Submitted:
20 June 2023
Posted:
27 June 2023
You are already at the latest version
Abstract
In bilaterians organism, the signaling of pyramidal neurons (PyrNs) is linked to the relative (prevalent) molecular chirality and physiological, perceptual, cognitive, and psychological functions and dysfunctions, providing the coupling of the central nervous system downstream and upstream evolutionary and developmental processes. The most apparent and discriminating morphological specificity of PyrNs is the geometry of the cell body. However, the question "why/how PyrNs soma gains the shape of quasi-tetrahedral symmetry" has never been explicitly articulated. If the basic function of PyrNs is sensory space perception, supporting the orientation, and movement coordination, then the pyramidal shape of soma is the best evolutionary-selected geometry to perform sensory-motor coupling. In biology, the impact of chiral symmetry (handedness) is evident at all levels of biological organization, from the prevalent symmetry of biological molecules to the morphology and function of bilateral organisms. How the tetrahedral symmetry of biomolecules is linked to the morphology and functions of bilateral organisms remains a challenging question. Cell chirality represents an intermediate point connecting two poles of biochirality. In this holistic perspective, examining the PyrNs' morphology-circuitry-function link is crucial to understanding the complex interaction between genetic, epigenetic, and environmental factors in the evolution of the CNS. The predictive power of our hypothesis can be partially expressed by the statement that the most integral and reliable biomarker of the neurodegenerative (including aging) and mental (including all variants of psychiatric illness) disorders would be the detection of the hemispheric asymmetry of D-amino acids (D-AAs) level in pyramidal neurons (PyrNs).
Keywords:
prevalent molecular chirality
; bilaterality
; pyramidal neuron
; evolution
Introduction
Space-time symmetry and relativity are indispensable forms of existence, which can neither be created nor destroyed, but only transformed.
The association of biological processes with mental health and illness points to the reciprocal relations between two o opposite poles of the biological hierarchy, molecular biology and the phenomenon of the conscious mind. Biological psychology (biopsychology, psychobiology, psychophysiology, neuropsychobiology) and biological psychiatry represent the branches of science dealing with this fundamental link. The molecular and cell physiology of pyramidal neurons (PyrNs) provides a natural bridge between molecular biology and the appearance of human thoughts.
At the cellular level, biological processes within CNS are grounded in the physiology and function of PyrNs [1]. In biology, many legitimate questions still need appropriate answers. But some questions have not been ever expressed. One such inquiry is why/how PyrN, implicated in the spatial navigation of bilaterians, gain quasi tetrahedral symmetry of soma presumably associated with the geometry of chiral carbon atoms and tetrahedral structure of water (Figure 1) [2,3,4,5,6]. The question should be resolved based on the understanding of the evolution of CNS (Figure 1) [7]. Unfortunately, the geometrical definition of soma shape was not explicitly clarified. Consequently, whether the pyramidal shape of soma was evolutionarily selected, or the current state is “an appendix of the CNS” has never been debated. Tetrahedral (sp3-hybridized) carbons with four different substituents are the principal components of biological molecules exhibiting homochirality. Evolutionary selected biochemical homochirality is based on the chiral stereoselectivity of biosynthetic and metabolic reactions [8,9]. Chirality is the right keyword in the search for the integrity of PyrNs origin, structure, and functions. Such an assumption agrees that chiral asymmetry appears in nature at all levels, from elementary particles and amino acids to mammals’ morphology and even galaxies’ levels [10,11,12]. The origin of chirality at the cellular level is attributed to biomolecular homochirality [12]. Molecular homochirality and cellular chirality are the internal “forces” driving an organism’s left–right asymmetric development [13,14] in bilaterians.
The hierarchical chain of chirality transfer from the atomic to the organism level is one of the challenging targets of contemporary biological sciences [15,16,17]. The attention to the possible all-embracing role of the symmetry determinant in biology (from the biomolecular chirality to the bilaterality of human cognitive function) has become a significant trend in cognitive neuroscience [12,18,19,20]. However, surprisingly, in this chirality-oriented stream of biological research, the question “why/how does PyrNs gain tetrahedral geometry?” remains unarticulated. Below, we introduce some relevant hypotheses based on the generalized view of biological symmetry. A broad view of biological symmetry assumes that chirality transfer from the molecular to macromolecular and cellular levels plays a vital role in physiology underlying perceptual and cognitive functions linked with bilateral organisms’ emotional and behavioral expression (Figure 2) [11,12,21,22]. The cellular and molecular mechanisms of psychological states are fast-developing branches of life science. From the time of Freud, Jung, and Assagioli, perceptual, cognitive, and behavioral functions have been considered the essential determinants of psychological processes [23,24,25]. Regardless of the view on the relationship between psychology’s cognitive and behavioral domains, it is a commonly agreed that both exhibit a hierarchy, with the bottom referencing the basic sensory and perceptual processes and the top referencing the higher levels of cognitive and executive functioning control. From the top view, the hierarchical structure is the chain of downstream processes, including motor functions, perceptual abilities, neuronal circuits, and underlying molecular biology. Interpreting sensory perception and motor functions within cellular and molecular biology relies significantly on the laws of space-time symmetry [12,26,27,28,29]. Following this logic, the shape and functions of PyrNs deserve specific attention.
Molecular Chirality
The notion of chirality (i.e., symmetry) is closely related to the concept of space. Following the intuitive view of Euclid and Galilei. Newton concluded that the external to the observer space is infinite, isotropic, uniform, perfectly penetrable, and immovable, where he located the relative motion and studied its variation [30,31]. The frequently discussed mystery of biological chirality [32] arose from the combination of intuitive illusions. Most essential is the assumption regarding the existence of the racemic prebiotic world and the premise of the absolute isotropic and homogenous continuum of space. The false assumptions inevitably lead to the wrong unsolvable question. The collective labor of modern scientific society delivers evidence that space-time symmetry and relativity (STSR) are the fundamental properties of nature [17] and primary determinants of life [12,33]. The advances in quantum mechanics and group theory confirm the long-time-repining intuitive assumption, influencing the minds of the greatest scientists from Gailey to Dirac, that the principle of invariance (i.e., symmetry) is “analogous” to the relativity principle [34]. It has become a guiding idea in modern physical [35,36,37] and biological sciences [12,38,39]. Narrowing attention to the spatial symmetry at the molecular level, we can state that majority of biological molecules (such as amino acids and sugars) and macromolecules (including DNA, RNA, proteins, and lipids) exhibit the prevalent form of spatial symmetry in three domains of geometry: chirality, fractality, and topology [40,41,42,43,44]. We will be focused on the effects of molecular chirality represented by two classes of stereo-isomers: chiral enantiomers (prevalent L-isoform) and achiral diastereomers (prevalent cis-stereo-form) (Figure 3). The range of chirality-specific phase-transitions at the molecular level include chiral inversion and spontaneous chiral symmetry breaking in diverse states of aggregation. The most studied biological molecules implicated in the effects of chirality are amino acids (AAs) and enzyme-substrate protein complexes in the CNS [45,46,47,48]. The prevalent molecular chirality of organisms at the protein level occurs through the fine-balanced interaction of L-(major) and D-(minor) isoforms. Up-regulation of D-AAs is implicated in organism aging [47,49], psychiatric disorders [50], and cancers [51,52]. The most known adverse impacts are attributed to the up-regulation of three AAs: D-aspartic acid, D-serine, and D-alanine. All of them (and, potentially, by D-glutamate) are agonists for NMDA (N-methyl-D-aspartate) receptors of PyrNs [53,54,55,56,57,58]. Notably, continuous spontaneous and induced assembly of homochiral proteins from simple to complex arrangements includes the tetrahedral complexes [59] (well- known in the hemoglobin conformational dynamics) [60,61].
Link of Physiological and Psychological Functions
The central point of association between brain physiology and psychology is the function of PyrNs, providing an opportunity to basis of sensory processes (including visual, auditory, gustatory, olfactory, and cutaneous systems) to participate in the higher cognitive functions (such as emotion, learning, and memory) [62,63]. This functional opportunity is based on the interaction of sub-populations of PyrNs observed in low-level sensory cortices including, primary somatosensory, visual, and auditory cortex, with PyrNs located in high-level areas such as the prefrontal cortex (PFC). Such a conceptual view allows us to trace a link between chirality-dependent molecular events in PyrNs and their role in perceptual, cognitive, and behavioral aspects of psychological functions (PFs) Object perception or object recognition is the process of meaningful interpretation of the sensory input underlying our ability to act in the world. Psychophysical and physiological evidence suggests that object recognition relay on perceptual constancy, referring to the fact conservation of perceived geometrical characteristics of objects during relative motion of subject and object. Perceptual constancy is the invariance of injects under spatial transformations and is closely associated with the mathematical definition of symmetries. Therefore, it is not accidental that CNS of bilateral animals possess the specific neuronal mechanism of symmetry/dissymmetry perception [64,65,66]. Prevalent molecular chirality plays pivotal role in human physiology. It can be illustrated by the regulation of intra-body dynamics of D-amino acids (D-AAs). It is well known that D-serine is involved in bio-chemical interaction of main organs of the body, including kidneys, liver, gut, urine, stomal, lung, and brain [7,9,67]. The dependence of CNS on D-AAs metabolism allows us to trace the interplay between physiology and complex of perceptual, cognitive, and psychological functioning [21].
Analyzing the types of personalities, Jung differentiated them based on the complex cognitive functions, including thinking, filing, sensation, intuition, and aesthetical judgment [23]. This approach assumes significant overlapping between concepts of cognitive and psychological functions, with the last one covering the areas of emotions and behavior [24,68,69,70,71]. The expressions psychological functions (PFs), faculties, abilities, agents, states, conditions, and types are widely used in psychological sciences and to a lesser degree, in clinical practice.
Bilateral Organism: Symmetry-Function Interplay
The conjunction of cognitive and psychological functions allows projecting all aspect of molecular biology and space-perception of bilateral organism on the area of psychology and psychopathology [71,72]. Such a view brings us to the understanding that the bilateral CNS of humans unavoidably exhibits bilateral patterns of physiological, cognitive, emotional, and behavioral functions underlying the psychological state of individuals (in health and disease conditions). It means that every individual has a window for the spontaneous and intended inputs influencing the state of bilateral CNS. This is practically exploited in bilateral/unilateral activation of the sensory system and corresponding cognitive functions in the healing of post-traumatic stress disorders [73] and mediating fear conditions [74]. The symmetry in biology is extended to the balanced arrangement of body parts around the 3-D axis. The Bilateria or bilaterians are animals revealing bilateral symmetry (chirality) from the embryonic state. Neuroimaging studies reveal hemispheric asymmetry of association and limbic fiber tracts in utero [75,76]. The body plan of Bilateria has right and left sides that are mirror images of each other. Evolutionary origin of biochirality points to several major contributing factors. Three of them, relevant to the topic of our discussion, are prevalent molecular chirality, the necessity of space-time orientation of moving body with the possibility to distinguish left and right (we point to the left/right because it is related to a left-right mirror symmetry at the molecular level), a sensory system capable of space-time localization of objects representing food or social partner. According to fossil evidence, bilateral body symmetry in animals took place about 500 million years ago. In the course of evolution, bilateral bogy inevitably leads to bilateral CNS. This fact convinces many scientists that the laterality of brain functions cannot be accidental [77]. Symmetry in CNS is the balanced and co-related spatial arrangement of molecular, cellular, and anatomical components providing the opportunity for the laterality of cognitive functions. The laterality of cognitive functions experiences the “symmetry pressure” from two sides: from the internal determinants and behavioral-environmental determinants. One is strongly associated with the genetic mechanism, and another evidently with epigenetic factors [78]. Epigenetics represents the network of molecular interfaces mediating gene-environmental interactions [79]. In the language of symmetry, any epigenetic mechanism is the factor (symmetry changing factor) breaking genetically imposed homochirality of ribosomal protein synthesis [80]. Exploring human brain lateralization with attention to molecular chirality [21], cell chirality [81], genetics and non-genetic factors in association with the underlying physical laws concerning space-time symmetry confirms that the origin of relative chiral homogeneity of biological molecules is somehow connected to the origin and evolution of life [10,19,81]. For moving bilateral organisms, persistent asymmetry in the activation of the sensory system (as the vision in the birds) becomes the environmental factor of the development of perception, cognition, and action, and, consequently, the object of intuitive attraction (attention). In humans, this attraction takes various shapes, from pragmatic skills of spatial orientation to aesthetic feeling. At the cellular level molecular chirality is known as the factor promoting neuronal proliferation [82]. Space-time perception is the most influential force driving the direction of individual organism development and the directionality of biological evolution [10,83]. Both ontogeny and phylogeny can be traced at the molecular, cellular and organism levels. In the case of humans, the interplay of prevalent molecular chirality and the sensory-motor functions is what drives evolution. Hence, the sensory-motor system’s mechanism supports higher cognitive abilities, consciousness, and psychological state. The human brain is the integrative bilaterally asymmetric machinery of space-time perception of moving organisms accompanied by executive function. In the CNS, the link of molecular chirality to the organism’s function at the cell level is mediated by multiple subtypes of neurons with unique morphologies, electrical properties, and molecular identities [84].
Pyramidal Neurons
PyrNs gain their name due to the easily recognizable geometrical shape of the soma. PyrNs soma shape is sharply distinct from other neuronal (Purkinje and granular cells) and non-neuronal (microglia, astrocytes, epithelial, or red blood cell) cell types. Based on this fact, it is reasonable to assume that the specificity of PNs functions accounts for this difference. The term PyrNs refers to all major classes of excitatory multipolar glutamatergic cell types sharing common (pyramidal-like) soma shape despite the different degree of deviation from the perfect geometrical form [85]. The detailed description of the PyrNs’ soma shape is rare and usually characterized as a “teardrop or rounded pyramid” [86], extremely elongated rod-shaped” [87], and mst frequently pyramid-shaped [88]. The base geometry of the pyramid (triangular or square) is never experimentally characterized or theoretically predicted. In such a situation, it is relevant to say that PyrNs soma has relative similarity with the pyramid’s geometry in general or tetrahedron, in particular. The possibility of a tetrahedral shape can be considered an evolutionary tendency partially deviated by impacts of the factors secondary to the sensory perception of the spatial environment. The genetic architecture of the structural left-right asymmetry of the bilateral human brain is evolutionary preserved for the optimum function of sensory perception [89]. At the cellular level, space-time perception occurs through the activity of PurNs. The functions of PyrNs includes the evaluation of the distance, direction, and left-right discrimination of movement. In agreement with this view, the spatial arrangement of the PyrNs firing was found to be affected by environmental geometry [90]. Notable, that the aging-related cognitive deficits are attributable to reduced activity of pyramidal neurons [91]. Left–Right discrimination of brain functions is well represented in the behavioral study [92,93]. This knowledge is supported by new results showing brain asymmetry at the molecular and cellular levels. but not sufficiently explores at the molecular and cellular levels. PyrNs are the primary excitatory multipolar cell type abundant in the brain cortex [94], hippocampus [95,96], and amygdala [97]. The heterogeneity of the PyrN family is defined by their distinct axonal projections, dendritic arborization, and types of receptors. The cortical circuit network predominantly comprises pyramidal-to-pyramidal neuron connections, yet their assembly during embryonic development has yet to be entirely understood. PyrNs of cortex are distributed in the sensory, motor, association, and executive areas and found in all cortical layers except layer I [1,98,99]. Each PyrN receives input from thousands of excitatory synapses segregated onto dendritic branches. It has been previously proposed that sophisticated neuronal circuits associated with non-linear properties of dendrites enable cortical neurons to recognize multiple independent patterns and robust sequence memory [100]. Dysfunctional PyrN circuitry has been associated with abnormalities of perception, cognition, and psychological condition. PyrN utilize as neurotransmitters amino acids L-glutamate [101] and D-serine [102]. The most significant contribution to the morphological asymmetry of brain hemispheres and left-right differentiation of neural pathways is attributed to PyrN. The fact that PyrN are the most populated neuronal type in the human cerebral cortex and hippocampus suggests their primary role in processing space-time information utilized in sensory-motor functions. PyrN signaling is necessary for normal development and essential functions of mature organisms [103]. While the distortion of neuronal geometry and formation of aberrant synapses are associated with pathological conditions [104], including impairment of visual perception [105] and mental retardation [106]. The bilateral cortex and hippocampus, containing the majority of PyrN, are studied in brain regions involved in a wide range of hemisphere-specific functions, including spatial coding, navigation, spatial memory, decision-making [107,108,109], and intelligence [110]. The evolutionary selected system for space-time information processing, including the morphology and spatial orientation of PyrN, is the fundamental feature underlying the function of CNS. Experimentally observed hemispheric asymmetry of synaptic morphology of Pyr Ns explains well-known functional laterality of human perceptual and cognitive functions [111,112,113,114,115].
The morphology of PyrNs concerning the function was the focus of long-term attention in neuroscience. The major studied structural features were dendritic arborization, synaptic connectivity, and axonal network [84,114,115,116]. Apical and basal segments of the dendritic tree, complemented by the relative orientation of presynaptic and postsynaptic neurons, were carefully studied [5,6,94,116,117,118,119,120,121,122]. Two dendritic arbors have distinct morphology, orientation and are involved in different synaptic circuits [94]. The dendritic orientation of PyrN is sublayer specific and exhibits dorsal-verbal and front-back differentiation [123]. The experimental parameters characterizing the cell body include soma size, spatial distributions, the density of soma, and pyramidal somatic integrative zones. The shape and spatial orientation of pyramidal soma has little attention, partly due to the void of reliable experimental control.
In bilateral organisms, beginning from C elegans, CNS contains PyrNs. During neurogenesis from the ventricular zone (VZ), before adopting pyramidal morphology, neurons pass through several intermediate stages (multipolar, bipolar) [124,125].
Presumably, many molecular correlates contribute to the pyramidal shape of soma, but all cell morphology alterations are supported/assisted by the molecular dynamics of homochiral enzyme-substrate complexes. A commonly accepted axiom is that molecular chirality drives cell chirality [126]. Experimental evidence suggests that such a chirality transfer occurs with the participation of diverse cytoskeleton-based long-lived structures. Indeed, it was shown that the interaction of the diverse family of cytoskeleton filaments (septin [127], actin [128,129], microtubules [130,131], and that neurofilaments (NF) [132] provides fundamental cell morphogenetic mechanisms, including the shape and spatial orientation of PyrNs soma [129]. It was shown that neurofilaments (NF), and other intermediate-filament proteins contain motifs in their N-terminal domains that bind unassembled tubulin. Peptides containing such motifs inhibit microtubules’ in vitro polymerization, leading to altered cell shapes [133]. This fact suggests that NF-microtubules interaction can contribute to the shape of pyramidal neuron soma. The homochirality of actin-myosin cytoskeletons allows the cells to develop polarity and left-right asymmetry [126]. However, the bidirectional impact of prevalent molecular chirality (internal determinant) and bilaterality of CNS (window to the external epigenetic factors) on the pyramidality of PyrNs soma has never been considered.
Notable that dendritic arborization exhibits cortical layer-dependent orientation preference towards the anterior orientation [120]. However, the spatial orientation of pyramidal soma in two brain hemispheres and their relation to space-time information processing have yet to be experimentally studied or theoretically discussed. Based on the lateralization of perceptual and cognitive functions, we can expect differential bilateral asymmetry in the morphology and orientation of PyrN. Indeed, currently, the hemispheric difference is experimentally observed in the number/volume [134] and synaptic organization [135]. Asymmetric hemispheric allocation of NMDA receptor subunits in hippocampal PyrN complements the whole picture [95,96]. The fact that PyrNs of the healthy human brain have a significantly greater density, larger size, and are more spherical in shape on the left- than on the right-side point to the meaningful link between two kinds of biological events [136]. Notable that bilateral asymmetry of brain activity indicates a state of the CNS system concerning mood and anxiety. For example, studies of brain EEG associated with PyrNs firing suggest that high levels of beta in the right hemisphere are associated with anxiety symptoms. In contrast, high levels of alpha in the left hemisphere indicate depressive features [136,137,138].
Conclusion
Our hypothesis brings highlights several formal consequences of two co-existent events: PyNs’ shape and bilateral localization [139]. Indeed, episodic memory, allowing mental navigation in space and time, is based on the hemisphere-asymmetrical activity of hippocampal PyrNs [108] At the same time, from a geometrical standpoint, PyN soma (with four non-equivalent vertexes) represents a highly asymmetric (chiral) tetragonal structure. The anatomical symmetry and asymmetry of the PyrN structure imposes fundamental information processing capabilities. The essential point is that two mirror images of pyramidal neurons (two seemingly identical PyrNs) are not superimposable (i.e., not identical). Furthermore, the chirality of PyrN provides a formal opportunity for hemisphere-specific information processing. Organic and non-organic molecular nano-scale complexes can spontaneously adopt various shapes, including square-pyramidal [140], triangular-prism [141,142], octahedral, icosahedral., and tetrahedral [143,144] geometry. The stability of such DNA, proteins, and lipids higher-order nano-scale structures [145,146] suggests the existence of molecular contribution to the shape of PyrNs. Notably, the common symmetry principles guide the external and internal determinants of cell morphology, assembly, and functions of intracellular molecules. Protein aggregate adopting tetrahedral symmetry creates shell-like architectures [147]. Such structures serve as enclosures for viral genomes or as internal determinants of cell shape. Another well-known example of tetrahedral protein assembly is globular hemoglobin (Hb) [148,149] (Hb is a highly conserved globular protein in all life forms and functionally tied to aerobic organisms utilizing oxygen from the atmosphere and delivering it to cells. The expression of mitochondrial hemoglobin (Hba-a2 and Hbb) in neurons (including nigral dopaminergic neurons, striatal γ-aminobutyric acid (GABA-ergic neurons, and cortical pyramidal neurons) was experimentally observed [150,151,152]). Presumably, the shape of the soma can adapt the demands of the cytoskeleton’s tetrahedral mesh-like gel, undergoing a chain of structural phase transitions [144,153,154]. However, additional experimental verification of such pathways is required. The co-appearance of all the above-mentioned spatial arrangements of PyrNs in the visual cortex suggests that the corresponding primary function is the involvement in the space-time orientation of moving organisms. In neuronal cells, the assembly of cytoskeleton proteins can be considered an internal determinant of soma’s shape.
The mystery is almost solved, and what remains now is the need for complex, goal oriented experimental evidence, including illuminating the link between molecular chirality (such as of L/D-actin cytoskeleton dynamics in dendritic spines or chirality of mGluR7 receptors/ligands complex) and the shape of PyrNs. We have introduced you to the spectrum of seemingly unrelated facts and thoughts, the fundamental link of which is currently hidden from the broad research audience. Now it is time for your own judgment regarding the hypothesis on the consideration and for a new experimental design.
The evolution of bilaterian CNS occurs through the differentiation of cell types and network organization. A line of phylogenomic evidence suggests that some organisms (such as sponges and placozoans) may benefit from the loss and/or modification of their neural cell types. But the mainstream of evolution demonstrates the benefit of the CNS for managing voluntary movement (events in the space-time domain). At the cellular level, the origin of CNS is characterized by the appearance of pyramidal neurons (PyrNs). Learning the congruence of the molecular and cellular events helps to understand the link between voluntary movement, space perception CNS, and PyrNs morphology.
Biologists have made long-term efforts to explain the common origin of the homochiral worth of DNA code and protein sequence resulting in the emergence of the human mind. Quite unexpectedly, for many scientists, it was realized that the Ariadne thread is the concept of symmetry [11,12]. On the other hand, all theories ignoring the symmetry principle occurred to be irrelevant. Indeed, the evolutionary history of life exhibits the chain of successive events linked by the apparent common association with the notion of symmetry. The four most significant are:
(I) the appearance of prevalent molecular chirality,
(II) bilateral organisms,
(III) pyramidal neyrons (3), and
(IV) the handedness of the human mind.
Events I, II, and IV are the long-term concern of the scientists. The appearance and evolution of PyrN (event III) attracts increasing attention but does not focus on the specific geometric shape of soma.
Author Contributions
Main contribution is done by Dyakin V.V.
Acknowledgments
The author gratefully thanks Nika Dyakina-Fagnano, for her significant contribution to text clarification (especially in the psychology-related parts) and Pamela Butler, Stephen Ginsberg, and Csaba Vadasz for the valuable suggestions and corrections.
Conflicts of Interest
The authors declare no conflict of interest. Main contribution is done by Dyakin V.V.
Abbreviations
Amino acids (AAs). Electroencephalogram (EEG). Pyramidal neurons (PyNs). Central Nervous system (CNS). Psychological functions (PFs). Space-time symmetry and relativity (STSR).
References
- Betz, W. Distinction of two nerve centers in the brain. Q J Microsc Sci 1875, 15, 190–192. [Google Scholar]
- Dahanayake, J.N. and Mitchell-Koch, K.R. Entropy connects water structure and dynamics in protein hydration layer. Phys.Chem.Chem.Phys., 2018, 20, 14765. [Google Scholar] [CrossRef] [PubMed]
- Marvan, T.; Polák, M.; Bachmann, T. and Phillips, W.A. Apical amplification—a cellular mechanism of conscious perception? Neuroscience of Consciousness, 2021, 2021(2), 2021. [CrossRef]
- Kumar, P.; Buldyrev, S.V. and H. Eugene, H. A tetrahedral entropy for water. PNAS. 2009, 106 (52) 22130-22134. [CrossRef]
- Wang, Y.; Ye, M.; Kuang, X.; Li, Y. and Hu, S. A simplified morphological classification scheme for pyramidal cells in six layers of primary somatosensory cortex of juvenile rats. IBRO Rep. 2018, 74–90. [CrossRef]
- Luine, V. and Frankfurt, M. Interactions between estradiol, BDNF and dendritic spines in promoting memory. Neuroscience. 2013, 239, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.M.M.; Meyer, K.A.; Santpere, G.; Gulden, F.O. and Sestan, N. Evolution of the Human Nervous System Function, Structure, and Development. Cell. 2017 Jul 13; 170(2): 226–247. [CrossRef]
- Mason, S.F. The development of concepts of chiral discrimination. Chirality 1989, 1(3), 183–191. [Google Scholar] [CrossRef]
- Bastings, J.J.A.J.; van Eijk, H.M.; Damink. S.W.O and Sander S. Rensen, S.S. D-amino Acids in Health and Disease: A Focus on Cancer. Nutrients 2019, 11(9), 2205. [CrossRef]
- Kondepudi, D. Chiral Asymmetry in Nature, Ch. 1 in Chiral Analysis (Second Edition). Advances in Spectroscopy, Chromatography and Emerging Methods 2018, pg. 3-28. [CrossRef]
- Cristadoro, G., Degli Esposti, M. & Altmann, E.G. The common origin of symmetry and structure in genetic sequences. Sci Reports 2018, 8, 15817. [CrossRef]
- Dyakin, V.V. Fundamental Cause of Bio-Chirality: Space-Time Symmetry—Concept Review. Symmetry 2023, 15(1), 79. [Google Scholar] [CrossRef]
- Inaki, M.; Liu, J. and Matsuno, K. Cell chirality: its origin and roles in left–right asymmetric development. Philos Trans R Soc Lond B Biol Sci. 2016, 371(1710). [CrossRef]
- Lin, YM. Creating chirality. Nat Chem Biol. 2008, 4, 330. [Google Scholar] [CrossRef]
- Takaoka, K.; Yamamoto, M.; and Hamada, H. Origin of body axes in the mouse embryo. Current Opinion in Genetics & Development 2007, 17(4}44-350. [CrossRef]
- Dyakin, V.V.; Lucas, J.; Dyakina-Fagnano, N.V.; Posner, E.P.; Vadasz, C. The Chain of Chirality Transfer as Determinant of Brain Functional Laterality. Breaking the Chirality Silence: Search for New Generation of Biomarkers; Relevance to Neurodegenerative Diseases, Cognitive Psychology, and Nutrition Science. Neurology and Neuroscience Research. 2017, 1(1),2. [CrossRef]
- Duan, P.; Cao, H.; Zhang, L.; Liu, M. Gelation induced supramolecular chirality: chirality transfer, amplification and application. Soft Matter. 2014, 10, 5428. [Google Scholar] [CrossRef]
- Vallortigara, G. The evolutionary psychology of left and right: costs and benefits of lateralization Dev Psychobiol. 2006 Sep;48(6):418-27. [CrossRef]
- Francks, C. Exploring human brain lateralization with molecular genetics and genomics Annals of The New York Academy of Sciences Issue: The Year in Cognitive Neuroscience. 2015, 1359, 1-13. [CrossRef]
- Stacho, M. and Manahan-Vaughan, D. Mechanistic flexibility of the retrosplenial cortex enables its contribution to spatial cognition. Trends in Neuroscience. 2022, 45(4), P284-296. 4. [CrossRef]
- Lee, T-W; Dolan, R.J.; and Critchley, H.D. Controlling Emotional Expression: Behavioral and Neural Correlates of Nonimitative Emotional Responses. Cerebral Cortex, 2008, 18(1), 104–113. [CrossRef]
- Yuan, J.; Lu, X.; Zhang, S.; Zheng, F.; Deng, Q.; Han, L. and Lu, Q. Molecular Chirality and Morphological Structural Chirality of Exogenous Chirality-Induced Liquid Crystalline Block Copolymers. Macromolecules 2022, 55, 5, 1566–1575. [CrossRef]
- [Jung. 1976] Book by C.G. Jung. Psychological Types (The Collected Works of C. G. Jung, Vol. 6) (Bollingen Series XX). (Part of the Jung’s Collected Works (#6) Series and Dzieła (#2) Series). Publisher: Princeton University Press. 1976. 1977.
- Assagioli, A. Dynamic Psychology and Psychosynthesis. Publisher: Psychosynthesis Research Foundation, inc 1958. Roberto Assagioli.
- Myers, S. The five functions of psychological type. Analytical Psychology 2016, 61(2), 183–202. [Google Scholar] [CrossRef]
- Pillai, A.S. and Jirsa, V.K. Perspective. Symmetry Breaking in Space-Time Hierarchies Shapes Brain Dynamics and Behavior. Neuron 2017, 94(5), 1010-1026. [CrossRef]
- Jirsa, V. and Sheheitli, H. Entropy, free energy, symmetry and dynamics in the brain. Journal of Physics: Complexity 2022 3(1), 015007. [CrossRef]
- Dyakin, V.V. and Uversky, V.N. Arrow of Time, Entropy, and Protein Folding: Holistic View on Biochirality. Int J Mol Sci. 2022, 23(7):3687. [CrossRef]
- Iohnston, I.G.; Dingle,K.; Greenbury, S.F.; Camargo, C.Q.; Doye , J.P.K.; Ahnert, S.E. and Louis, A.A. Symmetry and simplicity spontaneously emerge from the algorithmic nature of evolution. Proc Natl Acad Sci U S A. 2022, 119(11), e2113883119. [CrossRef]
- Jammer, M. Concepts of Space: The history of Theories of Space in Physics, 3rd ed.; Dover: New York, NY, USA, 1993. [Google Scholar]
- Capecchi, D. Development of the Concept of Space up to Newton Encyclopedia 2022, 2(3), 1528-1544. [CrossRef]
- Zinovyev et al. Manifestation of Supramolecular Chirality during Adsorption on CsCuCl3 and γ-Glycine Crystals. Symmetry 2023,15(2):498. [CrossRef]
- Ocklenburg & Mundorf. Ocklenburg S. and Mundorf, A. Symmetry and asymmetry in biological structures Proc. Natl. Acad. Sci. U.S.A. PNAS. 2022, 119 (28), e2204881119. [CrossRef]
- Tasson, J.D. What Do We Know About Lorentz Invariance? Rep. Prog. Phys. 2014, 77, 062901. [Google Scholar] [CrossRef]
- Comte, C. Symmetry, relativity and quantum mechanics. Nuov Cim B 1996, 111, 937–956. [Google Scholar] [CrossRef]
- Field, J.N. Space-Time Exchange Invariance: Special Relativity as a Symmetry Principle. American Journal of Physics 2001, 69, 569. [Google Scholar] [CrossRef]
- Ajaltouni, Z.J. Symmetry and relativity: From classical mechanics to modern particle physics. Natural Science. 2014, 6(4), Article ID:43343,7 pages. [CrossRef]
- Auffray, C. and Nottale, L. Review. Scale relativity theory and integrative systems biology: 1: Founding principles and scale laws. Progress in Biophysics and Molecular Biology 2008, 97(1), 79-114,. [CrossRef]
- Noble, R.; Tasaki, K.; Noble, P.J. and Noble, D. Biological Relativity Requires Circular Causality but Not Symmetry of Causation: So, Where, What and When Are the Boundaries? Front. Physiol. Sec. Integrative Physiology 2019, 10. [CrossRef]
- Cornelissen, J.J.L.M.; Rowan, A.E.; Nolte, R.J.M. and Sommerdijk, N.A.J.M. Chiral Architectures from Macromolecular Building Blocks Chem. Rev. 2001, 101, 12, 4039–4070. [CrossRef]
- Todoroff, N.; Kunze, J.; Schreuder, H.; Hessler, G.; Baringhaus, K-H. and Schneider, G. (2014). Fractal Dimensions of Macromolecular Structures. Mol Inform. 2014, 33(9): 588–596. [CrossRef]
- Wang, J., Panagiotou, E. The protein folding rate and the geometry and topology of the native state. Sci Rep 12, 6384. [CrossRef]
- Pandey, S., Mandal, S., Danielsen, M.B. et al. Chirality transmission in macromolecular domains. Nat Common 2022, 13, 76. [CrossRef]
- Kim, N.H., Choi, H., Shahzad, Z.M. et al. Supramolecular assembly of protein building blocks: from folding to function. Nano Convergence 2022, 9(4). [CrossRef]
- Dhanavade, M.J. and Sonawane K.D. Amyloid beta peptide-degrading microbial enzymes and its implication in drug design. 3 Biotech. 2020, 10(6), 247. [CrossRef]
- Reetz, M.T. and Garcia-Borràs, M. The Unexplored Importance of Fleeting Chiral Intermediates in Enzyme-Catalyzed Reactions. J. Am. Chem. Soc. 2021, 143, 37, 14939–14950. [CrossRef]
- Dyakin, V.V.; Dyakina-Fagnano, N.V.; Mcintire, L.B. and Uversky, V.N. Fundamental Clock of Biological Aging: Convergence of Molecular, Neurodegenerative, Cognitive and Psychiatric Pathways: Non-Equilibrium Thermodynamics Meet Psychology. Int J Mol Sci. 2022, 23(1), 285. [CrossRef]
- Dyakin, V.V.; Wisniewski T.M. and Lajtha, A. Chiral Interface of Amyloid Beta (Aβ): Relevance to Protein Aging, Aggregation and Neurodegeneration. Symmetry 2020, 12(4), 585. [CrossRef]
- Fujii, N.; Takata, T.; Fujii,N.; Aki K. & Sakaue, H. D-Amino Acid Residues in Proteins Related to Aging and Age-Related Diseases and a New Analysis of the Isomers in Proteins Chapter in the book (pg 241-245) by Yoshimura, T., Nishikawa, T., Homma, H. (eds) D-Amino Acids Springer, Tokyo. [CrossRef]
- Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Kitagishi,Y. and Matsuda, S. Omar Cauli, Academic Editor and Soraya L. Valles, Academic Editor D-Amino Acids as a Biomarker in Schizophrenia. Diseases. 2022, 10(1), 9. [CrossRef]
- Jacco J.A.J. Bastings, Hans M. van Eijk, Steven W. Olde Damink,and Sander S. Rensen. D-amino Acids in Health and Disease: A Focus on Cancer. Nutrients. 2019, 11(9): 2205. [CrossRef]
- Murtas, G. and Pollegioni, L. D-Amino Acids and Cancer: Friends or Foes? Int. J. Mol. Sci. 2023, 24(4), 3274. 4. [CrossRef]
- Wolosker, H., Balu, D.T. D-Serine as the gatekeeper of NMDA receptor activity: implications for the pharmacologic management of anxiety disorders. Transl Psychiatry 2020 10, 184. [CrossRef]
- Li, Y.; Han, H.; Yin, J.; Li, T.and Yina, Y. Role of D-aspartate on biosynthesis, racemization, and potential functions: A mini-review. Anim Nutr. 2018, 4(3): 311–315. [CrossRef]
- Saitoh, Y.; Katane, M.; Miyamoto, T.; Sekine, M.; Sakai-Kato, K. and Homma, H. D-Serine and D-Alanine Regulate Adaptive Foraging Behavior in Caenorhabditis elegans via the NMDA Recepto,r Journal of Neuroscience 2020, 40 (39) 7531-7544. [CrossRef]
- Kera, Y.; Aoyama, H.; Matsumura, H.; Hasegawa, Hisae Nagasaki, H. and Yamada, R. Presence of free d-glutamate and d-aspartate in rat tissues. Biochimica et Biophysica Acta (BBA) 1995, 1243(2), 282-286. [CrossRef]
- Mangas, A.; Coveñas,R.; Bodet, D.; Geffard, M.; Aguilar, L.A. and Yajeya, J. Immunocytochemical visualization of d-glutamate in the rat brain. Neuroscience 2007, 144(2), 654-664. [CrossRef]
- Lin CH, Yang HT, Lane HY. d-glutamate, d-serine, and d-alanine differ in their roles in cognitive decline in patients with Alzheimer’s disease or mild cognitive impairment. Pharmacol Biochem Behav 2019,185: 172760. [CrossRef]
- Luo, Q.; Hou, C.; Bai, Y.; Wang, R. and Liu, J. Protein Assembly: Versatile Approaches to Construct Highly Ordered Nanostructures. Chem. Rev. (American Chemical Society) 2016, 116, 22, 13571–13632. [CrossRef]
- Riccio, A.; Vitagliano, L.; di Prisco, G.; Zagari, A. and Mazzarella, L. The crystal structure of a tetrameric hemoglobin in a partial hemichrome state. Proc Natl Acad Sci U S A. 2002, 99(15), 9801–9806. [CrossRef]
- Ha, C-E. and Bhagavan, N.V. Hemoglobin Chapter in Essentials of Medical Biochemistry. eBook ISBN: 9780124166974. Sec Ed. Elsevier 2015.
- Goldstein, E. B. Crosstalk between psychophysics and physiology in the study of perception. In E. B. Goldstein (Ed.). Blackwell handbook of perception (pp. 1–23). Blackwell Publishing. 2001.
- Xu, X.; Hanganu-Opatz,I.L. and Bieler, M. Cross-Talk of Low-Level Sensory and High-Level Cognitive Processing: Development, Mechanisms, and Relevance for Cross-Modal Abilities of the Brain. Front. Neurorobot. 2020, 14. [CrossRef]
- Logothetis, N.K. and Pauls, J. Psychophysical and Physiological Evidence for Viewer-centered Object Representations in the Primate. Cerebral Cortex 1995, 3, 270–288. [Google Scholar] [CrossRef]
- Mascalzoni, E.; Osorio, D.; Regolinm L. and Giorgio Vallortigara, G. Symmetry perception by poultry chicks and its implications for three-dimensional object recognition. Proc Biol Sci. 2012. 7;279(1730), 841-6. [CrossRef]
- Pizlo, Z. and de Barros, J. A. The Concept of Symmetry and the Theory of Perception Front. Comput. Neurosci., 2021, 15. [Google Scholar] [CrossRef]
- Nakade, Y. ; Iwata,Y.; Furuichi, K.; Mita, M.; Hamase, K.; Konno, R. et al. Gut microbiota-derived D-serine protects against acute kidney injury. The Journal of Clinical Investigation Insight 2018 18, 3(20), e97957. [CrossRef]
- Freud, S. An outline of psychoanalysis. Std Edn. Vol. 23. London: Vintage. 1940.
- Wada, K.; Yamamoto, M. and Nakashima, K. Psychological function in aging. Nihon Rinsho (Janan) 2013, 71(10).1713-9.
- Bottaccioli, A.G.; Bologna, M. and Bottaccioli, F. Psychic Life-Biological Molecule Bidirectional Relationship: Pathways, Mechanisms, and Consequences for Medical and Psychological Sciences—A Narrative Review. Int. J. Mol. Sci. 2022, 23, 3932. [Google Scholar] [CrossRef]
- Corballis, M. C., & Beale, I. L. Bilateral symmetry and behavior. Psychological Review 1970, 77(5), 451–464. 5. [CrossRef]
- Delvenne, J-F.; Castronovo, J.; Demeyere, N. and umphreys, G.W. Bilateral Field Advantage in Visual Enumeration PLOS 2011. [CrossRef]
- Tamaki Amano and Motomi Toichi. Zoi Kapoula, Editor. The Role of Alternating Bilateral Stimulation in Establishing Positive Cognition in EMDR. Therapy: A Multi-Channel Near-Infrared Spectroscopy Study. PLoS One, 2016; 11, e0162735. [CrossRef]
- Boukezzi, S.; Silva, C.; Nazarian, B.; Rousseau, P-F.; Guedj, E. and Valenzuela-Moguillansky, C. Bilateral Alternating Auditory Stimulations Facilitate Fear Extinction and Retrieval. Front. Psychol. Sec. Psychology for Clinical Settings 2017, 8. [CrossRef]
- Kasprian G, Langs G, Brugger PC, Bittner M, Weber M, Arantes M, et al. The prenatal origin of hemispheric asymmetry: an in-utero neuroimaging study. Cereb Cortex 2011, 21, 1076–1083. [CrossRef]
- Mitter, C.; Prayer, D.; Brugger, P.C.; Weber, M. and Kaspria, G. In Vivo Tractography of Fetal Association Fibers. PLOS. ONE 2015, 10(3), e011953. 3. [CrossRef]
- Than, K. Symmetry in Nature: Fundamental Fact or Human Bias. Live Science. 2005. Available online: https://www.livescience.com/4002-symmetry-nature-fundamental-fact-human-bias.html.
- Gunturkun, O and Ocklenburg, S. Ontogenesis of Lateralization. Neuron, 2017, 94(2), 249–263. [CrossRef]
- Mehler, M.F. Epigenetic Principles and Mechanisms Underlying Nervous System Functions in Health and Disease. Prog Neurobiol. 2008, 86(4): 305–341. [CrossRef]
- Zion, E. and Chen, X. Breaking Symmetry: The Asymmetries in Epigenetic Inheritance. Biochem (Lond). 2021. 43(1): 14–19. [CrossRef]
- Wan, L.Q.; Chin, A.S.; Worley, K.E. and Ray, P. Cell chirality: emergence of asymmetry from cell culture. Philos Trans R Soc Lond B Biol Sci. 2016, 371(1710), 20150413. [CrossRef]
- Sun. Na.; Dou, X.; Tang, Z.; Zhang, D.; Ni, N.; Wang, J.; Gao, H.; Ju, Y.; Dai, X.; Zhao, C.; Gu, P.; Ji, J. and Feng, C. Bio-inspired chiral self-assemblies promoted neuronal differentiation of retinal progenitor cells through activation of metabolic pathway. Bioactive Materials 2021, 6(4), 990-997. 4. [CrossRef]
- Dong, C., Madar, A.D. & Sheffield, M.E.J. Distinct place cell dynamics in CA1 and CA3 encode experience in new environments. Nat Commun 2021, 12, 2977. [CrossRef]
- Graham, H.K. and Duan, X. Molecular mechanisms regulating synaptic specificity and retinal circuit formation. Wiley Interdiscip Rev Dev Biol. 2021, 10(1), e379. 1. [CrossRef]
- Matho,K.S; Huilgol, D.; Galbavy, W.; He, M.; Kim, G.; Xu An, Xu.; Lu, J. et al. Genetic dissection of the glutamatergic neuron system in cerebral cortex. Nature. 2021, 598, 7879):182-187. [CrossRef]
- Bekkers. J.M. Pyramidal neurons. Current biology Curr Biol. 2011, 21(24), R975. 24. [CrossRef]
- Banovac, I.; Sedmak, D.; Džaja, D.; Jalšovec, D.; Milošević, N.J.; Rašin, [M.R. and Petanjek, Z. Somato-dendritic morphology and axon origin site specify von Economo neurons as a subclass of modified pyramidal neurons in the human anterior cingulate cortex. Journal of Anatomy 2019, 235, 3, 651-666. [CrossRef]
- Johns, P. Neurons and glial cells. Chapter 5 in Clinical Neuroscience, 2014, 61-69. [CrossRef]
- Sha, Z.; Schijven, D.; Carrion-Castillo, A.; Joliot, M.; Mazoyer, B.; Fisher, S.E.; Crivello, F. and Francks, C. The genetic architecture of structural left-right asymmetry of the human brain. Nature Human Behaviour 2021, 5, 1226–1239. [CrossRef]
- Krupic, J.; Bauza, M.; Burton, S.; Barry,C. and O’Keefe, J. Grid cell symmetry is shaped by environmental geometry. Nature. 2015, 518(7538): 232–235. [CrossRef]
- M. Oh, M.M.; Simkin, D. and Disterhoft, J.F. Intrinsic Hippocampal Excitability Changes of Opposite Signs and Different Origins in CA1 and CA3 Pyramidal Neurons Underlie Aging-Related Cognitive Deficits. Front. Syst. Neurosci. 2016, 10. [CrossRef]
- Constant M. and Mellet, E. The Impact of Handedness, Sex, and Cognitive Abilities on Left–Right Discrimination: A Behavioral Study. Front. Psychol. 2018, 9, 405. [CrossRef]
- Merrick, C.M.; Dixon, T.C.; Breska, A.; Lin, J.; Chang, E.F.; King-Stephens, D.; Laxer, K.D.; Weber, P.B.; Carmena, J.; Knight, R.T. and Ivry, R.B. Left hemisphere dominance for bilateral kinematic encoding in the human brain. eLife 2022, 11, e69977. [CrossRef]
- Jiang, S., Guan, Y., Chen, S. et al. Anatomically revealed morphological patterns of pyramidal neurons in layer 5 of the motor cortex. Sci Rep 2020, 10, 7916. [CrossRef] [PubMed]
- Kawakami, R.; Shinohara, Y.; Kato, Y.; Sugiyama, H.; Shigemoto, R, and Itoauthors, I. Asymmetrical allocation of NMDA receptor ε2 subunits in hippocampal sircuitry. Science 2003, 300(5621), 990-994. [CrossRef]
- Shinohara, Y.; Hirase, H.; Watanabe, M, Makoto Itakura, M.; Masami Takahashi, M.and Shigemoto, R. Edited by Huganir,R.L. Left-right asymmetry of the hippocampal synapses with differential subunit allocation of glutamate receptors. PNAS. 2008,105 (49) 19498-19503. [CrossRef]
- Muller, J.F.; Mascagni, F. and McDonald. A.J. Pyramidal Cells of the Rat Basolateral Amygdala. Synaptology and Innervation by Parvalbumin-immunoreactive Interneurons. J Comp Neurol. 2006, 494(4): 635–650. [CrossRef]
- Lorente de Nó R, L. Cerebral cortex: architecture, intracortical connections and motor projections. In: Physiology of the nervous system, 3rd edn (Fulton JF, ed.), pp. 288–330. Oxford: Oxford University Press. 1949.
- Chan, C.H.; Godinho, L.N.; Thomaidou, D.; Tan, S.S. ; Gulisano, M and Parnavelas J. G. Emx1 is a Marker for Pyramidal Neurons of the Cerebral Cortex. Cerebral Cortex 2001, 11(12), 1191–1198. [Google Scholar] [CrossRef]
- Hawkins, J. and Ahmad, S. Hypothesis and theory article. Why Neurons Have Thousands of Synapses, a Theory of Sequence Memory in Neocortex. Front. Neural Circuits, 2016, 10. [Google Scholar] [CrossRef] [PubMed]
- Parnavelas, JG.; Dinopoulos, A. and Davies SW. The central visual pathways. In: Handbook of chemical neuroanatomy, vol. 7. Integrated systems of the CNS, Part II (Björklund A, Hökfelt T, Swanson LW, eds), pp. 1–164. Amsterdam: Elsevier. 1989.
- Dallérac, G.; Li, X.; Lecouflet, P.; Morisot, N.; Sacchi, S.; Asselot, R.; Thu Ha Pham, T.H. et al. Dopaminergic neuromodulation of prefrontal cortex activity requires the NMDA receptor coagonist D-serine. Proc Natl Acad Sci U S A. 2021, 118(23): e2023750118. [CrossRef]
- O’Keefe, J. and Krupic, J. Do hippocampal pyramidal cells respond to nonspatial stimuli? Physiological review 2021, 101(3), 1427-145. [CrossRef]
- Purpura, D.P. and Suzuki, K. Distortion of neuronal geometry and formation of aberrant synapses in neuronal storage disease. Brain Research 1976, 116(1), 1-2. [CrossRef]
- Adámek, P.; Langová, V. Horáček, J. Early-stage visual perception impairment in schizophrenia, bottom-up and back again. Schizophrenia 2022 8, 27. [CrossRef]
- Granato, A. and De Giorgio, A. Alterations of neocortical pyramidal neurons: turning points in the genesis of mental retardation. Front. Pediatr. 2014. [CrossRef]
- Rennó-Costa, C. and Adriano Tort, A.B.L. Place and Grid Cells in a Loop: Implications for Memory Function and Spatial Coding. J Neurosci. 2017, 37(34), 8062-8076. [CrossRef]
- Kitanishi, T. , Ito, H.T., Hayashi, Y. et al. Network mechanisms of hippocampal laterality, place coding, and goal-directed navigation. J Physiol Sci, 2017; 67, 247–258. [Google Scholar] [CrossRef]
- Zhong, S.; He, Y.; Shu, H. and Gaolang Gong, G. Developmental Changes in Topological Asymmetry Between Hemispheric Brain White Matter Networks from Adolescence to Young Adulthood. Cerebral Cortex, 2017; 7, 2560–2570. [Google Scholar] [CrossRef]
- Goriounova, N.A.; Heyer, D.B.; Wilbers, R.; Verhoog, M.B.; Giugliano, M.; Christophe Verbist, C. et al. Large and fast human pyramidal neurons associated with intelligence. Elife. 2018, 18(7), e41714. [CrossRef]
- Kawakami, R.; Shinohara, Y. ; Kato,Y.; Sugiyama, H.; Shigemoto, R. and Ito, I. Asymmetrical Allocation of NMDA Receptor ε2 Subunits in Hippocampal Circuitry. Science New Series 2003, 300(621), 990-994. [CrossRef]
- Sá, M.J.; Ruela, C. and Madeira, M.D. Dendritic right/left asymmetries in the neurons of the human hippocampal formation: a quantitative Golgi study. Arq Neuropsiquiatr 2007, 65(4B), 1105-13. [CrossRef]
- Shinohara, Y.; Hirase, H.; Watanabe, M. ; +2, and Ryuichi Shigemoto, R. Left-right asymmetry of the hippocampal synapses with differential subunit allocation of glutamate receptors. PNAS 2008, 105 (49) 19498-19503. [CrossRef]
- Goto, K.; Kurashima, R.; Gokan, H.; Inoue, N.; Ito, I. and Watanabe, S. Left−Right Asymmetry Defect in the Hippocampal Circuitry Impairs Spatial Learning and Working Memory in iv Mice. PLOS ONE 2010, 5(11), e15468.
- Ukai, H.; Kawahara, A.; Hirayama, K.; Show all 17 Ito Isao, I. ItPirB regulates asymmetries in hippocampal circuitry. PLOS ONE 2017, 12(6). [CrossRef]
- Luengo-Sanchez, S.; Bielza, C.; Benavides-Piccione, R.; Fernaud-Espinosa, I.; DeFelipe, J. and Larrañaga, P. A univocal definition of the neuronal soma morphology using Gaussian mixture models. Front. Neuroanat., 2015, 9. [CrossRef]
- Rockland, K.S. Pyramidal Neurons: Looking for the origins of axons. Evolutionary Biology. Neuroscience 2022. [CrossRef]
- Abrous, D.N. , Koehl, M. & Lemoine, M. A. Baldwin interpretation of adult hippocampal neurogenesis: from functional relevance to physiopathology. Mol Psychiatry 2022. 27, 383–402. [CrossRef]
- Leguey, I.; Benavides-Piccione, R.; Rojo, C.; Larrañaga, P.; Bielza, C. and DeFelipe, J. Neurons in the Rat Somatosensory Cortex. eNeuro. 2019, 5(6). [CrossRef]
- Weiler, S.; Nilo, D.G.; Bonhoeffer, T.; Hübener, M.; Rose, T. and Scheuss, V. Orientation and direction tuning align with dendritic morphology and spatial connectivity in mouse visual cortex. Current Biology 2020, 32(8), 1743-1753.e7. [CrossRef]
- Wahle, P.; Sobierajski, E.; Gasterstädt, I. Lehmann,N.; Weber, S.; Lübke, H.R.; Engelhardt, M.; Distler, C. and Meyer, G. Neocortical pyramidal neurons with axons emerging from dendrites are frequent in non-primates, but rare in monkey and human. eLife. 2022, 11, e76101. [CrossRef]
- Musall, S. , Sun, X.R., Mohan, H. et al. Pyramidal cell types drive functionally distinct cortical activity patterns during decision-making. Nat Neurosci 2023. [CrossRef]
- Park, J.; Papoutsi, A.; Ash, R.T. et al. Contribution of apical and basal dendrites to orientation encoding in mouse V1 L2/3 pyramidal neurons. Nat Commun 10, 5372 (2019). [CrossRef]
- Elaine, M. Pinheiro, E.M.; Xie, Z.; Norovich, A.L.; Vidaki, M.; Tsai, L-H. and Gertler, F.B. Lpd depletion reveals that SRF specifies radial versus tangential migration of pyramidal neurons. Nat Cell Biol. 2011, 13(8), 989–995. [CrossRef]
- Hobert, O. Development of left/right asymmetry in the Caenorhabditis elegans nervous system: From zygote to postmitotic neuron. Genesis 2014, 52(6), 528–543. [Google Scholar] [CrossRef]
- Tee, Y. , Shemesh, T., Thiagarajan, V. et al. Cellular chirality arising from the self-organization of the actin cytoskeleton. Nat Cell Biol 2015.17, 445–457. [CrossRef]
- Radler, M.R.; Liu, X.; Peng, M.; Doyle, B.; Toyo-Oka, K. and Spiliotis, E.T.Pyramidal neuron morphogenesis requires a septin network that stabilizes filopodia and suppresses lamellipodia during neurite initiation. Current Biology 2023, 33(3), P434-448.E8. [CrossRef]
- Inaki, M.; Sasamura. T. and Kenji Matsuno, K. Review. Cell Chirality Drives Left-Right Asymmetric Morphogenesis. Front. Cell Dev. Biol. 2018, 6. [CrossRef]
- Konietzny,A. ; Bär, J. and Mikhaylova, M. MINI REVIEW. Dendritic Actin Cytoskeleton: Structure, Functions, and Regulations. Front. Cell. Neurosci. Sec. Cellular Neurophysiology 2017, 11. [Google Scholar] [CrossRef]
- Satir, P. Chirality of the cytoskeleton in the origins of cellular asymmetry Philos Trans R Soc Lond B Biol Sci. 2016, 371(1710), 20150408. [CrossRef]
- Parato, J. and Bartolini, F.The microtubule cytoskeleton at the synapse Neurosci Lett. 2021, 14;753,135850. [CrossRef]
- Bocquet, A.; Berges, R.; Frank, R.; Robert, P.; Peterson, A.C. and Eyer, J. Neurofilaments Bind Tubulin and Modulate Its Polymerization. J Neurosci. 2009, 29(35), 11043–11054. [CrossRef] [PubMed]
- Uylings, H.B.; Jacobsen, A.M.; Zilles, K. and Amunts, K. Left-right asymmetry in volume and number of neurons in adult Broca’s area. Cortex. 2006, 42(4), 652-8. [CrossRef]
- El-Gaby, M.; Reeve, H.M. ; Lopes-dos-Santos,V.; Campo-Urriza, N.; Perestenko, P.V. Morley, A.; Strickland, L.A.M.; Lukács, I.P.; Paulsen, O. and Dupret, D. An Emergent Neural Coactivity Code for Dynamic Memory. Nat Neurosci. 2021, 24(5): 694–704. [CrossRef]
- Cullen, T.J.; Walker, M.A.; Eastwood, S.L.; Esiri, M.M.; Harrison, P.J. and Crow, T.J. Anomalies of asymmetry of pyramidal cell density and structure in dorsolateral prefrontal cortex in schizophrenia Br J Psychiatry 2006, 188:26-31. [CrossRef]
- Souta, R. An Introductory Perspective on the Emerging Application of qEEG. In the book Introduction to Quantitative EEG and Neurofeedback. Advsanced Theory and Applications. By Budzynski, T.H.; Washington, P.; Budzynski, H.K.; Washington, P.; Evans, J.R. and Abarbanel, A. Sec Ed. Acad. Press. 2009.
- Zhu, DY. , Cao, TT., Fan, HW. et al. The increased in vivo firing of pyramidal cells but not interneurons in the anterior cingulate cortex after neuropathic pain. Mol Brain. 2022, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Kong, X-Z.; Postema, M.C.; Guadalupe, T.; de Kovel, C.; Boedhoe, P.C.W.; Hoogman, M.; Mathias, S.R.; van Rooij, D.; Schijven, D.; Glahn, D.C.; Medland, S.E. et al., Mapping brain asymmety in health and disease through the ENIGMA consortium. Hum Brain Mapp. 2022, 43(1), 167–181. [CrossRef]
- Kay, J.W. and Phillips, W.A. Contextual Modulation in Mammalian Neocortex is Asymmetric. Symmetry 2020, 12(5), 815. [CrossRef]
- Tripathi, S. Dey,A.; Shanmugam, M.; Narayanan, R.S.and Chandrsekhar, V. Cobalt (II) Complexes as Single-Ion Magnets. Topics in Organometallic Chem 2019, 64. [CrossRef]
- Kagamiyama, H. and Hayashi, H. Crystallographic Structures. Branched-Chain Amino Acids, Part B. In book Methods in Enzymoligy. Elsevier 2000.
- Housecroft, C. E.; Sharpe, A. G. Inorganic Chemistry (2nd ed.). 2004. Prentice Hall. ISBN 978-0-13-039913-7.
- Gradišar, H.; Božič, S.; Doles, T.; Vengust, D.; Hafner-Bratkovič, I.; Mertelj, A.; Webb, B.; Šali, A.; Klavžar, S. and Roman Jerala, R. Design of a single-chain polypeptide tetrahedron assembled from coiled-coil segments. Nat Chem Biol. 2013, 9(6), 362–366. [CrossRef]
- Xie, N.; Liu, S.; Yang, X.; Xiaoxiao, He.; Huang, J. and Kemin Wang, K. DNA tetrahedron nanostructures for biological applications: biosensors and drug delivery. Analyst 2017, 18(142) 3322-3332. [CrossRef]
- Strychalski, W. 3D Computational Modeling of Bleb Initiation Dynamics. Front. Phys., Sec. Biophysics 2021, 9. [Google Scholar] [CrossRef]
- Yoo, S.H. and Lee, H-S. Foldectures: 3D Molecular Architectures from Self-Assembly of Peptide Foldamers. Acc. Chem. Res. 2017, 50, 4, 832–841. [Google Scholar] [CrossRef]
- Sasaki, E. , Böhringer, D., van de Waterbeemd, M. et al. Structure and assembly of scalable porous protein cages. Nat Commun 2017, 8, 14663. [Google Scholar] [CrossRef]
- Quaye, I.K. Extracellular hemoglobin: the case of a friend turned foe. Front Physiol. 2015; 6, 96. [CrossRef]
- Eaton, A.W. Retrospective on statistical mechanical models for hemoglobin allostery editors-pick J. Chem. Phys. The Journal of Chemical Physics 2022, 157, 184104. [Google Scholar] [CrossRef] [PubMed]
- Richter, F.; Meurers, B.H.; Zhu, C.; Medvedeva, V.P. and Chesselet, M-F. Neurons Express Hemoglobin α- and β-Chains in Rat and Human Brains. J Comp Neurol. 2009, 15(5): 538–547. [CrossRef]
- Walser, M.; Svensson, J.; Karlsson, Motalleb, R. ; Åberg, M.; Kuhn, H.G.; Isgaard, J. and Åberg, N.D. Growth Hormone and Neuronal Hemoglobin in the Brain—Roles in Neuroprotection and Neurodegenerative Diseases. Front. Endocrinol. Sec. Neuroendocrine Science 2021, 11. [CrossRef]
- Zheng, R.; Yan, Y.; Pu, J. and Zhang, B. Physiological and Pathological Functions of Neuronal Hemoglobin: A Key Underappreciated Protein in Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23(16), 9088. [CrossRef]
- Flynn, K.S. The cytoskeleton and neurite initiation. Bioarchitecture. 2013, 3(4): 86–109. [CrossRef]
- Giuliana Indelicato, Newton Wahome, Philippe Ringler. Shirley A. Müller, Mu-Ping Nieh, Peter Burkhard, Reidun Twarock. Principles Governing the Self-Assembly of Coiled-Coil Protein Nanoparticles. Biophysical Journal 2016, 110(3), P646-660. [CrossRef]
Figure 1.
(top). Transfer of symmetry constraints from molecular to cellular level. a) Tetrahedron–geometrical shape which can be transformed from chiral to achiral version. b) Tetrahedral (sp3-hybridized) carbons with four different substituents are the principal components of biological molecules exhibiting homochirality. c) Tetrahedral structure of water forming about 70 % of cytosol. d) L/D amino acids. e) Tetrahedral structure of hemoglobin (Hb) expressed in neuronal and red blood cells. f) Tetrahedral assembly of DNA. g) Pyramidal neuron. Adopted with changes from [5,6]. PNs were discovered by the Ukrainian anatomist and histologist Vladimir A. Betz (1834–1894) [1]. (bottom). Tree of life. Successive biologically essential evolutionary selections: I—prevalent molecular chirality, II—bilaterality, III—pyramidal neurons, IV—human mind. Adopted from Encyclopedia Britannica Inc. 2012 with alterations.
Figure 1.
(top). Transfer of symmetry constraints from molecular to cellular level. a) Tetrahedron–geometrical shape which can be transformed from chiral to achiral version. b) Tetrahedral (sp3-hybridized) carbons with four different substituents are the principal components of biological molecules exhibiting homochirality. c) Tetrahedral structure of water forming about 70 % of cytosol. d) L/D amino acids. e) Tetrahedral structure of hemoglobin (Hb) expressed in neuronal and red blood cells. f) Tetrahedral assembly of DNA. g) Pyramidal neuron. Adopted with changes from [5,6]. PNs were discovered by the Ukrainian anatomist and histologist Vladimir A. Betz (1834–1894) [1]. (bottom). Tree of life. Successive biologically essential evolutionary selections: I—prevalent molecular chirality, II—bilaterality, III—pyramidal neurons, IV—human mind. Adopted from Encyclopedia Britannica Inc. 2012 with alterations.
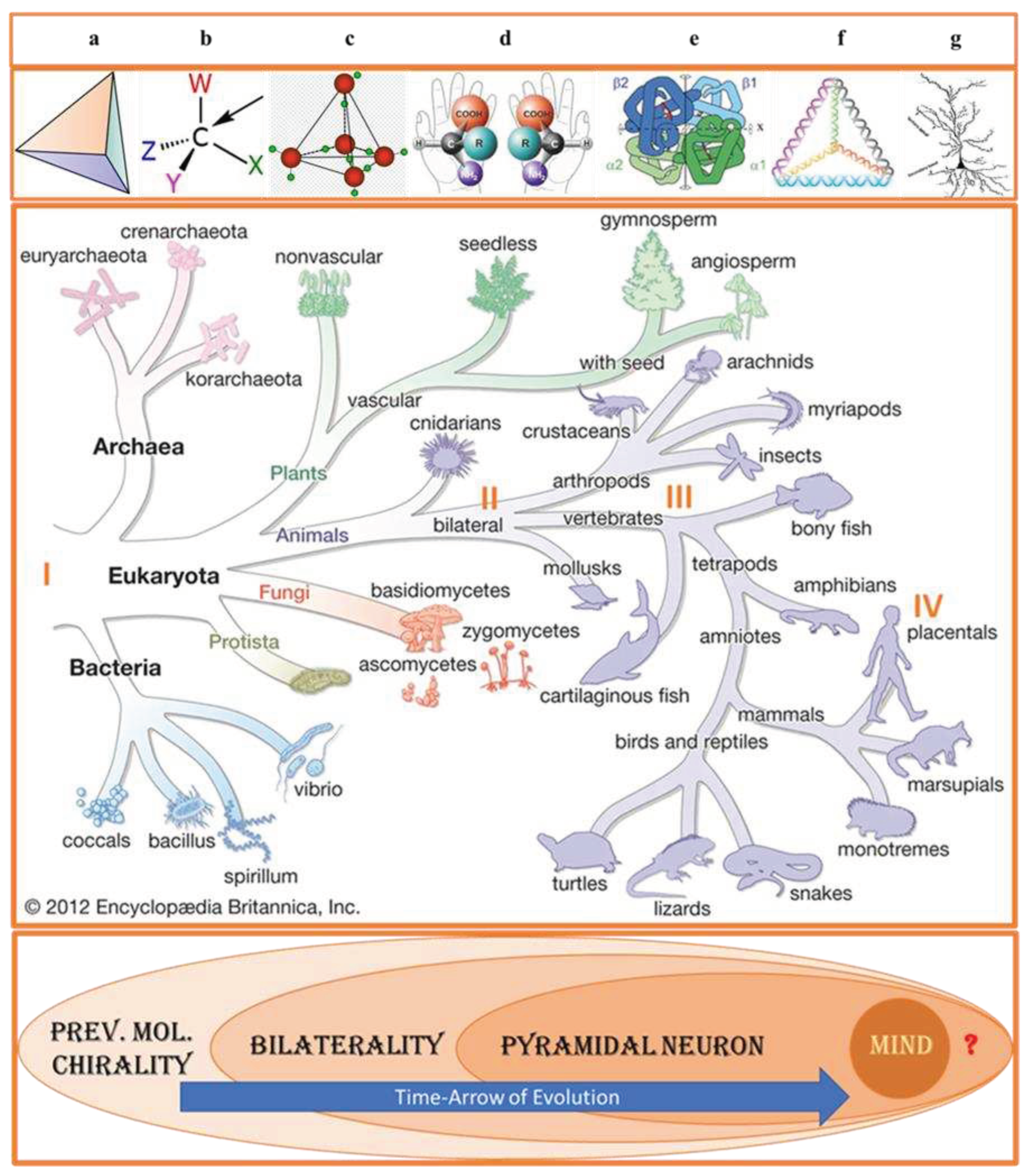
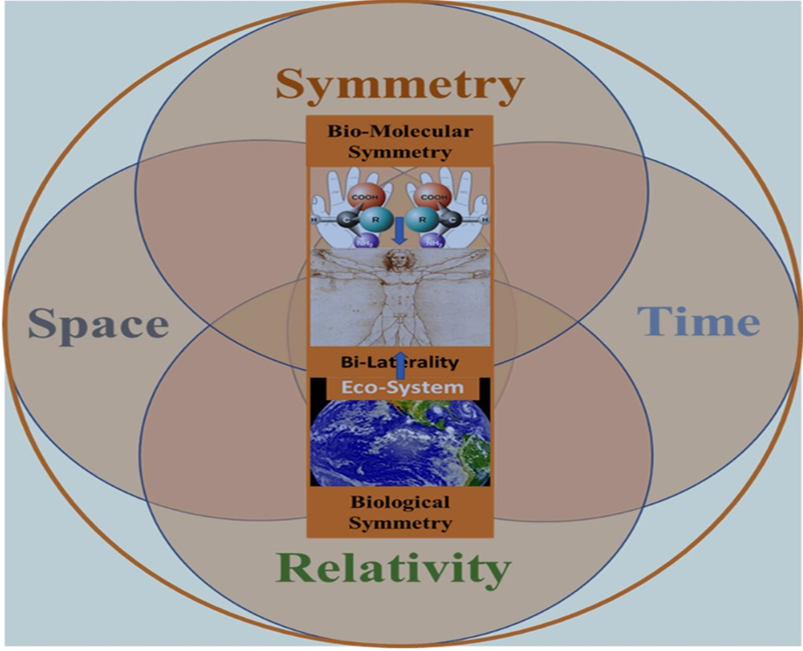
Figure 2.
Two pillars of human biology: homochirality and bilaterality. A broad view of biological symmetry assumes that chiral expression from the molecular to macromolecular and cellular levels plays a vital role in physiology underlying perceptual and cognitive function linked with the emotional and behavioral expression of bilateral organisms.
Figure 2.
Two pillars of human biology: homochirality and bilaterality. A broad view of biological symmetry assumes that chiral expression from the molecular to macromolecular and cellular levels plays a vital role in physiology underlying perceptual and cognitive function linked with the emotional and behavioral expression of bilateral organisms.

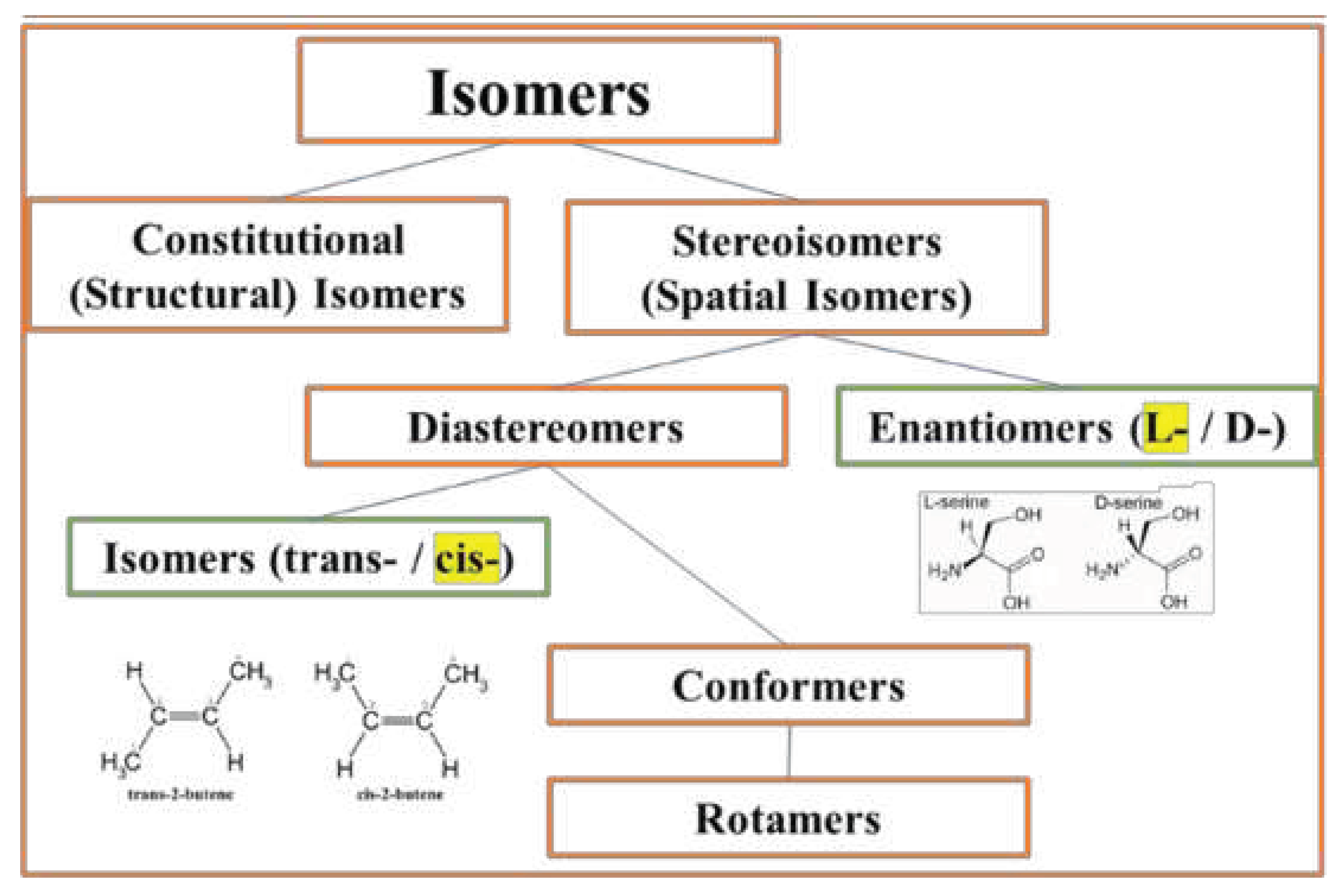
Figure 3.
At the molecular level, biological symmetry preference among stereoisomers exists in two forms: the preference for L-enantiomers (chiral) and cis diastereomers (achiral). While enantiomers can only come in pairs, many diastereomers exist for a given molecule.
Figure 3.
At the molecular level, biological symmetry preference among stereoisomers exists in two forms: the preference for L-enantiomers (chiral) and cis diastereomers (achiral). While enantiomers can only come in pairs, many diastereomers exist for a given molecule.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



