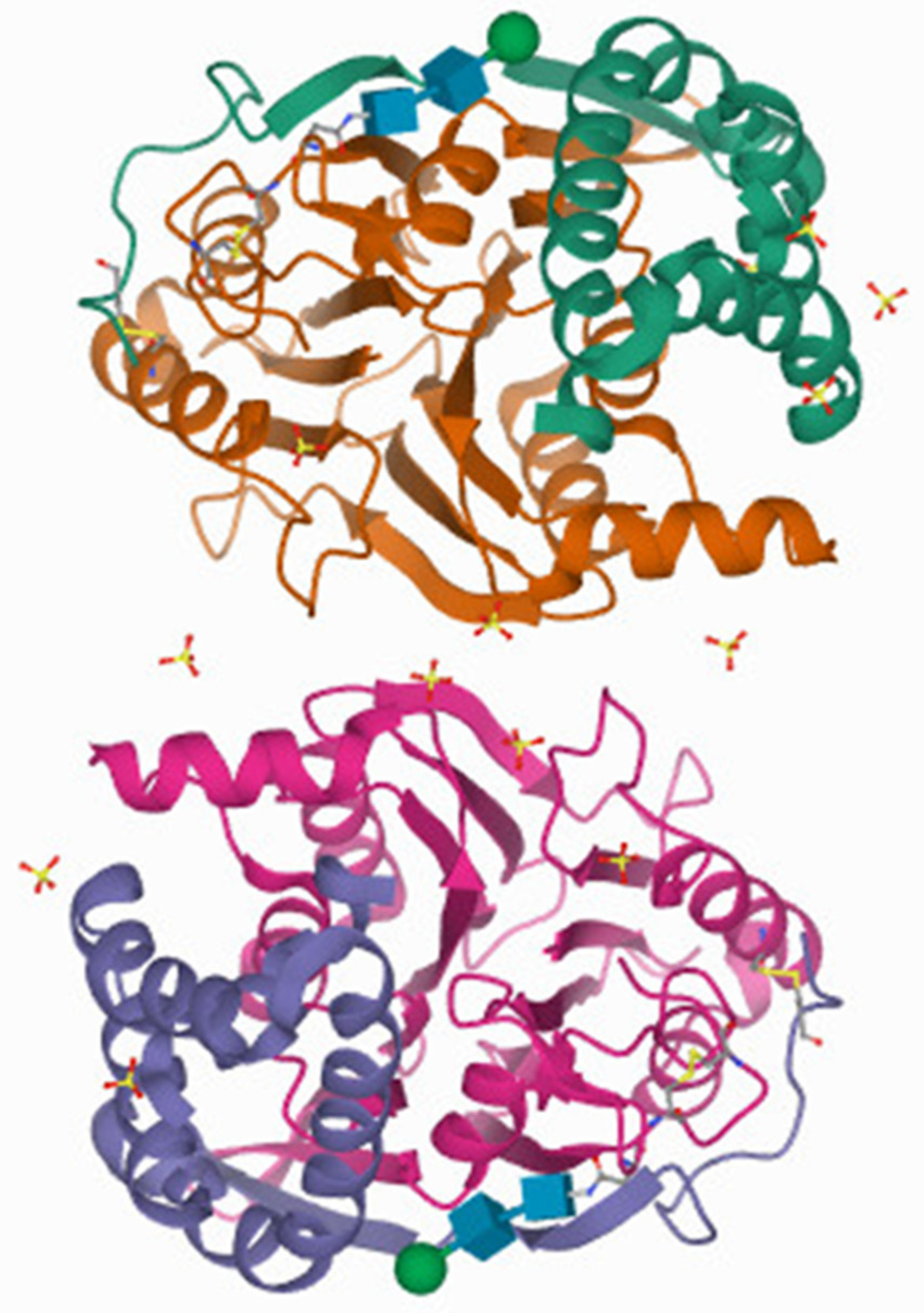
Figure 3.
Examples of AC inhibitors.
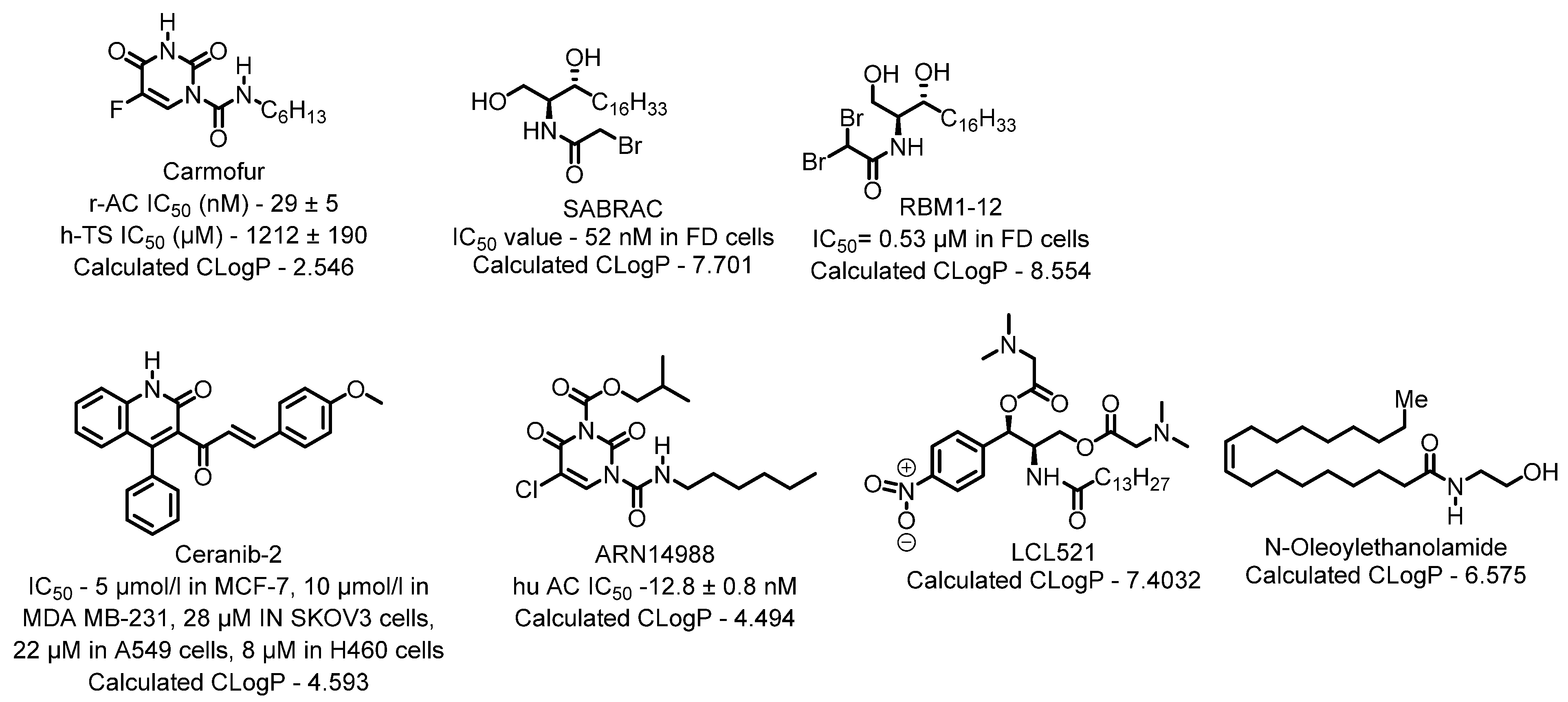
Figure 4.
Crystal structure of Sphingosine Kinase 1.
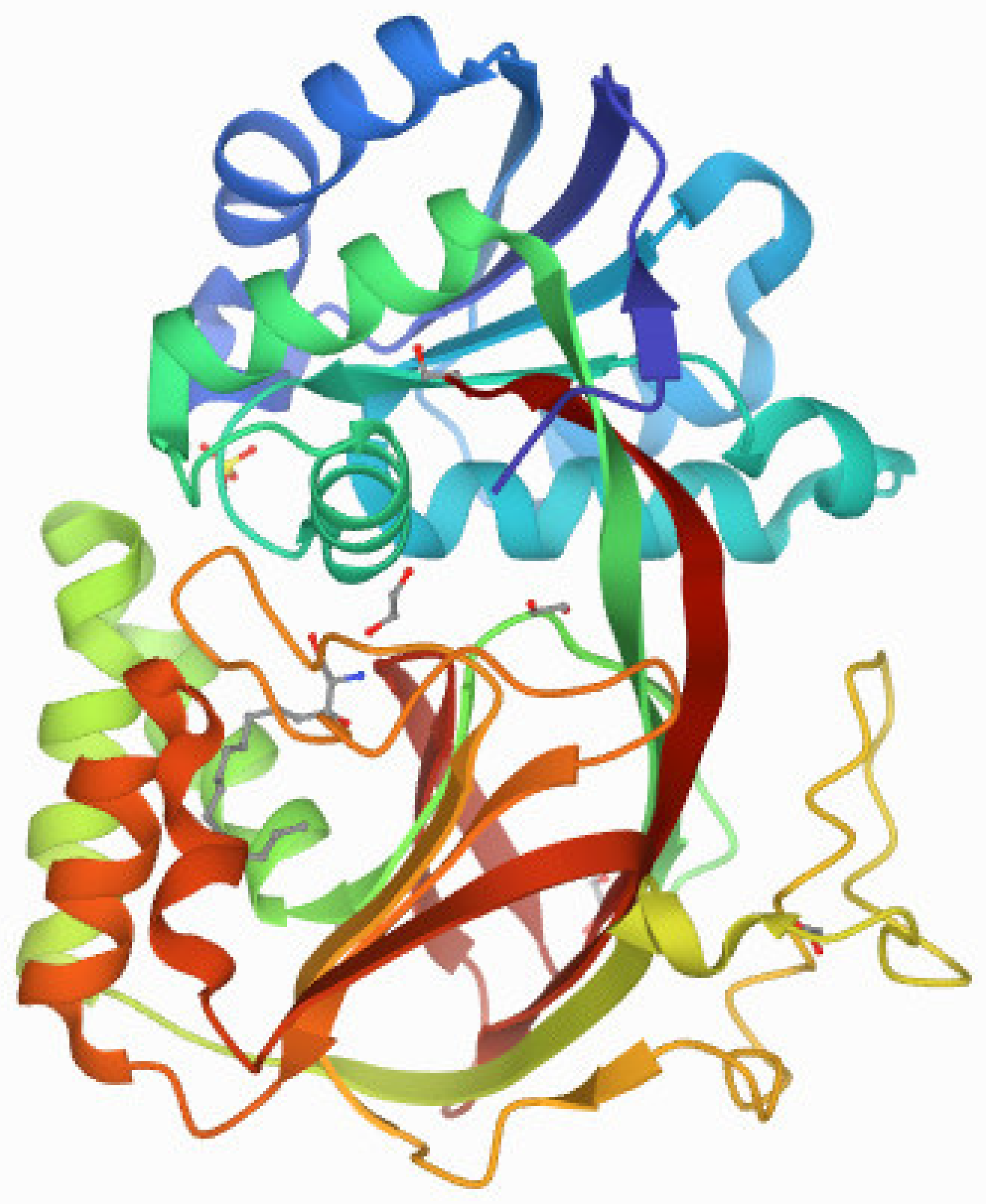
Figure 5.
Sphingosine Kinase 2 inhibitors.
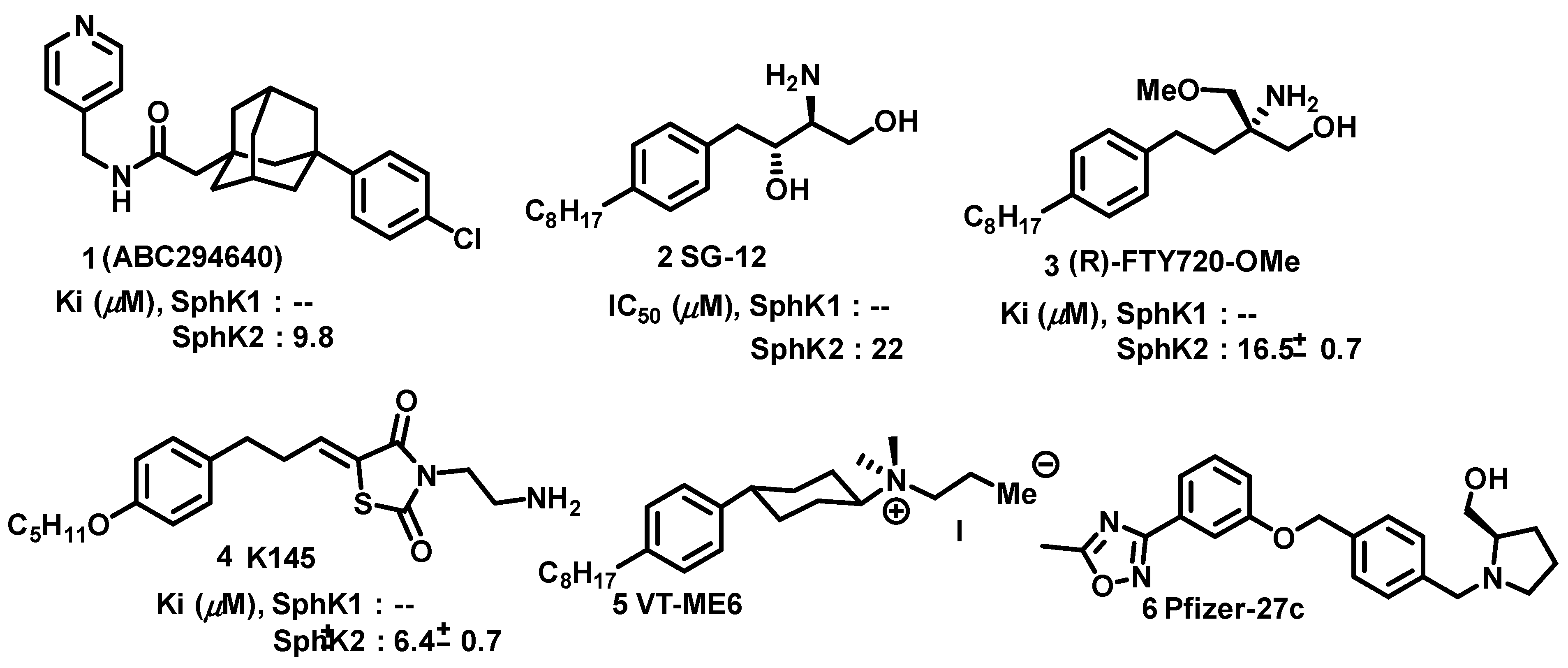
Figure 6.
Other Sphingosine Kinase Inhibitors.
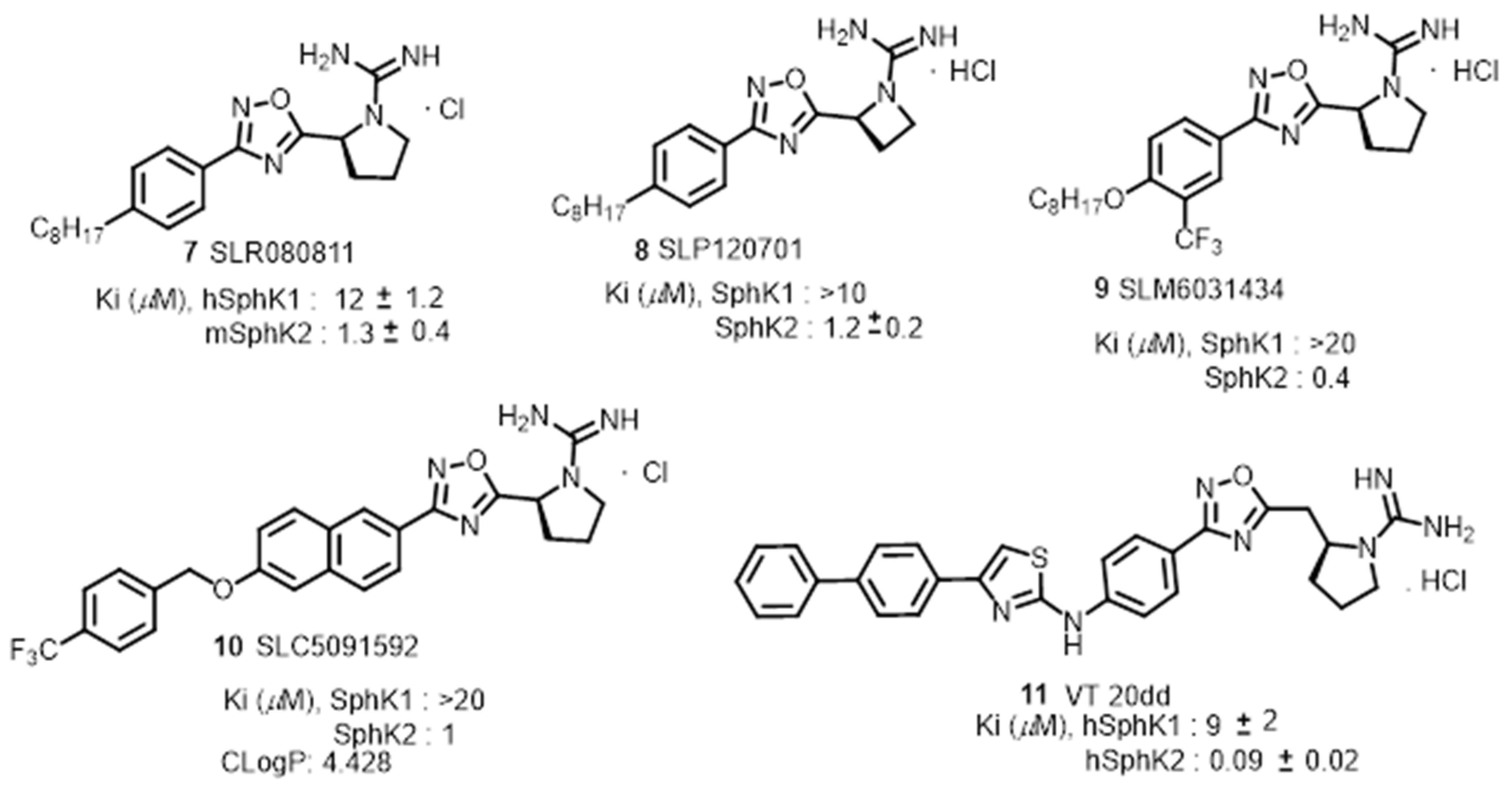
Figure 7.
Sphingomyelin Synthase Inhibitors.
Figure 8.
Chemical structure of non-sphingolipid analogs reported as DES1 inhibitors.
Figure 9.
Chemical structure of sphingolipid analogs reported as DES1 inhibitors.
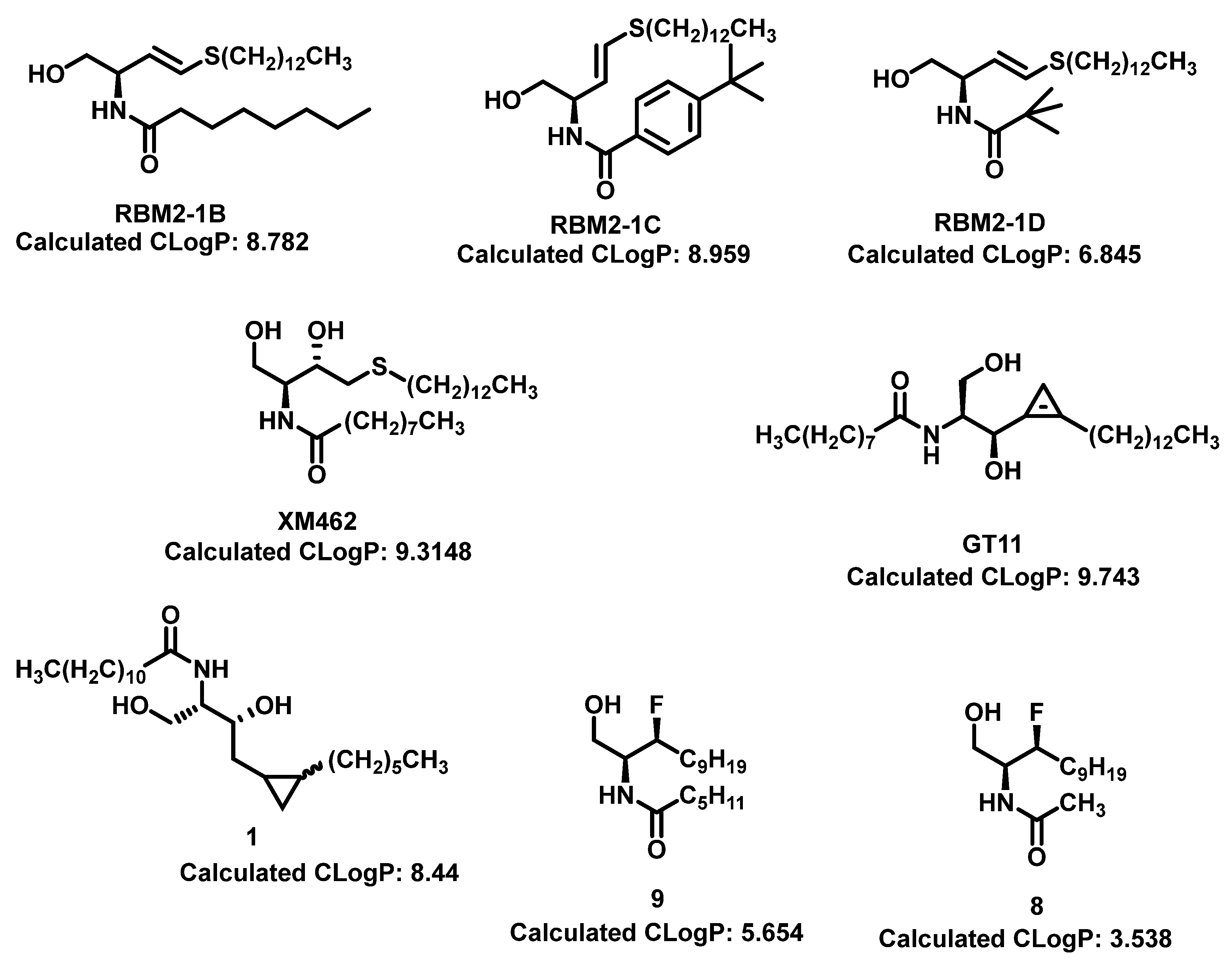
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



