
Submitted:
21 July 2023
Posted:
24 July 2023
You are already at the latest version
Abstract
Russia is one of the largest cereal grain exporters in the world, churning out 34,3 million tons of export grain (wheat, barley and oats) in 2021. Plant infectious pathogens continue to be among the main factors in yield loss in the field and are a danger to the grain exporting industry's ability to expand internationally. This is primarily due to phytosanitary restrictions imposed by nations that monitor the presence and absence of certain phytopathogens in imported goods. Phytosanitary measures prevent the spread of plant pathogens, thus cutting the cost of dealing with them, once the pathogens invade new agricultural regions. This paper is devoted to the detection and identification of bacteria in samples of grain crops of three regions in The Republic of Crimea. The objects of the study were bacterial isolates from plant samples particularly wheat, oats, barley and triticale. The study was conducted in 2021. The identification of the isolates was carried out by sequencing a section 16–23S of the rRNA amplified by PCR with 8UA/519B, 27f/907r and PSf/PSr primers. Nucleotide sequences were deciphered using the Bio Edit program and compared with sequences placed in GenBank (https://blast.ncbi.nlm.nih. gov). The result of identification was considered an organism with maximum similarity. As a result, 38 samples of grain crops were collected, 95 bacterial colonies were isolated, of which 68 were identified to genus level and 22 were identified to species level. Some of the phytopathogens identified include: Agrococcus jenensis, Pseudomonas sp. and Curtobacterium sp. Some of the bacteria identified are beneficial like Ochrobactrum sp. Erwinia sp. and Pantoea sp. had a frequency of 28.95%, with Pantoea agglomerans having a frequency of 18.42%. Ochrobactrum sp. had a frequency of 10.53%. Enterococcus mundtii an frequency of 5.26%. Information about the species composition of bacteria on grain crops can be used to determine the spread of bacteria and their diagnosis and for bioinformatic analysis of genomes in search of species-specific genetic markers.
Keywords:
Grain exports
; primers
; phyto-pathogens
; identification
; bacteria
; wheat
; barley
; oats
; Gene bank
1. Introduction
2020 World bank data indicated that Russia’s value-added agriculture expanded from $45.9 billion in 2000 to $66.2 billion in 2019 thus indicating a vast improvement. Grain production has increased significantly in the past years 2010 – 2019 [1]. This has allowed for improvement in the export value of grain from $10.6 billion in 2019 to $13 billion in 2021. Russia is currently one of the leading nations in grain export, trading about 34 million tons of grain (wheat, barley and maize) in 2021 alone, thus occupying a position of immense importance on the global market [2,3]. Grain export generates a lot of money and in 2021 alone the nation made 8 962 670 000 USD [2]. To date the main importers of Russian grain are Egypt and Turkey [4]. China currently accounts for 15% of the flour (73 thousand tons in 2020 and in 2021) exported by Russia [2]. Exportation of grain might be limited and restricted by the presence of unwanted or quarantine pests and pathogens.
All grain exports have to meet certain criteria, specified by the importing country and these have to be highlighted in a phytosanitary certificate by the exporting nation [3]. Phytosanitary requirements have to be observed to prevent the spread of diseases across nations and continents. In grain crops there is regulation of Pseudomonas syringae and Pseudomonas fuscovaginae. Quarantine bacterial pathogens of wheat include members of the genera Pseudomonas and Xanthomonas for example P. syringae and X. transluscens [5]. With these phytopathogens (Xanthomonas translucens, Pseudomonas syringae and Pseudomonas fuscovaginae) in mind and also given that the Republic of Crimea is a grain exporting region, from there grain samples were collected so as to detect and identify entire bacterial species within the plant samples, thus obtaining an estimation of their microbiome composition. Plant samples were collected, from them extracts were isolated and colonies were cultured so as to identify and detect various bacterial species within the samples and an estimate of the cultured bacterial composition of the plant samples was obtained. Bacteria have significant effects on plant growth, development, yield, health and nutrition thus an estimation of the microbiome composition of grain crops will help with the insight to all cultivated bacteria that can be found in grain crops [6]. The microbiome composition of crops varies depending on the crop thus the collection of different grain crops like barley and oats and not just wheat will help attain more comprehensive information on possible bacteria that make up a microbiome composition of grain crops. The species that can be present in the examined sample and from which the target species should be separated must all be known for bioinformatic prediction of a species-specific PCR target. PCR in conjunction with sequencing are widely used for identifying bacteria [7]. Prevention is better than cure and as such it is more important to prevent the spread of bacteria than having to try and control the damage caused to yield once they infest new areas. Hence, emphasis is stressed on the need to improve and develop efficient methods of bacterial detection [8]. The objective of this study were detection and identification of the cultivated bacteria in the samples collected from Belogorskij, Krasnogvardejskij and Simferopol‘skij regions of the Republic of Crimea.
2. Materials and Methods
The objects of the study were bacterial isolates from samples of oats, wheat, triticale and barley plants. Sampling was carried out from the 1st to the 3rd of June 2021 from three regions (Belogorskij, Krasnogvardejskij and Simferopol‘skij) of the Republic of Crimea. The plants upon sampling were in the initial phase of grain ripening known as the milky ripening stage. Analysis of the samples were conducted at the All-Russian Plant Quarantine Center. An analytical sample was prepared from each sample in accordance with the previously described technique [9] in the following way. To 5–10g of plant tissues chopped with sterilized scissors, 20 ml of phosphate-buffered saline was added (per 1 liter of distilled water 2.9g Na2HPO4*12H2O, 0.2g KH2PO4*2H2O, 8g NaCl and 0.2g KCl; pH 7.0–7.2), left on the shaker for 1 hour at 200 rpm, then liquid part of the sample was passed through filters with a pore size of 3–5 μl and centrifuged for 10 min at 4°C, 10000g. The supernatant was removed and the precipitate was suspended in 1 ml of phosphate-buffered saline. Sample preparation was carried out within 1 week after sample collection. Before preparation samples were stored at 4°C in the dark. Bacterial isolates were isolated on Yeast extract-dextrose-CaCO3 (YDC) [10] nutrient medium, for 5 to 7 days plated on three Petri dishes by the Drygalsky method, 20 µl of analytical samples. Using a sterile bacteriological loop, individual colonies were inoculated onto YDC nutrient medium. Some of them were not selected because they had morphological similarities to the colonies already selected. Different varieties of colony morphotypes grown on plates were selected. Small fragments of single colonies of the pure cultures thus obtained were collected using a sterile bacteriological loop and suspended in 200 μl of distilled water. The suspensions were used for DNA isolation. DNA of bacterial cultures was obtained using a commercial Proba-GS kit (ZAO Agro-Diagnostics, Russia) in accordance with the manufacturer’s instructions.
All DNA samples were tested by conventional PCR. Amplification was carried out on a T100 thermal cycler (Bio-Rad, United States) using oligonucleotides synthesized at CJSC Evrogen (Russia) and ready-made mixtures for PCR 5½ MasDDTaqMIX-2025 (CJSC "Dialat", Russia). The first test was performed with primers 8UA/519B (8UA: 5’-AGAGTTTGATCMTGGCTCAG-3’, 519B: 5’-GTATTACCGCGGCKGCTG-3’) for the 16–23S rRNA region [11]. The PCR mixture per reaction contained 14 µl of water, 5 µl of 5x MasDDTaqMIX-2025, 2 µl of each primer at a concentration of 10 µmol, and 2 µl of DNA. Amplification: 96 °C - 10 min; then 35 cycles: 95°C - 15 s, 55°C - 30 s, 72°C - 30 s; 72 °C -10 min. The presence of the PCR product was checked using horizontal electrophoresis in 1.5% agarose gel. Amplicon residues not used for electrophoresis were subjected to purification and sequencing as described above. DNA samples for which PCR products with primers 8UA/519B were of low concentration for sequencing or those whose results in Blasts did not give exact identification were subjected to PCR with primers 27F/907R (27F: 5’-AGAGTTTGATYMTGGCTCAG-3’, 907R: 5’-CCGTCAATTCMTTTGAGTTT-3’) for the 16–23S rRNA region [12]. PCR mixture per reaction contained 16 µl water, 5 µl of 5x MasDDTaqMIX-2025, 1 µl of each primer at a concentration of 10 µmol and 2 µl of DNA. Amplification program: 95 °C - 5 min; then 35 cycles: 95 °C - 15 s, 58 °C - 30 s, 72 °C - 60 s; then 72 °C - 5 minutes. In the absence of 880 bp PCR product further PCR was done using primers PSF/PSR (PSF: 5’-AGCCGTAGGGGAACCTGCGG-3’, PSR: 5’-TGACTGCCAAGGCATCCACC-3’) [13]. Several copies of the 610 bp sequence, amplifiable with the indicated primers are located in tRNA in bacteria of the genus Pseudomonas. PCR mixture per reaction contained 16 µl water, 5 µl of 5x MasDDTaqMIX-2025, 1 µl of each primer at a concentration of 10 µmol and 2 µl of DNA. Amplification program: 95 °C - 10 min; then 25 cycles: 95 °C - 20 s, 64 °C - 15 s, 72 °C - 15 s; then 72 °C - 2 minutes. The presence of the PCR product was checked using a gel-documenting system (Bio-Rad, USA) after electrophoretic separation of PCR products in 1.5% agarose gel. Thus, members of the genus Pseudomonas were identified amongst the isolates. The amplicons remaining in the tube were purified using the kit Gene JET PCR Purification Kit (Thermo Fisher Scientific, USA) and used for Sanger sequencing using the Big Dye Kit, BigDye®XTerminator™ Purification Kit (Thermo Fisher Scientific, USA) on genetic analyzer AB-3500 (Applied Biosystems, USA) according to an adapted method [14]. The presence of the PCR product was checked using horizontal electrophoresis in 1.5% agarose gel. Amplicon residues not used for electrophoresis were subjected to purification and sequencing as described above.
The sequencing results were processed using the BioEdit program (https:// bioedit.software.informer.com/). The decoded nucleotide sequences were compared using the BLAST service with the sequences available in GenBank (https:// blast.ncbi.nlm.nih.gov). The result of identification was considered the organism with the maximum similarity. (Max score), automatically calculated by the BLAST service based on the calculation of the Query coverage and Percent identity indicators. If several such organisms were found in a taxon, the oldest taxon was considered as the result of the identification process. For each identified species and genus, the frequency of occurrence (A) was calculated using the formula [15]: A = B/C*100%, where B is the number of samples on which a bacterium with a certain species was found, C is the total number of analyzed samples. When calculating the frequency of occurrence of bacterial genera, both isolates identified to the species level and those identified only at the genus level were taken into account.
3. Results
In three regions (Belogorskij, Krasnogvardejskij and Simferopol‘skij) of the Republic of Crimea, 38 samples of grain crops were collected including winter barley, winter wheat, oats, triticale and cereal legume mix as shown in Table 1. The sampling period for winter grain crops was in the milky ripening phase. There were no symptoms of bacterial diseases during sampling of winter crops on plants. A total of 38 samples of grain crops were taken (Table 1).

Table 1.
The results of samples collected of grain crops from Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea.
Table 1.
The results of samples collected of grain crops from Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea.
| Crop | Cultivar | Number of samples |
|---|---|---|
| Winter Wheat | Aksiniya Gubernator Dona Asket Anka Karavan Korona |
3 6 7 1 1 1 |
| Winter Barley | Onega Vosxod Rubezh Espada Master Toma |
5 3 2 1 1 1 |
| Oats | Verny‘j Skakun Podgorny‘j - |
1 1 1 1 |
| Cereal mix (Triticale; wheat and barley) | - | 1 |
| Cereal-legume mix | - | 1 |
N o t e. -means the cultivar is unknown. Amongst the samples collected 13 samples of Winter barley,19 samples of Winter Wheat, 4 samples of Oats, 1 sample of Cereal mix (Triticale; wheat and barley) and 1 sample of Cereal-legume mix.
The three regions (Belogorskij, Krasnogvardejskij and Simferopol‘skij) of the Republic of Crimea, 38 samples of grain crops were collected including winter barley, winter wheat, oats, triticale and cereal legume shown in the maps below:
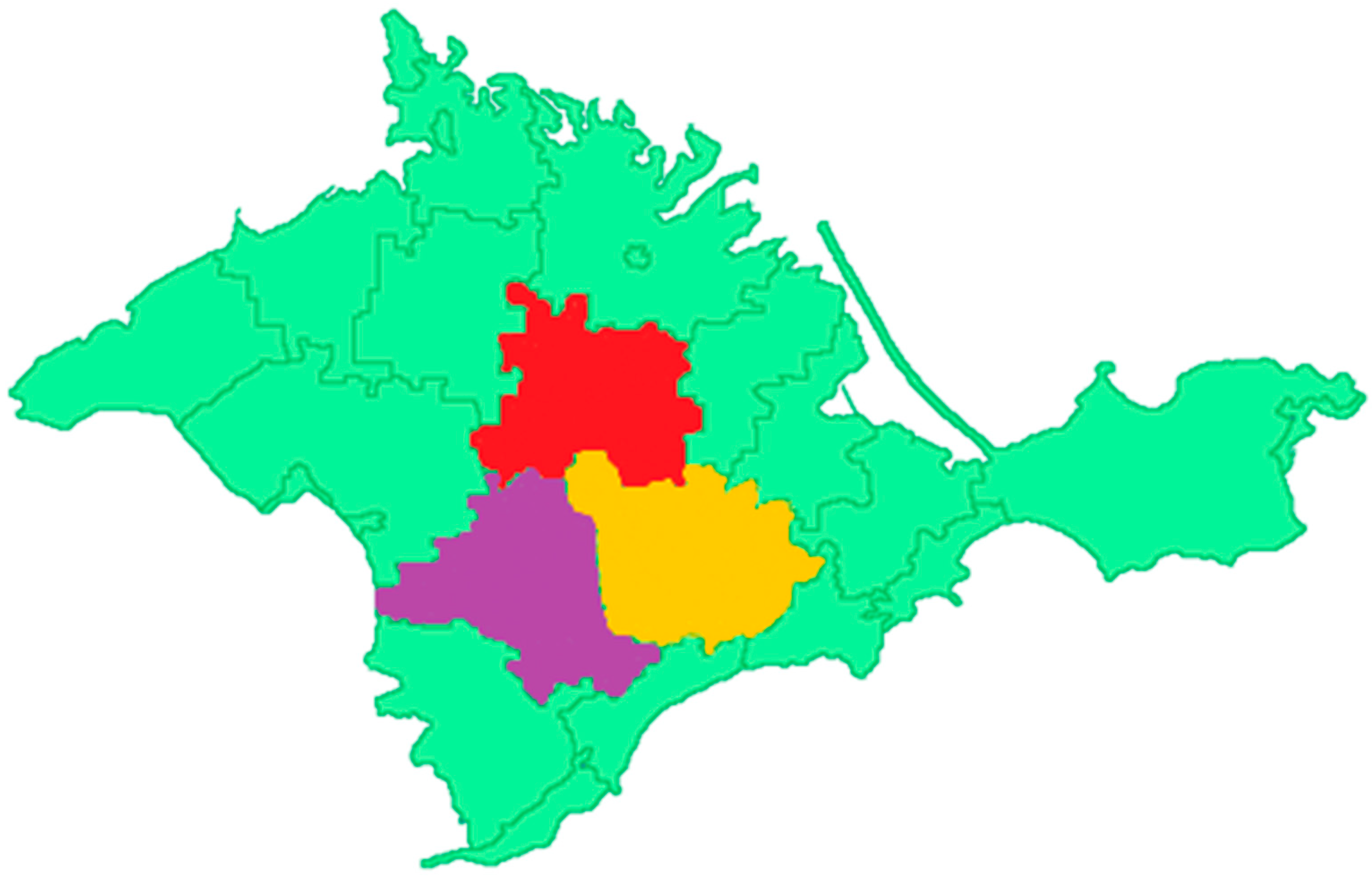
Figure 1.
A map of The Republic of Crimea, in orange- Belogorskij from which samples C1 -C19 were collected, in red- Krasnogvardejskij from which samples C20, C21. C24, C26, , C28, C33, C37, C38, C41, C42, C45, C46, C48, C51 and C53 were collected and in purple- Simferopol‘skij from which samples C54, C58, C59 and C60 where C refers to Crimea and the sample number collected.
Figure 1.
A map of The Republic of Crimea, in orange- Belogorskij from which samples C1 -C19 were collected, in red- Krasnogvardejskij from which samples C20, C21. C24, C26, , C28, C33, C37, C38, C41, C42, C45, C46, C48, C51 and C53 were collected and in purple- Simferopol‘skij from which samples C54, C58, C59 and C60 where C refers to Crimea and the sample number collected.
3.1. Bacteria identification
3 primer pairs used in identification of the bacteria, 8UA/519B, 27F/907R and PSF/PSR.
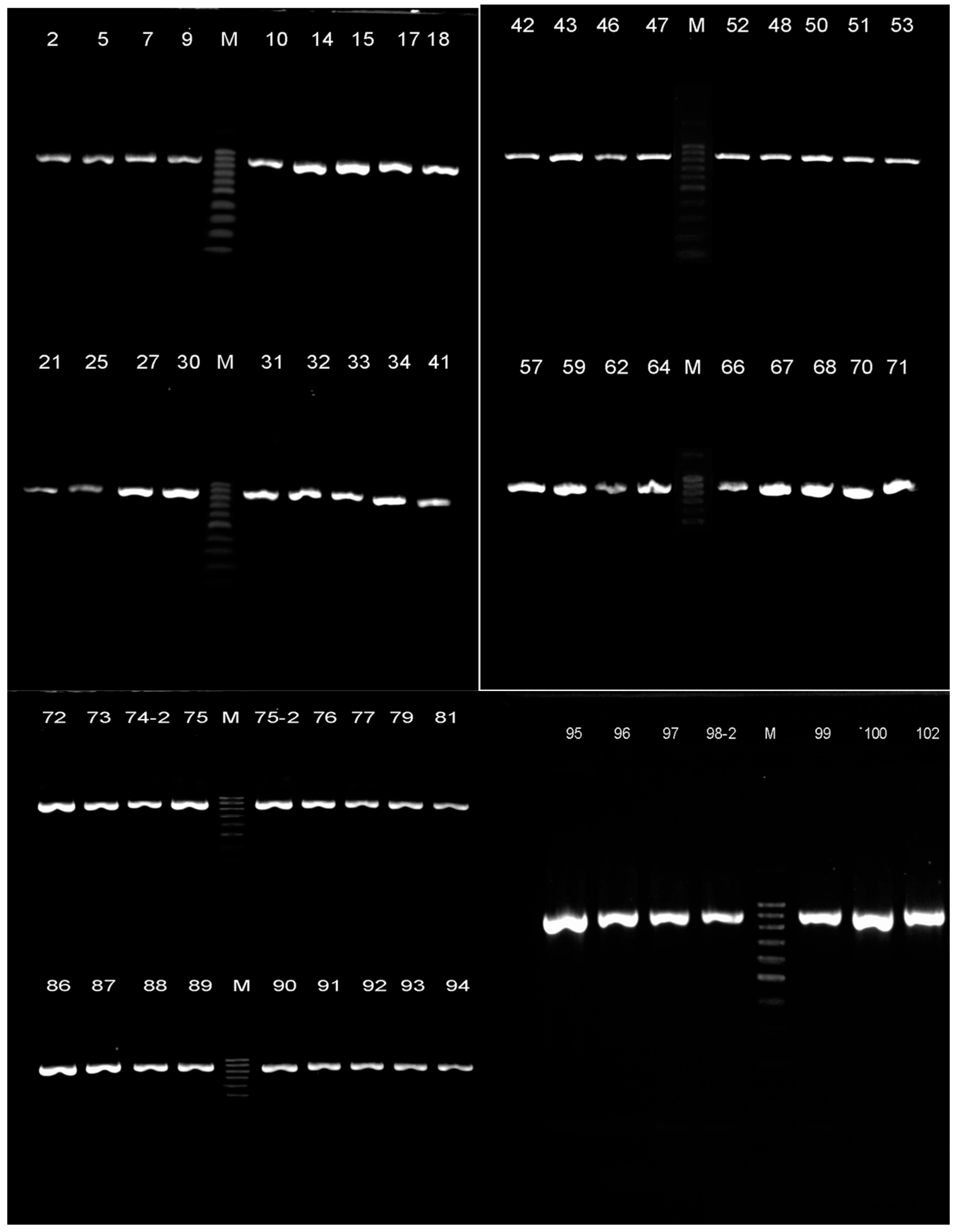
As a result of PCR with 8UA/519B primers, the 500 bp amplicon was obtained for 92 tested samples of DNA from bacterial cultures (Figure 2).
Figure 2.
PCR products with primers 8UA/519B (500 bp) obtained for DNA samples of bacterial isolates from cereal varieties:1-5 7-9 Winter Barley Onega, 10-12 Winter wheat Aksiniya, 14-15, 17 Winter wheat Asket, 18-22, 24 Winter barley Vosxod, 25-30 Winter wheat Gubernator Dona, 31-35 Winter barley Vosxod, 36, 38, 39 Winter wheat Asket, 41-42 Winter barley Onega, 43, 45-53, 55, 57-60 Winter wheat Gubernator Dona, 61-64 Winter wheat Asket, 66-75-1 Winter barley Onega, 75-2-79, 81-82 Winter wheat Asket, 83-85 Winter barley Rubezh, 86 Winter barley Espada, 87 Winter wheat Anka, 88 Winter wheat Karavan, 89, 90 Oats Verny‘j, 91, 92 Oats Skakun, 93, 94 Oats Podgorny‘j, 95-96 Winter barley Master, 97 Winter barley Rubezh, 98.1, 98.2 Winter barley Toma, 99 Triticale, 100 Cereal legume, and 102Winter wheat Korona; M Molecular weight marker GeneRuler 100 bp Plus (100-1000 bp), ("Thermo Fisher Scientific, USA) (Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
Figure 2.
PCR products with primers 8UA/519B (500 bp) obtained for DNA samples of bacterial isolates from cereal varieties:1-5 7-9 Winter Barley Onega, 10-12 Winter wheat Aksiniya, 14-15, 17 Winter wheat Asket, 18-22, 24 Winter barley Vosxod, 25-30 Winter wheat Gubernator Dona, 31-35 Winter barley Vosxod, 36, 38, 39 Winter wheat Asket, 41-42 Winter barley Onega, 43, 45-53, 55, 57-60 Winter wheat Gubernator Dona, 61-64 Winter wheat Asket, 66-75-1 Winter barley Onega, 75-2-79, 81-82 Winter wheat Asket, 83-85 Winter barley Rubezh, 86 Winter barley Espada, 87 Winter wheat Anka, 88 Winter wheat Karavan, 89, 90 Oats Verny‘j, 91, 92 Oats Skakun, 93, 94 Oats Podgorny‘j, 95-96 Winter barley Master, 97 Winter barley Rubezh, 98.1, 98.2 Winter barley Toma, 99 Triticale, 100 Cereal legume, and 102Winter wheat Korona; M Molecular weight marker GeneRuler 100 bp Plus (100-1000 bp), ("Thermo Fisher Scientific, USA) (Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).

As a result of PCR with 27F/907R primers, the 880 bp amplicon was obtained for 61 tested samples of DNA from bacterial cultures (Figure 2)
Figure 3.
PCR products with primers 27F/907R (880 bp) obtained for DNA samples of bacterial isolates from cereal varieties:2 5 7 9 Winter Barley Onega, 10 Winter wheat Aksiniya, 14,15, 17 Winter wheat Asket, 18 25 Winter barley Vosxod, 27, 30 Winter wheat Gubernator Dona, 31-34 Winter barley Vosxod, 41 42 Winter barley Onega, 43, 46-48, 50, 51, 53, 57,59 Winter wheat Gubernator Dona, 62, 64 Winter wheat Asket, 66-68, 70-73, 74-2, 75-1 Winter barley Onega, 75-2, 76,77, 79, 81 Winter wheat Asket, 86 Winter barley Espada, 87 Winter wheat Anka, 88 Winter wheat Karavan, 89, 90 Oats Verny‘j, 91, 92 Oats Skakun, 93, 94 Oats Podgorny‘j, 95-96 Winter barley Master, 97 Winter barley Rubezh, 98.2 Winter barley Toma, 99 Triticale, 100 Cereal legume, and 102Winter wheat Korona; M Molecular weight marker GeneRuler 100 bp Plus (100-1000 bp), ("Thermo Fisher Scientific, USA) (Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
Figure 3.
PCR products with primers 27F/907R (880 bp) obtained for DNA samples of bacterial isolates from cereal varieties:2 5 7 9 Winter Barley Onega, 10 Winter wheat Aksiniya, 14,15, 17 Winter wheat Asket, 18 25 Winter barley Vosxod, 27, 30 Winter wheat Gubernator Dona, 31-34 Winter barley Vosxod, 41 42 Winter barley Onega, 43, 46-48, 50, 51, 53, 57,59 Winter wheat Gubernator Dona, 62, 64 Winter wheat Asket, 66-68, 70-73, 74-2, 75-1 Winter barley Onega, 75-2, 76,77, 79, 81 Winter wheat Asket, 86 Winter barley Espada, 87 Winter wheat Anka, 88 Winter wheat Karavan, 89, 90 Oats Verny‘j, 91, 92 Oats Skakun, 93, 94 Oats Podgorny‘j, 95-96 Winter barley Master, 97 Winter barley Rubezh, 98.2 Winter barley Toma, 99 Triticale, 100 Cereal legume, and 102Winter wheat Korona; M Molecular weight marker GeneRuler 100 bp Plus (100-1000 bp), ("Thermo Fisher Scientific, USA) (Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
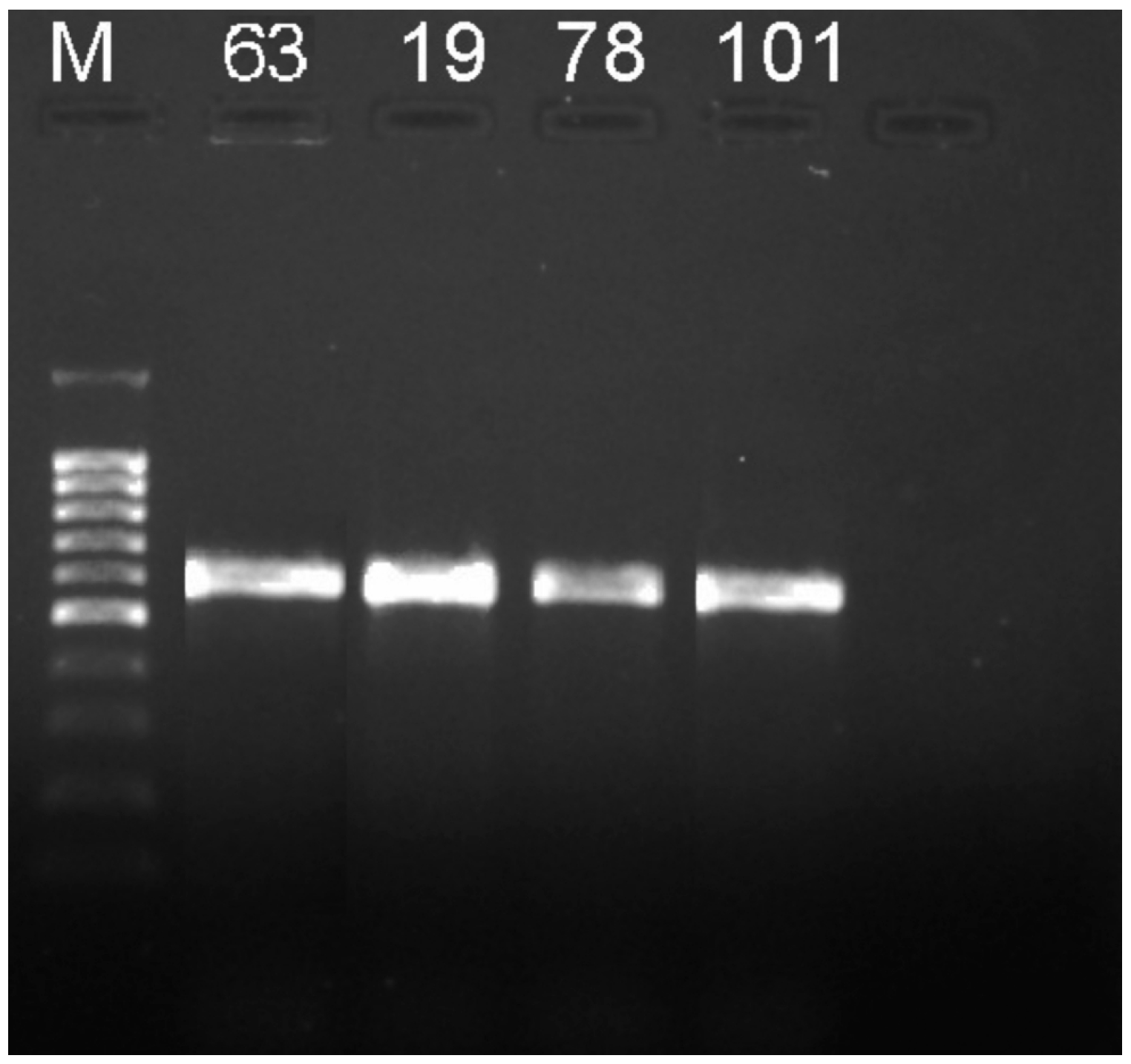
As a result of PCR with PSF/PSR primers, the 610 bp amplicon was obtained for 4 tested samples of DNA from bacterial cultures (Figure 4).
Figure 4.
PCR products with primers PSF/PSR (610 bp) obtained for DNA samples of bacterial isolates from cereal varieties:19 Winter barley Vosxod, 63 Winter wheat Asket, 78 Winter wheat Asket, 101 Oat (no name); M Molecular weight marker GeneRuler 100 bp Plus (100-1000 bp), ("Thermo Fisher Scientific, USA) (Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
Figure 4.
PCR products with primers PSF/PSR (610 bp) obtained for DNA samples of bacterial isolates from cereal varieties:19 Winter barley Vosxod, 63 Winter wheat Asket, 78 Winter wheat Asket, 101 Oat (no name); M Molecular weight marker GeneRuler 100 bp Plus (100-1000 bp), ("Thermo Fisher Scientific, USA) (Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
Purification, sequencing and processing, using the BioEdit program made it possible to extract nucleotide sequences for each of the selected isolates and align these sequences in the BLAST service (https://blast.ncbi.nlm.nih.gov). These methods also made it possible for the identification of Pantoea ananatis in a winter wheat sample which is well known for forming brownish lesions with clear margins and yellow halos on wheat leaves causing bacterial disease [16]. Table 2 provides examples of the results achieved.
Table 2.
The results of alignment of nucleotide sequences of bacterial isolates obtained from Wheat (Triticum aestivum) by Sanger sequencing (BLAST service, https://blast.ncbi.nlm.nih.gov; Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
Table 2.
The results of alignment of nucleotide sequences of bacterial isolates obtained from Wheat (Triticum aestivum) by Sanger sequencing (BLAST service, https://blast.ncbi.nlm.nih.gov; Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
| Crop | Cultivar | Isolate № | Primers | Result of identification |
|---|---|---|---|---|
| Winter wheat | Aksiniya | 10 | 27f-907r | Frigoribacterium sp. |
| 11 | 8UA-519B | Pantoea sp. | ||
| 12 | 8UA-519B | Pantoea sp. | ||
| Winter wheat | Aksiniya | 14 | 27f-907r | Ochrobactrum sp. |
| 15 | 27f-907r | Frigoribacterium sp. | ||
| Winter wheat | Aksiniya | 17 | 27f-907r | Pantoea agglomerans |
| Winter wheat | Gubernator Dona | 25 | 27f-907r | Arthrobacter sp. |
| 26 | 8UA-519B | Pantoea sp. | ||
| 27 | 27f-907r | Erwinia sp. | ||
| Winter wheat | Gubernator Dona | 28 | 27f-907r | Clavibacter michiganensis |
| 29 | 8UA-519B | Pantoea agglomerans | ||
| 30 | 27f-907r | Erwinia aphidicola | ||
| Winter wheat | Asket | 36 | 27f-907r | Pantoea sp. |
| 37 | 27f-907r | Erwinia sp. | ||
| 38 | 8UA-519B | Ochrobactrum sp. | ||
| 39 | 8UA-519B | Pantoea agglomerans | ||
| Winter wheat | Gubernator Dona | 43 | 27f-907r | Rosenbergiella sp. |
| 45 | 27f-907r | Pantoea ananatis | ||
| Winter wheat | Gubernator Dona | 46 | 27f-907r | Erwinia sp. |
| 47 | 27f-907r | Erwinia sp. | ||
| 48 | 27f-907r | Rosenbergiella sp. | ||
| 49 | 8UA-519B | Pantoea agglomerans | ||
| 50 | 27f-907r | Stenotrophomonas sp. | ||
| 51 | 27f-907r | Erwinia sp. | ||
| Winter wheat | Gubernator Dona | 52 | 27f-907r | Rosenbergiella sp. |
| 53 | 27f-907r | Erwinia sp. | ||
| Winter wheat | Gubernator Dona | 55 | 8UA-519B | Rosenbergiella sp. |
| 57 | 27f-907r | Erwinia sp. | ||
| 58 | 8UA-519B | Pantoea sp. | ||
| 59 | 27f-907r | Pantoea sp. | ||
| 60 | 8UA-519B | Pantoea sp. | ||
| Winter wheat | Asket | 61 | 8UA-519B | Uncultured bacterium |
| 62 | 27f-907r | Erwinia sp. | ||
| 63 | PSf- PSr | Pseudomonas sp. | ||
| 64 | 27f-907r | Exiguobacterium sp. | ||
| Winter wheat | Asket | 75-2 | 27f-907r | Pantoea sp. |
| Winter wheat | Asket | 76 | 27f-907r | Pantoea sp. |
| Winter wheat | Asket | 77 | 8UA-519B | Pantoea sp. |
| 78 | PSf- PSr | Pseudomonas sp. | ||
| Winter wheat | Asket | 79 | 27f-907r | Pantoea agglomerans |
| Winter wheat | Asket | 81 | 27f-907r | Plantibacter sp. |
| 82 | 27f-907r | Bacteria | ||
| Winter wheat | Anka | 87 | 27f-907 | Rosenbergiella sp. |
| Winter wheat | Karavan | 88 | 27f-907 | Rosenbergiella sp. |
| Winter wheat | Korona | 102 | 27f-907 | Pantoea agglomerans |
19 samples of Winter wheat were collected from which 45 colonies were isolated and identified with the use of 3 pairs of primers.
One sample of grain crops contained either a particular kind of detected bacterium, or a few detected bacteria (Table 3). Erwinia rhapontici was amongst some of the detected and identified bacterium in the samples of barley. E. rhapontici is known to cause premature death in infected plants [17]. Table 3 shows the list of the bacteria identified from the barley samples collected from The Republic of Crimea.
Table 3.
The results of alignment of nucleotide sequences of bacterial isolates obtained from Barley (Hordeum vulgare) by Sanger sequencing (BLAST service, https://blast.ncbi.nlm.nih.gov; Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
Table 3.
The results of alignment of nucleotide sequences of bacterial isolates obtained from Barley (Hordeum vulgare) by Sanger sequencing (BLAST service, https://blast.ncbi.nlm.nih.gov; Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
| Crop | Cultivar | Isolate № | Primers | Result of identification |
|---|---|---|---|---|
| Winter barley | Onega | 1 | 8UA-519B | Ochrobactrum sp. |
| 2 | 27f-907r | Erwinia aphidicola | ||
| 3 | 27f-907r | Rathayibacter festucae | ||
| 4 | 27f-907r | Arthrobacter sp. | ||
| 5 | 27f-907r | Arthrobacter sp. | ||
| 7 | 27f-907r | Stenotrophomonas maltophilia | ||
| 8 | 27f-907r | Erwinia sp. | ||
| 9 | 27f-907r | Bacteria | ||
| Winter barley | Vosxod | 18 | 27f-907r | Frigoribacterium sp. |
| 19 | PSf- PSr | Pseudomonas poae | ||
| 20 | 8UA-519B | Plantibacter flavus | ||
| 21 | 8UA-519B | Frigoribacterium sp. | ||
| 22 | 8UA-519B | Erwinia rhapontici | ||
| 24 | 8UA-519B | Erwinia rhapontici | ||
| Winter barley | Vosxod | 31 | 8UA-519B | Erwinia sp. |
| Winter barley | Vosxod | 32 | 27f-907r | Pantoea sp. |
| 33 | 27f-907r | Erwinia sp. | ||
| 34 | 27f-907r | Curtobacterium sp. | ||
| 35 | 27f-907r | Ochrobactrum sp. | ||
| Winter barley | Onega | 41 | 8UA-519B | Enterobacter sp. |
| 42 | 27f-907r | Uncultured bacterium | ||
| Winter barley | Onega | 66 | 27f-907r | Exiguobacterium sp. |
| 67 | 27f-907r | Stenotrophomonas sp. | ||
| 68 | 27f-907r | Stenotrophomonas sp. | ||
| Winter barley | Onega | 69 | 8UA-519B | Pseudomonas sp. |
| 70 | 27f-907r | Plantibacter sp. | ||
| 71 | 27f-907r | Stenotrophomonas sp. | ||
| 72 | 27f-907r | Stenotrophomonas sp. | ||
| 73 | 27f-907r | Agrococcus jenensis | ||
| Winter barley | Onega | 74-1 | 8UA-519B | Pantoea vagans |
| 74-2 | 27f-907r | Frigoribacterium sp. | ||
| 75-1 | 27f-907r | Rosenbergiella sp. | ||
| Winter barley | Rubezh | 83 | 27f-907r | Pantoea pleuroti |
| 84 | 27f-907 | Pantoea sp. | ||
| 85 | 8UA-519B | Microbacterium sp. | ||
| Winter barley | Espada | 86 | 27f-907 | Uncultured bacterium |
| Winter barley | Master | 95 | 27f-907 | Curtobacterium sp. |
| 96 | 27f-907 | Clavibacter michiganensis | ||
| Winter barley | Rubezh | 97 | 27f-907 | Microbacterium sp. |
| Winter barley | Toma | 98 | 8UA-519B | Enterococcus mundtii |
| 98-2 | 27f-907 | Frigoribacterium sp. |
13 samples of Winter barley were collected from which 41 colonies were isolated and identified with the use of 3 primers.
The bacteria identified in the study were either beneficial or harmful to grain plants and in some cases both. Pantoea agglomerans (Table 3) is known to cause bacterial blight in grain crops and yet some of its strains are beneficial to the rhizosphere of some plants [18,19,20]. The results of bacteria identified from oats, Triticale and cereal legume mix are shown in Table 3.
Table 3.
The results of alignment of nucleotide sequences of bacterial isolates obtained from Oats (Avena sativa) and 1 sample of Triticale and 1 sample of Cereal-legume by Sanger sequencing (BLAST service, https://blast.ncbi.nlm.nih.gov; Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
Table 3.
The results of alignment of nucleotide sequences of bacterial isolates obtained from Oats (Avena sativa) and 1 sample of Triticale and 1 sample of Cereal-legume by Sanger sequencing (BLAST service, https://blast.ncbi.nlm.nih.gov; Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021).
| Crop | Cultivar | Isolate № | Primers | Result of identification |
|---|---|---|---|---|
| Oats | Verny‘j | 89 | 27f-907 | Pantoea sp. |
| 90 | 27f-907 | Curtobacterium sp. | ||
| Oats | Skakun | 91 | 27f-907 | Rosenbergiella sp |
| 92 | 27f-907 | Pseudomonas sp. | ||
| Oats | Podgorny‘j | 93 | 27f-907 | Pantoea agglomerans |
| 94 | 27f-907 | Pantoea sp. | ||
| Oats | – | 101 | PSf- PSr | Pseudomonas sp. |
| Triticale, wheat, barley | – | 99 | 27f-907 | Arthrobacter sp. |
| Cereal-legume mix | – | 100 | 27f-907 | Frigoribacterium sp. |
4 samples of Winter Oats were collected from which 7 colonies were isolated and identified with the use of 2 primers and 1 sample of Triticale, and 1 sample of Cereal-legume were collected from which 2 colonies were isolated and identified with the use of 1 primer set (27f-907).
From the 38 samples collected in The Republic of Crimea, 95 bacterial colonies were isolated whose DNA was extracted, of these 68 were identified to genus level and 22 to species level.
The methods applied in the study made it possible to determine the frequency of occurrence of various bacteria at genera and a species level. Frequency of occurrence of the Pantoea and Erwinia genera was the highest at 28.95% (Table 4). The diversity of the bacteria is shown with the most common being Pantoea agglomerans, Pantoea ananatis, Pantoea vagans and Pantoea pleuroti with various frequency of occurrence. Table 4 shows the frequency of occurrence of all the detected and identified bacteria.
Table 4.
The frequency of occurrence of species and genera of bacteria in samples. of grain crops from Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021.
Table 4.
The frequency of occurrence of species and genera of bacteria in samples. of grain crops from Belogorskij, Krasnogvardejskij and Simferopol‘skij of the Republic of Crimea 2021.
| Genus | Frequency % | Species | Frequency % |
|---|---|---|---|
| Pantoea | 28.95 |
Pantoea agglomerans Pantoea ananatis Pantoea vagans Pantoea pleuroti |
18.42 2.63 2.63 2.63 |
| Erwinia | 28.95 |
Erwinia rhapontici Erwinia aphidicola |
2.63 5.26 |
| Rosenbergiella | 18.42 | - | - |
| Frigoribacterium | 15.79 | - | - |
| Stenotrophomonas | 7.89 | Stenotrophomonas maltophilia | 2.63 |
| Pseudomonas | 13.16 | Pseudomonas poae | 2.63 |
| Ochrobactrum | 10.53 | - | - |
| Arthrobacter | 10.53 | - | - |
| Uncultured bacterium | 7.89 | Uncultured bacterium - | 7.89 |
| Exiguobacterium | 5.26 | - | - |
| Curtobacterium | 7.89 | - | - |
| Microbacterium | 5.26 | - | - |
| Clavibacter | 5.26 | Clavibacter michiganensis | 5.26 |
| Bacteria | 5.26 | Bacteria | 5.26 |
| Rathayibacter | 2.63 | Rathayibacter festucae | 2.63 |
| Enterococcus | 5.26 | Enterococcus mundtii | 5.26 |
| Agrococcus | 2.63 | Agrococcus jenensis | 2.63 |
| Plantibacter | 5.26 | Plantibacter flavus | 2.63 |
| Enterobacter | 2.63 | - | - |
Note – means species were not identified within the given genus frequency of the various bacteria identified. For each identified species and genus, the frequency of occurrence (A) was calculated using the formula [13]: A = B/C*100%, where B is the number of samples on which a bacterium with a certain species was found, C is the total number of analyzed samples.
4. Discussion
From the 38 plant samples, 95 colonies were collected and 22 were identified to species level and other 68 were identified to genus level from which DNA was extracted and PCR was run and as a result the bacteria shown in Table 2, Table 3 and Table 4. Colony morphology, color shape and elevation we studied [21].
Pantoea sp., bacteria are mostly gram negative and form yellow, dark brown lesions on wheat [22]. This genus encompasses phytopathogenic bacteria and beneficial bacteria. Pantoea ananatis (strain TZ39) is a phytopathogen in rice where it causes bacterial blight [23]. Some Pantoea sp species have some strains that are beneficial towards plants particularly in the rhizosphere [24]. P. agglomerans (BSL 2) strains are a good example. The strains of this bacterium, in particular strains C9-1 and E325 are used as biocontrol agents commercially [9]. Pantoea sp. TW18 has been reviewed and is highly recommended for industrial use that is in remediation of radionuclides in environmental cleanup [24,25].
Pantoea agglomerans is a plant bacterium that exists both as an endophyte and epiphyte [26]. Some P. agglomerans strain causes disease like bacterial blight in grain crops including maize; wheat; cotton onion and some ornamental plants [18,19]. Pantoea agglomerans has some strains that are beneficial to plants as it contributes to the wholesomeness of the rhizosphere and is beneficial towards the host plants [20].
Pantoea ananatis is a gram-negative, facultatively anaerobic, rod shaped, aerobic bacterium. P. ananatis forms brownish lesions with clear margins and yellow halos on wheat leaves causing bacterial disease. Cereal leaf beetle (CLB, Oulema melanopus, Coleoptera, Chrysomelidae) is a vector of Pantoea ananatis and feeds also on, oat and barley. The pathogen is transmitted via oral secretion by insects [16]. Other vectors of P. ananatis are Diabrotica virgifera, tobacco thrips, onion thrips, cotton fleahoppers (Pseudatomoscelis seriatus), mulberry pyralid (Glyphodes pyloalis), ticks, lice, and fleas [27,28,29,30,31]. This phytopathogen was noted for causing mulberry bacterial wilt [32]. It is also noted for increasing crop yield of host plants as a result of promoting plant growth. [33].
Pantoea vagans is a hydrogen-oxidase positive, non-spore forming gram negative short-roded and oxidase negative bacterium [34]. This bacterium’s colonies are convex yellow-beige, smoothed edged colonies [35]. Previously known as a strain of Pantoea agglomerans it is a beneficial bacterium. It controls Erwinia amylovora which causes fire blight in apple and pear trees [36].
Pantoea pleuroti is a short-roded, non-spore forming, motile non-capsulated gram-negative bacterium that causes bacterial blight disease in Pleurotus eryngii an edible mushroom [37]. It grows well on tryptone glucose soy agar, forming yellow, convex, rough, round with entire margins. The bacteria grow well between 20- 37 0C and also at 40 oC and does not grow at 4, 10 and 44 oC [37].
Erwinia sp., bacteria are short rodded, non-spore forming facultative anaerobic gram-negative bacteria [38]. Some of the strains like Erwinia sp. (PR6) are actually beneficial as they promote plant growth [39]. The colonies are usually translucent, light beige circular colonies with soft margins. Erwinia sp is reported to have been found in wheat aphids in Russia [40]. Whilst some strains are beneficial some are harmful to plants. Its species mainly affect grain crops, vegetables and fruit [41].
Erwinia rhapontici was identified in this paper which is responsible for soft rot and pink seed [42]. In infected seeds it mainly causes premature plant death and poor plant emergence [17].
Erwinia aphidicola is a fermentative, oxidase negative bacterium [43]. When it infects seed poor germination is a common trait [40] It also causes bacterial leaf spot in common bean (Phaseolus vulgaris) and pea (Pisum sativum) [44].
Stenotrophomonas sp. is a gram-negative bacterium. It is a soil borne pathogen and also affects humans. Its hosts are both plants and soils. Usually located in either internal tissue like the root and stem vascular tissues and the external areas like the rhizosphere Bacterium pathogens belonging to this group are up to ten, including Stenotrophomonas koreensis, Stenotrophomonas rhizophila, Stenotrophomonas maltophilia, Stenotrophomonas acidaminiphila, Stenotrophomonas humi, Stenotrophomonas nitritireducens, Stenotrophomonas terrae and Stenotrophomonas chelatiphaga [45].
Stenotrophomonas. maltophilia is a beneficial bacterium to the ecosystem and to the health of the plant, thus making it of economic value. This species plays a vital role in plant and soil cycles thus making them very valuable to the soil ecosystem. Further laboratory work has seen this bacterium being used in the production of biomolecules, bioremediation and biocontrol. Some of its strains however have been noted to be pathogenic to humans with a compromised or weak immune system, this bacterium has been noted to be resistant to drugs. S. maltophilia is found either inside the plant that is vascular tissues of the stem and the roots, or outside the plant that is in the rhizosphere of the plants as it is involved in sulphur and nitrogen cycles. This bacterium has not only been noted to assist with plant growth but also assisting in the mitigation of abiotic stress within the plant and also in control of some pathogens [46] This bacterium is usually found in association with plants like oilseed, wheat, potatoes, various weeds and maize amongst others [45,47].
Pseudomonas sp. is a genus that has more than 150 species whose bacteria belong to the family Pseudomonadaceae and are rod shaped, facultative aerobes and gram negative [48]. The species of this genus are of concern as it occupies 20% of the bacteria identified in the grain excluding oats, triticale and barley. Pseudomonas spp comprise of phytopathogens like Pseudomonas putida and Pesudomonas syringae [49,50]. Pseudomonas syringae pv. atrofaciens causing a basal glume rot [51]. Not all species within this genus are harmful some of them are actually beneficial, to plant growth particularly Pseudomonas gramilis, Pseudomonas fluorescens, Pseudomonas migulae and Pseudomonas lini [52]
Pseudomonas poae is a beneficial microbe in the rhizosphere of plants. One study showed that in the rhizosphere of P. heterophylla, the presence of F. oxysporum promotes the growth of P. poae and also is responsible for attracting it [53]. This bacterium has been noticed to be beneficial in the conversion of phosphorous to make it available for plant uptake and in some cases also for nitrogen [54]. In enabling the uptake of nutrients by breaking them down this bacterium is known to enhance plant height [55].
Clavibacter michiganensis it is a gram-positive bacterium [56]. this bacterium affects mainly the xylem vessels on the plants [57]. It causes systematic vascular bacterial wilt of plants and leaf blight [58]. In wheat the bacterium is known to cause mosaic disease symptoms. It mainly affects tomatoes by causing bacterial canker. This bacterium transmitted via seed and bacteria infested plant residues [59]
Plantibacter flavus promotes plant growth. Two strains of the bacterial pathogen M251 and M259 were found to increase the length root-length and total biomass and promote shoot and root growth respectively. These two bacterial strains were found to have genes that are beneficial to plant pathways namely auxin and cytokinin biosynthesis and 1-aminocyclopropane-1-carboxylate deaminase production due to the 70 actinobacterial putative plant-associated genes [60].
Microbacteriaceae bacterium are mainly aerobic, rod shaped, motile, gram positive, mesophilic and a non-spore forming bacteria [60]. They are widely dispersed in diverse terrestrial and aquatic settings and frequently live inside of plants as pathogens or endophytes [61]. In plants afflicted by various nematodes, numerous putative novel species of multiple genera in this family were discovered alongside representatives of recognized species [62]. The leaves of various herbaceous and woody plants infected with nematodes, leaf-mining insects, and plant parasitic mites were found to include Microbacteriacea strains in both affected and unaffected areas [63].
Curtobacterium sp., has been identified not just an endophyte in plants but also a plant disease causative agent in grain plants including rice and sorghum and in citrus like grapevine and strawberries [64]. This genus includes the species like Curtobacterium flaccumfaciens this is a seed borne, motile, aerobic, gram positive bacterium whose mortility is aided by flagella whose colonies are usually shiny, round, yellow and smooth in some cases even convex on Potato Dextrose Agar (PDA) [65,66,67]. This pathogen is of economic significance as it infects both juvenile and aged plants by causing wilting because it affects the vascular system of plants [68]. Meloidogyne incognita is a nematode that facilitates this pathogen in entering the plant via wounds. It is responsible for diseases mainly in beans like bean bacterial wilt [69].
Rathayibacter festucae mainly affects wheat and cause yellow bacterial slime in wheat kennels on the leaves and stems leading to gumming diseases. This bacterium produces rose orange colonies [70]. The bacteria are known to cause yellow bacterial slime on host plants’ leaves, seedheads and sterms therefore causing gumming diseases [71]. Some bacterial pathogens like Rathayibacter sp are transmitted by nematode and upon infection they cause decay and more openings for more bacteria to infect the plants [70].
Agrococcus jenensis is a yellow pigment, non-spore forming, yellow pigment, gram positive and non-motile bacterium [72]. It grows between 15 to 37oC with optimal growth at 30oC at 42oC no growth is noted and a pH of 5 to 11 with an optimal pH of 7.5 [72]. In its genome this bacterium has alkanesulfonate assimilation genes. These are commonly used in paints and thus it is also classified as a beneficial pathogen[73].
Enterococcus mundtii is a bacterium that can be isolated from plant surfaces It is a catalase negative, gram-positive, facultative anaerobic bacterium that can survive in extreme conditions of temperatures in the range 5-65 0C and pH 4.5-10 and also in other hostile environments [74]. This bacterium is also known as a lactic acid bacteria (LAB). LAB species aid in silage fermentation and as such influence silage quality [75]. LAB species produce bacteriocins these can be used in food presevation and most importantly in the control of Listeria monocytogenes which are known to infect vegetables via the roots and are a hazard to both human and animal health. Application of E. mundtii strains (WFE3, WFE20 and WFE31) to soil has reduced levels of Listeria monocytogenes in soil thus aiding in the improvement of nitrate nitrogen (NO3−-N) levels of soil [76].
Rosenbergiella sp. are oxidase negative, gram-negative, facultatively anaerobic, motile, rod-shaped bacteria that will not grow beyond 36 °C with optimum growth at 28–30 °C. It forms yellow-orange-pigmented colonies on nutrient media [77]. They are known to produce high levels of IAA (indole acetic acid) and thus improving plant germination and the growth rate of plants [78].
Frigoribacterium sp., are aerobic, short rodded, non-motile gram positive bacteria that produce white colonies. Optimum conditions for growth are between 8 pH level and 25 - 30 °C, though growth has been noted even at 10 to 45 °C and pH levels of 5 – 10 [79,80].
Ochrobactrum sp. are salt tolerant, ammonia oxidizing bacteria. They are beneficial bacteria at a commercial level wherethey are used to ammend soils. These bacteria are also cold resistant thus making it very useful in ammonia degradation for nitrogen fixation [81].
Arthrobacter sp. are aerobic, gram-stain-positive, non-spore-forming bacteria that is soil borne. Bacteria strains within this genome aid plant growth by assisting in phosphate solubilization [82]. They only grow within a pH range of 5 to 11 and a temperature of 10 °C to 37 °C , however growth optimums for temperature and pH are 25 °C and 7 respectively. On nutrient media it produces cream circular convex colonies [83].
Exiguobacterium sp.are bacteria that can survive in a wide range of temperatures that has proven to be vital in biodegradation of plastic [84]. These bacteria considered beneficial as they help improve soil nitrogen content by harboring nitrogen fixing traits [85]. Some of its bacterial strains (Exiguobacterium sp. WS1-12) were also noted to have a positive effect on root hair formation thus a positive effect on plant growth including grain (wheat) [82].
Plantibacter sp. aregram-positive bacteria of varying sizes that is rod shaped and produces yellow pigments [86].
5. Conclusion
38 samples of grain crops were collected from three regions of Crimea from which we isolated 95 bacterial colonies were isolated, 68 were identified to genus level, 22 were identified to species level using the molecular genetic diagnostic methods described in this paper. Among them, phytopathogens include Agrococcus jenensis, Pseudomonas sp. and Curtobacterium sp., Pantoea ananatis and Clavibacter michiganensis. Erwinia rhapontici is included in the A1 list of the Euro-Mediterranean Plant Protection Organization (EPPO) meaning it is a missing or un-dected quarantine organism in Brazil and thus regulated by phytosanitary requirements of Brazil, Sudan, Colombia and Mali (https://fsvps.gov.ru/ru). Pantoea ananatis is included in the alarm list of the North American Plant Protection Organization (NAPPO). Whilst Pseudomonas sp. is not mentioned as quarantine pathogen some of the genera within this species are considered to be Quarantine pathogens like Pseudomonas syringae in Mexico, Taiwan and Colombia. P. syringae is currently under regulation but not considered a Quarantine pathogen in countries like Great Britain, Egypt and Zimbabwe. Clavibacter michiganensis is currently on the alarm list of Guinea (https://fsvps.gov.ru/ru, https://gd.eppo.int/). Bacteria with economically useful properties were also isolated and identified: Pantoea agglomerans, Arthrobacter sp., Exiguobacterium sp., Ochrobactrum sp., Rosenbergiella sp., Enterococcus mundtii, Agrococcus jenensis, Plantibacter flavus, Pantoea vagans, Pseudomonas poae and Stenotrophomonas. Maltophilia. According to the literature, Frigoribacterium sp., Plantibacter sp. and Enterobacter sp. neither have pronounced harmful nor beneficial properties. The highest frequency of occurrence, 28.9%, was noted in species belonging to the genus Pantoea and Erwinia. Representatives of the genera Rosenbergiella (18.42%), Frigoribacterium (15.79%), Pseudomonas (13.16%), Ochrobactrum and Arthrobacter (10.53%) are also characterized by a high frequency of occurrence. The obtained experimental data on the species composition of bacteria on grain crops can be used in the analysis of the spread of bacterioses in the territory of the Russian Federation and their diagnostics, as well as for the bioinformatic analysis of bacterial genomes in the search for species specific.
Funding
No funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Eurasian Research Institute (ERI) 2020. An Overview of Agricultural Development of Russia. https://www.eurasian-research.org/publication/an-overview-of-agricultural-development-of-russia/. Accessed 30.12.2022.
- Kapoguzov, E.A., Chupin, R.I., Aleshchenko, V.V. and Bykov, A.A. cereals export factors and impact on wheat price in Russian regions. J. Sib. Fed. Univ. Humanit. soc. sci., 2021/14(12), 1782–1794. DOI: 10.17516/1997-1370-0858. [CrossRef]
- FAO. 2017 Phytosanitary certificates. International Plant Protection Convention https://www.ippc.int/static/media/files/publication/en/2017/10/ISPM_12_2014_En_2017-10-26_InkAm.pdf Accessed 30.12.2022.
- Observatory of Economic Complexity (OEC) 2023 https://oec.world/en/profile/bilateral-product/wheat/reporter/rus?redirect=true Accessed 15.05.2023.
- Fitzgerald TL, Powell JJ, Schneebeli K, Hsia MM, Gardiner DM, Bragg JN, McIntyre CL, Manners JM, Ayliffe M, Watt M, Vogel JP, Henry RJ, Kazan K. Brachypodium as an emerging model for cereal-pathogen interactions. Ann Bot. 2015 Apr;115(5):717-31. doi: 10.1093/aob/mcv010. PMID: 25808446; PMCID: PMC4373291. [CrossRef] [PubMed]
- Sadia Latif, Sameeda Bibi, Rabia Kouser, Hina Fatimah, Saba Farooq, Samar Naseer, Rizwana Kousar, Characterization of bacterial community structure in the rhizosphere of Triticum aestivum L. Genomics, 2020 112(6) 4760-4768.
- Guilbaud C, Morris CE, Barakat M, Ortet P, Berge O. Isolation and identification of Pseudomonas syringae facilitated by a PCR targeting the whole P. syringae group. FEMS Microbiol Ecol. 2016 Jan;92(1): fiv146. doi: 10.1093/femsec/fiv146. Epub 2015 Nov 25. PMID: 26610434. [CrossRef] [PubMed]
- FAO 2023. Global body adopts new measures to stop the spread of plant pests https://www.fao.org/news/story/en/item/1118322/icode/. Accessed 19.03.2023.
- Slovareva O.Y. Detection and identification of wheat and barley phytopathogens in the Russian Federation. Microbiology Independent Research Journal MIR Journal. 2020;7(1):13-23. https://doi.org/10.18527/2500-2236-2020-7-1-13-23. [CrossRef]
- Dutta B, Gitaitis RD, Sanders FH, Booth C, Smith S, Langston DB. First Report of Bacterial Leaf Spot of Pumpkin Caused by Xanthomonas cucurbitae in Georgia, United States. Plant Dis. 2013 Oct;97(10):1375. doi: 10.1094/PDIS-03-13-0317-PDN. PMID: 30722167. [CrossRef] [PubMed]
- Honda-Takinami R., Hata J., Matsuoka K., Hoshi S., Koguchi T., Sato Y., Akaihata H., Kataoka M., Ogawa S., Nishiyama K., Suzutani T., Kojima Y. Association between the presence of bacteria in prostate tissue and histopathology in biopsies from men not complaining of lower urinary tract symptoms. Fukushima Journal of Medical Science, 2022, 68(3): 161-167 (doi: 10.5387/fms.2022-34). [CrossRef]
- Mao DP, Zhou Q, Chen CY, Quan ZX. Coverage evaluation of universal bacterial primers using the metagenomic datasets. BMC Microbiol. 2012 May 3; 12:66. doi: 10.1186/1471-2180-12-66. PMID: 22554309; PMCID: PMC3445835. [CrossRef]
- Kazempour M.N, Kheyrgoo M., Pedramfar H., Rahimian H. Isolation and identification of bacterial glum blotch and leaf blight on wheat (Triticum aestivum L.) in Iran. African Journal of Biotechnology, 2010, 9(20): 2860-2865.
- Белкин Д.Л., Бoндаренкo Г.Н., Яремкo А.Б., Уварoва Д.А. Метoд секвенирoвания в видoвoй идентификации карантинных вредных oрганизмoв. Карантин растений. Наука и практика, 2019, 28(2): 31-34.
- Минаева Л.П., Самoхвалoва Л.В., Завриев С.К., Стахеев А.А. Первoе выявление гриба Fusarium coffeatum на территoрии Рoссийскoй Федерации. Сельскoхoзяйственная биoлoгия, 2022, 57(1): 131-140 (doi: 10.15389/agrobiology.2022.1.131rus). [CrossRef]
- Krawczyk, K.; Foryś, J.; Nakonieczny, M.; Tarnawska, M.; Bereś, P.K. Transmission of Pantoea ananatis, the causal agent of leaf spot disease of maize (Zea mays), by western corn rootworm (Diabrotica virgifera virgifera LeConte). Crop Prot. 2020.
- Harada H, Oyaizu H, Kosako Y, Ishikawa H. Erwinia aphidicola, a new species isolated from pea aphid, Acyrthosiphon pisum. J Gen Appl Microbiol. 1997 Dec;43(6):349-354. doi: 10.2323/jgam.43.349. PMID: 12501306. [CrossRef] [PubMed]
- Bruisson S, Zufferey M, L’Haridon F, Trutmann E, Anand A, Dutartre A, De Vrieze M, Weisskopf L. Endophytes and Epiphytes from the Grapevine Leaf Microbiome as Potential Biocontrol Agents Against Phytopathogens. Frontiers in Microbiology. 2019. 10.3389/fmicb.2019.02726 ISSN=1664-302X. [CrossRef]
- Yang Y, Yang J, Wu WM, Zhao J, Song Y, Gao L, Yang R, Jiang L. "Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 2. Role of Gut Microorganisms". Environ. Sci. Technol. 2015 49 (20): 12087–93. Bibcode:2015EnST...4912087Y. doi: 10.1021/acs.est.5b02663. PMID 26390390. [CrossRef]
- Carobbi A, Di Nepi S, Fridman CM, Dar Y, Ben-Yaakov R, Barash I, Salomon D, Sessa G. An antibacterial T6SS in Pantoea agglomerans pv. betae delivers a lysozyme-like effector to antagonize competitors. Environ Microbiol. 2022 Oct;24(10):4787-4802. doi: 10.1111/1462-2920.16100. Epub 2022 Jun 20. PMID: 35706135; PMCID: PMC9796082. [CrossRef]
- van Teeseling MCF, de Pedro MA and Cava F. Determinants of Bacterial Morphology: From Fundamentals to Possibilities for Antimicrobial Targeting. Front. Microbiol.2017. 8:1264. doi: 10.3389/fmicb.2017.01264. [CrossRef]
- Krawczyk K, Wielkopolan B, Obrępalska-Stęplowska A. Pantoea ananatis, A New Bacterial Pathogen Affecting Wheat Plants (Triticum aestivum L.) in Poland. Pathogens. 2020 Dec 21;9(12):1079. doi: 10.3390/pathogens9121079. PMID: 33371529; PMCID: PMC7767503. [CrossRef]
- Yu L, Yang C, Ji Z, Zeng Y, Liang Y, Hou Y. Complete Genomic Data of Pantoea ananatis Strain TZ39 Associated with New Bacterial Blight of Rice in China. Plant Dis. 2022 Feb;106(2):751-753. doi: 10.1094/PDIS-08-21-1845-A. Epub 2022 Feb 9. PMID: 34597149. [CrossRef]
- Tahir M, Khan MB, Shahid M, Ahmad I, Khalid U, Akram M, Dawood A, Kamran M. Metal-tolerant Pantoea sp. WP-5 and organic manures enhanced root exudation and phytostabilization of cadmium in the rhizosphere of maize. Environ Sci Pollut Res Int. 2022 Jan;29(4):6026-6039. doi: 10.1007/s11356-021-16018-3. Epub 2021 Aug 25. PMID: 34431061. [CrossRef] [PubMed]
- Zhang Z, Liu H, Song W, Ma W, Hu W, Chen T, Liu L. Accumulation of U(VI) on the Pantoea sp. TW18 isolated from radionuclide-contaminated soils. J Environ Radioact. 2018 Dec; 192:219-226. doi: 10.1016/j.jenvrad.2018.07.002. Epub 2018 Jul 6. PMID: 29982006. [CrossRef] [PubMed]
- Dutkiewicz J, Mackiewicz B, Kinga Lemieszek M, Golec M, Milanowski J. Pantoea agglomerans: a mysterious bacterium of evil and good. Part III. Deleterious effects: infections of humans, animals and plants. Ann Agric Environ Med. 2016 Jun 2;23(2):197-205. doi: 10.5604/12321966.1203878. PMID: 27294620. [CrossRef] [PubMed]
- Gitaitis, R.D.; Walcott, R.R.; Wells, M.L.; Perez, J.C.D.; Sanders, F.H. Transmission of Pantoea ananatis, Causal Agent of Center Rot of Onion, by Tobacco Thrips, Frankliniella fusca. Plant Dis. 2003, 87, 675–678.
- Gitaitis, R.D.; Walcott, R.R.; Wells, M.L.; Perez, J.C.D.; Sanders, F.H. Transmission of Pantoea ananatis, Causal Agent of Center Rot of Onion, by Tobacco Thrips, Frankliniella fusca. Plant Dis. 2003, 87, 675–678.
- Coutinho, T.A.; Venter, S.N. Pantoea ananatis: An unconventional plant pathogen. Mol. Plant Pathol. 2009, 10, 325–335.
- Krawczyk, K.; Wielkopolan, B.; Obrępalska-Stęplowska, A. Pantoea ananatis, A New Bacterial Pathogen Affecting Wheat Plants (Triticum L.) in Poland. Pathogens 2020, 9, 1079.
- Murrell, A.; Dobson, S.J.; Yang, X.; Lacey, E.; Barker, S.C. A survey of bacterial diversity in ticks, lice and fleas from Australia. Parasitol. Res. 2003, 89, 326–334.
- Yuan T, Huang Y, Luo L, Wang J, Li J, Chen J, Qin Y, Liu J. Complete Genome Sequence of Pantoea ananatis strain LCFJ-001, Isolated from Bacterial Wilt Mulberry. Plant Dis. 2023 Jan 23. doi: 10.1094/PDIS-10-22-2473-A. Epub ahead of print. PMID: 36691281. [CrossRef]
- Usuda Y, Nishio Y, Nonaka G, Hara Y. Microbial Production Potential of Pantoea ananatis: From Amino Acids to Secondary Metabolites. Microorganisms. 2022 May 31;10(6):1133. doi: 10.3390/microorganisms10061133. PMID: 35744651; PMCID: PMC9231021. [CrossRef]
- Chen X, Sun Q, Tang L, Guo T, Huang S, Mo J, Li Q. First Report of Bacterial Necrosis Caused by Pantoea vagans in Mango in China. Plant Dis. 2022 Nov 9. doi: 10.1094/PDIS-08-22-1950-PDN. Epub ahead of print. PMID: 36350723. [CrossRef] [PubMed]
- Brady CL, Venter SN, Cleenwerck I, Engelbeen K, Vancanneyt M, Swings J, Coutinho TA. Pantoea vagans sp. nov., Pantoea eucalypti sp. nov., Pantoea deleyi sp. nov. and Pantoea anthophila sp. nov. Int J Syst Evol Microbiol. 2009 Sep;59(Pt 9):2339-45. doi: 10.1099/ijs.0.009241-0. Epub 2009 Jul 20. PMID: 19620357. [CrossRef] [PubMed]
- Silvana Díaz Herreraa, Cecilia Grossi, Myriam Zawoznika, María Daniela Groppa. Wheat seeds harbour bacterial endophytes with potential as plant growth promoters and biocontrol agents of Fusarium graminearum. Microbiological Research. 2016 37–43.
- Ma Y, Yin Y, Rong C, Chen S, Liu Y, Wang S, Xu F. Pantoea pleuroti sp. nov., Isolated from the Fruiting Bodies of Pleurotus eryngii. Curr Microbiol. 2016 Feb;72(2):207-212. doi: 10.1007/s00284-015-0940-5. Epub 2015 Nov 19. PMID: 26581526. [CrossRef] [PubMed]
- Pan MK, Feng GD, Yao Q, Li J, Liu C, Zhu H. Erwinia phyllosphaerae sp. nov., a novel bacterium isolated from phyllosphere of pomelo (Citrus maxima). Int J Syst Evol Microbiol. 2022 Apr;72(4). doi: 10.1099/ijsem.0.005316. PMID: 35416765. [CrossRef] [PubMed]
- Sagar A, Sing A., Labhane N., Riyazuddin R., Marker S., Parihar D.K and Ramteke P.W. Native bacterium Erwinia sp. (PR16) enhances growth and yield of wheat variety AAI-W6 under reduced level of NPK. International Journal of Life Sciences and Applied Sciences. 2020, 2 27-36.
- Campillo T, Luna E, Portier P, Fischer-Le Saux M, Lapitan N, Tisserat NA, Leach JE. Erwinia iniecta sp. nov., isolated from Russian wheat aphid (Diuraphis noxia). Int J Syst Evol Microbiol. 2015 Oct;65(10):3625-3633. doi: 10.1099/ijsem.0.000466. PMID: 26198254. [CrossRef] [PubMed]
- Morohoshi T, Nameki K, Someya N. Comparative genome analysis reveals the presence of multiple quorum-sensing systems in plant pathogenic bacterium, Erwinia rhapontici. Biosci Biotechnol Biochem. 2021 Jul 23;85(8):1910-1914. doi: 10.1093/bbb/zbab104. PMID: 34100908. [CrossRef] [PubMed]
- Krawczyk, K.; Wielkopolan, B.; Obrępalska-Stęplowska, A. Pantoea ananatis, A New Bacterial Pathogen Affecting Wheat Plants (Triticum L.) in Poland. Pathogens 2020, 9, 1079.
- Halpern M, Fridman S, Atamna-Ismaeel N, Izhaki I. Rosenbergiella nectarea gen. nov., sp. nov., in the family Enterobacteriaceae, isolated from floral nectar. Int J Syst Evol Microbiol. 2013 Nov;63(Pt 11):4259-4265. doi: 10.1099/ijs.0.052217-0. Epub 2013 Jul 5. PMID: 23832968. [CrossRef] [PubMed]
- Asif M, Eudes F, Randhawa H, Amundsen E, Yanke J, Spaner D. Cefotaxime prevents microbial contamination and improves microspore embryogenesis in wheat and triticale. Plant Cell Rep. 2013 Oct;32(10):1637-46. doi: 10.1007/s00299-013-1476-4. Epub 2013 Jul 30. PMID: 23896731. [CrossRef] [PubMed]
- Ryan RP, Monchy S, Cardinale M, Taghavi S, Crossman L, Avison MB, Berg G, van der Lelie D, Dow JM. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat Rev Microbiol. 2009 Jul;7(7):514-25. doi: 10.1038/nrmicro2163. PMID: 19528958. [CrossRef] [PubMed]
- Haque MM, Mosharaf MK, Khatun M, Haque MA, Biswas MS, Islam MS, Islam MM, Shozib HB, Miah MMU, Molla AH, Siddiquee MA. Biofilm Producing Rhizobacteria with Multiple Plant Growth-Promoting Traits Promote Growth of Tomato Under Water-Deficit Stress. Front Microbiol. 2020 Nov 26;11 :542053. doi: 10.3389/fmicb.2020.542053. PMID: 33324354; PMCID: PMC7727330. [CrossRef]
- Gilligan PH, Lum G, VanDamme PAR, Whittier S. Murray PR, Baron EJ, Jorgensen JH, et al. (eds.). Burkholderia, Stenotrophomonas, Ralstonia, Brevundimonas, Comamonas, Delftia, Pandoraea, and Acidivorax. In: Manual of Clinical Microbiology (8th ed.). 2003 ASM Press, Washington, DC. pp. 729–748. ISBN 978-1-55581-255.
- Euzéby JP. List of Bacterial Names with Standing in Nomenclature: a folder available on the Internet. Int J Syst Bacteriol. 1997 Apr;47(2):590-2. doi: 10.1099/00207713-47-2-590. PMID: 9103655. [CrossRef] [PubMed]
- Anzai Y, Kim H, Park JY, Wakabayashi H, Oyaizu H. Phylogenetic affiliation of the pseudomonads based on 16S rRNA sequence. Int J Syst Evol Microbiol. 2000 Jul;50 Pt 4:1563-1589. doi: 10.1099/00207713-50-4-1563. PMID: 10939664. [CrossRef] [PubMed]
- Arnold DL, Preston GM. Pseudomonas syringae: enterprising epiphyte and stealthy parasite. Microbiology (Reading). 2019 Mar;165(3):251-253. doi: 10.1099/mic.0.000715. Epub 2018 Nov 14. PMID: 30427303. [CrossRef] [PubMed]
- Krawczyk, K.; Wielkopolan, B.; Obrępalska-Stęplowska, A. Pantoea ananatis, A New Bacterial Pathogen Affecting Wheat Plants (Triticum L.) in Poland. Pathogens 2020, 9, 1079.
- Diggle SP, Whiteley M. Microbe Profile: Pseudomonas aeruginosa: opportunistic pathogen and lab rat. Microbiology (Reading). 2020 Jan;166(1):30-33. doi: 10.1099/mic.0.000860. Erratum in: Microbiology (Reading). 2021 Aug;167(8): PMID: 31597590; PMCID: PMC7273324. [CrossRef]
- Padda KP, Puri A, Chanway C. Endophytic nitrogen fixation - a possible ’hidden’ source of nitrogen for lodgepole pine trees growing at un-reclaimed gravel mining sites. FEMS Microbiol Ecol. 2019 Nov 1;95(11): fiz172. doi: 10.1093/femsec/fiz172. PMID: 31647534. [CrossRef] [PubMed]
- Yuan QS, Wang L, Wang H, Wang X, Jiang W, Ou X, Xiao C, Gao Y, Xu J, Yang Y, Cui X, Guo L, Huang L, Zhou T. Pathogen-Mediated Assembly of Plant-Beneficial Bacteria to Alleviate Fusarium Wilt in Pseudostellaria heterophylla. Front Microbiol. 2022 Mar 30;13: 842372. doi: 10.3389/fmicb.2022.842372. PMID: 35432244; PMCID: PMC9005978. [CrossRef]
- Irshad U, Yergeau E. Bacterial Subspecies Variation and Nematode Grazing Change P Dynamics in the Wheat Rhizosphere. Front Microbiol. 2018 Sep 5;9: 1990. doi: 10.3389/fmicb.2018.01990. PMID: 30233510; PMCID: PMC6134019. [CrossRef]
- Oh EJ, Hwang IS, Park IW, Oh CS. Comparative Genome Analyses of Clavibacter michiganensis Type Strain LMG7333T Reveal Distinct Gene Contents in Plasmids from Other Clavibacter Species. Front Microbiol. 2022 Feb 1;12: 793345. doi: 10.3389/fmicb.2021.793345. PMID: 35178040; PMCID: PMC8844524. [CrossRef]
- Eichenlaub R, Gartemann KH. The Clavibacter michiganensis subspecies: molecular investigation of gram-positive bacterial plant pathogens. Annu Rev Phytopathol. 2011; 49:445-64. doi: 10.1146/annurev-phyto-072910-095258. PMID: 21438679. [CrossRef] [PubMed]
- Baek KY, Lee HH, Son GJ, Lee PA, Roy N, Seo YS, Lee SW. Specific and Sensitive Primers Developed by Comparative Genomics to Detect Bacterial Pathogens in Grains. Plant Pathol J. 2018 Apr;34(2):104-112. doi: 10.5423/PPJ.OA.11.2017.0250. Epub 2018 Apr 1. PMID: 29628816; PMCID: PMC5880354. [CrossRef]
- Shree P. Thapa, Sivakumar Pattathil, Michael G. Hahn, Marie-Agnès Jacques, Robert L. Gilbertson, and Gitta Coaker. Genomic Analysis of Clavibacter michiganensis Reveals Insight into Virulence Strategies and Genetic Diversity of a Gram-Positive Bacterial Pathogen. Molecular Plant-Microbe Interactions. 2017 30 (10), 786-802.
- Lymperopoulou DS, Coil DA, Schichnes D, Lindow SE, Jospin G, Eisen JA, Adams RI. Draft genome sequences of eight bacteria isolated from the indoor environment: Staphylococcus capitis strain H36, S. capitis strain H65, S. cohnii strain H62, S. hominis strain H69, Microbacterium sp. strain H83, Mycobacterium iranicum strain H39, Plantibacter sp. strain H53, and Pseudomonas oryzihabitans strain H72. Stand Genomic Sci. 2017 Jan 31; 12:17. doi: 10.1186/s40793-017-0223-9. PMID: 28163826; PMCID: PMC5282799. [CrossRef]
- Lymperopoulou DS, Coil DA, Schichnes D, Lindow SE, Jospin G, Eisen JA, Adams RI. Draft genome sequences of eight bacteria isolated from the indoor environment: Staphylococcus capitis strain H36, S. capitis strain H65, S. cohnii strain H62, S. hominis strain H69, Microbacterium sp. strain H83, Mycobacterium iranicum strain H39, Plantibacter sp. strain H53, and Pseudomonas oryzihabitans strain H72. Stand Genomic Sci. 2017 Jan 31; 12:17. doi: 10.1186/s40793-017-0223-9. PMID: 28163826; PMCID: PMC5282799. [CrossRef]
- Tarlachkov SV, Starodumova IP, Dorofeeva LV, Prisyazhnaya NV, Roubtsova TV, Chizhov VN, Nadler SA, Subbotin SA, Evtushenko LI. 2021. Draft genome sequences of 28 actinobacteria of the family Microbacteriaceae associated with nematode-infected plants. Microbiol Resour Announc 10: e01400-20. https://doi.org/10.1128/MRA.01400-20. [CrossRef]
- Tarlachkov SV, Ospennikov YV, Demidov AV, Starodumova IP, Dorofeeva LV, Prisyazhnaya NV, Chizhov VN, Subbotin SA, Evtushenko LI. Draft Genome Sequences of 9 Actinobacteria from the Family Microbacteriaceae Associated with Insect- and Nematode-Damaged Plants. Microbiol Resour Announc. 2022 Oct 20;11(10):e0048722. doi: 10.1128/mra.00487-22. Epub 2022 Aug 31. PMID: 36043868; PMCID: PMC9584284.]. [CrossRef]
- Mayer E, Dörr de Quadros P, Fulthorpe R. Plantibacter flavus, Curtobacterium herbarum, Paenibacillus taichungensis, and Rhizobium selenitireducens Endophytes Provide Host-Specific Growth Promotion of Arabidopsis thaliana, Basil, Lettuce, and Bok Choy Plants. Appl Environ Microbiol. 2019 Sep 17;85(19): e00383-19. doi: 10.1128/AEM.00383-19. PMID: 31350315; PMCID: PMC6752021. [CrossRef]
- Dimkić I, Bhardwaj V, Carpentieri-Pipolo V, Kuzmanović N, Degrassi G. The chitinolytic activity of the Curtobacterium sp. isolated from field-grown soybean and analysis of its genome sequence. PLoS One. 2021 Nov 3;16(11): e0259465. doi: 10.1371/journal.pone.0259465. PMID: 34731210; PMCID: PMC8565777. [CrossRef]
- Kariluoto S, Edelmann M, Herranen M, Lampi AM, Shmelev A, Salovaara H, Korhola M, Piironen V. Production of folate by bacteria isolated from oat bran. Int J Food Microbiol. 2010 Sep 30;143(1-2):41-7. doi: 10.1016/j.ijfoodmicro.2010.07.026. Epub 2010 Aug 11. PMID: 20708290. [CrossRef] [PubMed]
- Bastas KK and Sahin F. Evaluation of seedborne bacterial pathogens on common bean cultivars grown in central Anatolia region. Turkey. European Journal of Plant Pathology, 2017 147, 239–253.
- EFSA Panel on Plant Health (EFSA PLH Panel); Jeger M, Bragard C, Caffier D, Candresse T, Chatzivassiliou E, Dehnen-Schmutz K, Gilioli G, Grégoire JC, Jaques Miret JA, MacLeod A, Navajas Navarro M, Niere B, Parnell S, Potting R, Rafoss T, Rossi V, Urek G, Van Bruggen A, Van der Werf W, West J, Winter S, Tegli S, Hollo G, Caffier D. Pest categorisation of Curtobacterium flaccumfaciens pv. flaccumfaciens. EFSA J. 2018 May 31;16(5): e05299. doi: 10.2903/j.efsa.2018.5299. PMID: 32625922; PMCID: PMC7009624. [CrossRef]
- Mei L, Piao Z, Hu J, Shi L, Bai Y, Yin S. Lysinimonas yzui sp. nov., isolated from cattail root soil from mine tailings. Int J Syst Evol Microbiol. 2020 Mar;70(3):2003-2007. doi: 10.1099/ijsem.0.004013. PMID: 32234114. [CrossRef] [PubMed]
- Dorofeeva LV, Evtushenko LI, Krausova VI, Karpov AV, Subbotin SA, Tiedje JM. Rathayibacter caricis sp. nov. and Rathayibacter festucae sp. nov., isolated from the phyllosphere of Carex sp. and the leaf gall induced by the nematode Anguina graminis on Festuca rubra L., respectively. Int J Syst Evol Microbiol. 2002 Nov;52(Pt 6):1917-1923. doi: 10.1099/00207713-52-6-1917. PMID: 12508848. [CrossRef] [PubMed]
- Dorofeeva LV, Evtushenko LI, Krausova VI, Karpov AV, Subbotin SA, Tiedje JM. Rathayibacter caricis sp. nov. and Rathayibacter festucae sp. nov., isolated from the phyllosphere of Carex sp. and the leaf gall induced by the nematode Anguina graminis on Festuca rubra L., respectively. Int J Syst Evol Microbiol. 2002 Nov;52(Pt 6):1917-1923. doi: 10.1099/00207713-52-6-1917. PMID: 12508848. [CrossRef] [PubMed]
- Dhanjal S, Kaur I, Korpole S, Schumann P, Cameotra SS, Pukall R, Klenk HP, Mayilraj S. Agrococcus carbonis sp. nov., isolated from soil of a coal mine. Int J Syst Evol Microbiol. 2011 Jun;61(Pt 6):1253-1258. doi: 10.1099/ijs.0.024745-0. Epub 2010 Jul 2. PMID: 20601492. [CrossRef] [PubMed]
- Lau KJX, Junqueira ACM, Uchida A, Purbojati RW, Houghton JNI, Chénard C, Wong A, Kolundžija S, Clare ME, Kushwaha KK, Putra A, Gaultier NE, Heinle CE, Premkrishnan BNV, Vettah VK, Drautz-Moses DI, Schuster SC. Complete Genome Sequence of Agrococcus sp. Strain SGAir0287, Isolated from Tropical Air Collected in Singapore. Microbiol Resour Announc. 2019 Aug 8;8(32): e00616-19. doi: 10.1128/MRA.00616-19. PMID: 31395637; PMCID: PMC6687924. [CrossRef]
- Fisher K, Phillips C. The ecology, epidemiology and virulence of Enterococcus. Microbiology (Reading). 2009 Jun;155(Pt 6):1749-1757. doi: 10.1099/mic.0.026385-0. Epub 2009 Apr 21. PMID: 19383684. [CrossRef]
- Yimin Cai. Identification and Characterization of Enterococcus Species Isolated from Forage Crops and Their Influence on Silage Fermentation. Journal of Dairy Science. 1999. 82 (11) 2466-2471.
- Gaetano Guida, Raimondo Gaglio, Alessandro Miceli, Vito Armando Laudicina, Luca Settanni. Biological control of Listeria monocytogenes in soil model systems by Enterococcus mundtii strains expressing mundticin KS production. Applied Soil Ecology. 2022. 170.
- Halpern M, Fridman S, Atamna-Ismaeel N, Izhaki I. Rosenbergiella nectarea gen. nov., sp. nov., in the family Enterobacteriaceae, isolated from floral nectar. Int J Syst Evol Microbiol. 2013 Nov;63(Pt 11):4259-4265. doi: 10.1099/ijs.0.052217-0. Epub 2013 Jul 5. PMID: 23832968. [CrossRef] [PubMed]
- Caneschi WL, Felestrino ÉB, Fonseca NP, Villa MM, Lemes CGC, Cordeiro IF, Assis RAB, Sanchez AB, Vieira IT, Kamino LHY, do Carmo FF, Garcia CCM, Moreira LM. Brazilian Ironstone Plant Communities as Reservoirs of Culturable Bacteria with Diverse Biotechnological Potential. Front Microbiol. 2018 Jul 23; 9:1638. doi: 10.3389/fmicb.2018.01638. PMID: 30083146; PMCID: PMC6064971. [CrossRef]
- Li AH, Liu HC, Xin YH, Kim SG, Zhou YG. Glaciihabitans tibetensis gen. nov., sp. nov., a psychrotolerant bacterium of the family Microbacteriaceae, isolated from glacier ice water. Int J Syst Evol Microbiol. 2014 Feb;64(Pt 2):579-587. doi: 10.1099/ijs.0.052670-0. Epub 2013 Oct 24. PMID: 24158943. [CrossRef] [PubMed]
- Wang HF, Zhang YG, Chen JY, Guo JW, Li L, Hozzein WN, Zhang YM, Wadaan MAM, Li WJ. Frigoribacterium endophyticum sp. nov., an endophytic actinobacterium isolated from the root of Anabasis elatior (C. A. Mey.) Schischk. Int J Syst Evol Microbiol. 2015 Apr;65(Pt 4):1207-1212. doi: 10.1099/ijs.0.000081. Epub 2015 Jan 21. PMID: 25609679. [CrossRef] [PubMed]
- Poszytek K, Karczewska-Golec J, Ciok A, Decewicz P, Dziurzynski M, Gorecki A, Jakusz G, Krucon T, Lomza P, Romaniuk K, Styczynski M, Yang Z, Drewniak L, Dziewit L. Genome-Guided Characterization of Ochrobactrum sp. POC9 Enhancing Sewage Sludge Utilization-Biotechnological Potential and Biosafety Considerations. Int J Environ Res Public Health. 2018 Jul 16;15(7):1501. doi: 10.3390/ijerph15071501. PMID: 30013002; PMCID: PMC6069005. [CrossRef]
- Patel, J.K., Gohel, K., Patel, H. et al. Wheat Growth Dependent Succession of Culturable Endophytic Bacteria and Their Plant Growth Promoting Traits. Curr Microbiol 78, 4103–4114 (2021).
- Li X, Li S, Wu Y, Li J, Xing P, Wei G, Shi P. Arthrobacter rhizosphaerae sp. nov., isolated from wheat rhizosphere. Arch Microbiol. 2022 Aug 6;204(9):543. doi: 10.1007/s00203-022-03150-y. PMID: 35932431. [CrossRef] [PubMed]
- Yang Y, Yang J, Wu WM, Zhao J, Song Y, Gao L, Yang R, Jiang L. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 2. Role of Gut Microorganisms. Environ Sci Technol. 2015 Oct 20;49(20):12087-93. doi: 10.1021/acs.est.5b02663. Epub 2015 Oct 1. PMID: 26390390. [CrossRef] [PubMed]
- Collins MD, Lund BM, Farrow JA, Schleifer KH (1983). "Chemotaxonomic study of an alkaliphilic bacterium, Exiguobacterium aurantiacum gen nov., sp. nov". J. Gen. Microbiol. 1983 129 (7): 2037–2042.
- Lymperopoulou DS, Coil DA, Schichnes D, Lindow SE, Jospin G, Eisen JA, Adams RI. Draft genome sequences of eight bacteria isolated from the indoor environment: Staphylococcus capitis strain H36, S. capitis strain H65, S. cohnii strain H62, S. hominis strain H69, Microbacterium sp. strain H83, Mycobacterium iranicum strain H39, Plantibacter sp. strain H53, and Pseudomonas oryzihabitans strain H72. Stand Genomic Sci. 2017 Jan 31; 12:17. doi: 10.1186/s40793-017-0223-9. PMID: 28163826; PMCID: PMC5282799. [CrossRef]
- Wang C, Wu W, Wei L, Feng Y, Kang M, Xie Y, Zong Z. Enterobacter wuhouensis sp. nov. and Enterobacter quasihormaechei sp. nov. recovered from human sputum. Int J Syst Evol Microbiol. 2020 Feb;70(2):874-881. doi: 10.1099/ijsem.0.003837. PMID: 31702537. [CrossRef] [PubMed]
- Zhang Z, Liu H, Karani H, Mallen J, Chen W, De A, Mani S, Tang JX. Enterobacter sp. Strain SM1_HS2B Manifests Transient Elongation and Swimming Motility in Liquid Medium. Microbiol Spectr. 2022 Jun 29;10(3): e0207821. doi: 10.1128/spectrum.02078-21. Epub 2022 Jun 1. PMID: 35647691; PMCID: PMC9241836. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



