
Submitted:
28 June 2023
Posted:
06 July 2023
You are already at the latest version
Abstract
Caries are lesions caused by acidic compounds derived from the metabolism of bacteria such as streptococci. It is not merely the presence of acidogenic bacteria that causes caries but an imbalance in the oral biofilm microhabitat and its behavior. One factor underlying biofilm formation is quorum sensing (QS), that in gram-positives relies on peptidic molecules, whereas in gram-negatives is driven by small diffusible signals. Strategies based on QS inhibition have been proposed as alter-natives to antimicrobial therapies. Here, we investigate the antibiofilm potential of the lactonase enzyme Aii20J, previously reported as effective against biofilms of periodontal disease origin. We generated in vitro polymicrobial biofilms dominated by gram-positive taxa using supragingival samples from caries-free and caries-active children. The effect of Aii20J on the biofilms was evalu-ated regarding its biomass-reducing ability, as assessed with crystal violet assay, and its effects on the bacterial composition of the polymicrobial biofilms, as assessed by 16S community profiling. We describe significant biomass reductions upon Aii20J exposure without significant changes in bac-terial composition at the genus level. Our results support the use of Aii20J to prevent oral biofilm formation while highlighting the influence of the polymicrobial environment on bacterial commu-nities and their response to antibiofilm treatments.
Keywords:
oral biofilm
; caries
; quorum sensing
; quorum quenching
; acylhomoserine lactones
; Streptococcus
; 16S community profiling
; caries-free children
; caries-active children
Introduction
Caries is an oral disease that affects children and adults worldwide. The most recent estimate points to 520 million children suffering from caries in deciduous teeth and 2000 million people with caries in permanent teeth in 2019 [1]. When left untreated, caries can progress into more severe diseases, such as root canal infections, and eventually lead to bloodstream infections and endocarditis.
The classical specific plaque hypothesis highlighted the etiological role of bacterial taxa such as Streptococcus mutans as main players in dental caries. Nevertheless, the ecological plaque hypothesis describes caries as a polymicrobial disease resulting from an imbalance in the healthy oral biofilm [2,3]. This imbalance is commonly due to an increased intake of fermentable sugars and carbohydrates that bacteria metabolize, producing acidic compounds. A lowered pH maintained in time eventually surpasses the homeostatic mechanisms of the healthy oral biofilm through the selection of acidogenic and aciduric microorganisms, shifting towards a cariogenic biofilm [4,5,6]. Thus, it is now accepted that the functional activities within oral biofilms have a critical role in the progression of diseases, complementing that of the microbiome structure [3,7,8].
The search for novel strategies for the prevention and treatment of oral diseases has pointed out the possibility of interfering with cariogenic virulence factors without affecting bacterial viability [9,10,11,12,13]. One of these factors is bacterial communication, also known as quorum sensing (QS). The presence of the main QS pathways has been described in caries-associated species and species retrieved from carious lesions. Autoinducer peptides, typically used by gram-positives, have been described in several streptococci, where QS mainly depends on the competence-stimulating peptide (CSP). Interference with the CSP signaling system in Streptococcus mutans has been shown to inactivate virulence factors and impair the persistence of this bacterium in the oral biofilm [14,15]. The autoinducer-2 (AI-2) has already been described in several oral pathogens, specifically streptococci [16,17]. Furthermore, the use of a cellular extract from Tenacibaculum sp. 20J active against AI-2 reduced S. mutans biofilm formation in vitro, possibly by inhibiting AI-2 signaling [9]. Gram-negative’s N-acyl-homoserine lactones (AHLs) had been considered non-relevant in the oral environment [16,18,19,20]. Nevertheless, AHLs have been found in saliva and tooth samples [12,21], and bacterial strains isolated from caries have been shown to produce AHLs [22,23]. Moreover, bacterial genes predicted to be related to AHL biosynthesis have been described in the saliva metagenomes of caries-active children [24]. In addition, two studies on in vitro oral biofilms showed that exposure to AHLs and modified AHLs reduced lactic acid accumulation without affecting bacterial growth [25,26]. More recently, the use of the quorum quenching (QQ) enzyme Aii20J, highly active against AHLs, was described as highly effective against biofilm formation in samples retrieved from patients with periodontal disease [27]. Interestingly, this previous study also found a great effect of the Aii20J enzyme on in vitro biofilms that were largely dominated by streptococci [27]. Altogether, this evidence points to the possibility of interfering with AHL-mediated communication mechanisms to modulate oral biofilm formation, even in bacterial populations where gram-negative bacteria are found only in low proportions.
This study aimed to characterize the effect of the QQ enzyme Aii20J on in vitro biofilms from caries-free and caries-active children. The microbiome structure of the initial supragingival biofilms was investigated by 16S rRNA sequencing, and these supragingival samples were used to generate in vitro biofilms in the presence of the enzyme. The effects of Aii20J were investigated regarding biofilm mass (crystal violet -CV- assay) and microbial composition (Illumina sequencing of the 16S rRNA gene) of the in vitro biofilms. This study describes reductions in biomass of in vitro biofilms generated from samples of caries-free and caries-active children upon treatment with Aii20J. These observations were not accompanied by changes in the microbial diversity of the biofilms, as previously described for biofilms derived from adult patients with periodontal disease [27].
2. Results and Discussion
2.1. Microbial composition of supragingival biofilm samples from caries-free and caries-active children
Supragingival biofilm was sampled from either caries-free or caries-active children (Table 1). To investigate the microbiome structure of these samples, their genomic DNA (gDNA) was extracted to sequence the V3-V4 hypervariable regions of the 16S rRNA gene.
Figure 1A,B shows the relative abundance of the most abundant genera identified in supragingival biofilm samples from caries-free and caries-active children. Streptococci were found in relative abundances ranging from 2 to 34% among subjects. Such proportions are similar to those compiled in the Human Oral Microbiome Database (HOMD) based on the studies of Eren et al. [28] and of Segata et al. [29], with reported relative abundances of 15 – 30% for the genus Streptococcus in supragingival plaque samples. Besides Streptococcus, the most relatively abundant genera were Leptotrichia, Veillonella, Fusobacterium, Capnocytophaga, Prevotella, Neisseria, Corynebacterium, Porphyromonas, and Selenomonas. Yet, supragingival biofilms comprised a much higher number of taxa, with 38 – 71 total genera identified among samples. The less relatively abundant genera comprised proportions of up to 40% of the supragingival biofilms (Figure 1B). The only exception was sample Ca8, taken from a caries-active subject, in which the predominant genera were Lactococcus and Serratia, with relative abundances of 34% and 32%, respectively. Besides, this was the only sample in which these two genera were identified (Figure 1A). The principal components analysis (PCA) of the supragingival biofilm samples separated subject Ca8 from the others (Figure S1). After excluding sample Ca8, the microbiota structure of the supragingival biofilms was analyzed again (Figure 1C). The new PCA displayed an almost complete overlapping between samples from caries-free (yellow) and caries-active children (blue) (Figure 1C). Thus, within this collection of supragingival biofilm samples, the microbiota of caries-free and caries-active children are not significantly different. Sample Ca8 has been excluded from subsequent analyses for consistency purposes. The observations presented in Figure 1C align with those of Belda-Ferre et al. [30], who did not find significant differences in the microbiome structure of supragingival plaque samples collected from healthy adults and patients with caries. It is relevant to note that Belda-Ferre et al. [30] used supragingival plaque samples taken from intact tooth surfaces, as in the present study, and not directly from the carious lesions. Interestingly, the protein profiles of those subgingival samples were clustered according to the caries status of the subjects [30]. Indeed, in recent years the emphasis has been put on the functional activities within oral microbial communities rather than on their microbiota distribution, as the former has been described to account for a greater impact on disease evolution [3,8].
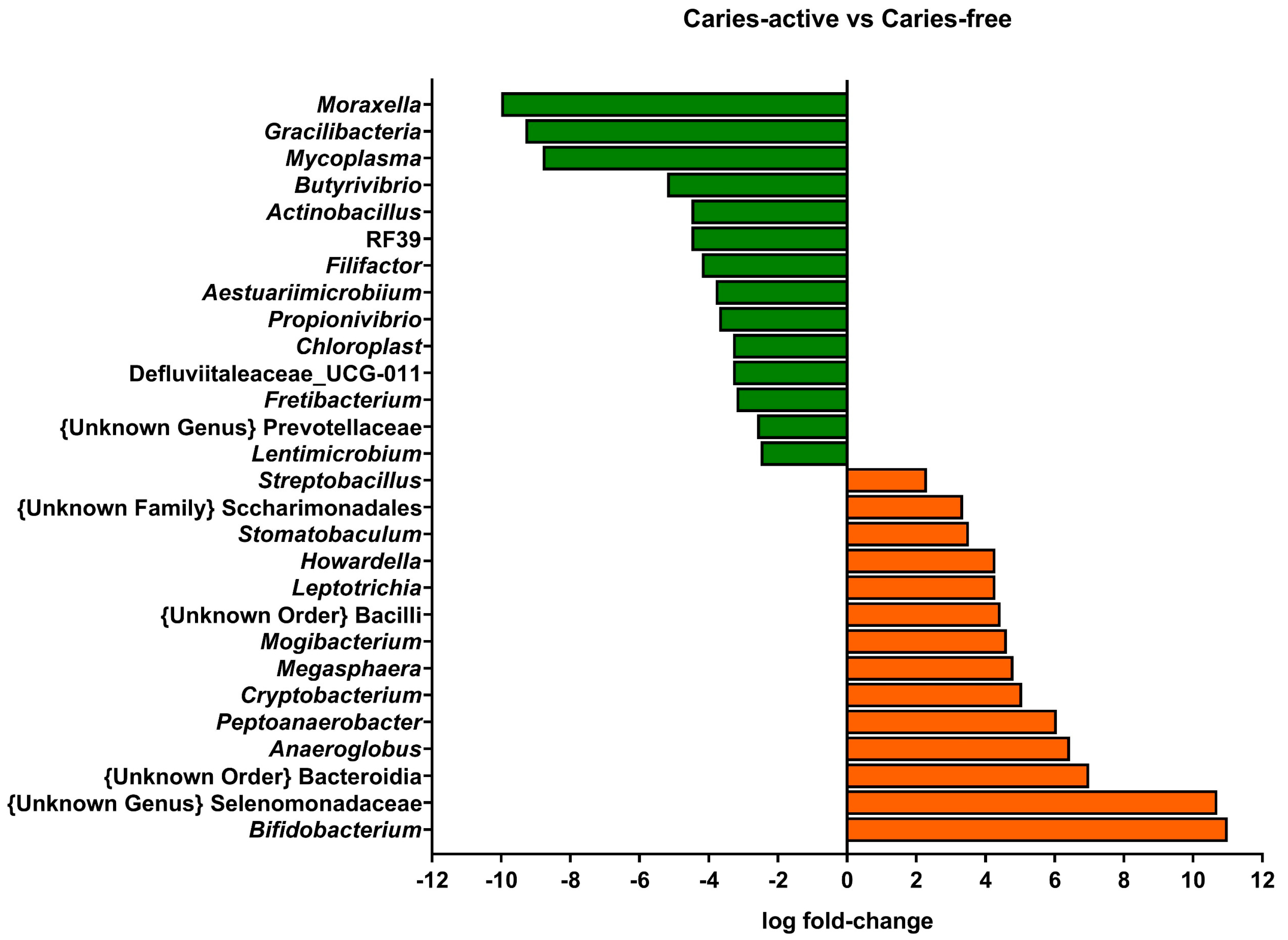
Interestingly, when examining the microbial composition of the samples at the genus level with a linear discriminant analysis (LDA), genera present at very low relative abundances showed significant differential abundances between caries-free and caries-active subjects (Figure 2). Figure 2 shows positive fold-change values for genera that were more abundant in samples from children with active caries relative to caries-free children, whereas negative fold-change values represent the opposite case.
Genera classically associated with caries, such as Streptococcus, did not present relevant differences in relative abundance between caries-free and caries-active subjects, pointing to metabolic activities rather than specific taxon abundance as the most relevant parameter that differentiates cariogenic and healthy biofilms. Instead, low-abundance genera did present significant differences between caries-free and caries-active children. The genus Bifidobacterium was increased in caries-active biofilm samples. Bifidobacterium members can be highly acidogenic, initiating caries or aiding their progression [10,31]. Nevertheless, their increased presence in caries-active subjects can also be derived from a selection of acidogenic bacteria in an acidified environment. The genus Moraxella was significantly increased in caries-free samples compared to caries-active samples. This observation aligns with the study of Belda-Ferre et al. [32], in which they described a tendency of Gammaproteobacteria members to be more common in healthy subjects. The overall homogeneity of the principal genera identified among biofilms of subjects with different oral health statuses reinforces the need for proteomic or metatranscriptomic studies that allow the observation of differences in functional activities among samples and the description of the role of low-abundance taxa that are differentially represented in caries-free and caries-active subjects.
Figure 2.
Linear Discriminant Analysis of the genera identified in supragingival biofilm samples from children with active caries, compared to caries-free children. Genera with log fold-change values lower than 2 (green bars) were considered as significantly reduced in caries-active subjects, and genera with log fold-change values higher than 2 (orange bars) were significantly increased in the same subjects.
Figure 2.
Linear Discriminant Analysis of the genera identified in supragingival biofilm samples from children with active caries, compared to caries-free children. Genera with log fold-change values lower than 2 (green bars) were considered as significantly reduced in caries-active subjects, and genera with log fold-change values higher than 2 (orange bars) were significantly increased in the same subjects.
2.2. In vitro biofilm formation from caries-free and caries-active children
In vitro biofilms were generated using supragingival biofilm samples as inocula. Supragingival biofilms from either caries-free or caries-active children were inoculated in aerobic conditions in the Amsterdam Active Attachment biofilm model (AAA model) [26,33] in McBain medium and, in parallel, in McBain supplemented with 0.2% sucrose. The use of McBain medium permits the generation of biofilms containing up to 40% relative abundance of gram-negative taxa [27]. In contrast, the use of McBain supplemented with sucrose has been established as a successful in vitro setting for developing cariogenic polymicrobial biofilms [34].
Figure 3 shows the values of total attached biofilm mass derived from supragingival samples grown without Aii20J (untreated controls). Biomass values, as assessed with CV assay, were heterogeneous among different subjects in both culture media used (Figure 3A,B). Comparison of the samples grouped according to the oral health status of the subjects revealed no statistically significant differences between biofilm formation abilities of samples from caries-free and caries-active children in neither of the culture media used (p = 0,8412 for biofilms grown in McBain; p = 0,4143 for biofilms grown in McBain-sucrose). These results showed that interindividual differences had a greater influence on the in vitro biofilm formation abilities of the samples than the oral health status of the subject. This observation aligns with previous reports on in vitro biofilms obtained from saliva and subgingival samples [27]. Here, the initial investigation of the microbial composition of the inocula used to generate the biofilms did not reveal significant differences between subjects (Figure 1). Nevertheless, these microbial populations have different biofilm formation abilities upon in vitro cultivation (Figure 3). These results align with the increasing importance given to functional activities within microbial populations in addition to their bacterial composition [8,35]. Biomass quantification of biofilms grown in McBain ranged between 0.1 and 0.4 in absorbance (OD590nm) (Figure 3A), whereas biomass quantification of biofilms grown in McBain-sucrose was higher and ranged between 0.2 and 1.2 (Figure 3B). These results obtained with CV assay subscribe to those of Janus et al. [34]. Using the same culture media as in the present study, they described that saliva-derived biofilms cultivated in the presence of sucrose achieved higher growth values and rates than biofilms cultivated without sucrose [34].
2.3. Effect of Aii20J on supragingival-derived biofilms from caries-free and caries-active children
To evaluate the antibiofilm potential of Aii20J, biofilm mass values obtained in the presence of the enzyme were compared to that of untreated controls. Figure 4A,B displays values of biofilm quantification after treatment with Aii20J, represented as the percentage of biofilm mass achieved relative to untreated controls in the two culture conditions studied. Overall, biofilm mass was reduced in the presence of Aii20J. In biofilms grown in McBain, macroscopical differences were observed in 40% of caries-free biofilms and 33% of caries-active biofilms (Figure 4A). Additionally, statistically significant differences were observed for 20% of caries-free and 22% of caries-active biofilms (t-test) (Figure 4A). In biofilms grown in the presence of sucrose (Figure 4B), macroscopical differences were observed in 50% of caries-free and 55% of caries-active biofilms, and statistically significant differences were found in 40% of caries-free and 11% of caries-active biofilms (Figure 4B). The effect of Aii20J on the biofilms was further analyzed by grouping the samples according to the oral health status of the subjects. Wilcoxon tests comparing biomass values of untreated biofilms with biomass values after treatment with Aii20J revealed statistically significant differences upon Aii20J-treatment in the caries-free biofilms grown in McBain (inserts in Figure 4A), and both the caries-free and caries-active biofilms grown in McBain-sucrose (inserts in Figure 4B). Altogether, these results indicate that biofilms grown in the presence of sucrose respond to Aii20J with greater biomass reductions than those grown without sucrose. The selection of saccharolytic bacteria present in the initial supragingival biofilms is expected with the use of a culture medium supplemented with sucrose. Thus, the presence of Aii20J seems to have had a greater effect on these bacteria than on bacteria selected in the medium without sucrose.
To investigate the microbial composition of the biofilms that responded to the Aii20J treatment with the greatest biomass reductions, gDNA from biofilms grown in McBain-sucrose was extracted, and the 16S rRNA gene was sequenced. Figure S2 shows the most relatively abundant genera identified in the biofilms generated in the presence of sucrose, under exposure to Aii20J, and as untreated controls.
Overall, in vitro biofilms displayed a much lower diversity than the initial supragingival samples. The most abundant genus among the in vitro biofilms was Streptococcus, with 86.6 – 99.9% relative abundance, followed by Granulicatella and Neisseria, with 0 – 4% and 0 – 12% relative abundances, respectively (Figure S2). The exceptions to this tendency were biofilms Ca9 and Ca10, which comprised 33 – 58% Streptococcus relative abundance. In these samples, Veillonella was identified in a 16 – 56% relative abundance, and Neisseria was found in < 0.1 – 21% relative abundance (Figure S2). Overall, biofilms from caries-free samples comprised 2 – 6 identified genera (samples NCa1 – Nca3 and NCa7 – Nca10), with some samples reaching up to 24 genera (sample NCa5), whereas in most caries-active samples, 10 – 16 genera were identified (samples Ca1, Ca3 – Ca5, Ca8 – Ca10). When comparing the microbiota composition of untreated biofilms and biofilms exposed to Aii20J, no significant differences were found in caries-free or caries-active groups (data not shown).
Further analyses of the microbial composition at the species level were performed on biofilms that responded to the treatment with Aii20J with important reductions in biomass. The 16S rRNA gene of these samples was sequenced by PacBio, and the taxonomical assignment of these full-length sequences was analyzed. Surprisingly, among the amplicon sequence variants (ASVs) belonging to the genus Streptococcus, we observed a significant decrease of Streptococcus mitis in almost all samples treated with Aii20J (t-test, α = 0.05) (Figure 5). In parallel, the relative abundance of Streptococcus cristatus and Streptococcus salivarius increased in the same samples (Figure 5). Similar results have been reported by Muras et al. [12]; the use of Aii20J on polymicrobial biofilms dominated by streptococci resulted in a decrease in the relative abundance of Streptococcus vestibularis, while the relative abundance of Streptococcus oralis subsp. dentisani increased [12]. Interestingly, pure streptococci cultures are not affected by the QQ enzyme Aii20J [9]. This evidence suggests that streptococci might interact with intergeneric cues (possibly AHLs or AHL-like molecules) produced by other taxa present in the polymicrobial biofilms and that the function of these intergeneric cues changes upon Aii20J exposure, as suggested in a previous study performed in our laboratory [27]. Additionally, several studies have already pointed out a role of AHLs on gram-positive taxa [36,37].
Altogether, our results show that Aii20J significantly reduces biofilms that are vastly dominated by gram-positive taxa, specifically streptococci. The observed changes in biofilm behavior are probably due to the surrounding environment of the polymicrobial biofilms generated. These results further support the use of Aii20J as an antibiofilm strategy for the prevention and treatment of oral diseases and a potential alternative or adjuvant to antimicrobial therapy. Furthermore, our results point towards the potential interactions of gram-positive taxa and intergeneric cues produced in polymicrobial environments.
3. Materials and Methods
3.1. Subject recruitment and ethics statements
A cohort of ten caries-free children (female/male (F:M): 4:6, mean age: 8.7 ± 3.2 years), and ten caries-active children (F:M: 5:5, mean age: 7.8 ± 1.5 years) were recruited from a private practice in Santiago de Compostela (ES) between December 2021 and July 2022 (Table 1). Orthodontic appliances and the use of antibiotics up to one month prior to sampling were the exclusion criteria set for this study.
The investigation protocol 2021/495 for patient recruitment and sample handling, modified in November 2021, was approved by the Ethical Committee of Clinical Investigations of Galicia (Xunta de Galicia, ES). Written informed consent was obtained from the legal guardians of all participants.
3.2. Sample collection and growth conditions
Supragingival plaque samples were taken from ten caries-free children (codes NCa1 – NCa10) and ten caries-active children (codes Ca1 – Ca10) (Table 1) by scrapping with a curette the vestibular and lingual surfaces of four dental pieces. Both deciduous and permanent pieces were sampled. Supragingival plaque was stored in 1.5 mL of thioglycolate medium (Merck Millipore) at 4 °C and processed promptly. Samples (n = 20) were vortexed for 30 s, and 100 μL were set aside and frozen at -80 °C for gDNA extraction. The remaining volume was diluted 1:53 [38] in either McBain or McBain supplemented with 0.2% sucrose (McBain-sucrose). Samples were grown in the AAA model, and when necessary, Aii20J was added at 20 µg/mL. Biofilms were grown in aerobiosis at 37 °C for 24 hours, refreshing culture media and treatments at 12 hours.
3.3. Strains and culture conditions
Chromobacterium subtsugae CV026, formerly Chromobacterium violaceum CV026, was used in this study. C. subtsugae CV026 is able to respond to AHLs with acyl chains between 4 and 8 carbon atoms in length by producing a violet pigment but does not produce endogenous AHLs [39]. C. subtsugae CV026 was routinely plated in Luria Bertani (LB) with 25 μg/mL of kanamycin and grown at 30 °C.
3.4. Production and purification of the QQ enzyme Aii20J
The QQ enzyme Aii20J, an AHL-lactonase enzyme obtained from the bacterium Tenacibaculum sp. strain 20J [40,41], was obtained as previously described [42]. Briefly, protein expression of the recombinant E. coli BL21(DE3)pLysS containing Aii20J was induced when suspensions reached 0.6 OD600nm by the addition of 1 mM Isopropyl-D-thiogalatopyranoside (IPTG) followed by further overnight incubation at 22 °C with gentle orbital shaking. The induced cells were pelleted, resuspended in phosphate buffered saline (PBS), and then lysed by sonication. Imidazole was added at a final concentration of 20 mM to prevent non-specific binding during later purification steps. Purification of Aii20J was achieved using the His GraviTrap affinity column protein purification kit (GE Healthcare) following the manufacturer's instructions. Briefly, the columns have a nickel-based medium for the purification of histidine-tagged proteins by immobilized metal affinity chromatography. The first step was the equilibration of the column with a phosphate buffer (10 mM Na2HPO4·2 H2O, 10 mM Na2H2PO4·H2O, 500 mM NaCl) containing imidazole in the same concentration as in the sample (20 mM) to prevent binding of host cell proteins with exposed histidines to the nickel-based medium. After sample flow-through, a washing step was performed with the 20 mM-imidazole buffer. Last, elution of the affinity-bound protein was achieved with a 500 mM-imidazole buffer. Remaining imidazole was then removed from the eluted sample by dialysis with D-tubes (MWCO 10 kDa) (Merck Millipore) in sterile Milli-Q water.
The QQ enzyme Aii20J was added to the assays to a standardized working titer of 20 µg/mL. The AHL-degradation capacity of the enzyme was routinely verified using chromogenic assays (see section 3.5).
3.5. Quorum Quenching activity solid plate assay
Solid plate bioassays were used to assess the AHL-degradation activity of the QQ enzyme Ai20J, purified and in biofilm supernatants after incubation. Ten-fold serial dilutions of the purified enzyme were prepared until reaching a titer of 0.02 μg/mL of Aii20J. The established minimum active concentration (MAC) of Aii20J is 2 μg/mL [43]. For biofilm supernatants treated with Aii20J at a titer of 20 μg/mL, 100 μL were transferred to a fresh microtube at the end of the incubation time. Both the purified Aii20J and biofilm supernatants containing Aii20J were incubated with 10 μM of C6-HSL for 3 hours at 22 °C. PBS (pH 6.5) with 10 μM of C6-HSL was used as negative control [44]. The bioassays were carried out as described elsewhere [27].
3.6. Biofilm generation and quantification
In vitro biofilms derived from supragingival samples were generated using a modification of the AAA model [26,33], as previously described [27]. Briefly, custom stainless-steel lids for 12-well culture plates (VWR) were used to allow the insertion of glass coverslips (18×18 mm) (Menzel Gläser, Braunschweig, DE) [26,27]. The wells were filled with 3 mL of inocula and, when needed, Aii20J was added to the standardized titer of 20 µg/mL. Sterile Milli-Q water was added to negative control biofilms. After the incubation time, biofilms quantification was performed by CV assay [27]. After CV staining, biofilms were visually examined [27]. Absorbance of CV-stained biofilms was measured at 590 nm in a Multiskan SkyHigh (Thermo Scientific). Experiments were performed in triplicate.
3.7. Investigation of the microbial composition of biofilms
3.7.1. Genomic DNA extraction
Genomic DNA for sequencing analysis was obtained from initial supragingival biofilm samples and biofilms grown in the AAA model. For the latter, biofilms were harvested by 15 minute-sonication of the glass coverslips in 5 mL of sterile PBS. Genomic DNA extraction was done using the "DNeasy PowerBiofilm Kit" (Qiagen), following the manufacturer's instructions [27], and DNA concentration was measured using a Qubit 4 Fluorometer (Thermo Scientific).
3.7.2. Library preparation and microbiome analysis
The library preparation and sequencing were performed in the Foundation for the Promotion of Health and Biomedical Research of Valencia Region (FISABIO) (ES). Genomic DNA (5 ng/µL in Tris 10 mM pH 8.5) was used to amplify the V3-V4 hypervariable regions of the 16S rRNA gene. The libraries were prepared according to Illumina’s protocol and sequenced using a 2x300 base pair paired-end run on a MiSeq Sequencer. Quality assessment was performed using the program prinseq-lite [45]. The analysis and clustering of the sequences into amplicon sequence variants (ASVs) was done with the pipeline DADA2 [46]. Classification of the ASVs to the genus level was done with the SILVA database [47]. Computations and statistics were performed in R [48] using knitr, knitcitations, markdown, biostrings, and vegan [49,50,51,52,53].
A selection of biofilms that responded to the Aii20J treatment with reductions in their biomass was additionally used to sequence the full 16S rRNA gene in a PacBio Sequel II sequencer. Data derived from the sequencing were obtained using an ad-hoc pipeline written in RStatistics environment [48]. Sequence data were analyzed using the qiime2 pipeline [54].
A LDA of the identified taxa was performed with the packages Phyloseq and DESeq2 [55]. Only differences with a log fold-change, either below 2 or above 2, were considered relevant.
3.8. Statistical analyses
Statistical analyses were performed using GraphPad Prism 9.5.1 (GraphPad, San Diego, CA, USA, www.graphpad.com). The normality of the datasets' distribution was tested using Shapiro-Wilk tests. Two-tailed Student's t-tests (referred to in the text as t-tests) for normally distributed samples and Mann-Whitney tests for non-normally distributed samples were performed to determine the statistical significance of the differences between control and Aii20J-treated biofilms within the same subject. In addition, Wilcoxon tests were performed to assess the significance of the reductions in biofilm formation in the presence of Aii20J throughout samples originated from different subjects and grown in the same conditions. Significant differences were determined with an α = 0.05 for all the statistical analyses.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Principal Components Analysis (PCA) plots of the microbial composition at the species level of supragingival biofilms. Figure S2: Relative abundance at the genus level of in vitro biofilms obtained from supragingival biofilm samples.
Author Contributions
Conceptualization, Ana Otero; Formal analysis, Ana Parga and Sabela Balboa; Funding acquisition, Ana Otero; Investigation, Ana Parga and Sabela Balboa; Methodology, Ana Parga, Paz Otero-Casal and Ana Otero; Project administration, Ana Otero; Resources, Paz Otero-Casal and Ana Otero; Supervision, Ana Otero; Writing – original draft, Ana Parga and Ana Otero; Writing – review & editing, Ana Parga, Sabela Balboa, Paz Otero-Casal and Ana Otero. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Xunta de Galicia (ES), doctoral grant number ED481A-2019/194; and Instituto de Salud Carlos III (ES), PDTS project number DTS-21/00015. The APC was funded by Instituto de Salud Carlos III (ES) (PDTS project number DTS21/00015).
Institutional Review Board Statement
The study was conducted under the Declaration of Helsinki and approved by the Ethical Committee of Clinical Investigations of Galicia (Xunta de Galicia, ES) (protocol code 2021/495, approved on November 2021).
Informed Consent Statement
Informed consent was obtained from the legal guardians of all subjects involved in the study.
Acknowledgments
The authors thank Dr. Vanessa Blanc (DENTAID, ES) for her help and discussions regarding the sampling methodology and patient selection criteria.
Conflicts of Interest
The Aii20J enzyme used in this study is protected by the following patent: Otero, A., Romero, M., and Mayer, C. 2016. Peptide with quorum-sensing inhibitory activity, polynucleotide that encodes said peptide, and the uses thereof. PCT/ES2014/070569. The remaining authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Network, G.B.o.D.C. Global Burden of Disease Study 2019 (GBD 2019); Institute of Health Metrics and Evaluation (IHME): Seattle, 2020, 2020. [Google Scholar]
- Marsh, P.D.; Lewis, M.A.O.; Rogers, H.; Williams, D.W.; Wilson, M. Marsh and Martin's oral microbiology, 6th ed.; Elsevier: Edinburgh; New York, 2016; p. 261. [Google Scholar]
- Simón-Soro, A.; Mira, A. Solving the etiology of dental caries. Trends in Microbiology 2015, 23, 76–82. [Google Scholar] [CrossRef]
- Bradshaw, D.J.; Lynch, R.J.M. Diet and the microbial aetiology of dental caries: new paradigms. International Dental Journal 2013, 63, 64–72. [Google Scholar] [CrossRef]
- Sheiham, A.; James, W.P.T. Diet and Dental Caries: The Pivotal Role of Free Sugars Reemphasized. Journal of Dental Research 2015, 94, 1341–1347. [Google Scholar] [CrossRef]
- Boisen, G.; Davies, J.R.; Neilands, J. Acid tolerance in early colonizers of oral biofilms. BMC Microbiology 2021, 21, 45. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Nyvad, B. The Role of Bacteria in the Caries Process: Ecological Perspectives. Journal of Dental Research 2011, 90, 294–303. [Google Scholar] [CrossRef]
- Duran-Pinedo, A.E.; Frias-Lopez, J. Beyond microbial community composition: functional activities of the oral microbiome in health and disease. Microbes and Infection 2015, 17, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Muras, A.; Mayer, C.; Romero, M.; Camino, T.; Ferrer, M.D.; Mira, A.; Otero, A. Inhibition of Steptococcus mutans biofilm formation by extracts of Tenacibaculum sp. 20J, a bacterium with wide-spectrum quorum quenching activity. Journal of Oral Microbiology 2018, 10, 1429788. [Google Scholar] [CrossRef] [PubMed]
- Philip, N.; Suneja, B.; Walsh, L.J. Ecological Approaches to Dental Caries Prevention: Paradigm Shift or Shibboleth? Caries Research 2018, 52, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Muras, A.; Otero, A. Breaking Bad: Understanding How Bacterial Communication Regulates Biofilm-Related Oral Diseases. In Trends in Quorum Sensing and Quorum Quenching: New Perspectives and Applications, 1st ed.; Rai, V.R., Bai, J.A., Eds. CRC Press/Taylor & Francis Group: Boca Raton, 2020. [Google Scholar]
- Muras, A.; Otero-Casal, P.; Blanc, V.; Otero, A. Acyl homoserine lactone-mediated quorum sensing in the oral cavity: a paradigm revisited. Scientific Reports 2020, 10, 9800. [Google Scholar] [CrossRef] [PubMed]
- Parga, A.; Otero, A. Ecological modulation of oral biofilms: novel approaches to treat and prevent oral diseases. In Oral Biofilms in Health and Disease; Springer Series on Biofilms; Springer: Berlin/Heidelberg, Germany, in press. [Google Scholar]
- Cvitkovitch, D.G.; Li, Y.-H.; Ellen, R.P. Quorum sensing and biofilm formation in Streptococcal infections. The Journal of Clinical Investigation 2003, 112, 1626–1632. [Google Scholar] [CrossRef]
- Qi, F.; Kreth, J.; Lévesque, C.M.; Kay, O.; Mair, R.W.; Shi, W.; Cvitkovitch, D.G.; Goodman, S.D. Peptide pheromone induced cell death of Streptococcus mutans. FEMS microbiology letters 2005, 251, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Frias, J.; Olle, E.; Alsina, M. Periodontal Pathogens Produce Quorum Sensing Signal Molecules. Infection and Immunity 2001, 69, 3431–3434. [Google Scholar] [CrossRef] [PubMed]
- Rickard, A.H.; Palmer, R.J.; Blehert, D.S.; Campagna, S.R.; Semmelhack, M.F.; Egland, P.G.; Bassler, B.L.; Kolenbrander, P.E. Autoinducer 2: a concentration-dependent signal for mutualistic bacterial biofilm growth. Molecular Microbiology 2006, 60, 1446–1456. [Google Scholar] [CrossRef]
- Jakubovics, N.; Kolenbrander, P. The road to ruin: the formation of disease-associated oral biofilms: Formation of oral biofilms. Oral Diseases 2010, 16, 729–739. [Google Scholar] [CrossRef]
- Huang, R.; Li, M.; Gregory, R.L. Bacterial interactions in dental biofilm. Virulence 2011, 2, 435–444. [Google Scholar] [CrossRef]
- Guo, L.; He, X.; Shi, W. Intercellular communications in multispecies oral microbial communities. Frontiers in Microbiology 2014, 5. [Google Scholar] [CrossRef]
- Kumari, A.; Pasini, P.; Daunert, S. Detection of bacterial quorum sensing N-acyl homoserine lactones in clinical samples. Analytical and Bioanalytical Chemistry 2008, 391, 1619–1627. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.Y.; Tan, W.-S.; Khan, S.A.; Chew, H.P.; Kasim, N.H.A.; Yin, W.-F.; Chan, K.-G. Unusual multiple production of N-acylhomoserine lactones a by Burkholderia sp. strain C10B isolated from dentine caries. Sensors 2014, 14, 8940–8949. [Google Scholar] [CrossRef]
- Goh, S.-Y.; Khan, S.A.; Tee, K.K.; Abu Kasim, N.H.; Yin, W.-F.; Chan, K.-G. Quorum sensing activity of Citrobacter amalonaticus L8A, a bacterium isolated from dental plaque. Scientific Reports 2016, 6, 20702. [Google Scholar] [CrossRef] [PubMed]
- Aleti, G.; Baker, J.L.; Tang, X.; Alvarez, R.; Dinis, M.; Tran, N.C.; Melnik, A.V.; Zhong, C.; Ernst, M.; Dorrestein, P.C.; et al. Identification of the Bacterial Biosynthetic Gene Clusters of the Oral Microbiome Illuminates the Unexplored Social Language of Bacteria during Health and Disease. mBio 2019, 10, e00321–00319. [Google Scholar] [CrossRef] [PubMed]
- Janus, M.M.; Crielaard, W.; Zaura, E.; Keijser, B.J.; Brandt, B.W.; Krom, B.P. A novel compound to maintain a healthy oral plaque ecology in vitro. Journal of Oral Microbiology 2016, 8, 32513. [Google Scholar] [CrossRef] [PubMed]
- Muras, A.; Mayer, C.; Otero-Casal, P.; Exterkate, R.A.M.; Brandt, B.W.; Crielaard, W.; Otero, A.; Krom, B.P. Short-Chain N -Acylhomoserine Lactone Quorum-Sensing Molecules Promote Periodontal Pathogens in In Vitro Oral Biofilms. Applied and Environmental Microbiology 2020, 86, e01941–01919. [Google Scholar] [CrossRef] [PubMed]
- Parga, A.; Muras, A.; Otero-Casal, P.; Arredondo, A.; Soler-Ollé, A.; Àlvarez, G.; Alcaraz, L.D.; Mira, A.; Blanc, V.; Otero, A. The quorum quenching enzyme Aii20J modifies in vitro periodontal biofilm formation. Frontiers in Cellular and Infection Microbiology 2023, 13. [Google Scholar] [CrossRef]
- Eren, A.M.; Borisy, G.G.; Huse, S.M.; Mark Welch, J.L. Oligotyping analysis of the human oral microbiome. Proceedings of the National Academy of Sciences 2014, 111, E2875–E2884. [Google Scholar] [CrossRef]
- Segata, N.; Haake, S.K.; Mannon, P.; Lemon, K.P.; Waldron, L.; Gevers, D.; Huttenhower, C.; Izard, J. Composition of the adult digestive tract bacterial microbiome based on seven mouth surfaces, tonsils, throat and stool samples. Genome Biology 2012, 13, R42. [Google Scholar] [CrossRef]
- Belda-Ferre, P.; Williamson, J.; Simón-Soro, Á.; Artacho, A.; Jensen, O.N.; Mira, A. The human oral metaproteome reveals potential biomarkers for caries disease. Proteomics 2015, 15, 3497–3507. [Google Scholar] [CrossRef]
- Meurman, J.H. Probiotics: do they have a role in oral medicine and dentistry? European Journal of Oral Sciences 2005, 113, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Belda-Ferre, P.; Alcaraz, L.D.; Cabrera-Rubio, R.; Romero, H.; Simón-Soro, A.; Pignatelli, M.; Mira, A. The oral metagenome in health and disease. The ISME Journal 2012, 6, 46–56. [Google Scholar] [CrossRef]
- Exterkate, R.A.M.; Crielaard, W.; Ten Cate, J.M. Different Response to Amine Fluoride by Streptococcus mutans and Polymicrobial Biofilms in a Novel High-Throughput Active Attachment Model. Caries Research 2010, 44, 372–379. [Google Scholar] [CrossRef]
- Janus, M.M.; Keijser, B.J.F.; Bikker, F.J.; Exterkate, R.A.M.; Crielaard, W.; Krom, B.P. In vitro phenotypic differentiation towards commensal and pathogenic oral biofilms. Biofouling 2015, 31, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Solbiati, J.; Frias-Lopez, J. Metatranscriptome of the oral microbiome in health and disease. Journal of Dental Research 2018, 97, 492–500. [Google Scholar] [CrossRef]
- Qazi, S.; Middleton, B.; Muharram, S.H.; Cockayne, A.; Hill, P.; O'Shea, P.; Chhabra, S.R.; Cámara, M.; Williams, P. N-acylhomoserine lactones antagonize virulence gene expression and quorum sensing in Staphylococcus aureus. Infection and Immunity 2006, 74, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Parga, A.; Manoil, D.; Brundin, M.; Otero, A.; Belibasakis, G.N. Gram-negative quorum sensing signalling enhances biofilm formation and virulence traits in gram-positive pathogen Enterococcus faecalis. Journal of Oral Microbiology 2023, 15, 2208901. [Google Scholar] [CrossRef]
- Mira, A.; Buetas, E.; Rosier, B.; Mazurel, D.; Villanueva-Castellote, Á.; Llena, C.; Ferrer, M.D. Development of an in vitro system to study oral biofilms in real time through impedance technology: validation and potential applications. Journal of Oral Microbiology 2019, 11, 1609838. [Google Scholar] [CrossRef]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef]
- Romero, M.; Martin-Cuadrado, A.-B.; Roca-Rivada, A.; Cabello, A.M.; Otero, A. Quorum quenching in cultivable bacteria from dense marine coastal microbial communities. FEMS Microbiology Ecology 2011, 75, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Muras, A.; Mayer, C.; Buján, N.; Magariños, B.; Otero, A. In vitro quenching of fish pathogen Edwardsiella tarda AHL production using marine bacterium Tenacibaculum sp. strain 20J cell extracts. Diseases of Aquatic Organisms 2014, 108, 217–225. [Google Scholar] [CrossRef]
- Mayer, C.; Muras, A.; Romero, M.; López, M.; Tomás, M.; Otero, A. Multiple quorum quenching enzymes are active in the nosocomial pathogen Acinetobacter baumannii ATCC17978. Frontiers in Cellular and Infection Microbiology 2018, 8, 310. [Google Scholar] [CrossRef]
- Mayer, C.; Romero, M.; Muras, A.; Otero, A. Aii20J, a wide-spectrum thermostable N-acylhomoserine lactonase from the marine bacterium Tenacibaculum sp. 20J, can quench AHL-mediated acid resistance in Escherichia coli. Applied Microbiology and Biotechnology 2015, 99, 9523–9539. [Google Scholar] [CrossRef] [PubMed]
- Muras, A.; López-Pérez, M.; Mayer, C.; Parga, A.; Amaro-Blanco, J.; Otero, A. High Prevalence of Quorum-Sensing and Quorum-Quenching Activity among Cultivable Bacteria and Metagenomic Sequences in the Mediterranean Sea. Genes 2018, 9, 100. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nature Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research 2013, 41, D590–596. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A language and environment for statistical computing, 4.0.2; R Foundation for Statistical Computing: Vienna, 2022. [Google Scholar]
- Allaire, J.J.; Horner, J.; Marti, V.; Porte, N. markdown: Markdown rendering for R; 0.7.4; 2014.
- Boettiger, C. knitcitations: Citations for knitr markdown files, 1.0.5; 2014.
- Xie, Y. Dynamic documents with R and Knitr; CRC Press, Taylor & Francis: Boca Raton, FL, USA, 2014; p. 190. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O'Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community ecology package, 2.4-4; 2017.
- Pagès, P.A. Biostrings; Bioconductor: 2017.
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nature Biotechnology 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biology 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Relative abundance of the most abundant bacterial genera identified in the supragingival biofilm samples and Principal Components Analysis (PCA) plots of their microbiota structure. Supragingival biofilms were sampled from the vestibular and lingual surfaces from caries-free (n = 10) and caries-active children (n = 10). (A). The relative abundance of the most relatively abundant genera identified in the samples is represented on the y-axis. Genera detected in lower relative abundance are grouped in the category "Other". Supragingival samples from caries-free children (NCa) are shown on the left part of the x-axis, and samples from caries-active children (Ca) are on the right part of the x-axis. (B). The relative abundance of the genera grouped under the category "Other" identified in relative abundances higher than 0.5%. Genera detected in relative abundances lower than 0.5% appear again under the category "Other". (C). PCA plot of the 19 supragingival biofilm samples after removing sample Ca8 (n = 10 caries-free children, represented by yellow circles; n = 9 caries-active children, represented by blue circles).
Figure 1.
Relative abundance of the most abundant bacterial genera identified in the supragingival biofilm samples and Principal Components Analysis (PCA) plots of their microbiota structure. Supragingival biofilms were sampled from the vestibular and lingual surfaces from caries-free (n = 10) and caries-active children (n = 10). (A). The relative abundance of the most relatively abundant genera identified in the samples is represented on the y-axis. Genera detected in lower relative abundance are grouped in the category "Other". Supragingival samples from caries-free children (NCa) are shown on the left part of the x-axis, and samples from caries-active children (Ca) are on the right part of the x-axis. (B). The relative abundance of the genera grouped under the category "Other" identified in relative abundances higher than 0.5%. Genera detected in relative abundances lower than 0.5% appear again under the category "Other". (C). PCA plot of the 19 supragingival biofilm samples after removing sample Ca8 (n = 10 caries-free children, represented by yellow circles; n = 9 caries-active children, represented by blue circles).
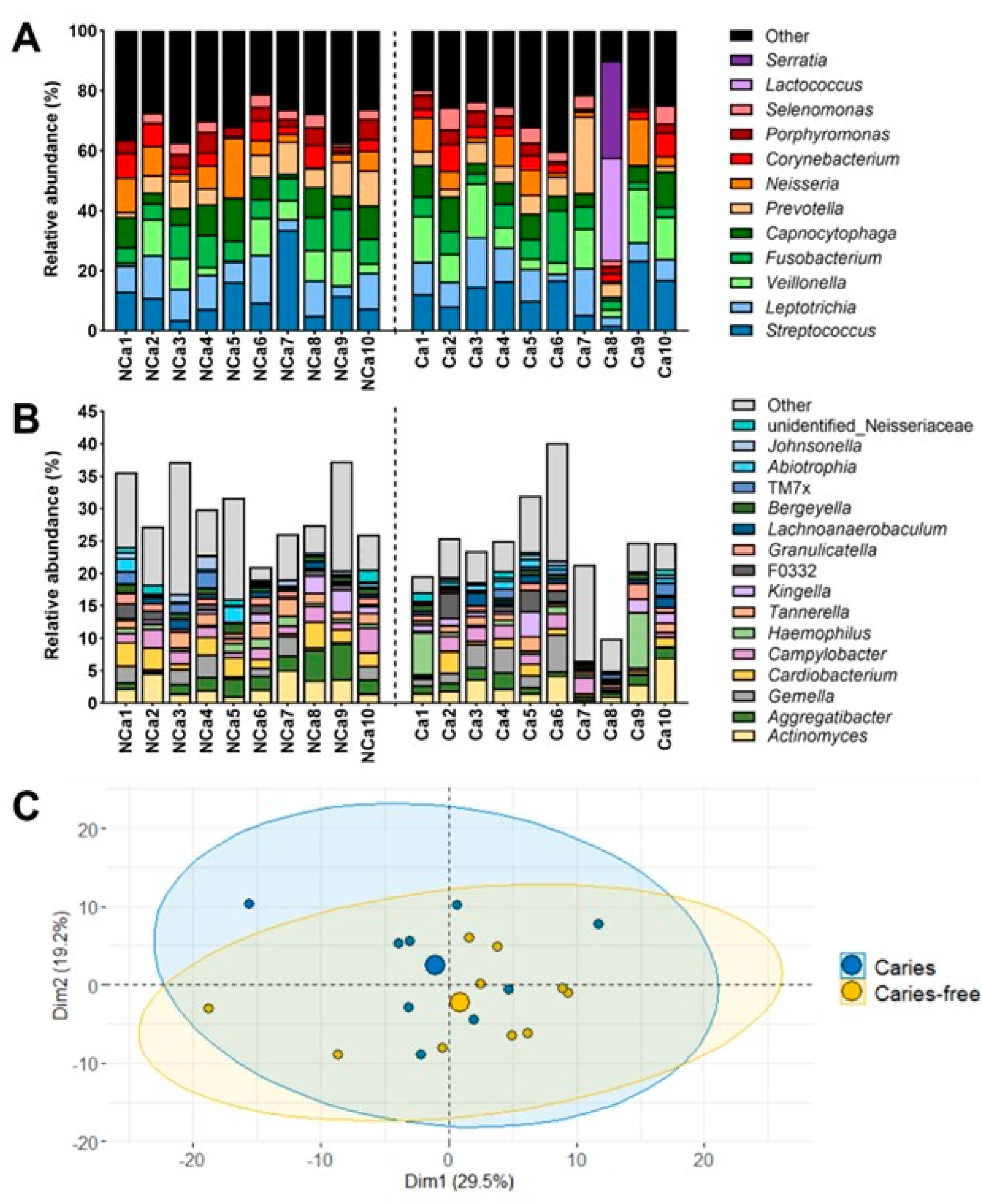
Figure 3.
Quantification of in vitro supragingival-derived biofilms. Histograms represent the biofilm mass achieved by each sample without the Aii20J treatment (untreated controls), assessed by CV assays (y-axis). Results are represented as the mean of OD590nm measurements, with error bars for standard deviation (n = 3). White bars represent biofilms from samples of caries-free children, whereas grey bars represent biofilms from samples of caries-active children (x-axis). Supragingival samples were grown in McBain medium (A) and McBain medium supplemented with 0.2% sucrose (B).
Figure 3.
Quantification of in vitro supragingival-derived biofilms. Histograms represent the biofilm mass achieved by each sample without the Aii20J treatment (untreated controls), assessed by CV assays (y-axis). Results are represented as the mean of OD590nm measurements, with error bars for standard deviation (n = 3). White bars represent biofilms from samples of caries-free children, whereas grey bars represent biofilms from samples of caries-active children (x-axis). Supragingival samples were grown in McBain medium (A) and McBain medium supplemented with 0.2% sucrose (B).
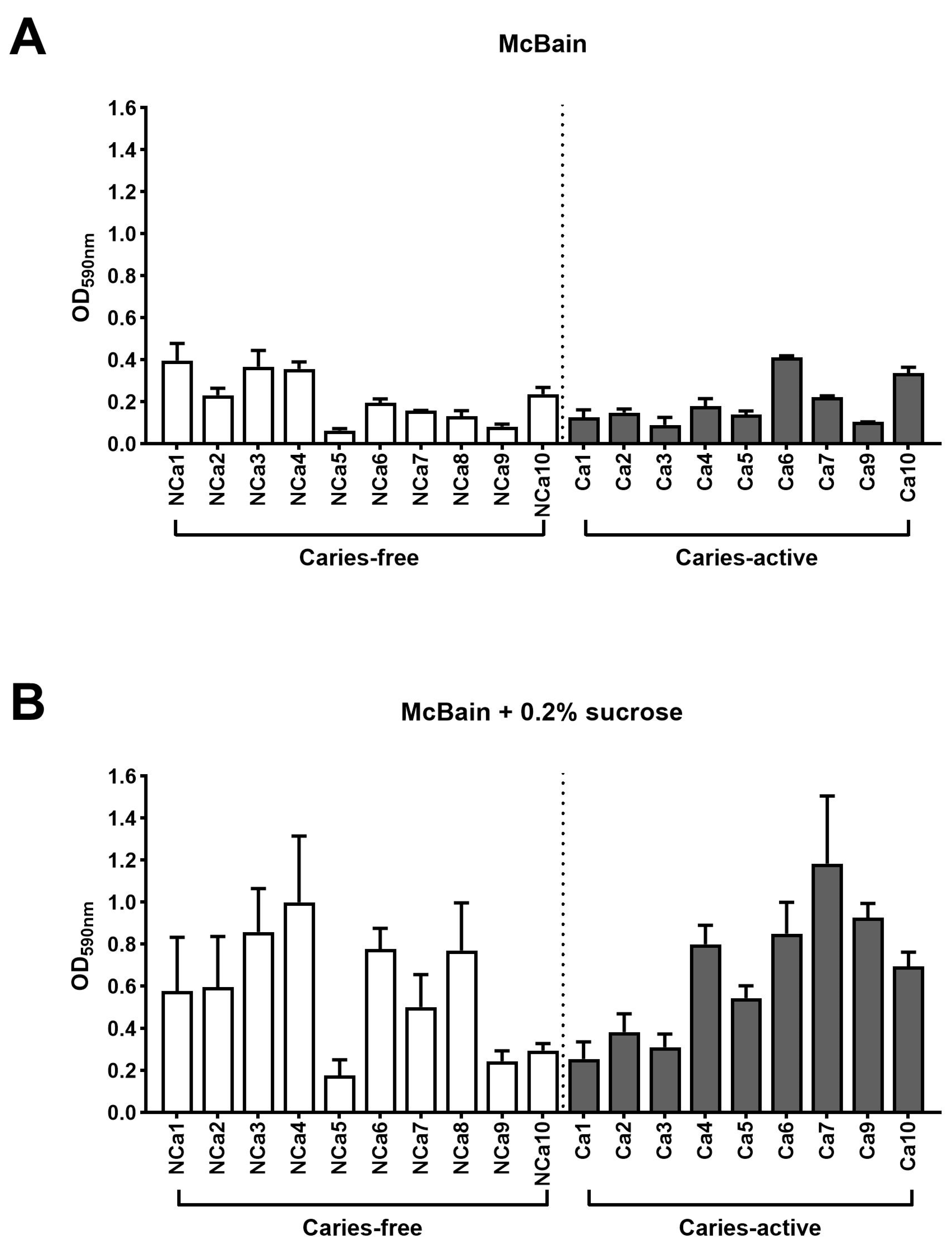
Figure 4.
In vitro biofilm formation of supragingival-derived biofilm samples in the presence of Aii20J. Histograms represent the biofilm mass achieved in Aii20J-treated samples as a percentage of the biofilm formed in untreated controls, assessed by CV assay (y-axis). Results are expressed as mean, with error bars for standard deviation (n = 3). Supragingival biofilm samples used to inoculate the in vitro biofilms were obtained from caries-free (light green bars) and caries-active children (dark green bars). Biofilms were generated in McBain medium (A) and McBain supplemented with 0.2% sucrose (B). Diamonds (♦) mark macroscopical differences between Aii20J-treated and untreated biofilms. Statistical significance of pairwise comparisons between each Ai20J-treated biofilm and its untreated control is displayed as *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001 (t-tests, α = 0.05). Boxplots are graphed with the median and interquartile range, with whiskers ranging from min. to max. of absolute values of biomass (OD590nm) in untreated and Aii20J-treated samples from caries-free (n = 10) and caries-active subjects (n = 9). Statistical significance of differences in biomass upon Aii20J treatment are marked with asterisks (*) (Wilcoxon tests, *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001).
Figure 4.
In vitro biofilm formation of supragingival-derived biofilm samples in the presence of Aii20J. Histograms represent the biofilm mass achieved in Aii20J-treated samples as a percentage of the biofilm formed in untreated controls, assessed by CV assay (y-axis). Results are expressed as mean, with error bars for standard deviation (n = 3). Supragingival biofilm samples used to inoculate the in vitro biofilms were obtained from caries-free (light green bars) and caries-active children (dark green bars). Biofilms were generated in McBain medium (A) and McBain supplemented with 0.2% sucrose (B). Diamonds (♦) mark macroscopical differences between Aii20J-treated and untreated biofilms. Statistical significance of pairwise comparisons between each Ai20J-treated biofilm and its untreated control is displayed as *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001 (t-tests, α = 0.05). Boxplots are graphed with the median and interquartile range, with whiskers ranging from min. to max. of absolute values of biomass (OD590nm) in untreated and Aii20J-treated samples from caries-free (n = 10) and caries-active subjects (n = 9). Statistical significance of differences in biomass upon Aii20J treatment are marked with asterisks (*) (Wilcoxon tests, *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001).
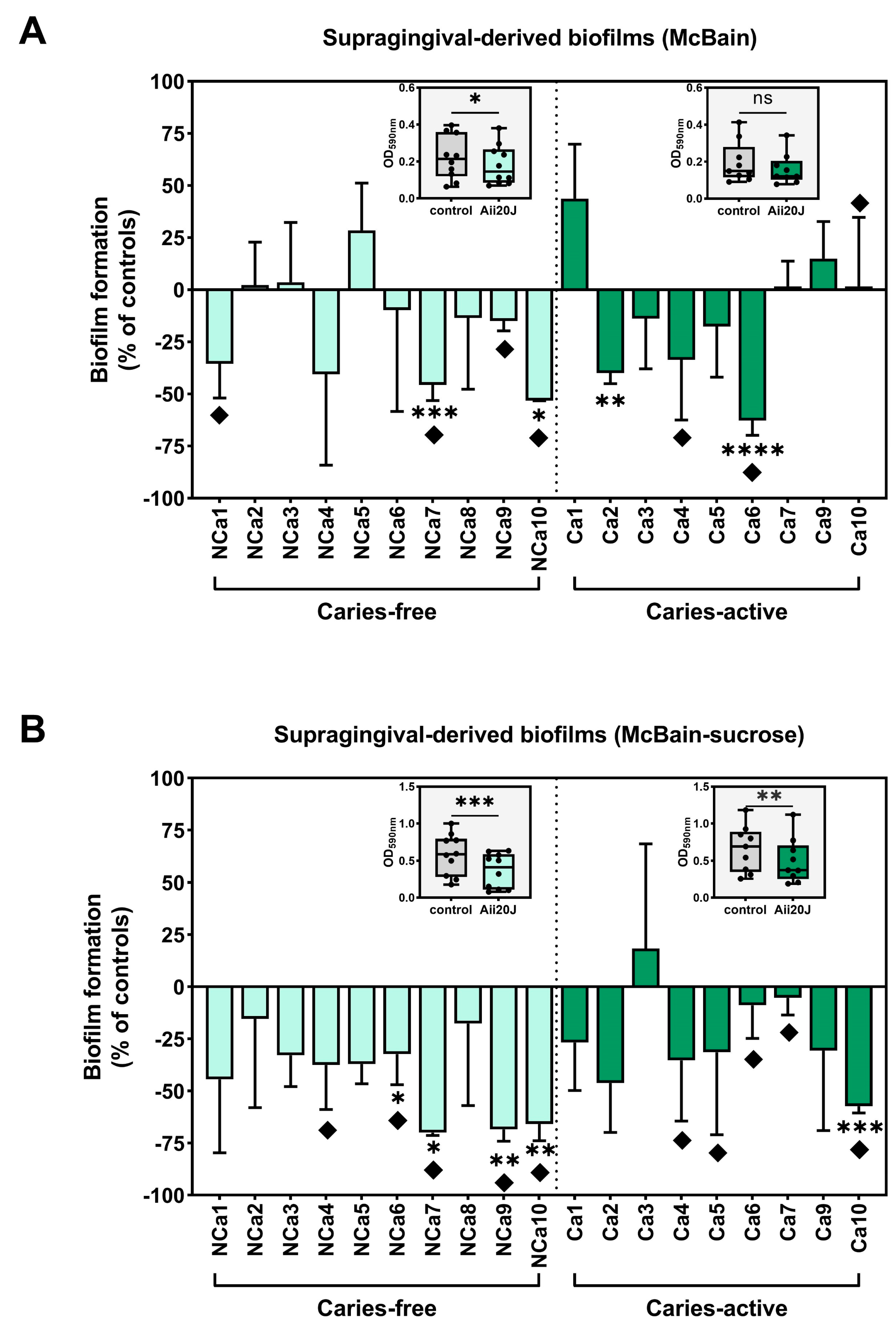
Figure 5.
Relative abundance of Streptococcus species in in vitro biofilms obtained from supragingival biofilm samples. Full-length 16S rRNA gene sequences were obtained by PacBio sequencing. Histograms show the relative abundance of the species identified in biofilms that were reduced in biomass upon exposure to the enzyme Aii20J.
Figure 5.
Relative abundance of Streptococcus species in in vitro biofilms obtained from supragingival biofilm samples. Full-length 16S rRNA gene sequences were obtained by PacBio sequencing. Histograms show the relative abundance of the species identified in biofilms that were reduced in biomass upon exposure to the enzyme Aii20J.
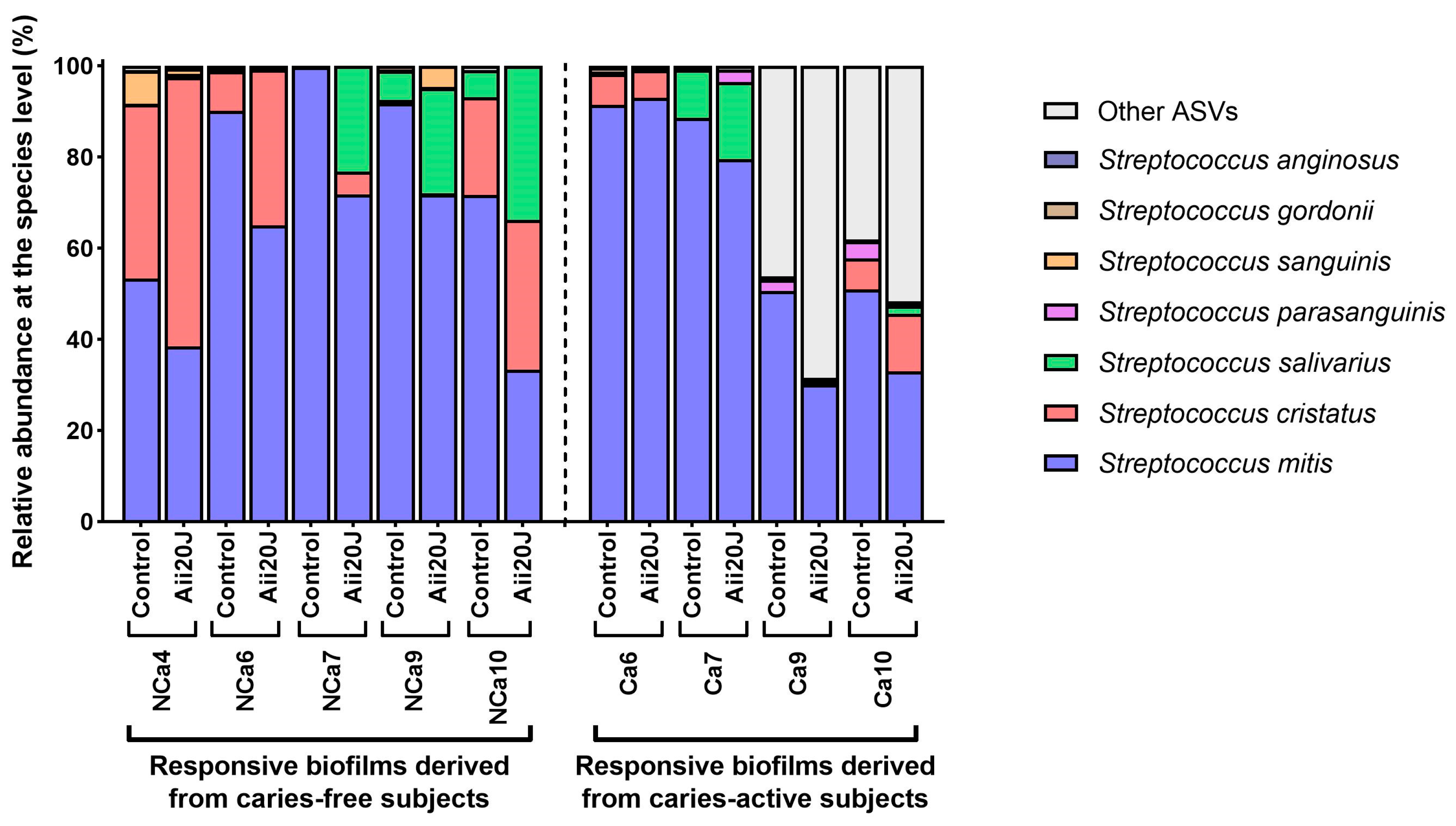

Table 1.
Classification of the oral health status of the subjects (n = 20). F: female. M: male. N/A: not available.
Table 1.
Classification of the oral health status of the subjects (n = 20). F: female. M: male. N/A: not available.
| Code | Age | Sex | Classification |
|---|---|---|---|
| NCa1 | 3 | M | Caries-free |
| NCa2 | 11 | F | Caries-free |
| NCa3 | 13 | M | Caries-free |
| NCa4 | 11 | M | Caries-free |
| NCa5 | N/A | M | Caries-free |
| NCa6 | 9 | F | Caries-free |
| NCa7 | 8 | F | Caries-free |
| NCa8 | 7 | M | Caries-free |
| NCa9 | 5 | F | Caries-free |
| NCa10 | 11 | M | Caries-free |
| Ca1 | 6 | M | Caries-active |
| Ca2 | 9 | M | Caries-active |
| Ca3 | 7 | F | Caries-active |
| Ca4 | 6 | M | Caries-active |
| Ca5 | 7 | M | Caries-active |
| Ca6 | 9 | M | Caries-active |
| Ca7 | 6 | F | Caries-active |
| Ca8 | 10 | F | Caries-active |
| Ca9 | 9 | F | Caries-active |
| Ca10 | 9 | F | Caries-active |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



